effects of invasion by introduced versus native conifers on coastal heathland vegetation
TRANSCRIPT

Journal of Vegetation Science && (2012)
Effects of invasion by introduced versus native coniferson coastal heathland vegetation
Heidi I. Saure, Vigdis Vandvik, Kristian Hassel & Ole R. Vetaas
Keywords
Aspect; Canopy effect; Natural afforestation;
Picea sitchensis; Pinus sylvestris; PRC;
Semi-natural landscapes; Single tree impact;
Species composition; Species richness
Abbreviations
DCA = detrended correspondence analysis;
CCA = constrained correspondence analysis;
GLMM = generalized linear mixed model;
PRC = principal response curves.
Nomenclature
Lid & Lid (2005) ; Hill et al. (2006) ; Soderstrom
et al. (2002)
Received 7 November 2011
Accepted 26 September 2012
Co-ordinating Editor: Sam Scheiner
Saure, H.I. (corresponding author, heidi.
[email protected]): NLA University College, PO
Box 74 Sandviken, N-5812, Bergen,Norway
Saure, H.I. & Vandvik, V. (vigdis.
[email protected]): Department of Biology,
University of Bergen, PO Box 7803, N-5020,
Bergen, Norway
Hassel, K. ([email protected]):
Systematics and Evolution Group, Museum of
Natural History and Archaeology, Section of
Natural History, Norwegian University of
Science and Technology, N-7491, Trondheim,
Norway
Vetaas, O.R. ([email protected]):
Department of Geography, University of
Bergen, PO Box 7802, N-5020, Bergen, Norway
Abstract
Questions: What are the effects of conifer invasion on plant species composi-
tion and richness of coastal heathlands; do effects differ between heathlands
invaded by introduced or native coniferous tree species?
Location: Threatened coastal Calluna heathlands, western Norway.
Methods:We compared the effects of invasion by introduced Sitka spruce (Picea
sitchensis (Bong.) Carr.) and native Scots pine (Pinus sylvestris L.) on heathland
plant communities by focusing on the local impacts of single coniferous tree can-
opies. For each of 58 target coniferous trees (2–4-m tall), 2-m transects were laid
out in a north and south orientation from the tree stem to beyond the tree can-
opy, and control transects were laid out in open heathland adjacent to the target
trees. Vascular plants and bryophytes were recorded within 0.0625 m2 contigu-
ous quadrats along the transects. Principal response curves (PRC) and general-
ized linear mixed models (GLMMs) were used to investigate tree canopy effects
on species composition and richness.
Results: Invasion by coniferous trees induced considerable changes in the
coastal heathland vegetation: distinct microcommunitites developed beneath
the canopies of relatively young, single coniferous trees in the heathlands. There
was amarked difference in subcanopy species composition under the introduced
Sitka spruce relative to the native Scots pine; forest floor species had colonized
to a greater extent under Sitka spruce, replacing the light-demanding species
characteristic of the coastal heathland vegetation.
Conclusion: Our study showed that conifer invasion induces considerable and
rapid changes in heathland vegetation, and that the effects were stronger under
the introduced conifer. This suggests that Sitka spruce invasionmay pose a more
immediate threat to the characteristic coastal heathland vegetation and flora
than succession based on native conifers. The open coastal heathlands along the
Atlantic coast of Western Europe are a threatened landscape of high conserva-
tion value, and our results suggest that the spread of Sitka spruce into these
landscapes should be controlled, especially in heathlands that are targeted for
conservation.
Introduction
Coastal heathlands are semi-natural, treeless landscapes
that extend almost 3600 km along the Atlantic coast of
Western Europe, from Portugal in the south to northern
Norway in the north. The heathlands are dominated by
ericaceous species, in particular Calluna vulgaris, and they
harbour a characteristic light-demanding oceanic flora, but
are often relatively species-poor (Gimingham 1961;
Fremstad 1997; Loidi et al. 2010). As key cultural land-
scapes of Western Europe that harbour characteristic habi-
tats and biodiversity, coastal heathlands are important
targets for nature conservation (Webb 1998; Council of
Europe 2000).
Journal of Vegetation ScienceDoi: 10.1111/jvs.12010© 2012 International Association for Vegetation Science 1

The coastal heathlands are man-made landscapes that
developed gradually from 6000 to about 1000 yrs ago, fol-
lowing forest clearance (Kaland 1986; Prøsch-Danielsen &
Simonsen 2000; Holden et al. 2007; Hjelle et al. 2010).
Mild winters allowed outdoor grazing on the evergreen
C. vulgaris year round, and traditional management
included regular burning to improve the pasture value of
C. vulgaris, as well as cutting of vegetation and peat for fod-
der and fuel. This land-use regime effectively prevented
forest regeneration and maintained the dynamics of the
heathland ecosystem (Webb 1998; Kvamme et al. 2004).
At their greatest extent, coastal heathlands covered sev-
eral million hectares (Webb 1998), but more than 80% of
the European heathlands have been lost over the last
150 yrs and heathlands are now classified as threatened
(UK BAP 1999; Kvamme et al. 2004; Lindgaard & Henrik-
sen 2011). Land conversion, such as commercial forest
plantations or pasture improvements, and airborne nitro-
gen deposition contribute to degeneration or loss of coastal
heathlands (van Breemen & van Dijk 1988; Webb 1998;
Holden et al. 2007), but a key driver is abandonment of
the traditional land-use regime (Webb 1998; Plieninger
et al. 2006). Abandoned heathlands are invaded by native
tree species such as birch (Betula pubescens and B. pendula)
and Scots pine (Pinus sylvestris L.; Mitchell et al. 1999), and
the resulting succession has strong impacts on understorey
species composition and diversity (Mitchell et al. 1997;
Curt et al. 2003). As the canopy closes, the characteristic
heathland flora may decline or be gradually replaced by
more shade-tolerant forest species (Hester et al. 1991;
Sørensen & Tybirk 2000).
Invasion by introduced coniferous tree species can
accelerate successional rates in heathlands (Christensen &
Johnsen 2001; Catling & Carbyn 2005), and may decrease
native species diversity (Richardson 1998; Higgins et al.
1999). Sitka spruce (Picea sitchensis (Bong.) Carr.), native to
the west coast of North America, is widely planted
throughout Europe (Carrillo-Gavilan & Vila 2010). It starts
reproducing early (5–20 yrs) (Artsdatabanken 2009;
Vikane unpublished data) and can thus quickly invade
abandoned heathlands. Such invasions are currently
observed in Norway (Maren & Nilsen 2008; Thorvaldsen
2011). Sitka spruce forms dense stands that are likely to
have stronger effects on understorey species composition
and diversity than the more open native Scots pine stands
(Ferris et al. 2000), but we are not aware of any studies
that have investigated the effect of invasion of Sitka spruce
on coastal heathland vegetation.
This study investigates the impact of invasion by two
conifers, the introduced Sitka spruce and the native Scots
pine, on plant species richness and composition of coastal
heathlands in four sites in western Norway. Tree invasion
is still at an early stage in our study area, with young coni-
fers scattered within otherwise open heathlands, creating
a “patchy” environment. Single tree canopies significantly
impact environmental conditions, such as light, soil nutri-
ents and moisture conditions, and thus affect species
composition (Zinke 1962; Vetaas 1992; Wallrup et al.
2006; Jager et al. 2007), and the impact of single trees
may be used to forecast consequences of future canopy
closure (Jager et al. 2007). In the early stage of invasion,
effects operate at fine spatial scales (Zinke 1962; Amiotti
et al. 2000). We therefore study the fine-scale effects of
single coniferous trees on heathland vegetation. This
study was performed at 60 °N, where the southern sides
of the coniferous tree canopies receive more solar radia-
tion than the northern side, resulting in differences in
light, temperature and soil moisture conditions (Geier-
Hayes et al. 1995; Hutchinson et al. 1999). To assess the
effects of this fine-scale variability we laid out transects
oriented north and south from the stem of each target
tree. We ask: (1) what are the effects of coniferous inva-
sion on plant species composition and richness of coastal
heathlands; and (2) are there differences in the effects
depending on whether the heathlands are invaded by
introduced or native coniferous tree species? In particular,
we assess how far along the gradient from the tree stems
the canopy effects can be detected, and we explore how
these effects are related to local environmental effects of
the canopy by comparing responses along the north- and
south-oriented transects.
Methods
Study area
Four study sites were selected within coastal heathlands in
western Norway (Fig. 1, Table 1). The sites are within the
boreonemoral bioclimatic vegetation zone and the hyper-
oceanic vegetation section (Moen 1998). The climate is
oceanic with mild winters (average temperatures
of the coldest mo>1 °C), high precipitation
(1300–2000 mm�yr�1) and a relatively long growing sea-
son (more than 200 d>5 °C; DNMI 2011). The prevailing
wind direction is from south to southeast (Flesland meteo-
rological station near sites 1–3; DNMI 2011) and south to
west (Skudenes II meteorological station, near site 4;
DNMI 2011). Bedrock consists of dioritic and granitic
gneisses (site 1 and 3), diorite and monzodiorite (site 2)
andmonzonite (site 4; NGU 2012).
The investigated sites have characteristic coastal Calluna
heathland vegetation (Fremstad 1997) with a dense field
layer dominated by Calluna vulgaris and characterized by
ericaceous species (e.g. Vaccinium spp., Empetrum nigrum
and Erica tetralix) and graminoids (e.g. Molinia caerulea,
Eriophorum vaginatum, Trichophorum cespitosum, Avenella
flexuosa and Carex spp.). Juniperus communis is the most
Journal of Vegetation Science2 Doi: 10.1111/jvs.12010© 2012 International Association for Vegetation Science
Effects of introduced versus native conifer invasion H.I.Saure et al.

common species of a scanty shrub layer. Bryophyte cover
is high, and is dominated by Hypnum cupressiforme/jutlandi-
cum (coll.), Pleurozium schreberi, Hylocomium splendens,
Sphagnum species, Rhytidiadelphus loreus and R. squarrosus.
The study sites showed no sign of recent burning or cut-
ting, and grazing pressure is low; outwintered sheep were
present in site 2 and there were signs of low-intensity deer
grazing in site 3. Natural woodlands are present in the sur-
rounding landscape, and Appendix S1 provides informa-
tion on the local woodland species pool (municipalities
near the city of Bergen, western Norway). At all sites, Sitka
spruce is spreading into heathlands from adjacent planta-
tions (large plantations or narrow strips of shelter plant-
ing). Mean height of Sitka spruce trees in the heathlands
was 4.6 m, and mean age of fertile Sitka spruce (i.e. carry-
ing cones) was 14.7 yrs. Seedling density in a 100-m zone
bordering plantations was 0.55 individuals m�2 (Vikane,
unpubl.). Seedlings of other shrubs and trees, e.g. Sorbus
acuparia and native Scots pine, were also recorded, but at
much lower densities than Sitka spruce.
Sampling
Fieldwork was carried out in July–September 2009.
Vegetation was sampled within 5 9 5 m blocks centred on
2–4-m tall target trees of Sitka spruce or Scots pine in Callu-
na wet heath vegetation (Fremstad 1997), avoiding
slopes>12°, bare rock and wet depressions, and with dis-
tance between target coniferous trees >10 m. These criteria
resulted in selection of blocks with tree cover of ca. 15%
and where the target coniferous tree was the only
tree>2 m.
Within each block, we placed three 2-m transects; two
canopy transects and one open heathland control transect
(hereafter referred toas control).Canopy transectswere laid
out 180 °S and 360 °N from the target coniferous tree stem
(hereafter referred to as the south and north transects). The
control was placed parallell to the canopy transects, with
the mid-point in line with the stem and at a distance from
the edge of the target coniferous tree canopy of at least
0.5 m. Controlswere randomly allocated to the east orwest
side of the target tree, unless one of the sideswas unsuitable
according to the rejection criteria givenabove.
Transects were divided into eight sample plots of
0.25 9 0.25 m, which were analysed for vascular plant
and bryophyte abundance (recorded as frequency in four
subplots). Similarly, we recorded cover of the target conif-
erous tree in each plot. Target coniferous trees varied
somewhat in size, but all canopy transects ran at least
0.5 m (two sample plots) beyond the target coniferous tree
canopy. This transect length was assumed to be long
enough for the outer sample plots to be unaffected by the
target tree canopy.
We sampled 38 blocks with Sitka spruce as the target
coniferous tree, and 20 blocks with Scots pine, which
resulted in 174 transects and 1392 transect plots.
Table 1. Study sites on large, coastal islands of western Norway.
Site 1 Site 2 Site 3 Site 4
Local name Palsvarden Hille Stura Kvalheimsneset
Municipality Karmøy Austevoll Øygarden Radøy
County Rogaland Hordaland Hordaland Hordaland
Latitude, longitude 59 °10 ′N, 5 °13 ′E 60 °7 ′N, 5 °4 ′E 60 °37 ′N, 4 °50 ′E 60 °41 ′N, 4 °54 ′E
Climate station Skudenes II & III Storebø Tjelstø Manger
Mean annual temperature (°C) 7.6 7.0 7.2 7.2
Mean annual precipitation (mm) 1295 1875 1975 1540
Area (ha) 16.95 10.87 2.97 5.58
Currently grazed (+) (“+”) (“+”)
Mean age fertile sitka spruce (yr/SD) 18.0 (4.7) 12.9 (3.5) 12.4 (3.0) 12.0 (2.1)
Blocks sitka spruce/Scots pine (number) (10/5) (10/5) (8/5) (10/5)
Fig. 1. Maps showing former distribution (ca. 1850) of the European
heathlands (Diemont & Kaland, in Haaland 2002) and the study sites within
the coastal heathland zone (1–4) in western Norway. Table 1 provides
further information on the investigated areas.
Journal of Vegetation ScienceDoi: 10.1111/jvs.12010© 2012 International Association for Vegetation Science 3
H.I.Saure et al. Effects of introduced versus native conifer invasion

Data analysis
A detrended correspondence analysis (DCA; Hill & Gauch
1980) with down-weighting of rare species showed that
the first gradient was rather long (3.9 SD), and constrained
correspondence analysis (CCA) was chosen to investigate
relationships between species composition and environ-
ment. We used partial CCA in conjunction with principal
response curves (PRC; van den Brink & ter Braak 1999) to
test and quantify the effects of the target trees on the
heathland vegetation. PRC was originally developed for
analysing repeated-measures design experiments, and is
commonly used in ecological research (see e.g. Frampton
et al. 2000; Savadogo et al. 2009; Nakatani et al. 2011).
We use PRC in a spatial context, i.e. comparing commu-
nity composition along spatial transects, rather than over
time, following the general procedures of Heegaard &
Vandvik (2004), adapted for our data as follows: to analyse
spatial data using the PRC approach, we need two sets of
categorical variables, one describing the spatial gradient (in
our case the eight distances along the trasects), and one
describing the “controls” and “treatments” we want to
compare (in our case control transects vs. the four different
categories of canopy transects represented by tree species
9 aspect). In the spatial PRC analysis, blocks and distance
classes are coded as “covariables”, and the interactions
between distance and each of the treatment variables are
coded as “environmental variables”. The resulting PRC
analysis thus estimates the compositional differences of
each of the treatments from the control transects at each of
the eight distances from target tree stems. Within this gen-
eral framework, effects of different coniferous tree species
were tested by coding canopy transect distances under
each of Sitka spruce and Scots pine as different treatments
and partialling out the main effect of canopy transects.
Effect of aspect was tested by coding canopy transect dis-
tances for each aspect (north and south) as treatment and
partialling out the main effect of canopy transect distance
and blocks. Finally, the interactive effect of coniferous tree
species and aspect was tested by coding canopy transect
distances for each combination of aspect (north and south)
and coniferous tree species (Sitka spruce and Scots pine) as
treatments, and partialling out the main effects of canopy
transects, coniferous tree species, aspect and blocks.
Important features of the method are the PRC diagram
and the species scores. The coding used in PRC standardizes
the control to be zero-valued for all times, i.e. a horizontal
line in the PRC diagram. The PRC scores for each of the
“treatments” through time represent compositional differ-
ences from the control, and the species scores reflect the
influence of particular species on the overall community
response described by the PRC scores through time. Species
with high positive scores are positively correlated, species
with negative scores respond oppositely, and species with
near zero scores are indifferent to the effects reflected by
the PRC axes (Microcomputer Power, Ithaca, NY, USA).
The PRC diagram,which represents the total effect of conif-
erous trees (including differences betweeen tree species
and aspect) was tested by coding each combination of
coniferous tree species and aspect along the canopy tran-
sect as treatments, while partialling out the effect of blocks.
A generalized linear mixedmodel (GLMM) tested differ-
ences in the dependent variable, species richness (species
number per plot, including vascular plants and bryo-
phytes), in response to the fixed factorial effects of trees vs.
controls, native vs. introduced conifers, north vs south
canopy transects, and distance to stem. The GLMM was
run with the function lmer (package lme4) and fitted by
the Laplace approximation. Blocks were included as ran-
dom effects, and the model was simplified by backwards
elimination starting from the full model with three-way
interactions. A Poisson distribution of errors was assumed
for the species richness response variable, thus the GLMM
was runwith a log-link function. Following the PRC analy-
ses, the effects were plotted as contrasts between canopy
transects and their respective controls with distance to
stem.Welch two-sample t-tests were run post-hoc to assess
the significance of differences in species richness between
canopy transects and controls, and the different transect
categories (tree species, aspect) at each specific distance
from stems.
The number of species unique to the canopy transects
(Sitka spruce north transect, Sitka spruce south
transect, Scots pine north transect and Scots pine south
transect) and their respective controls are displayed to indi-
cate howmany species are likely to have established or dis-
appeared after development of coniferous tree canopy.
Ordinations were run using CANOCO 4.5 for Windows.
All other analyses were run by using R (R Foundation for
Statistical Computing, Vienna, AT).
Results
Species composition
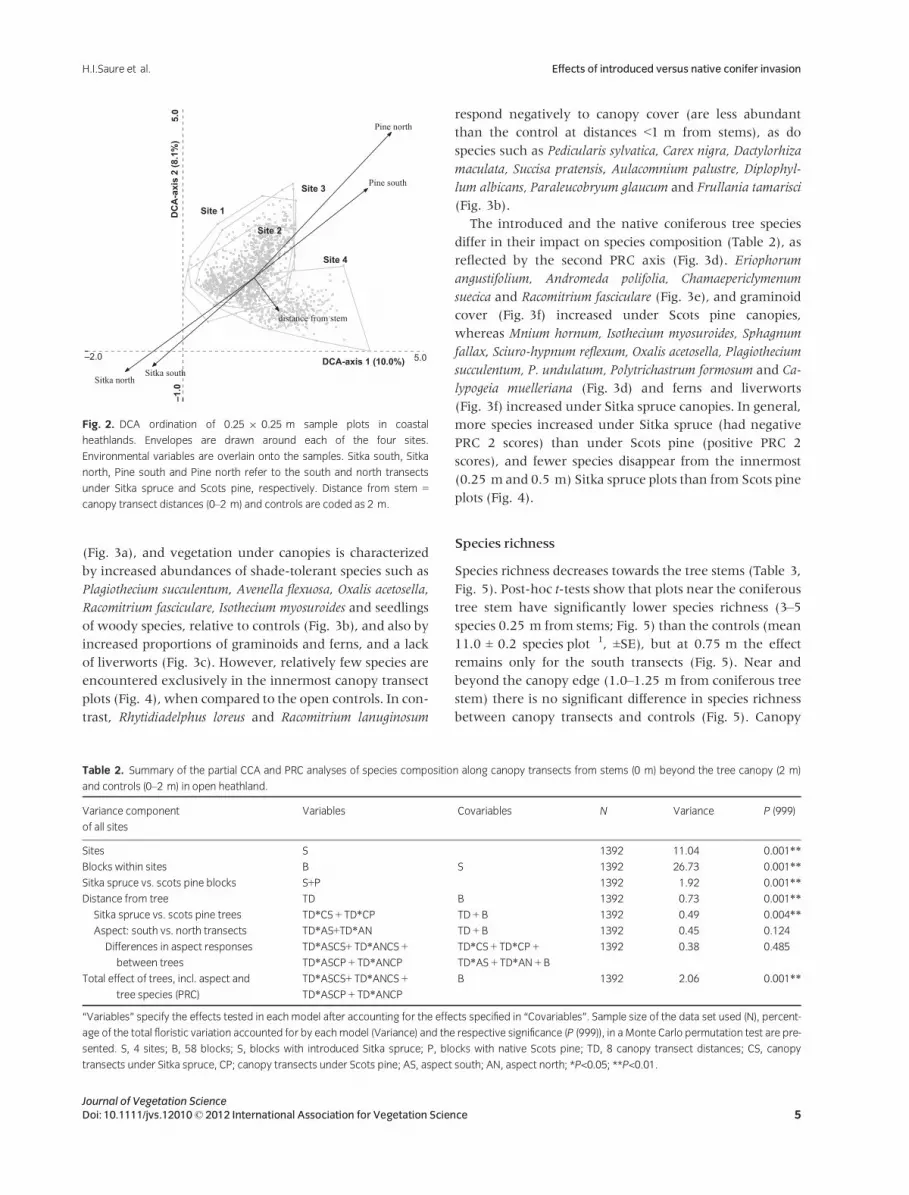
The DCA ordination displays a continuous variation in spe-
cies composition among the sites, blocks and plots in our
data (Fig. 2), and indicates that species composition varies
both between and within sites. Within each site, both
coniferous tree species and distance from stem seem to
impact species composition, whereas south and north tran-
sects point in the same direction for each coniferous tree
species, indicating relatively low impact of aspect on spe-
cies composition (Fig. 2), which is also confirmed in the
PRC analyses (Table 2).
The overall impact of coniferous tree canopies on the
heathland community is reflected along the first PRC axis
Journal of Vegetation Science4 Doi: 10.1111/jvs.12010© 2012 International Association for Vegetation Science
Effects of introduced versus native conifer invasion H.I.Saure et al.
(Fig. 3a), and vegetation under canopies is characterized
by increased abundances of shade-tolerant species such as
Plagiothecium succulentum, Avenella flexuosa, Oxalis acetosella,
Racomitrium fasciculare, Isothecium myosuroides and seedlings
of woody species, relative to controls (Fig. 3b), and also by
increased proportions of graminoids and ferns, and a lack
of liverworts (Fig. 3c). However, relatively few species are
encountered exclusively in the innermost canopy transect
plots (Fig. 4), when compared to the open controls. In con-
trast, Rhytidiadelphus loreus and Racomitrium lanuginosum
respond negatively to canopy cover (are less abundant
than the control at distances <1 m from stems), as do
species such as Pedicularis sylvatica, Carex nigra, Dactylorhiza
maculata, Succisa pratensis, Aulacomnium palustre, Diplophyl-
lum albicans, Paraleucobryum glaucum and Frullania tamarisci
(Fig. 3b).
The introduced and the native coniferous tree species
differ in their impact on species composition (Table 2), as
reflected by the second PRC axis (Fig. 3d). Eriophorum
angustifolium, Andromeda polifolia, Chamaepericlymenum
suecica and Racomitrium fasciculare (Fig. 3e), and graminoid
cover (Fig. 3f) increased under Scots pine canopies,
whereas Mnium hornum, Isothecium myosuroides, Sphagnum
fallax, Sciuro-hypnum reflexum, Oxalis acetosella, Plagiothecium
succulentum, P. undulatum, Polytrichastrum formosum and Ca-
lypogeia muelleriana (Fig. 3d) and ferns and liverworts
(Fig. 3f) increased under Sitka spruce canopies. In general,
more species increased under Sitka spruce (had negative
PRC 2 scores) than under Scots pine (positive PRC 2
scores), and fewer species disappear from the innermost
(0.25 m and 0.5 m) Sitka spruce plots than from Scots pine
plots (Fig. 4).
Species richness
Species richness decreases towards the tree stems (Table 3,
Fig. 5). Post-hoc t-tests show that plots near the coniferous
tree stem have significantly lower species richness (3–5
species 0.25 m from stems; Fig. 5) than the controls (mean
11.0 ± 0.2 species�plot�1, ±SE), but at 0.75 m the effect
remains only for the south transects (Fig. 5). Near and
beyond the canopy edge (1.0–1.25 m from coniferous tree
stem) there is no significant difference in species richness
between canopy transects and controls (Fig. 5). Canopy
Table 2. Summary of the partial CCA and PRC analyses of species composition along canopy transects from stems (0 m) beyond the tree canopy (2 m)
and controls (0–2 m) in open heathland.
Variance component
of all sites
Variables Covariables N Variance P (999)
Sites S 1392 11.04 0.001**
Blocks within sites B S 1392 26.73 0.001**
Sitka spruce vs. scots pine blocks S+P 1392 1.92 0.001**
Distance from tree TD B 1392 0.73 0.001**
Sitka spruce vs. scots pine trees TD*CS + TD*CP TD + B 1392 0.49 0.004**
Aspect: south vs. north transects TD*AS+TD*AN TD + B 1392 0.45 0.124
Differences in aspect responses
between trees
TD*ASCS+ TD*ANCS +
TD*ASCP + TD*ANCP
TD*CS + TD*CP +
TD*AS + TD*AN + B
1392 0.38 0.485
Total effect of trees, incl. aspect and
tree species (PRC)
TD*ASCS+ TD*ANCS +
TD*ASCP + TD*ANCP
B 1392 2.06 0.001**
“Variables” specify the effects tested in each model after accounting for the effects specified in “Covariables”. Sample size of the data set used (N), percent-
age of the total floristic variation accounted for by eachmodel (Variance) and the respective significance (P (999)), in a Monte Carlo permutation test are pre-
sented. S, 4 sites; B, 58 blocks; S, blocks with introduced Sitka spruce; P, blocks with native Scots pine; TD, 8 canopy transect distances; CS, canopy
transects under Sitka spruce, CP; canopy transects under Scots pine; AS, aspect south; AN, aspect north; *P<0.05; **P<0.01.
–2.0 5.0
–1.0
5.0
distance from stem
Sitka southSitka north
Pine south
Pine north
Site 1
Site 3
Site 2
Site 4
DCA-axis 1 (10.0%)
DC
A-a
xis
2 (8
.1%
)
Fig. 2. DCA ordination of 0.25 9 0.25 m sample plots in coastal
heathlands. Envelopes are drawn around each of the four sites.
Environmental variables are overlain onto the samples. Sitka south, Sitka
north, Pine south and Pine north refer to the south and north transects
under Sitka spruce and Scots pine, respectively. Distance from stem =
canopy transect distances (0–2 m) and controls are coded as 2 m.
Journal of Vegetation ScienceDoi: 10.1111/jvs.12010© 2012 International Association for Vegetation Science 5
H.I.Saure et al. Effects of introduced versus native conifer invasion

effects on species richness were more pronounced in the
south transects than in north transects (Table 3); south
transects have lower species richness than north transects
at all distances (often >1.0 species; Table 3), and the nega-
tive effect of canopy on species richness also persisted
somewhat further from the stems in the south transects
(Fig. 5). There were no differences in species richness
between vegetation under Sitka spruce and Scots pine can-
opies (Table 3).
Discussion
We found that distinct microcommunities had emerged
beneath relatively young and isolated coniferous trees
PRC-1 Positive species scores
9%6%
6%
9%
26%
44%
0%
PRC-1 Negative species scores
1%4%
8%
11%
42%
32%
2%
PRC-2 Positive species scores0%
5%
2%
12%
21%
43%
17%
PRC-2 Negative species scores
6% 6%
2%
13%
13%
37%
23%
Ferns Woody Low shrub Herbs Graminoids Mosses Liverworts
(a) (b) (c)
(d) (e) (f)
Fig. 3. PRC diagram (a & d) visualising the total effect of trees on species composition in coastal heathlands, relative to controls, along a transect from the
stems (0 m) and beyond the canopy (2 m). Species scores of the most strongly affected species (species scores >|0.2|) are shown to the right (b & e). Rare
species (<8 occurrences) and indifferent species are not displayed. The relative abundance of plant groups (c & f) along PRC axes 1 and 2 is displayed,
counting all species with scores >|0.1|. Plant group ‘ferns’ also include Lycopodiaceae.
Journal of Vegetation Science6 Doi: 10.1111/jvs.12010© 2012 International Association for Vegetation Science
Effects of introduced versus native conifer invasion H.I.Saure et al.

(2–4-m tall) that have invaded coastal heathlands. Canopy
effects differed between the introduced and the native
coniferous tree species, and between the north and south
side of the trees. Whereas coniferous tree species had
strong effects on understorey species composition, transect
orientation primarily affected species richness.
Community dynamics beneath coniferous canopies
The pronounced changes in vegetation beneath canopies
of the invading conifers, relative to open heaths, are seen
mainly as an increased occurrence and abundance of
shade-tolerant forest species and a reduction in light-
demanding heathland species. Species sorting (sensu
Leibold et al. 2004) along the environmental gradient
imposed by the developing canopies is likely to be the
dominant process in the emergence of distinct vegetation
beneath the invading coniferous tree. Shading is known to
be an important explanatory factor for species richness
reductions towards tree stems (Jager et al. 2007), and even
small single trees, like the conifers in this study, may
induce shade light gradients on heathland vegetation
(Piessens et al. 2006). Spruce canopies are known to be
denser than Scots pine canopies (Hill 1979; Ferris et al.
2000), and the degree of species sorting (i.e. the degree of
environmental control of spatial dynamics) is clearly stron-
ger beneath Sitka spruce than beneath Scots pine. Thus,
contrasting microcommunities have developed beneath
the two conifers; species associated with Scots pine are
light-demanding heathland species (e.g. Eriophorum angus-
tifolium and Andromeda polifolia), while species correlated
with Sitka spruce canopies are typical, shade-tolerant for-
est species (e.g. Sciuro-hypnum reflexum, Oxalis acetosella,
Isotheciummyosuroides and Plagiothecium undulatum).
Species richness decreased beneath canopies of the
invading conifers. This corresponds to other studies where
species richness is reduced in formerly treeless habitats
after invasion of introduced tree species (Richardson et al.
Sitk
a sp
ruce
Northern transects
0
5
10
15
20
25
Can
opy
tran
sect
s
25
20
15
10
5
0
Con
trol
sSouthern transects
0
5
10
15
20
25
30
30
25
20
15
10
5
0
Scot
s pi
ne
0
5
10
15
20
25
30
35
Can
opy
tran
sect
s
35
3025201510
5
0
0.25 0.5 0.75 1 1.25 1.5 1.75 2
Con
trol
s
05
10152025303540
4035302520151050
0.25 0.5 0.75 1 1.25 1.5 1.75 2
Distance from stem (m)Distance from stem (m)
Fig. 4. Number of species (vascular plants and bryophytes) unique to canopy transects or controls at different distances along the transects.
Journal of Vegetation ScienceDoi: 10.1111/jvs.12010© 2012 International Association for Vegetation Science 7
H.I.Saure et al. Effects of introduced versus native conifer invasion

1989; Jager et al. 2007), but Dickie et al. (2011) found
increasing species richness in grasslands that were invaded
by an introduced and a native tree species. In plantation
forests, species richness has been found to be lower under
Norway spruce (Picea abies) than under Scots pine and
birch (Wallrup et al. (2006). Interestingly, we found no
difference in subcanopy species richness between the two
coniferous tree species. This can partially be accounted for
by increased colonization by forest species under Sitka
spruce, indicating that dispersal conditions must be good
within the heathland landscape. The importance of dis-
persal is supported by the the higher proportions of forest
ferns and liverworts under Sitka spruce, as the small and
numerous propagules may give these groups a dispersal
advantage relative to other forest species (Tryon 1970; Ha-
jek et al. 2011).
South-facing aspect seems to enhance the canopy effect
on species richness as significant differences from controls
extend further away from the stems in the south transects.
Other studies from northern latitudes frequently report
increased species richness in southern canopy transects
(Lopez-Pintor et al. 2006) and southern forest edges
(Honnay et al. 2002), which is typically attributed to
higher levels of solar radiation and thus warmer and drier
microclimates on the south-facing sides of the trees or
forests at these latitudes (Geier-Hayes et al. 1995). In
contrast, we found lower species richness on the south side
of the coniferous trees. This may be accounted for by the
cool and humid regional climate, resulting in a regional
species pool that is dominated by species that are adapted
to the cooler and more humid environments found on the
north side of the coniferous trees (Vevle 2000; Lid & Lid
2005; Atherton et al. 2010).
From a landscape perspective, the vegetation under
these invading conifers may be regarded as small, develop-
ing, but already quite compositionally distinct “forest un-
derstorey patches” within the matrix of coastal heathlands,
or as forest understorey metacommunites (sensu Leibold
et al. 2004). Similarly, small-scale metacommunities in
boreal forest ecosystems has been documented on rock
boulders that host a set of cryptogam species not found in
the forest matrix (Virtanen & Oksanen 2007). The devel-
opment of these distinct metacommunities is driven by
strong species sorting combined with dispersal of shade-
tolerant forest specialist species (especially bryophytes)
from nearby forest patches (see above), but could poten-
tially have been cancelled out by strong mass effects (sensu
Shmida & Wilson 1985; Leibold et al. 2004), as the propa-
gule rain from the surrounding heathland vegetation is
likely to be substantial. Evidently, the species sorting
resulting from microenvironmental and other effects of
the young conifers are too strong to be obscured by propa-
gule rain from the heathland matrix. Mass effects may still
be buffering the impact of the invading conifers however
(see also Shmida & Ellner 1984; Lichstein & Pacala 2011;
Alexander et al. 2012), which would imply that the rate of
compositional change under trees may increase as the tree
density in the landscape increases and the heathland
matrix becomes relatively less abundant.
Implications for heathland vegetation following conifer
invasion
Scots pine is the only native conifer in western Norway
(Gjerde 1993), thus the spread of Sitka spruce from planta-
Table 3. Generalized linear mixed model testing the fixed effects of tree
invasion, tree species, canopy transect aspect and distance to stem on
total species richness.
Estimate SE z-value P-value
Intercept 2.39 0.05 48.01 <0.001***
Distance along transects �0.01 0.02 �0.41 0.69 ns
Sitka spruce (vs. scots pine) 0.01 0.05 0.15 0.88 ns
North transects
(vs. control)
�0.27 0.05 �5.94 <0.001***
South transects
(vs. control)
�0.43 0.05 �9.22 <0.001***
Distance along
north transects
0.20 0.04 5.58 <0.001***
Distance along
south transects
0.25 0.04 7.04 <0.001***
Blocks were included as random variables and a Poisson distribution of
errors was assumed. Y = number of species, D = distance along tran-
sects, tree = introduced Sitka spruce and native Scots pine, treatment
(treat) = south transects, north transects and open controls, B = blocks
each containing a target tree and the associated treatment transects. The
model was simplified by backward elimination Y~D*tree*treat + (1|B).
Bold highlights significant effects.
–6.00
–5.00
–4.00
–3.00
–2.00
–1.00
0.00
1.00
2.00
0.25 0.5 0.75 1 1.25 1.5 1.75 2
Distance from stem (m)
Mea
n sp
ecie
s no
. dev
ianc
e fr
om c
ontr
ol
Sitka southSitka northPine southPine northControl
Fig. 5. Species richness in canopy transects, shown as difference in mean
species number at each transect type and distance, relative to the
controls. Controls are not significantly different from each other, and
overall mean control species number is 11. Large symbols have
significantly different species richness (P < 0.05) than their respective
controls. See Table 3 for tests of significance for the different main effects
and interactions.
Journal of Vegetation Science8 Doi: 10.1111/jvs.12010© 2012 International Association for Vegetation Science
Effects of introduced versus native conifer invasion H.I.Saure et al.

tions means that a “new” type of conifer with a denser and
lower canopy has entered the heathlands. Sitka spruce
invasion into semi-natural landscapes, such as the coastal
heathlands investigated in this study, is too recent to have
resulted in closed-canopy Sitka spruce forests (Peterken
2001).We therefore do not knowhow the introduced Sitka
spruce will impact characteristic heathland plant species
and diversity as forest succession proceeds, but impacts by
single trees at an early invasion stage can be used to predict
consequences of later invasion stages (Jager et al. 2007).
The establishment of shade-tolerant forest species,
which was more pronounced beneath the introduced Sitka
spruce than beneath the native Scots pine, is at the cost of
abundance or even presence of typical heathland species
(Hester et al. 1991). Dense Sitka spruce plantations rapidly
out-shade most vascular plants, but ferns and bryophytes
survive the low-light regime at varying levels of cover and
diversity (Hill 1979; French et al. 2008). After canopy
closure, the understorey within naturalized Sitka spruce
forest is therefore likely to show much less resemblance to
the coastal heathlands.
Old-growth, natural Sitka spruce forests along the Paci-
fic coast of North America have a well-developed under-
storey, with shrubs, ferns, herbs and a very well-developed
bryophyte layer (Peterson et al. 1997). Similarly, thinning
of Sitka spruce plantations allows some colonization of
woodland species (Wallace & Good 1995; French et al.
2008). As naturalized Sitka spruce forests in western Nor-
way mature, probably 50–120 yrs after canopy closure
(Banner et al. 1989; Ferris et al. 2000), the stands may
therefore become more structurally diverse, and are likely
to facilitate a more diverse understorey vegetation and
more native tree species than are found within current
Sitka spruce plantations, reflecting the local species pool of
woodland species.
In the investigated heathlands, vegetation beneath
native Scots pine was characterized by open heathland
species, indicating that future changes in understorey com-
position might be less dramatic in Scots pine forests com-
pared to Sitka spruce forests. This is supported by other
studies that report similarities in understorey species com-
position between heathland and Scots pine forests (Peter-
ken 2001; Curt et al. 2003), and suggests that heathland
invaded by Scots pine may have a higher restoration
potential than those invaded by Sitka spruce.
The West European coastal heathlands are important
for conservation (Council of Europe 2000), not only for
their characteristic biodiversity, but also for their historical
and aesthetic values (Norwegian Ministry of the Environ-
ment 2006–2007). We have shown that the invader
species makes a difference to vegetation development after
coniferous tree invasion in the coastal heathlands. To facil-
itate future restoration of coastal heathlands, Sitka spruce
invasion should be controlled in areas of potential
restoration interest. Short generation time (5–20 yrs) and
high seed production (Richardson & Rejmanek 2004)
means that Sitka spruce has a considerable invasion poten-
tial. Removal of adjacent Sitka spruce plantations and
invading individuals within the coastal heathlands will
therefore be necessary to control its spread.
Acknowledgements
We thank J.H. Vikane for sharing data on Sitka spruce
populations, S. Vogler, R. Smith, L. Hagenlund, R. Ramirez
and S. Rumpf for fieldwork and technical assistance; J H.
Vikane, J.-A. Grytnes and J. Kapfer for useful advice on
data analyses; B. Helle, C. Aasebø and L.G. Velle for art-
work and C. Jenks for linguistic correction. The project
was supported by NLA University College and the Norwe-
gian Research Council, project no.184099.
References
Alexander, H.M., Foster, B.L., Ballantyne, F., Collins, C.D., An-
tonovics, J. & Holt, R.D. 2012. Metapopulations and meta-
communities: combining spatial and temporal perspectives
in plant ecology. Journal of Ecology 100: 88–103.
Amiotti, N.M., Zalba, P., Sanchez, L.F. & Peinemann, N. 2000.
The impact of single trees on properties of loess-derived
grassland soils in Argentina. Ecology 81: 3283–3290.
Artsdatabanken 2009. Sitkagran (Picea sitchensis). Artsdataban-
kens faktaark ISSN 1504-9140. pp. 3.
Atherton, I., Bosanquet, S. & Lawley, M. 2010. Mosses and liver-
worts of Britain and Ireland – a field guide. British Bryological
Society, Plymouth, UK.
Banner, A.J., Pojar, J., Schwab, J.W. & Trowbridge, R. 1989.
Vegetation and soils of the Queen Charlotte Islands: recent
impacts of development. In: Schudder, G.E. & Gessler, N.
(eds.) The outer shores. pp. 261–279. Queen Charlotte Islands
MuseumPress, Skidegate, BC, CA.
van Breemen, N. & van Dijk, H.F.G. 1988. Ecosystem effects of
atmospheric deposition of nitrogen in the Netherlands. Envi-
ronmental Pollution 54: 249–274.
van den Brink, P.J. & ter Braak, C.J.F. 1999. Principal response
curves: analysis of time-dependent multivariate responses of
biological community to stress. Environmental Toxicology and
Chemistry 18: 138–148.
Carrillo-Gavilan, M.A. & Vila, M. 2010. Little evidence of inva-
sion by alien conifers in Europe. Diversity and Distributions 16:
203–213.
Catling, P.M. & Carbyn, S. 2005. Invasive Scots pine, Pinus sylves-
tris, replacing Corema, Corema conradii, heathland in the
Annapolis Valley, Nova Scotia. Canadian Field-Naturalist 119:
237–244.
Christensen, S. & Johnsen, I. 2001. The lichen-rich coastal
heath vegetation on the isle of Anholt, Denmark – con-
Journal of Vegetation ScienceDoi: 10.1111/jvs.12010© 2012 International Association for Vegetation Science 9
H.I.Saure et al. Effects of introduced versus native conifer invasion

servation and management. Journal of Coastal Conservation
7: 13–22.
Council of Europe 2000. European landscape convention. Stras-
bourg, FR.
Curt, T., Prevosto, B., Klesczewski, M. & Lepart, L. 2003. Post-
grazing Scots pine colonization of mid-elevation heathlands:
population structure, impact on vegetation composition and
diversity. Annals of Forest Science 60: 711–724.
Dickie, I.A., Yeates, G.W., St. John,M.G., Stevenson, B.A., Scott,
J.T., Rillig, M.C., Peltzer, D.A., Orwin, K.H., Kirschbaum, M.
U.F., Hunt, J.E., Burrows, L.E., Barbour, M.M. & Aislabie, J.
2011. Ecosystem service and biodiversity trade-offs in two
woody successions. Journal of Applied Ecology, 48: 926–934.
DNMI 2011. http://eklima.met.no.
Ferris, R., Peace, A.J., Humphrey, J.W. & Broome, A.C. 2000.
Relationships between vegetation, site type and stand struc-
ture in coniferous plantations in Britain. Forest Ecology and
Management 136: 35–51.
Frampton, G.K., van den Brink, P.J. & Gould, P.J.L. 2000.
Effects of spring drought and irrigation on farmland ar-
thropods in southern Britain. Journal of Applied Ecology
37: 865–883.
Fremstad, E. 1997. Vegetasjonstyper i Norge. NINA-temahefte 12:
1–279.
French, L.J., Smith, G.F., Kelly, D.L., Mitchell, F.J.G., O’Don-
oghue, S., Iremonger, S.F. & McKee, A.-M. 2008. Ground
flora communities in temperate oceanic plantation forests
and the influence of silvicultural, geographic and edaphic
factors. Forest Ecology and Management 255: 476–494.
Geier-Hayes, K., Hayes, M.A. & Basford, D.D. 1995. Determining
individual tree shade length: a guide for silviculturists. Gen-
eral technical report, United States Department of Agricul-
ture, Forest Service, Intermountain Research Station.
Gimingham, C.H. 1961. North European heath communities: a
‘network of variation’. Journal of Ecology 49: 655–694.
Gjerde, I. 1993. Skogbruk og fauna pa Vestlandet: betydningen av tres-
lagsskifte for forekomst og fordeling av skogshabitat. Skogforsk,
Oslo, NO.
Haaland, S. 2002. Fem tusen ar med flammer. Det europeiske lyng-
heilandskapet. Vigmostad & Bjørke AS, NO.
Hajek, M., Rolecek, J., Cottenie, K., Kintrova, K., Horsak, M.,
Poulickova, A., Hajkova, P., Frankova, M. & Dite, D. 2011.
Environmental and spatial controls of biotic assemblages in a
discrete semi-terrestrial habitat: comparison of organisms
with different dispersal abilities sampled in the same plots.
Journal of Biogeography 38: 1683–1693.
Heegaard, E. & Vandvik, V. 2004. Climate change affects the out-
come of competitive interactions – an application of principal
response curves.Oecologia 139: 459–466.
Hester, A.J., Miles, J. & Gimingham, C.H. 1991. Succession from
heather moorland to birch woodland. I. Experimental alter-
ation of specific environmental conditions in the field. Jour-
nal of Ecology 79: 303–315.
Higgins, S.I., Richardson, D.M., Cowling, R.M. & Trinder-Smith,
T.H. 1999. Predicting the landscape-scale distribution of alien
plants and their threat to plant diversity. Conservation Biology
13: 303–313.
Hill, M.O. 1979. The development of a flora in even-aged planta-
tions.In: Ford, E.D., Malcolm, D.C. & Atterson, J. (eds.) The
Ecology of even-aged forest plantations. pp. 175–192. Institute of
Terrestrial Ecology, Cambridge, UK.
Hill, M.O. & Gauch, H.G. 1980. Detrended correspondence anal-
ysis: an improved ordination technique. Plant Ecology 42: 47–
58.
Hill, M.O., Bell, N., Bruggeman-Nannenga, M.A., Brugues,
M., Cano, M.J., Enroth, J., Flatberg, K.I., Frahm, J.P.,
Gallego, M.T., Garilleti, R., Guerra, J., Hedenas, L., Holy-
oak, D.T., Hyvonen, Ignatov, M.S., Lara, F., Mazimpaka,
V., Munoz, J. & Soderstrom, L. 2006. An annotated
checklist of the mosses of Europe and Macaronesia. Jour-
nal of Bryology, 28: 198–267.
Hjelle, K.L., Halvorsen, L.S. & Overland, A. 2010. Heathland
development and relationship between humans and envi-
ronment along the coast of western Norway through time.
Quaternary International 220: 133–146.
Holden, J., Shotbolt, L., Bonn, A., Burt, T.P., Chapman, P.J.,
Dougill, A.J., Fraser, E.D.G., Hubacek, K., Irvine, B., Kirkby,
M.J., Reed, M.S., Prell, C., Stagl, S., Stringer, L.C., Turner, A.
& Worrall, F. 2007. Environmental change in moorland
landscapes. Earth-Science Reviews 82: 75–100.
Honnay, O., Verheyen, K. & Hermy, M. 2002. Permeability of
ancient forest edges for weedy plant species invasion. Forest
Ecology andManagement 161: 109–122.
Hutchinson, T., Boerner, R., Iverson, L., Sutherland, S. & Suther-
land, E. 1999. Landscape patterns of understory composition
and richness across a moisture and nitrogen mineralization
gradient in Ohio (U.S.A.) quercus forests. Plant Ecology 144:
177–189.
Jager, H., Tye, A. & Kowarik, I. 2007. Tree invasion in naturally
treeless environments: impacts of quinine (Cinchona pubes-
cens) trees on native vegetation in Galapagos. Biological Con-
servation 140: 297–307.
Kaland, P.E. 1986. The origin and management of Norwegian
coastal heaths as reflected by pollen analysis. In: Berhe, K.E.
(eds.) Anthropogenic indicators in pollen diagrams. pp. 37–52.
A.A. Belkema, Rotterdam, NL.
Kvamme, M., Kaland, P.E. & Brekke, N.G. 2004. Conservation and
management of north european coastal heathlands. Case study :the
heathland centre, Lygra, western Norway. Heathguard, The
Heathland Centre, Lygra, NO.
Leibold, M.A., Holyoak, M., Mouquet, N., Amarasekare, P.,
Chase, J.M., Hoopes, M.F., Holt, R.D., Shurin, J.B., Law, R.,
Tilman, D., Loreau, M. & Gonzalez, A. 2004. The metacom-
munity concept: a framework for multi-scale community
ecology. Ecology Letters 7: 601–613.
Lichstein, J. & Pacala, S. 2011. Local diversity in heterogeneous
landscapes: quantitative assessment with a height-structured
forest metacommunitymodel. Theoretical Ecology 4: 269–281.
Lid, J. & Lid, D.T. 2005. Norsk Flora. 7. utgave ved Reidar Elven. Det
Norske Samlaget, Oslo, NO.
Journal of Vegetation Science10 Doi: 10.1111/jvs.12010© 2012 International Association for Vegetation Science
Effects of introduced versus native conifer invasion H.I.Saure et al.

Lindgaard, A. & Henriksen, S. 2011. Norsk rødliste for naturtyper
2011. Artsdatabanken, Trondheim, NO.
Loidi, J., Biurrun, I., Campos, J.A., Garcıa-Mijangos, I. & Herre-
ra, M. 2010. A biogeographical analysis of the European
Atlantic lowland heathlands. Journal of Vegetation Science 21:
832–842.
Lopez-Pintor, A., Sal, A.G. & Benayas, J.M.R. 2006. Shrubs as a
source of spatial heterogeneity – the case of Retama sphaero-
carpa in Mediterranean pastures of central Spain. Acta Oeco-
logica 29: 247–255.
Maren, I.E. & Nilsen, L.S. 2008. Kystlyngheier i midt- og nord-
norge. Blyttia 66: 9–25.
Mitchell, R.J., Marrs, R.H., Duc, M.G.L. & Auld, M.H.D. 1997.
A study of succession on lowland heaths in Dorset, southern
England: changes in vegetation and soil chemical properties.
Journal of Applied Ecology 34: 1426–1444.
Mitchell, R.J., Marrs, R.H., Le Duc, M.G. & Auld, M.H.D. 1999.
A study of the restoration of heathland on successional sites:
changes in vegetation and soil chemical properties. Journal
of Applied Ecology 36: 770–783.
Moen, A. 1998. Nasjonalatlas for Norge: vegetasjon. Statens Kart-
verk, Hønefoss, NO.
Nakatani, A.S., Martines, A.M., Nogueira, M.A., Fagotti, D.f.S.L.,
Oliveira, A.G., Bini, D., Sousa, J.P. & Cardoso, E.J. B.N.
2011. Changes in the genetic structure of bacteria andmicro-
bial activity in an agricultural soil amended with tannery
sludge. Soil Biology and Biochemistry, 43: 106–114.
NGU 2012. Berggrunnsgeologidatabasen, Norges Geologiske
Undersøkelse. http://www.ngu.no/kart/kartkatalog/
Norwegian Ministry of the Environment 2006–2007. The gov-
ernment‘s environmental policy and the state of the envi-
ronment in Norway. Report No. 26.
Peterken, G.F. 2001. Ecological effects of introduced tree species
in Britain. Forest Ecology andManagement 141: 31–42.
Peterson, E.B., Peterson, N.M., Weetman, G.F. & Martin, P.J.
1997. Ecology and management of sitka spruce, emphasizing its
natural range in British Columbia. UBC Press, Vancouver, CA.
Piessens, K., Honnay, O., Devlaeminck, R. & Hermy, M. 2006.
Biotic and abiotic edge effects in highly fragmented heath-
lands adjacent to cropland and forest. Agriculture, Ecosystems
& Environment 114: 335–342.
Plieninger, T., Hochtl, F. & Spek, T. 2006. Traditional land-use
and nature conservation in European rural landscapes. Envi-
ronmental Science and Policy 9: 317–321.
Prøsch-Danielsen, L. & Simonsen, A. 2000. Palaeoecological
investigations towards the reconstruction of the history of
forest clearances and coastal heathlands in south-western
Norway. Vegetation History and Archaeobotany 9: 189–204.
Richardson, D.M. 1998. Forestry trees as invasive aliens. Conser-
vation Biology 12: 18–26.
Richardson, D.M. & Rejmanek, M. 2004. Conifers as invasive
aliens: a global survey and predictive framework. Diversity
and Distributions 10: 321–331.
Richardson, D.M., Macdonald, I.A.W. & Forsyth, G.G. 1989.
Reductions in plant species richness under stands of alien
trees and shrubs in the fynbos biome. South African Forestry
Journal 149: 1–8.
Savadogo, P., Tigabu, M., Sawadogo, L. & Oden, P.C. 2009.
Examination of multiple disturbances effects on herbaceous
vegetation communities in the Sudanian savanna-woodland
of West Africa. Flora - Morphology, Distribution, Functional
Ecology of Plants 204: 409–422.
Shmida, A. & Ellner, S. 1984. Coexistence of plant species with
similar niches. Plant Ecology 58: 29–55.
Shmida, A. &Wilson,M.V. 1985. Biological determinants of spe-
cies diversity. Journal of Biogeography 12: 1–20.
Soderstrom, L., Urmi, E. & Vana, J. 2002. Distribution of hepati-
cae and anthocerotae in Europe andMacaronesia. Lindbergia
27: 3–48.
Sørensen, M.M. & Tybirk, K. 2000. Vegetation analysis along a
successional gradient from heath to oak forest. Nordic Journal
of Botany 20: 537–546.
Thorvaldsen, P. 2011. Skjøtselsplan for kystlynghei pa Tarva i Bjugn
kommune. Bioforsk Rapport/ Bioforsk Report, Bioforsk Vest
Fureneset, Norway.
Tryon, R. 1970. Development and evolution of fern floras of oce-
anic islands. Biotropica 2: 76–84.
UK BAP 1999. Website http://www.ukbap.org.uk/UKPlans.
aspx?ID=462(originalUKBAPTranches1and2(1995-1999).
Vetaas, O.R. 1992. Micro-site effects of trees and shrubs in dry
savannas. Journal of Vegetation Science 3: 337–344.
Vevle, O. 2000. Ellenbergs økologiske faktortal for norske planter.
Høgskolen i Telemark, Bø, Norway.
Virtanen, R. & Oksanen, J. 2007. The effects of habitat connec-
tivity on cryptogam richness in boulder metacommunity.
Biological Conservation 135: 415–422.
Wallace, H.L. & Good, J.E.G. 1995. Effects of afforestation on
upland plant communities and implications for vegetation
management. Forest Ecology and Management 79: 29–46.
Wallrup, E., Saetre, P. & Rydin, H. 2006. Deciduous trees affect
small-scale floristic diversity and tree regeneration in conifer
forests. Scandinavian Journal of Forest Research 21: 399–404.
Webb, N.R. 1998. The traditional management of European
heathlands. Journal of Applied Ecology 35: 987–990.
Zinke, P.J. 1962. The pattern of influence of individual forest
trees on soil properties. Ecology 43: 130–133.
Supporting Information
Additional supporting information may be found in the
online version of this article:
Appendix S1. Species list from coastal heathlands,
Sitka spruce plantations and Scots pine forests.
Journal of Vegetation ScienceDoi: 10.1111/jvs.12010© 2012 International Association for Vegetation Science 11
H.I.Saure et al. Effects of introduced versus native conifer invasion