differential expression and localization of cftr and enac in mouse endometrium during...
TRANSCRIPT

www.elsevier.com/locate/cellbi
Cell Biology International 28 (2004) 433e439
Differential expression and localization of CFTRand ENaC in mouse endometrium during pre-implantation
Jian Zhi Yang, Louis Chukwuemeka Ajonuma, Lai Ling Tsang, Sun Yee Lam,Dewi Kenneth Rowlands, Lok Sze Ho, Chen Xi Zhou,
Yiu Wa Chung, Hsiao Chang Chan)
Epithelial Cell Biology Research Center, Department of Physiology, Faculty of Medicine,
The Chinese University of Hong Kong, Shatin, NT, Hong Kong SAR
Received 23 September 2003; revised 9 February 2004; accepted 16 March 2004
Abstract
Interaction between the cystic fibrosis transmembrane conductance regulator (CFTR), a CAMP-activated Cl� channel, and
epithelial NaC channel (ENaC) has been proposed as the major mechanism regulating uterine fluid absorption and secretion.Differential expression of these ion channels may give rise to dynamic changes in the fluid environment affecting variousreproductive events in the female reproductive tract. This study investigated the expression and localization of CFTR and ENaC
during the pre-implantation period. Semi-quantitative reverse transcriptase polymerase chain reaction and immunohistochemistrywere used to study the expression and localization of CFTR and ENaC in uteri collected from mature superovulated female mice.RT-PCR showed maximal ENaC and CFTR expression on day 3 after mating. Maximal immunoreactivity was also observed forboth ENaC and CFTR on day 3 after mating. However, ENaC was immunolocalized to the apical membrane of both luminal and
glandular epithelia, while CFTR was predominantly found in the stromal cells rather than the epithelial cells. Differential expressionand localization of CFTR and ENaC provide a molecular mechanism by which maximal fluid absorption can be achievedimmediately prior to implantation, to ensure the immobilization of the blastocyst necessary for implantation.
� 2004 International Federation for Cell Biology. Published by Elsevier Ltd. All rights reserved.
Keywords: CFTR; ENaC; Endometrium; Pre-implantation; Immunohistochemistry; RT-PCR
1. Introduction
Implantation is a complex and dynamic process thatis initiated by the adhesion between the embryonictrophectoderm and the epithelial cells of the uterineendometrium. This rate-limiting process in reproductionremains a poorly understood phenomenon, but it isthought to be greatly influenced by the uterine fluid,which undergoes dynamic changes during the endome-trial cycle and reproductive events (Long and Evans,1998). The disappearance of luminal fluid in the
) Corresponding author. Tel.: C852-26096839; fax: C852-
26035022.
E-mail address: [email protected] (H.C. Chan).
1065-6995/$ - see front matter � 2004 International Federation for Cell Bi
doi:10.1016/j.cellbi.2004.03.011
pre-implantation period, a characteristic feature ofmany species, is considered to be a mechanism leadingto the ‘‘closure’’ of the lumen, thereby enablingembryos to be held in contact with uterine epitheliumbefore initiation of implantation. While a couple ofmechanisms, such as endocytosis (Parr, 1983) andcharacteristic cytoplasmic projections known as ‘‘pino-podes’’ that appear on the luminal surface beforeimplantation (Nardo et al., 2002; Nikas et al., 1995),have long been thought to play a role in fluidabsorption during implantation, recent studies suggestthat the dynamic changes in uterine fluid may belargely determined by the interaction of epithelial ionchannels in the endometrium (Chan et al., 2002).
Fluid movement across epithelia is secondary to themovement of solutes, particularly ions. NaC and Cl�
ology. Published by Elsevier Ltd. All rights reserved.

434 J.Z. Yang et al. / Cell Biology International 28 (2004) 433e439
conductance in the apical membrane of epithelia isessential for electrolyte and fluid absorption andsecretion, respectively. Active NaC absorption drivesCl� counter-ion and fluid out of the lumen into theblood, while active Cl� secretion drives both NaC andfluid from the plasma into the lumen (Chan et al.,2000). Recent studies on cultured mouse endometrialepithelia have demonstrated the involvement of thecystic fibrosis transmembrane conductance regulator(CFTR), a cAMP-activated Cl� channel, in mediatinga number of neurohormonal-regulated secretory re-sponses in the uterus (Chan et al., 1997a, 1999;Deachapunya and O’Grady, 1998; Fong and Chan,1998; Matthews et al., 1998). On the other hand,previous studies have shown that uterine fluid volumeand its NaC concentration during pre-implantation arerelatively low compared to the other stages, suggestingthat NaC and water are reabsorbed, leading to the‘‘closure’’ of the lumen to promote successful embryoimplantation (Hoversland and Weitlauf, 1981; vanWinkle et al., 1983).
Recent molecular and electrophysiological studieshave shown the expression of an epithelial NaC channel(ENaC), which consists of three homologous subunits(a, b and g) and is inhibited by amiloride, as well as anamiloride-blockable NaC-dependent short-circuit cur-rent (Isc), in primary cultured mouse (Chan et al.,1997a, 1997b) and human uterine epithelia (Matthewset al., 1998). Amiloride-sensitive and NaC-dependentfluid absorption has recently been demonstrated in ratuterine glands by confocal microscopy, indicating theinvolvement of ENaC in mediating the fluid absorptionresponsible for the closure of the uterine lumen andimmobilization of the blastocyst necessary for implan-tation (Naftalin et al., 2002).
Previous studies in our laboratory have demon-strated differential expression of CFTR and ENaC atdifferent stages of the estrus cycle (Chan et al., 2002),providing a molecular basis for the long observedcyclic changes in uterine fluid volume (Long andEvans, 1998). Functional interaction of these ionchannels has also been demonstrated (Chan et al.,2001), suggesting that the interplay between CFTR andENaC may provide a mechanism for regulating thedynamic changes in uterine fluid. We hypothesized thatthe disappearance of uterine fluid during implantationmay be a result of down-regulation of CFTR and up-regulation of ENaC, leading to a net enhancement offluid absorption. The present study investigated theexpression and localization of ENaC subunits andCFTR during implantation using semi-quantitativereverse transcriptase polymerase chain reaction (RT-PCR) and immunohistochemistry, respectively. Theresults demonstrated differential expression of theseion channels during pre-implantation, indicating theirroles in this complex process.
2. Materials and methods
2.1. Materials
CFTR mouse monoclonal antibody was purchasedfrom Neomarkers, Fremont, CA, USA. ENaC (rabbitanti-epithelial sodium channel affinity purified polyclonalantibody)gwas purchased fromChemicon International,a was purchased from Zymed Lab, phosphate-bufferedsaline (PBS), reverse transcriptase polymerase chainreaction (RT-PCR) kit and primers were purchased fromGIBCO, Invitrogen, Grand Island, NY, USA. Pregnantmare’s serum gonadotropin (PMSG) was bought fromSigma Chemical Co., St. Louis, MO, USA, and humanchorionic gonadotropin (hCG; Pregnyl) was purchasedfrom Organon, Oss, Netherlands.
2.2. Animals and ovulation induction
Mature (8e10 weeks) male and female ICR micewere used in this study. Mice were kept in theLaboratory Animal Service Centre of the ChineseUniversity of Hong Kong prior to experiments, fedlaboratory chow and water ad libitum, maintainedunder controlled conditions of 12 h light and 12 h darkat 21e23 (C and 70e80% humidity. Experiments wereconducted in accordance with the university guidelineson animal experimentation.
Female mice received an intraperitoneal (IP) injectionof 10 IU of PMSG followed by an additional IP injectionof 10 IU of hCG 48 h later. Female mice were co-cagedwith sexually mature male ICR mice overnight. Femalemice were examined for copulation plugs the followingmorning and those that exhibited them were selected aspregnant and caged separately. Uteri were collected fromboth mated and control (superovulated but unmated)mice themorning after hCGadministration on days 1, 3, 4and 5. Uteri were immediately snap frozen in liquidnitrogen and stored at�70 (C forRNA isolationandRT-PCR, or fixed in 4% formalin for immunohistochemistry.
2.3. Semi-quantitative RT-PCR
Semi-quantitative RT-PCR was performed five timeson mRNA obtained from mice uteri at different stages.The specific oligonucleotide primers for GAPDH were:GAC CAC AGT CCA TGA CAT CAC TGC (sense)and GCT GTT GAA GTC GCA GGA GAC AAC(antisense), corresponding to nucleotides 565e904 withexpected cDNA of 340 bp. The specific oligonucleotideprimers for CFTR were: CAT CTT TGG TGT TTCCTA TGA TG (sense) and GTA AGG TCT CAG TTAGAA TTG AA (antisense), corresponding to nucleo-tides 1655e2135 with expected cDNA of 481 bp;a-ENaC: TCA CTT CAG CAC ATC TTC CACAGC TGC (sense) and GTA TCT GCC TAG CTG

435J.Z. Yang et al. / Cell Biology International 28 (2004) 433e439
GTC CAA GTG GGA (antisense), corresponding tonucleotides 2171e2960 with expected cDNA of 790 bp;g-ENaC: GAC TCT CTT CCT GAC ACA AAT GGTCCT (sense) and ACA CAC ATT CTC ACA CATACA CAT ACT (antisense), corresponding to nucleo-tides 2070e2793 with expected cDNA of 724 bp. Theconditions were: denaturation at 94 (C for 45 s; anneal-ing at 53 (C, 58 (C, 62 (C, 53 (C for 60 s; extension at72 (C for 60 s; 25, 30, 33 cycles for GAPDH, CFTR andENaC, respectively. Optimal amplification cycles aredetermined based on the linear relationship betweenthe amount of PCR product detected and the numberof amplification cycles. The intensity of the bands ofCFTR and ENaC subunits was normalized to that ofGAPDH, which was amplified simultaneously. Experi-ments in the absence of reverse transcriptase wereconducted as negative controls.
2.4. Immunohistochemistry
Tissues were fixed overnight in 4% formalin andembedded in paraffin wax after graded alcohol de-hydration. Five-micrometer sections were cut usinga Reichert-Jung Biocut Rotary Microtome 1130(Germany) and dried onto Superfrost microscope slides(Fisher brand, Fisher scientific), deparaffinized in xyleneand rehydrated in graded ethanol. Endogenous perox-idase activity was quenched using 3% hydrogenperoxide incubation for 30 min. The slides were thenplaced in a cooling jar with sodium citrate buffer ( pH 6)for antigen retrieval for 5 min. After cooling, slides wererinsed in 1 ! PBS and blocking solution was added for30 min and washed. Slides were then incubated over-night with the primary antibody (CFTR 1:1000; ENaC1:500, a and g subunits). Immunostaining was per-formed using a biotinylated secondary antibody,a horseradish peroxidaseestreptavidin conjugate anda substrateechromogen mixture. Reaction was revealedby incubation with 3,3-diaminobenzene hydrochloridesubstrate (DAB, Vector Laboratories, Inc., CA, USA)for another 15 min and counterstained with Harrishematoxylin. Tissue sections were observed under anOlympus epifluorescence microscope (Olympus IX-70,Japan). Brown deposits indicated sites of positiveimmunostaining. The experiments were repeated at leastthree times. Negative controls were included for alltissue sections by omitting the primary antibody.
3. Results
3.1. Uterine CFTR and ENaC expressionby semi-quantitative RT-PCR
The expressions of uterine CFTR and ENaC mRNAin mated and unmated mice were examined 1e5 days
after PMSG and hCG administration using semi-quantitative RT-PCR analysis. Experiments were re-peated at least five times in each group. As shown inFig. 1, ENaC a and g subunits in the uteri of mated ratsexhibited an expression pattern different from that ofunmated ones, in that the expression was greatlyenhanced on days 3 and 4 after mating compared tothat in unmated ones. CFTR expression in matedanimals was also different from that of unmated ones,with a maximum level of expression on day 3 in matedanimals, but a minimum level observed in the unmatedanimals on the same day (Fig. 1).
3.2. Immunohistochemistry
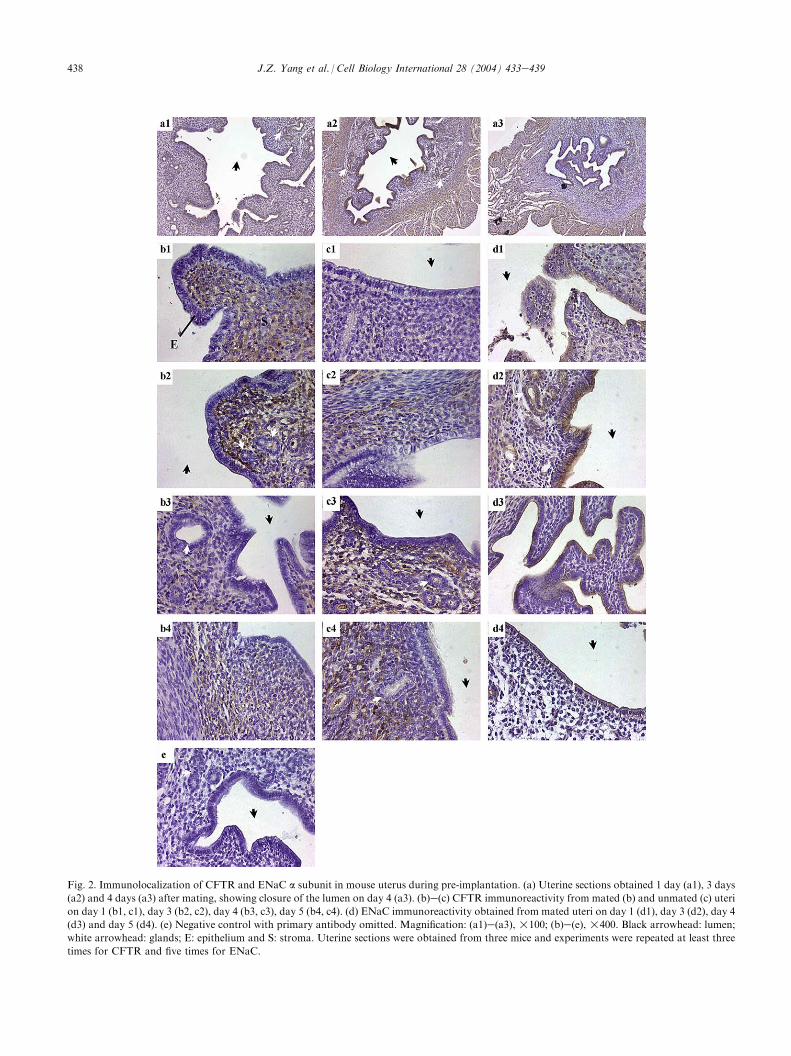
The uterine sections showed that the opening ofthe uterine lumen progressively decreased with time,with nearly complete closure observed on day 4(Fig. 2a1ea3), consistent with the characteristic ofimplantation. ENaC immunoreactivity was predomi-nantly localized to the apical side of both luminal andglandular epithelia in both mated and unmated uteri,with the strongest intensity seen 3 days after mating(Fig. 2d1ed4). In contrast to ENaC, CFTR immuno-reactivity was predominantly found in the stromalcells, but not the epithelial cells (neither luminal norglandular), in both mated and unmated uteri(Fig. 2b1ec4). Similar to the expression patternobserved by RT-PCR, the endometrium 3 days aftermating exhibited strongest CFTR immunoreactivity(Fig. 2b2), whereas the weakest immunoreactivity wasobserved in unmated uteri on the same day (Fig. 2c2).
4. Discussion
The present study demonstrated differential expres-sion of ENaC and CFTR during pre-implantation inmice, providing a molecular mechanism underlying thecharacteristic fluid absorption observed before implan-tation. ENaC mRNA expression, as demonstrated byRT-PCR, correlated well with its immunoreactivity,showing a maximal level on day 3, the day beforeimplantation (day 4), and a gradual decline in expres-sion afterwards. The immunolocalization of ENaC isconsistent with its localization found in other tissues(Blaug et al., 2001; Smith et al., 1998). The apicallocalization of ENaC enables influx of NaC, which canthen be pumped out of the cells by NaeK-ATPase,driving fluid absorption along the osmotic gradient.
The expression of ENaC in both luminal andglandular epithelia suggests that both epithelia areinvolved in NaC and fluid absorption. Consistent withthis finding, amiloride-sensitive and NaC-dependentfluid absorption has recently been observed in ratuterine glands (Naftalin et al., 2002). The fact that
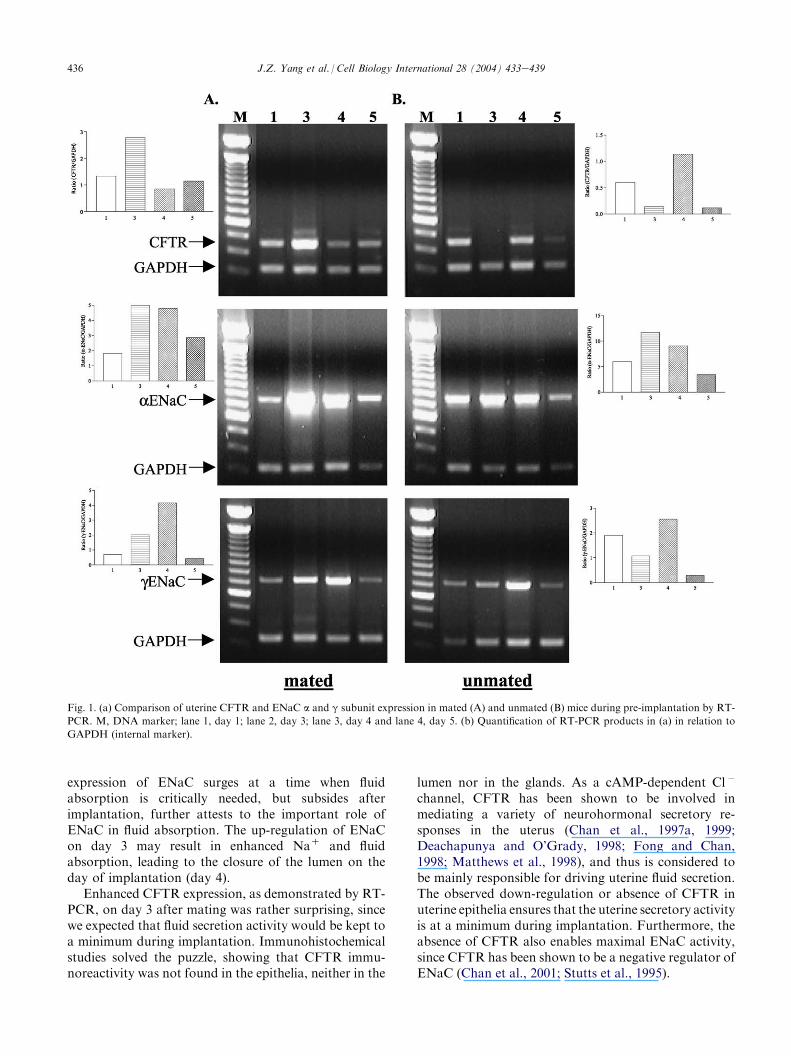
436 J.Z. Yang et al. / Cell Biology International 28 (2004) 433e439
Fig. 1. (a) Comparison of uterine CFTR and ENaC a and g subunit expression in mated (A) and unmated (B) mice during pre-implantation by RT-
PCR. M, DNA marker; lane 1, day 1; lane 2, day 3; lane 3, day 4 and lane 4, day 5. (b) Quantification of RT-PCR products in (a) in relation to
GAPDH (internal marker).
expression of ENaC surges at a time when fluidabsorption is critically needed, but subsides afterimplantation, further attests to the important role ofENaC in fluid absorption. The up-regulation of ENaCon day 3 may result in enhanced NaC and fluidabsorption, leading to the closure of the lumen on theday of implantation (day 4).
Enhanced CFTR expression, as demonstrated by RT-PCR, on day 3 after mating was rather surprising, sincewe expected that fluid secretion activity would be kept toa minimum during implantation. Immunohistochemicalstudies solved the puzzle, showing that CFTR immu-noreactivity was not found in the epithelia, neither in the
lumen nor in the glands. As a cAMP-dependent Cl�
channel, CFTR has been shown to be involved inmediating a variety of neurohormonal secretory re-sponses in the uterus (Chan et al., 1997a, 1999;Deachapunya and O’Grady, 1998; Fong and Chan,1998; Matthews et al., 1998), and thus is considered tobe mainly responsible for driving uterine fluid secretion.The observed down-regulation or absence of CFTR inuterine epithelia ensures that the uterine secretory activityis at a minimum during implantation. Furthermore, theabsence of CFTR also enables maximal ENaC activity,since CFTR has been shown to be a negative regulator ofENaC (Chan et al., 2001; Stutts et al., 1995).
437J.Z. Yang et al. / Cell Biology International 28 (2004) 433e439
1 3 4 5
1 3 4 5
1 3 4 5
1 3 4 5
1 3 4 5
1 3 4 50
1
2
3
0.0
0.5
1.0
1.5
0
1
2
3
4
5
0
5
10
15
0
1
2
3
4
5
0
1
2
3
Rat
io (
CFT
R/G
APD
H)
Rat
io (
α-E
NaC
/GA
PDH
)R
atio
(γ-
EN
aC/G
APD
H)
Rat
io (
γ-E
NaC
/GA
PDH
)R
atio
(α-
EN
aC/G
APD
H)
Rat
io (
CFT
R/G
APD
H)
Fig. 1 (continued ).
Taken together, the demonstrated differential expres-sion of CFTR and ENaC during pre-implantationsuggests that maximal fluid absorption prior to implan-tation may be a result of: (1) enhanced NaC absorptiondue to up-regulated ENaC expression; (2) reduced fluidsecretion due to down-regulation of CFTR; and (3)maximal ENaC activity, and thus fluid absorption, dueto removal of negative regulation in the absence ofCFTR. The differential expression of CFTR and ENaCduring the estrous cycle, as previously observed (Chanet al., 2002), and during implantation, as presentlydemonstrated, provides a mechanism whereby theendometrium can switch from fluid secretion to fluidabsorption, or vice versa, depending on physiologicalneeds, e.g. secretion for sperm transport and absorptionfor implantation.
Although it is not clear whether PMSG or hCG hasany direct effect on the expression of these ion channels,the results observed here could be explained by theaction of progesterone, which is secreted upon ovulationunder the stimulation of the superovulation agents.Down-regulation of CFTR and up-regulation of ENaChave been observed in rodents during diestrus, whenprogesterone levels are high (Chan et al., 2002).Progesterone has also been shown to inhibit CFTR
expression (Mularoni et al., 1995) and stimulate ENaCexpression (Tsang et al., 2001). Therefore, it isanticipated that the physiological processes in theendometrium after ovulation, which are involved inthe preparation for implantation under the influence ofprogesterone would include down-regulation of CFTRand up-regulation of ENaC, in addition to the well-known maximal growth of endometrial thickness andsecretory glands. The differences in the expressionpatterns between the mated and unmated animals,though both have been stimulated by superovulationagents, suggest that the embryo may also play a role inthe expression regulation of these ion channels. Whetherthe endometrium is responding to a signaling substancereleased from the embryo or directly to the physicalpresence of the embryo remains to be elucidated.
The most unexpected finding in the present study isthe first-time observation of CFTR expression in thestromal region of the endometrium. CFTR is normallyexpressed in epithelial cells (Hayslip et al., 1997;Mularoni et al., 1995), but is also found in lymphocytes(Krause et al., 1992). Therefore, the lymphocytes presentin the stromal region could be partly responsible for theimmunoreactivity of CFTR observed, but this cannot befully accounted for by the extensive and uniform
438 J.Z. Yang et al. / Cell Biology International 28 (2004) 433e439
Fig. 2. Immunolocalization of CFTR and ENaC a subunit in mouse uterus during pre-implantation. (a) Uterine sections obtained 1 day (a1), 3 days
(a2) and 4 days (a3) after mating, showing closure of the lumen on day 4 (a3). (b)e(c) CFTR immunoreactivity from mated (b) and unmated (c) uteri
on day 1 (b1, c1), day 3 (b2, c2), day 4 (b3, c3), day 5 (b4, c4). (d) ENaC immunoreactivity obtained from mated uteri on day 1 (d1), day 3 (d2), day 4
(d3) and day 5 (d4). (e) Negative control with primary antibody omitted. Magnification: (a1)e(a3), !100; (b)e(e), !400. Black arrowhead: lumen;
white arrowhead: glands; E: epithelium and S: stroma. Uterine sections were obtained from three mice and experiments were repeated at least three
times for CFTR and five times for ENaC.

439J.Z. Yang et al. / Cell Biology International 28 (2004) 433e439
staining throughout the stroma observed. The finding ofCFTR in the stroma of both mated and unmated uteri,both of which have been stimulated by superovulationagents, tends to suggest an action of the superovulationagents. Indeed, drugs for ovarian hyperstimulation havebeen shown to up-regulate CFTR in the uterus in vivo(Ajonuma et al., unpublished data).
However, the difference in the time course and levelsof expression and immunoreactivity between mated andunmated animals suggests that events involved in theprocess of implantation do affect CFTR expression inthe stroma. It is interesting to note that CFTRregulation in epithelial cells appears to differ from thatin stromal cells. Cytokines released during the process ofimplantation may be responsible, at least in part, for up-regulated stromal expression of CFTR, since interleukin1b has been shown to up-regulate CFTR expression(Cafferata et al., 2000). CFTR expression in the stromasuggests that it may play additional role(s) other than asan ion channel, since the stroma is known to be involvedin decidualization and angiogenesis during peri-implan-tation. The possible role of CFTR in these processesrequires further investigation.
In summary, the demonstrated differential expressionand localization of CFTR and ENaC during implanta-tion provides a molecular mechanism by which maximalfluid absorption can be achieved immediately prior toimplantation, to ensure the immobilization of theblastocyst necessary for implantation.
References
Blaug S, Hybiske K, Cohen J, Firestone GL, Machen TE, Miller SS.
ENaC and CFTR-dependent ion and fluid transport in mammary
epithelia. Am J Physiol 2001;281:c633e48.
Cafferata EG, Gonzalez-Guerrico AM, Giodano L, Pivetta OH,
Santa-Coloma TA. Interleukin-1 regulates CFTR expression in
human intestinal T84 cells. Biochim Biophys Acta 2000;1500:
241e8.
Chan HC, Fong SK, So SC, Chung YW, Wong PY. Stimulation of
anion secretion by beta-adrenoceptors in the mouse endometrial
epithelium. J Physiol 1997a;501:517e25.
Chan HC, Liu CQ, Fong SK, Law SH, Leung PS, Leung PY, et al.
Electrogenic ion transport in the mouse endometrium: functional
aspects of the cultured epithelium. Biochim Biophys Acta 1997b;
1356:140e8.
Chan LN, Chun YW, Leung PS, Liu CQ, Chan HC. Activation of an
adenosine 3#, 5#-cyclic monophosphate-dependent conductance in
response to neurohormonal stimuli in mouse endometrial epithelial
cells: the role of cystic fibrosis transmembrane conductance
regulator. Biol Reprod 1999;60:374e80.Chan LN, Wang XF, Tsang LL, Liu CQ, Chan HC. Suppression of
CFTR-mediated CL-secretion by enhanced expression of epithelial
NaC channels in mouse endometrial epithelium. Biochem Biophys
Res Commun 2000;276:40e4.
Chan LN, Wang XF, Tsang LL, So SC, Chung YW, Liu CQ, et al.
Inhibition of amiloride-sensitive NaD absorption by activation of
CFTR in mouse endometrial epithelium. Pflugers Arch 2001;
443(Suppl. 1):S132e6.Chan LN, Tsang LL, Rowlands DK, Rochelle LG, Boucher RC, Liu
CQ, et al. Distribution and regulation of ENaC subunit and CFTR
mRNA expression in murine female reproductive tract. J Membr
Biol 2002;185:165e76.Deachapunya C, O’Grady SM. Regulation of chloride secretion across
porcine endometrial epithelial cells by prostaglandins E2. J Physiol
1998;508:31e47.
Fong SK, Chan HC. Regulation of anion secretion by prostaglandin
E2 in the mouse endometrial epithelial epithelium. Biol Reprod
1998;58:1020e5.
Hayslip CC, Hao E, Usala SJ. The cystic fibrosis transmembrane
conductance regulator gene is expressed in the human endocervix
throughout the menstrual cycle. Fertil Steril 1997;67:636e40.
Hoversland RC, Weitlauf HM. The volume of uterine fluid in
‘‘implanting’’ and ‘‘delayed implanting’’ mice. J Reprod Fertil
1981;62:105e9.
Krause RD, Berta G, Rado TA, Bubin JK. Antisense oligonucleotides
to CFTR confer a cystic fibrosis phenotype on B-lymphocytes. Am
J Physiol 1992;263:C1147e51.
Long JA, Evans HM. The oestrus cycle in the rat and its associated
phenomena. In: Leuschner AO, editor. Memoirs of the University
of California, vol. 6. Los Angeles, CA; 1998.
Matthews CJ, McEwan GT, Redfern CP, Thomas EJ, Hirst BH.
Absorptive apical amiloride-sensitive NaC conductance in human
endometrial epithelium. J Physiol 1998;513:443e52.
Mularoni A, Beck L, Sadir R, Adessi GL, Nicollier M. Down-
regulation by progesterone of CFTR expression in endometrial
epithelial cells: a study by competitive RT-PCR. Biochem Biophys
Res Commun 1995;217:1105e11.
Naftalin RJ, Thiagarajah JR, Pedley KC, Pocock VJ, Milligan SR.
Progesterone stimulation of fluid absorption by the rat uterine
gland. Reproduction 2002;123:633e8.
Nardo LG, Sabatini L, Rai R, Nardo F. Pinopode expression during
human implantation. Eur J Obstet Gynecol Reprod Biol 2002;101:
104e8.
Nikas G, Drakakis P, Loutradis D, Mara-Skoufari C, Koumantakis E,
Michalas S, et al. Uterine pinopodes as markers of the ‘nidation
window’ in cycling women receiving exogenous oestradiol and
progesterone. Hum Reprod 1995;10:1208e13.
Parr MB. Relationship of uterine closure to ovarian hormones and
endocytosis in the rat. J Reprod Fertil 1983;68:185e8.Smith PR, Macklre SA, Weiser PC, Brooker DR, Ahn YJ, Harte BJ,
et al. Expression and localization of epithelial sodium channel in
mammalian urinary bladder. Am J Physiol 1998;274:F91e6.
Stutts MJ, Canessa CM, Olsen JC, Hamrick M, Cohn JA, Rossier BC,
et al. CFTR as a CAMP-dependent regulator of sodium channels.
Science 1995;269:847e50.
Tsang LL, Chan LN, Liu CQ, Chan HC. Effect of phenol red and
steroid hormones on cystic fibrosis transmembrane conductance
regulator in mouse endometrial epithelial cells. Cell Biol Int 2001;
25:1021e4.
van Winkle LJ, Campione AL, Webster DP. Sodium ion concen-
trations in uterine flushings from ‘‘implanting’’ and ‘‘delayed
implanting’’ mice. J Exp Zool 1983;226:321e4.