effect of argemone oil and argemone alkaloid, sanguinarine on sertoli–germ cell coculture
TRANSCRIPT

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Toxicology Letters 186 (2009) 104–110
Contents lists available at ScienceDirect
Toxicology Letters
journa l homepage: www.e lsev ier .com/ locate / tox le t
Effect of argemone oil and argemone alkaloid, sanguinarineon Sertoli–germ cell coculture
Vivek Mishra, Daya Krishna Saxena, Mukul Das ∗
Indian Institute of Toxicology Research, (Formerly Industrial Toxicology Research Centre), Council of Scientific and Industrial Research,Post Box#80, Lucknow 226 001, India
a r t i c l e i n f o
Article history:Received 19 November 2008Received in revised form23 December 2008Accepted 6 January 2009Available online 16 January 2009
Keywords:Sertoli–germ cell cocultureCytotoxicityApoptosisRat
a b s t r a c t
Several incidences of reduction in the fertility (sperm count) have been reported in India and world-wide as well. Adulteration of food and consumption of adulterated mustard oil with argemone oil (AO)are presumed to be the factors for reduction in sperm count. In the present study we have studied theexfoliation of germ cells from Sertoli cell, its viability after detachment, cytotoxicity and execution ofapoptosis via mitochondrial pathway for different concentration of AO, argemone alkaloid (AA) and itsmajor constituent sanguinarine (SA). A dose dependent increase in germ cell detachment and decrease inviability of detached germ cells were observed (P < 0.05). A significant inhibition was observed via 3-(4,5-dimethylthiazol-2-yl)-2,5-dipehyl tetrazolium bromide (MTT) assay in the proliferative activity of germcell and leakage of cytosolic enzyme was observed via Lactate dehydrogenase(LDH) assay (P < 0.05). Atime and dose dependent inhibition of mitochondrial membrane potential was observed (P < 0.05). Treat-ment of Sertoli–germ cells with the lowest concentration of AO/AA and SA for 24 h resulted in 5.2- , 4.4-and 3.6-fold increase in the percentage of early apoptotic cells, respectively. This increase was enhancedto 8.3, 4.75 and 5.81-fold, respectively at 48 h in detached germ cells undergoing early apoptosis. Theseresults suggest that alterations in germ cell apoptosis by a disruption in contact mediated communicationbetween the Sertoli cells and germ cells, may subsequently lead to testicular impairment.
© 2009 Elsevier Ireland Ltd. All rights reserved.
1. Introduction
Reproductive aging in the male is characterized by the dra-matic changes in the seminiferous epithelium that eventuallylead to decrease in steroidogenesis and spermatogenesis (Wanget al., 1993). The successful production of male gametes requiresextensive and precise interactions between germ cells and the sur-rounding testicular somatic (Sertoli) cells. These interactions aremanifested by the changes in numerous Sertoli cell functions thatparallel the maturation of germ cells as they progress through theseminiferous epithelial cycle (Parvinen, 1982). Testicular dysfunc-tion may result from any disturbance in the normal interaction
Abbreviations: AOa, rgemone oil; AA, argemone alkaloid; SA, sanguinar-ine; TJs, tight junctions; BTB, blood testis barrier; AJs, adherens junctions; ESs,ectoplasmic specializations; EMEM, eagle’s minimum essential medium; MEM, min-imum essential medium; HBSS, hank’s balanced salt solution; FBS, Fetal bovineserum; LDH, lactate dehydrogenase; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-dipehyltetrazolium bromide; DMSO, dimethyl sulphoxide; JC-15, 5′ ,6,6′-tetrachloro-1,1′ ,3,3′-tetraethylbenzimidazol carbocyanine iodide; NADH, nicotinamide adeninedinucleotide, reduced form; NAD+, nicotinamide adenine dinucleotide, oxidizedform; PBS, phosphate buffer saline.
∗ Corresponding author. Tel.: +91 522 2613786; fax: +91 522 2628227.E-mail address: [email protected] (M. Das).
between Sertoli and germ cells responsible for maintaining theintegrity of the germinal epithelium (Foster, 1988). This cellularevent, which regulates the movement as well as the developmentof germ cell, is also accompanied by intermittent disassembly andreassembly of cellular junctions such as tight junctions (TJs) amongSertoli cells that form the blood testis barrier (BTB), the cell–cellactin-based adherens junctions (AJs) between Sertoli and germ cellssuch as ectoplasmic specializations (ESs), a testis-specific AJ type,and cell–cell intermediate filament-based desmosome like junc-tions between Sertoli and germ cells (Russell and Peterson, 1985;Cheng and Mruk, 2002.
Mustard oil from Brassica nigra seeds is the predominant cook-ing medium used in the northern parts of India especially, theGangetic basin (Khan, 2001). Mustard oil is also used conven-tionally for skin and hair massage in this region. Consumptionof argemone contaminated mustard oil even for a short durationleads to a clinical condition collectively referred to as EpidemicDropsy (Sarkar, 1926; Lal and Roy, 1937; Chopra et al., 1939). Sev-eral outbreaks of argemone poisoning have been reported in thedifferent Indian subcontinents as well as other areas of the globeviz a viz Madagascar, Mauritius, Rangoon, and South Africa (Sainani,1976; Das and Khanna, 1997). Though, sporadic incidences of thisdisease have been reported for over last 100 years, the largest epi-demic occurred in 1998 at New Delhi, India affecting thousands of
0378-4274/$ – see front matter © 2009 Elsevier Ireland Ltd. All rights reserved.doi:10.1016/j.toxlet.2009.01.006

Author's personal copy
V. Mishra et al. / Toxicology Letters 186 (2009) 104–110 105
Fig. 1. Structure of sanguinarine and dihydrosanguinarine.
people of which over 3000 people were admitted to various hospi-tals and 65 lost their lives (Das and Khanna, 1997; Das and Khanna,1998). Argemone oil (AO) is known to cause dilatation of the smallarterioles and capillaries, leading to the leakage of serum proteinsresulting in altered osmosis, edema formation and increased cap-illary damage (Chaudhuri, 1959; Das et al., 2005a). Experimentaland clinical studies suggest that reactive oxygen species (ROS) areinvolved in the toxicity of AO, thereby leading to a decrease inthe bio-antioxidant pool and antioxidant defense capacity (Daset al., 1991, 2005a; Upreti et al., 1991). The ROS may also playa vital role in exerting genotoxicity and carcinogenicity of arge-mone oil and sanguinarine (Ansari et al., 2004, 2005; Das et al.,2005b). The toxic effects of AO are attributed to the presence ofinterconvertible methylene dioxy phenyl alkaloids, sanguinarine(SA)/dihydro-sanguinarine (Sarkar, 1948) (Fig. 1), which have alsobeen implicated to cause cellular macromolecular damage (Das etal., 2005a).
It has been reported that there exists a direct correlationbetween genotoxicity and reproductive toxicity for various xeno-biotics (Tekeuchi et al., 1997; Sekiguchi et al., 2002; Yang et al.,2005) For example, acrylamide which has been shown to formmicronuclei and chromosomal aberrations, also resulted in reducedtesticular and epididymidis weights as well as reduced sperm count(Adler et al., 1993; Higashikuni et al., 1994; Yang et al., 2005)Similarly 1- and 2-bromopropane also shows genotoxic and repro-ductively toxic response in Korean workers (Tekeuchi et al., 1997;Sekiguchi et al 2002).
The reduction in male fertility rate which may result fromlowering of sperm counts has been a major concern throughoutthe world (Irvine et al., 1996; Marimuthu et al., 2003; Shanna etal., 2003; Jensen et al., 2002). Reports have suggested that thereduction in male fertility rate may be due to interaction withenvironment or chemical exposure (Sharpe and Skakkebaek, 1993;Danish Environmental Protection Agency, 1995). We argued thatconsumption of adulterated food may be one of the etiological fac-tors. Because of argemone oil and its active ingredient, sanguinarinehas been shown to possess genotoxic potential (Ansari et al., 2004,2005), the effect of these was undertaken on rat Sertoli–germ cellscoculture to evaluate the reproductive toxicity.
2. Materials and methods
2.1. Chemicals
Standard sanguinarine, collagenase (type I), DNAse I, Eagle’s minimum essen-tial medium (EMEM or MEM), Hank’s balanced salt solution (HBSS), fetalbovine serum (FBS), trypan blue, lactate dehydrogenase (LDH), 3-(4,5-dimethyl-thiazol-2-yl)-2,5-dipehyl tetrazolium bromide (MTT), dimethyl sulphoxide (DMSO),5,5′ ,6,6′-tetrachloro-1,1′ ,3,3′-tetraethylbenzimidazol carbocyanine iodide (JC-1),Annexin V-FITC apoptosis detection kit were procured from Sigma–Aldrich, St. Louis,USA. Tissue culture dishes were obtained from Tarson, India.
2.2. Preparation of sanguinarine alkaloid from argemone oil
Argemone mexicana seeds were procured from outskirts of Lucknow city, UttarPradesh, India. The seeds were crushed and the oil was extracted with the helpof Soxhlet apparatus using n-hexane16. The hexane containing argemone oil wasfiltered under vacuum through a Buchner funnel containing glass wool and the sol-vent was distilled at 30 ◦C under vacuum in Buchii Rotavapor-R. The oil obtained wasstored in amber glass bottle under nitrogen atmosphere. The yield of argemone oilfrom its seeds was 35% (v/w). Alkaloid was precipitated as an orange-coloured salt bypassing dry hydrochloride acid gas through argemone oil. The precipitated alkaloidwas purified through resolution on an alumina column followed by crystallizationfrom alcohol:benzene mixture. The alkaloid, sanguinarine content was found to be0.54% in oil (Sarkar, 1948) and purity was assessed.
2.3. Isolation of Sertoli–germ cells from testis of rats
Sertoli–germ cells were isolated from four wistar albino rats of 28 days old.Testes were excised from the capsule. Testes were then chopped with a sharp knifein the HBSS medium. Chopped tissue in medium was centrifuged at 1000 rpm for2 min and pellets were collected. The washing step was repeated and pellet collected(Adhikari et al., 2000).
2.4. Primary culture of Sertoli–germ cells and cell viability assay
Pellet was treated with 30 ml of 0.1% collagenase for 20 min and centrifuged at1000 rpm for 2 min. The resulting pellet was treated with 15 ml of 0.25% trypsinsolution containing 150 �g DNAse for 15 min. Washing was done two times withminimum essential medium (MEM). Finally cells were grown in MEM supplementedwith 10% FBS (Adhikari et al., 2000). Non-exfoliated cocultures (germ cells attachedwith Sertoli cells) were maintained at 37 ◦C with 95% humidity in CO2 incubatorcontaining 5% CO2. The cell density was adjusted to 1.0–1.5 × 106 cells/ml and theviability of the freshly isolated cells was performed by 1:1 dilution of the cell suspen-sion with 0.4% trypan blue solution. This mixture was loaded on to the chambers ofa haemocytometer. After 1–2 min, number of stained cells and total number of cellswere counted in haemocytometer. The number of unstained cells represents viablecells and calculated as percentage of total cells. The non-exfoliated coculture cellsafter 24 h of plating were treated with different concentrations of test sample (AO-0.1 �l/ml, 1.5 �l/ml and 2.5 �l/ml, argemone alkaloid (AA) and SA-1 �g/ml, 15 �g/mland 25 �g/ml, respectively) for 24 h and 48 h time period. The concentrations of AAand SA correspond to the content of the doses used for AO.
2.5. Assessment of germ cell exfoliation
Exfoliated (detached) cells were pelleted at 1000 rpm for 5 min from the mediumand resuspended in an appropriate volume of serum free medium. A known volumewith equal number of cells was poured in the wells of plate along with and withoutthe test sample and detachment of germ cells from the Sertoli–germ cell monolayerwas counted using a haemocytometer (Sinha et al., 1999).
2.6. Viability of detached cells
The viability of detached cells was studied by staining with trypan blue (1:1)and counted by a haemocytometer as described above (Sinha et al., 1999).
2.7. Cell proliferation inhibition assay
The anti-proliferative effects of argemone oil (AO)/alkaloid (AA)/SA onSertoli–germ cells were determined by the MTT dye uptake method (Darzynkiewiczet al., 1992). Briefly, the cells (10,000/well) were incubated in the presence andabsence of test sample (triplicate) in a 96-well plate in a final volume of 0.1 ml forthe indicated time periods at 37 ◦C. A 10 �l MTT (5 mg/ml PBS) was then added tothe wells, and the plates were further incubated for 4 h. The plate was centrifuged at1200 rpm for 10 min and 100 �l of DMSO was added after removing the supernatant,to dissolve the formazan formed. The absorbance was read at 530 nm after 5 min, ina micro plate reader (Synergy HT of BIO-TEK International, USA).
Author's personal copy
106 V. Mishra et al. / Toxicology Letters 186 (2009) 104–110
2.8. Measurement of LDH in the medium
The leakage of the cytosolic enzyme LDH into the medium after argemoneoil/alkaloid/sanguinarine treatment was performed according to the method ofKornberg (Kornberg, 1955). Media from treated and control plates were collectedand centrifuged to pellet the cells. The clear medium was used for the measure-ment of enzyme. Briefly, the measurement of LDH in culture supernatants gives aquantitative value of loss of cell viability. The activity of LDH could be measuredas the reduction of pyruvate to lactate. The reduction is coupled to the oxidationof nicotinamide adenine dinucleotide, reduced form (NADH) to Nicotinamide ade-nine dinucleotide, oxidized form (NAD+), which is followed spectrophotometricallyat 340 nm. Because NADH has a high absorbance at 340 nm compared with NAD+,the reaction is measured as the rate of decrease in absorbance at 340 nm. The LDHrelease was calculated by the formula:
LDH release = (Test Sample − Low Control)(High Control − Low Control)
× 100
Low Control: normal cells in 200 �l assay medium; High Control: normal cells in200 �l assay medium containing 1% Triton X-100; Test Sample: normal cells in 200 �lassay medium containing test substance
2.9. Mitochondrial membrane potential Assay
For the detection of mitochondrial membrane potential, argemone oil/alkaloidand sanguinarine treated cells were incubated with JC-1 (5 �g/ml final concentra-tion) for 20 min in dark at 37 ◦C, harvested and suspended in PBS. The mitochondrialmembrane potential was measured at respective excitation and emission wave-length of 530 and 590 nm, on a micro plate reader (Synergy HT of BIO-TEKInternational, USA).
2.10. Assessment of apoptotic and necrotic cells
The apoptotic and necrotic cell distribution was analysed by Annexin-V bindingand PI uptake. Positioning of quadrants on Annexin-V/PI dot plots was performedand living cells (Annexin-V−/PI−), early apoptotic/primary apoptotic cells (Annexin-V + /PI−), late apoptotic/secondary apoptotic cells (Annexin-V + /PI+) and necroticcells (Annexin-V−/PI+) were distinguished (Vermes et al., 1995). Therefore, thetotal apoptotic proportion included the percentage of cells with fluorescenceAnnexin-V + /PI− and Annexin-V+/PI+. Briefly, the cells were treated with argemoneoil/alkaloid/sanguinarine for 24 h and 48 h. The harvested cells were suspended in1 ml binding buffer (1×). An aliquot of 100 �l was incubated with 5 �l Annexin-V-FITC and 5 �l PI for 15 min in dark at room temperature and 400 �l binding buffer(1×) was added to each sample. The FITC and PI fluorescence were measured throughFL-1 filter (530 nm) and FL-2 filter (585 nm), respectively, and 10,000 events wereacquired on flow cytometer (Becton–Dickinson, USA).
3. Results
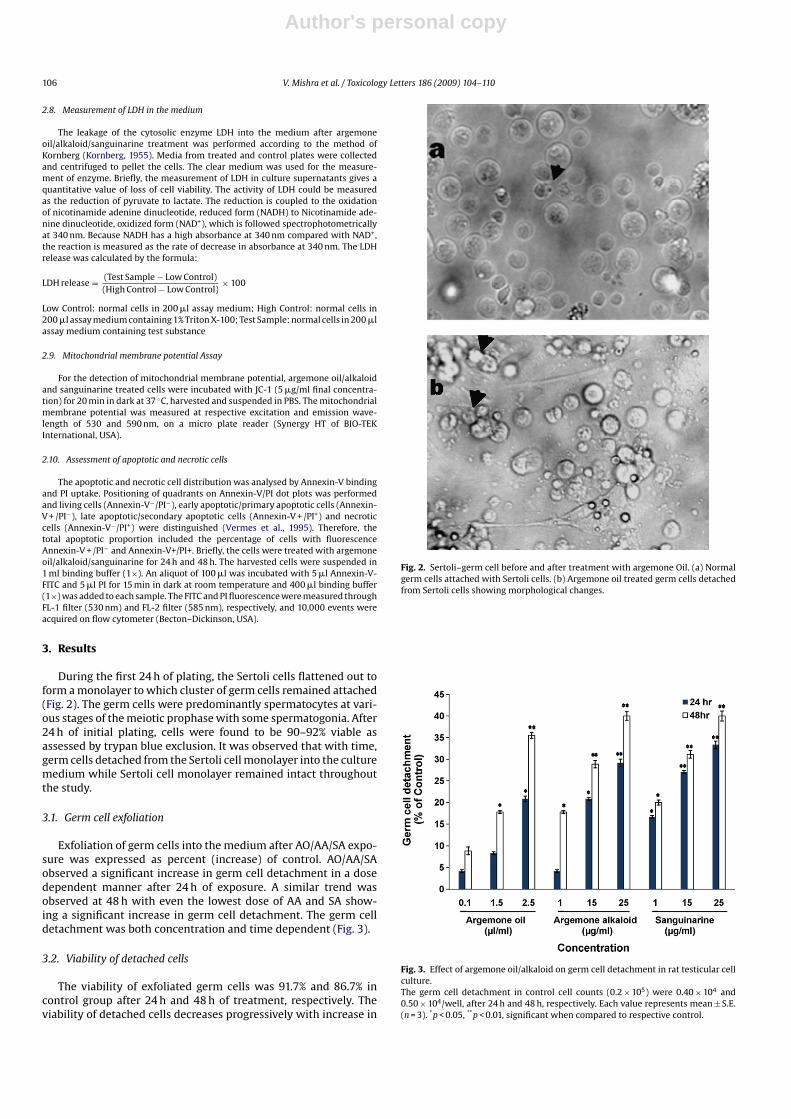
During the first 24 h of plating, the Sertoli cells flattened out toform a monolayer to which cluster of germ cells remained attached(Fig. 2). The germ cells were predominantly spermatocytes at vari-ous stages of the meiotic prophase with some spermatogonia. After24 h of initial plating, cells were found to be 90–92% viable asassessed by trypan blue exclusion. It was observed that with time,germ cells detached from the Sertoli cell monolayer into the culturemedium while Sertoli cell monolayer remained intact throughoutthe study.
3.1. Germ cell exfoliation
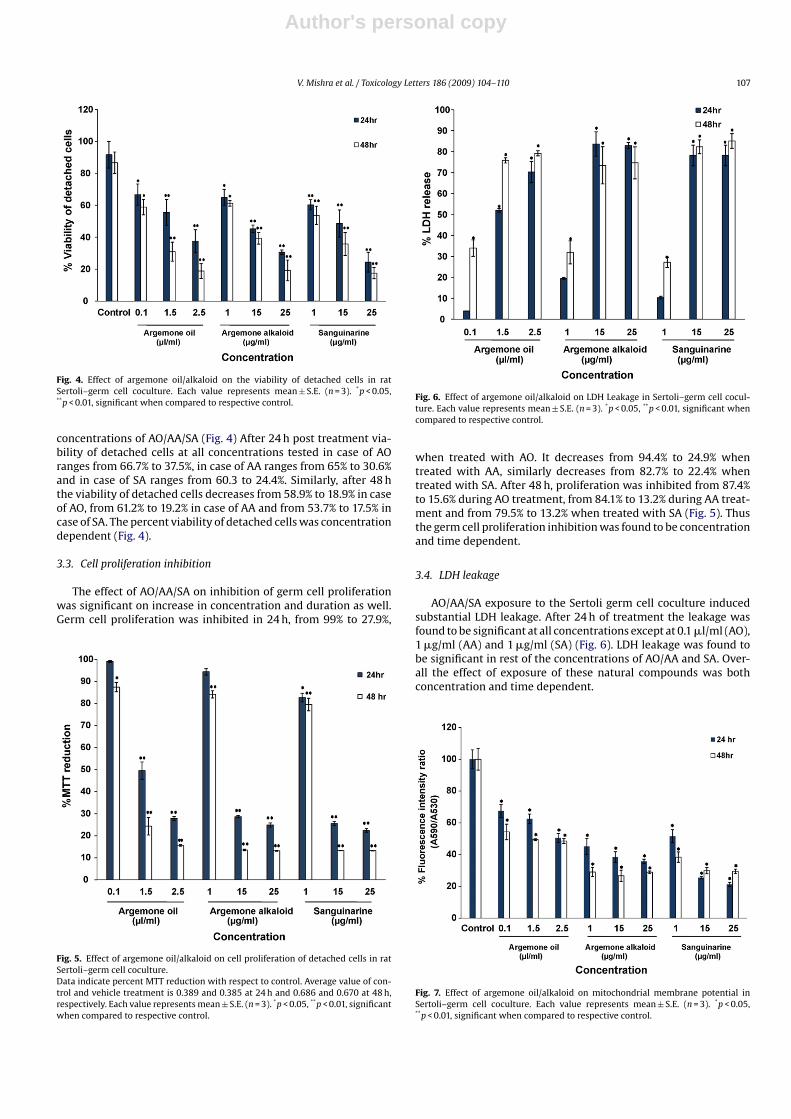
Exfoliation of germ cells into the medium after AO/AA/SA expo-sure was expressed as percent (increase) of control. AO/AA/SAobserved a significant increase in germ cell detachment in a dosedependent manner after 24 h of exposure. A similar trend wasobserved at 48 h with even the lowest dose of AA and SA show-ing a significant increase in germ cell detachment. The germ celldetachment was both concentration and time dependent (Fig. 3).
3.2. Viability of detached cells
The viability of exfoliated germ cells was 91.7% and 86.7% incontrol group after 24 h and 48 h of treatment, respectively. Theviability of detached cells decreases progressively with increase in
Fig. 2. Sertoli–germ cell before and after treatment with argemone Oil. (a) Normalgerm cells attached with Sertoli cells. (b) Argemone oil treated germ cells detachedfrom Sertoli cells showing morphological changes.
Fig. 3. Effect of argemone oil/alkaloid on germ cell detachment in rat testicular cellculture.The germ cell detachment in control cell counts (0.2 × 105) were 0.40 × 104 and0.50 × 104/well, after 24 h and 48 h, respectively. Each value represents mean ± S.E.(n = 3). *p < 0.05, **p < 0.01, significant when compared to respective control.
Author's personal copy
V. Mishra et al. / Toxicology Letters 186 (2009) 104–110 107
Fig. 4. Effect of argemone oil/alkaloid on the viability of detached cells in ratSertoli–germ cell coculture. Each value represents mean ± S.E. (n = 3). *p < 0.05,**p < 0.01, significant when compared to respective control.
concentrations of AO/AA/SA (Fig. 4) After 24 h post treatment via-bility of detached cells at all concentrations tested in case of AOranges from 66.7% to 37.5%, in case of AA ranges from 65% to 30.6%and in case of SA ranges from 60.3 to 24.4%. Similarly, after 48 hthe viability of detached cells decreases from 58.9% to 18.9% in caseof AO, from 61.2% to 19.2% in case of AA and from 53.7% to 17.5% incase of SA. The percent viability of detached cells was concentrationdependent (Fig. 4).
3.3. Cell proliferation inhibition
The effect of AO/AA/SA on inhibition of germ cell proliferationwas significant on increase in concentration and duration as well.Germ cell proliferation was inhibited in 24 h, from 99% to 27.9%,
Fig. 5. Effect of argemone oil/alkaloid on cell proliferation of detached cells in ratSertoli–germ cell coculture.Data indicate percent MTT reduction with respect to control. Average value of con-trol and vehicle treatment is 0.389 and 0.385 at 24 h and 0.686 and 0.670 at 48 h,respectively. Each value represents mean ± S.E. (n = 3). *p < 0.05, **p < 0.01, significantwhen compared to respective control.
Fig. 6. Effect of argemone oil/alkaloid on LDH Leakage in Sertoli–germ cell cocul-ture. Each value represents mean ± S.E. (n = 3). *p < 0.05, **p < 0.01, significant whencompared to respective control.
when treated with AO. It decreases from 94.4% to 24.9% whentreated with AA, similarly decreases from 82.7% to 22.4% whentreated with SA. After 48 h, proliferation was inhibited from 87.4%to 15.6% during AO treatment, from 84.1% to 13.2% during AA treat-ment and from 79.5% to 13.2% when treated with SA (Fig. 5). Thusthe germ cell proliferation inhibition was found to be concentrationand time dependent.
3.4. LDH leakage
AO/AA/SA exposure to the Sertoli germ cell coculture inducedsubstantial LDH leakage. After 24 h of treatment the leakage wasfound to be significant at all concentrations except at 0.1 �l/ml (AO),1 �g/ml (AA) and 1 �g/ml (SA) (Fig. 6). LDH leakage was found tobe significant in rest of the concentrations of AO/AA and SA. Over-all the effect of exposure of these natural compounds was bothconcentration and time dependent.
Fig. 7. Effect of argemone oil/alkaloid on mitochondrial membrane potential inSertoli–germ cell coculture. Each value represents mean ± S.E. (n = 3). *p < 0.05,**p < 0.01, significant when compared to respective control.
Author's personal copy
108 V. Mishra et al. / Toxicology Letters 186 (2009) 104–110
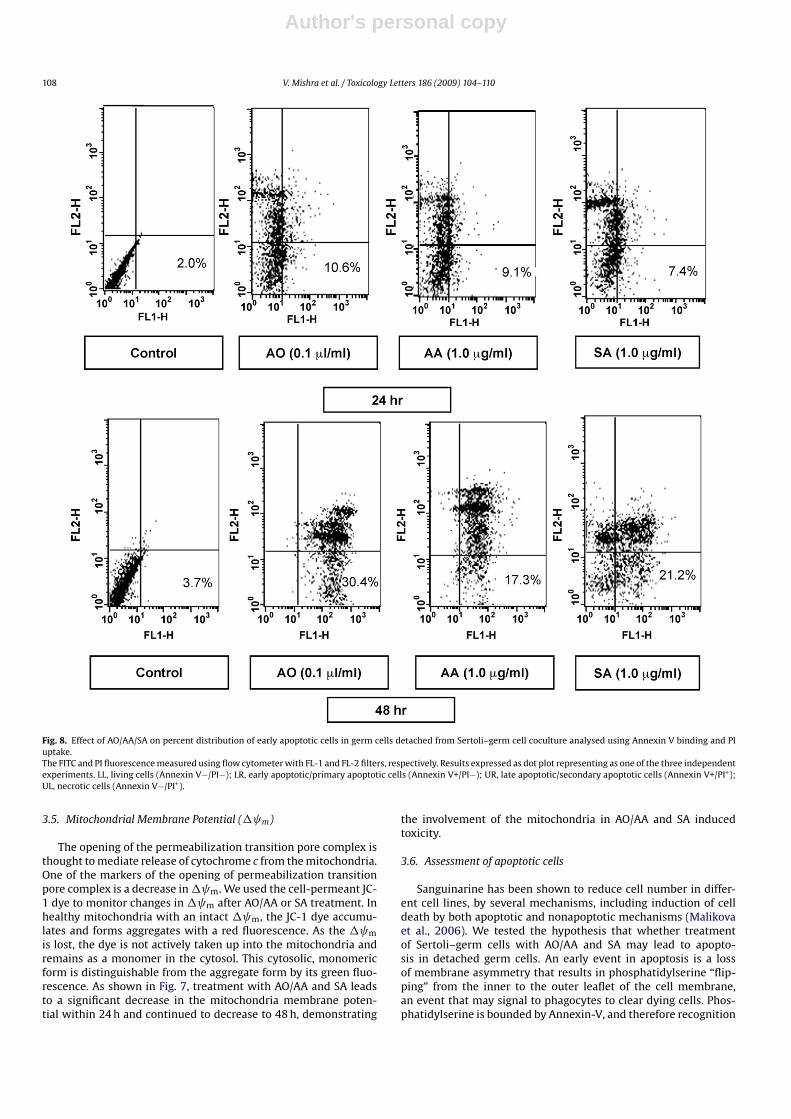
Fig. 8. Effect of AO/AA/SA on percent distribution of early apoptotic cells in germ cells detached from Sertoli–germ cell coculture analysed using Annexin V binding and PIuptake.The FITC and PI fluorescence measured using flow cytometer with FL-1 and FL-2 filters, respectively. Results expressed as dot plot representing as one of the three independentexperiments. LL, living cells (Annexin V−/PI−); LR, early apoptotic/primary apoptotic cells (Annexin V+/PI−); UR, late apoptotic/secondary apoptotic cells (Annexin V+/PI+);UL, necrotic cells (Annexin V−/PI+).
3.5. Mitochondrial Membrane Potential (� m)
The opening of the permeabilization transition pore complex isthought to mediate release of cytochrome c from the mitochondria.One of the markers of the opening of permeabilization transitionpore complex is a decrease in� m. We used the cell-permeant JC-1 dye to monitor changes in� m after AO/AA or SA treatment. Inhealthy mitochondria with an intact � m, the JC-1 dye accumu-lates and forms aggregates with a red fluorescence. As the � m
is lost, the dye is not actively taken up into the mitochondria andremains as a monomer in the cytosol. This cytosolic, monomericform is distinguishable from the aggregate form by its green fluo-rescence. As shown in Fig. 7, treatment with AO/AA and SA leadsto a significant decrease in the mitochondria membrane poten-tial within 24 h and continued to decrease to 48 h, demonstrating
the involvement of the mitochondria in AO/AA and SA inducedtoxicity.
3.6. Assessment of apoptotic cells
Sanguinarine has been shown to reduce cell number in differ-ent cell lines, by several mechanisms, including induction of celldeath by both apoptotic and nonapoptotic mechanisms (Malikovaet al., 2006). We tested the hypothesis that whether treatmentof Sertoli–germ cells with AO/AA and SA may lead to apopto-sis in detached germ cells. An early event in apoptosis is a lossof membrane asymmetry that results in phosphatidylserine “flip-ping” from the inner to the outer leaflet of the cell membrane,an event that may signal to phagocytes to clear dying cells. Phos-phatidylserine is bounded by Annexin-V, and therefore recognition

Author's personal copy
V. Mishra et al. / Toxicology Letters 186 (2009) 104–110 109
of Annexin-V binding via a FITC-labeled Annexin-V serves as a use-ful marker of early apoptotic events. Treatment of Sertoli–germcells with the lowest concentration of AO/AA and SA for 24 hresulted in 5.2-, 4.4- and 3.6-fold increase in the percentage ofAnnexin-(+) cells, respectively (Fig. 8). After 48 h treatment withthe same concentration of AO/AA and SA resulted in a greaterincrease (8.3, 4.75 and 5.81-fold, respectively) in detached germcells undergoing early apoptosis. These data suggest that theincrease in germ cell detachment and decrease in cell viability,are the results of greater induction of apoptosis by AO/AA and SA(Fig. 8).
4. Discussion
Xenobiotics can disturb testicular function by interfering withseveral pathways operating within the testis. Testicular lesionsmay be characterized by disruption of normal interaction betweenSertoli cells and germ cells, a process that is responsible for main-taining the integrity of the epithelium (Ritzen et al., 1981) leadingto early detachment of spermatocytes and spermatids from thegerminal epithelium. The present study suggests that significantshedding of germ cells from the Sertoli cell monolayer occurredwhen cocultures of Sertoli and germ cells were treated with differ-ent concentrations of AO/AA or SA. This suggests that AO/AA or SAmay disrupt the normal interactions between germ cells and Ser-toli cells leading to cell detachment. The increased cell detachmentof germ cells from Sertoli cells has been shown to be responsiblefor disturbed spermatogenesis, decreased sperm count and reducedsperm production rate (Saxena et al., 1987, 1988; Sokol, 1990). Germcell loss from the Sertoli cell monolayer has also been induced by anumber of chemicals which cause testicular damage in vivo. Certainphthalate esters, trifluoro acetaldehyde, methoxy ethoxy acetic acidand N-butoxy acetic acid have been reported to induce concentra-tion dependent germ cell detachment in vitro which correlated wellwith their activity in vivo (Foster et al., 1987; Gray and Beamand,1984; Lloyd et al., 1988). The mechanism by which germ cell exfoli-ation is produced in culture is not yet clear. However, in the presentstudy, it is quite possible that germ cell exfoliation is due to directtoxic action of AO/SA on Sertoli–germ cell coculture, as the differ-ence in the viability of the detached germ cells from the control andtreated plates was significant.
Perturbation of cell membrane integrity and LDH leakage dueto plasma membrane damage are the important parameters forassessing cytotoxicity of chemicals. In the present study AO/AA orSA induced significant LDH leakage and cell membrane integrityloss in the medium at both the time intervals. To investigatethe consequences of separation of germ cells from Sertoli cells,the role of apoptosis in AO/AA/SA induced germ cell death wasstudied by two methods: the mitochondrial membrane potential(� m) and the assessment of apoptotic cells via Annexin-V (FITC).Since mitochondria plays a central role in the intrinsic pathway ofapoptosis, stimuli that activates this pathway leads to increasedpermeability of the outer mitochondrial membrane and decreasedtransmembrane potential, release of cytochrome c, and productionof reactive oxygen species, which ultimately lead to activation ofdownstream effector caspases (Kaufmann and Earnshaw, 2000).Here, we demonstrate that treatment with AO/AA or SA resultsin a decrease in � m at 24 h, and continues to decrease till 48 h.Decrease in � m has been followed by further signs of apoptosisexecution as measured by loss of membrane symmetry (Annexin-V staining) (Amanda et al., 2004). Treatment of Sertoli–germ cellswith AO/AA or SA results in loss of membrane integrity of detachedgerm cells during 24 h and continues to 48 h, which correlates withthe increased loss of cell viability of detached germ cells. It has beenproposed that filament extensions are involved in maintaining the
integrity of Sertoli cell–germ cell contacts (Aumuller et al., 1992).Therefore, it is quite likely that collapse in vimentin filaments maylead to apoptosis in germ cell via disruption in contact mediatedcommunication between the Sertoli cells and germ cells, whichmay subsequently lead to testicular impairment following AO/SAexposure.
Conflict of interest
The authors declare that there are no conflicts of interest.
Acknowledgements
We are grateful to the Director of our Centre for his keen inter-est in the study. One of us (VM) is thankful to Council of Scientificand Industrial Research (CSIR)/University Grant Commission (UGC),New Delhi for the award of Junior Research Fellowship. We are alsothankful to Mr. Ram Narayan for technical assistance and central cellculture laboratory for providing the facility. Financial assistance ofCSIR Supra institutional Project SIP-08 is gratefully acknowledged.The manuscript is IITR communication # 2714.
References
Adhikari, N., Sinha, N., Saxena, D.K., 2000. Effect of lead on Sertoli–germ cell cocultureof rat. Toxicol. Lett. 116, 45–49.
Adler, I.D., Zouh, R., Schmid, E., 1993. Perturbation of cell division by acrylamide invitro and in vivo. Mutat. Res. 301, 249–254.
Amanda, P.S., Bittman, R., Burow, M.E., Clejan, S., Elliott, S., Hammond, T., Tang, Y.,Beckman, B.S., 2004. Novel ceramide analogs as potential chemotherapeuticagents in breast cancer. J. Pharmacol. Exp. Ther. 309, 523–532.
Ansari, K.M., Chauhan, L.K., Dhawan, A., Khanna, S.K., Das, M., 2004. Unequivocal evi-dence of genotoxic potential of argemone oil in mice. Int. J. Cancer 112, 890–895.
Ansari, K.M., Dhawan, A., Khanna, S.K., Das, M., 2005. In vivo DNA damaging potentialof sanguinarine alkaloid, isolated from argemone oil, using alkaline comet assayin mice. Food Chem. Toxicol. 43, 147–153.
Aumuller, G., Schulze, C., Viebahn, C., 1992. Intermediate filaments in sertoli cells.Microsc. Res. Tech. 20, 50–72.
Chaudhuri, R.N., 1959. Clinical and experimental research. Bull. Cal. School Trop.Med. 7, 157–165.
Cheng, C.Y., Mruk, D., 2002. Cell junction dynamics in the testis: their implicationin sertoli–germ cell interactions and male contraceptive development. Physiol.Rev. 82, 825–874.
Chopra, R.N., Pasricha, C.L., Goyal, R.K., Lal, S., Sen, A.K., 1939. The experimental pro-duction of syndrome of Epidemic Dropsy in man. Indian Med. Gaz. 74, 193–198.
Danish Environmental Protection Agency, 1995. Male reproductive health andenvironmental chemicals with oestrogenic effects. Copenhagen: Ministry ofEnvironment and Energy.
Darzynkiewicz, Z., Bruno, S., Del, B.G., Gorczyca, W., Hotz, M.A., Lassota, P., 1992.Features of apoptotic cells measured by flow cytometry. Cytometry 13, 795–808.
Das, M., Ansari, K.M., Dhawan, A., Shukla, Y., Khanna, S.K., 2005b. Correlation of DNAdamage in epidemic dropsy patients to carcinogenic potential of argemone oiland isolated sanguinarine alkaloid in mice. Int. J. Cancer 117, 709–717.
Das, M., Babu, K., Reddy, N.P., Srivastava, L.M., 2005a. Oxidative damage of plasmaproteins and lipids in epidemic dropsy patients: alterations in antioxidant status.Biochim. Biophys. Acta 1722, 209–217.
Das, M., Khanna, S.K., 1998. Epidemic Dropsy. The National Med. J. Ind. 11, 207–208.Das, M., Khanna, S.K., 1997. Clinicoepidemiological, toxicological, and safety evalua-
tion studies on argemone oil. Crit. Rev. Toxicol. 27, 273–297.Das, M., Upreti, K.K., Khanna, S.K., 1991. Biochemical toxicology of argemone oil: role
of reactive oxygen species in iorn catalysed lipid peroxidation. Bull. Environ.Cont. Toxicol. 46, 422–430.
Foster, P.M.D., 1988. Testicular organisation and biochemical function. In: Lamb, J.C.,Foster, P.M.D. (Eds.), Physiology and Toxicology of Male Reproduction. AcademicPress, New York, p. 713.
Foster, P.M.D., Lloyd, S.C., Blackburn, D.M., 1987. Comparison of the in vivo and invitro testicular effects produced by methoxy, ethoxy and N-butoxy acetic acid inthe rat. Toxicology 43, 17–30.
Gray, T.J.B., Beamand, J.A., 1984. Effect of some phthalate esters and other testiculartoxins on primary cultures of testicular cells. Food Chem. Toxicol. 22, 123–131.
Higashikuni, N., Hara, M., Nakagawa, S., Sutou, S., 1994. 2-(2-Furyl)-3-(5-nitro-2-furyl) acrylamide (AF-2) is a weak in vivo clastogen as revealed by themicronucleus assay. Mutat. Res. 320, 149–156.
Irvine, S., Cawood, E., Richardson, D., MacDonald, E., Aitken, J., 1996. Evidence ofdeteriorating semen quality in the United Kingdom: birth cohort study in 577men in Scotland over 11 years. Br. Med. J. 312, 467–471.
Jensen, T.K., Carlsen, E., Jorgensen, N., Berthelsen, J.G., Keiding, N., Christensen, K.,2002. Poor semen quality may contribute to recent decline in fertility rates.Hum. Reprod. 17, 1437–1440.

Author's personal copy
110 V. Mishra et al. / Toxicology Letters 186 (2009) 104–110
Kaufmann, S.H., Earnshaw, W.C., 2000. Induction of apoptosis by cancer chemother-apy. Exp. Cell Res. 256, 42–49.
Khan, J.S., 2001. Blended mustard oil for health benefits. In: Bhatnagar, A.K., Singh,H.B., Prakash, S. (Eds.), Mustard Research and Promotion Consortium New Delhi,pp. 132–138.
Kornberg, A., 1955. Lactate dehydrogenase of muscle. In: Colowick, S.P., Kaplan, N.O.(Eds.), Methods in Enzymology, 1. Academic Press, New York, pp. 441–454.
Lal, R.B., Roy, S.C., 1937. Investigation into epidemiology of Epidemic Dropsy. I. Intro-ductory notes and historical survey. Indian Med. Gaz. 25, 163–167.
Lloyd, S.C., Blackburn, D.M., Foster, P.M.D., 1988. Triflouoroethanol and its oxidativemetabolite. Comparison of in vivo and in vitro effects in rat testis. Toxicol. Appl.Pharmacol. 92, 390–401.
Malikova, J., Zdarilova, A., Hlobilkova, A., 2006. Effects of sanguinarine and chelery-thrine on the cell cycle and apoptosis. Biomed. Pap. Med. Fac. Palacky Olomouc.Czech. Repub. 150, 5–12.
Marimuthu, P., Kapilashrami, M.C., Misra, M.M., Singh, G., 2003. Evaluation of trendin semen analysis for 11 years in subjects attending a fertility clinic in India.Asian J Androl. 5, 221–225.
Parvinen, M., 1982. Regulation of the seminiferous epithelium. Endocr. Rev. 3,404–417.
Ritzen, E.M., Hansson, V., French, F.S., 1981. The Sertoli cell. In: Burger, H., Kretser,D.D. (Eds.), The Testis. Raven Press, New York, p. 177.
Russell, L.D., Peterson, R.N., 1985. Sertoli cell junctions: morphological and functionalcorrelates. Int. Rev. Cytol. 94, 177–211.
Sainani, G.S., 1976. Epidemic Dropsy. In: Ahuja, M.M.S (Ed.), Progress In ClinicalMedicine In India, Ist ed. Arnold Heineman, New Delhi, pp. 92–105.
Sarkar, S.L., 1926. Katakar Oil Poisoning. Indian Med. Gaz 61, 62–63.Sarkar, S.N., 1948. Isolation from argemone oil of dihydrosanguinarine and san-
guinarine: toxicity of sanguinarine. Nature 162, 265–266.
Saxena, D.K., Lal, B., Chandra, S.V., 1988. Age dependent testicular changes in leadexposed rats. J. Environ. Biol. 9, 218–231.
Saxena, D.K., Srivastava, R.S., Lal, B., Chandra, S.V., 1987. The effect of lead exposureon the testis of growing rats. Exp. Pathol. 31, 249–252.
Sekiguchi, S., Suda, M., Zhai, Y.L., Honma, T., 2002. Effects of 1-bromopropane, 2-bromopropane, and 1, 2-dichloropropane on the estrous cycle and ovulation inF344 rats. Toxicol. Lett. 126, 41–49.
Shanna, H.S., Charlene, B., Erma, Z.D., Liu, F., Kruse, R.L., Hatch, M., Redmon, J.B., Wang,C., Overstreet, J.W., 2003. Geographic differences in semen quality of fertile US.males. Environ. Health Perspect. 111, 414–420.
Sharpe, R.M., Skakkebaek, N.E., 1993. Are oestrogens involved in falling sperm countsand disorders of male reproductive tract? Lancet 341, 1392–1396.
Sinha, N., Adhikari, N., Narayan, R., Saxena, D.K., 1999. Cytotoxic effect of endosulfanon rat Sertoli–germ cell coculture. Reprod. Toxicol. 13, 291–294.
Sokol, R.Z., 1990. The effect of duration of exposure on the expression of lead toxicityon the male reproductive axis. J. Androl. 11, 521–526.
Tekeuchi, Y., Ichihara, G., Kamijima, M., 1997. A review on toxicity of 2-bromopropane: Mainly on its reproductive toxicity. J. Occup. Med. 39, 179–191.
Upreti, K.K., Das, M., Khanna, S.K., 1991. Role of antioxidants and scavengers onargemone oil induced toxicity. Arch. Environ. Contam. Toxicol. 20, 531–537.
Vermes, I., Haanen, C., Steffens-Nakken, H., Reutelingsperger, C., 1995. A novel assayfor apoptosis. Flow cytometric detection of phosphatidylserine expression onearly apoptotic cells using fluorescein labeled Annexin V. J. Immunol. Methods184, 39–45.
Wang, C., Leung, A., Sinha-Hikim, A.P., 1993. Reproductive aging in the male Brown-Norway rat: a model for the human. Endocrinology 133, 2773–2781.
Yang, H.J., Lee, S.H., Yong, J., Choi, J.H., Chang, H.H., Lee, M.H., 2005. Genotoxicity andtoxicological effects of acrylamide on reproductive system in male rats. J. Vet.Sci. 6, 103–110.