build-up and release from proactive interference during chronic ethanol consumption in mice: a...
TRANSCRIPT

Behavioural Brain Research, 25 (1987) 63-74 Elsevier 63
BBR 00682
Build-up and release from proactive interference during ethanol consumption in mice" a behavioral and
neuroanatomical study
chronic
D a n i e l B e r a c o c h e a 1, L a u r e n t L e s c a u d r o n 2, A n t o i n e T a k o 1, A la in V e r n a 2 a n d R o b e r t
J a f f a r d 1 JLaboratoire de Psvehophysiologie, C.N.R.S. UA 339, and 2Laboratoire de Cytologie, Universit~ de Bordeaux I, Talence (France)
(Received 1 January 1987) (Revised version received 10 March 1987)
(Accepted 17 March 1987)
Key words. Spontaneous alternation; Memory; Korsakoff syndrome; Ethanol; Mammillary body; Hippocampus; Interference; Retrieval
Male mice of the BALB/c strain were given a solution of 15% ethanol as their only source of fluid during either 24 or 48 weeks. They were submitted to a sequential alternation (SA) task in a T-maze (6 successive trials). It was found that 48 but not 24 weeks of alcohol administration lead to a deficit as compared to pair-fed or tap-water controls. Whereas experimental mice performed as well as controls on the first 3 choices, they exhibited a gradual decrease in the SA rate on subsequent trials. We suggest that this deficit might result from an exaggerated vulnerability to proactive interference (PI). In order to further test this hypothesis, a second experiment investigated whether a between-trials variation of context of the maze would increase performance. It was found that the SA rate improved as soon as the variation was provided (5th trial). We suggest that the deficit of experimental mice results from an impairment of retrieval processes. A neuroanatomical study was conducted to quantify cell losses resulting from 8, 24 or 48 weeks of ethanol treatment in the mammillary bodies (MM) or the hippocampus (HPC). At the time of appearance of the deficit, MM exhibited a - 32% cellular loss, whereas this was only - 18% in the HPC. This result emphasises the importance of MM lesion in memory deficits resulting from long-term alcohol consumption.
I N T R O D U C T I O N
Chronic ethanol consumption in man has been found to result in brain damage and associated learning and recent memory impairments 7,8a8, 23,24 Although the dorsomedial nucleus of the thalamus (MDT) and the mammillary bodies (MM) of the hypothalamus have been the specific structures most often associated with the alcoho- lic patient's amnesic symptoms, the question still remains of whether a combined thalamic-hypo-
thalamic lesion is needed to cause amnesia 6,2°, 21,28,30. In recent experiments aimed at producing an animal model of this pathology, we found that, in mice, chronic ethanol consumption for 6 months induced an accelerated rate of forgetting in a spatial delayed alternation task 1,2. Parallel neuroanatomical investigations showed that alco- hol-treated mice exhibited cellular losses which were much more pronounced in the MM than in either the hippocampus or thalamic nuclei 5a6-18. The fact that experimental lesions of the MM
Correspondence." D. Beracochea, Laboratoire de Psychophysiologie, C.N.R.S. UA 339, Universit6 de Bordeaux I, avenue des Facult6s, 33405 Talence Cedex, France.
0166-4328/87/$03.50 ¢c) 1987 Elsevier Science Publishers B.V. (Biomedical Division)

64
alone was sufficient to induce the same behavioral impairment as long-term ethanol consumption, led us to suggest that damage to the MM was mainly responsible for the alcohol-induced mem- ory impairment 3"4. However, compared to 6 months alcohol treatment, the complete experi- mental lesion of the MM was found to result in an additional deficit. Thus, in contrast to alcohol- treated mice, experimentally MM-lesioned sub- jects exhibited a spontaneous alternation (SA) deficit when tested with a sequential but not discrete procedure in a T-maze; more precisely, their alternation rate was found to decrease as the number of trials increased, suggesting an increas- ed vulnerability to proactive interference (PI) 3"4.
The present study was designed to investigate whether an alcohol treatment of longer duration than 6 months would lead to a more severe cellu- lar loss in the MM and, in parallel, to the same sequential alternation deficit as we found after a complete experimental lesion of this nucleus. Moreover, additional behavioral analyses were designed in an attempt to qualify the nature (ac- quisition-storage-retrieval) of the deficit. Finally, changes in neuronal density in the hippocampus were also measured, since this structure has been found to play an important role in SA behavior.
EXPERIMENT1
The first experiment was aimed at comparing SA behavior of mice subjected to either 6 or 12 months of chronic ethanol consumption by using a sequential test procedure. As pointed out above, such a procedure allowed to distinguish between the ability to alternate per se (2nd trial of a series), from the progressive inability to do so as the number of trials increased. In the latter case, repetitive testing (i.e. sequences of more than two trials) constitutes a potent source of PI, since a correct response on trial N (which depends on information received in trial N-1) may be influenc- ed by all previous information received from trial 1 to N-211. Accordingly, mice were submitted to daily sessions of 6 trials each with an intertrial interval (ITI) of 30 s, conditions which were previously shown to induce a trial-dependent
decrease of SA in MM-lesioned mice but not in control subjects 3"4.
Materials and Methods
Animals The animals were male mice of the BALB/c Orl
strain approximately 6 weeks old at the time of receipt. They were housed in cages of 15 subjects, matched for weight and placed in a colony room (ambient temperature, 22 + 1 °C; automatic light cycle, 08.00-20.00 h) with free access to food and water for 3 weeks before the beginning of ethanol administration.
Alcohol administration A total of 120 subjects were used and assigned
to 3 groups. Animals of the two experimental groups (n = 60) were given as their only source of fluid an increasing progression of ethanol solu- tions as follows: 5~o (v/v) solution for the first week; 10~o (v/v) the second week and 15~o (v/v) for the remaining time. The solutions were mixed from 95~o ethanol and supplemented with sac- charose (30 g/l). They were freely available to subjects in two 250-ml bottles. Dry food was also freely available throughout the experiments (Extra-Labo, Pietrement, France). Twice weekly, the subjects were weighed and the quantity of food and ethanol solutions consumed were measured; at the same time, the animals were replaced in clean cages with fresh ethanol solu- tions and dry food. Mice of the first control group (n = 30) were pair-fed with an isocaloric solution of dextri-maltose and dry food. Animals assigned to the second control group (n = 30) had ad libi- tum access to dry food and tap water.
Animals of the 1st and 2nd experimental groups received ethanol for 24weeks and 48 weeks respectively. At the end of this period, water was progressively substituted for ethanol (or dextri-maltose) by steps of 5 ~o (v/v) a week. Behavioral testing began following a 4-week complete withdrawal period.
Behavioral testing The experiments were run blind. All testing was
conducted in a T-maze constructed of wood. The

stem and arms were 35 cm long, 10 cm wide and 25 cm high. The starting box (10 x 12 cm) was separated from the stem by a vertical sliding door. Horizonta l sliding doors were placed at the entrance of each arm. Three and two days before the beginning of testing for SA, animals were given two free exploration sessions of 5 min each in order to familiarize them with the conditions they would encounter in the alternation test. Then, subjects were submitted to a session of 6 successive free trials each day. To begin a trial, the subject was placed in the start box and after 30 s, the door to the stem was opened. When the mouse entered one of the arms, the door to that arm was closed. The chosen arm and the time which elapsed between opening the door and choosing the arm (choice latency) were noted. After a 30-s confinement in the chosen arm, the mouse was removed and placed in the start box for the second trial. Visible traces of urine and faeces were removed from the stem and arms between trials. Behavioral testing was conducted on mice ran- domly selected in each group. Alcohol-treated mice (24 weeks: n = 15; 48 weeks: n = 20) and their pair-fed (n = 8 and n = 10 respectively) and water (n = 7 and n = 10) controls were submitted to 4 daily sessions of 6 successive trials.
Statistical analysis
Differences between groups were analysed using repeated measures analysis of variance (ANOVA). The MacNemar test and the M a n n - W h i t n e y U-test were used for other com- parisons.
65
Results
Nutrients intake and body weight. As shown in Table I, there were no statistically significant differences between group body weights (F2.28 = 0.143). Moreover, the duration of the treatments does not modify the evolution of body
weights in the experimental and pair-fed groups (F2,28 = 0.56). The number of calories daily ingested as dry food was similar between pair-fed and ethanol groups, but inferior to that consumed by the water group (F2,28 = 0.02). However , the total amount of calories daily ingested was quite similar for all groups (F2,28 = 0.305).
Behavior
Results for the 24 weeks and 48 weeks of treat- ment are summarized as followed:
24 weeks: no significant differences were ob- served between the pair-fed and water groups (ANOVAs, F < 0 . 8 ) ; thus, they were pooled (control group, n = 15) for statistical analysis. The mean rate of spontaneous alternation (6 trials x 4 sessions) of alcohol (78.3%) and con- trol subjects (75.6~o) was not statistically differ- ent ( F I . 2 8 = 0.84); moreover the evolution of SA rates across successive trials was the same
(F4,112 = 0.22). 48 weeks: no significant differences were ob-
served between the pair-fed and water groups (ANOVAs, F < 0.35) thus they were also pooled (control group, n = 20) for statistical analysis. Alcohol treatment induced a decrease in the mean
rate of SA (63.5~o vs 76.5~o, fl,3~ = 19.4,
TABLE I
Mean caloric intake during the 48 weeks of diets administration and mean body weights (+_ S.E.M.) at the start (lst week) and at the end (24 weeks or 48 weeks) of treatments followed by a 4-week withdrawal period
Body weights (g)
1st Week 24 Weeks
Cah)ric intake/@
48 Weeks Dry food Dextri Sucrose Ethanol Total
Water 22.4 +. 1.4 34.2 _+ 1.8 34.2 + 1.5 408 Pair-fed 21.1 + 1.4 34.7 + 2.6 35.7 _+ 2.0 267 Ethanol 21.0 +_ 2.1 34.9 + 2.3 34.8 _+ 2.7 267
- - - 408 160 - - 427 - 60 100 427

66
TABLE I1
Median choices latency on each trial in the usual condition (same context) and with a context variation occurring on trials 5 (cardboard added: variation-ON) and 6 (carboard removed: variation-OFF)
Trial 2nd 3rd 4th 5th 6th
ALCOHOL Usual 19 22 24 16 17 Variation 52* 32*
CONTROLS Usual 18 22 22 18 17 Variation 56* 38*
* Variations-ON and -OFF produced for each group a signifi- cant increase in latencies (P < 0.02). In the same situation (i.e. usual or variation) there were no between-groups differ- ences.
P < 0.001). However this impairment was not uniform, since experimental subjects exhibited a progressive decrease in SA rates across succes- sive trials (interaction x treatment x trials, F4.~52 = 3.98, P = 0.004). Thus, while no signifi- cant between-group differences were observed on the 2nd trial of the session (alcohol: 85.0 + 4.1% ; controls: 81.3 + 4.0 %), additional trials resulted in a dramatic impairment of SA in alcohol as compared to control mice ( P < 0.01 for trials 4-6). An analysis performed on trials 4 -6 revealed that the deficit of alcohol-treated mice is much more pronounced when the N trial was preceded (trial N-I) by an alternated choice (for example, N-2: right and N-l : left) than by a repeated choice (right-right) (respectively 49.6% and 66.7%, 22 = 6.86, P < 0.01); this phenome- non was not observed in control subjects (76.4% vs 83.3%, X 2= 1.34, P < 0 . 2 0 ) . Finally, no between-groups differences were observed for choice latencies (see Table II).
Discussion
Our results show that chronic ethanol treat- ment produces an impairment of SA which criti- cally depends on both the duration of ethanol administration and the use of a sequential test procedure. The absence of any behavioral impair- ment after 24 weeks of alcohol treatment confirms
our previous findings 1"2, while the observed dis- ruption of SA behavior after 48 weeks of ethanol administration agrees with previous reports showing that alcohol treatment disrupts SA in rodents 34. However, it seems highly important to emphasize that the deficit presently observed closely depends on the repetition of trials within the same session and, as such, would more likely result from an exaggerated vulnerability to PI than to an impairment of the innate tendency to alternate. Thus, on one hand, experimental mice alternated at normal rates on the 2nd and 3rd trials, additional trials being required to reveal the deficit; on the other hand, the deficit is mostly observed after alternated choices, a condition which obviously increases the PI interference effect 11
E X P E R I M E N T 2
Previous experiments using the same procedure have shown that MM-lesioned mice exhibited the same deficit as that we found in 48-weeks alcohol- treated mice, namely a progressive decrease in SA as the number of trials increases 3'4. In the above- mentioned previous experiment, we determined to establish whether providing the MM-lesioned animals with an extrinsic cue on one trial (5th trial) would increase the rate of SA on the sub- sequent (6th) trial. Such an assumption was based on the fact that, in amnesic patients, proactive interference between successive lists of words was substantially reduced when each list was associat- ed with a different context 36. Unexpectedly, the performance of MM-lesioned subjects was found to be markedly improved as soon as the cue was provided (5th trial), thus suggesting an action on retrieval p r o c e s s e s 3"4. The aim of the present experiment was to determine whether such a variation of context would have the same effect on the deficit exhibited by alcohol-treated mice (48 weeks). Indeed, experiments currently in pro- gress in our laboratory indicate that this proce- dure (context variation) does not obligatorily reverse this type of deficit; more precisely, neither aged-deficient animals nor normal mice tested with a longer ITI are aided by the context variation procedure (unpublished observations).
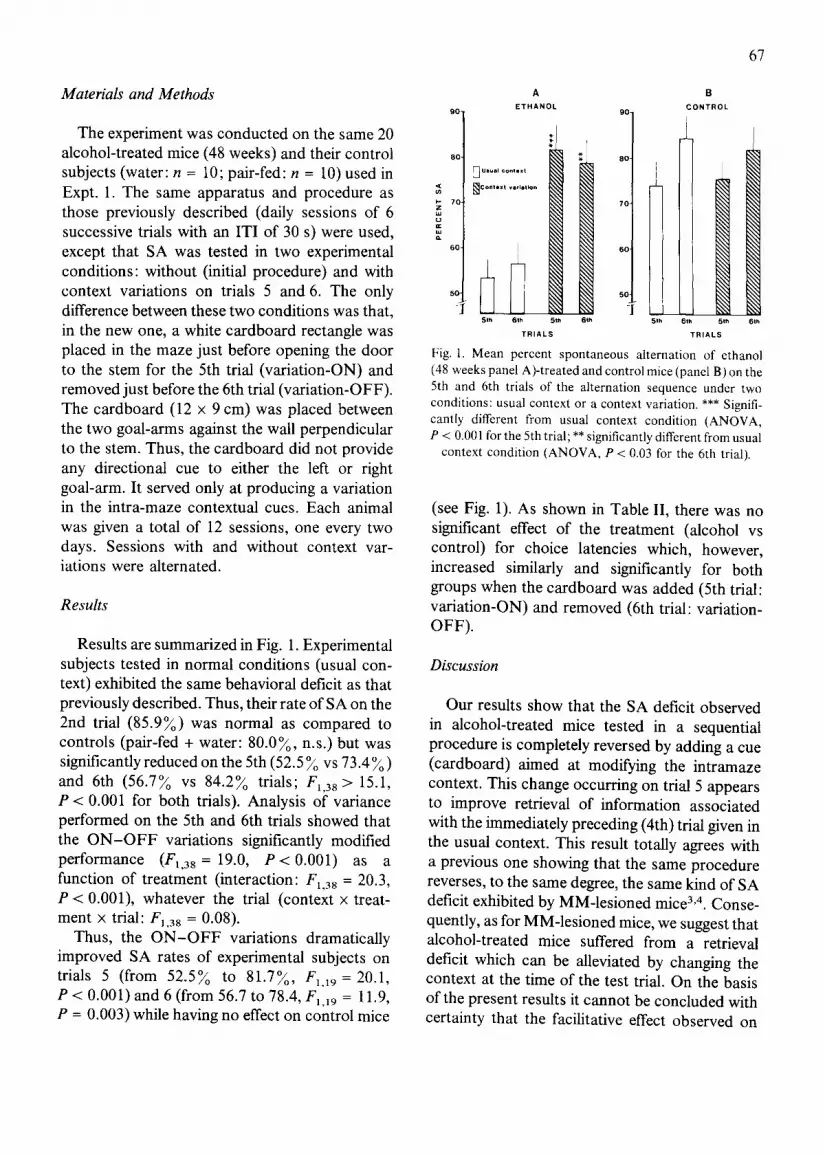
9 0
Materials and Methods
The experiment was conducted on the same 20 alcohol-treated mice (48 weeks) and their control subjects (water: n = 10; pair-fed: n = 10) used in Expt. 1. The same apparatus and procedure as those previously described (daily sessions of 6 successive trials with an ITI of 30 s) were used, except that SA was tested in two experimental conditions: without (initial procedure) and with context variations on trials 5 and 6. The only difference between these two conditions was that, in the new one, a white cardboard rectangle was placed in the maze just before opening the door to the stem for the 5th trial (variation-ON) and removed just before the 6th trial (variation-OFF). The cardboard (12 x 9 cm) was placed between the two goal-arms against the wall perpendicular to the stem. Thus, the cardboard did not provide any directional cue to either the left or right goal-arm. It served only at producing a variation in the intra-maze contextual cues. Each animal was given a total of 12 sessions, one every two days. Sessions with and without context var- iations were alternated.
Results
Results are summarized in Fig. 1. Experimental subjects tested in normal conditions (usual con- text) exhibited the same behavioral deficit as that previously described. Thus, their rate of SA on the 2nd trial (85.9~o) was normal as compared to controls (pair-fed + water: 80.0~o, n.s.) but was significantly reduced on the 5th (52.5 ~o vs 73.4 ~o ) and 6th (56.7~o vs 84.2~o trials; FL38 > 15.1, P < 0.001 for both trials). Analysis of variance performed on the 5th and 6th trials showed that the O N - O F F variations significantly modified p e r f o r m a n c e (F1,38 = 19.0, P<0 .001 ) as a function of treatment (interaction: F1,38 = 20.3, P < 0.001), whatever the trial (context x treat- ment x trial: F1,38 = 0 .08) .
Thus, the O N - O F F variations dramatically improved SA rates of experimental subjects on trials 5 (from 52.5~o to 81.7~o, F],19 = 20.1, P < 0.001) and 6 (from 56.7 to 78.4, Fl,19 = 11.9, P = 0.003) while having no effect on control mice
8 0
50"
7o
uJ
m o.
6O
m
A B E T H A N O L C O N T R O L
9 0
~ usual context
~ Context vaaltbon
I
5th 6th 5t~ 6th
TRIALS
67
5 th 6th 5th 6th
TRIALS
Fig. I. Mean percent spontaneous alternation of ethanol (48 weeks panel A)-treated and control mice (panel B) on the 5th and 6th trials of the alternation sequence under two conditions: usual context or a context variation. *** Signifi- cantly different from usual context condition (ANOVA, P < 0.001 for the 5th trial; ** significantly different from usual
context condition (ANOVA, P < 0.03 for the 6th trial).
(see Fig. 1). As shown in Table II, there was no significant effect of the treatment (alcohol vs control) for choice latencies which, however, increased similarly and significantly for both groups when the cardboard was added (5th trial: variation-ON) and removed (6th trial: variation- OFF).
Discussion
Our results show that the SA deficit observed in alcohol-treated mice tested in a sequential procedure is completely reversed by adding a cue (cardboard) aimed at modifying the intramaze context. This change occurring on trial 5 appears to improve retrieval of information associated with the immediately preceding (4th) trial given in the usual context. This result totally agrees with a previous one showing that the same procedure reverses, to the same degree, the same kind of SA deficit exhibited by MM-lesioned mice 3,4. Conse- quently, as for MM-lesioned mice, we suggest that alcohol-treated mice suffered from a retrieval deficit which can be alleviated by changing the context at the time of the test trial. On the basis of the present results it cannot be concluded with certainty that the facilitative effect observed on

68
trial 6 does result from the removal of the cardboard (variation-OFF); however, previous experiments conducted on MM-lesioned mice have been conclusive on this particular (but here of quite minor interest) point. Moreover, one can note that variation-OFF, which consists of returning to the usual context, is likely to be perceived by the animals as a variation of context since, as for the just preceding variation-ON, it produces a significant increase in choice latencies. Finally, from a behavioural point of view, alcohol- treated and MM-lesioned mice exhibit the same SA deficit which can be reversed in the same way, suggesting that both suffered from the same basic memory (retrieval) impairment. Since long-term alcohol consumption has been found to produce lesions in several brain structures, among which the M M 16, the purpose of Expt. 3 was to deter- mine whether the amnesic effect of chronic ethanol administration can be explained by damage to MM. In fact, this third experiment is the continuation of neuroanatomical investi- gations already engaged by our group either for other brain structures or for shorter periods of ethanol treatment 5'16-1s.
EXPERIMENT 3
The aim of the experiment was to evaluate MM and hippocampal (HPC) damage as a function of the duration of ethanol administration. Indeed, firstly, MM is the cerebral structure whose dam- age seems to be associated with (even if not the cause of) the amnesic Korsakoff syndrome 6'~3' 2~,32. Secondly, lesions of the hippocampus are generally considered to induce a quite different type of amnesia (bitemporal as opposed to diencephalic amnesia29); more importantly, HPC lesions have been shown to disrupt SA in rodents ~°. Accordingly, we compared the evolu- tion of cell loss in these two structures after 8, 24 and 48 weeks of alcohol administration.
Materials and Methods
Animals Subjects were randomly taken from the 24- and
48-weeks alcohol-treated groups (n = 5 for each
group) and their controls (pair-fed: n = 3 ; water: n = 2). The same procedure was used for animals which had received ethanol for only 8 weeks (n = 5) and for their pair-fed (n = 3) and water (n = 2) controls.
Methods The experiment was run blind. Animals were
coded, weighed, anaesthetized and killed. Brains were removed, weighed, and immersed for 3 days in Bouin-Holland f'Lxative. Then, they were dehy- drated, embedded in paraffin, and cut into serial frontal 7-#m thick sections. Sections were stained with erythrosin-Toluidine blue. Quantitative de- termination of the neuronal density was perform- ed as follows: for each brain, counting was per- formed on 9 frontal sections separated by 56 #m for MM, and on 30 frontal sections also separated by 56/~m in the hippocampus. For each section, quantitative analysis was made by counting all neurons showing a clearly defined nucleus within a rectangular frame of 120 × 178 mm for MM and HPC sections superimposed on a television screen and centered on the studied structure.
Results
Brain weights There were no significant differences between
alcohol-treated and pair-fed mice brain weights, which however were slightly inferior (1.8-5.7 ~o ) to those of the water groups (0.4129 + 0.003 g, 0.4184 + 0.001 g and 0.4277 + 0.009 g for, re- spectively, alcohol, pair-fed and water groups).
Neuroanatomy Hippocampus. For quantitative analysis, the
hippocampus was divided into anterior and pos- terior parts. Neuronal densities were not signifi- cantly different between pair-fed and water groups (F~.9 = 0.74) so that these two groups were pooled (control group, n = 5) for statistical analysis. ANOVA showed that ethanol adminis- tration led to a significant cellular loss in the hippocampus as a whole (F1,24 = 103.6, P < 0.001). As shown in Table III, this loss of cells is a function of treatment duration (F2.24 = 28.0, P < 0.001). The effect of ethanol is

69
T A B L E III
Mean neuronal density observed in the anterior (AH) and posterior hippocampus (PH) following 8, 24 or 48 weeks ethanol treatments and in control (pair-fed or tap-water) groups
The cellular loss is expressed in % (compar i son be tween e thanol and pooled control groups) . A H , anter ior h i p p o c a m p u s ; PH ,
pos ter ior h ippocampus .
Groups 8 Weeks 24 Weeks 48 Weeks
A H PH A H PH A H PH
Wate r 52.0 _+ 1.9 59.9 _+ 0.75 53.0 _+ 0.87 59.3 + 1.51 50.6 + 0.76 58.27 + 0.81
Pair-fed 51.4 _+ 0.57 61.1 + 0.7 49.9 + 1.1 58.6 + 0.21 50.3 + 2.6 59.20 + 0.85 Ethanol 50.3 + 2.0 59.0 _+ 1.6 50.6 + 2.1 52.2 + 1.8 41.9 _+ 3.2 45.5 + 2.8
Cellular loss - 2.7% - 2.4% - 1.5% - 11.4% - 17% - 22.5%
much more pronounced in the anterior than in the ventral hippocampus (F1.8 = 13.8, P = 0.006), although at 48 weeks cellular losses in these two parts were not statistically different (F1,8 = 0.94, see Fig. 2).
Mammillary bodies. For the quantitative analy- sis, the median MM was divided into 3 parts: anterior, median and posterior. Neuronal den- sities were not significantly different between pair- fed and water groups ( E l , 9 = 1.06, P > 0.35), so that these two groups were pooled (control group, n = 5) for statistical analysis. ANOVA showed that ethanol administration led to a highly signifi- cant cellular loss in MM as a whole (F1,24 = 34.4, P < 0.001). As shown in Table IV, this loss of cells is a function of treatment duration (F2,24 = 35.1, P < 0.001) and is more important in the posterior, as compared to the median and anterior parts of the structure (F2,48 = 19.0, P < 0.01).
Discussion
Our results show that the effects of the ethanol treatment were function both of duration and of the studied structures. We have found that the cellular loss observed in both the hippocampus and mammillary bodies was greater for the longest duration (48 weeks) as compared to other ones (8 or 24 weeks). We also found that the mammillary bodies exhibited more damage than
the hippocampus, whatever the treatment du- ration considered (8, 24 or 48 weeks). Our results agree well with findings of other groups showing that long-term administration of ethanol in rodents induces cellular losses and alteration of dendritic spines in hippocampus 25,2v,33 and other brain areas 5'31. The question remains to know whether or not simultaneous damage to several structures is necessary to induce behavioral and/or cognitive impairments. One of the main interesting results of our study was the demon- stration that such treatment did not affect brain structures with the same severity. This differential effect is of great importance to identify structures which could be principally associated with memo- ry disturbances induced in alcohol-related amnesia. More explicitly, we have shown in a previous report that 24 weeks ethanol adminis- tration induced a delayed alternation deficit for long but not short delays 1'2. The neuroanatomical analysis we performed showed that this treatment hardly damaged thalamic structures (anteroven- tral, anterodorsal and mediodorsal nuclei) and the nucleus basalis magnocellularis 5, whereas, as it is shown in Table IV, MM exhibited the greatest cellular loss. Moreover, experimental lesions re- stricted to the MM resulted in a very similar behavioral impairment (see Introduction). Taken together, these results emphasise the importance of the mammillary bodies in the alcoholic memory pathology. This conclusion agrees with clinical observation S 6" 19,20.

70
E~
h~
.!
-~1 + 1 + l ~
~ ( ' ~ ~
J r l ~-L + 1 ~ "
+1 +1 +l ~
+1 +~ +1 ~
+1 +~ + i ~
÷t +t +i ~
L ~ ~ I r~ ,p~
+t +l +t ~

7!
Fig. 2. Photomicrographs of anatomically matched frontal sections through the mouse anterior (A,B) and posterior (C,D) hippocampus and median mammillary nucleus (E,F), illustrating morphological alterations induced by 48 weeks of ethanol administration. B,D,F: ethanol-treated mouse. A,C,E: pair-fed mouse. SO, stratum oriens; SP, stratum pyramidale; SR, stratum
radiatum. Bar = 20 ~m.

72
GENERAL DISCUSSION
Our main results can be summarized as follows. The first experiment shows that chronic ethanol consumption in mice induces an impair- ment of spatial alternation in a T-maze: a result which agrees with previous findings 34. However, this deficit depends both on the duration of ethanol treatment and on the testing procedure used. Thus, on one hand, when tested after 24 weeks of ethanol administration, experimental subjects behave normally, while after 48 weeks of treatment they are significantly impaired; on the other hand, a sequential test procedure is required to observe a deficit, 48-weeks alcohol-treated mice being only impaired on the last 3 trials of a 6-trials session performed with an ITI of 30 s. The second experiment shows that providing experi- mental mice with a change in intramaze contex- tual cues (cardboard added, 5th trial; or removed, 6th trial) completely alleviates the sequential test-dependent alternation deficit on these two trials. The third experiment shows that whatever the duration of ethanol treatment (8 weeks to 48 weeks), the median mammillary nucleus exhi- bits a more important cellular loss than the hippo- campus.
Our current hypothesis is that long-term ethanol consumption impairs alternation because it increases the animal's vulnerability to interfer- ence. Indeed, one can postulate that the main problem the animal appears to have when required to alternate on more than two successive trials (sequential test procedure) is to segregate, at a given trial (N), the last information (target trial: N- l ) from the previous one (interfering trials: N-2, N-3 ...). The same pattern of deficit has recently been observed in a spatial delayed non- matching to sample task performed in an auto- mated radial maze. In such a situation, alcohol- treated mice were found to perform as well as controls on the first test, but were markedly impaired when submitted to additional tests occurring at 1-min between-test intervals (Tako et al., in preparation). Finally, as far as the com- parison can be made, our results resemble those reported for amnesic Korsakoff patients who ex- hibit a slightly more pronounced build-up of PI
than controls in a proactive inhibition paradigm ~. It has been suggested that PI build-up and release may reflect processes that occur primarily at the time of initial encoding; namely, the repetition of similar items would lead to a decrement in atten- tion which would be responsible for a decrease in memory performance 9,26. The results obtained on the second experiment do not support this hypo- thesis. It has been found that a variation of the context of the maze occurring on the last two trials of the sequence alleviate the deficit. More precisely, a variation of context on the 5th test trial improved retrieval of information associated with the immediately preceding (4th) trial which has been carried out under normal context condi- tions (the change of context was also effective on the 6th trial). This clearly shows that under normal context conditions, the encoding of infor- mation is not impaired (4th trial), but rather an impairment of the retrieval processes seems to be responsible for the deficit (compare 5th trial under normal and variation of context conditions: Fig. 2). The fact that such a context variation was particularly efficient when the test trial followed alternated choices (the condition which provided the greatest interference), suggests that changing the context led the animal to more actively reconstruct the sequence of past events. Such an interpretation agrees well with recent psychologi- cal studies performed on normal subjects and with the proposals that the amnesic deficits in the Korsakoff syndrome stem from a basic failure to sustain the searching operations involved in nor- mal remembering 22, to a premature closure of the problem-solving activity 3° or, as stated for other forms of amnesia, to a failure of effortful cognitive operations at the time of retrieval ~9"35
Our anatomical results show that chronic alcohol consumption induces significant neuronal loss in both the hippocampus and MM. Within these two structures the treatment does not affect all regions with the same severity. Thus, on one hand, the posterior part of the median mammilla- ry nucleus is more sensitive to alcohol than the anterior one, while, on the other hand, the posterior hippocampus exhibits significantly higher cell losses than the anterior part of the structure. Taken as a whole, these cell losses are

function of the duration of ethanol treatment, the MM exhibiting first a significant loss of cells (posterior part) as soon as after 8 weeks of treat- ment, 24 weeks being required to observe a similar phenomenon in the posterior hippocampus. Fi- nally, cells of the median mamillary nucleus seem to be much more sensitive than the hippocampal cells to ethanol and, as shown in previous experi- ments, than either thalamic nuclei (anteroventral, anterodorsal and mediodorsal nuclei) or the nucleus basalis magnocellularis 5. These results agree with anatomical investigations performed on brains of amnesic Korsakoff patients, since in all cases which have been reported, MM is the more frequently damaged brain structure 6'32.
It has been suggested that MM lesion would play an important role in retrieval deficits 4"~2"19. The present neuroanatomical findings support this assumption, since MM exhibit a greater cellu- lar loss than the hippocampus at the time of the appearance of the deficits. However, one cannot totally exclude a possible involvement of the hippocampus in the sequential alternation deficit, since it appears that, during 24 and 48 weeks of ethanol administration, the mean cellular loss is more pronounced in the hippocampus (from 6.5~o to 20~o at 24 and 48 weeks respectively) than in the MM (from 18.8~o to 26.2~o at 24 and 48 weeks respectively). However, it seems diffi- cult to attribute to damage observed in the hippo- campus an important role in the presently observed deficit. On the one hand, it has been shown that the complete lesion of the different subfields of this structure by ibotenic acid injections do not lead to severe memory deficits in a place task; such a deficit was observed only after combined lesions of both the hippocampus and the subiculum15; on the other hand, the pattern of SA deficit observed in the present study does not resemble that observed following hippo- campal lesion. It has effectively been observed that hippocampal lesion disrupts SA 'as a rule'J°'~4; namely it was found that on two successive trials (lst and 2nd), rodents were unable to alternate (Left-Right) but rather exhi- bited a perseverative response (Left-Left) 1°,~4. This was not the case in our study, since ethanol- treated mice exhibited a rate of SA similar to that
73
observed in control mice on the first 3 successive trials of the series. Additional trials were required to observe a deficit.
The fact that experimental lesions of the MM induce the very same sequential alternation deficit as that observed in alcohol-treated mice, and that MM is the most sensitive structure to alcohol treatment (at least when compared with hippo- campus and thalamic nuclei), indicate that MM damage may be principally responsible for the deficit. However, current experiments are in pro- gress to determine whether or not combined moderate damage of both MM and ventral hippo- campus could induce a sequential alternation deficit as compared to a single complete lesion of MM or the hippocampus.
ACKNOWLEDGEMENTS
This study was supported by a grant from 'Haut Comit6 d'Etude et d'Information sur l'alcoolisme', 1984-36, and by the C.N.R.S.
REFERENCES
1 Beracochea, D., Durkin, T.P. and Jaffard, R., On the involvement of the central cholinergic system in memory deficits induced by long-term ethanol consumption in mice, Pharmacol. Biochem. Behav., 24 (1986) 519-524.
2 Beracochea, D. and Jaffard, R., Memory deficits subse- quent to chronic consumption of alcohol in mice: an analysis based on spontaneous alternation behavior, Behav. Brain Res., 15 (1985) 15-25.
3 Beracochea, D. and Jaffard, R., Mammillary bodies lesions in mice induce memory deficits which resemble those of the amnesic Korsakoff syndrome. In B. Will, P. Schmitt and J.C. Dalrymple-Alford (Eds.), Brain Plastici- ty, Learning and Memory, Plenum, 1985, p. 546.
4 Beracocbea, D. and Jaffard, R., Impairment of spon- taneous alternation behavior in sequential test proce- dures following mammillary bodies lesions in mice. Evidence for time-dependent interference-related memo- ry deficits, Behav. Neurosci., 101 (1987) 101-111.
5 Beracochea, D., Lescaudron, L., Verna, A. and Jaffard, R., Neuroanatomical effects of chronic ethanol con- sumption on dorsomedial and anterior thalamic nuclei and on substantia innominata in mice, Neurosci. Lett., 73 (1987) 81-84.
6 Brion, S. and Mikol, J., Atteinte du noyau lat6ral dorsal du thalamus et syndrome de Korsakoff alcoolique, J. Neurol. Sci., 38 (1978) 249-261.

74
7 Butters, N., Cermak, L.S., Montgomery, K. and Adinolfi, A., Some comparisons of the memory deficits of chronic alcoholics and patients with Korsakoffs disease, Alcohol Clin. Exp. Res., 1 (1977) 73-80.
8 Butters, N. and Cermak, L.S., Alcoholic's Korsakoffs syn- drome. An Information-Processing Approach to Amnesia, Academic, New York, 1980.
9 Dillon, R.F., Locus of proactive interference effects in short-term memory, J. Exp. Psychol., 99 (1973) 75-81.
10 Douglas, R.J., The development of hippocampal function: implications for theory and for therapy. In R.L. Isaacson and K.H. Pribram (Eds.), The Hippocampus, Vol. 2, Plenum, New York, 1975, pp. 327-361.
11 Grant, D.S., Intertrial interference in rat short-term memory, J. Exp. Psychol., 7 (1981) 217-227.
12 Greene, E. and Naranjo, J.N., Thalamic role in spatial memory, Behav. Brain Res., 19 (1986) 123-131.
13 Irle, E. and Markowitsch, H.J., Widespread neuro- anatomical damage and learning deficits following chronic alcohol consumption or vitamin B~ (thiamine) deficiency in rats, Behav. Brain Res., 9 (1983) 277-294.
14 Isaacson, R.J., The Limbic System, 2nd edn., Plenum, 1982, 387 pp.
15 Jarrard, L.E., Selective hippocampal lesions and be- havior: implications for current research and theorizing. In R.L. Isaacson and K.H. Pribram (Eds.), The Hippo- campus, Vol. 4, Plenum, New York, 1986, pp. 93-126.
16 Lescaudron, L., Beracochea, D., Verna, A. and Jaffard, R., Chronic ethanol consumption induces neuronal loss in mammillary bodies of the mouse: a quantitative analy- sis, Neurosci. Lett., 50 (1984) 151-155.
17 Lescaudron, L., Seguela, P., Geffard, M. and Verna, A., Effects of long-term ethanol consumption on GABAergic neurons in the mouse hippocampus: a quantitative immu- nocytochemical study, Drug, Alcohol Dependence, 18 (1986) in press.
18 Lescaudron, L. and Verna, A., Effects of chronic ethanol consumption on pyramidal neurons of the mouse dorsal and ventral hippocampus: a quantitative histological analysis, Exp. Brain Res., 58 (1985) 362-367.
19 Lhermitte, F. and Signoret, J.L., Analyse neuro- psychologique et diff6rentiation des syndromes amndsi- ques, Rev. Neurol., 126 (1972) 161-178.
20 Mair, W.G.P., Warrington, E.K. and Weiskrantz, L., Memory disorders in Korsakoff's psychosis: a neuropathological and neuropsychological investigation of two cases, Brain, 102 (1979) 749-783.
21 Markowitsch, H.J. and Pritzel, M., The neuropathology of amnesia, Prog. Neurobiol., 25 (1985) 189-288.
22 Meissner, N.W., Learning and memory in the Korsakoff syndrome, J. Neuropsychiatry, 4 (1968) 6-20.
23 Oscar-Berman, M. and Zola-Morgan, S.M., Compara- tive neuropsychology and Korsakoffs syndrome. I. Spa- tial and visual reversal learning, Neuropsychologia, 18 (1980) 499-512.
24 Parson, O.A., Neuropsychological deficits in alcoholics: facts and fancies, Alcoholism, l (1977) 51-56.
25 Phillips, S.C. and Cragg, B.G., Chronic consumption of alcohol by adult mice: effect on hippocampal cells and synapses, Exp. Neurol., 80 (1983) 218-226.
26 Radtke, R.C. and Grove, E.K., Proactive inhibition in short-term memory: availability or accessibility? J. Exp. Psychol. : Human Learning and Memory, 3 (1977) 78-91.
27 Riley, J.N. and Walker, D.W., Morphological alterations in hippocampus after long-term alcohol consumption in mice, Science, 201 (1978) 646-648.
28 Signoret, J.L. and Goldberg, G., Troubles de memoire lors des 16sions du thalamus chez l'homme, Rev. Neurol., 142 (1986) 445-448.
29 Squire, L.S., The neuropsychology of human memory, Ann. Rev. Neurosci., 5 (1982) 241-273.
30 Talland, G.A., Deranged memory, Academic, New York, 1965, 356 pp.
31 Tavares, M.A., Paula-Barbosa, M.M., Alcohol-induced granule cell loss in the cerebellar cortex of the adult rat, Exp. Neurol., 78 (1982) 574-582.
32 Victor, M., Adams, R.D. and Collins, G.H., The Wernicke-Korsakoff Syndrome, Davis, Philadelphia, PA, 1971.
33 Walker, D.W., Barnes, D.E., Zornetzer, S.F., Hunter, B.E. and Kubanis, P., Neuronal loss in hippocampus induced by prolonged ethanol consumption in rats, Science, 209 (1980) 711-713.
34 Walker, D.W., Hunter, B.E. and Abraham, W.C., Neuroanatomical and functional deficits subsequent to chronic ethanol administration in animals, Alcohol Clin. Exp. Res., 5 (1981) 267-281.
35 Weingartner, H., Psychobiological determinants of memory failures. In L.R. Squire and N. Butters (Eds.), Neuropsychology of Memory, The Guilford Press, U.K. 1984, pp. 203-212.
36 Winocur, G. and Kinsbourne, M., Contextual cueing as an aid to Korsakoff amnesics, Neuropsychologia, 16 (1978) 671-682.