disruption of grazer populations as a contributing factor to the initiation of the te brown tide...
TRANSCRIPT

Lhnrrol. Ocerrno~,:. 42(5, part 2), 1997, 1215-1222
0 1997, by the American Socicly or Llnmology and Oceanography, Inc.
Disruption of grazer populations as a contributing factor to the initiation of the Te brown tide algal bloom
xas
Edward J. Buskey, Paul A. Montagna, Anthony F. Amos, and Terry E. Whitledge Marine Science Institute, 750 Channelview Dr., University of Texas at Austin, Port Aransas, Texas 78373-5015
Abstract Disruption of grazer populations, or mismatches between phytoplankton blooms and population dynamics of their
planktonic and benthic grazers, may play an important role in the initiation of some harmful algal blooms. The Laguna Madre of Texas has experienced a persistent, nearly monospecific phytoplankton bloom since January 1990 commonly referred to as the “brown tide.” Whole-water samples collected in the area where the bloom began reveal the pattern of bloom initiation and dispersion during the first few months of the bloom. Enumeration of microzooplankton from these same samples also reveals that protozoan grazer populations were depressed and nearly eliminated before the bloom began. Benthic biomass and species diversity also declined before the onset of the bloom. The disruption of planktonic and benthic grazers may have been due to extreme hypersaline conditions caused by an extended period of drought (salinities > 60%0). A massive fish kill caused by an abnormal period of below freezing temperatures released a pulse of nutrients into an already disturbed environment allowing this nuisance bloom to become established. Planktonic and benthic grazers have been unable to contribute to the demise of this bloom, which has persisted for >7 yr without interruption.
Harmful algal blooms in estuarine and neritic environ- ments have been increasing in frequency in recent years (An- derson 1989; Smayda 1989; Hallegraeff 1993); this increase has been related to the increasing anthropogenic influences on coastal waters, especially alterations in nutrient supplies and nutrient ratios in coastal waters. The role of phytoplank- ton grazers in the population dynamics of these blooms is not well understood, but it seems likely that declines in graz- er populations may contribute to the initiation of some harm- ful algal blooms and that grazers and pathogens may play a role in terminating blooms.
Zooplankton grazing has often been suggested to play an important role in phytoplankton bloom dynamics. Early studies focused on copepods and diatom blooms. Fleming (1939) suggested that diatom populations could be controlled by grazers, and Riley (1946; 1947) produced a mathematical model that demonstrated the potential of grazer populations to maintain low populations of phytoplankton. Martin (1965) suggested that the spring bloom of diatoms in Narragansett Bay was able to become established in part due to low abun- dances of grazers and that the winter-spring bloom ended when rising spring temperatures become favorable to zoo- plankton reproduction. Benthic grazers may also help control phytoplankton biomass, especially in shallow environments (Cloern 1982; Officer et al. 1982). However, remarkably lit- tle is known about the role of grazers in the bloom dynamics of harmful algal bloom species.
A dense bloom of a small unidentified alga has persisted
Acknowledgments This work was supported by the Texas Higher Education Coor-
dinating Board (grants 3658-264, TATP-426, and 003658-019) and by the National Science Foundation (OCE 95-29750).
Some field samples were collected by Dean Stockwell and Rick Kalke. Technical assistance was provided by Cammie Hyatt, Jay Peterson, Chris Collumb, and Mary Conley.
This is University of Texas Marine Science Institute Contribution 975.
uninterrupted in Laguna Madre, Texas, since January 1990. The organism responsible for this lengthy bloom is a small phytoplankton species (4-5-pm diam) which is presently be- ing formally described (DeYoe et al. in prep.). It is similar in morphology and pigments to the northeast “brown tide” organism Aureococcus anopha&erens (Stockwell et al. 1993) which has been responsible for recurrent blooms in Narragansett Bay and Long Island Sound (Cosper et al. 1987). Molecular data from 18s rRNA sequences indicate that both species belong to the newly recognized class Pe- lagophyceae (Anderson et al. 1993), but the two species are different enough to be placed in separate genera (DeYoe et al. 1995). This alga is usually present at densities ranging from 0.5 to 5X106 cells ml-’ (Buskey et al. 1996). Laguna Madre is a large (2.15 X 10” ha), shallow (avg depth, 1.2 m), often hypersaline coastal lagoon (Armstrong 1987) that con- tains extensive seagrass habitat (Quammen and Onuf 1993). The brown tide bloom has severely discolored the previously clear waters of Laguna Madre and reduced the distribution of seagrasses in deeper waters due to attenuation of light (Dunton 1994; Onuf 1996). Unlike blooms of A. anophag- efferens, which have appeared sporadically in several loca- tions for periods of weeks to months, the Texas brown tide has persisted without interruption for >7 yr.
Most field studies of toxic or nuisance phytoplankton blooms begin well after the initiation of the bloom, making it difficult to assess the environmental and biological factors that may have contributed to the initiation of the bloom. A previously published study (Buskey and Stockwell 1993) ex- amined the effects of the brown tide bloom on zooplankton populations in lower Baffin Bay and upper Laguna Madre after the bloom had spread to these regions in June 1990. Analysis of previously unexamined water samples from up- per Baffin Bay has allowed us to identify the area where the bloom began and the time of the bloom’s initiation to within a month. By examining the changes in physical and biolog- ical parameters leading up to the initiation of the bloom, an understanding of factors contributing to its initiation can be
1215

1216 Buskey et al.
reached. A period of drought in the region caused increases in salinity up to 70%0 in some parts of Laguna Madre. This increase in salinity corresponds to a decline in microzoo- plankton populations preceding the bloom, which would be major grazers of phytoplankton in this size range. There was .also a concordant decrease in benthic filter-feeders prior to the bloom. The decline of grazer populations may have helped this bloom become established.
Although the first signs of the alga can be recognized from samples collected in August 1989 (thousands of cells per ml), the bloom did not reach high density (millions of cells per ml) until after an unusually severe freeze in December 1989. Freezes are periodic but infrequent occurrences in the south Texas coastal area. Occasionally, exceptionally strong cold fronts cause dramatic changes in air temperature and accompanying strong winds rapidly cool shallow waters, causing kills of, marine organisms. Should one front follow soon after another, the effects are even more pronounced. Such was the case in December 1989, with a massive die- off of fish and other marine organisms following cold fronts on 16 and 23 December (DeYoe and Suttle 1994).
Materials and methods
The intensity of the December 1989 freeze was recorded with General Oceanics model 6011 mark II current meters which measure current speed and direction, water tempera- ture, and conductivity. These were located at the Aransas Pass Ship Channel at the University of Texas Marine Science Institute Pier Laboratory, the Gulf Intracoastal Waterway at the northern end of the Laguna Madre (Sta. 1) and in the upper Laguna Madre near the mouth of Baffin Bay (Sta. 9; Fig. 1). A weather station was located at Sta. 9. Data on all instruments were collected every 10 min.
Temperature and salinity of surface and bottom waters were measured in the field with a SeaBird model SBE 19 CTD profiler. Discrete water samples for nutrient analysis were collected immediately below the surface by hand. The samples were collected in clean polyethylene bottles and im- mediately chilled with ice and held in the dark. Nutrient analyses were determined on the chilled samples with a seg- mented-flow automated chemical analyzer by methods de- veloped for marine and estuarine waters (Whitledge et al. 198 I). Replicate subsurface whole-water samples were col- lected at 14 stations in Baffin Bay and the upper Laguna Madre (Fig. I). Samples were preserved with - 10% acid Lugol’s iodine (Stoecker et al. 1994).
Estimates of brown tide cell densities were based on rep- licate cell counts using a hemocytometer and a Wild m20 microscope at 200X magnification for dense samples col- lected after January 1990. Microzooplankton (protozoa, ro- tifers, copepod nauplii) and lower density brown tide sam- ples (August-December 1989) were enumerated from whole-water samples by the Utermahl method, in which 5- 20 ml of sample (depending on plankton density) was settled into a well slide and counted with an inverted microscope. The brown tide alga cannot be positively identified with light microscopy based on morphological characteristics alone. Cells counted as brown tide possessed their characteristic
Fig. 1. Map of Baffin Bay and the upper Laguna Madrc show- ing the sampling sites.
size and shape, but these counts may be subject to error, especially at low densities before the bloom began.
Benthic macrl3faunal abundance and community structure were measured ;it four stations: 8 in the grass bed (8g), 8 in the sand (8s), 9, and 15 (Fig. 1). These stations are synon- ymous with 18Sfg, 189s, 6, and 24 in previous publications (Montagna et al. 1993) The benthos were analyzed from sed- iment cores takl=n by hand by divers within a 2-m radius (Montagna and Kalke 1992). The cores were 6.7-cm diam- eter, covering an area of 35.4 cm2, and taken to a depth of 10 cm. Animals were extracted with a 0.5~mm-mesh sieve, and identified to species level. Diversity was calculated with Hill’s diversity number one (N I) (Hill 1973)-a measure of the effective number of species in a sample-and indicates the number of abundant species. It is calculated as the ex- ponentiated form of the Shannon diversity index (H’), where Nl = exp H’. As diversity decreases, N 1 will tend toward 1. The Shannon index is the average uncertainty per species in an infinite community made up of species with known proportional abundances (Shannon and Weaver 1949).
Results
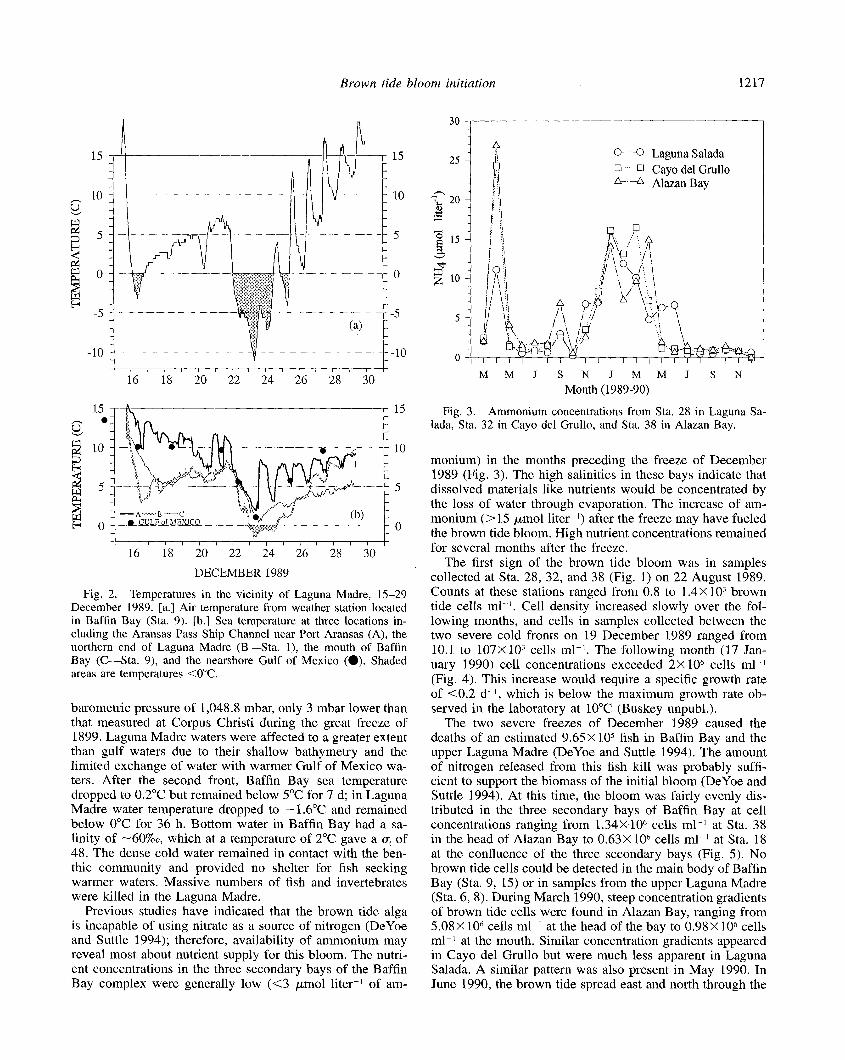
The first cold front on 15-l 6 December 1989 dropped air temperature by 24” overnight to a low of -3°C at Baffin Bay (Fig. 2). Air temperature remained below freezing for 16 h on 16 December. The next front passed through on December 21. The low air temperature at Baffin Bay was - 11.1 “C early in the morning of 23 December. Air tempcr- ature remained below freezing for 33.8 h at Baffin Bay. The swash zone on gulf beaches was frozen and the margins of all bays were frozen for several meters from shore. This cold front was one of the strongest on record, with a surface ’
Brown tide bloom initiation 1217
-10 _-- -- ---- - - --- -_
,,,,,I I I I I I I I I I I
16 18 20 22 24 26 28 30
15
10
5
0
-5
-10
--__--
II I I I I I II I I I III
16 18 20 22 24 26 28 30
DECEMBER 1989
Fig. 2, Temperatures in the vicinity of Laguna Madre, 15-29 December 1989. [a.] Air temperature from weather station located in Bafiin Bay (Sta. 9). Lb.] Sea temperature at three locations in- cluding the Aransas Pass Ship Channel near Port Aransas (A), the northern end of Laguna Madre (B-Sta. l), the mouth of Baffin Bay (C-Sta. 9), and the nearshore Gulf of Mexico (0). Shaded areas are temperatures <O”C.
barometric pressure of 1,048.8 mbar, only 3 mbar lower than that measured at Corpus Christi during the great freeze of 1899. Laguna Madre waters were affected to a greater extent than gulf waters due to their shallow bathymetry and the limited exchange of water with warmer Gulf of Mexico wa- ters. After the second front, Baffin Bay sea temperature dropped to 0.2”C but remained below 5°C for 7 d; in Laguna Madre water temperature dropped to - 1.6”C and remained below 0°C for 36 h. Bottom water in Baffin Bay had a sa- linity of -60%0, which at a temperature of 2°C gave a UV of 48. The dense cold water remained in contact with the ben- thic community and provided no shelter for fish seeking warmer waters. Massive numbers of fish and invertebrates were killed in the Laguna Madre.
Previous studies have indicated that the brown tide alga is incapable of using nitrate as a source of nitrogen (DeYoe and Suttle 1994); therefore, availability of ammonium may reveal most about nutrient supply for this bloom. The nutri- ent concentrations in the three secondary bays of the Baffin Bay complex were generally low (<3 firno1 liter-’ of am-
o--O Laguna Salada q . . -a Cayo de1 Gmllo A--LJ Alazan Bay
MMJSNJMMJSN Month (1989-90)
Fig. 3. Ammonium concentrations from Sta. 28 in Laguna Sa- lada, Sta. 32 in Cayo de1 Grullo, and Sta. 38 in Alazan Bay.
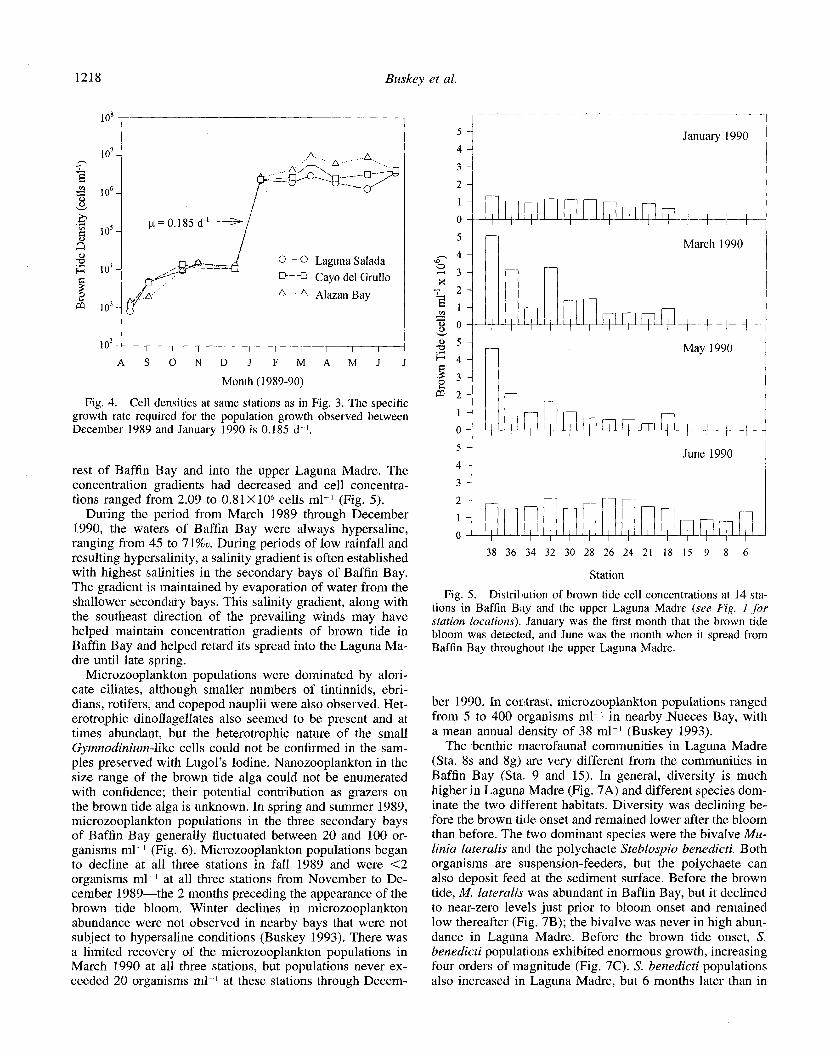
monium) in the months preceding the freeze of December 1989 (Fig. 3). The high salinities in these bays indicate that dissolved materials like nutrients would be concentrated by the loss of water through evaporation. The increase of am- monium (> 15 pmol liter I) after the freeze may have fueled the brown tide bloom. High nutrient concentrations remained for several months after the freeze.
The first sign of the brown tide bloom was in samples collected at Sta. 28, 32, and 38 (Fig. 1) on 22 August 1989. Counts at these stations ranged from 0.8 to 1.4X 1 O3 brown tide cells ml-‘. Cell density increased slowly over the fol- lowing months, and cells in samples collected between the two severe cold fronts on 19 December 1989 ranged from 10.1 to 107X lo3 cells ml-‘. The following month (17 Jan- uary 1990) cell concentrations exceeded 2X 1 O6 cells ml I (Fig. 4). This increase would require a specific growth rate of CO.2 d- I, which is below the maximum growth rate ob- served in the laboratory at 10°C (Buskey unpubl.).
The two severe freezes of December 1989 caused the deaths of an estimated 9.65X lo5 fish in Baffin Bay and the upper Laguna Madre (DeYoe and Suttle 1994). The amount of nitrogen released from this fish kill was probably suffi- cient to support the biomass of the initial bloom (DeYoe and Suttle 1994). At this time, the bloom was fairly evenly dis- tributed in the three secondary bays of Baffin Bay at cell concentrations ranging from 1.34X 10h cells ml-l at Sta. 38 in the head of Alazan Bay to 0.63 X lo6 cells ml I at Sta. 18 at the confluence of the three secondary bays (Fig. 5). No brown tide cells could be detected in the main body of Baffin Bay (Sta. 9, 15) or in samples from the upper Laguna Madre (Sta. 6, 8). During March 1990, steep concentration gradients of brown tide cells were found in Alazan Bay, ranging from 5.08X lo6 cells ml I at the head of the bay to 0.98X 10” cells ml-’ at the mouth. Similar concentration gradients appeared in Cayo de1 Grullo but were much less apparent in Laguna Salada. A similar pattern was also present in May 1990. Tn June 1990, the brown tide spread east and north through the
1218 Buskey et al.
.Q 2 IO5
p= 0.185 d-’ +
a” I @--o Laguna Salada 0-u Cayo de1 Grullo
A A Alazan Bay
lo2 ] I I I I I I I I I I ASONDJFMAMJJ
Month (1989-90)
Fig. 4. Cell densities at same stations as in Fig. 3. The specific growth rate required for the population growth observed between December 1989 and January 1990 is 0.185 d-l.
rest of Baffin Bay and into the upper Laguna Madre. The concentration gradients had decreased and cell concentra- tions ranged from 2.09 to 0.8 1 X lo6 cells ml-l (Fig. 5).
During the period from March 1989 through December 1990, the waters of Baffin Bay were always hypersaline, ranging from 45 to 71%0. During periods of low rainfall and resulting hypersalinity, a salinity gradient is often established with highest salinities in the secondary bays of Baffin Bay. The gradient is maintained by evaporation of water from the shallower secondary bays. This salinity gradient, along with the southeast direction of the prevailing winds may have helped maintain concentration gradients of brown tide in Baffin Bay and helped retard its spread into the Laguna Ma- dre until late spring.
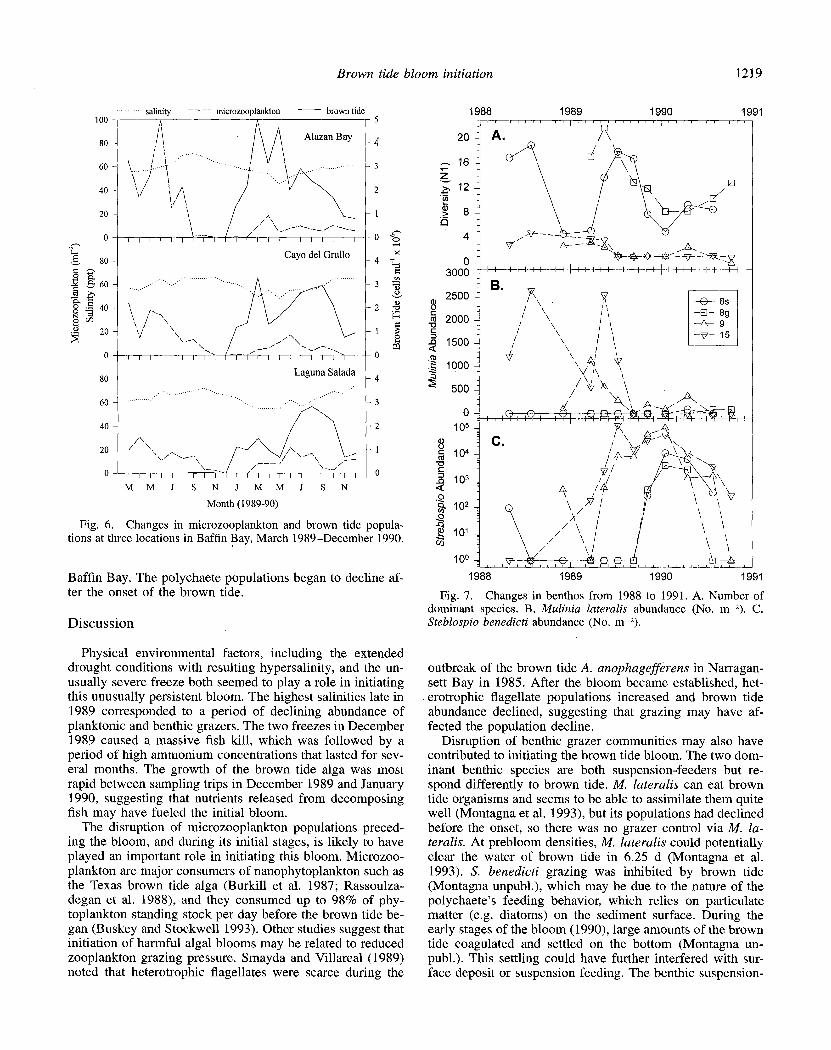
Microzooplankton populations were dominated by alori- cate ciliates, although smaller numbers of tintinnids, ebri- dians, rotifers, and copepod nauplii were also observed. Het- erotrophic dinoflagellates also seemed to be present and at times abundant, but the heterotrophic nature of the small Gymnodinium-like cells could not be confirmed in the sam- ples preserved with Lugol’s Iodine. Nanozooplankton in the size range of the brown tide alga could not be enumerated with confidence; their potential contribution as grazers on the brown tide alga is unknown. In spring and summer 1989, microzooplankton populations in the three secondary bays of Baffin Bay generally fluctuated between 20 and 1.00 or- ganisms ml- I (Fig. 6). Microzooplankton populations began to decline at all three stations in fall 1989 and were <2 organisms ml I at all three stations from November to De- cember 1989-the 2 months preceding the appearance of the brown tide bloom. Winter declines in microzooplankton abundance were not observed in nearby bays that were not subject to hypersaline conditions (Buskey 1993). There was a limited recovery of the microzooplankton populations in March 1990 at all three stations, but populations never ex- ceeded 20 organisms ml- I at these stations through Decem-
January 1990
March 1990
May 1990
+--H-+-
June 1990
38 36 34 32 30 28 26 24 21 18 15 9 8 6
Station
Fig. 5. Distritlution of brown tide cell concentrations at 14 sta- tions in Baffin Bay and the upper Laguna Madre (see Fig. I for station locations). January was the first month that the brown tide bloom was detected, and June was the month when it spread from Baffin Bay throughout the upper Laguna Madre.
ber 1990. In cortrast, microzooplankton populations ranged from 5 to 400 organisms ml-l in nearby Nueces Bay, with a mean annual density of 38 ml-’ (Buskey 1993).
The benthic macrofaunal communities in Laguna Madre (Sta. 8s and 8g) are very different from the communities in Baffin Bay (Sta. 9 and 15). In general, diversity is much higher in Laguna Madre (Fig. 7A) and different species dom- inate the two different habitats. Diversity was declining be- fore the brown tide onset and remained lower after the bloom than before. The two dominant species were the bivalve Mu- linia lateralis and the polychaete Steblospio benedicti. Both organisms are suspension-feeders, but the polychaete can also deposit feed at the sediment surface. Before the brown tide, M. lateralis was abundant in Baffin Bay, but it declined to near-zero levels just prior to bloom onset and remained low thereafter (Fig. 7B); the bivalve was never in high abun- dance in Laguna Madre. Before the brown tide onset, S. benedicti populatj ons exhibited enormous growth, increasing four orders of magnitude (Fig. 7C). S. benedicti populations also increased in Laguna Madre, but 6 months later than in
Brown tide bloom initiation 1219
“’ salinity - --- microzooplankton - brown tide I A A , 5
1988 1989 1990 1991
/\A Alazan Bay c
4
0+
Cayo de1 Grullo
80 -
I
. . . . . . .’ 60 “”
. . .’ ‘.... ,, ;’
40 -I / \ l-2
MMJSNJMMJSN
Monlh (1989-90)
Fig. 4. Changes in microzooplankton and brown tide popula- tions at three locations in Baffin Bay, March 1989-December 1990.
Baffin Bay. The polychaete populations began to decline af- ter the onset of the brown tide.
Discussion
Physical environmental factors, including the extended drought conditions with resulting hypersalinity, and the un- usually severe freeze both seemed to play a role in initiating this unusually persistent bloom. The highest salinities late in 1989 corresponded to a period of declining abundance of planktonic and benthic grazers. The two freezes in December 1989 caused a massive fish kill, which was followed by a period of high ammonium concentrations that lasted for sev- eral months. The growth of the brown tide alga was most rapid between sampling trips in December 1989 and January 1990, suggesting that nutrients released from decomposing fish may have fueled the initial bloom.
The disruption of microzooplankton populations preced- ing the bloom, and during its initial stages, is likely to have played an important role in initiating this bloom. Microzoo- plankton are major consumers of nanophytoplankton such as the Texas brown tide alga (Burkill et al. 1987; Rassoulza- degan et al. 1988), and they consumed up to 98% of phy- toplankton standing stock per day before the brown tide be- gan (Buskey and Stockwell 1993). Other studies suggest that initiation of harmful algal blooms may be related to reduced zooplankton grazing pressure. Smayda and Villareal (1989) noted that heterotrophic flagellates were scarce during the
$j 1000
s 500
0 105
8 gj IO4 7 2 IO3 a
1988 1989 1990 1991
Fig. 7. Changes in benthos from 1988 to 1991. A. Number of dominant species. B. Mulinia lateralis abundance (No. m *). C. Steblospio benedicti abundance (No. m2).
outbreak of the brown tide A. anophageffkrens in Narragan- sett Bay in 1985. After the bloom became established, het- erotrophic flagellate populations increased and brown tide abundance declined, suggesting that grazing may have af- fected the population decline.
Disruption of benthic grazer communities may also have contributed to initiating the brown tide bloom. The two dom- inant benthic species are both suspension-feeders but re- spond differently to brown tide. M. lateralis can eat brown tide organisms and seems to be able to assimilate them quite well (Montagna et al. 1993), but its populations had declined before the onset, so there was no grazer control via M. Za- teralis. At prebloom densities, 44. lateralis could potentially clear the water of brown tide in 6.25 d (Montagna et al. 1993). S. benedicti grazing was inhibited by brown tide (Montagna unpubl.), which may be due to the nature of the polychaete’s feeding behavior, which relies on particulate matter (e.g. diatoms) on the sediment surface. During the early stages of the bloom (1990), large amounts of the brown tide coagulated and settled on the bottom (Montagna un- publ.). This settling could have further interfered with sur- face deposit or suspension feeding. The benthic suspension-

1220 Buskey et al.
feeders that could have consumed brown tide algae seem to have suffered, leading to a breakdown in grazer control. The bivalve that could eat the alga was not present, and the poly- chaete that could not eat it was present but still suffered population declines.
The growth of an algal species must exceed the losses to sinking, mixing, advection, disease, grazing, and other sources of mortality if bloom formation is to occur. The La- guna Madre is a shallow system (avg depth, - 1.2 m) with little water exchange with the Gulf of Mexico. Turnover time for the waters of the Laguna Madre are estimated to be >l yr (Shormann 1992), so losses of phytoplankton due to the physical effects of mixing and advection are extremely small. Losses to sinking are resuspended, by wind-driven mixing in these shallow waters. Planktonic grazers have had only a minor impact on Texas brown tide algal populations. The spread of the established brown tide bloom into the upper Laguna Madre was followed by substantial declines in populations of both mesozooplankton, dominated by the copepod Acartia tonsa, and microzooplankton (Buskey and Stockwell 1993). Reduction in gut pigment content, adult body size, and egg release rates for A. tonsa indicate that the brown tide was an inadequate food and was not heavily grazed (Buskey and Stockwell 1993). Field incubations us- ing the dilution method (Landry and Hassett 1982) indicate that community grazing rates were low for microzooplank- ton populations on the brown tide algae (Buskey and Stock- well 1993). Laboratory studies also support the nutritional inadequacy of this alga to several species of planktonic graz- ers; several protozoan species grow poorly on the brown tide alga compared to other similarly sized species and growth is inhibited for other species in its presence (Buskey and Hyatt 1995). The inability of grazers to terminate this bloom in >7 yr supports the notion that grazers cannot eliminate a well-established bloom that has sufficient resources to sup- port phytoplankton growth.
There is conflicting evidence on the role of grazers in controlling established blooms of dinoflagellates; some stud- ies conclude that, when abundant, grazers could potentially control dinoflagellate blooms (e.g. Turner and Anderson 1983; Watras et al. 1985; Sellner et al. 1991) and others suggest a lesser impact of planktonic grazers (e.g. Sellner and Olson 1985; Sellner and Brownlee 1990). Similarly, there has been conflicting evidence of the role of grazers in the dynamics of Phaeocystis blooms. Verity and Smayda (1989) concluded that grazing by Acartia and similarly sized copepods had little effect on Phaeocystis blooms because colonies were generally too large to consume and solitary cells were too small. However, these copepods may indi- rectly contribute to Phaeocystis blooms by removing com- peting diatoms. Protozoan grazers may, however, be impor- tant grazers on solitary Phaeocystis cells. (Admiraal and Venekamp 1986; Weisse and Schefffel-Moser 1990), and co- pepod grazing on protozoa can reduce grazing pressure and enhance Phaeocystis blooms (Hansen et al. 1993). Uye (1986) estimated the grazing impact of the copepod com- munity on blooms of Chattonella antiqua, based on natural abundances of C. antiqua and its copepod grazers, and lab- oratory-determined grazing rates. Under peak bloom condi- tions, copepods would have little impact on the bloom; how-
ever, Uye concluded that grazing might be important during the initial phases of a bloom.
From an ecological viewpoint, it seems odd that assem- blages of multiple species of algae are the rule in marine planktonic envil-onments, which appear to be homogeneous. This is Hutchinson’s (196 1) “paradox of the plankton.” In a homogeneous environment, interspecific competition should have led to dominance by the competitively superior species. Richerson et al. (1970) have argued that planktonic environments are not homogeneous, but on a small scale are heterogeneous in terms of nutrients, grazers, and other fac- tors, and a single species never has enough of a competitive advantage to dominate the environment. From this perspec- tive, plankton blooms such as brown or red tides are the null case for the paradox of the plankton (Slobodkin 1989); the conditions that normally allow a variety of competing spe- cies to coexist have somehow lapsed, and a single species of algae dominates the environment. In the case of the Texas brown tide alg2.l bloom in the Laguna Madre, conditions allowing for the dominance of this single species have per- sisted for >7 yr. The disruption of the ecological balance before the bloom and the enduring persistence of the bloom demonstrate that hypersaline lagoons with low flushing rates are particularly fragile environments.
Although some of the harmful effects of the brown tide algal bloom have been documented (Buskey and Hyatt 1995; Onuf 1996) and we have proposed here an explanation for the factors contributing to initiation of the bloom, the most important unanswered question remains: why has the bloom persisted for so :ong? The answer may lie in part in the low flushing rates 01’ the Laguna Madre (Shormann 1992), but clearly this is not the complete answer because other aquatic systems with low flushing rates (e.g. lakes) are not generally plagued by persLstent phytoplankton blooms for periods of years. The answer may also lie in the extreme competitive ability of this alga in a harsh hypersaline environment. This alga grows well at salinities of 50%0 and greater, while some of its protozoan grazers show reduced growth under hyper- saline condition; (Buskey unpubl. data). Little is known about the nutrient requirements or kinetics of this alga other than its apparent inability to utilize nitrate (DeYoe and Suttle 1994), but it may grow well under conditions that would limit the growth IIf diatoms, as has been found for the related species A. anophagefierens (Keller and Rice 1989). This alga may also produce allelopathic chemicals suppressing the growth of other algal species. At least part of the explanation for the persistence of this bloom seems to lie in the failure of grazers to bring it under control. This failure may be due in part to the toxic effects and growth inhibition it causes on some species of planktonic grazers (Buskey and Hyatt 1995) and also to increased predation pressure by mesozoo- plankton on microzooplankton and rare larger phytoplankton species, with few alternative food choices in the appropriate size range.
References
ADMIKAAL, W., AI\D L. A. H. VENEKAMP. 1986. Significance of tintinnid grazing during blooms of Phaeocystis youchetii (Hap-

Brown tide bloom initiation 1221
tophyceae) in Dutch coastal waters. Ncth. J. Sea Res. 20: 61- 66.
ANIXIRSON, D. M. 1989. Toxic algal blooms and red tides: A global perspective, p. 1 l-16. Zn Red tides: Biology, environmental science, and toxicology, Proc. 1st Int. Symp. on Red Tides: Elsevier.
ANDI!RSC)N, R. A., G. W. SAUNDERS, M. I? PASKIND, AND J. P, Snx- TON. 1993. Ultrastructure and 18s rRNA gene sequence for Pelagomonas calceolata gen. et sp. nov. and the description of a new algal class, the Pclagophyccae classis nov. J. Phycol29: 701-715.
ARMSTIIONG, N. E. 1987. The ecology of open-bay bottoms of Texas: A community profile. U.S. Fish. Wildl. Serv. Biol. Rep. 85 (7.12).
BURKIIL, I? H., R. E C. MANTOURA, C. A. LI~EWELI,YN, AND N. J. l? OWENS. 1987. Microzooplankton grazing and selectivity of phytoplankton in coastal waters. Mar. Biol. 93: 581-590.
BUSKEY, E. J. 1993. Annual pattern of micro- and mesozooplank- ton abundance and biomass in a subtropical estuary. J. Plank- ton Rcs. 15: 907-924.
- AND C. J. HYATT. 1995. Effects of the Texas (USA) “blown tide” alga on planktonic grazers. Mar. Ecol. Prog. Ser. 126: 285-292.
-, S. STEWART, J. PETERWN, AND C. COLLUMB. 1996. Cur- rent status and historical trends of brown tide and red tide phytoplankton blooms in the Corpus Christi Bay National Es- tuary Program study area. Texas Nat. Resour. Conserv. Comm. Publ. CCBNEP-07.
AND D. A. STOCKWEIL. 1993. Effects of a persistent “blown tide” on zooplankton populations in the Laguna Madre of south Texas, p. 659-666. Zn Toxic phytoplankton blooms in the sea. Proc. 5th Int. Conf. on Toxic Marine Phytoplankton. Elscvier.
CLOERN, J. E. 1982. Does the benthos control phytoplankton bio- mass in south San Francisco Bay? Mar. Ecol. Prog. Ser. 9: 191-202.
COSPBR, E. M., AND OTHERS. 1987. Recurrent and persistent brown tide blooms perturb a coastal marine ecosystem. Estuaries 10: 284-290.
DEYOE, H. R., A. M. CHAN, AND C. A. SUTTL,E. 1995. Phylogeny of Aureococcus anophageflerens and a morphologically similar bloom forming algae from Texas as determined by 18s rRNA sequence analysis. J. Phycol. 31: 413-418.
AND C. A. SUTTLE. 1994. The inability of the Texas “blown tide” alga to use nitrate and the role of nitrogen in the initiation of a persistent bloom of this organism. J. Phycol. 30: 800-806.
DIJNTON, K. H. 1994. Seasonal growth and biomass of the sub- tropical Halodule wrightii in relation to continuous measure- ments of underwater irradiance. Mar. Biol. 120: 479-489.
FI~EMING, R. H. 1939. The control of diatom populations by graz- ing. J. Cons. Cont. Int. Explor. Mer. 14: 210-227.
HAI~LEGIIAEFI;, G. M. 1993. A review of harmful algal blooms and their apparent global increase. Phycologia 32: 79-99.
HANSEN, E C., M. RIXKERMAN, W. C. M. KL,EIN BRETEI,ER, AND R. RIEGMAN. 1993. Phaeocystis blooming enhanced by co- pepod predation on protozoa: Evidence from incubation ex- periments. Mar. Ecol. Prog. Ser. 102: 5 l-57.
HII,L, M. 0. 1973. Diversity and evenness: A unifying notation and its consequences. Ecology 54: 427-432.
HUTCMINSON, G. E. 1961. The paradox of the plankton. Am. Nat. 95: 137-145.
KELJXR, A. A., AND R. L. RICE. 1989. Effects of nutrient enrich- ment on natural populations of the brown tide phytoplankton Aureococcus anophagefferens (Chrysophyseae). J. Phycol. 25: 636-646.
LANDRY, M. R., AND R. P. HASSETT. 1982. Estimating the grazing impact of marinc micro-zooplankton. Mar. Biol. 67: 283-288.
MARTIN, J. H. 1965. Phytoplankton-zooplankton relationships in Narragansett Bay. Limnol. Oceanogr. 10: 185-19 1.
MONTAGNA, P A., AND R. D. KALKB. 1992. The effect of fresh- water inflow on meiofaunal and macrofaunal populations in the Guadalupe and Nueccs estuaries, Texas. Estuaries 15: 266-285.
-, D. A. STOCKWELL, AND R. D. KALKB. 1993. Dwarf surf- clam Mulinea lateralis (Say, 1822) populations and feeding during the Texas brown tide. J. Shellfish Res. 12: 833-842.
OFFICER, C. B., T. J. SMAYDA, AND R. MANN. 1982. Benthic filter feeding: A natural eutrophication control. Mar. Ecol. Prog. Ser. 9: 203-2 10.
ONUF, C. P 1996. Seagrass responses to long-term light reduction by brown tide in upper Laguna Madre, Texas: Distribution and biomass patterns. Mar. Ecol. Prog. Ser. 138: 219-23 1.
QUAMMEN, M. L., AND C. P, ONUF;. 1993. Laguna Madre: Scagrass changes continue decades after salinity reduction. Estuaries 16: 302-3 10.
RAssouL%ArXCJAN, E, M. LAVAL-PEIJT~, AND R. W. SHEI.DON. 1988. Partitioning of the food ration of marine ciliates between pica- and nanoplankton. Hydrobiologia 159: 75-88.
RICYIERSON, I?, R. ARMS’I’RONG, AND C. R. GOLDMAN. 1970. Con- tcmporancous disequilibrium, a new hypothesis to explain the “paradox of the plankton.” Proc. Natl. Acad. Sci. 67: 1710- 1714.
RILEY, G. A. 1946. Factors controlling phytoplankton populations on Georgcs Bank. J. Mar. Res. 6: 54-73.
- 1947. A theoretical analysis of the phytoplankton popu- lation of Georges Bank. J. Mar. Res. 6: 104-l 13.
SULLNER, K. G., AND D. C. BIIOWNLEE. 1990. Dinoflagellate-mi- crozooplankton interactions in Chesapeake Bay, p. 22 l-226. Zn Toxic marine phytoplankton: Proc. 4th Int. Conf. Elsevier.
-, R. V. LACOUTURE, S. J. CIBIK, A. BKINDLEY, AND S. G. BROWNIXE. 1991. Importance of a winter dinoflagellate-mi- croflagellate bloom in the Patuxent River estuary. Estuaries Coastal Shelf Sci. 32: 27-42.
, AND M. M. OIZWN. 1985. Copepod grazing in red tides of Chesapeake Bay, p. 245-250. Zn Toxic dinoflagellates: Proc. 3rd Int. Conf. Elsevier.
SIIANNON, C. E., AND W. WEAVER. 1949. The mathematical theory of communication. Univ. Illinois.
SHORMANN, D. E. 1992. The effects of freshwater inflow and hy- drography on the distribution of brown tide in south Texas. M.A. thesis, Univ. Texas at Austin. 112 p.
SLOBODKIN, L. B. 1989. The null case of the paradox of the plank- ton, p. 449-483. Zn E. M. Cosper et al. [eds.], Novel phyto- plankton blooms: Causes and impacts of recurrent brown tides and other unusual blooms. Springer.
SMAYDA, T. J. 1989. Primary production and the global epidemic of phytoplankton blooms in the sea: A linkage?, p. 449-483. Zn E. M. Cosper et al. [eds.], Novel phytoplankton blooms: Causes and impacts of recurrent brown tides and other unusual blooms. Springer.
-, AND T. A. VII~LAREAL. 1989. The 1985 “brown tide” and the open phytoplankton niche in Narragansett Bay during sum- mcr, p. 159-187. Zn E. M. Cospcr et al. [eds.], Novel phyto- plankton blooms: Causes and impacts of recurrent brown tides and other unusual blooms. Springer.
STOCKWELL, D. A., E. J. BUSKEY, AND T. E. W~IITLEDGE. 1993. Studies on conditions conducive to the development and maintenance of a persistent “brown tide” in Laguna Madre, Texas, p. 693-698. Zn Toxic phytoplankton blooms in the sea. Proc. 5th Int. Conf. on Toxic Marine Phytoplankton. Elsevier.

1222 Buskey et al.
STOECKER, D. K., D. J. GIFKIRD, AND M. Purr. 1994. Preservation of marine planktonic ciliates: Losses and cell shrinkage during fixation. Mar. Ecol. Prog. Ser. 110: 293-299.
TURNER, J. T, AND D. M. ANDERSON. 1983. Zooplankton grazing during dinoflagellate blooms in a Cape Cod embayment, with observations of predation upon tintinnids by copepods. Mar. Ecol. 4: 359-374.
UYE, S. 1986. Impact of copepod grazing on the red-tide flagellate Chattonella antiqua. Mar. Biol. 92: 35-43.
VERITY, F? G., AND T J. SMAYDA. 1989. Nutritional value of Phaeocystis pouchetii (Prymnesiophyceae) and other phy-
toplankton for Acartia spp. (Copepoda): Ingestion, egg pro- duction, and growth of nauplii. Mar. Biol. 100: 161-171.
WATRAS, C. J., V. C. GARCON, R. J. OLSON, S. W. CHISHOLM, AND D. M. ANDERSON. 1985. The effects of zooplankton grazing on estuarine blooms of the toxic dinoflagellate Gonyaulax ta- marensis. J. Plankton Res. 7: 891-908.
WEISSE, T., ANC~ U. SCHEFFEL-MijsER. 1990. Growth and grazing loss rates in single-celled Phaeocystis sp. (Primnesiophyceae). Mar. Biol. 1106: 153-158.
WHITLEDGE, T. EL, S. C. MALLOY, C. J. PATTON, AND C. D. WIRICK. 198 1. Automated nutrient analyses in seawater. Brookhaven Natl. Lab. Formal Rep. BNL 51398.