cytology of vicia species. x. karyotype evolution and phylogenetic implication in vicia species of...
TRANSCRIPT

ORIGINAL ARTICLE
Cytology of Vicia species. X. Karyotype evolutionand phylogenetic implication in Vicia species of the sectionsAtossa, Microcarinae, Wiggersia and Vicia
M. Ruffini Castiglione & M. Frediani & M. T. Gelati &C. Ravalli & G. Venora & P. Caputo & R. Cremonini
Received: 28 June 2010 /Accepted: 21 October 2010# Springer-Verlag 2010
Abstract Automated karyotype analyses and sequence ofrDNA spacers have been analysed for the species belongingto sections Atossa, Microcarinae, Wiggersia and Vicia.Karyomorphological parameters, based on Rec, Syi and TF% indices, have been determined and evidenced that, interm of symmetry, the karyotype of Vicia lathyroides wasthe most asymmetric one. A multivariate analysis using 34karyological parameters, in addition to the symmetryindices, has been carried out and the correspondingdendrogram of linkage distances showed six differentgroups. Molecular investigations on the inclusive group instudy by employing ITS DNA sequences indicated adifferent pattern of relationships. The cladistic analysiscombining the molecular data set with karyological param-eters evidenced that the species of sections Vicia and Atossa
join closely to each other in a paraphyletic group, whichincludes the monophyletic section Wiggersia. Therefore,our karyological and molecular data provide informationabout the phylogenetic position of the analysed speciesinside the subgenus Vicia and are discussed in relation toprevious results obtained by morphology, isozymes andribosomal genes analyses.
Keywords Automated karyotype analysis . Genes codingfor rRNA . Phylogeny . Vicia species
Introduction
The genus Vicia, including approximately 190 species(ILDIS 1999), is a member of the legume tribe Vicieae(Fabaceae). Ball (1968) divided the genus into foursections: Vicia, Cracca, Ervum and Faba while Kupicha(1976) subsequently recognised two subgenera, Vicilla andVicia, with 17 and five sections, respectively. SubgenusVicia contains fewer species than Vicilla but includes themore agriculturally important species. Elucidating thetaxonomic relationship between the crops and their wildrelatives is important, especially in connection with theexploitation of the germplasm of wild relatives as a sourceof novel characteristics to be introduced into the crop plantsby wide crosses.
More recently, Maxted (1993), on the basis of pheneticstudy, subdivided the subgenus Vicia into nine series, 38species, 14 subspecies and 22 varieties, and substituted thesection Faba sensu Kupicha with three distinct section:Bithynicae, Faba and Narbonensis.
Various aspects of the classification proposed by Maxted(1993) have been recently confirmed with different meth-odological approaches (Fennell et al. 1998; Potokina et al.
Handling Editor: Peter Nick
M. Ruffini Castiglione (*) :R. CremoniniDipartimento di Biologia, Università di Pisa,Via L. Ghini 5,56126, Pisa, Italye-mail: [email protected]
M. Frediani :M. T. GelatiDipartimento di Agrobiologia ed Agrochimica,Università della Tuscia,Via S. C. de Lellis,01100, Viterbo, Italy
C. Ravalli :G. VenoraStazione Sperimentale per la Granicoltura,Via Bouganvillea 20,95041, Caltagirone (CT), Italy
P. CaputoDipartimento delle Scienze Biologiche,Università di Napoli Federico II,Via Foria 233,80139, Napoli, Italy
ProtoplasmaDOI 10.1007/s00709-010-0232-7

1999; Venora et al. 2000; Leht and Jaaska 2002; Frediani etal. 2005; Caputo et al. 2006; Choi et al. 2006; Jaaska andLeht 2007; Endo et al. 2008). To improve understanding ofthe relationships within subgenus Vicia, we started theanalysis of its nine sections by cytological, karyologicaland molecular methods. Results for sections Bithynicae,Faba, Narbonensis, Peregrinae and Hypechusa and par-tially for section Atossa have been already published(Venora et al. 2000; Frediani et al. 2004; Frediani et al.2005; Caputo et al. 2006; Ruffini Castiglione et al. 2007and 2009). In the present work, our attention was focusedon sections Atossa, Microcarinae, Wiggersia and Vicia. Thesection Atossa includes four species (Vicia oroboides, Viciabalansae, Vicia abbreviata and Vicia sepium) belonging tothree series. This section is widely spread from Iceland toJapan, but the distribution is largely due to V. sepiumbecause the other species are much more restricted. Thesection Microcarinae includes only Vicia dionysiensis, anendemic species in Southern Syria, restricted to basaltic andvolcanic soils. The two species of section Wiggersia areVicia cuspidata, restricted to Southeast Europe and South-west Asia, and Vicia lathyroides, that makes wider thedistribution of the section to include North Africa andWestern Europe. The section Vicia includes five species intwo series. The centre of distribution is Southeast Europeand Southwest Asia, while two species, Vicia pyrenaica andVicia qatmensis, are restricted to Spain and Syria, respec-tively. This section includes one of the most commontemperate weeds (Vicia sativa) having many subspecieswith different chromosome numbers (2n=10, 12, 14). Also
the present work on the four sections above described wasperformed following a multidisciplinary approach.
The use of modern image analysis systems hasallowed the production of detailed karyotypes of plantspecies employed for interspecific comparison (Venora etal. 2008). Moreover, multivariated analysis using severalkaryomorphological parameters in addition to symmetryindices allows the construction of the dendrograms oflinkage distance rather resolved and very useful tocompare and to include in phylogenetic trees obtained bymolecular elaborations.
The data generated from the comparison of rDNAsequences and karyomorphological parameters havebeen utilised for evaluating the karyotype evolutionand phylogenetic relationships of the species belongingto the residual sections that supplement the subgenusVicia.
Materials and methods
Plant material
The names, source and accession numbers of the selectedspecies are listed in Table 1.
Karyomorphometry
Slides were prepared according to Venora et al. (1991). Atleast five metaphases for each of five seedlings were
Table 1 Names, accessions numbers, source and origin of the analysed Vicia species
Section Series Species Accessionnumber
Sourcea Origin
Atossa (Alef.) Asch.& Graebener Pseudovicilla Maxted V. oroboides Wulfen in Jacq. Botanic garden Ljubljana Republicof Slovenia
Truncatulae (B.Fedtsch. Ex Radzhi )Maxted
V. balansae Boiss. No seeds
Atossa Asch. & Graebener V. sepium var. sepium L. PI 440768 USDA Kazakhstan
Microcarinae Maxted V. dionysiensis Mout IG 63377 ICARDA Syria
Wiggersia Maxted V. cuspidata Boiss. IG 63208 ICARDA Syria
V. lathyroides L. PI 422500 USDA Germany
Vicia L. Vicia L. V. pyrenaica Pourret Vic 69/43 IPK Spain
V. sativa subsp. sativa L. PI 253426 USDA Spain
V. barbazitaeTen. & Guss.
No seeds
Grandiflorae B. Fedtsch. Ex Radzhi V. qatmensis Gomb. IG 64141 ICARDA Syria
V. grandiflora Scop. IG 64144 ICARDA Syria
a ICARDA International Centre for Agricultural Research in the Dry Areas, Aleppo, Syria
USDA United States Department of Agriculture, Pullman, WA, USA
IPK Institute of Plant Genetics and Crop Plant Research, Gatersleben, Federal Republic of Germany
M. Ruffini Castiglione et al.

analysed. Microscopic investigation was carried out by aZeiss Axioplan 2 microscope (Carl Zeiss Jena GmbH, Jena,Federal Republic of Germany) connected to a KS400 Zeissimage analysis system, with the dedicated karyotypingsoftware Ikaros 3.4 (Metasystem GmbH, Altlussheim,Federal Republic of Germany).
The automated image analysis system ensures a highdegree of accuracy since measurements are taken directly inthe first image of the chromosomes (Venora et al. 1991);therefore, this method allows a precise determination of thekaryomorphological indices (TF%, Rec and SYi). Totallengths of short arms, long arms and satellites weremeasured with the computer system. The classification ofStebbins (1971), the TF% index (Huziwara 1962), and theRec and SYi indices (Greilhuber and Speta 1976) werecomputed for all the species.
The TF% index is expressed by the ratio between thesum of the lengths of the short arms of individualchromosomes and the total length of the complement; theRec index expresses the average of the ratios between thelength of each chromosome and that of the longest one; theSYi value indicates the ratio between the average length ofthe short arms and the average length of the long arms.
Chromosome pairs were grouped by using clusteranalysis following ANOVA (Scott and Knott 1974);nomenclature by Levan et al. (1964) was followed,excluding the satellite length in computing arm ratios.
A hierarchical cluster analysis using Euclidean distances(SPSS release 14 Inv 1989-2006 statistic package) was alsoemployed to compare all the karyological data of theanalysed species.
DNA sequencing analysis
Nuclear DNA was extracted and purified from secondaryroots produced after decapitation of the primary one andfrom young leaves of plantlets. The entire region includingITS1, 5.8S and ITS2 rDNA was amplified by standardpolymerase chain reaction (PCR). Amplification wascarried out using the following parameters: 95°C for5 min, followed by 35 cycles of 95°C for 1 min, 55°C for2 min, 72°C for 2 min, and finally 72°C for 5 min.
The primers, designed by using the conserved regions ofthe 18S and 26S ribosomal genes, were: 18Sdir 5′-CGTAACAAGGTTTCCGTAGG-3′ and 26Scom 5′-AGCGGGTAGTCCCGCCTGA-3′. The PCR-amplifiedDNA fragments showed a single band when examined onagarose gel. The PCR products were sequenced directly forboth strands by using the automated sequencer ABI Prism310 (Applied Biosystems, USA).
ITS rDNA sequences of the species of sectionsMicrocarinae, Vicia, Wiggersia and Atossa, including V.oroboides, previously investigated by Ruffini Castiglione
et al. (2009), were aligned by using the Clustal 1.83software with default values (Thompson et al. 1994).Alignments were carried out as daughter processes ofBioedit (Hall 1999), which was also used for sequenceediting and manipulation.
A cladistic analysis was carried out by employing thedata handling environment Winclada (Nixon 1999),running Nona (Goloboff 1999) as a daughter process.Settings included: a maximum storage space of 10,000 trees(hold 10,000); a tree storage space per iteration of 100(hold/100); 100 iterations of the default algorithm(mult*100) followed by deep branch swapping on thefound trees (max). Cladograms were further manipulatedwith Winclada, which was also used for 1,000 replicas of abootstrap analysis (Felsenstein 1985); Nona wasemployed for computation of Bremer support (Bremer1994). A second cladistic analysis was carried out addingalso the karyological parameters reported in Table 5. Onlyparameters from code n. 4 to code n. 37 were added. Thiscladistic analysis, which included also 34 continuouscharacters, was carried out by using the TNT software(Goloboff et al. 2003), using the same parameters asabove. The karyological characters were regarded asordered, according to the software specifications. Boot-strap and Bremer support for this second investigationwere computed with TNT.
Results
Karyotyping and karyomorphometry
The chromosome number and the nuclear DNA content ofVicia species are reported in Table 2. Most analysed speciespossess 14 chromosomes, while four species have 12chromosomes: V. dionysiensis (sect. Microcarinae),V. cuspidata and V. lathyroides (sect. Wiggersia) and V.sativa subsp. sativa (sect. Vicia). The reported DNAcontents evidence genome size diversity, even within eachsection, ranging from 7.1 pg (V. oroboides) to 27.60 pg (V.dionysiensis).
Figure 1 shows idiograms of haploid complement of theanalysed species of sect. Atossa (a), sect. Microcarinae (b),sect. Wiggersia (c) and sect. Vicia (d).
All karyological data are reported in Table 3; the Rec,SYi as well as the TF% indices are summarised in Table 4.
The species belonging to the section Atossa present thecomplement 2n=14 and two different karyotype formulas.Inside this section, V. sepium var. sepium shows the highestvalues for SYi and TF% indices and V. oroboides thehighest value for Rec index.
The sections Microcarinae and Wiggersia include one andtwo species respectively, all possessing 2n=12 chromosomes.
Cytology of Vicia species
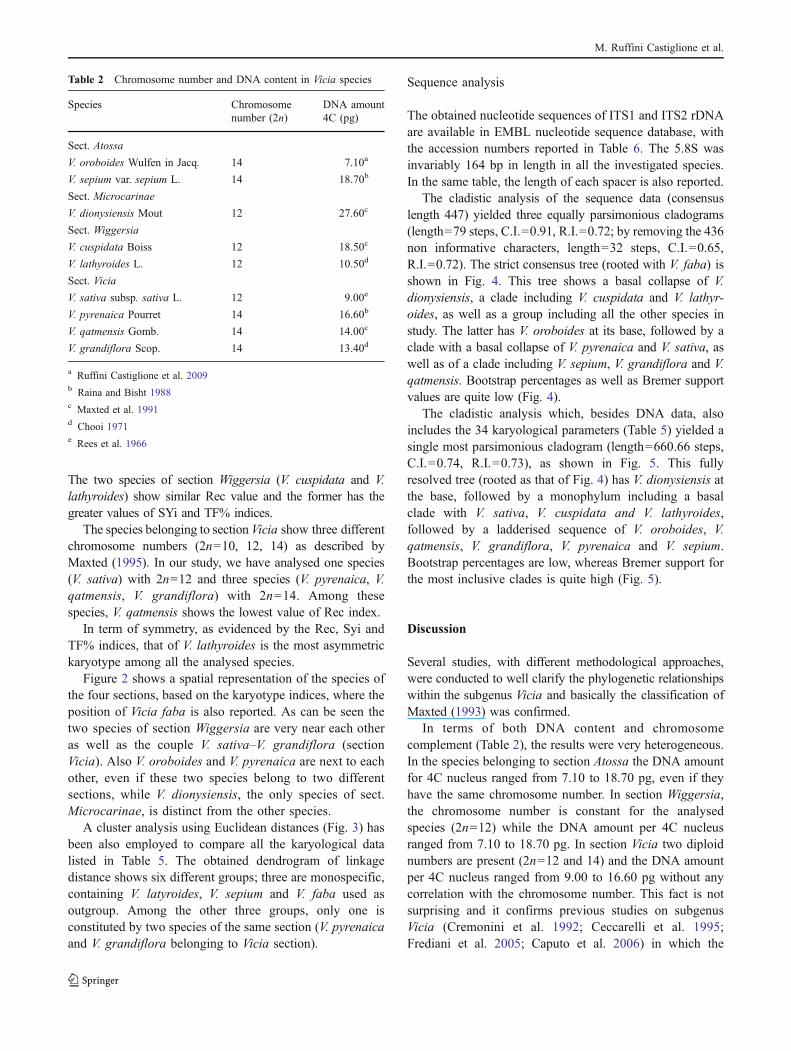
The two species of section Wiggersia (V. cuspidata and V.lathyroides) show similar Rec value and the former has thegreater values of SYi and TF% indices.
The species belonging to section Vicia show three differentchromosome numbers (2n=10, 12, 14) as described byMaxted (1995). In our study, we have analysed one species(V. sativa) with 2n=12 and three species (V. pyrenaica, V.qatmensis, V. grandiflora) with 2n=14. Among thesespecies, V. qatmensis shows the lowest value of Rec index.
In term of symmetry, as evidenced by the Rec, Syi andTF% indices, that of V. lathyroides is the most asymmetrickaryotype among all the analysed species.
Figure 2 shows a spatial representation of the species ofthe four sections, based on the karyotype indices, where theposition of Vicia faba is also reported. As can be seen thetwo species of section Wiggersia are very near each otheras well as the couple V. sativa–V. grandiflora (sectionVicia). Also V. oroboides and V. pyrenaica are next to eachother, even if these two species belong to two differentsections, while V. dionysiensis, the only species of sect.Microcarinae, is distinct from the other species.
A cluster analysis using Euclidean distances (Fig. 3) hasbeen also employed to compare all the karyological datalisted in Table 5. The obtained dendrogram of linkagedistance shows six different groups; three are monospecific,containing V. latyroides, V. sepium and V. faba used asoutgroup. Among the other three groups, only one isconstituted by two species of the same section (V. pyrenaicaand V. grandiflora belonging to Vicia section).
Sequence analysis
The obtained nucleotide sequences of ITS1 and ITS2 rDNAare available in EMBL nucleotide sequence database, withthe accession numbers reported in Table 6. The 5.8S wasinvariably 164 bp in length in all the investigated species.In the same table, the length of each spacer is also reported.
The cladistic analysis of the sequence data (consensuslength 447) yielded three equally parsimonious cladograms(length=79 steps, C.I.=0.91, R.I.=0.72; by removing the 436non informative characters, length=32 steps, C.I.=0.65,R.I.=0.72). The strict consensus tree (rooted with V. faba) isshown in Fig. 4. This tree shows a basal collapse of V.dionysiensis, a clade including V. cuspidata and V. lathyr-oides, as well as a group including all the other species instudy. The latter has V. oroboides at its base, followed by aclade with a basal collapse of V. pyrenaica and V. sativa, aswell as of a clade including V. sepium, V. grandiflora and V.qatmensis. Bootstrap percentages as well as Bremer supportvalues are quite low (Fig. 4).
The cladistic analysis which, besides DNA data, alsoincludes the 34 karyological parameters (Table 5) yielded asingle most parsimonious cladogram (length=660.66 steps,C.I.=0.74, R.I.=0.73), as shown in Fig. 5. This fullyresolved tree (rooted as that of Fig. 4) has V. dionysiensis atthe base, followed by a monophylum including a basalclade with V. sativa, V. cuspidata and V. lathyroides,followed by a ladderised sequence of V. oroboides, V.qatmensis, V. grandiflora, V. pyrenaica and V. sepium.Bootstrap percentages are low, whereas Bremer support forthe most inclusive clades is quite high (Fig. 5).
Discussion
Several studies, with different methodological approaches,were conducted to well clarify the phylogenetic relationshipswithin the subgenus Vicia and basically the classification ofMaxted (1993) was confirmed.
In terms of both DNA content and chromosomecomplement (Table 2), the results were very heterogeneous.In the species belonging to section Atossa the DNA amountfor 4C nucleus ranged from 7.10 to 18.70 pg, even if theyhave the same chromosome number. In section Wiggersia,the chromosome number is constant for the analysedspecies (2n=12) while the DNA amount per 4C nucleusranged from 7.10 to 18.70 pg. In section Vicia two diploidnumbers are present (2n=12 and 14) and the DNA amountper 4C nucleus ranged from 9.00 to 16.60 pg without anycorrelation with the chromosome number. This fact is notsurprising and it confirms previous studies on subgenusVicia (Cremonini et al. 1992; Ceccarelli et al. 1995;Frediani et al. 2005; Caputo et al. 2006) in which the
Table 2 Chromosome number and DNA content in Vicia species
Species Chromosomenumber (2n)
DNA amount4C (pg)
Sect. Atossa
V. oroboides Wulfen in Jacq. 14 7.10a
V. sepium var. sepium L. 14 18.70b
Sect. Microcarinae
V. dionysiensis Mout 12 27.60c
Sect. Wiggersia
V. cuspidata Boiss 12 18.50c
V. lathyroides L. 12 10.50d
Sect. Vicia
V. sativa subsp. sativa L. 12 9.00e
V. pyrenaica Pourret 14 16.60b
V. qatmensis Gomb. 14 14.00c
V. grandiflora Scop. 14 13.40d
a Ruffini Castiglione et al. 2009b Raina and Bisht 1988c Maxted et al. 1991d Chooi 1971e Rees et al. 1966
M. Ruffini Castiglione et al.
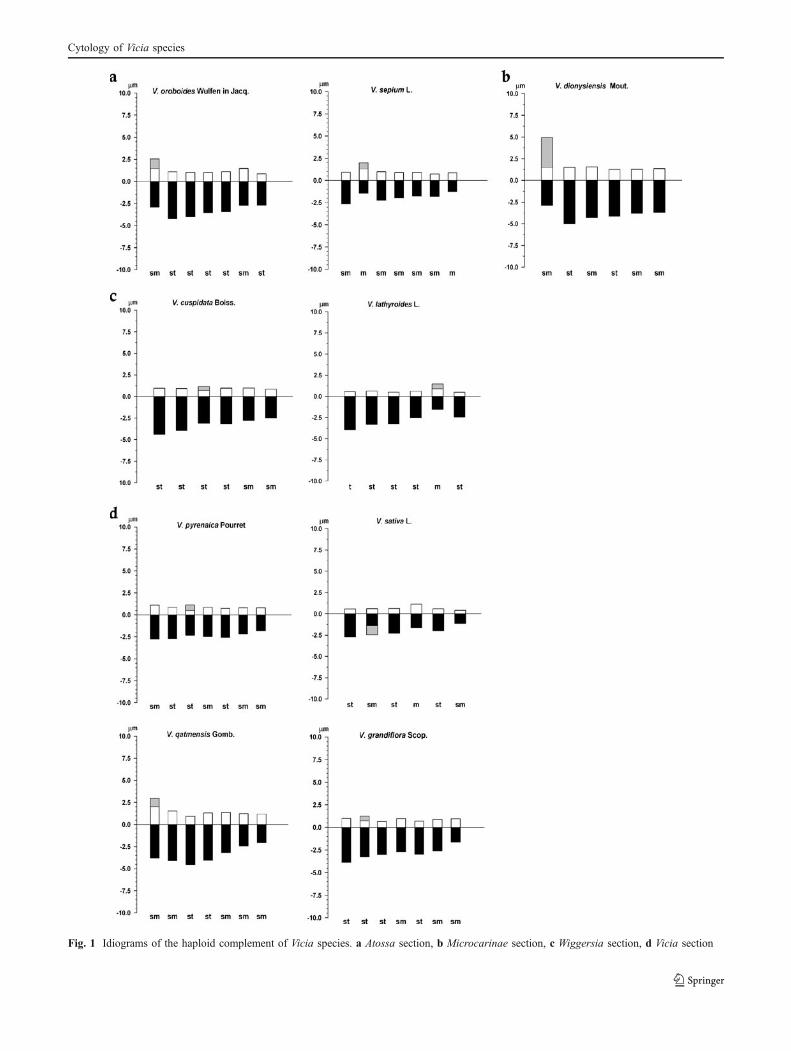
Fig. 1 Idiograms of the haploid complement of Vicia species. a Atossa section, b Microcarinae section, c Wiggersia section, d Vicia section
Cytology of Vicia species
genome size often seems to vary independently to theevolution and diversification of the species within thesections. Indeed genome expanding and downsizing pro-cesses may both contribute to the stability of complementsamong a group of species (Knight and Ackerly 2002) andmay represent an evolutive benefit in Angiosperms (Adamsand Wendel 2005, Bennetzen 2005, Leitch and Bennett2007).
As regards karyotype parameters determination (Tables 3and 4), our method allows an accurate determination of thekaryomorphological indices which can be considered to bedirectly correlated with the evolution of the karyotype asreported for Cicer (Venora et al. 1995), Vigna (Venora et al.1998), and many species of Vicia (Venora et al. 1999 and2000, Kotseruba et al. 2000; Frediani et al. 2005, Caputo etal. 2006, Ruffini Castiglione et al. 2007 and 2009). Theanalysis of the indices of symmetry and the related spatialrepresentation (Table 4 and Fig. 2) also point to the mixingof the species belonging to sections Atossa and Vicia; onlythe two species of section Wiggersia are distinct but closeto each other.
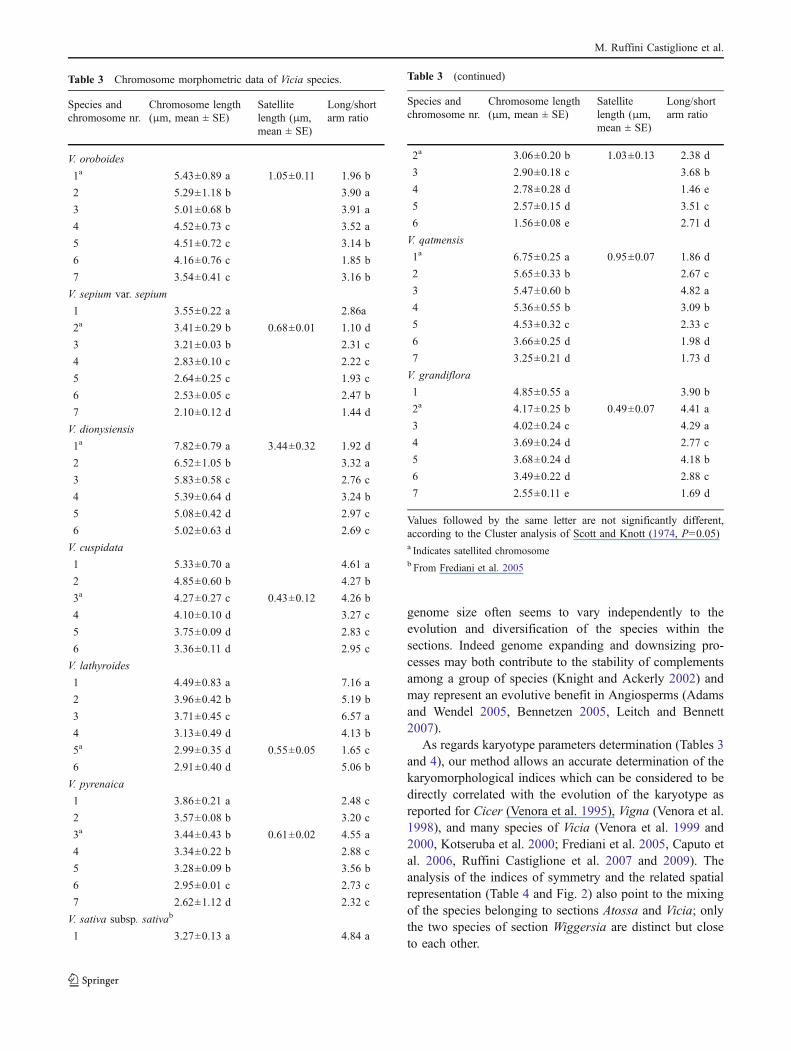
Table 3 Chromosome morphometric data of Vicia species.
Species andchromosome nr.
Chromosome length(μm, mean ± SE)
Satellitelength (μm,mean ± SE)
Long/shortarm ratio
V. oroboides
1a 5.43±0.89 a 1.05±0.11 1.96 b
2 5.29±1.18 b 3.90 a
3 5.01±0.68 b 3.91 a
4 4.52±0.73 c 3.52 a
5 4.51±0.72 c 3.14 b
6 4.16±0.76 c 1.85 b
7 3.54±0.41 c 3.16 b
V. sepium var. sepium
1 3.55±0.22 a 2.86a
2a 3.41±0.29 b 0.68±0.01 1.10 d
3 3.21±0.03 b 2.31 c
4 2.83±0.10 c 2.22 c
5 2.64±0.25 c 1.93 c
6 2.53±0.05 c 2.47 b
7 2.10±0.12 d 1.44 d
V. dionysiensis
1a 7.82±0.79 a 3.44±0.32 1.92 d
2 6.52±1.05 b 3.32 a
3 5.83±0.58 c 2.76 c
4 5.39±0.64 d 3.24 b
5 5.08±0.42 d 2.97 c
6 5.02±0.63 d 2.69 c
V. cuspidata
1 5.33±0.70 a 4.61 a
2 4.85±0.60 b 4.27 b
3a 4.27±0.27 c 0.43±0.12 4.26 b
4 4.10±0.10 d 3.27 c
5 3.75±0.09 d 2.83 c
6 3.36±0.11 d 2.95 c
V. lathyroides
1 4.49±0.83 a 7.16 a
2 3.96±0.42 b 5.19 b
3 3.71±0.45 c 6.57 a
4 3.13±0.49 d 4.13 b
5a 2.99±0.35 d 0.55±0.05 1.65 c
6 2.91±0.40 d 5.06 b
V. pyrenaica
1 3.86±0.21 a 2.48 c
2 3.57±0.08 b 3.20 c
3a 3.44±0.43 b 0.61±0.02 4.55 a
4 3.34±0.22 b 2.88 c
5 3.28±0.09 b 3.56 b
6 2.95±0.01 c 2.73 c
7 2.62±1.12 d 2.32 c
V. sativa subsp. sativab
1 3.27±0.13 a 4.84 a
Table 3 (continued)
Species andchromosome nr.
Chromosome length(μm, mean ± SE)
Satellitelength (μm,mean ± SE)
Long/shortarm ratio
2a 3.06±0.20 b 1.03±0.13 2.38 d
3 2.90±0.18 c 3.68 b
4 2.78±0.28 d 1.46 e
5 2.57±0.15 d 3.51 c
6 1.56±0.08 e 2.71 d
V. qatmensis
1a 6.75±0.25 a 0.95±0.07 1.86 d
2 5.65±0.33 b 2.67 c
3 5.47±0.60 b 4.82 a
4 5.36±0.55 b 3.09 b
5 4.53±0.32 c 2.33 c
6 3.66±0.25 d 1.98 d
7 3.25±0.21 d 1.73 d
V. grandiflora
1 4.85±0.55 a 3.90 b
2a 4.17±0.25 b 0.49±0.07 4.41 a
3 4.02±0.24 c 4.29 a
4 3.69±0.24 d 2.77 c
5 3.68±0.24 d 4.18 b
6 3.49±0.22 d 2.88 c
7 2.55±0.11 e 1.69 d
Values followed by the same letter are not significantly different,according to the Cluster analysis of Scott and Knott (1974, P=0.05)a Indicates satellited chromosomeb From Frediani et al. 2005
M. Ruffini Castiglione et al.
Also the multivariate analysis using 34 karyologicalparameters (Table 5) in addition to the indices TF%, SYiand Rec produces a dendrogram (Fig. 3) which confirmsthe grouping of the species according not to the sections butto the chromosome numbers, as it is possible also toobserve at about linkage distance 10. It is highlighted alsothat V. faba, used as outgroup, is in a clade faraway of allthe studied species.
Molecular investigations on the inclusive group in study,by employing ITS DNA sequences indicate a pattern ofrelationships (Fig. 4) which is divergent from thatsuggested by karyology. However, in this case, only sectionWiggersia is clearly monophyletic and sect. Vicia is
included in a monophylum, which, however, also includesV. sepium, a member of sect. Atossa. Anyway from thesame figure, it is also possible to observe that the species ofsection Vicia are divided in two groups, according toMaxted (1993); indeed, this author splitted up the sectionVicia into two series: series Vicia, which contains Viciabarbazitae, V. pyrenaica and V. sativa (the latter two basallycollapsed in their clade according to our Fig. 4) and seriesGrandiflorae, which contain V. grandiflora and V. qatmen-sis (which are sister species in Fig. 4). The two series aremorphologically distinguished from each other by corollacolour and size, legume shape and size and by the length ofthe hilum relative to the circumference of the seed.
The simultaneous analysis on the ITS data and the 34karyological parameters (Fig. 5) produces an entirelyresolved pattern of relationships in which, as in thepreviously mentioned case, section Wiggersia is monophy-letic. No other monophylum corresponding to establishedsections is apparent in the cladogram; in particular, sections
Fig. 3 Hierarchical cluster analysis. Dendrogram of Vicia species(using linkage between groups) based on the 37 chromosomeparameters listed in Table 5
Fig. 2 Karyotype symmetry of Vicia species. Section Faba, 1 Viciafaba; section Atossa, 2 V. sepium var. sepium, 3 V. oroboides; sectionVicia, 4 V. qatmensis, 5 V. pyrenaica, 6 V. sativa subsp. sativa, 7 V.grandiflora; section Wiggersia, 8 V. cuspidata, 9 V. lathyroides;section Microcarinae, 10 V. dionysiensis
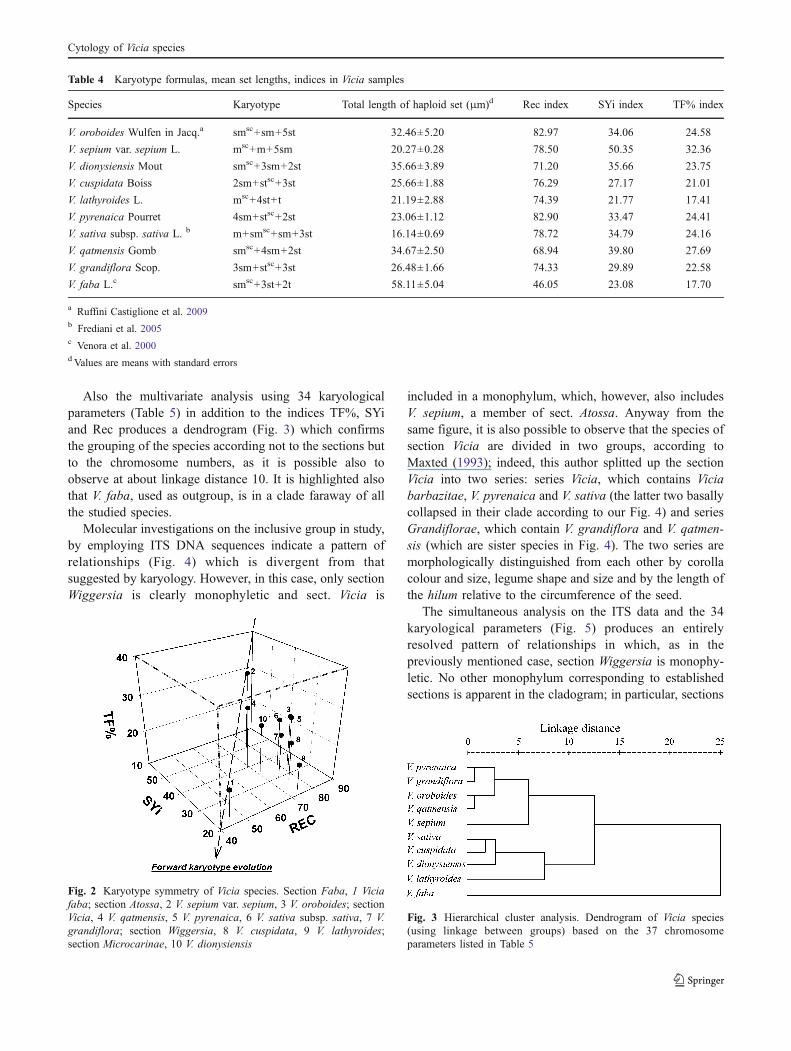
Table 4 Karyotype formulas, mean set lengths, indices in Vicia samples
Species Karyotype Total length of haploid set (μm)d Rec index SYi index TF% index
V. oroboides Wulfen in Jacq.a smsc+sm+5st 32.46±5.20 82.97 34.06 24.58
V. sepium var. sepium L. msc+m+5sm 20.27±0.28 78.50 50.35 32.36
V. dionysiensis Mout smsc+3sm+2st 35.66±3.89 71.20 35.66 23.75
V. cuspidata Boiss 2sm+stsc+3st 25.66±1.88 76.29 27.17 21.01
V. lathyroides L. msc+4st+t 21.19±2.88 74.39 21.77 17.41
V. pyrenaica Pourret 4sm+stsc+2st 23.06±1.12 82.90 33.47 24.41
V. sativa subsp. sativa L. b m+smsc+sm+3st 16.14±0.69 78.72 34.79 24.16
V. qatmensis Gomb smsc+4sm+2st 34.67±2.50 68.94 39.80 27.69
V. grandiflora Scop. 3sm+stsc+3st 26.48±1.66 74.33 29.89 22.58
V. faba L.c smsc+3st+2t 58.11±5.04 46.05 23.08 17.70
a Ruffini Castiglione et al. 2009b Frediani et al. 2005c Venora et al. 2000d Values are means with standard errors
Cytology of Vicia species
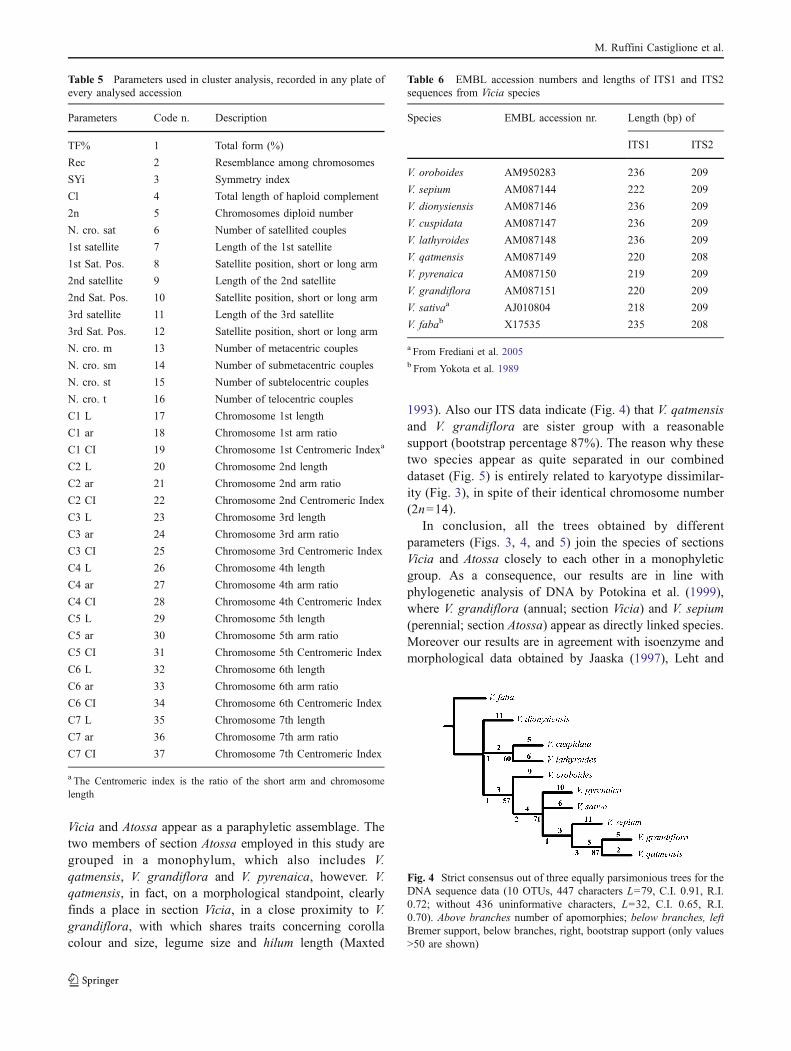
Vicia and Atossa appear as a paraphyletic assemblage. Thetwo members of section Atossa employed in this study aregrouped in a monophylum, which also includes V.qatmensis, V. grandiflora and V. pyrenaica, however. V.qatmensis, in fact, on a morphological standpoint, clearlyfinds a place in section Vicia, in a close proximity to V.grandiflora, with which shares traits concerning corollacolour and size, legume size and hilum length (Maxted
1993). Also our ITS data indicate (Fig. 4) that V. qatmensisand V. grandiflora are sister group with a reasonablesupport (bootstrap percentage 87%). The reason why thesetwo species appear as quite separated in our combineddataset (Fig. 5) is entirely related to karyotype dissimilar-ity (Fig. 3), in spite of their identical chromosome number(2n=14).
In conclusion, all the trees obtained by differentparameters (Figs. 3, 4, and 5) join the species of sectionsVicia and Atossa closely to each other in a monophyleticgroup. As a consequence, our results are in line withphylogenetic analysis of DNA by Potokina et al. (1999),where V. grandiflora (annual; section Vicia) and V. sepium(perennial; section Atossa) appear as directly linked species.Moreover our results are in agreement with isoenzyme andmorphological data obtained by Jaaska (1997), Leht and
Fig. 4 Strict consensus out of three equally parsimonious trees for theDNA sequence data (10 OTUs, 447 characters L=79, C.I. 0.91, R.I.0.72; without 436 uninformative characters, L=32, C.I. 0.65, R.I.0.70). Above branches number of apomorphies; below branches, leftBremer support, below branches, right, bootstrap support (only values>50 are shown)
Table 5 Parameters used in cluster analysis, recorded in any plate ofevery analysed accession
Parameters Code n. Description
TF% 1 Total form (%)
Rec 2 Resemblance among chromosomes
SYi 3 Symmetry index
Cl 4 Total length of haploid complement
2n 5 Chromosomes diploid number
N. cro. sat 6 Number of satellited couples
1st satellite 7 Length of the 1st satellite
1st Sat. Pos. 8 Satellite position, short or long arm
2nd satellite 9 Length of the 2nd satellite
2nd Sat. Pos. 10 Satellite position, short or long arm
3rd satellite 11 Length of the 3rd satellite
3rd Sat. Pos. 12 Satellite position, short or long arm
N. cro. m 13 Number of metacentric couples
N. cro. sm 14 Number of submetacentric couples
N. cro. st 15 Number of subtelocentric couples
N. cro. t 16 Number of telocentric couples
C1 L 17 Chromosome 1st length
C1 ar 18 Chromosome 1st arm ratio
C1 CI 19 Chromosome 1st Centromeric Indexa
C2 L 20 Chromosome 2nd length
C2 ar 21 Chromosome 2nd arm ratio
C2 CI 22 Chromosome 2nd Centromeric Index
C3 L 23 Chromosome 3rd length
C3 ar 24 Chromosome 3rd arm ratio
C3 CI 25 Chromosome 3rd Centromeric Index
C4 L 26 Chromosome 4th length
C4 ar 27 Chromosome 4th arm ratio
C4 CI 28 Chromosome 4th Centromeric Index
C5 L 29 Chromosome 5th length
C5 ar 30 Chromosome 5th arm ratio
C5 CI 31 Chromosome 5th Centromeric Index
C6 L 32 Chromosome 6th length
C6 ar 33 Chromosome 6th arm ratio
C6 CI 34 Chromosome 6th Centromeric Index
C7 L 35 Chromosome 7th length
C7 ar 36 Chromosome 7th arm ratio
C7 CI 37 Chromosome 7th Centromeric Index
a The Centromeric index is the ratio of the short arm and chromosomelength
Table 6 EMBL accession numbers and lengths of ITS1 and ITS2sequences from Vicia species
Species EMBL accession nr. Length (bp) of
ITS1 ITS2
V. oroboides AM950283 236 209
V. sepium AM087144 222 209
V. dionysiensis AM087146 236 209
V. cuspidata AM087147 236 209
V. lathyroides AM087148 236 209
V. qatmensis AM087149 220 208
V. pyrenaica AM087150 219 209
V. grandiflora AM087151 220 209
V. sativaa AJ010804 218 209
V. fabab X17535 235 208
a From Frediani et al. 2005b From Yokota et al. 1989
M. Ruffini Castiglione et al.

Jaaska (2002) and Jaaska and Leht (2007) which demon-strate that Vicia and Atossa sections form a monophyleticgroup, basally related to V. lathyroides, as it also occurs inour analysis where V. cuspidata is present in the same cladewith V. lathyroides. Overall, our results are broadlycongruent with the classification of Maxted (1993); forexample, species of section Wiggersia, which were segre-gated by the author from section Vicia on account of thelarger size of the plant, subsessile legumes and sculpturedseed testa, do form a monophylum which is a possible sistergroup to sections Vicia and Atossa. Very little can be saidon the position of V. dionysiensis, as it belongs to themonospecific section Microcarinae. This Syrian endemic,although displaying the morphological traits which allowplacement in subgenus Vicia (i.e., stipular nectaries), isunique in the subgenus since it has a keel definitely shorterthan wings and standard, a general growth habit similar tothe species of sect. Atossa and a general resemblance tospecies of section Hypechusa (Maxted 1993).
In conclusion, this report on sections Atossa, Vicia,Microcarinae and Wiggersia as well as our previous studieson the other sections of subgenus Vicia and analogousstudies present in the literature, clearly demonstrate that theclassification of the subgenus is still uncertain; probablyonly a simultaneous investigation of all sections ofsubgenus Vicia with other parameters, will give informationfor more sound phylogenetic and taxonomic conclusions.
Conflict of interest The authors declare that they have no conflict ofinterest.
References
Adams KL, Wendel JF (2005) Polyploidy and genome evolution inplants. Curr Op Pl Biol 8:135–141. doi:10.1016/j.pbi.2005.01.001
Ball PW (1968) Vicia L. In: Tutin TG, Heywood VH, Burges NA,Valentine DH, Walters SM, Webb DA (eds) Flora Europaea.Cambridge University Press, Cambridge, pp 129–136
Bennetzen JL (2005) Trasposable elements, gene creation and genomerearrangement in flowering plants. Curr Opin Genet Dev 15:381–386. doi:10.1016/j.gde.2005.09.010
Bremer K (1994) Branch support and tree stability. Cladistics 10:295–304. doi:10.1111/j.1096-0031.1994.tb00179.x
Caputo P, Frediani M, Venora G, Ravalli C, Ambrosio M, CremoniniR (2006) Nuclear DNA contents, rDNAs and karyotypeevolution in subgenus Vicia. III. The heterogeneous sectionHypechusa. Protoplasma 228:167–177. doi:10.1007/s00709-006-0158-2
Ceccarelli M, Minelli S, Maggini F, Cionini PG (1995) Genome sizevariation in Vicia faba. Heredity 74:180–187. doi:10.1038/hdy.1995.26
Chooi WY (1971) Variation in nuclear DNA content in the genusVicia. Genetics 68:195–211
Choi BH, Seok DI, Endo Y, Ohashi H (2006) Phylogeneticsignificance of stylar features in genus Vicia (Leguminosae): ananalysis with molecular phylogeny. J Plant Res 119:513–523.doi:10.1007/s10265-006-0015-6
Cremonini R, Funari S, Mazzuca S (1992) Cytology of Viciaspecies. I. Nuclear structure and DNA content. Chromatin 1:19–26
Endo Y, Choi BH, Ohashi H, Delgado-Salinas A (2008) Phylogeneticrelationships of new world Vicia (Leguminosae) inferred fromnrDNA internal transcribed spacer sequences and floral charac-ters. Syst Bot 33:356–363. doi:10.1600/036364408784571536
Felsenstein J (1985) Confidence limits on phylogenies: an approachusing the bootstrap. Evolution 39:783–791. doi:10.2307/2408678
Fennell SR, Powell W, Wright F, Ramsay G, Wangh R (1998)Phylogenetic relationships between Vicia faba (Fabaceae) andrelated species inferred from chloroplast trnL sequences. PlantSyst Evol 212:247–259. doi:10.1007/BF01089741
Frediani M, Maggini F, Gelati MT, Cremonini R (2004) RepetitiveDNA sequences as probe for phylogenetic analysis in Viciagenus. Caryologia 57:379–388
Frediani M, Caputo P, Venora G, Ravalli C, Ambrosio M, CremoniniR (2005) Nuclear DNA contents, rDNAs, and karyotypeevolution in Vicia subgenus Vicia: II. Section Peregrinae.Protoplasma 226:181–190. doi:10.1007/s00709-005-0114-6
Goloboff P (1999). Nona. Instruction manual. Published by the author,S. M. de Tucumán, Argentina
Goloboff. P, Farris JS, Nixon K. (2003). TNT (Tree analysis usingNew Technology), Version 1.0. Published by the authors,Tucumán, Argentina
Greilhuber J, Speta F (1976) C-banded karyotypes in the Scillahohenacheri group: S. persica and puschkinia (Liliaceae). PlantSyst Evol 126:149–188. doi:10.1007/BF00981669
Hall TA (1999) BioEdit: a user-friendly biological sequencesalignment editor and analysis program for Windows 95/98/NT.Nucl Acid Symp Ser 41:95–98
Huziwara Y (1962) Karyotype analysis in some genera of CompositaeVIII: further studies on the chromosomes of Aster. Amer J Bot49:116–119. doi:10.2307/2439026
ILDIS (1999) International Legume Database and informationService. http://www.ildis.org/
Jaaska V (1997) Isoenzyme diversity and phylogenetic affinities inVicia subgenus Vicia (Fabaceae). Genet Res Crop Evol 44:557–574. doi:10.1023/A:1008630003045
Jaaska V, Leht M (2007) Phylogenetic relationships between andwithin section Hypechusa, Narbonensis and Peregrinae of genusVicia (Fabaceae) based on evidence from isozymes andmorphology. Centr Europ J Biol 2:137–155. doi:10.2478/s11535-007-0008-z
Fig. 5 Single most parsimonious tree for the combined matrixincluding DNA sequence data and 34 karyological parameters(10 OTUs, 481 characters, 34 of which continuous (L=660.66 steps,C.I.=0.74, R.I.=0.73)). Below branches, left Bremer support; belowbranches, right bootstrap support (only values >50 are shown)
Cytology of Vicia species

Knight CA, Ackerly DD (2002) Variation in nuclear DNA contentacross environmental gradients: a quantile regression analysis.Ecol Lett 5:66–76. doi:10.1046/j.1461-0248.2002.00283
Kotseruba V, Venora G, Blangiforti S, Ruffini Castiglione M,Cremonini R (2000) Cytology of Vicia species. IX. NuclearDNA amount, chromatin organization and computer aidedkaryotyping of a Russian accession of Vicia faba L. Caryologia55:195–204
Kupicha FK (1976) The infrageneric structure of Vicia. Not R BotGard Edinb 34:278–326
Leht H, Jaaska V (2002) Cladistic and phenetic analysis of relation-ships in Vicia subgenus Vicia by morphology and isozymes. PlantSyst Evol 232:237–260. doi:10.1007/s006060200045
Leitch IJ, Bennett MD (2007) Genome size and its uses: the impact offlow cytometry. In: Dolozel J, Greilhuber J, Suda J (eds) Flowcytometry with plant cells. Wiley-VCH, Weinheim, pp 153–176
Levan A, Freda K, Sandberg AA (1964) Nomenclature for centro-meric position chromosomes. Hereditas 52:201–220
Maxted N (1993) A phenetic investigation of Vicia L. subgenus Vicia(Leguminosae, Vicieae). Bot J Linn Soc 111:155–182.doi:10.1006/bojl.1993.1013
Maxted N (1995) An ecogeographical study of Vicia subgenus Vicia.Systematic and ecogeographic studies on crop genepools. IPGRI,Rome, Italy, p 8
Maxted N, Callimassia MA, Bennett MD (1991) Cytotaxonomicstudies of Eastern Mediterranean Vicia species (Leguminosae).Plant Syst Evol 177:221–234. doi:10.1007/BF00937959
Nixon KC (1999) Winclada (beta) ver. 0.9.9. Published by the author,Ithaca, New York, USA
Potokina EK, Tomooka N, Vaughan DA, Alexandrova T, Xu RQ(1999) Phylogeny of Vicia subgenus Vicia (Fabaceae) based onanalysis of RAPDs and RFLP of PCR amplified chloroplastgenes. Genet Res Crop Evol 46:149–161. doi:10.1023/A:1008640322198
Raina SN, Bisht MS (1988) DNA amount and chromatin compactnessin Vicia. Genetica 77:65–77. doi:10.1007/BF00058550
Rees H, Cameron FM, Hazarika MH, Jones GH (1966) Nuclearvariation between diploid angiosperms. Nature 211:828–830.doi:10.1038/211828a0
Ruffini Castiglione M, Frediani M, Gelati MT, Ravalli C, Venora G,Caputo P, Cremonini R (2007) Cytological and molecular charac-terization of Vicia esdraelonensis Warb: & Eig: a rare taxon.Protoplasma 231:151–159. doi:10.1007/s00709-007-0256-9
Ruffini Castiglione M, Frediani M, Ravalli C, Venora G, Cremonini R(2009) Cytological characterization of Vicia oroboides Wulfen inJacq. Protoplasma 236:21–27. doi:10.1007/s00709-009-0047-6
Scott AJ, Knott M (1974) A cluster method for grouping means in theanalysis of variance. Biometrics 30:505–512. doi:10.2307/2529204
Stebbins L (1971) Chromosomal evolution in higher plants. EdwardArnold (ed), London, UK
Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTAL W:improving the sensitivity of progressive multiple sequencealignment through sequence weighting, position-specific gappenalties and weight matrix choice. Nucl Acids Res 22:4673–4680. doi:10.1093/nar/22.22.4673
Venora G, Blangiforti S, Cremonini R (1998) Karyotype analysis oftwelve species belonging to genus Vigna. Cytologia 64:117–127
Venora G, Ravalli C, Cremonini R (2008) The karyotype as a tool toidentify plant species. Vicia species belonging to Vicia subgenus.Caryologia 61:300–319
Venora G, Conicella C, Errico A, Saccardo F (1991) Karyotyping inplant by an image analysis system. J Genet Breed 45:233–240
Venora G, Ocampo B, Singh K, Saccardo F (1995) Karyotype of theKabuli-type chickpea (Cicer arietinum L.) by image analysissystem. Caryologia 48:147–155
Venora G, Blangiforti S, Ruffini Castiglione M, Black Samuelsson S,Cremonini R (1999) Cytology of Vicia species. 8. Nuclear DNAcontent, chromatin organization and karyotype evolution in Viciapisiformis L. populations. Caryologia 52:105–115
Venora G, Blangiforti S, Frediani M, Maggini F, Gelati MT, RuffiniCastiglione M, Cremonini R (2000) Nuclear DNA contents,rDNAs, chromatin organization and karyotype evolution in Viciasect. faba. Protoplasma 213:118–125. doi:10.1007/BF01280512
Yokota V, Kawata T, Iida Y, Kato A, Tamifuji S (1989) Nucleotidesequences of the 5.8S rRNA genes and internal transcribedspacer regions in carrot and broad bean ribosomal DNA. J MolEvol 29:294–301. doi:10.1007/BF02103617
M. Ruffini Castiglione et al.