differential expression of the mango alcohol dehydrogenase gene family during ripening
TRANSCRIPT

Phytochemistry 71 (2010) 1485–1494
Contents lists available at ScienceDirect
Phytochemistry
journal homepage: www.elsevier .com/locate /phytochem
Differential expression of the mango alcohol dehydrogenase gene familyduring ripening
Rajesh K. Singh a, Vidhu A. Sane a,*, Aparna Misra a, Sharique A. Ali b, Pravendra Nath a
a Plant Gene Expression Lab., National Botanical Research Institute, Lucknow 226 001, Indiab Saifia Science College Biotechnology Dept., Bhopal, India
a r t i c l e i n f o a b s t r a c t
Article history:Received 6 March 2010Received in revised form 18 May 2010Available online 2 July 2010
Keywords:Mangifera indicaAlcohol dehydrogenaseRipeningEthyleneABAFruit
0031-9422/$ - see front matter � 2010 Elsevier Ltd. Adoi:10.1016/j.phytochem.2010.05.024
* Corresponding author. Tel.: +91 522 2297959; faxE-mail address: [email protected] (V.A. Sa
Alcohol dehydrogenases play an important role during fruit ripening and aroma production. Three full-length cDNAs (MiAdh1, 2 and 3) encoding alcohol dehydrogenases were obtained from mango fruit pulpusing RT-PCR approaches. All three members displayed strong homology in the coding region when com-pared at the protein and nucleotide levels, however showed variations in untranslated regions. Expres-sion patterns of these ADHs were different during fruit development and ripening. MiADH1 andMiADH2 transcripts accumulated at the onset of ripening in mango fruit whereas MiADH3 accumulatedduring early development of fruit. Expression analysis also indicated that mango ADHs were responsiveto ethylene but regulated differently by ABA. MiADH1 was induced by ABA treatment whereas MiADH2transcript was negatively regulated by ABA. MiADH3 did not respond to ABA in ripening fruit. Differencesin substrate specificity for NADH and NADPH were also observed between the three enzymes. Total ADHenzyme activity correlated positively with increased transcript levels at the initiation of ripening.
� 2010 Elsevier Ltd. All rights reserved.
1. Introduction
Plants produce alcohols and aldehydes by the action of alcoholdehydrogenases (EC 1.1.1.1). Alcohol dehydrogenases in plantshave been encoded by small gene family comprised generallytwo or three members (Thompson et al., 2007), with the exceptionof Arabidopsis (Chang and Meyerowitz, 1986). Most of the alcoholdehydrogenases so far characterized are dimeric enzymes and be-long to the medium-size zinc-containing class comprising of �370residues. ADHs are involved in mediating stress responses inplants, mainly in anaerobiosis and other stresses like dehydration,low temperature, or chemical treatments (Matton et al., 1990;deBruxelles et al., 1996; Dolferus et al., 1994; Peters and Frenkel,2004). ADH genes are also expressed in plant tissues in a develop-mentally-regulated manner, particularly during fruit ripening (Vander Straeten et al., 1991; Chen and Chase, 1993; Speirs et al., 1998,2002; Echeverria et al., 2004) and have been shown to play a majorrole in flavour development in ripe tomato fruit (Speirs et al., 1998)and it is suggested that low activity in the fruit could lead to re-duced/altered flavour (Speirs et al., 2002). Besides tomato, detailedstudies related to role of Adh genes in relation to ripening and ar-oma synthesis are limited only to grapes and melon (Tesniere andVerries, 2000; Zaldivar et al., 2009; Defilippi et al., 2009;Manrıquez et al., 2006). However, in apple (Echevarria et al.,
ll rights reserved.
: +91 522 2205839.ne).
2004) and avocado (Loulakalis et al., 2006) contrary to grape andmelon, ADHs have been characterized at the biochemical level onlyand not at the molecular level.
Mango (Mangifera indica L.) is one of the most popular fruitcrops of tropical and sub-tropical regions. India accounts forapproximately 65% of the total worldwide mango production andboasts of 1000 or more cultivars that differ in fruit size, shape, col-our, aroma, taste and ripening patterns (Salvi and Gunjate, 1988).Mono- and/or sesquiterpene hydrocarbons are the major constitu-ents of Indian mango cultivars but other volatiles like alcohols,aldehydes, esters, furanones and lactones which vary in qualitativeand quantitative terms in different cultivars (Pandit et al., 2009)are also responsible for the distinct flavour and taste of differentmango cultivars. Despite the diversity in flavour and aroma, mangois poorly worked out at molecular level with respect to aroma re-lated genes. In the present study, we isolated and analyzed threemembers of ADH gene family from mango fruit and show that in-spite of the high sequence homology shared by these genes theyare expressed differentially and are regulated by different cues.
2. Results
2.1. Identification, sequencing and phylogenetic analysis of mangoalcohol dehydrogenases
Three mango alcohol dehydrogenases designated as MiADH1,MiADH2, and MiADH3 were cloned using mango fruit pulp cDNAas template. Nucleotide sequence identity amongst these three

1486 R.K. Singh et al. / Phytochemistry 71 (2010) 1485–1494
alcohol dehydrogenases was in the range of 70–90%. The 30UTR ofthe three genes also shared a lot of sequence identity. MiADH1 (147nt) had 70% sequence identity with MiADH2 and MiADH3 for 30UTR(Fig. 1A) while MiADH2 (180 nt) and MiADH3 (129 nt) had a com-mon 30UTR up to a region of 129 nt. However, MiADH2 had anextension of 51 nt in its 30UTR that was absent in MiADH3.
The encoded putative proteins of MiADH1 (382 aa), MiADH2(382 aa) and MiADH3 (379 aa) belonged to medium chain zincADHs. MiADH3 had a nine-base deletion near the 50end, which re-sulted in a three amino acid residue shorter polypeptide (Fig. 2a).Most of the residues in the ORF of these ADHs were identical. Allthree proteins had a predicted molecular mass of 41 kDa and iso-electric points of 6.3 (MiADH1), 6.1 (MiADH2) and 5.86 (MiADH3),based on bioinformatic analysis (EXPASY). All the three MiADHproteins contained features that were essentially conserved withmedium-chain alcohol dehydrogenases. Comparison of these pro-teins with other medium-chain alcohol dehydrogenases revealedthat the positions of cysteines and histidine residues involved inthe binding to the two zinc atoms (C-48, -178, H-70 and C-100,103, 106, 114 for MiADH1, MiADH2 and C-45, -175, H-67 and C-97, -100, -103, -111 for MiADH3) were conserved (Eklund andRamaswamy, 2008; Yokoyama and Harry, 1993). The amino-acidresidues implicated in the specificity towards NAD+ as co-factorin the dehydrogenase reaction viz. T-182, V-207, D-227 were alsofound to be conserved in mango fruit ADHs (Jornvall et al., 1987).
A phylogenetic tree was generated from the alignment of thededuced amino acid sequences of three mango ADHs with 31homologues from other plant species using a short chain ADH(SDR) of Chlamydomonas reinhardtii as outgroup (Fig. 2B). MiADH1,MiADH2 and MiADH3 aligned within the same sub-lineage group.
2.2. Structural organization of MiADH genes
Because of the high sequence conservation in the cDNA of thesethree Adhs, we also studied the genomic sequences. The genomicsequences of the three ADH genes mango varied in length fromthe initiation codon to termination codon (MiAdh1-3549 bp,MiAdh2-3428 bp and MiAdh3-2539 bp, respectively). Both MiAdh1
Fig. 1A. Nucleotide sequence alignment fo
Fig. 1B. Schematic diagrams of the structure of three MiADH genes. Black boxes represecodon and stop codons, respectively.
and 2 were composed of 10 exons and nine introns as reportedfor other plant Adhs. MiAdh3, however, was made up of nine exonsand eight introns (Fig. 1B). The first intron (after 37 nucleotidesfrom the initiation codon in MiADH1 and 2) was absent fromMiADH3 leading to a larger exon of 165 nt in MiADH3. Except forthis, other exons were of identical sizes and conserved in locationin all the three MiADHs. The intron sizes varied generally from 79to 981 bp (Fig. 1B). Intron 1 was the longest in both MiADH1 and2 (981 bp in MiADH1 and 865 bp in MiADH2). Sequence analysessuggested that intron sequences of MiADH2 and 3 shared greatersimilarity with each other than with MiADH1.
2.3. Differential transcript accumulation of mango ADHs duringripening
Transcript accumulation of different ADHs in mango was stud-ied during the course of ethylene induced ripening of harvestedmango for 7 days. A basal level of expression of MiADH transcriptwas detected in case of all the three ADHs on day 0 in ethylene un-treated fruit. The transcript levels of MiADH1 shot up after 24 h inethylene treated fruit, however, from day 2 onwards levels ofMiADH1 transcript decreased from day 1 levels and were thenmaintained throughout the progression of ripening (Fig. 3A). Theexpression of the other two ADH genes was low compared toMiADH1 although the pattern of transcript accumulation ofMiADH2 and 3 was similar to that of MiADH1. Of the three MiADH3transcript showed the lowest steady state levels during the courseof mango ripening. Since these three ADHs showed very high se-quence identity even at nucleotide level, we checked the expres-sion of these transcripts by semi-quantitative PCR at highannealing temperature using gene-specific primers to ensure thatthe transcript patterns were not due to cross hybridization.MiADH3 transcript levels were low and did not give clear resultswhen 20 cycles of PCR were carried out as in case of MiADH1 and2. So for all semi-quantitative experiments related to MiADH3, 35cycles of PCR were carried out. Our results of semi-quantitativePCRs matched with the northern analysis (Fig. 3B) and confirmedthat all the three ADHs are expressed at different levels during
r 50 and 30UTR of three MiADHs cDNA.
nt exons and empty boxes represent introns. ‘ATG’ and ‘TAG’ indicate the initiator

Fig. 2A. Amino acid sequence comparison among MiADH1–3. Identical amino acids are shaded in black, and similar amino acids are shaded in gray. Sequences were alignedusing Clustal W program and presented using BOX shade program.
R.K. Singh et al. / Phytochemistry 71 (2010) 1485–1494 1487
mango softening. The expression was also checked in 1-MCP + eth-ylene treated fruits. All the three MiADHs showed transcript accu-mulation in 1-MCP treated fruits but the levels were far lowercompared to the levels in ethylene treated fruits especially onday 1 post treatment (Fig. 3C and D). Since the patterns were samein northern blots and semi-quantitative PCR experiments, furtherstudies were carried out using semi-quantitative PCR.
The expression patterns of MiADH1, 2 and 3 were also analyzedin developing fruit (developmental stages D1–D4, as described inExperimental), flower and vegetative tissues like leaf and stem. Ba-sal levels of MiADH1 were observed in developing fruit. The levelswere similar to those found in fully mature green mango. The tran-script was absent in leaf and flower but expressed in stem. MiADH2transcript expressed at basal levels in different stages of develop-ing fruit and was also expressed in leaves. Unlike MiADH1 andMiADH2, expression of MiADH3 was high throughout the progres-sion of fruit development and expansion. Moreover, MiADH3 wasalso specific to fruit as no transcript was found in vegetative tissues(Fig. 4A).
Since mango is a climacteric fruit which requires ethylene forinitiation of ripening, we checked whether these three ADHs areregulated by ethylene. Exposure of mature green unripe fruits toshort interval ethylene treatment (0.5, 1.5, 3 and 5 h ethyleneexposure) caused a rapid increase in transcript accumulation ofMiADH1 and MiADH2 within 30 min of ethylene exposure. Ethyleneinduction of MiADH2 expression was also observed in mangoleaves. However, ethylene had no effect on the transcript accumu-lation of MiADH3 even after 5 h exposure (Fig. 4B).
Since ripening is also affected by other hormones like ABA andIAA, we studied the regulation of these genes in fruits by treatmentwith hormones like ABA (Parikh et al., 1990) and IAA in combina-tion with or without ethylene. The real time qRT-PCR results re-vealed that MiADH1 was induced by ABA as in case of ethylene(though fold expression was lower in case of ABA) and co-treat-ment of fruits with ethylene and ABA increased the levels of mRNAover and above those by either of the hormone alone on day 1. Incontrast, ABA treatment strongly repressed MiADH2 and even basalexpression was lost after 1 day of ABA treatment. However, co-treatment with ethylene was able to overcome the inhibitory effectof ABA although transcript levels did not reach the same levels asin only ethylene treated fruits (Fig. 5). MiADH3 seemed to be insen-sitive to treatment with ABA/ethylene and continued to be ex-pressed at a basal level. None of the three ADHs weresignificantly affected by auxin (IAA), however, ethylene-IAA co-treatment resulted in higher expression as compared to treatmentwith auxin alone (Fig. 5).
2.4. ADH activity in fruit
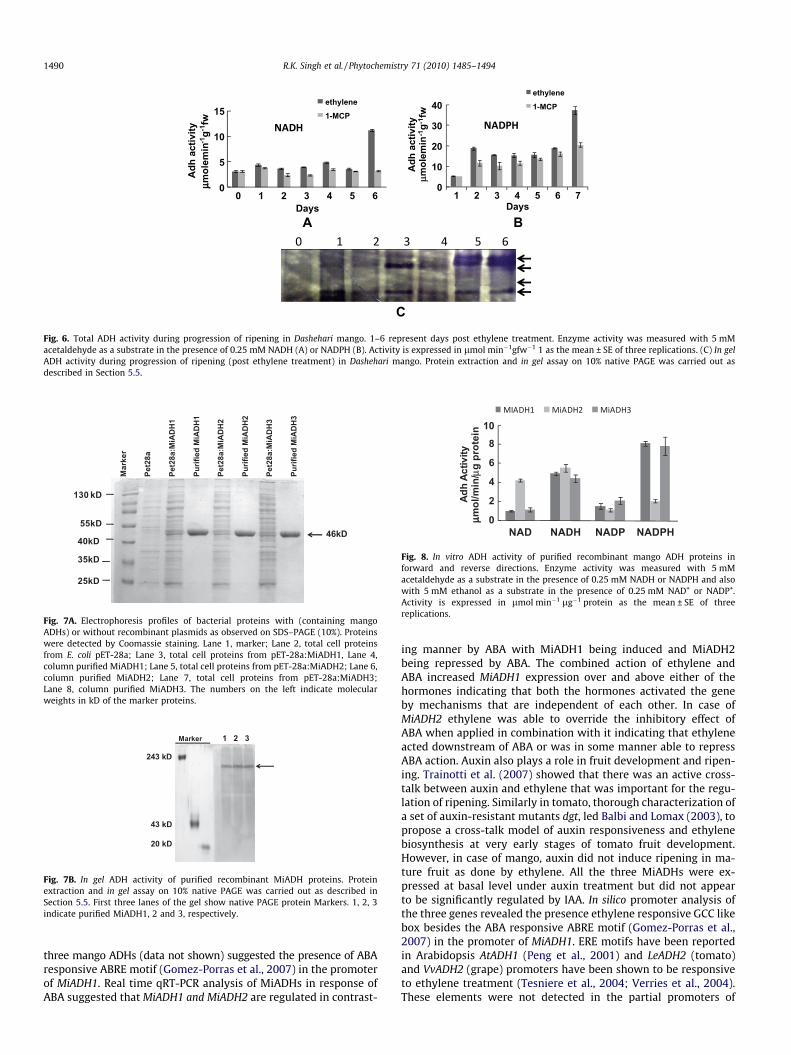
Considering the ripening related increase in ADH transcripts,we measured ADH activity in Dashehari fruit extracts during thecourse of ripening with acetaldehyde as a substrate in the presenceof NADH or NADPH. The fruit activity was higher in the presence ofNADPH than NADH (Fig. 6). The NADPH dependent activity in-creased almost fourfold on day 1 post ethylene treatment and con-tinued to be maintained at those levels until day 5. Thereafter on

Fig. 2B. Phylogenetic analysis of mango ADHs amino acid sequences with otherAdhs sequences belonging to the medium-chain zinc-binding type. Full-lengthprotein sequences were aligned using CLUSTAL-W and a phylogenetic tree wasconstructed using PHYLIP with the PROTPARS program. Numbers above thebranches indicate bootstrap values (1000 replicates). Chlamydomonas reinhardtiiCrSdr was used as out group. Sequences are: Arabidopsis thaliana At-ADH1(NP_177837.1); Brassica oleracea BoAdh (AAF23532.1); (EDO98712.1); Citrus xparadisi CpAdh (AAY86033.1); Cucumis melo CmAdh1 (ABC02081.1) and CmAdh2(ABC02082.1); Fragaria x ananassa FaAdh (CAA33613.1) and FaSdh (BAF75466.1);Lotus japonicus LjAdh (CAG30579.1); Malus domestica MdAdh (CAA88271.1);Medicago truncatula MtAdh (ACJ84419); Nicotiana tabacum NtAdh (CAA57446.1);Petunia x hybrida PhAdh2 (AAO74898.1); Pisum sativum PsAdh (P12886.1); Populustrichocarpa PtAdh (XP_002328464.1); Ricinus communis RcAdh (EEF36243.1);Solanum lycopersicum SlAdh2 (CAA54450.1) and SlAdh1 (AAB33481.2); Solanumtuberosum StAdh (CAA37333.1); Vitis vinifera VvAdh1 (AAG01381.1), VvAdh2(AAG01382.1), VvAdh3 (AAG01383.1) and VvAdh6 (AAF44335.1).
1488 R.K. Singh et al. / Phytochemistry 71 (2010) 1485–1494
day 6 it increased another two folds in ethylene treated fruit. Anincrease in activity in 1-MCP treated fruit was also observed onday 1. However, this increase was much less than ethylene treatedfruits rising only up to 60% of activity in ethylene treated fruit.Thereafter there was a gradual increase in activity up to day 6(Fig. 6B). In contrast to the NADPH dependant activity, the NADHdependant activity did not show any major change until day 5.On day 6, however, there was a sudden fourfold increase in activityonly in ethylene treated fruit (Fig. 6A). We also checked the ADHisozymes during progression of ripening on native PAGE gel byactivity staining. Two major and one minor (between slowestand fastest bands) isoforms appeared from day 1 onwards. Late
in ripening, on day 5 and day 6 a new slow migrating form ap-peared (Fig. 6C). The activity of all these forms increased withthe progression of ripening. Since ADHs can exist in dimeric homoand heteromeric forms, it is difficult to speculate which forms arebeing expressed during mango ripening. Since in gel assays did notgive much information on three MiADHs, we expressed recombi-nant pET-28a:MiADH1, pET-28a:MiADH2 and pET-28a:MiADH3in a heterologous system [Escherichia coli BL21 (DE3)]. Recombi-nant proteins expressed after IPTG induction at 16 �C in the solublefraction were analyzed on SDS–PAGE. For all the three pET-28a:MiADH constructs the presence of an additional band in solu-ble extracts was revealed by Coomassie staining when compared tothe non-recombinant control pET-28a. Purification of the recombi-nant proteins by Ni–NTA columns resulted in single band ofapproximately 46 kD (ADH and His tag; Fig. 7A). To see if thesethree purified recombinant MiADHs were active, in gel andin vitro activity assays were performed. In gel assays using NADP+
and ethanol resulted in single activity band of approximately180 kD in all the three cases (Fig. 7B). In vitro assays carried outwith three purified ADH proteins for acetaldehyde and ethanol.MiADH1 and MiADH3 preferred NADPH (acetaldehyde) whereasMiADH2 showed greater affinity towards NADH as substrate(Fig. 8). Our results suggested that though MiADH enzymes func-tion both in forward and reverse directions they prefer reductionof acetaldehyde to ethanol under in vitro conditions (Fig. 8).
3. Discussion
At the onset of ripening the vast array of volatile compoundsproduced by fruits are responsible for their flavour and aromacharacteristics (Dudareva et al., 2004). Although the pathways foraroma biosynthesis have not yet characterized in detail, alcoholdehydrogenases have been implicated in formation of aroma re-lated alcohols (Speirs et al., 1998). The presence of more than1000 varieties of mango that differ in aroma makes mango aninteresting system for study of aroma and led us to characterizeADHs in mango for their role in ripening. Our results suggest theexistence of at least three different ADHs in mango fruit that sharehigh homology at nucleotide and amino acid level in the coding re-gion. Despite this, the promoter regions of these genes were diver-gent suggesting that the three ADH genes might be underregulation of different signals. The genes are clubbed with mediumchain dehydrogenases from dicot plants. As in mango, the exis-tence of multiple ADHs had also been reported from other fruitplants. These include two ADH genes from melon, three in tomatoand four in grape (one of which is probably a pseudogene). Of thethree tomato genes, LeADH2 was expressed in ripening fruit (Long-hurst et al., 1994) while the rest were expressed in anthers (Inger-soll et al., 1994). Two melon ADH genes (Cm-ADH1, Cm-ADH2)expressed during fruit ripening although these belonged to differ-ent ADH-subfamilies (Manrıquez et al., 2006). Of the three func-tional grape ADHs, VvADH1 and VvADH3 were prominentlyexpressed during fruit development whereas VvADH2 expressedmostly during ripening. In mango, all three ADHs were expressedin fruit during ethylene induced ripening although their relativetranscript levels varied a lot. The expression of all three mangoADHs showed a rapid but transient increase on day 1 post ethylenetreatment followed by a decrease in subsequent days particularlyin MiADH2 and MiADH3. These results are quite similar to thoseof melon where both the genes showed a transient but sharp in-crease on 39 DAP when ethylene production was at maximumand declined thereafter. Although MiADH1 also showed a decreasein subsequent stages, it continued to remain at much higher levelsas compared to MiADH2 and MiADH3. The expression was ethyleneregulated and ripening related as evident from the decrease in
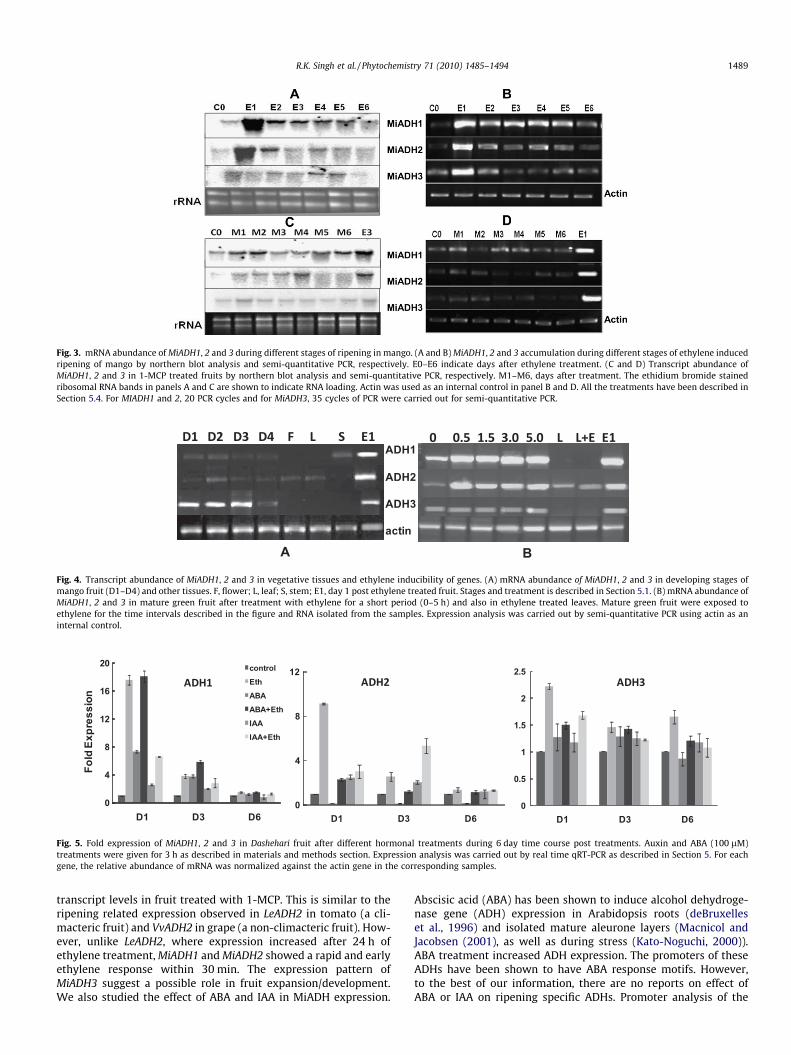
Fig. 3. mRNA abundance of MiADH1, 2 and 3 during different stages of ripening in mango. (A and B) MiADH1, 2 and 3 accumulation during different stages of ethylene inducedripening of mango by northern blot analysis and semi-quantitative PCR, respectively. E0–E6 indicate days after ethylene treatment. (C and D) Transcript abundance ofMiADH1, 2 and 3 in 1-MCP treated fruits by northern blot analysis and semi-quantitative PCR, respectively. M1–M6, days after treatment. The ethidium bromide stainedribosomal RNA bands in panels A and C are shown to indicate RNA loading. Actin was used as an internal control in panel B and D. All the treatments have been described inSection 5.4. For MIADH1 and 2, 20 PCR cycles and for MiADH3, 35 cycles of PCR were carried out for semi-quantitative PCR.
ADH1
ADH2
ADH3
actin
A B
Fig. 4. Transcript abundance of MiADH1, 2 and 3 in vegetative tissues and ethylene inducibility of genes. (A) mRNA abundance of MiADH1, 2 and 3 in developing stages ofmango fruit (D1–D4) and other tissues. F, flower; L, leaf; S, stem; E1, day 1 post ethylene treated fruit. Stages and treatment is described in Section 5.1. (B) mRNA abundance ofMiADH1, 2 and 3 in mature green fruit after treatment with ethylene for a short period (0–5 h) and also in ethylene treated leaves. Mature green fruit were exposed toethylene for the time intervals described in the figure and RNA isolated from the samples. Expression analysis was carried out by semi-quantitative PCR using actin as aninternal control.
0
0.5
1
1.5
2
2.5
D1 D3 D6
Fold
Exp
ress
ion
0
4
8
12
D1 D3 D60
4
8
12
16
20
D1 D3 D6
controlEthABAABA+EthIAAIAA+Eth
Fig. 5. Fold expression of MiADH1, 2 and 3 in Dashehari fruit after different hormonal treatments during 6 day time course post treatments. Auxin and ABA (100 lM)treatments were given for 3 h as described in materials and methods section. Expression analysis was carried out by real time qRT-PCR as described in Section 5. For eachgene, the relative abundance of mRNA was normalized against the actin gene in the corresponding samples.
R.K. Singh et al. / Phytochemistry 71 (2010) 1485–1494 1489
transcript levels in fruit treated with 1-MCP. This is similar to theripening related expression observed in LeADH2 in tomato (a cli-macteric fruit) and VvADH2 in grape (a non-climacteric fruit). How-ever, unlike LeADH2, where expression increased after 24 h ofethylene treatment, MiADH1 and MiADH2 showed a rapid and earlyethylene response within 30 min. The expression pattern ofMiADH3 suggest a possible role in fruit expansion/development.We also studied the effect of ABA and IAA in MiADH expression.
Abscisic acid (ABA) has been shown to induce alcohol dehydroge-nase gene (ADH) expression in Arabidopsis roots (deBruxelleset al., 1996) and isolated mature aleurone layers (Macnicol andJacobsen (2001), as well as during stress (Kato-Noguchi, 2000)).ABA treatment increased ADH expression. The promoters of theseADHs have been shown to have ABA response motifs. However,to the best of our information, there are no reports on effect ofABA or IAA on ripening specific ADHs. Promoter analysis of the
0
5
10
15
Adh
act
ivity
µµmol
emin
-1g-1
fw0 1 2 3 4 5 6
Days
ethylene1-MCP
0
10
20
30
40
Adh
act
ivity
µmol
emin
-1g-1
fw
1 2 3 4 5 6 7Days
ethylene
1-MCP
A B
C
Fig. 6. Total ADH activity during progression of ripening in Dashehari mango. 1–6 represent days post ethylene treatment. Enzyme activity was measured with 5 mMacetaldehyde as a substrate in the presence of 0.25 mM NADH (A) or NADPH (B). Activity is expressed in lmol min�1gfw�1 1 as the mean ± SE of three replications. (C) In gelADH activity during progression of ripening (post ethylene treatment) in Dashehari mango. Protein extraction and in gel assay on 10% native PAGE was carried out asdescribed in Section 5.5.
Mar
ker
Pet2
8a
Pet2
8a:M
iAD
H1
Purif
ied
MiA
DH
1
Pet2
8a:M
iAD
H2
Purif
ied
MiA
DH
2
Pet2
8a:M
iAD
H3
Purif
ied
MiA
DH
3
46kD
130 kD
55kD
40kD
35kD
25kD
Fig. 7A. Electrophoresis profiles of bacterial proteins with (containing mangoADHs) or without recombinant plasmids as observed on SDS–PAGE (10%). Proteinswere detected by Coomassie staining. Lane 1, marker; Lane 2, total cell proteinsfrom E. coli pET-28a; Lane 3, total cell proteins from pET-28a:MiADH1, Lane 4,column purified MiADH1; Lane 5, total cell proteins from pET-28a:MiADH2; Lane 6,column purified MiADH2; Lane 7, total cell proteins from pET-28a:MiADH3;Lane 8, column purified MiADH3. The numbers on the left indicate molecularweights in kD of the marker proteins.
Marker 1 2 3
243 kD
43 kD
20 kD
Fig. 7B. In gel ADH activity of purified recombinant MiADH proteins. Proteinextraction and in gel assay on 10% native PAGE was carried out as described inSection 5.5. First three lanes of the gel show native PAGE protein Markers. 1, 2, 3indicate purified MiADH1, 2 and 3, respectively.
0
2
4
6
810
NAD NADH NADP NADPH
Adh
Activ
ityµ µm
ol/m
in/µ
g pr
otei
n
Fig. 8. In vitro ADH activity of purified recombinant mango ADH proteins inforward and reverse directions. Enzyme activity was measured with 5 mMacetaldehyde as a substrate in the presence of 0.25 mM NADH or NADPH and alsowith 5 mM ethanol as a substrate in the presence of 0.25 mM NAD+ or NADP+.Activity is expressed in lmol min�1 lg�1 protein as the mean ± SE of threereplications.
1490 R.K. Singh et al. / Phytochemistry 71 (2010) 1485–1494
three mango ADHs (data not shown) suggested the presence of ABAresponsive ABRE motif (Gomez-Porras et al., 2007) in the promoterof MiADH1. Real time qRT-PCR analysis of MiADHs in response ofABA suggested that MiADH1 and MiADH2 are regulated in contrast-
ing manner by ABA with MiADH1 being induced and MiADH2being repressed by ABA. The combined action of ethylene andABA increased MiADH1 expression over and above either of thehormones indicating that both the hormones activated the geneby mechanisms that are independent of each other. In case ofMiADH2 ethylene was able to override the inhibitory effect ofABA when applied in combination with it indicating that ethyleneacted downstream of ABA or was in some manner able to repressABA action. Auxin also plays a role in fruit development and ripen-ing. Trainotti et al. (2007) showed that there was an active cross-talk between auxin and ethylene that was important for the regu-lation of ripening. Similarly in tomato, thorough characterization ofa set of auxin-resistant mutants dgt, led Balbi and Lomax (2003), topropose a cross-talk model of auxin responsiveness and ethylenebiosynthesis at very early stages of tomato fruit development.However, in case of mango, auxin did not induce ripening in ma-ture fruit as done by ethylene. All the three MiADHs were ex-pressed at basal level under auxin treatment but did not appearto be significantly regulated by IAA. In silico promoter analysis ofthe three genes revealed the presence ethylene responsive GCC likebox besides the ABA responsive ABRE motif (Gomez-Porras et al.,2007) in the promoter of MiADH1. ERE motifs have been reportedin Arabidopsis AtADH1 (Peng et al., 2001) and LeADH2 (tomato)and VvADH2 (grape) promoters have been shown to be responsiveto ethylene treatment (Tesniere et al., 2004; Verries et al., 2004).These elements were not detected in the partial promoters of

R.K. Singh et al. / Phytochemistry 71 (2010) 1485–1494 1491
MiADH2 and MiADH3. A complex interplay of hormones is knownto affect fruit development and ripening with auxin and GA beingimportant during fruit expansion and ABA and ethylene for ripen-ing (Srivastava and Handa, 2005). In Dashehari mango, additionalcomplexities are seen due to differential ripening patterns withinthe fruit. Ripening is initiated from the stone and moves graduallytowards the peel and is earlier near the pedicel and later at thebottom. The differential spatial regulation of ripening by hormonesmay selectively affect the expression of one or the other ADHs. Italso needs to be work out whether the contrasting effects of ABAon MiADH1 and MiADH2 help in maintaining the net levels ofADH in fruit during ripening. These results indicate a complexhormonal regulation of the genes during fruit development andripening.
Interestingly, when ADH activity was analyzed in vitro and by ingel assay in mango fruit extracts, it revealed an increase duringprogression of ripening and softening. The mango fruit activitywas higher in the presence of NADPH than NADH as reported formelon and olive ADHs (Manrıquez et al., 2006; Salas and Sanchez,1998). The sudden increase in ADH activity in presence of NADPHon day 1 matches the transcript patterns of MiADH1 and MiADH2.However, the second peak in activity on day 6 (which is observedin NADPH as well as NADH) does not correlate with the transcriptpatterns of any of the three mango ADHs indicating that otherADH(s) that utilize either NADPH or NADH or both may be acti-vated at late ripening stages. Studies on recombinant proteins alsoindicate that MiADH1 might be active during initial ripening as itshowed preference for NADPH. Previous studies have shown thataroma associated ADHs are mainly NADPH dependent while thoseassociated with stress preferably use NADH. However, presence oftwo NADPH- and NADH-dependent ADHs in strawberry receptaclewith broad substrate specificities has been shown by Mitchell andJelenkovic (1995). The group observed that NADH-dependent ADHshowed higher activity against branched alcohols and nonaromaticaldehydes and an increase in activity during ripening, which wascorrelated with characteristic aroma of strawberry. The selectiveincrease in NADPH dependent ADH activity (which is correlatedwith MiADH1 and 2 transcript patterns) indicates that these ADHsmight be related to aroma development in fruit. Since fruit pulpdeterioration begins on day 6 only in ethylene treated fruit butnot as much in control or 1-MCP fruit, the changes associated withthis deterioration may be perceived as stress and may activatestress related ADHs (as evident from a sudden increase in NADH-dependent ADH). The deterioration of the fruit is also associatedwith changes in flavour and aroma that might explain the increasein NADPH activity. Speirs and co workers tested the relationship ofADH activity and fruit ripeness/softness in six different cultivars oftomato and showed that ADH activity was directly correlated withthe rate of softening and suggested that increased ADH activityduring softening might be a function of decreasing internal oxygenconcentration (Speirs et al., 2002). In gel activity shows the appear-ance of two forms of ADH on day 1 that may possibly representMiADH1 and 2 or a combination of both. The levels of these formsincrease dramatically on day 6 along with the appearance of anadditional band. This may represent the NADH-dependent ADH.
4. Conclusion
In this paper, we demonstrated that three mango ADH isogenes,although exhibiting high sequence homology, are expressed differ-entially during fruit development and ripening. The three isoformsrespond differently to other hormones like ABA and IAA in additionto ripening related hormone ethylene suggesting distinct meta-bolic roles for these genes. Differential ADH expression at tran-script and activity levels suggests tight adaptation of the fruit tothe developmental events. Further studies related to promoters
of these MiADHs will lead to understanding of regulation of thesegenes at different time points and under different stress conditions.
5. Experimental
5.1. Material and treatments
All the experiments were carried out with Dashehari mango (M.indica). Mature unripe Dashehari mangoes (12–15 cm long with ahard stone) were harvested from a local mango orchard at Luc-know. Ripening was initiated by exposing the fruits to exogenousethylene (100 ll/l) for 24 h in a closed 10 L chamber and then al-lowed to ripen for 6 days at 23 �C in air only (Sane et al., 2005).Pulp tissue was harvested every 24 h, frozen in liquid nitrogenand stored at �70 �C until further use. For short duration ethylenetreatment, mature unripe mangoes were exposed to ethylene(100 ll/l) and pulp tissue was collected at time intervals of 30,60, 90 min, 3 h and 5 h. Fruits that were allowed to undergopost-harvest ripening in air for 6 days without any exogenous eth-ylene treatment were treated as control and pulp tissue harvestedas mentioned earlier (Sane et al., 2005; Chourasia et al., 2006). For1-methylcyclopropene (1-MCP) (an ethylene perception inhibitor)treatment, mature unripe fruits were exposed to 100 ll/l 1-MCPfor 12 h followed immediately by 100 ll/l ethylene treatment for24 h and then kept at 23 �C in air as in earlier cases. Four stagesof developing fruit D1 (1–1.5 cm), D2 (3 cm), D3 (6 cm) and D4(10 cm) were also included in the study along with leaf, flowerand stem tissues for expression analysis. For ABA and IAA treat-ment, fruits were dipped in a solution containing 100 lM ABA orIAA in 0.2% teepol (detergent) and vacuum infiltrated for 2 h. Con-trol fruits were infiltrated with 0.2% teepol.
5.2. RNA extraction, reverse transcription-PCR and sequencing
RNA was extracted from various tissues using cetryl trimethylammonium bromide as described by Chourasia et al. (2006,2008). RNA from day 4 ethylene-treated mango pulp (treated withRNase free DNase) was used for cDNA synthesis. Reverse transcrip-tion was carried out using 30 RACE with the adapter primer (30AP:50GGCCACGCGTC GACTAGTACTTTTTTTTTTTTTTTTT-30) and MuM-LV reverse transcriptase (MBI, Fermentas) according to manufac-turer’s instruction. To amplify the Adh gene fragment frommango, degenerate primers AdhF1 (50-GAR GCW GSA GGB ATWGTG GAR AGT GT-30) and AdhF2 (50-GGV ACT TCC ACA CCT TCAGYG ART ACA-30) and AdhR1 (50-CTY CKR TCV ACH CCD CCA TYNGTC AT-30) AdhR2 (50 GAY YGT KGC WCC TTC HGC ARC MGCAAG-30) were designed. PCR-amplified fragments were cloned inpTZ57R/T vector using the InsT/A cloning Kit (MBI, Fermentas).Sequencing was outsourced to MWG DNA sequencing services,Germany. Gene-specific primers were designed for 30 RACE (AdhF3and AdhF4) based on the sequence of partial clone and these incombination with 30AP primer were used to obtain the 30end. Toget the upstream sequence of MiAdh1, SMART RACE cDNA amplifi-cation kit (Clontech Laboratories, Inc., Palo Alto, CA) was used.cDNA was prepared by reverse transcription using the 30AP alongwith the (SMART) primer (50AAGCAGTGGTAACAACGCAGAGTACGCGGG-30). Fragments were amplified using this cDNA and two gene-specific primers AdhR3 (50-CAGTGAACACAGGAAGAACATGGTCTC-30)and AdhR4 (50-AGATCAGTTACACCTTCCCCTACACTCC-30). Amplified50 RACE product was cloned and sequenced. These partial frag-ments were then joined to get a composite sequence. Two specificprimers (F0 and R0) were synthesized out of the composite se-quence to amplify the full-length cDNA fragment of 1155 bp frag-ment designated as MiAdh1 (Ac. No. GU233766). MiAdh2 wasobtained using AdhF4 and 30AP primer. This fragment differed in

1492 R.K. Singh et al. / Phytochemistry 71 (2010) 1485–1494
sequence from the MiAdh1 sequence and specific primers (Adh2R2and Adh2R3R) were designed for extension towards 50end. A genespecific primer MiAdh2F0 was then designed based on the se-quence of this Partial fragment and along with AdhR0 resulted infull length MiAdh2 ORF (Ac. No. GU233767). In the process ofobtaining genomic sequence of MiAdh1 by genome walking incombination with genome walker adapter primers (AP1 andAP2), we obtained MiAdh3 partial sequence. For this a genomewalker library was constructed using Universal genome walkerkit (Clontech) according to manufacturer’s protocol. Based on thegenomic sequence, gene specific forward Adh3Fo primer (contain-ing first ATG codon) was designed and used in combination with30AP to get the cDNA fragment containing the entire ORF and 30UTRfor MiAdh3 (Ac. No. GU233768).
The genome walker library was also used to isolate promoter ofMiAdh1, 2 and 3 (GU233769, GU233770, GU233771, respectively)using gene-specific primers and genome walker adapter primerAP1 and AP2. Sequences of all the primers used for amplificationof different ADH genes are given in Table 1.
5.3. Sequence analysis
Homology searches and signal peptide predictions were per-formed using the Basic Local Alignment Search Tool (BLAST, NCBIBethesda, MD) and SIGNALP version 1.1 (Center for Biological Se-quence Analysis, Technical University of Denmark) respectively.Multiple alignments were carried out using CLUSTALW program.Phylogenetic analyses of putative mango ADH proteins were car-ried out using Phylip 3.5c package employing parsimony and boot-strap analysis (100 replicates). Promoter analysis was carried outby PLANT CARE program.
5.4. Northern analysis and semi-quantitative PCR and real time PCR
Total RNA was isolated from mango pulp from control (ethyleneuntreated), different days post ethylene treated fruit and 1-MCPtreated fruit. RNA from fruit pulp from different developmentalstages and vegetative tissues were also isolated. RNA gel blot anal-yses were carried out as previously described (Sane et al., 2005).Total RNA (40 lg) from each sample was resolved on a 1.2% form-aldehyde agarose gel as described by Sambrook et al. (1989) andmodified in the Qiagen Oligotex handbook (2002) protocol forRNA electrophoresis. RNA was transferred to Hybond-N nylonmembrane (Amersham-Pharmacia, Biotech, UK). Blots were pre-hybridized at 42 �C for 6–8 h in hybridization buffer (50% formam-
Table 1List of primers.
ADHF1 50GARGCWGSAGGBATWGTGGARAGTGT 30
30ADHF2 50GGVACTTCCACACCTTCAGYGARTACA 30
ADHR1 50CTYCKRTCVACHCCDCCATYNGTCAT 30
ADHR2 50GAYYGTKGCWCCTTCHGCARCMGCAAG 30
ADHF3 50TGAAGCTAAGAAGTTTGGCTGCACAGA 30
ADHR3 50CAGTGAACACAGGAAGAACATGGTCTC 30
ADHF4 50GACAAACCTGTTCAACAGGTTATTGCT 30
ADHR4 50AGATCAGTTACACCTTCCCCTACACTCC 30
ADHF0bam 50GGATCCATGTCTAACAAAGTCGGTCAA 30
ADHR0hin 50AAGCTTCTAATCCTCCATGCGAATGATG 30
ADH2R1 50GAATGCATCATCTTTGTTTGGAACACC 30
ADH2R2 50AACTGCAACACCCCAACCATCATGGAC 30
ADH2F0 50TCTTGAGGATCCATGTCTAACACAGCT 30
ADH3F0 50GCTGTGGGATCCATGTCTCATTGGCTG 30
ADH250F(�1) 50TCTTGAAACAAAATGTCTAACAC 30
ADH230R10 50CAGAAGCAAAGTAGACCGGAAA 30
ADH230R2 50AGAAACAAAAGCACACAAAACA 30
ADH3Fo modified 50TGGGATCCATGTCTCATTGGCTGTATT 30
ADH3F(�1) 50TCTTCCTCATATATTGGGATTTGC 30
ide, 6X SSC, 5X Denhardt’s solution, 0.1% SDS and 100 lg/mL dena-tured calf thymus DNA). An 1149 bp long MiAdh1 cDNA fragmentcontaining complete ORF was labeled with a 32P-dCTP using ran-dom priming method and used as a probe for northern blots (Sam-brook et al., 1989). Other probes were prepared using the part ofORF and 30UTR of each gene. This included a region of 1120 bpfor MiAdh2 and 1269 bp for MiAdh3. Hybridization was carriedout overnight at 42 �C in hybridization buffer containing the radi-olabeled probe. The autoradiograms were developed after exposingthe blots to X-ray films (Fuji) at �70 �C for 3–4 days. Hybridizationexperiments for each gene were replicated at least twice with dif-ferent RNA samples.
Since the nucleotide sequences within the ORF of these threeADHs were highly homologous, transcript analysis was also carriedout by semi-quantitative PCR. For this, semi-quantitative PCR wascarried out using cDNA prepared from 500 ng DNA free RNA fromdifferent fruit and vegetative tissues. PCR was performed using thegene-specific primers for each gene and actin as an internal con-trol. Twenty cycles were carried out for MiADH1 and 2 and 35 cy-cles for MiADH3. Care was taken in designing primers so that theamplification was gene specific (data not shown).
Real-time qRT-PCR was performed in a 20 ll reaction volumeusing SYBR GREEN PCR Master Mix (PE-Applied Biosystems, FosterCity, CA, USA) on an ABI PRISM 7000 sequence-detection systemaccording to manufactures instructions. RT-PCR conditions wereas follows: 50 �C for 2 min, followed by 95 �C for 10 min, then 40cycles of 95 �C for 15 s and 60 �C for 1 min. All RT-PCR experimentswere run in triplicate. For each sample, a Ct (threshold sample) va-lue was calculated from the amplification curves by selecting theoptimal DRn (emission of reporter dye over starting backgroundfluorescence) in the exponential portion of the amplification plot.Relative fold differences were calculated based on the comparativeCt method using the actin as an internal standard. To determinerelative fold differences for each sample in each experiment, theCt values for all MiADH genes were normalized to the Ct valuefor actin and was calculated relative to a calibrator ([Control day0 unripe fruit (C)] for all the ADHs) using the formula 2�DDCt.
5.5. Protein extraction and enzyme activity
One gram of frozen pulp tissue from different stages of ripeningwas used for protein extraction. For each stage, tissue was pooledfrom three individual fruits. Three independent experiments werecarried out with pooled samples. Protein was extracted as de-scribed by Kanellis et al. (1991), with slight modifications. Tissueswere ground to a fine powder in a pestle and mortar with liquidnitrogen and homogenized in 1 ml extraction buffer (100 mMTris–HCl, pH 8.0, 500 mM NaCl, 5 mM EDTA, 5 mM sodium dithio-nite, 2 mM PMSF and 10% glycerol). The homogenate was centri-fuged at 10,000g for 20 min at 4 �C in a Sorvall RC5C centrifuge(Sorvall Instruments Co., Inc., Newton, CT). The supernatant thusobtained was desalted through a Sephadex G-50 column(Amersham-Pharmacia, Sweden) and the desalted sample wasused for enzyme assays. In vitro Adh activity was determined asdescribed by Manrıquez et al. (2006) in a substrate solutioncontaining 5 mM acetaldehyde, 50 mM sodium phosphate buffer,pH 5.8, and 0.25 mM NADH or NADPH. For the assay, 25 ll of en-zyme extract (25 lg protein) was added to 1 ml of substrate solu-tion and incubated for 90 min at 37 �C. The change in absorbancewas measured at 340 nm on a spectrophotometer (PharmaciaUltrospec 3000; Amersham Biosciences, Piscataway, NJ). Spectro-photometric readings at 0 min and after 90 min of incubation weretaken for each sample. Enzyme blank and substrate blank in thereaction mixture were used as negative controls. The results wereexpressed in enzyme units per gram of fresh weight and one unit ofthe enzyme activity was defined as the amount of enzyme that

R.K. Singh et al. / Phytochemistry 71 (2010) 1485–1494 1493
converts 1 lmol of NADH/NADPH to NAD+/NADP+ per minute un-der the assay conditions described. Three independent experi-ments were carried out for each set. For in gel experiments,desalted protein samples were separated on 10% native-PAGE inTris–glycine buffer pH 9.0 (Laemmli, 1970) at 40 mA for 90 minat 4 �C. After electrophoresis, gel was stained for ADH activityusing stain containing 1 M Tris–HCl, pH 9.0, 1.5 mM NAD+ orNADP+, 0.25 mM NBT or MTT, 0.025 mM PMSF and 5% (v/v) ethanol(Duncan et al., 1996). Staining was carried out at 37 �C for 30–45 min till the appearance of the bands. Protein was quantifiedas described by Peterson (1977), with BSA as a standard.
5.6. Hetrologous expression of MiADHs in bacteria
The mango ADH cDNAs comprising of start codon and stop co-don were amplified by specific forward and reverse primers havingBamHI site. Amplified products were cloned first in pTZ57R/T vec-tor and sequenced. Complete ORF fragments for all the three ADHswere obtained by BamHI digestion, and ligated to pET28a (Nova-gen) vector to give, respectively, pET28a-MiADH1, pET28a-MiADH2 and pET28a-MiADH3. E. coli BL21(DE3) cells were usedfor isopropyl-bthiogalactopyranoside (IPTG)-induced expressionof these constructs. Recombinant bacteria and E. coli BL21 (DE3)cells transformed with pET-28a (as a control), were grown at37 �C in LB medium containing 50 lg/ml ampicillin and 30 lg/mlChloramphenicol and induced with 0.4 mM IPTG. After an induc-tion period of 3 h at 16 �C, the cells were harvested, washed andsuspended in buffer A [20 mM Tris–HCl extraction buffer, pH 7.2,0.5 mM DTT, 1 mM PMSF and 20% (v:v) glycerol]. Repetitive freezeand thaw method was applied for protein extraction. Extracts werecleared by centrifugation (12,000g, 15 min at 4 �C). The over ex-pressed proteins were purified by using Ni–NTA column (BangloreGenei, India) as per the manufacturer’s guide lines. Recombinantproteins were eluted at a concentration of 200 mM imidazole.5 lg of total soluble protein was electrophoresed on a 12%, SDS–PAGE (Laemmli, 1970). Protein (5 lg) was mixed with 2� SDS–PAGE sample loading buffer [glycerol 20% (v/v), Tris–Cl (pH 6.8)0.1 M, SDS 4% (w/v), DTT 0.2 M, bromophenol blue 0.2% (w/v)]and denatured at 95 �C prior to loading. The electrophoresis wascarried out in Mini Protean III Dual slab cell system (Bio-Rad) atconstant current of 16 mA/gel. Gels were stained with coomassieblue R-250.
In gel assays and in vitro assays were also carried out with puri-fied proteins as described in Section 5.5.
Acknowledgements
We are thankful to Central Institute for Subtropical Horticulture,Lucknow, for the mango samples. Senior Research Fellowship pro-vided to Rajesh K. Singh by CSIR, India is gratefully acknowledged.
References
Balbi, V., Lomax, T.L., 2003. Regulation of early tomato fruit development by thediageotropica gene. Plant Physiol. 131, 186–197.
Chang, C., Meyerowitz, E.M., 1986. Molecular cloning and DNA sequence of theArabidopsis thaliana alcohol dehydrogenase gene. Proc. Natl. Acad. Sci. USA 83,1408–1412.
Chen, A.R.S., Chase Jr., T., 1993. Alcohol dehydrogenase 2 and pyruvatedecarboxylase induction in ripening and hypoxic tomato fruit. Plant Physiol.Biochem. 31, 875–885.
Chourasia, A., Sane, V.A., Nath, P., 2006. Differential expression of pectate lyaseduring ethylene induced post harvest softening of mango (Mangifera indica var.Dashehari). Physiol. Plantarum 128, 546–555.
Chourasia, A., Sane, V.A., Singh, R.K., Nath, P., 2008. Isolation and characterization ofthe MiCel1 gene from mango: ripening related expression and enhancedendoglucanase activity during softening. Plant Growth Regul. 56, 117–127.
deBruxelles, G.L., Peacock, W.J., Dennis, E.S., Dolferus, R., 1996. Abscisic acid inducesthe alcohol dehydrogenase gene in Arabidopsis. Plant Physiol. 111, 381–391.
Defilippi, B.G., Manriquez, D., Luengwilai, K., et al., 2009. Aroma volatiles:biosynthesis and mechanisms of modulation during fruit ripening. Adv. BotlRes. 50, 1–37.
Dolferus, R., De Bruxelles, G., Dennis, E.S., Peacock, W.J., 1994. Regulation of theArabidopsis Adh gene by anaerobic and other environmental stresses. Ann. Bot.74, 301–308.
Dudareva, N., Pichersky, E., Gershenzon, J., 2004. Biochemistry of plant volatiles.Plant Physiol. 135, 1893–1902.
Duncan, R., Colin, S., Bolwell, G.P., 1996. Inducible UDP-glucose dehydrogenase fromFrench bean (Phaseolus vulgaris L.) locates to vascular tissue and has alcoholdehydrogenase activity. Biochem. J. 313, 311–317.
Echeverria, G., Graell, J., Lopez, M.L., Lara, I., 2004. Volatile production, quality, andaroma-related enzyme activities during maturation of ‘‘Fuji” apples.Postharvest Biol. Technol. 31, 217–227.
Eklund, H., Ramaswamy, S., 2008. Three-dimensional structures of MDR alcoholdehydrogenases. Cell. Mol. Life Sci. 65, 3907–3917.
Gomez-Porras, J.L., Riano-Pachon, D.M., Dreyer, I., Mayer, J.E., Mueller-Roeber, B.,2007. Genome-wide analysis of ABA-responsive elements ABRE and CE3. BMCGenomics 8, 260–273.
Ingersoll, J.C., Rothenberg, M., Liedl, B.E., Folkerts, K., Garvin, D., Hanson, M.R., Doyle,J.J., Mutschler, M.A., 1994. A novel anther-expressed adh-homologous gene inLycopersicon esculentum. Plant Mol. Biol. 26, 1875–1891.
Jornvall, H., Persson, B., JeVery, J., 1987. Characteristics of alcohol/polyoldehydrogenases: the zinc-containing long-chain alcohol dehydrogenases. Eur.J. Biochem. 167, 195–201.
Kanellis, A.K., Solomos, T., Roubelakis-Angelakis, K.A., 1991. Suppression of cellulaseand polygalacturonase and induction of alcohol dehydrogenase isoenzymes inavocado fruit mesocarp subjected to low oxygen stress. Plant Physiol. 96, 269–274.
Kato-Noguchi, H., 2000. Abscisic acid and hypoxic induction of anoxia tolerance inroots of lettuce seedlings. J. Exp. Bot. 51 (352), 1939–1944.
Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of thehead of bacteriophage T4. Nature 277, 680–685.
Longhurst, T., Lee, E., Hinde, R., Brady, C., Speirs, J., 1994. Structure of the tomatoAdh2 gene and Adh2 pseudogenes, and a study of Adh2 gene expression in fruit.Plant Mol. Biol. 26, 1073–1084.
Loulakakis, C.A., Hassan, M., Gerasopoulos, D., Kanellis, A.K., 2006. Effects of lowoxygen on in vitro translation products of poly(A)+ RNA, cellulase and alcoholdehydrogenase expression in preclimacteric and ripening-initiated avocadofruit. Postharvest Biol. Technol. 39, 27–37.
Macnicol, P.K., Jacobsen, J.V., 2001. Regulation of alcohol dehydrogenase geneexpression in barley aleurone by gibberellin and abscisic acid. Physiol. Plant 111(4), 533–539.
Manrıquez, D., El-Sharkawy, I., Flores, F.B., El-Yahyaoui, F., Regad, F., Bouzayen, M.,Latche, A., Pech, J.C., 2006. Two highly divergent alcohol dehydrogenases ofmelon exhibit fruit ripening-specific expression and distinct biochemicalcharacteristics. Plant Mol. Biol. 61, 675–685.
Matton, D.P., Constable, P., Brisson, N., 1990. Alcohol dehydrogenase geneexpression in potato following elicitor and stress treatment. Plant Mol. Biol.14, 775–783.
Mitchell, W.C., Jelenkovic, G., 1995. Characterizing NAD- and NADP-dependentalcohol dehydrogenase enzymes of strawberries. J. Am. Soc. Hortic. Sci. 120 (5),798–801.
Pandit, S.S., Chidley, H.G., Kulkarni, R.S., Pujari, K.H., Giri, A.P., Gupta, V.S., 2009.Cultivar relationships in mango based on fruit volatile profiles. Food Chem. 114,363–372.
Parikh, H.R., Nair, G.M., Modi, V.V., 1990. Some structural changes during ripening ofmangoes (Mangifera indica var. Alphonso) by abscisic acid treatment. Ann. Bot.65, 121–127.
Peng, H.P., Chan, C.S., Shih, M.C., Yang, S.F., 2001. Signalling events in the hypoxicinduction of alcohol dehydrogenase gene in Arabidopsis. Plant Physiol. 126,742–749.
Peters, J., Frenkel, C., 2004. Relationship between alcohol dehydrogenase activityand low-temperature in two maize genotypes, Silverado F1 and Adh1–Adh2–doubly null. Plant Physiol. Biochem. 42, 841–846.
Peterson, G.L., 1977. A simplification of the protein assay method of Lowry et al.which is more generally applicable. Anal. Biochem. 83, 346–356.
Salas, J.J., Sanchez, J., 1998. Alcohol dehydrogenases from olive (Olea Europea) fruit.Phytochemistry 48 (1), 35–40.
Salvi, M.J., Gunjate, R.T., 1988. Mango breeding work in the Konkan region ofMaharashtra state. Acta Horticulturae 231, 100–102.
Sambrook, T., Fritsch, E.F., Maniatis, T., 1989. Molecular Cloning, ALaboratory Manual. Cold Spring Harbour Laboratory Press, Cold SpringHarbour.
Sane, V.A., Chourasia, A., Nath, P., 2005. Softening in mango (Mangifera indica var.Dashehari) is correlated with the expression of the early ethylene responsive,ripening related expansion gene, MiExpA1. Postharvest Biol. Technol. 38, 223–230.
Speirs, J., Correll, R., Cain, P., 2002. Relationship between ADH activity, ripeness andsoftness in six tomato cultivars. Sci. Hortic. 93, 137–142.
Speirs, J., Lee, E., Holt, K., Kim, Y.D., Scott, N.S., Loveys, B., Schuch, W., 1998. Geneticmanipulation of alcohol dehydrogenase levels in ripening tomato fruit affectsthe balance of some flavour aldehydes and alcohols. Plant Physiol. 117, 1047–1058.
Srivastava, A., Handa, A.K., 2005. Hormonal regulation of tomato fruit development:a molecular perspective. J. Plant Growth Regul. 24, 67–82.

1494 R.K. Singh et al. / Phytochemistry 71 (2010) 1485–1494
Straeten, D.V.D., Rodrigues, R.A., Gielen, P.J., Montagu, M.V., 1991. Tomato alcoholdehydrogenase expression during fruit ripening and under hypoxic conditions.FEBS 293, 39–42.
Tesniere, C., Pradal, M., El-Kereamy, A., Torregrosa, L., Chatelet, P., Roustan, J.P.,Chervin, C., 2004. Involvement of ethylene signalling in a non-climacteric fruit:new elements regarding the regulation of ADH expression in grapevine. J. Exp.Bot. 55, 2235–2240.
Tesniere, C., Verries, C., 2000. Molecular cloning and expression of cDNAs encodingalcohol dehydrogenases from Vitis vinifera L. during berry development. PlantSci. 157, 77–88.
Thompson, C.E., Salzano, F.M., Norberto de Souza, O., Freitas, L.B., 2007. Sequenceand structural aspects of the functional diversification of plant alcoholdehydrogenases. Gene 396, 108–115.
Trainotti, L., Tadiello, A., Casadoro, G., 2007. The involvement of auxin in theripening of climacteric fruits comes of age: the hormone plays a role of its ownand has an intense interplay with ethylene in ripening peaches. J. Exp. Bot. 58(12), 3299–3308.
Verries, C., Pradal, M., Chatelet, P., Torregrosa, L., Tesniere, C., 2004. Isolation andanalysis of the promoter of VvAdh2, a grapevine (Vitis vinifera L.) ripening-related gene. Plant Sci. 167, 1067–1074.
Yokoyama, S., Harry, D.E., 1993. Molecular phylogeny and evolutionary rates ofalcohol dehydrogenases in vertebrates and plants. Mol. Biol. Evol. 10, 1215–1226.
Zaldivar, C., Rivera-Cabrera, F., et al., 2009. Effect of refrigerated storage on aromaand alcohol dehydrogenase activity in tomato fruit. Postharvest Biol. Technol.54 (2), 93–100.