effects of endothelin receptor antagonists on the progression of diabetic nephropathy
TRANSCRIPT

Inflammation, Vol. 18, No. 6, 1994
EFFECTS OF ENDOTHELIN RECEPTOR ANTAGONISTS ON BRADYKININ-INDUCED
INCREASES IN MACROMOLECULAR EFFLUX
W I L L I A M G. M A Y H A N and I S R A E L R U B I N S T E I N
Departments of Physiology and Biophysics and Internal Medicine University of Nebraska Medical Center
Omaha, Nebraska 68198-4575
Abstract--The goal of this study was to determine the effects of endothelin receptor antagonists on agonist-induced increases in macromolecular extravasation in the ham- ster cheek pouch in vivo. We used intravital fluorescent microscopy and fluorescein isothiocyanate dextran (FITC-dextran; mol wt = 70 K) to examine extravasation from postcapillary venules in response to bradykinin and endothelin before and fol- lowing application of inhibitors of endothelin receptors (ETAn and ETA). Increases in extravasation of macromolecules were quantitated by counting the number of venular leaky sites. Bradykinin (0.5 and 1.0 /~M) and endothelin-1 (0.01 and 0.1 nM) produced a dose-related increase in the number of venular leaky sites and super- fusion of PD 142893 (ETAB antagonist), and PD 147953 and BQ-123 (ETA antago- nists) significantly decreased bradykinin- and endothelin-induced responses. Addition of calcium to the superfusate restored bradykinin-induced increases in venular leaky sites in the presence of endothelin receptor antagonism. Thus, the findings of the present study suggest that endothelin receptor antagonists abrogate bradykinin- and endothelin-induced increases in macromolecular efltux from postcapillaiy venules. The mechanism for the effects of endothelin receptor antagonists appears to be related to inhibition of the ETA receptor which, in turn, alters the mobilization of calcium across venular endothelium.
INTRODUCTION
Endo the l i a l ce l ls syn thes ize and re lease a var ie ty o f va soac t ive subs t ances
inc lud ing p ros t ag l and ins , e n d o t h e l i u m - d e r i v e d re lax ing fac tor (ni tr ic ox ide) and
endo the l ins . P rev ious s tudies h a v e sugges ted that p ro s t ag l and ins and ni t r ic ox ide
p lay an impor t an t role in the regu la t ion o f v a s c u l a r reac t iv i ty and pe rmeab i l i t y
(1 -6 ) . H o w e v e r , s ince the d i scove ry o f endo t he l i n , m o s t research has focused
on the effects o f endo the l i n s o n the regu la t ion o f v a s c u l a r tone and t i ssue b l o o d
633
0360-3997/94/1200-0633507.00/0 �9 1994 Plenum Publishing Corporation

634 Mayhan and Rubenstein
flow (7-10). Several recent studies have begun to investigate the effects of endothelins on vascular permeability. For the most part, these studies have examined the direct effects of endothelin on vascular permeability (11-15) or the effects of endothelin on agonist-induced increases in vascular permeability (16-18).
Recently, the mechanisms that govem the effects of endothelin on endo- thelium and vascular muscle have been investigated using various specific inhib- itors of endothelin-receptors (ET A and ETB). These studies suggest that endothelin can activate receptors on endothelium (ETa) and vascular muscle (ETA) to modulate vascular reactivity (8, 19, 20). In addition, two studies suggested that endothelin increases permeability of peripheral tissues via acti- vation of ET A receptors (21, 22). However, no studies have examined the role of ET A and ET B receptors on the modulation of permeability of the microcir- culation in vivo in response to inflammatory mediators. Thus, the first goal of the present studies was to examine the effects of endothelin receptor antagonists on endothelin-1- and bradykinin-induced increases in macromolecular efflux from venules in vivo.
We (23) and others (24-27) have shown that modulation of external calcium concentration influences the formation of venular leaky sites and microvascular permeability during normal and stimulated states. In addition, investigators have shown that endothelins affect calcium mobilization to influence contraction of vascular smooth muscle (7, 9, 28, 29). Thus, the second goal of this study was to examine whether alterations in the external calcium concentration could mod- ulate the effects of endothelin receptor antagonists on bradykinin-induced increases in macromolecular efflux from venules in vivo.
MATERIALS AND METHODS
Preparation of Animals. Adult, male Syrian hamsters weighing between 100 and 130 g were anesthetized (pentobarbital sodium, 60 mg/kg body wt, intraperitoneal) and a tracheotomy was performed to facilitate spontaneous breathing. Supplemental anesthesia was administered at a dose of 20-40 mg/kg/hr intravenously.
The left femoral vein was cannulated for the purpose of injecting the intravascular tracer ffuorescein isothiocyanate dextran (FITC-dextran; mol wt 70,000 D). The left femoral artery was cannulated for the purpose of measuring arterial pressure, which was constant throughout the exper- iment.
To visualize the microcirculation of the cheek pouch, we used a method that we have described
previously (3, 23, 30, 31). Briefly, the cheek pouch was spread over a small plastic baseplate and an incision was made in the skin to expose the cheek pouch membrane. An avascular layer of connective tissue was excised to expose the microvessels of the cheek pouch. An upper chamber was then positioned over the baseplate to provide a reservoir for the suffusion fluid. This arrangement

Endothelin Antagonists and Permeability 635
forms a triple-layered complex: the baseplate, the upper chamber, and the cheek pouch microcir- culation exposed between these two plates.
The cheek pouch microcirculation was continuously suffused with a bicarbonated buffer (36- 38~ The cheek pouch chamber also was connected via a three-way valve to an infusion pump, which allowed for infusion of agonists into the suffusate. This method, which we have used pre- viously (3, 23, 30, 31), allowed us to maintain a constant temperature and pH of the suffusion fluid during infusion of agonists.
Evaluation of Microvascular Permeability. Alterations in microvascular transport were quan- titated by counting the number of microvascular leaky sites, which occurred exclusively around postcapillary venules, as we have reported previously (3, 23, 30, 31). The number of leaky sites (number per 0.11 cm 1) was determined under control conditions (before application of agonists) and at 1, 3, 5, 7, 10, 15, 20, and 30 min during and following application of agonists. The number of leaky sites observed in two microscopic fields (0.11 cmZ/microscopic field), chosen at random, under control conditions and during application of agonists was averaged, and we report responses as the average of these two microscopic fields (3, 5, 32).
Effects of ETAB inhibition. We examined the effects of PD 142893 (0.1 tzM) on bradykinin- induced formation of venular leaky sites (N = 5). Successive doses of bradykinin (0.5 and 1.0 #M) were superfused over the cheek pouch microcireulation for 5 rain. There was a 30 to 40-min recovery period between the application of each concentration of bradykinin. The number of venular leaky sites was determined as described above. After initially testing responses of the cheek pouch micro- circulation to bradykinin, we then started a continuous superfusion of PD 142893 (0.1 /~M) over the cheek pouch microcirculation. Previous studies have shown that PD 142893 inhibits the ETAB receptor (33), and in preliminary studies we determined that this concentration of PD 142893 was efficacious in inhibiting responses of the cheek pouch microcirculation to endothelin-1. Thirty min- utes after starting superfusion of PD 142893, we again examined the effects of bradykinin on the formation of venular leaky sites.
Effects of ET A inhibition. First, we examined the effects of PD 147953 (0.1 #M; N = 7) on bradykinin-induced formation of venular leaky sites. PD 147953 is similar to FR 139317, a potent ET A receptor antagonist (33). Successive doses of bradykinin (0.5 and 1.0 #M) were super- fused over the cheek pouch microcirculation for 5 min. The number of venular leaky sites was determined as described above. After initially testing responses of the cheek pouch microcirculation to bradykinin, we then started a continuous superfusion of PD 147953 (0.1 tzM) over the cheek pouch microcirculation. Thirty minutes after starting superfusion of PD 147953, we again examined the effects of bradykinin on the formation of venular leaky sites.
To determine the specificity of endothelin receptor antagonism on bradykinin-induced increases in venular leaky site formation, we examined the effect of PD 147953 on endothelin-induced for- mation of venular leaky sites (N = 7). Thus, in these studies we first examined the effects of endothelin (0.01 and 0.1 nM) on venular leaky site formation. Then, we applied PD 147953 (0.1 #M) to the cheek pouch microeirculation. Thirty minutes after starting superfusion of PD 147953, we again examined the effects of endothelin-1 on venular leaky site formation.
In another series of experiments, we examined the effects of BQ-123 (1.0 tzM; N = 4) on bradykinin-induced formation of venular leaky sites. BQ-123 is a highly selective ET A receptor antagonist (33, 34). Successive doses of bradykinin (0.5 and 1.0 t~M) were superfused over the cheek pouch microcirculation for 5 min. The number of venular leaky sites was determined as described above. After initially testing responses of the cheek pouch microcirculation to bradykinin, we then started a continuous superfusion of BQ-123 over the cheek pouch microcirculation. Thirty minutes after starting superfusion of BQ-123, we again examined the effects of bradykinin on the formation of venular leaky sites.
Effects of Extracellular Calcium. We examined whether addition of excess calcium to the suffusate during application of PD 147953 could affect venular leaky site formation in response to

636 Mayhan and Rubenstein
bradykinin. Thus, in these studies, we first examined the formation of venular leaky sites in response to bradykinin during superfusion with a standard calcium-containing buffer (1.5 mM). Then we started a continuous supeffusion of PD 147953 (0.1 /zM; N = 5) over the cheek pouch microcir- culation. Concomitant with the application of PD 147953, we suffused the cheek pouch microcir- culation with buffer containing twice-normal calcium (3.0 mM). Thirty minutes after starting suffusion of PD 147953 and the buffer containing twice-normal calcium, we again examined the formation of venular leaky sites in response to bradykinin.
Effects of Repeated Application of Bradykinin. To determine whether inhibition of venular leaky sites by PD 142893, PD 147953, and BQ-123 in response to bradykinin was related to tachyphylaxis to bradykinin, we examined the effects of repeated application of bradykinin on the formation of venular leaky sites. Thus, in these studies (N = 8), a similar protocol was completed as that described for endothelin receptor antagonists. Bmdykinin (0.5 and 1.0 #M) was superfused over the cheek pouch microcirculation for 5 min, and the number of venular leaky sites was determined as described above. Then, instead of suffusing an endothelin receptor antagonist across the cheek pouch microcirculation, we suffused vehicle. Thirty minutes after starting superfusion of vehicle, we again examined the effects of bmdykinin on the formation of venular leaky sites.
Statistical Analysis. Paired t tests were used to compare responses before and after appli- cation of PD 142892, PD 147953, BQ-123 or vehicle A P value of 0.05 was considered to be significant.
R E S U L T S
Effects of ETAB Inhibition. Under control condit ions, no leaky sites were
visible in the cheek pouch. Topical application of bradykin in produced a sig- nificant concentra t ion-dependent increase in the n u m b e r of venular leaky sites. Al l leaky sites occurred around postcapillary venules . The m ax im um number
o f leaky sites occurred wi thin 5 - 1 0 min after starting suffusion of bradykinin , and all leaky sites resolved during the recovery period. These findings are similar to that which we and others have reported previously (5, 31, 35-38) . Applicat ion
of PD 142893 did not produce venu la r leaky sites, but PD 142893 significantly inhibi ted venular leaky site formation in response to bradykin in (Figure 1).
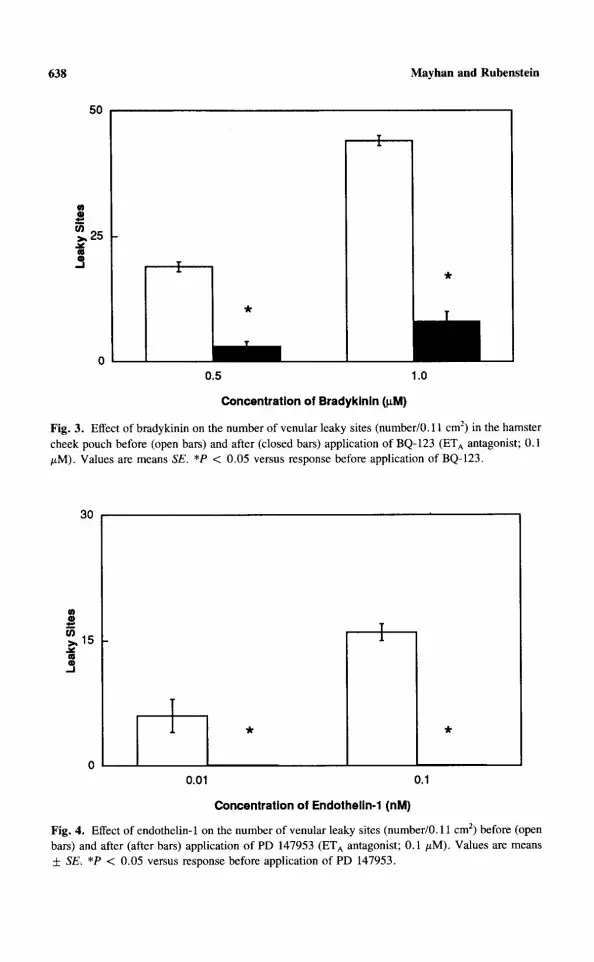
Effects of ETA Inhibition. Un d e r control condi t ions, no leaky sites were visible in the cheek pouch. Topical applicat ion of b radykin in produced a con- centra t ion-dependent increase in the number of venular leaky sites (Figures 2 and 3). Appl icat ion of PD 147953 or BQ-123 did not produce venular leaky
sites, but PD 147953 (Figure 2) and BQ-123 (Figure 3) significantly inhibited venular leaky site formation in response to bradykinin.
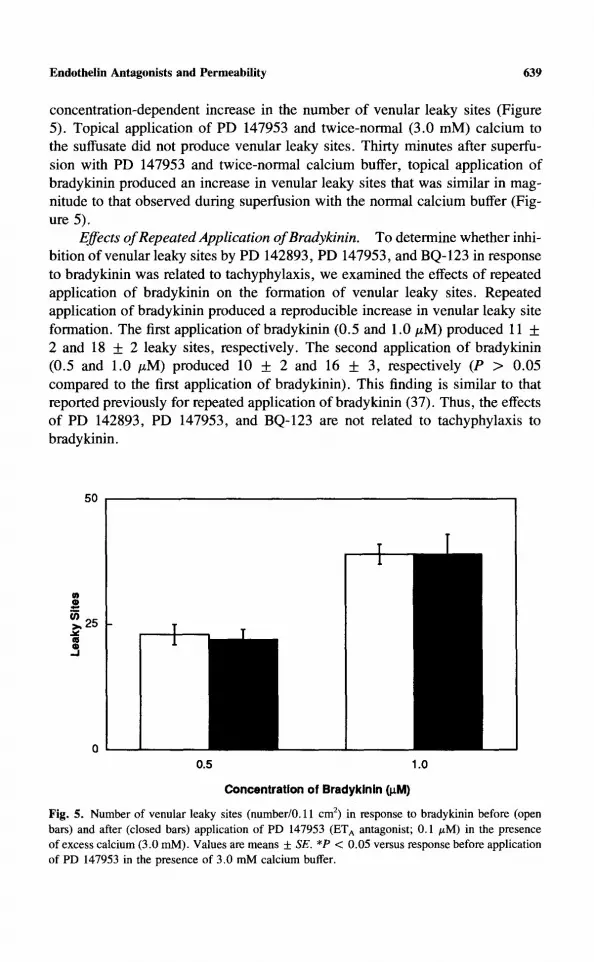
Topical applicat ion of endothel in produced an increase in the n u m b e r of venular leaky sites. Appl icat ion of PD 147953 did not produce venular leaky sites, but PD 147953 abolished venular leaky site formation in response to endothel in (Figure 4).
Effects of Extracellular Calcium. Un d e r control condit ions, no leaky sites were visible in the cheek pouch. Topical application of bradykin in produced a

Endothelin Antagonists and Permeability
30
637
m
~15 m @
J 0.5 1.0
.k
Concentration of Bradykinin (I~M)
Fig. 1. Effect of bradykinin on the number of venular leaky sites (number/0.11 cm 2) in the hamster cheek pouch before (open bars) and after (closed bars) application of PD 142893 (ETAB antagonist; 0.1 ftM). Values are means + SE. P < 0.05 versus response before application of PD 142893.
30
.ag lu
.k
0.5 1.0
Concentration of Bradykinln (I~M)
Fig. 2. Effect of bradykinin on the number of venular leaky sites (number/0.11 cm 2) in the hamster cheek pouch before (open bars) and after (closed bars) application of PD 147953 (ETA antagonist; 0.1 ftM). Values are means + SE. *P < 0.05 versus response before application of PD 147953.
638
50
Mayhan and Rubenstein
w
~>,25 Jlr M
0.5 1.0
Concentration of Bradyklnln (~M)
Fig. 3. Effect of bradykinin on the number of venular leaky sites (number/0.11 cm 2) in the hamster cheek pouch before (open bars) and after (closed bars) application of BQ-123 (ETA antagonist; 0.1 /zM). Values are means SE. *P < 0.05 versus response before application of BQ-123.
30
Q
r m ,,,J
I 0 I 0.01 0.1
Concentration of Endothelln-1 (nM)
Fig. 4. Effect of endothelin-1 on the number of venular leaky sites (number/0.11 cm 2) before (open bars) and after (after bars) application of PD 147953 (ET A antagonist; 0.1 /~M). Values are means • SE. *P < 0.05 versus response before application of PD 147953.
Endothelin Antagonists and Permeability 639
concentration-dependent increase in the number o f venular leaky sites (Figure 5). Topical application of PD 147953 and twice-normal (3.0 mM) calcium to the suffusate did not produce venular leaky sites. Thirty minutes after superfu- sion with PD 147953 and twice-normal calcium buffer, topical application of bradykinin produced an increase in venular leaky sites that was similar in mag- nitude to that observed during superfusion with the normal calcium buffer (Fig- ure 5).
Effects of Repeated Application of Bradykinin. To determine whether inhi- bition of venular leaky sites by PD 142893, PD 147953, and BQ-123 in response to bradykinin was related to tachyphylaxis, we examined the effects of repeated application of bradykinin on the formation of venular leaky sites. Repeated application of bradykinin produced a reproducible increase in venular leaky site formation. The first application of bradykinin (0.5 and 1.0/~M) produced 11 _ 2 and 18 ___ 2 leaky sites, respectively. The second application o f bradykinin (0.5 and 1.0 /zM) produced 10 + 2 and 16 _ 3, respectively (P > 0.05 compared to the first application of bradykinin). This finding is similar to that reported previously for repeated application of bradykinin (37). Thus, the effects of PD 142893, PD 147953, and BQ-123 are not related to tachyphylaxis to bradykinin.
50
~., 2s
Q , . I
0 0.5 1.0
Concentration of Bradyklnln (p.M)
Fig. 5. Number of venular leaky sites (number/0.11 cm 2) in response to bradykinin before (open bars) and after (closed bars) application of PD 147953 (ET A antagonist; 0.1 /~M) in the presence of excess calcium (3.0 mM). Values are means + SE. *P < 0.05 versus response before application of PD 147953 in the presence of 3.0 mM calcium buffer.

640 Mayhan and Rubenstein
DISCUSSION
There are two major new findings of the present study. First, in vivo extravasation of macromolecules from postcapillary venules in response to bra- dykinin and endothelin can be inhibited by antagonists of ETAB and ETA recep- tors. We found that PD 142893, PD 147953, and BQ-123 inhibited venular leaky site formation in response to bradykinin and endothelin. Second, the effects of endothelin receptor antagonists on bradykinin-induced increases in venular leaky site formation appear to be related to their effects on extracellular calcium mobilization. We found that application of additional calcium to the superfusate could reverse the effects of PD 147953 on bradykinin-induced venular leaky site formation. Our findings suggest a unique role for endothelin receptor antagonists in modulating microvascular permeability in vivo.
Consideration of Methods. The hamster cheek pouch has been used as a model to study the effects of inflammatory mediators on microvascular perme- ability (5, 23, 30, 31, 38-40). In the present study, we used in vivo fluorescent microscopy to examine extravasation of FITC-dextran 70 K from venules con- tained within the microcirculation of the hamster cheek pouch. This allowed us to determine the precise location of macromolecular efflux during stimulation with bradykinin and endothelin. We and others have used the evaluation of formation of venular leaky sites as a means to quantitate changes in macromo- lecular transport (3, 23, 30, 31, 38-41). We have suggested that the formation of venular leaky sites contributes to increases in microvascular permeability, since an increase in clearance of FITC-dextran from the cheek pouch was directly related to the formation of venular leaky sites (23).
We examined macromolecular extravasation of FITC-dextran from venules in response to application of bradykinin before and after treatment with endoth- elin receptor antagonists (PD 142893, PD 147953, and BQ-123). We found that topical application of endothelin receptor antagonists significantly inhibited macromolecular extravasation from venules in response to bradykinin. We con- sidered the possibility that impairment of bradykinin-induced increases in ven- ular leaky sites after treatment with ETAB and ET A receptor antagonists may be related to tachyphylaxis to bradykinin. However, repeated application of bra- dykinin produced reproducible increases in venular leaky sites. Thus, inhibition of bradykinin-induced venular leaky site formation by endothelin receptor antag- onists can not be attributed to tachyphylaxis.
We considered the possibility that inhibition of bradykinin-induced increases in venular leaky sites by endothelin receptor antagonists may be related to an effects on calcium mobilization across venular endothelium. We (23) and others (24-27) have shown that modulation of external calcium concentration influ- ences the formation of venular leaky sites and microvascular permeability during

Endothelin Antagonists and Permeability 641
normal and stimulated states. These previous studies (23-27) suggest that increases in permeability of venules require a transmembrane flux of calcium, presumably for contraction of endothelial cells, to provide a pathway for the movement of macromolecules. Investigators have shown that endothelins affect calcium mobilization to influence contraction of vascular smooth muscle (7, 9, 28, 29). The present studies are the first to suggest that endothelin receptor antagonists also may affect the mobilization of calcium across venular endothe- lium to modulate microvascular permeability.
Consideration of Previous Studies. Several studies have examined the effects of endothelin on vascular permeability (11-15). These investigators (11- 15) have suggested that intravascular and intradermal injection of endothelin-1 produce an increase in permeability of many vascular tissues, including skin, liver, pancreas, stomach, lung, kidney, and intestine. Thus, in addition to its pronounced effects of vascular reactivity, endothelin-1 produces an increase in vascular permeability. The results of the present study confirm the findings of previous investigators (11-15). We found that topical application of endothelin produced an increase in extravasation of macromolecules from venules contained within the hamster cheek pouch microcirculation in vivo. In addition, our study extends previous findings (11-15) by suggesting that endothelin acts specifically on postcapillary venules to increase macromolecular efflux.
In addition to studies that have examined the direct effects on endothelin on vascular permeability, other investigators have examined the effects of endothelin-1 on agonist-induced increases in vascular permeability (16-18). These investigators (16-18) report that endothelin-1 suppresses agonist-induced increases in vascular permeability. The mechanism by which endothelin-1 inhib- its agonist-induced increases in vascular permeability was not directly investi- gated, but was speculated to be related to the effect of endothelin-1 on vascular diameter, a direct effect on endothelin on vascular permeability, and/or an effect of endothelin on nerves or cells that synthesize/release substances to modulate vascular permeability (16, 18).
Other recent studies have examined the role of endothelin receptors in changes in vascular permeability in response to endothelin-1 (21, 22). These studies suggcst that increases in vascular permeability in response to intravas- cular administration of endothelin-1 could be inhibited by treatment with BQ- 123, an ETA antagonist (21, 22). Thus, these studies (21, 22) suggest that changes in vascular permeability in response to endothelin-1 are related to acti- vation of ET A receptors. The findings of the present study are in agreement with the previous studies (21, 22). We found that endothelin-l-induced formation of venular leaky sites could be inhibited by application of another ETA receptor antagonist, PD 147953. In addition, we extend findings of previous studies (21, 22) by examining the effects endothelin receptor antagonists on bradykinin- induced increases in macromolecular efflux. We found that endothelin receptor

642 Mayhan and Rubenstein
antagonists inhibit format ion o f venular leaky sites in response to bradykinin.
Fur thermore , we examined a potential mechan i sm for the effects o f endothel in
receptor antagonists on macromolecu la r efflux. W e found that addit ion o f extra-
cel lular ca lc ium to the suffusate could restore bradykinin- induced formation o f
venular leaky sites in the presence o f an ETA antagonist . This suggests that the
effects o f endothel in receptor antagonists on macromolecu la r efflux in the periph-
eral microci rcula t ion may be related to alterations in ca lc ium mobi l iza t ion across postcapil lary venular endothel ium.
In conclus ion, we suggest that endothel in receptor antagonists inhibit the
format ion o f venu la r leaky sites in response to bradykinin. In addit ion, the effects
o f endothel in receptor antagonists on venular leaky site format ion in response
to bradykinin could be reversed by addit ion o f excess extracel lular calcium.
Thus, inhibi t ion o f the ET A receptor, prevent ing the mobi l iza t ion o f ca lc ium
across postcapil lary venular endothel ium, appears to be important for inhibit ion
o f bradykinin- induced format ion o f venular leaky sites by endothel in receptor
antagonists. Our findings suggest a unique role for endothel in receptor antago-
nists on the regulat ion o f bradykinin- induced increases in microvascular perme- ability in v ivo.
Acknowledgments--The authors would like to thank Glenda Sharpe for excellent technical assistance. The authors also would like to thank Steve Haleen from Parke-Davis Pharmaceuticals for helpful suggestions and the gift of PD 142893, PD 147953, and BQ-123. This study was supported, in part, by National Institutes of Health grants HL-40781 and DE-10347, Research Career Develop- ment Awards HL-02124 and DE-00355, a Seed Grant (94-08) from the University of Nebraska College of Medicine, a Grant-in-Aid from the American Heart Association; National Affiliate (91006230), a Grant-in-Aid from the American Heart Association; Nebraska Affiliate (9307792S), and support from Parke-Davis Pharmaceuticals.
R E F E R E N C E S
1. FURCHGOTT, R. F., and P. M. VANHOUTTE. 1989. Endothelium-derived relaxing and contracting factors. FASEB J. 3:2007-2018.
2. JOYNER, W. L., E. SVENSJO, and K. E. ARFORS. 1979. Simultaneous measurements of macro- molecular leakage and arteriolar blood flow as altered by PGE~ and B2-receptor stimulant in the hamster cheek pouch. Microvasc. Res. 18:301-310.
3. YONG, T., and W. G. MAYHAN. 1992. Effect of prostaglandin El on leukotriene C4-induced increases in vascular permeability of hamster cheek pouch. Inflammation 16:159-167.
4. PECK, M. J., P. J. PIPER, and T. J. WILLIAMS. 1981. The effect of leukotrienes Ca and D4 on the microvasculature of guinea-pig skin. Prostaglandins 21:315-321.
5. MAYHAN, W. G. 1992. Role of nitric oxide in modulating permeability of the hamster cheek pouch in response to adenosine 5'-diphosphate and bradykinin. Inflammation 16:295-305.
6. MONCADA, S. 1990. The first Robert Furchgott lecture: From endothelium-dependent relaxation to the L-arginine: NO pathway. Blood Vessels 27:208-217.

Endothelin Antagonists and Permeability 643
7. MASAKI, T. 1993. Endothelins: Homeostatic and compensatory actions in the circulatory and endocrine systems. Endocrine Rev. 14:256-268.
8. SAKURAI, T., and K. GOTO. 1993. Endothelins. Vascular actions and clinical implications. Drugs 46:795-800.
9. CHABRIER, P. E., and P. BRAQUET. 1990. Endothelin. Horm. Res. 34:169-174. 10. RANDALL, M. D. 1991. Vascular activities of the endothelins. Pharrnacol. Ther. 50:73-93. 11. BRAIN, S. D., G. THOMAS, D. C. CROSSMAN, R. FULLER, and M. K. CHURCH. 1992. Endoth-
elin-1 induces a histamine-dependent flare in vivo, but does not activate human skin mast cells in vitro. Br. J. Clin. Pharmacol. 33:117-120.
12. FILEP, J. G., M. G. SIROIS, A. ROUSSEAU, A. FOURNIER, and P. SIROIS. 1991. Effects of endothelin-1 on vascular permeability in the conscious rat: Interactions with platelet-activating factor. Br. J. Pharmacol. 104:797-804.
13. SIROIS, M. G., J. G. FILEP, A. ROUSSEAU, A. FOURNIER, G. E. PLANTE, and P. SIROIS. 1992. Endothelin-1 enhances vascular permeability in conscious rats: Role of thromboxane A2. Eur. J. Pharmacol. 214:119-125.
14. LEHOUX, S., G. E. PLANTE, M. G. SIROIS, P. SIROIS, and P. D'ORLI~ANS-JUSTE. 1992. Phos- phoramidon blocks big-endothelin-1 but not endothelin-1 enhancement of vascular permeability in the rat. Br. J. Pharmacol. 107:996-1000.
15. FILEP, J. G., G. E. FOLDES-FILEP, A. ROUSSEAU, P. SIROIS, and A. FOURNIER. 1993. Vascular responses to endothelin-1 following inhibition of nitric oxide synthesis in the conscious rat. Br. J. Pharmacol. 110:1213-1221.
16. BRAIN, S. D., D. C. CROSSMAN, Z. L. BUCKLEY, and T. J. WILLIAMS. 1989. Endothelin-l: Demonstration of potent effects on the microcirculation of humans and other species. J. Car- diovasc. Pharmacol. 13:S147-S149.
17. CHANDLER, C. L., A. R. MOORE, F. M. DESA, D. W. HOWAT, and D. A. WILLOUGHBY. 1989. Anti-inflammatory effects of endothelin-1. J. Cardiovasc. Pharmacol. 13:$218-$219.
18. CHANDER, C. L., A. R. MOORE, F. M. DESA, D. HOWAT, and D. A. WILLOUGHBY. 1988. The local modulation of vascular permeability by endothelial cell derived products. J. Pharm. Pharmacol. 40:745-746.
19. HAYNES, W. G., and D. J. WEBB. 1993. The endothelin family of peptides: Local hormones with diverse roles in health and disease. Clin. Sci. 84:485-500.
20. RUBANYI, G. M., and L. H. P. BOTELHO. 1991. Endothelins. FASEB J. 5:2713-2720. 21. FILEP, J. G., M. G. SIROIS, E. F(3LDES-FILEP, A. ROUSSEAU, G. E. PLANTE, A. FOURNIER, M.
YANO, and P. SIROIS. 1993. Enhancement by endothelin-1 of microvascular permeability via the activation of ET A receptors. Br. J. Pharmacol. 109:880-886.
22. FILEP, J. G., E., FI3LDES-FILEP, A. ROUSSEAU, A. FOURNIER, P. SIROIS, and M. YANO. 1992. Endothelian-1 enhances vascular permeability in the rat heart through the ET A receptor. Eur. J. Pharmacol. 219:343-344.
23. MAYHAN, W. G., and W. L. JOYNER. 1984. The effect of altering the external calcium con- centration and a calcium channel blocker, verapamil, on microvascular leaky sites and dextran clearance in the hamster cheek pouch. Microvasc. Res. 28:159-179.
24. SVENSJO, E., and G. J. GREGA. 1986. Evidence for endothelial cell mediated regulation of macromolecular permeability by postcapillary venules. Fed. Proc. 45:89-95.
25. OLESEN, S. P. 1987. Regulation of ion permeability in frog brain venules, significance of calcium, cyclic nucleotides and protein kinase C. J. Physiol. 387:59-68.
26. PAUL, J., A. Y. BEKKER, and W. N. DURAN. 1990. Calcium entry blockage prevents leakage of macromolecules induced by ischemia-reperfusion in skeletal muscle. Circ. Res. 66:1636- 1642.
27. CURRY, F. E. 1992. Modulation of venular microvessel permeability of calcium influx into endothelial cells. FASEB J. 6:2456-2466.

644 Mayham and Rubenstein
28. JOSHUA, I. G. 1990. Endothelin-induced vasoconstriction of small resistance vessels in the microcirculation of the rat cmmaster muscle. Microvasc. Res. 40:191-198.
29. YANAGISAWA, M., H. KURIHARA, S. KIMURA, Y. TOMOBE, M. KOBAYASHI, Y. MITSUI, Y. YAZAKI, K. GOTO, and T. MASAKI. 1988. A novel potent vasoconstrictor peptide produced by vascular endothelial cells. Nature 332:411-415.
30. MAYHAN, W. G., G. SAHAGUN, R. SPECTOR, and D. D. HEISTAD. 1986. Effects of leukotriene C4 on the cerebral microvasculature. Am. J. Physiol. 251:H471-H474.
31. MURRAY, M. A., D. D. HEISTAD, and W. G. MAYHAN. 1991. Role of protein kinase C in bradykinin-induced increases in microvascular permeability. Circ. Res. 68:1340-1348.
32. MAYHAN, W. G. 1993. Role of nitric oxide in leukotriene C4-induced increases in microvascular transport. Am. J. Physiol. 265:H409-H414.
33. DOHERTY, A. M., W. L. ConY, P. L. DEPUE, J. X. HE, L. A. WAITE, D. M. LEONARD, N. L. LEITZ, D. T. DUDLEY, S. T. RAPUNDALO, G. P. HINGORANIS, S. J., HALEEN, D. M. LADOUCEUR, K. E. HILL, M. A. FLYNN, and E. E. REYNOLDS. 1993. Structure-activity rela- tionships of C-terminal endothelin hexapeptide antagonists. J. Med. Chem. 36:2585-2594.
34. IHARA, M., K. NOGUCHI, T. 8AEKI, T. FUKURODA, S. TSUCHIDA, S. KIMURA, T. FUKAMI, K. ISHIKAWA, M. NISHIKIBE, and M. YANO. 1991. Biological profiles of highly potent novel endoth- elin antagonists selective for the ETA receptor. Life Sci. 50:247-255.
35. YONG, T., X. P. GAD, S. KOIZUMI, J. M. CONLON, S. I. RENNARD, W. G. MAYHAN, and I. RUBINSTEIN. 1992. Role of peptidases in bradykinin-induced increase in vascular permeability in vivo. Circ. Res. 70:952-959.
36. SVENSJO, E., K. E. ANDERSSON, E. BOUSKELA, F. Z. G. A. CYRINO, and S. LINDGREN. 1993. Effects of two vasodilatory phosphodiesterase inhibitors on bradykinin-induced permeability in the hamster cheek pouch. Agents Actions 39:35-41.
37. SVENSJO, E., and W. L. JOYNER. 1984. The effects of intermittent and continuous stimulation of microvessels in the cheek pouch of hamsters with histamine and bradykinin on the devel- opment of venular leaky sites. Microcirc. Endothel. Lymph. 1:381-396.
38. SVENSJO, E. 1978. Bradykinin and prostaglandin El, E2 and F2 alpha induced macromolecular leakage in the hamster cheek pouch. Prostaglandins Med. 1:397-410.
39. SVENSJO, E. 1978. Characterization of leakage of macromolecules in postcapillary venules. Acta Univ. Upsaliensis 34:1-42.
40. SVENSJO, E., K. E. ARFORS, R. M. RAYMOND, and G. J. GREGA. 1979. Morphological and physiological correlation of bradykinin-induced macromolecular efflux. Am. J. Physiol. 236:H600-H606.
41. BORIC, M. P., J. S. ROBLERO, and W. N. DURAN. 1987. Quantitation of bradykinin-induced microvascular leakage of FITC-dextran in rat cremaster muscle. Microvasc. Res. 33:397-412.