ethanol consumption in the female long–evans rat
TRANSCRIPT

Alcohol 26 (2002) 103–113
0741-8329/02/$ – see front matter © 2002 Elsevier Science Inc. All rights reserved.PII:
S0741-8329(01)00203-8
Ethanol consumption in the female Long–Evans rat:a modulatory role of estradiol
Matthew M. Ford*, J. Charles Eldridge, Herman H. Samson
Center for the Neurobehavioral Study of Alcohol, Department of Physiology & Pharmacology, Wake Forest University School of Medicine, Medical Center Boulevard, Winston-Salem, NC 27157-1083, USA
Received 16 April 2001; received in revised form 2 October 2001; accepted 10 November 2001
Abstract
The examination of
various gonadal hormone manipulations on ethanol intake in human subjects and in rodent models has resulted indisparate findings. In the present study, we examined the effects of ovariectomy and subsequent estradiol (E
2
) replacement on ethanolintake in a within-subject design, as well as assessed the relevance of reproductive status on the efficacy of an E
2
stimulus in eliciting
consumption. Female Long–Evans rats (
n
�
24) were given access to 10% ethanol and water in a continuous-access paradigm. After es-tablishment of baseline intake values, rats were divided into four groups: sham/placebo (Shm
�
P), sham/estradiol (Shm
�
E
2
), ovariecto-mized/placebo (Ovx
�
P), and ovariectomized/estradiol (Ovx
�
E
2
). Rats in the Ovx
�
P group were found to have a large and permanentdecline in ethanol intake that persisted more than 3 months postsurgery. Administration of E
2
to Ovx
�
E
2
rats was associated with resto-ration of ethanol consumption to baseline levels. When Shm
�
E
2
and Ovx
�
E
2
groups were compared, reproductive status was found to be adetermining factor in the efficacy of E
2
to elicit ethanol intake. Together, these findings provide evidence that ovarian hormones, particularlyestradiol, exert activational effects on estrogen-responsive substrates to modulate ethanol consumption in the adult female rat. © 2002Elsevier Science Inc. All rights reserved.
Keywords:
Ethanol self-administration; Continuous access; Female rat; Ovariectomy; Estradiol replacement
1. Introduction
The majority of studies with human subjects that haveaddressed ethanol–gonadal hormone interactions have fo-cused primarily on the effects of moderate ethanol con-sumption on gonadal hormone levels and hypothalamic-pituitary-gonadal (HPG) axis integrity. Ovarian hormonemodulation of ethanol consumption and the effects of thesehormones on the pharmacological and behavioral sensitivityto ethanol, on the other hand, have remained understudied.Methodological constraints have made it difficult to assessethanol consumption patterns across the menstrual cycle inhuman beings (Gill, 1997). Individual variation in the hor-monal profiles exhibited in women across the menstrual cy-cle necessitates the use of a within-subject design, whichfew studies have incorporated (Gill, 1997). In one recentstudy, a within-subject design was implemented. However,there was no detectable difference in ethanol consumptionacross the menstrual cycle (Holdstock & de Wit, 2000). Theinvestigators advocated that several limitations hindered
their results; that is, variable drinking histories of the womenstudied, insufficient sample size, and the absence of a test phasewhen estradiol levels were elevated (Holdstock & de Wit,2000).
Animal models may offer a means for arriving at a pre-cise understanding of ovarian hormone action on ethanol in-take by avoiding many of the pitfalls inherent to studieswith human subjects. The female rat provides a usefulmodel of ethanol self-administration because it exhibitsboth spontaneous and cyclic ovulation (Yen et al., 1999),possesses cyclic hormone profiles that resemble those of thehuman menstrual cycle (with the exception of a truncatedluteal phase), and is thought to have similar neural circuitrymediating the neurobiological effects of ethanol (Koob,1992; Koob et al., 1998). In addition, rats can be investigatedwithin an experimenter-controlled environment, thereby en-hancing the probability that the underlying effects of an en-dogenous hormonal state on ethanol consumption are delin-eated (Roberts et al., 1998).
Sex differences in ethanol consumption have been ob-served in several rat strains, with females typically consum-ing more than males (Juárez & Barrios de Tomasi, 1999;Lancaster & Spiegel, 1992; Li & Lumeng, 1984). It is be-
* Corresponding author. Tel.:
�
1-336-716-8595; fax:
�
1-336-716-8501.
E-mail address:
[email protected] (M.M. Ford).Editor: T. R. Jerrells

104
M.M. Ford et al. / Alcohol 26 (2002) 103–113
lieved that these gender differences in consumption can beattributed, at least in part, to an assembly of sexually dimor-phic neural substrates whose sensitivity to ethanol varies inrelation to cyclic ovarian hormone levels (Almeida et al.,1998; Kelly et al., 1999; Lancaster, 1994; Lancaster et al.,1996). Lancaster et al. (1996) have documented an increasein ethanol intake throughout a peripubertal transition periodthat encompasses the establishment of estrous cycles. Thereports of an enhancement of operant responding for etha-nol during diestrus day (Roberts et al., 1998) and an attenu-ation of ethanol intake on proestrus day (Forger & Morin,1982) are additional pieces of evidence that indicate thatovarian hormone environment and the estrous cycle may beimportant determinants of ethanol consumption in the fe-male rat.
Several studies have been conducted to assess the effectsof ovariectomy (ovex) and estradiol replacement on ethanolconsumption in rodents. Hilakivi-Clarke (1996) reportedthat outbred CD-1 mice that were 30 days postovex showeda small decrease in intake of a 5% ethanol solution in com-parison with findings for sham controls. In this same study,60 days of 17
�
-estradiol pellet treatment in ovariectomizedmice did not alter ethanol intake from levels observed inovariectomized controls (Hilakivi-Clarke, 1996). In a sepa-rate study, ovex (1–2 weeks postsurgery) in Wistar rats wasnot associated with a change in intake of a 12% ethanol so-lution, whereas ovariectomized rats receiving daily injec-tions of 17
�
-estradiol had significantly decreased ethanolintakes when compared with findings for rats with intactovaries (Almeida et al., 1998). A large dose (300
�
g) ofethinyl estradiol was shown to be associated with sup-pressed intake of a 5% ethanol solution in intact femaleSprague–Dawley rats (Messiha, 1981). Similarly, results ofadditional studies have shown that administration of largedoses (5 or 100
�
g) of estradiol benzoate led to an attenua-tion of ethanol intake in ovariectomized rats (Aschkenasy-Lelu, 1960; Sandberg et al., 1982). Forger and Morin (1982)observed that ovariectomized Sprague–Dawley rats hadgreatly attenuated intake and preference for 4% ethanol. Incontrast, Cailhol and Mormède (2001) concluded that ovexfailed to alter the maintenance of ethanol self-administra-tion in the three rat strains studied.
The disparity of findings among studies of ovex and es-tradiol replacement may be due, in part, to the species andstrain of rodent selected, to the dose and route of hormoneadministration chosen, to the reproductive status (intactovaries, days postovex, or weeks to months postovex) of thetest animals studied, or to the concentration of ethanol solu-tion presented. Importantly, none of the aforementionedstudies determined a baseline ethanol intake before the as-signment of animals to experimental groups, introducing thepossibility that intake variability was not evenly distributedamong groups. Collection of an intake baseline would alsohave supported a within-subject analysis of the experimen-tal manipulations. The purpose of the present study was toexamine the effects of various gonadal hormone manipula-
tions on ethanol intake by female rats in a within-subject de-sign, to delineate further the modulatory role that ovariansteroids play in ethanol consumption. Furthermore, estra-diol replacement regimens were administered to rats withintact ovaries as well as to ovariectomized rats to determinewhether reproductive (hormonal) status is associated withan altered efficacy of estradiol stimulus on ethanol con-sumption.
2. Materials and methods
2.1. Animals
Twenty-four female, Long–Evans rats, each weighingbetween 190 and 220 g, were acquired from Harlan SpragueDawley, Inc. (Indianapolis, IN). The rats were housed sin-gly in hanging wire cages with artificial lights on between0500 and 1900 and given ad libitum access to rat chow andwater (except where noted). The rats were acclimated tohome cages and handled daily for at least 6 days before thestart of the experiment. The Animal Care and Use Commit-tee of Wake Forest University School of Medicine approvedall procedures in accordance with the guidelines of the Insti-tutional Care and Use Committee of the National Instituteon Drug Abuse, National Institutes of Health, and the
Guidefor the Care and Use of Laboratory Animals
(Institute ofLaboratory Animal Resources, Commission on Life Sci-ences, National Research Council, 1996).
2.2. Experimental design
Before the initial preference test, 10% ethanol (10E) waspresented as the only fluid for 3 days. Experimental phase Iinvolved the presentation of 10E and tap water in a two-bot-tle-choice, continuous-access paradigm for 14 days, as pre-viously described (Tolliver et al., 1988). Bottle positionswere alternated daily between left and right sides to mini-mize the effects of side preference on ethanol consumption.All bottles were removed daily at 0830 and replaced at1030. During this time-out period, fluid volumes were mea-sured and the rats were weighed. Water and 10E were re-plenished every other day. At the conclusion of the prefer-ence test, rats were assigned to either a sham-operated or anovariectomized group (
n
�
12 for each) on the basis of theirmean baseline ethanol intakes (grams per kilogram of bodyweight per day). Intake baseline values were ranked fromlargest (#1) to smallest (#24). Odd-ranked drinkers were as-signed to the ovariectomized pool, and even-ranked drink-ers were designated to the sham pool. After surgery, all ratswere allowed a 3-day recovery period without access to 10Eto prevent any potential interaction between ethanol and thesurgical anesthetic used.
Experimental phase II included a 24-day evaluation ofpostsurgical ethanol intake with access to both 10E and wa-ter. After this assessment, sham-operated and ovariecto-mized rats were divided further into subgroups (
n
�
6) onthe basis of their postsurgical ethanol intakes. A similar

M.M. Ford et al. / Alcohol 26 (2002) 103–113
105
ranking procedure was implemented as described above tocreate the following experimental groups: sham/placebo(Shm
�
P), sham/estradiol (Shm
�
E
2
), ovariectomized/pla-cebo (Ovx
�
P), and ovariectomized/estradiol (Ovx
�
E
2
). Atthe beginning of phase III, each rat received either a placeboor a 17
�
-estradiol pellet implant (0.25 mg; 21-day release).All rats were allowed a 2-day recovery period in the ab-sence of 10E before being reevaluated in a two-bottle-choice paradigm for an additional 64 days. Phase IV of theexperiment encompassed the period that elapsed during andafter the washout of implant-supported estradiol activity.
2.3. Surgery: ovariectomy and implants
The rats were anesthetized with halothane (4.0%) andmaintained on this inhalant (2.0%) throughout surgery (20–25min) by means of a fitted nose cone. All instruments were au-toclaved before use, and the procedure was conducted in asterile environment. Bilateral (1.5-cm) incisions were madein the flanks. Ovaries were excised after ligation at the tip ofthe uterine horn. Sham operations were conducted underidentical conditions, but without ligature application or ovaryremoval. The presence or absence of ovaries was confirmedduring autopsy. The subcutaneous implantation of pellets atthe nape of the neck was conducted under similar anestheticconditions used for ovariectomy.
2.4. Drugs
Ethanol (95% [vol./vol.]) was diluted in tap water to aconcentration of 10% (vol./vol.). Placebo and 17
�
-estradiolpellets (0.25 mg; 21-day release) were purchased from In-novative Research of America (Sarasota, FL). The pelletwas selected for its zero-order-release kinetics and ability tomaintain 17
�
-estradiol plasma concentrations in the mid- toupper-physiological range (30–40 pg/ml). Halothane waspurchased from Halocarbon Laboratories (River Edge, NJ).
2.5. Data analysis and presentation
Phase I baseline measures are reported as 8-day means
�
standard error of the mean (S.E.M.) (representative of thelast 8 days of preference testing before surgery). The base-line body weights and all parameters during phases II–IVare presented as 4-day means
�
S.E.M. These 4-day periodsare referred to as intervals in the subsequent text. The re-sults are depicted in figures as four experimental groupsthroughout all phases, despite that these groupings were notempirically determined until the end of phase II. This wasdone to facilitate the analysis of across-phase effects foreach group.
All experimental parameters were analyzed by using theGB-STAT version 6.5 software package (Dynamic Micro-systems, Inc., Silver Spring, MD). A two-way repeatedmeasure analysis of variance (ANOVA) (factors day andgroup) was carried out to assess differences in baseline val-
ues among the experimental groups. A two-way repeatedmeasure ANOVA (factors interval and surgery) was used todetect significant differences among groups during phase II.Similarly, differences among groups throughout phases IIIand IV were analyzed by three-way repeated measure ANOVA(factors interval, surgery, and pellet). Within-group analysesacross phases were examined by two-way repeated measureANOVA (factors interval and group). The Tukey multiple com-parisons procedure was implemented to determine pair-wisedifferences if a significant main effect occurred. In all cases, in-tervals 1 and 7 (corresponding to the 4-day periods immedi-ately after surgery and pellet implantation) were excludedfrom statistical analyses to isolate the gonadal hormone ef-fects from the deprivation-like effect that was apparent atthese intervals.
3. Results
3.1. Phase I: baseline
The baseline ethanol intake (Fig. 1) and the baseline eth-anol preference ratio (Fig. 2) for all rats averaged 4.86
�
0.39 g/kg/day (range, 1.61–8.88 g/kg/day) and 0.47
�
0.04(range, 0.16–0.79), respectively. There was a statisticallysignificant main effect of day for both baseline intakes[
F
(7,140)
�
2.54,
P
�
.05] and baseline preference ratios[
F
(7,140)
�
2.88,
P
�
.01]. The baseline water intake (Fig.3) and the baseline body weight (Fig. 4) for all rats averaged15.73
�
1.11 ml/day (range, 5.88–24.67 ml/day) and 235
�
2 g (range, 214–254 g), respectively. A statistically signifi-cant main effect of day was found for water intake baselines[
F
(7,140)
�
3.62,
P
�
.01] and body weight baselines[
F
(3,60)
�
12.19,
P
�
.001]. There was no statistically sig-nificant group effect detected for any parameter evaluated.
3.2. Phase II: surgery (ovex vs. sham)
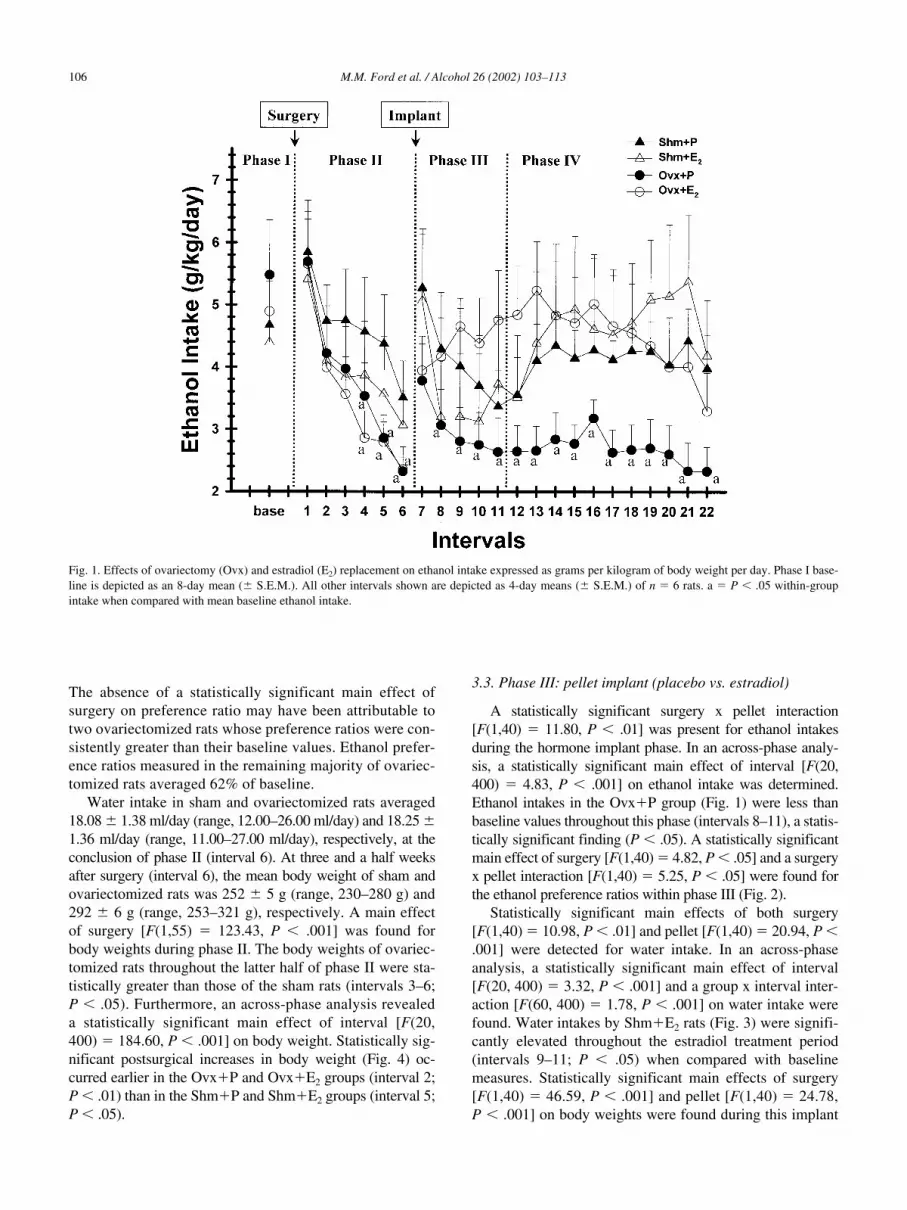
Three and a half weeks after surgery (interval 6), themean ethanol intake in sham rats (Shm
�
P and Shm
�
E
2
groups) was 3.28
�
0.39 g/kg/day (range, 1.11–6.28 g/kg/day). At the same postsurgical period, ovariectomized rats(Ovx
�
P and Ovx
�
E
2
groups) exhibited a mean ethanolintake of 2.34
�
0.20 g/kg/day (range, 1.42–3.74 g/kg/day). A statistically significant main effect of surgery[
F
(1,55)
�
11.28,
P
�
.01] was found for ethanol intakesduring this phase. In an across-phase analysis, a statisti-cally significant main effect of interval [
F
(20, 400)
�
4.83,
P
�
.001] was also determined. The Ovx
�
P andOvx
�
E
2
groups had significantly attenuated ethanol in-takes as early as 2 weeks postovex (intervals 4–6;
P
�
.05)when compared with findings for their respective baselineintakes (Fig. 1). By three and a half weeks after surgery(interval 6) the sham and ovariectomized rats had meanethanol preference ratios of 0.37
�
0.04 (range, 0.12–0.65) and 0.33
�
0.03 (range, 0.16–0.56), respectively.
106
M.M. Ford et al. / Alcohol 26 (2002) 103–113
The absence of a statistically significant main effect ofsurgery on preference ratio may have been attributable totwo ovariectomized rats whose preference ratios were con-sistently greater than their baseline values. Ethanol prefer-ence ratios measured in the remaining majority of ovariec-tomized rats averaged 62% of baseline.
Water intake in sham and ovariectomized rats averaged18.08
�
1.38 ml/day (range, 12.00–26.00 ml/day) and 18.25
�
1.36 ml/day (range, 11.00–27.00 ml/day), respectively, at theconclusion of phase II (interval 6). At three and a half weeksafter surgery (interval 6), the mean body weight of sham andovariectomized rats was 252
�
5 g (range, 230–280 g) and292
�
6 g (range, 253–321 g), respectively. A main effectof surgery [
F
(1,55)
�
123.43,
P
�
.001] was found forbody weights during phase II. The body weights of ovariec-tomized rats throughout the latter half of phase II were sta-tistically greater than those of the sham rats (intervals 3–6;
P
�
.05). Furthermore, an across-phase analysis revealeda statistically significant main effect of interval [
F
(20,400)
�
184.60,
P
�
.001] on body weight. Statistically sig-nificant postsurgical increases in body weight (Fig. 4) oc-curred earlier in the Ovx
�
P and Ovx
�
E
2
groups (interval 2;
P
�
.01) than in the Shm
�
P and Shm
�
E
2
groups (interval 5;
P
�
.05).
3.3. Phase III: pellet implant (placebo vs. estradiol)
A statistically significant surgery x pellet interaction[
F
(1,40)
� 11.80, P � .01] was present for ethanol intakesduring the hormone implant phase. In an across-phase analy-sis, a statistically significant main effect of interval [F(20,400) � 4.83, P � .001] on ethanol intake was determined.Ethanol intakes in the Ovx�P group (Fig. 1) were less thanbaseline values throughout this phase (intervals 8–11), a statis-tically significant finding (P � .05). A statistically significantmain effect of surgery [F(1,40) � 4.82, P � .05] and a surgeryx pellet interaction [F(1,40) � 5.25, P � .05] were found forthe ethanol preference ratios within phase III (Fig. 2).
Statistically significant main effects of both surgery[F(1,40) � 10.98, P � .01] and pellet [F(1,40) � 20.94, P �.001] were detected for water intake. In an across-phaseanalysis, a statistically significant main effect of interval[F(20, 400) � 3.32, P � .001] and a group x interval inter-action [F(60, 400) � 1.78, P � .001] on water intake werefound. Water intakes by Shm�E2 rats (Fig. 3) were signifi-cantly elevated throughout the estradiol treatment period(intervals 9–11; P � .05) when compared with baselinemeasures. Statistically significant main effects of surgery[F(1,40) � 46.59, P � .001] and pellet [F(1,40) � 24.78,P � .001] on body weights were found during this implant
Fig. 1. Effects of ovariectomy (Ovx) and estradiol (E2) replacement on ethanol intake expressed as grams per kilogram of body weight per day. Phase I base-line is depicted as an 8-day mean (� S.E.M.). All other intervals shown are depicted as 4-day means (� S.E.M.) of n � 6 rats. a � P � .05 within-groupintake when compared with mean baseline ethanol intake.

M.M. Ford et al. / Alcohol 26 (2002) 103–113 107
phase. In addition, a surgery x pellet interaction [F(1,40) �6.12, P � .05] was detected for body weight. Estradiol re-placement was associated with reduced body weights in theOvx�E2 group (Fig. 4).
3.4. Phase IV: postimplant
Statistically significant main effects of surgery [F(1,110) � 13.63, P � .001] and pellet [F(1, 110) � 27.25, P �.001] were found for ethanol intakes during phase IV. A sta-tistically significant surgery x pellet interaction [F(1, 110) �8.37, P � .01] also occurred for this measure. In an across-phase analysis, a statistically significant main effect of in-terval [F(20, 400) � 4.83, P � .001] on ethanol intake wasdetermined. Ethanol intakes by the Ovx�P group (Fig. 1)were significantly attenuated throughout phase IV (intervals12–22; P � .05) when compared with baseline intakes, indi-cating that ovex was associated with a permanent decline inethanol intake. A surgery x pellet interaction [F(1, 110) �4.69, P � .05] was determined for ethanol preference ratioswithin this hormone washout phase. Ethanol preference ra-tios for the Shm�E2 group (Fig. 2) gradually recovered tobaseline levels during the first 2 weeks of the hormonewashout phase.
Statistically significant main effects of surgery [F(1,110) �4.85, P � .05] and pellet [F(1,110) � 26.29, P � .001]were found for water intakes during phase IV. An across-
phase analysis revealed a statistically significant main effectof interval [F(20, 400) � 3.32, P � .001] for water intake.In comparison with baseline measures, water intakes inShm�E2 rats (Fig. 3) were significantly elevated at the be-ginning of the hormone washout period (intervals 12–13;P � .05). Statistically significant main effects of interval[F(10,110) � 2.77, P � .01], surgery [F(1,110) � 137.85,P � .001], and pellet [F(1,110) � 51.47, P � .001] wereobserved for body weights during phase IV. A statisticallysignificant surgery x pellet interaction [F(1,110) � 7.14,P � .01] was also found for this measure.
3.5. Patterns of ethanol intake
Although identifiable estrous cycle-like (4- to 5-day) pat-terns of ethanol intake were not consistently observed inrats with ovaries intact, occasional periods of cyclic drink-ing were exhibited: throughout phase II for the Shm�E2 ratwhose drinking profile is depicted in Fig. 5B and through-out phase I for the Ovx�P rat whose drinking profile isshown in Fig. 5C. Ovex (as early as 2 weeks postsurgery)was associated with decreases in daily intake variabilitywhen daily ethanol intakes were compared with baselineprofiles. This phenomenon was apparent in the Ovx�P ratwhose drinking profile is represented in Fig. 5C when com-paring intake during phases II through IV to baseline pat-terns and in the Ovx�E2 rat whose drinking profile is de-
Fig. 2. Effects of ovariectomy (Ovx) and estradiol (E2) replacement on ethanol preference ratio determined as the 10% ethanol (10E) volume divided by thetotal volume of 10E and water. Phase I baseline is depicted as an 8-day mean (� S.E.M.). All other intervals shown are depicted as 4-day means (� S.E.M.)of n � 6 rats.

108 M.M. Ford et al. / Alcohol 26 (2002) 103–113
picted in Fig. 5D when comparing intake during phase II tobaseline profiles. In contrast, the Shm�P rats exhibitedconsistent variability in ethanol intakes across all experi-mental phases (as exemplified in Fig. 5A).
4. Discussion
In the current study, ovex was associated with a statisti-cally significant decline in ethanol intake by 3 weeks aftersurgery, and this effect was found to be permanent in ratsthat remained in a hormone-deprived state. This findingcontradicts results of some previous reports (Almeida et al.,1998; Cailhol & Mormède, 2001) in which no change inethanol intake was observed after ovex. The differences inexperimental approach among studies may help explain thedisparity of findings. The collection of intake baselines be-fore ovex in our study supported a within-subject design,whereas in previous studies these measures were not ob-tained. In most cases, we observed that rats with the largestbaseline ethanol intakes exhibited the greatest reductions inintake after ovex. The converse was true in rats with lowbaseline intakes. This apparently graded-consumption re-sponse to ovex within our rat cohort underscores the impor-tance of properly balancing ethanol intakes among experi-mental groups before surgical manipulation. It is possiblethat the experimental groups compared by a between-sub-
ject design in previous studies (Almeida et al., 1998; Cail-hol & Mormède, 2001) were insufficiently balanced to de-tect an ovex effect. Also, in the present study the quality ofthe ethanol stimulus (10% [vol./vol.] solution) remainedconstant throughout acquisition and maintenance periods ofexposure. In both the study by Cailhol and Mormède andthe study by Almeida and colleagues, the acquisition of eth-anol consumption was conducted while the concentration ofthe ethanol solution was increased in a step-wise fashion.The absence of stable conditions throughout this acquisitionprocedure may have similarly confounded the detection ofan ovex effect on ethanol consumption.
Ovariectomy was not associated with statistically sig-nificant alterations in water intake within the Ovx�P andOvx�E groups across phase II. Furthermore, the consis-tent discrepancy in water intake between Ovx�P andShm�P groups during phases III–IV mirrored initial base-line differences (Fig. 3), indicating that the lack of statis-tically significant changes in water intake held true forovariectomized rats more than 3 months postsurgery. Con-sistent with findings in previous reports (Mohamed &Abdel-Rahman, 2000; Tarttelin & Gorski, 1973), ovex ledto significant body weight gain over sham control values.This change in body weight, however, did not influence thepostovex decline in the milliliters of ethanol consumed(data not shown).
Fig. 3. Effects of ovariectomy (Ovx) and estradiol (E2) replacement on water intake expressed as milliliters per day. Phase I baseline is depicted as an 8-daymean (� S.E.M.). All other intervals shown are depicted as 4-day means (� S.E.M.) of n � 6 rats. a � P � .05 within-group intake when compared withmean baseline water intake.

M.M. Ford et al. / Alcohol 26 (2002) 103–113 109
During E2 replacement to the Ovx�E2 group, ethanol in-takes were restored to presurgical baseline levels (Fig. 1;phases III–IV). In contrast, results of other studies in ratshave shown decreases in ethanol intake of ovariectomizedrats administered either 5 �g of E2 for 17 days (Almeida etal., 1998) or 5 �g of estrogen benzoate for 20 days (Sand-berg et al., 1982). However, the findings of these two stud-ies were somewhat ambiguous. In the study by Almeida andcolleagues, E2 treatment began before ethanol presentation,making it necessary to compare the consumption by theovariectomized group to that of controls with ovaries intactwhile the ethanol solution concentration was incrementallymanipulated. Although Sandberg and collaborators admin-istered hormone to ovariectomized rats that had severalmonths of drinking experience, the treatment was associatedwith an initial suppression of ethanol intake. However, in-take that followed repeated administration of estrogen ben-zoate was shown to return to levels observed in untreatedovariectomized rats. The disparity of results among theseearlier reports and the current study could also be due to theroute of administration or to the estrogen dose selected. Inour laboratory, we have found that daily E2 injections of 5�g/kg support supraphysiological plasma concentrations ofthis hormone, as well as that a distinctive rise and subse-quent decline in E2 concentrations are exhibited after ad-ministration (unpublished observations, M. M. Ford, J. C.Eldridge, & H. H. Samson, 2001). In the current study, pel-
let implants were used to provide stable plasma E2 concen-trations within a physiological range.
The efficacy of E2 administration on ethanol consump-tion was decidedly dependent on the reproductive status ofthe rat (Fig. 1; phase III). A hormone stimulus that had littleto no effect on Shm�E2 rat ethanol intakes was shown con-vincingly to be associated with restoration of ethanol con-sumption of Ovx�E2 rats to their baseline intake levels.This finding would support the suggestion that estrogen-responsive substrates facilitating this behavior were sensi-tized owing to hormone deprivation. Dependency on repro-ductive status in the determination of the responsiveness toan E2 stimulus has been demonstrated previously in the con-trol of luteinizing hormone release and in the activation oflordosis behavior (McGinnis et al., 1981). McGinnis andcolleagues purport that the magnitude and duration of alter-ations in nuclear estrogen receptors (ERs) determine thephysiological or behavioral efficacy of an estrogen stimu-lus. Augmentation of ER mRNA levels has been demon-strated within several hypothalamic brain regions of rats af-ter ovex (Shughrue et al., 1992). It is possible that thesensitization of Ovx�E2 rats to the E2-invoked ethanol con-sumption may have been attributable to an elevated nuclearER state within the neuronal circuitry mediating this behav-ior. In addition, the enactment of an autoregulatory processby which E2 up-regulates or down-regulates ER levels in aconcentration-dependent manner (Mohamed & Abdel-Rah-
Fig. 4. Effects of ovariectomy (Ovx) and estradiol (E2) replacement on body weight expressed as grams. Phase I baseline and all other intervals are depicted as4-day means (� S.E.M.) of n � 6 rats. a � P � .05, initially occurring significant within-group comparison when compared with mean baseline ethanolintake; b � P � .05 between-group comparison when ovariectomized rats were compared with sham rats.

110 M.M. Ford et al. / Alcohol 26 (2002) 103–113
man, 2000) may help to explain the underlying ramifica-tions of the prevailing endogenous hormone environmenton the sensitivity of this ethanol consumption response.
Estradiol may alter ethanol consumption through severalmechanisms of action. Estradiol has been shown to exertimmediate (seconds to minutes) effects by altering neuronalexcitability, modulating cAMP signal transduction cascades,and interacting with G-protein-coupled receptors (McEwen& Alves, 1999). Prolonged (hours to days) effects of E2,through a classic genomic mechanism, may result in tran-script expression changes that are relevant to ethanol con-sumption. Alterations in taste reactivity (Wade & Zucker,1970), neurotransmitters and their respective receptors (Lan-caster et al., 1996; McEwen & Alves, 1999), ingestive behav-ior regulation (Kalra et al., 1999), and energy metabolism
(Wade et al., 1985) may be some of the potential mecha-nisms by which E2 exerts its modulatory effects on ethanolconsumption.
The elevated ethanol intakes rapidly achieved after E2
implants to the Ovx�E2 group persisted throughout the firsthalf of the hormone washout period (Fig. 1; phase IV). Thepersistence of ethanol consumption may have resulted fromthe accumulation of E2-inducible gene products within thetarget neurons that control this behavior (McGinnis et al.,1981). In the absence of further hormonal support, these E2-inducible products would degrade over time, and only whendeclining below a set threshold would a correspondingchange in the affected behavior be apparent. Similarly, thistype of delayed response to the discontinuation of hormonalsupport may help explain why sizeable decreases in ethanol
Fig. 5. Effects of ovariectomy (Ovx) and estradiol (E2) replacement on daily consumption patterns of ethanol expressed as grams per kilogram of body weightper day. The drinking profiles depicted are the daily intake patterns of individual rats that were representative of their respective experimental group: sham/placebo (Shm�P) (panel A; rat #1-13), sham/estradiol (Shm�E2) (panel B; rat #1-17), ovariectomized/placebo (Ovx�P) (panel C; rat #1-20), and ovariecto-mized/estradiol (Ovx�E2) (panel D; rat #1-1).

M.M. Ford et al. / Alcohol 26 (2002) 103–113 111
consumption were not found until more than 2 weeks post-ovex.
Interestingly, the finding that E2 elicited augmented wa-ter intakes in both Shm�E2 and Ovx�E2 groups (Fig. 3)seems to indicate that this hormone perhaps acts throughdistinct mechanisms in its modulation of water as opposedto ethanol intake. Mohamed and Abdel-Rahman (2000)have demonstrated that autoregulation of ER by E2 is regionand tissue specific. This finding is substantiated by the ob-servation that mRNA expression of ER� in rats 12 weekspostovex is up-regulated in some tissues and remains un-changed in others, as well as that E2 treatment is associatedwith the restoration of ER� mRNA levels to sham rat (intactovaries) levels in only those regions previously demonstrat-ing an up-regulation. It is possible that the substrates regu-lating water consumption possess ER populations that arenot subject to the same degree of autoregulatory control asthose affiliated with ethanol intake modulation. The differ-ential distribution and regulation of ER� and ER� isoforms,which likely alter brain function and behavior in distinctmanners (Österlund et al., 1998), may also help explain thisdichotomy. In addition, water turnover has been shown toincrease in female rats, both ovariectomized rats and thosewith ovaries intact, treated 14 weeks or longer with estradiolbenzoate (Carlberg et al., 1984). It is important to note thatthe onset of increased water intake in the Ovx�E2 rats wasdelayed when compared with that of the Shm�E2 group.This difference may have been due to the correspondingethanol consumption profiles exhibited by each group, espe-cially when taking into consideration that ethanol consump-tion induces diuresis (Eisenhofer & Johnson, 1982).
In this study, female rats with ovaries intact consumedtwice as much ethanol as that of several cohorts of male rats(4.71 � 0.27 vs. 2.24 � 0.18 g/kg/day, respectively) previ-ously examined in this laboratory under similar experimen-tal conditions (Files et al., 1994; Samson et al., 1991, 1992).This observation is in agreement with results of earlier stud-ies that have shown similar gender differences across nu-merous rat strains (Juárez & Barrios de Tomasi, 1999; Lan-caster & Spiegel, 1992; Li & Lumeng, 1984). The ethanolintake of female rats 3 months postovex (2.33 � 0.39 g/kg/day; interval 22) closely matched historical male intake val-ues. Lancaster and Spiegel (1992) have reported gender dif-ferences in the daily patterns of beer (10% ethanol) intake inLong–Evans rats, with female rats exhibiting larger andmore variable intakes compared with those of male rats.Five of six Ovx�P rats in this experiment expressed de-creased amounts of variability in daily ethanol intakes asearly as 2 weeks postovex (refer to Fig. 5C). Thus, in thecurrent study, ovex was associated with a reduction in bothethanol intake and daily intake variability in female rats.These findings are consistent with results of earlier studies(Findlay et al., 1979; Tarttelin & Gorski, 1973), document-ing a reduction of variability in fluid drinking patterns afterovex. This altered consumption pattern arguably resemblesmore closely the reported drinking patterns of male rats
(Lancaster & Spiegel, 1992) than the patterns of female ratswith ovaries intact, supporting the suggestion that reproduc-tive status and fluctuations in ovarian hormones may con-tribute greatly to gender differences in ethanol consumptionby the Long–Evans strain.
Ethanol and water intakes were expressed as 4-daymeans � S.E.M. to reduce the potential effects of estrouscycle on the variability in these measures. Although it hasbeen documented extensively that female rats normally ex-hibit estrous cycles of 3–5 days in length, the 4-day cycle isconsidered the most prevalent variant (Everett, 1989). Thisprecaution was adopted in consideration of results of a pre-vious study in which a cyclic regulation of ethanol con-sumption across the estrous cycle was observed, with intakeand preference ratio being lowest on proestrus (Forger &Morin, 1982). In the current study, consistent cyclical drink-ing patterns were not observed in female rats with ovariesintact (Fig. 5A–B). The lack of such patterns is in agree-ment with results of a recent study that showed no change inethanol consumption across various menstrual cycle phasesin human subjects (Holdstock & de Wit, 2000). It is possi-ble that the detection of discernable differences in intakeacross the estrous cycle is reliant on the concentration ofethanol used, as well as that the 10% solution used in thecurrent study was too aversive (and consumed as too smallof a volume). When a 4% ethanol solution was used, whichwas found to be optimally preferred across a wide range ofconcentrations, estrous cycle patterns were observed (Forger& Morin, 1982). Furthermore, some rats in our study exhib-ited obvious side preferences, which resulted in alternatingdays of binge and abstinence behavior as the ethanol solu-tion was moved daily from side to side. The development ofside preferences may have confounded the observation ofcyclic drinking patterns in this study.
The peaks in ethanol intakes that occurred at intervals 1and 7 may be attributable either to a deprivation-like effectresulting from the imposed postsurgical periods of ethanolabstinence or to surgical stress. The ethanol deprivation ef-fect is not typically observed after this short of an absti-nence period (Heyser et al., 1997), but it has been shown tooccur after 3 days of abstinence in a continuous-access pro-cedure (Spanagel et al., 1996). Abstinence periods were ap-plied to prevent potential ethanol interactions with the sur-gical anesthetic used. Recent evidence from our laboratoryhas shown that these transient peaks in ethanol intake stilloccur after surgery when continuous access to 10E is main-tained throughout the perioperative period (unpublished ob-servations, M. M. Ford, J. C. Eldridge, & H. H. Samson,2001), supporting the suggestion that these peaks are relatedmore to surgical stress than to ethanol deprivation. Regard-less of the cause, the changes in ethanol and water intakebetween intervals 1 and 2 and between intervals 7 and 8 arelikely due to a reduction of the surgical stress, the ethanoldeprivation effect, or both.
Recent survey estimates indicate that approximately 25%of postmenopausal women receive some form of estrogen

112 M.M. Ford et al. / Alcohol 26 (2002) 103–113
replacement therapy (Wysowski et al., 1995). A single etha-nol challenge has been shown to elevate estradiol levels asmuch as threefold (of the intended target range) in womenreceiving hormone replacement therapy (HRT) (Ginsburg etal., 1996), thereby increasing the potential risk of breastcancer (Purohit, 1998). Although not yet addressed in stud-ies with human subjects, HRT in postmenopausal womenmay raise an important health issue in women who consumealcoholic beverages. The results of this study seem to indi-cate that HRT augments ethanol intake to preestablishedbaseline levels and above. In the human condition, this maytranslate to postmenopausal women receiving HRT drink-ing levels of ethanol that resemble their premenopausal lev-els. It is possible that the same level of moderate ethanolconsumption throughout subsequent life stages (premeno-pause followed by postmenopause) may have considerablydifferent health consequences for women (Gill, 2000).
In summary, ovex was associated with a significant de-cline in ethanol intake, and subsequent E2 administrationwas associated with a restoration of ethanol consumption tosham levels. Reproductive status was found to be a deter-mining factor in the responsiveness of an ethanol drinkingresponse to E2. Together, these findings provide evidencethat ovarian hormones, particularly E2, exert activational ef-fects on hormone-inducible substrates to modulate ethanolconsumption in the adult female rat. The latency of ethanolintake changes after hormone removal (either through ovexor discontinuation of E2 replacement) seems to indicate thatE2 has prolonged effects on this behavior, perhaps through agenomic mechanism of action. Additional studies will be re-quired to dissect the mechanisms by which E2 regulates eth-anol intake in the female rat.
Acknowledgments
This research was supported by grants P50AA11997 (toHHS) and T32AA07565 (to MMF) from the National Instituteon Alcohol Abuse and Alcoholism. We would like to thankDr. William Sonntag for the use of his surgical facilities andSharla Flohr for her assistance with surgical procedures.
References
Almeida, O. F. X., Shoaib, M., Deicke, J., Fischer, D., Darwish, M. H., &Patchev, V. K. (1998). Gender differences in ethanol preference and in-gestion in rats. The role of the gonadal steroid environment. J Clin In-vest 101, 2677–2685.
Aschkenasy-Lelu, P. (1960). Relation entre l’effect inhibiteur des oestro-genes sur la consommation d’alcohol du rat et leur action genitale. ArchSci Physiol (Paris) 14, 165–174.
Cailhol, S., & Mormède, P. (2001). Sex and strain differences in ethanoldrinking: effects of gonadectomy. Alcohol Clin Exp Res 25, 594–599.
Carlberg, K. A., Fregly, M. J., & Fahey, M. (1984). Effects of chronic es-trogen treatment on water exchange in rats. Am J Physiol 247, E101–E110.
Eisenhofer, G., & Johnson, R. H. (1982). Effect of ethanol ingestion onplasma vasopressin and water balance in humans. Am J Physiol 242,R522–R527.
Everett, J. W. (1989). Neurobiology of Reproduction in the Female Rat: AFifty-Year Perspective. Berlin: Springer-Verlag.
Files, F. J., Lewis, R. S., & Samson, H. H. (1994). Effects of continuousversus limited access to ethanol on ethanol self-administration. Alcohol11, 523–531.
Findlay, A. L., Fitzsimons, J. T., & Kucharczyk, J. (1979). Dependence ofspontaneous and angiotensin-induced drinking in the rat upon theoestrous cycle and ovarian hormones. J Endocrinol 82, 215–225.
Forger, N. G., & Morin, L. P. (1982). Reproductive state modulates ethanolintake in rats: effects of ovariectomy, ethanol concentration, estrous cy-cle and pregnancy. Pharmacol Biochem Behav 17, 323–331.
Gill, J. (1997). Women, alcohol and the menstrual cycle. Alcohol Alcohol32, 435–441.
Gill, J. (2000). The effects of moderate alcohol consumption on femalehormone levels and reproductive function. Alcohol Alcohol 35, 417–423.
Ginsburg, E. S., Mello, N. K., Mendelson, J. H., Barbieri, R. L., Teoh, S.K., Rothman, M., Gao, X., & Sholar, J. W. (1996). Effects of alcoholingestion on estrogens in postmenopausal women. JAMA 276, 1747–1751.
Heyser, C. J., Schulteis, G., & Koob, G. F. (1997). Increased ethanol self-administration after a period of imposed ethanol deprivation in ratstrained in a limited access paradigm. Alcohol Clin Exp Res 21, 784–791.
Hilakivi-Clarke, L. (1996). Role of estradiol in alcohol intake and alcohol-related behaviors. J Stud Alcohol 57, 162–170.
Holdstock, L., & de Wit, H. (2000). Effects of ethanol at four phases of themenstrual cycle. Psychopharmacology (Berl) 150, 374–382.
Institute of Laboratory Animal Resources, Commission on Life Sciences,National Research Council. (1996). Guide for the Care and Use of Lab-oratory Animals. Washington, DC: National Academy Press.
Juárez, J., & Barrios de Tomasi, E. (1999). Sex differences in alcoholdrinking patterns during forced and voluntary consumption in rats. Al-cohol 19, 15–22.
Kalra, S. P., Dube, M. G., Pu, S., Xu, B., Horvath, T. L., & Kalra, P. S.(1999). Interacting appetite-regulating pathways in the hypothalamicregulation of body weight. Endocr Rev 20, 68–100.
Kelly, S. J., Ostrowski, N. L., & Wilson, M. A. (1999). Gender differencesin brain and behavior: hormonal and neural bases. Pharmacol BiochemBehav 64, 655–664.
Koob, G. F. (1992). Drugs of abuse: anatomy, pharmacology and functionof reward pathways. Trends Pharmacol Sci 13, 177–184.
Koob, G. F., Roberts, A. J., Schulteis, G., Parsons, L. H., Heyser, C. J.,Hyytiä, P., Merlo-Pich, E., & Weiss, F. (1998). Neurocircuitry targetsin ethanol reward and dependence. Alcohol Clin Exp Res 22, 3–9.
Lancaster, F. E. (1994). Gender differences in the brain: implications forthe study of human alcoholism. Alcohol Clin Exp Res 18, 740–746.
Lancaster, F. E., Brown, T. D., Coker, K. L., Elliott, J. A., & Wren, S. B.(1996). Sex differences in alcohol preference and drinking patternsemerge during the early postpubertal period. Alcohol Clin Exp Res 20,1043–1049.
Lancaster, F. E., & Spiegel, K. S. (1992). Sex differences in pattern ofdrinking. Alcohol 9, 415–420.
Li, T.-K., & Lumeng, L. (1984). Alcohol preference and voluntary alcoholintakes of inbred rat strains and the National Institutes of Health hetero-geneous stock of rats. Alcohol Clin Exp Res 8, 485–486.
McEwen, B. S., & Alves, S. E. (1999). Estrogen actions in the central ner-vous system. Endocr Rev 20, 279–307.
McGinnis, M. Y., Krey, L. C., MacLusky, N. J., & McEwen, B. S. (1981). Ste-roid receptor levels in intact and ovariectomized estrogen-treated rats: anexamination of quantitative, temporal and endocrine factors influencing theefficacy of an estradiol stimulus. Neuroendocrinology 33, 158–165.
Messiha, F. S. (1981). Steroidal actions and voluntary drinking of ethanolby male and female rats. Prog Biochem Pharmacol 18, 205–215.
Mohamed, M. K., & Abdel-Rahman, A. A. (2000). Effect of long-termovariectomy and estrogen replacement on the expression of estrogenreceptor gene in female rats. Eur J Endocrinol 142, 307–314.

M.M. Ford et al. / Alcohol 26 (2002) 103–113 113
Österlund, M., Kuiper, G. G., Gustafsson, J. A., & Hurd, Y. L. (1998).Differential distribution and regulation of estrogen receptor-alpha and-beta mRNA within the female rat brain. Brain Res Mol Brain Res 54,175–180.
Purohit, V. (1998). Moderate alcohol consumption and estrogen levels inpostmenopausal women: a review. Alcohol Clin Exp Res 22, 994–997.
Roberts, A. J., Smith, A. D., Weiss, F., Rivier, C., & Koob, G. F. (1998).Estrous cycle effects on operant responding for ethanol in female rats.Alcohol Clin Exp Res 22, 1564–1569.
Samson, H. H., Schwarz-Stevens, K., Tolliver, G. A., Andrews, C. M., &Files, F. J. (1992). Ethanol drinking patterns in a continuous-access op-erant situation: effects of ethanol concentration and response require-ments. Alcohol 9, 409–414.
Samson, H. H., Tolliver, G. A., & Schwarz-Stevens, K. (1991). Ethanolself-administration in a nonrestricted access situation: effect of ethanolinitiation. Alcohol 8, 43–53.
Sandberg, D., David, S., & Stewart, J. (1982). Effects of estradiol benzoateon the pattern of eating and ethanol consumption. Physiol Behav 29,61–65.
Shughrue, P. J., Bushnell, C. D., & Dorsa, D. M. (1992). Estrogen receptormessenger ribonucleic acid in female rat brain during the estrous cycle:
a comparison with ovariectomized females and intact males. Endocri-nology 131, 381–388.
Spanagel, R., Hölter, S. M., Allingham, K., Landgraf, R., & Zieglgäns-berger, W. (1996). Acamprosate and alcohol: I. Effects on alcohol in-take following alcohol deprivation in the rat. Eur J Pharmacol 305,39–44.
Tarttelin, M. F., & Gorski, R. A. (1973). The effects of ovarian steroids onfood and water intake and body weight in the female rat. Acta Endo-crinol (Copenh) 72, 551–568.
Tolliver, G. A., Sadeghi, K. G., & Samson, H. H. (1988). Ethanol preferencefollowing the sucrose-fading initiation procedure. Alcohol 5, 9–13.
Wade, G. N., Gray, J. M., & Bartness, T. J. (1985). Gonadal influences onadiposity. Int J Obes 9 Supplement 1, 83–92.
Wade, G. N., & Zucker, I. (1970). Hormonal modulation of responsivenessto an aversive taste stimulus in rats. Physiol Behav 5, 269–273.
Wysowski, D. K., Golden, L., & Burke, L. (1995). Use of menopausal es-trogens and medroxyprogesterone in the United States, 1982–1992.Obstet Gynecol 85, 6–10.
Yen, S. S. C., Jaffe, R. B., & Barbieri, R. L. (1999). Reproductive Endocri-nology: Physiology, Pathophysiology, and Clinical Management (4thed.). Philadelphia: W. B. Saunders.