cyanobacterial h2 production — a comparative analysis
TRANSCRIPT

12 Springer
Dear Author: Please find attached the final pdf file of your contribution, which can be viewed using the Acrobat Reader, version 3.0 or higher. We would kindly like to draw your attention to the fact that copyright law is also valid for electronic products. This means especially that:
• You may print the file and distribute it amongst your colleagues in the scientific community for scientific and/or personal use.
• You may make your article published by Springer-Verlag available on your personal home page provided the source of the published article is cited and Springer-Verlag and/or other owner is mentioned as copyright holder. You are requested to create a link to the published article in Springer's internet service. The link must be accompanied by the following text: "The original publication is available at springerlink.com". Please use the appropriate DOI for the article. Articles disseminated via SpringerLink are indexed, abstracted and referenced by many abstracting and information services, bibliographic networks, subscription agencies, library networks and consortia.
• Without having asked Springer-Verlag for a separate permission your institute/your company is not allowed to place this file on its homepage.
• You may not alter the pdf file, as changes to the published contribution are prohibited by copyright law.
• Please address any queries to the production editor of the journal in question, giving your name, the journal title, volume and first page number.
Yours sincerely, Springer-Verlag

ORIGINAL ARTICLE
Kathrin Schutz Æ Thomas Happe Æ Olga Troshina
Peter Lindblad Æ Elsa Leitao Æ Paulo Oliveira
Paula Tamagnini
Cyanobacterial H2 production — a comparative analysis
Received: 29 April 2003 / Accepted: 15 August 2003 / Published online: 15 October 2003� Springer-Verlag 2003
Abstract Several unicellular and filamentous, nitrogen-fixing and non-nitrogen-fixing cyanobacterial strainshave been investigated on the molecular and the physi-ological level in order to find the most efficient organismsfor photobiological hydrogen production. These strainswere screened for the presence or absence of hup and hoxgenes, and it was shown that they have different sets ofgenes involved in H2 evolution. The uptake hydrogenasewas identified in all N2-fixing cyanobacteria, and some ofthese strains also contained the bidirectional hydroge-nase, whereas the non-nitrogen fixing strains only pos-sessed the bidirectional enzyme. In N2-fixing strains,hydrogen was mainly produced by the nitrogenase as aby-product during the reduction of atmospheric nitrogento ammonia. Therefore, hydrogen production wasinvestigated both under non-nitrogen-fixing conditionsand under nitrogen limitation. It was shown that thehydrogen uptake activity is linked to the nitrogenaseactivity, whereas the hydrogen evolution activity of thebidirectional hydrogenase is not dependent or evenrelated to diazotrophic growth conditions. With regardto large-scale hydrogen evolution by N2-fixing
cyanobacteria, hydrogen uptake-deficient mutants haveto be used because of their inability to re-oxidize thehydrogen produced by the nitrogenase. On the otherhand, fermentative H2 production by the bidirectionalhydrogenase should also be taken into account in furtherinvestigations of biological hydrogen production.
Keywords Cyanobacterium Æ Hydrogen ÆHydrogenase Æ Nitrogenase Æ Photobiological H2
production Æ Screening (H2 production)
Abbreviations Chl: chlorophyll Æ MV: methyl viologen
Introduction
Photobiological production of H2 by microorganisms isof great public interest since it promises a renewableenergy carrier from nature�s most plentiful resources,solar energy and water. Cyanobacteria and green algaeare the only organisms currently known to be capable ofboth oxygenic photosynthesis and hydrogen production(Houchins 1984; Happe and Naber 1993). In cyano-bacteria (formerly called ‘‘blue-green algae’’), hydrogenis produced (i) by a light-dependent reaction catalysedby nitrogenase or (ii) in dark anaerobic conditions by ahydrogenase (Rao and Hall 1996; Hansel and Lindblad1998), while in the green algae hydrogen is producedphotosynthetically (Melis and Happe 2001).
Cyanobacteria comprise a large and diverse group ofoxygenic photoautotrophic prokaryotes, in which manystrains are able to reduce atmospheric dinitrogen toammonia. In some filamentous cyanobacteria, nitrogenfixation is restricted to specialized cells (called hetero-cysts), which differentiate as consequence of nitrogenstarvation. In cyanobacteria, up to three enzymes canbe directly involved in hydrogen metabolism: nitroge-nase(s) which produces H2 concomitantly with nitrogenfixation, a membrane-bound uptake hydrogenase whichre-oxidizes the H2 evolved by nitrogenase, and thebidirectional hydrogenase that catalyses both oxidation
Planta (2004) 218: 350–359DOI 10.1007/s00425-003-1113-5
E. Leitao Æ P. Oliveira Æ P. Tamagnini (&)Institute for Molecular and Cell Biology, University of Porto,Rua do Campo Alegre 823, 4150-180 Porto, PortugalE-mail: [email protected]: +351-22-6099157
K. Schutz Æ T. HappeBiochemie der Pflanzen, AG Photobiotechnologie,Ruhr-Universitat-Bochum, 44780 Bochum, Germany
O. Troshina Æ P. LindbladDepartment of Physiological Botany, EBC,Uppsala University, Villavagen 6, 752 36 Uppsala, Sweden
O. TroshinaInstitute of Basic Biological Problems,Russian Academy of Sciences,142 290 Pushchino, Moscow region, Russia
P. Oliveira Æ P. TamagniniDepartment of Botany, Faculty of Sciences, University of Porto,Rua do Campo Alegre 1191, 4150-181 Porto, Portugal

of molecular hydrogen and reduction of protons (forreviews, see Houchins 1984; Schulz 1996; Bergman et al.1997; Appel and Schulz 1998; Hansel and Lindblad1998; Tamagnini et al. 2002).
Two different NiFe-hydrogenases have been found incyanobacteria, and both have been in part characterizedon a biochemical and molecular basis. The membrane-bound uptake hydrogenase transfers/donates electronsfrom H2 to the photosynthetic and the respiratoryelectron transport chains, thereby regaining energy andreducing equivalents lost as H2 during nitrogen fixation(Houchins 1984). Mutational analyses support theprobability that the highly conserved dimeric uptakehydrogenase re-oxidizes the hydrogen gas evolved by thenitrogenase during nitrogen fixation (Happe et al. 2000;Lindberg et al. 2002; Lindblad et al. 2002; Masukawaet al. 2002). The reversible, bidirectional hydrogenasecatalyzes both uptake and evolution of molecularhydrogen and shows high similarities to the NAD+-reducing hydrogenase of, for example, Ralstoniaeutropha. The heterotetrameric enzyme consists of anH2-oxidizing dimer HoxYH and a diaphorase part,HoxFU, transferring electrons to NAD+ (Boison et al.1996). In contrast to the known NAD+-reducinghydrogenases from other bacteria, a putative fifth sub-unit (HoxE) was recently determined by isolating thenative protein as well as the corresponding gene cluster(Schmitz et al. 2002). A number of reports suggest afunction for the bidirectional hydrogenase in disposingof excess of reducing equivalents during dark anaerobicconditions (Stal and Moezelaar 1997; Troshina et al.2002) or during periods of adaptation to higher lightintensities (Appel et al. 2000), although the high affinityof this hydrogenase for H2 may suggest a function forthe hydrogenase in recapturing exogenous H2 undercertain conditions (Houchins 1984).
In filamentous heterocystous cyanobacteria, hydro-gen evolution is mainly catalysed by the nitrogenase inthe heterocysts. The conventional Mo-dependent en-zyme complex consists of a dinitrogenase reductase (Feprotein) and a dinitrogenase (FeMo protein) subunit(Haselkorn and Buikema 1992), the latter of whichcatalyzes nitrogen reduction. The biochemical featuresof cyanobacterial nitrogenase and the structural genes(nifHDK) are very similar to the nitrogenase of otherdiazotrophic organisms (Bohme 1998). Alternative ni-trogenases have been found in A. variabilis and othercyanobacterial strains (Kentemich et al. 1988, 1991;Bishop and Premakumar 1992; Thiel 1993; Thiel et al.1995, 1997), but they differ from the conventional en-zyme physically, chemically, and by their catalyticproperties, and they seem to allocate a higher proportionof electrons for the reduction of protons to hydrogen(Tsygankov et al. 1998).
Several filamentous strains have been investigatedregarding the presence or absence of DNA sequenceshomologous to different hydrogenase genes (Tamagniniet al. 2000). Physiological studies on cyanobacterialhydrogen metabolism have been limited to a few strains
like the nitrogen-fixing filamentous heterocystous An-abaena sp. PCC 7120 (Lindblad et al. 2002), A. varia-bilis (Serebriakova et al. 1994; Tsygankov et al. 1999;Happe et al. 2000; Borodin et al. 2000; Fedorov et al.2001), and Nostoc punctiforme PCC 73102 (Oxelfeltet al. 1995; Tamagnini et al. 1997; Lindberg et al. 2002)as well as the non-N2-fixing strains Synechococcus sp.PCC 6301 (Schmitz and Bothe 1996), Synechocystis sp.PCC 6803 (Appel et al. 2000), and Spirulina platensis(Aoyama et al. 1997). Until now it has been difficult tomake comparisons among these results since the exper-iments were performed in several groups, the strainswere grown under different conditions, and the mea-surements were performed in different ways.
The present study therefore focuses on a physiologi-cal and molecular investigation of the hydrogenmetabolism of different unicellular and filamentous cy-anobacteria strains in order to find the most promisingcandidates for large-scale photobiological hydrogenproduction. Whenever it was possible, the strains werecultivated under identical growth conditions.
Materials and methods
Organisms and growth conditions
All cyanobacterial strains used in this work are listed in Table 1. Theculture collections from which the cyanobacteria were obtained arethe following: ATCC, American Type Culture Collection (Rock-ville, MD, USA); PCC, Pasteur Culture Collection (Paris, France);CCAP, Culture Collection of Algae and Protozoa (Ambleside, UK);HCC, Culture Collection of the Hawaii Natural Energy Institute,University of Hawaii at Manoa (Honolulu, HI, USA).
The cyanobacterial cultures were grown in glass tubes con-taining 200 ml BG110, or BG11 medium (when stated) (Castenholz1988) supplemented with 10 mM Hepes–NaOH (pH 7.5) at 25 �C,under continuous irradiance of about 100 lmol photons m)2 s)1
from two sides or, when stated, in an alternating light/dark cycle(12/12 h). Cultures were bubbled with air containing 1% (v/v) CO2
or with air only. The medium for the Anabaena variabilis hup)
mutant (AVM13) contained additionally 50 lg ml)1 of neomycin.The Anabaena sp. PCC 7120 mutant (AMC414) was maintained inmedium containing 1 lg ml)1 of streptomycin. However, since themutant is stable (see Lindblad et al. 2002), experimental cells weregrown without the addition of the antibiotic.
To de-repress nitrogenase activity in Leptolyngbya sp. PCC73110, a 3-day-old BG11-grown culture was transferred to BG110and bubbled with air and CO2 (2%) for 24 h, before sparging withargon for another 24 h.
Optical microscopy and image processing
Light micrographs were obtained with an Olympus BX50 micro-scope equipped with an Olympus DP10 digital camera. Imageprocessing was performed using Adobe Photoshop 6.0 (AdobeSystems Incorporated, USA).
Nitrogenase activity (C2H2 reduction), H2 productionand H2 uptake assays
Nitrogenase activity was measured using the acetylene-reductionassay. Acetylene-reduction and H2-production assays were carriedout in glass vials (14 ml) by incubation of the samples (2 ml) under
351

continuous irradiance of 100 lmol photons m)2 s)1 from the bot-tom or, when needed, in darkness at 25 �C using a rotary shaker.The gas phase contained argon in the case of H2 production and20% (v/v) C2H2 in air or argon for C2H2 reduction. The gas phasewas analyzed by gas chromatography (Oxelfelt et al. 1995; Happeet al. 2000).
Bidirectional hydrogenase activity was assayed in cell suspen-sions by determining the rate of methyl viologen (MV)-dependentH2 evolution by gas chromatography (Tamagnini et al. 1997). Thereaction mixture contained 50 mM K-phosphate buffer (pH 7.0),5 mM MV and 10 mM sodium dithionite. Activities are expressedas nmol of H2, or C2H4, produced per lg chlorophyll (Chl) a andper h. H2 uptake was measured using a Hansatech DW1 H2-elec-trode (Hansatech, Norfolk, UK) as described previously (Oxelfeltet al. 1995). The H2 uptake activity is expressed as nmol of H2
consumed per lg Chl a and per h. For Gloeothece, the activities arereferred to the protein content since the polysaccharide sheatharound the cells (see Fig. 1b) prevents an accurate determination ofthe Chl a content.
Chlorophyll a and protein content
The total Chl a content of the cells was determined spectrophoto-metrically at 663 nm in 90% (v/v) methanol extracts (Mackinney1941). The protein content was estimated by using CoomassieBrilliant Blue G-250 with bovine serum albumin (BSA; Sigma) asstandard (Peterson 1983).
Isolation of DNA, PCR, and other molecular-biological techniques
Genomic DNA was extracted, and PCR was carried out using thethermal cycler GeneAmp PCR system 2400 (Perkin-Elmer)according to methods described previously (Tamagnini et al. 1997).In order to reduce the extracellular polysaccharides (see Fig. 1b) of
Gloeothece sp. ATCC 27152, the pelleted cells were washed twiceprior to standard DNA extraction (washing buffer: 50 mM Tris–HCl with 50 mM NaCl and 5 mM EDTA, pH 8.0; Fiore et al.2000). After precipitation, the DNA was washed once with thebuffer mentioned above, precipitated and collected using a pipettetip, before being resuspended in water (Oxelfelt et al. 2002).
The PCR primers and PCR-generated probes are listed inTable 2. The oligonucleotide sequences have already been published(Tamagnini et al. 2000; Hansel et al 2001), with the exception ofGloS1A (5¢-gctaggagacaatcttcaac-3¢), LMH4¢A (5¢-ggctatctccttaa-cgaaga-3¢), and LMH4¢B (5¢-ggctaactgacgcagat-3¢). The identities ofall PCR-generated DNA probes were confirmed by sequencing.
Agarose-gel electrophoresis was performed by standard proto-cols with the use of 1· TAE or 1· TBE buffer (Sambrook et al.1989). DNA fragments were isolated from agarose gels using theQIAEX II Gel Extraction Kit (Qiagen) according to the manu-facturer�s instructions. Sequencing of the purified PCR product wasperformed using the ABI Prism BigDye Terminator CycleSequencing Ready Reaction Kit following the manufacturer�sprotocol, the thermal cycler mentioned above, and an ABIPrism 310 or 377 Genetic Analyser.
For Southern hybridizations, about 500 ng genomic DNA wasdigested with the restriction endonucleases EcoRI, HindIII or acombination of HindIII and EcoRI. The separation and blotting ofthe DNA was performed according to standard procedures. TheDNA probes were labelled with the AlkPhos Direct labelling kit(Amersham Pharmacia Biotech), and hybridization was carried outat 57 �C. The chemiluminescence of alkaline phosphatase wasdetected using the CDP-Star detection reagent (Amersham Phar-macia Biotech), and by exposing the membranes for 4–24 h to anX-ray film (Ortho CP-G Plus; AGFA). In each Southern experi-ment a positive control was included to confirm the specificity ofthe DNA probe and to verify the overall conditions of therespective hybridization. Results from Southern hybridizationsperformed previously were obtained using 32P-labelled probes(Tamagnini et al. 2000). However, no significant differences wereobserved in a control experiment using the two methods.
Table 1 Presence/absence of sequences homologous to hup (uptake hydrogenase), hox(bi-directional hydrogenase), and hypF (hydrog-enases accessory) genes in selected cyanobacterial strains.P1 toP10 Different probes; for details see Table 2. Bold indicates homologousprobe.np Not performed,WT wild type
Organisma hup genes hox genes hypF References
hupS hupL hoxY hoxH
UnicellularGloeothece sp. ATCC 27152 +P1, P3 +P4 )P6, P7 )P8, P9 )P10 This workSynechocystis sp. PCC 6803 ) ) + + + Kaneko et al. 1996;
Nakamura et al. 1998Filamentous non-heterocystousLeptolyngbya sp. PCC 73110 +/)P1 +P4, P5 )P7 )P9 )P10 This work; Tamagnini et al. 2000Lyngbya majuscula CCAP 1446/4 + P2, P3 + P5 + P7 )P8, P9 +P10 This work
Filamentous heterocystousAnabaena variabilis ATCC 29413 (WT) +P1 +P4 + P6, P7 + P8, P9 +P10 This work; Schmitz et al. 1995;
Tamagnini et al. 2000;Happe et al. 2000
A. variabilis ATCC 29413 (mutant AVM13) b b + + np Happe et al. 2000Anabaena sp. PCC 7120 (WT) + P1 + P4 +P6 +P8, P9 + Carrasco et al. 1995; Tamagnini
et al. 2000; Kaneko et al. 2001Anabaena sp. PCC 7120 (mutant AMC414) c c + + np Carrasco et al. 1998;
Lindblad et al. 2002Nostoc punctiforme PCC 73102 +P1, P2, P3 +P4, P5 )P6 )P8, P9 + P10 Oxelfelt et al. 1998;
Tamagnini et al. 1997, 2000Nostoc sp. Mitsui 38901 (HCC 1048) +P1 +P4 )P6 )P8, P9 +P10 This work; Tamagnini et al. 2000Nostoc sp. Mitsui 56111 (HCC 1061) +P1 +P4 +P6 +P8 +P10 This work; Tamagnini et al. 2000Nostoc sp. Mitsui 91911 (HCC 1075) +P1 +P4 +P6 +P8 +P10 This work; Tamagnini et al. 2000
aAll cyanobacterial strains are N2-fixing, with the exception ofSynechocystis sp. PCC 6803bHydrogen-uptake-deficient mutant in which the completehupS, and the 5¢ end of hupL were deletedcHydrogen-uptake-deficient mutant by knockout of the xisC gene responsible for the hupL rearrangement
352

Results
In order to find an efficient strain for photobiologicalhydrogen production, several cyanobacteria (see Table 1and Fig. 1) were selected according to specific criteria: (i)morphological diversity ranging from unicellular to fila-mentous heterocystous strains, i.e. temporal versus spa-tial separation of photosynthesis and nitrogen fixation;(ii) available genomes (Synechocystis sp. PCC 6803,Anabaena/Nostoc sp. PCC 7120, Nostoc punctiforme
ATCC 29133/PCC 73102); (iii) genetic diversity withrespect to hydrogenases (Nostoc punctiforme PCC 73102,Nostoc sp. Mitsui 38901, Nostoc sp. Mitsui 56111,Nostoc sp.Mitsui 91911); and (iv) existence of hydrogen-uptake-deficient mutants ofAnabaena sp. PCC 7120 andAnabaena variabilis ATCC 29413.
Initially, the cyanobacterial strains were screened forthe presence of sequences homologous to both hydrog-enase structural genes (hup and hox), and to thehydrogenase(s) accessory gene hypF by heterologous andhomologous Southern hybridizations. The results ob-
Fig. 1a–k Light micrographs ofseveral cyanobacteria used inthis study. All strains are N2-fixing, with the exception ofSynechocystis sp. PCC 6803.The cyanobacterial cultures,from which these images wereobtained, were grown in BG110,with the exception ofSynechocystis sp. andLeptolyngbya sp. (BG11), andL. majuscula (ASW:BG; Allen1968), at 25 �C underalternating light/dark cycles(16/8 h). a, b Unicellularstrains. a Synechocystis sp.PCC 6803. b Gloeothece sp.ATCC 27152; the colonialstructure is maintained by afirm sheath. c, f Filamentousnon-heterocystous strains. cLeptolyngbya sp. PCC 73110.d–f Mat-forming Lyngbyamajuscula CCAP 1446/4. dGeneral view of the culture. eDetail of trichome breakage. fApical part of a trichomeenclosed by a heavy sheath. g–kFilamentous heterocystousstrains. g Anabaena sp. PCC7120. h, i Anabaena variabilisATCC 29413 wild type (h) andmutant AVM13 (i). j Nostocpunctiforme PCC 73102. kNostoc sp. Mitsui 91911 (HCC1075). het Heterocyst. a, c, d, h,i, k Bright field; b, g, j phasecontrast; e, f dark field. Bars =10 lm
353
tained in the present study were combined with dataavailable in the literature and from genome databases,and are summarized in Table 1. As expected, the uptake-hydrogenase structural genes (hupSL) were identified inall N2-fixing cyanobacteria, whereas the structural genesencoding the hydrogenase part of the bidirectionalenzyme were not present in all strains. As confirmed bythe genome sequence, hoxYH was absent from Nostocpunctiforme PCC 73102 (Tamagnini et al. 1997; Meekset al. 2001), and most probably absent from the Nostocsp. Mitsui 38901, as well as the filamentous non-het-erocystous Leptolyngbya sp. PCC 73110, and theunicellular Gloeothece sp. ATCC 27152. However, aresidual bidirectional hydrogenase activity could bedetected in the latter strain (see below).
The accessory gene hypF could be detected in allfilamentous heterocystous cyanobacteria tested, inde-pendently of their hydrogenase content, i.e. the gene wasdetected in strains lacking the bidirectional enzyme (seeTable 1; Hansel et al. 2001) as well as in Synechocystissp. PCC 6803, a strain without the uptake hydrogenase.The heterologous probe obtained from Nostoc puncti-forme PCC 73102 failed to hybridize with genomicDNA from Leptolyngbya sp. PCC 73110 and Gloeot-hece sp. ATCC 27152. However, since only one probewas used it is premature to exclude the presence of hypFin these two strains.
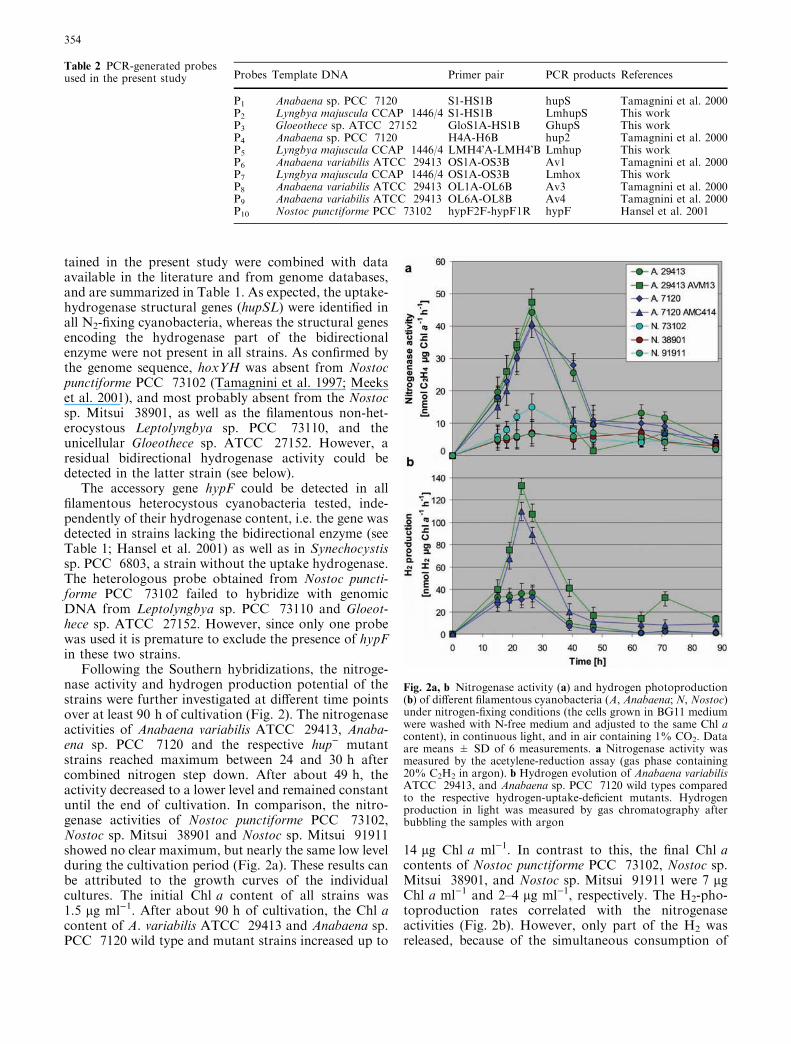
Following the Southern hybridizations, the nitroge-nase activity and hydrogen production potential of thestrains were further investigated at different time pointsover at least 90 h of cultivation (Fig. 2). The nitrogenaseactivities of Anabaena variabilis ATCC 29413, Anaba-ena sp. PCC 7120 and the respective hup) mutantstrains reached maximum between 24 and 30 h aftercombined nitrogen step down. After about 49 h, theactivity decreased to a lower level and remained constantuntil the end of cultivation. In comparison, the nitro-genase activities of Nostoc punctiforme PCC 73102,Nostoc sp. Mitsui 38901 and Nostoc sp. Mitsui 91911showed no clear maximum, but nearly the same low levelduring the cultivation period (Fig. 2a). These results canbe attributed to the growth curves of the individualcultures. The initial Chl a content of all strains was1.5 lg ml)1. After about 90 h of cultivation, the Chl acontent of A. variabilis ATCC 29413 and Anabaena sp.PCC 7120 wild type and mutant strains increased up to
14 lg Chl a ml)1. In contrast to this, the final Chl acontents of Nostoc punctiforme PCC 73102, Nostoc sp.Mitsui 38901, and Nostoc sp. Mitsui 91911 were 7 lgChl a ml)1 and 2–4 lg ml)1, respectively. The H2-pho-toproduction rates correlated with the nitrogenaseactivities (Fig. 2b). However, only part of the H2 wasreleased, because of the simultaneous consumption of
Fig. 2a, b Nitrogenase activity (a) and hydrogen photoproduction(b) of different filamentous cyanobacteria (A, Anabaena; N, Nostoc)under nitrogen-fixing conditions (the cells grown in BG11 mediumwere washed with N-free medium and adjusted to the same Chl acontent), in continuous light, and in air containing 1% CO2. Dataare means ± SD of 6 measurements. a Nitrogenase activity wasmeasured by the acetylene-reduction assay (gas phase containing20% C2H2 in argon). b Hydrogen evolution of Anabaena variabilisATCC 29413, and Anabaena sp. PCC 7120 wild types comparedto the respective hydrogen-uptake-deficient mutants. Hydrogenproduction in light was measured by gas chromatography afterbubbling the samples with argon
Table 2 PCR-generated probesused in the present study Probes Template DNA Primer pair PCR products References
P1 Anabaena sp. PCC 7120 S1-HS1B hupS Tamagnini et al. 2000P2 Lyngbya majuscula CCAP 1446/4 S1-HS1B LmhupS This workP3 Gloeothece sp. ATCC 27152 GloS1A-HS1B GhupS This workP4 Anabaena sp. PCC 7120 H4A-H6B hup2 Tamagnini et al. 2000P5 Lyngbya majuscula CCAP 1446/4 LMH4�A-LMH4�B Lmhup This workP6 Anabaena variabilis ATCC 29413 OS1A-OS3B Av1 Tamagnini et al. 2000P7 Lyngbya majuscula CCAP 1446/4 OS1A-OS3B Lmhox This workP8 Anabaena variabilis ATCC 29413 OL1A-OL6B Av3 Tamagnini et al. 2000P9 Anabaena variabilis ATCC 29413 OL6A-OL8B Av4 Tamagnini et al. 2000P10 Nostoc punctiforme PCC 73102 hypF2F-hypF1R hypF Hansel et al. 2001
354

H2 by the uptake hydrogenase. In contrast to this, theuptake-hydrogenase-deficient mutant strains of A. vari-abilis ATCC 29413 and Anabaena sp. PCC 7120evolved several times more H2 than the wild types(Fig. 2b) and other heterocystous strains included in thepresent study (data not shown). Unlike the mutantstrains, showing no H2 consumption, other heterocys-tous Anabaena and Nostoc wild-type strains showed H2
uptake activities ranging from 14 to 50 nmol (lgChl a))1 h)1.
An H2-electrode was used to compare the H2 pro-duction of A. variabilis ATCC 29413 and Anabaena sp.PCC 7120 with that of the respective mutants (data notshown). These measurements were performed usingbatch cultures grown in an orbital shaker, under N2-fixing conditions at 25 �C in an alternating light/darkcycle (16/8 h). In the wild types we could only detect anet H2 uptake in light, while a clear H2 production wasobserved for both mutants. Similar results were previ-ously obtained for Nostoc punctiforme ATCC 29133and its hydrogenase-free mutant (Lindberg et al. 2002).In agreement with the results obtained by gas chroma-tography, the A. variabilis mutant AVM13 showedhigher H2 production rates than the Anabaena sp. PCC7120 mutant, reaching 28 nmol (lg Chl a))1 h)1.
The filamentous non-heterocystous cyanobacteriumLyngbya majuscula was found to be able to grow dia-zotrophically in continuous light and air, but culturesshowed only low acetylene-reduction activities whenmeasured in light and air (data not shown). However,when air was replaced with an atmosphere of argon thelevels of acetylene reduction were higher (Table 3).Interestingly, the rates of nitrogenase activity were onlyhalf of that in L. majuscula cultures under a 12/12 hlight/dark photocycle (Table 3). The levels of H2 pho-
toproduction under an atmosphere of argon were verylow, but under dark anaerobic conditions L. majusculashowed H2-evolution activity (Table 3) that startedimmediately after a change to anaerobic conditions andwas observed in cultures grown in air without pre-liminary anaerobic adaptation. This cyanobacteriumalso showed high H2 consumption measured by using anH2-electrode (Table 3), which may be the result of thecombined activity of two enzymes: a bidirectional andan uptake hydrogenase. High levels of bidirectionalhydrogenase activity, measured as H2 evolution fromreduced MV, were detected in aerobically grown N2-fixing L. majuscula (Table 4).
Unlike Lyngbya, Leptolyngbya was not able to growand fix nitrogen aerobically. Nitrogenase activity couldbe de-repressed in Leptolyngbya by nitrogen limitationunder micro-oxic conditions. However, the rates ofacetylene reduction and in vivo H2 production in thelight were quite low [5.7 nmol C2H4 (lg Chl a)
)1 h)1 and5.9 nmol H2 (lg Chl a))1 h)1, respectively]. The H2-uptake activity was 15.1 nmol H2 (lg Chl a))1 h)1 in thelight, whereas there was no activity in the dark. MV-dependent H2 production could not be measured at all,in agreement with the negative results from the Southernblot hybridizations (Table 1).
The unicellular cyanobacterium Gloeothece sp.ATCC 27152 is known to be able to fix N2 aerobicallyin the dark phase, when grown in alternating light–darkcycles (Gallon et al. 1988; Reade et al. 1999). Acetylenereduction, H2 photoproduction, and hydrogen uptakewere assayed in Gloeothece samples taken from the darkphase (4 h) of cultures grown under 12/12 h light/darkcycles. The highest nitrogenase activities were measuredin the light under argon. Activities were lower whenassays were done aerobically in light or in darkness
Table 3 Acetylene reduction, H2 evolution and H2 uptake [nmol (lg Chl a))1 h)1] in Lyngbya majuscula CCAP 1446/4 grown underdifferent conditions. n.d. Not determined
Growth conditions C2H2 reduction(light, argon)
H2 evolution H2 uptake
Lightb Darkc Lightb Darkc
Constant light 11.5 1 1.4 20.5 42.4Light/Dark, 12/12 h from light phasea 5.1 0.9 5.3 16.9 24.5Light/Dark, 12/12 h from dark phasea 6.1 n.d. 4.8 17.5 33.7
aCells for measurements were taken 4 h into the light or dark phase during growth under alternating light/dark (12/12 h) cyclebMeasurements performed in lightcMeasurements performed in darkness
Table 4 Methyl viologen-dependent H2-evolution of Lyngbya majuscula CCAP 1446/4,Anabaena variabilis ATCC 29413, Anabaena sp.PCC 7120, andSynechocystis sp. PCC 6803. WT Wild type
Strain Growth conditions MV-dependent H2
evolution[nmol H2
(lg Chl a))1 h)1]
Lyngbya majuscula CCAP 1446/4 BG110, constant light, bubbling with air 61.4Anabaena variabilis ATCC 29413 (WT) BG11, constant light, bubbling with air and 1% CO2 42.3Anabaena sp. PCC 7120 (WT) BG11, constant light, bubbling with air and 1% CO2 39.7Synechocystis sp. PCC 6803 BG11 (1 mM NaNO3), constant light, bubbling with air and 1% CO2 235.5
355
(Table 5). No nitrogenase-mediated H2 photoevolutionwas observed, most likely due to the consumption by anactive uptake hydrogenase (Table 5). A residual level[1.01 nmol H2 (mg protein))1 min)1] of MV-dependentH2 evolution (bidirectional hydrogenase activity) wasdetected in the culture, in contrast to the lack of ahybridization signal when probes for hox genes wereused (Table 1).
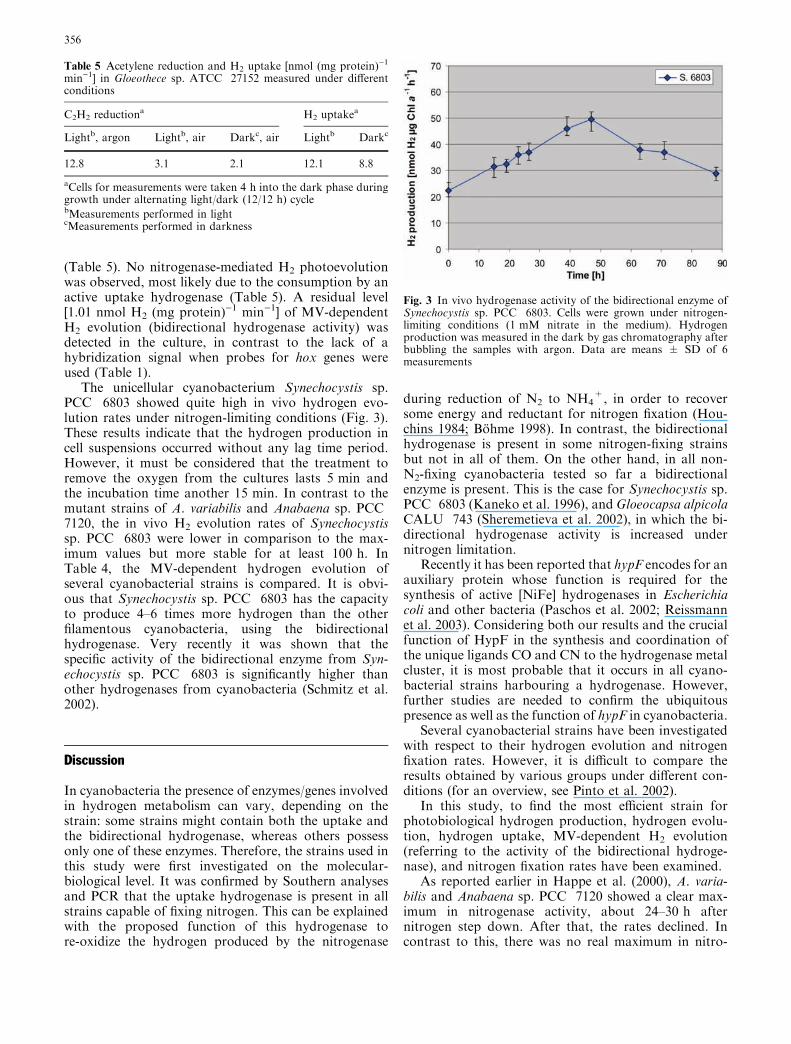
The unicellular cyanobacterium Synechocystis sp.PCC 6803 showed quite high in vivo hydrogen evo-lution rates under nitrogen-limiting conditions (Fig. 3).These results indicate that the hydrogen production incell suspensions occurred without any lag time period.However, it must be considered that the treatment toremove the oxygen from the cultures lasts 5 min andthe incubation time another 15 min. In contrast to themutant strains of A. variabilis and Anabaena sp. PCC7120, the in vivo H2 evolution rates of Synechocystissp. PCC 6803 were lower in comparison to the max-imum values but more stable for at least 100 h. InTable 4, the MV-dependent hydrogen evolution ofseveral cyanobacterial strains is compared. It is obvi-ous that Synechocystis sp. PCC 6803 has the capacityto produce 4–6 times more hydrogen than the otherfilamentous cyanobacteria, using the bidirectionalhydrogenase. Very recently it was shown that thespecific activity of the bidirectional enzyme from Syn-echocystis sp. PCC 6803 is significantly higher thanother hydrogenases from cyanobacteria (Schmitz et al.2002).
Discussion
In cyanobacteria the presence of enzymes/genes involvedin hydrogen metabolism can vary, depending on thestrain: some strains might contain both the uptake andthe bidirectional hydrogenase, whereas others possessonly one of these enzymes. Therefore, the strains used inthis study were first investigated on the molecular-biological level. It was confirmed by Southern analysesand PCR that the uptake hydrogenase is present in allstrains capable of fixing nitrogen. This can be explainedwith the proposed function of this hydrogenase tore-oxidize the hydrogen produced by the nitrogenase
during reduction of N2 to NH4+, in order to recover
some energy and reductant for nitrogen fixation (Hou-chins 1984; Bohme 1998). In contrast, the bidirectionalhydrogenase is present in some nitrogen-fixing strainsbut not in all of them. On the other hand, in all non-N2-fixing cyanobacteria tested so far a bidirectionalenzyme is present. This is the case for Synechocystis sp.PCC 6803 (Kaneko et al. 1996), and Gloeocapsa alpicolaCALU 743 (Sheremetieva et al. 2002), in which the bi-directional hydrogenase activity is increased undernitrogen limitation.
Recently it has been reported that hypF encodes for anauxiliary protein whose function is required for thesynthesis of active [NiFe] hydrogenases in Escherichiacoli and other bacteria (Paschos et al. 2002; Reissmannet al. 2003). Considering both our results and the crucialfunction of HypF in the synthesis and coordination ofthe unique ligands CO and CN to the hydrogenase metalcluster, it is most probable that it occurs in all cyano-bacterial strains harbouring a hydrogenase. However,further studies are needed to confirm the ubiquitouspresence as well as the function of hypF in cyanobacteria.
Several cyanobacterial strains have been investigatedwith respect to their hydrogen evolution and nitrogenfixation rates. However, it is difficult to compare theresults obtained by various groups under different con-ditions (for an overview, see Pinto et al. 2002).
In this study, to find the most efficient strain forphotobiological hydrogen production, hydrogen evolu-tion, hydrogen uptake, MV-dependent H2 evolution(referring to the activity of the bidirectional hydroge-nase), and nitrogen fixation rates have been examined.
As reported earlier in Happe et al. (2000), A. varia-bilis and Anabaena sp. PCC 7120 showed a clear max-imum in nitrogenase activity, about 24–30 h afternitrogen step down. After that, the rates declined. Incontrast to this, there was no real maximum in nitro-
Table 5 Acetylene reduction and H2 uptake [nmol (mg protein))1
min)1] in Gloeothece sp. ATCC 27152 measured under differentconditions
C2H2 reductiona H2 uptake
a
Lightb, argon Lightb, air Darkc, air Lightb Darkc
12.8 3.1 2.1 12.1 8.8
aCells for measurements were taken 4 h into the dark phase duringgrowth under alternating light/dark (12/12 h) cyclebMeasurements performed in lightcMeasurements performed in darkness
Fig. 3 In vivo hydrogenase activity of the bidirectional enzyme ofSynechocystis sp. PCC 6803. Cells were grown under nitrogen-limiting conditions (1 mM nitrate in the medium). Hydrogenproduction was measured in the dark by gas chromatography afterbubbling the samples with argon. Data are means ± SD of 6measurements
356

genase activity of the other studied filamentous hetero-cystous cyanobacteria. This might be due to the partic-ular growth phase of the cultures, because after about30 h the A. variabilis ATCC 29413 and Anabaena sp.PCC 7120 strains were in the beginning of the expo-nential growth phase whereas the other strains grewmuch slower. Nevertheless, all cultures were incubatedunder saturating light conditions, so that the lowernitrogenase activities are not the result of light limita-tion. The hydrogen production correlates with thenitrogenase activity, but the rates of H2 production inthe wild types are quite low because of the activity of theuptake hydrogenase. This explains the importance ofhup) mutants, which have been studied in more detail. Itcould be shown that due to the deletion of a part of thehupSL genes or the xisC gene the mutants produced(and evolved) molecular hydrogen when grown in air(Carrasco et al. 1998; Happe et al. 2000; Lindberg et al.2002; Lindblad et al. 2002). However, with the con-struction of a hup)/hox) double mutant in Anabaena sp.PCC 7120 the hydrogen evolution could not be furtherincreased. Interestingly, the hoxH) single mutantexhibited even a lower production of hydrogen than thewild type, whereas the values for the hupL) single andthe hupL)/hoxH) double mutant were nearly identicaland several times higher than in the other strains(Masukawa et al. 2002). By investigating the durabilityof H2 production of Anabaena sp. PCC 7120 wild typeand the uptake-hydrogenase-deficient mutant in a 1-lphoto-bioreactor it was shown that hydrogen evolutionunder argon or air atmosphere by the mutant is possiblefor more than 10 days without adding new medium ornutrients (Lindblad et al. 2002).
Unlike the heterocystous strains, the investigated fil-amentous non-heterocystous and unicellular N2-fixingstrains showed low or only traces of H2 produced bynitrogenase due to low nitrogenase activities as well tothe presence of active hydrogenases. Only Leptolyngbyasp. PCC 73110 evolved some H2 in light under anaer-obic conditions. However, the rate of H2 evolution waslow in this case compared with that observed in theheterocystous strains, and it could be obtained only inanaerobic nitrogen-limited cultures. In addition, L. ma-juscula, which is able to fix N2 aerobically in light and toprotect nitrogenase from O2 inactivation, does notevolve H2 due to efficient recycling by active hydrogen-ases. However, this strain could be used for H2 pro-duction during fermentation due to the presence of aconstitutively expressed bidirectional hydrogenase.
Another promising approach for biological hydrogenproduction is the use of a unicellular cyanobacteriumlike Synechocystis sp. PCC 6803. The rates for H2
evolution under nitrogen-limiting conditions in the darkare quite high, and they are comparable to those mea-sured for heterocystous cyanobacteria and green algae(Melis and Happe 2001). The production rates are about10 times higher when an artificial electron donor likereduced MV is added, which shows the potential of the
Synechocystis system. Gloeocapsa alpicola CALU 743,another unicellular non-nitrogen-fixing cyanobacterium,performs fermentation of endogenously stored glycogenunder dark anoxic conditions, with formation ofhydrogen, acetate and CO2 as the main products. Therates for this process are comparable to those producedby the nitrogenase-based system of heterocystous cy-anobacteria and the hydrogenase-based system of greenalgae, indicating a unique possibility for hydrogen pro-duction by cyanobacteria such as G. alpicola (Troshinaet al. 2002).
In conclusion, for effective large-scale hydrogen pro-duction several requirements should be fulfilled, such ashigh H2 evolution rates and stability. In the case of N2-fixing cyanobacteria, hydrogen-uptake-deficient mutantshave to be used because of their inability to re-oxidizethe hydrogen produced by the nitrogenase. On the otherhand, fermentative H2 production by the bidirectionalhydrogenase emerges as a valid alternative deservingfurther investigation.
Acknowledgements The work of the Portuguese team was finan-cially supported by FCT and FSE (III Quadro Comunitario deApoio), and PRAXIS/P/BIA/13238/98. Financial support from theEuropean Science Foundation Program on Cyanobacterial Nitro-gen Fixation (CYANOFIX), COST Action 841, the Swedish En-ergy Agency, and the Swedish Research Council is gratefullyacknowledged. The cyanobacterial strains Gloeothece sp. ATCC27152 and Lyngbya majuscula CCAP 1446/4 were kindly providedby Professors J.R. Gallon and P.C. Wright, respectively. We thankDr. Helena Carvalho for assistance with the optical microscopywork, and Fredrik Oxelfelt for the Gloeothece DNA-extractionmethod.
References
Allen MM (1968) Simple conditions for growth of unicellular blue–green algae on plates. J Phycol 4:1–4
Aoyama K, Uemura I, Miyake J, Asada Y (1997) Fermentativemetabolism to produce hydrogen gas and organic compoundsin a cyanobacterium, Spirulina platensis. J Ferment Bioeng83:17–20
Appel J, Schulz R (1998) Hydrogen metabolism in organisms withoxygenic photosynthesis: hydrogenases as important regulatorydevices for a proper redox poising? J Photochem Photobiol B:Biol 47:1–11
Appel J, Phunpruch S, Steinmuller K, Schulz R (2000) The bidi-rectional hydrogenase of Synechocystis sp. PCC 6803 works asan electron valve during photosynthesis. Arch Microbiol173:333–338
Bergman B, Gallon JR, Rai AN, Stal LJ (1997) N2 fixation by non-heterocystous cyanobacteria. FEMS Microbiol Rev 19:139–185
Bishop PE, Premakumar R (1992) Alternative nitrogen fixationsystems. In: Stacey G, Burris RH, Evans HJ (eds) Biologicalnitrogen fixation. Chapman and Hall, New York, pp 736–762
Bohme H (1998) Regulation of nitrogen fixation in heterocyst-forming cyanobacteria. Trends Plant Sci 3:346–351
Boison G, Schmitz O, Mikheeva L, Shestakov S, Bothe H (1996)Cloning, molecular analysis and insertional mutagenesis of thebidirectional hydrogenase genes from the cyanobacteriumAnacystis nidulans. FEBS Lett 394:153–158
Borodin VB, Tsygankov AA, Rao KK, Hall DO (2000) Hydrogenproduction by Anabaena variabilis PK84 under simulated out-door conditions. Biotechnol Bioeng 69:478–485
357

Carrasco CD, Buettner JA, Golden JW (1995) Programmed DNArearrangement of a cyanobacterial hupL gene in heterocystous.Proc Natl Acad Sci USA 92:791–795
Carrasco CD, Garcia JS, Golden JW (1998) Programed DNArearrangement of a hydrogenase gene during Anabaena het-erocyst development. In: Zaborsky OR, Benemann JR,Matsunaga T, Miyake J, San Pietro A (eds) BioHydrogen.Plenum Press, New York, pp 203–207
Castenholz RW (1998) Culturing methods for cyanobacteria.Methods Enzymol 167:68–93
Fedorov AS, Tsygankov AA, Rao KK, Hall DO (2001) Productionof hydrogen by an Anabaena variabilis mutant in a photobior-eactor under aerobic outdoor conditions. In: Miyake J,Matsunaga T, San Pietro A (eds) Biohydrogen II. Elsevier,Oxford pp 223–228
Fiore MF, Moon DH, Tsai SM, Lee H, Trevors JT (2000) Mini-prep DNA isolation from unicellular and filamentous cyano-bacteria. J Microbiol Methods 39:159–169
Gallon JR, Perry SM, Rajab TMA, Flayesh KAM, Chaplin AE(1988) Metabolic changes associated with the diurnalpattern of N2 fixation in Gloeothece. J Gen Microbiol134:3079–3087
Hansel A, Lindblad P (1998) Towards optimization of cyanobac-teria as biotechnologically relevant producers of molecularhydrogen, a clean and renewable energy source. Appl MicrobiolBiotechnol 50:153–160
Hansel A, Axelsson R, Lindberg P, Troshina OY, Wunschiers R,Lindblad P (2001) Cloning and characterization of a hyp genecluster in the filamentous cyanobacterium Nostoc sp. strainPCC 73102. FEMS Microbiol Lett 201:59–64
Haselkorn R, Buikema WJ (1992) Nitrogen fixation in cyanobac-teria, In: Stacey G, Burris RH, Evans HJ (eds) Biologicalnitrogen fixation. Chapman & Hall, London, pp 166–190
Happe T, Naber JD (1993) Isolation, characterization and aminoacid sequence of hydrogenase from the green algae Chlamydo-monas reinhardtii. Eur J Biochem 214:475–478
Happe T, Schutz K, Bohme H (2000) Transcriptional and muta-tional analysis of the uptake hydrogenase of the filamentouscyanobacterium Anabaena variabilis ATCC 29413. J Bacteriol182:1624–1631
Houchins JP (1984) The physiology and biochemistry of hydrogenmetabolism in cyanobacteria. Biochim Biophys Acta 768:227–255
Kaneko T, Sato S, Kotani H, Tanaka A, Asamizu E, Nakamura Y,Miyajima N, Hirosawa, M, Sugiura M, Sasamoto S, Kimura T,Hosouchi T, Matsuno A, Muraki A, Nakazaki N, Naruo K,Okumura S, Shimpo S, Takeuchi C, Wada T, Watanabe A,Yamada M, Yasuda M, Tabata S (1996) Sequence analysis ofthe genome of the unicellular cyanobacterium Synechocystis sp.strain PCC 6803. II. Sequence determination of the entiregenome and assignment of potential protein-coding regions.DNA Res 30:185–209
Kaneko T, Nakamura Y, Wolk CP, Kuritz T, Sasamoto S,Watanabe A, Iriguchi M, Ishikawa A, Kawashima K, KimuraT, Kishida Y, Kohara M, Matsumoto M, Matsuno A, MurakiA, Nakazaki N, Shimpo S, Sugimoto M, Takazawa M,Yamada M, Yasuda M, Tabata S (2001) Complete genomicsequence of the filamentous nitrogen-fixing cyanobacteriumAnabaena sp. strain PCC 7120. DNA Res 8:205–213
Kentemich T, Danneberg G, Hundeshagen B, Bothe H (1988)Evidence for the occurrence of the alternative, vanadium-con-taining nitrogenase in the cyanobacterium Anabaena variabilis.FEMS Microbiol Lett 51:19–24
Kentemich T, Haverkamp G, Bothe H (1991) The expression of athird nitrogenase in the cyanobacterium Anabaena variabilis. ZNaturforsch Teil C 46:217–222
Lindberg P, Schutz K, Happe T, Lindblad P (2002) A hydrogen-producing, hydrogenase-free mutant strain of Nostocpunctiforme ATCC 29133. Int J Hydrogen Energy 27:1291–1296
Lindblad P, Christensson K, Lindberg P, Fedorov A, Pinto F,Tsygankov A (2002) Photoproduction of H2 by wildtype An-abaena PCC 7120 and a hydrogen uptake deficient mutant:from laboratory experiments to outdoor culture. Int J Hydro-gen Energy 27:1271–1281
Masukawa H, Mochimaru M, Sakurai H (2002) Disruption of theuptake hydrogenase gene, but not the bidirectional hydrogenasegene, leads to enhanced photobiological hydrogen productionby the nitrogen-fixing cyanobacterium Anabaena sp. PCC7120. Appl Microbiol Biotechnol 58:618–624
Mackinney G (1941) Absorption of light by chlorophyll solutions.J Biol Chem 140:315–322
Meeks JC, Elhai J, Thiel T, Potts M, Larimer F, Lamerdin J,Predki P, Atlas R (2001) An overview of the genome of Nostocpunctiforme, a multicellular, symbiotic cyanobacterium. Pho-tosynth Res 70:85–106
Melis A, Happe T (2001) Hydrogen production. Green algae as asource of energy. Plant Physiol 127:740–748
Nakamura Y, Kaneko T, Hirosawa, M, Miyajima N, Tabata S(1998) CyanoBase, a www database containing the completegenome of Synechocystis sp. strain PCC 6803. Nucleic AcidsRes 20:63–67
Oxelfelt F, Tamagnini P, Salema R, Lindblad P (1995) Hydrogenuptake in Nostoc strain PCC 73102: effects of nickel, hydrogen,carbon and nitrogen. Plant Physiol Biochem 33:617–623
Oxelfelt F, Tamagnini P, Lindblad P (1998) Hydrogen uptake inNostoc sp. strain PCC 73102. Cloning and characterization ofa hupSL homologue. Arch Microbiol 169:267–274
Oxelfelt F, Oliveira P, Leitao E, Barros M, Cardoso P, TamagniniP (2002) Uptake hydrogenases in the unicellular cyanobacteriaGloeothece ATCC 27152 and Cyanothece ATCC 51142. 5thEuropean workshop on the molecular biology of cyanobacteria.Stockholm, Sweden
Paschos A, Bauer A, Zimmermann A, Zehelein E, Bock A (2002)HypF, a carbamoyl phosphate-converting enzyme involved in[NiFe] hydrogenase maturation. J Biol Chem 277:49945–49951
Peterson GL (1983) Determination of total protein. MethodsEnzymol 91:95–119
Pinto F, Troshina O, Lindblad P (2002) A brief look at threedecades of research on cyanobacterial hydrogen evolution. Int JHydrogen Energy 27:1209–1215
Rao KK, Hall DO (1996) Hydrogen production by cyanobacteria:potential, problems and prospects. J Mar Biotechnol 4:10–15
Reade JPH, Dougherty LJ, Rogers LJ, Gallon JR (1999) Synthesisand proteolytic degradation of nitrogenase in cultures of theunicellular cyanobacterium Gloeothece strain ATCC 27152.Microbiology 145:1749–1758
Reissmann S, Hochleitner E, Wang H, Paschos A, Lottspeich F,Glass RS, Bock A (2003) Taming of a poison: biosynthesis ofthe NiFe-hydrogenase cyanide ligands. Science 299:1067–1070
Sambrook J, Fritsch EF, Maniatis T (1989) Molecular cloning: alaboratory manual, 2nd edn. Cold Spring Harbor LaboratoryPress, New York, USA
Schmitz O, Bothe H (1996) The diaphorase subunit HoxU of thebidirectional hydrogenase as electron transferring protein incyanobacterial respiration? Naturwissenschaften 83:525–527
Schmitz O, Boison G, Hilscher R, Hudeshagen B, Zimmer W,Lottspeich F, Bothe H (1995) Molecular biological analysis of abi-directional hydrogenase from cyanobacteria. Eur J Biochem233:266–276
Schmitz O, Boison G, Salzmann H, Bothe H, Schutz K, Wang SH,Happe T (2002) HoxE — a subunit specific for the pentamericbidirectional hydrogenase complex (HoxEFUYH) of cyano-bacteria. Biochim Biophys Acta 1554:66–74
Schulz R (1996) Hydrogenases and hydrogen production ineukaryotic organisms and cyanobacteria. J Mar Biotechnol4:16–22
Serebriakova L, Zorin NA, Lindblad P (1994) Reversible hydrog-enase in Anabaena variabilis ATCC 29413: Presence andlocalization in non-N2-fixing cells. Arch Microbiol 161:140–144
358

Sheremetieva ME, Troshina OY, Serebryakova LT, Lindblad P(2002) Identification of hox genes and analysis of their tran-scription in the unicellular cyanobacterium Gloeocapsa alpicolaCALU 743 growing under nitrate-limiting conditions. FEMSMicrobiol Lett 214:229–33
Stal LJ, Moezelaar R (1997) Fermentation in cyanobacteria.FEMS Microbiol Rev 21:170–211
Tamagnini P, Troshina O, Oxelfelt F, Salema R, Lindblad P (1997)Hydrogenases in Nostoc sp. strain PCC 73102, a strain lackinga bidirectional enzyme. Appl Environ Microbiol 63:1801–1807
Tamagnini P, Costa J-L, Almeida L, Oliveira M-J, Salema R,Lindblad P (2000) Diversity of cyanobacterial hydrogenases, amolecular approach. Curr Microbiol 40:356–361
Tamagnini P, Axelsson R, Lindberg P, Oxelfelt F, Wunschiers R,Lindblad P (2002) Hydrogenases and hydrogen metabolism ofcyanobacteria. Microbiol Mol Biol Rev 66:1–20
Thiel T (1993) Characterization of genes for an alternative nitro-genase in the cyanobacterium Anabaena variabilis. J Bacteriol175:6276–6286
Thiel T, Lyons EM, Erker JC, Ernst A (1995) A second nitrogenasein vegetative cells of a heterocyst-forming cyanobacterium. ProcNatl Acad Sci USA 92:9358–9362
Thiel T, Lyons EM, Erker JC (1997) Characterization of genes fora second Mo-dependent nitrogenase in the cyanobacteriumAnabaena variabilis. J Bacteriol 179:5222–5225
Troshina O, Serebryakova L, Sheremetieva M, Lindblad P (2002)Production of H2 by the unicellular cyanobacterium Gloeocapsaalpicola CALU 743 during fermentation. Int J Hydrogen En-ergy 27:1283–1289
Tsygankov AA, Serebryakova LT, Sveshnikov DA, Rao KK,Gogotov IN, Hall DO (1998) Hydrogen photoproduction bythree different nitrogenases in whole cells of Anabaena vari-abilis and the dependence on pH. Int J Hydrogen Energy22:859–867
Tsygankov AA, Borodin VB, Rao KK, Hall DO (1999) H2 Pho-toproduction by batch culture of Anabaena variabilis ATCC29143 and its mutant PK84 in a photobioreactor. BiotechnolBioeng 64:709–715
359