conservation of the sinai hairstreak butterfly
TRANSCRIPT

CONSERVATION OF THE SINAI HAIRSTREAK BUTTERFLY
ANDREW POWER B.A. (HONS)
Thesis submitted to the University of Nottingham for the
degree of Master of Research
January 2013

i
Abstract
The Sinai Hairstreak Satyrium jebelia (Nakamura 1975) is one of only two butterflies listed as
endemic to Egypt (and since the recent unpublished discovery of a Saudi population of the
Sinai Baton Blue, in fact may be the only endemic). It occurs only in the mountains of the St
Katherine Protectorate in the South Sinai, and has remained unstudied since its discovery in
1974. It is likely to be Critically Endangered and at considerable risk of extinction. For the first
time, the distribution of both the Sinai Hairstreak and its larval food plant, Rhamnus
dispermus, are mapped. This study also provides the first population estimate of the Sinai
Hairstreak, using Mark-Release-Recapture methods, as well as information on the behaviour
and conservation requirements of this rare endemic butterfly.
The total world population of Sinai Hairstreaks is estimated to be 1,010 individuals; this
population is divided into six smaller sub-populations of varying size. Recapture data
revealed that Sinai Hairstreaks have high powers of dispersal. This coupled with the relatively
close proximity of the sub-populations may indicate the existence of a single panmictic
population. Due to the small size of the population and the limited amount of suitable
habitat available, the Sinai Hairstreak is at considerable risk of extinction and according to
IUCN criteria should be classed as Critically Endangered.
Tree size, the percentage of healthy green leaves, plant diversity and the presence of fruit on
larval food plant were all found to be significant in explaining the presence of Sinai
Hairstreaks on Rhamnus dispermus. All of these factors can be considered to be a function of
habitat quality. No immediate threats to the Sinai Hairstreak are evident. The larval food
plant grows in remote locations that are at low risk from human disturbance. The local
people do not harvest or use the larval food plant in any way and the effect of grazing is
likely to be minimal. The most significant threat facing the Sinai Hairstreak is global warming.
If climate change predictions for Egypt are correct, the quality of habitat and plant diversity
will decrease in the St Katherine Protectorate as the temperatures increases.
The information from this study provides a framework for future work and a foundation for
the establishment of a long-term conservation strategy for the Sinai Hairstreak.
Annotated lists of dragonfly and bird species recorded during the study period in the St
Katherine Protectorate are included in the appendices. The dragonfly species the Southern
Skimmer Orthetrum brunneum was recorded for only the third time in the Sinai.

ii
Acknowledgements
Firstly I would like to thank my supervisor Francis Gilbert, whose passion for the Sinai is the
reason I chose to come to Nottingham. I would also like to thank the Sinai fieldwork team –
Luke McLeod, Olivia Norfolk, Katy Thompson & Lisa Gecchele who provided me with advice
and plenty of laughs throughout my time in Egypt and Nottingham. I thank the EEAA and the
Protectorate management for permission to work in the park, Farag and Fox Camp for
providing an excellent base camp and Prof. Samy Zalat for logistical help.
I am particularly grateful to have had Suleiman Abusada as my guide in Egypt, like brothers
we squabbled and joked throughout our many days and nights in the mountains. No matter
where or when I wanted to work, or for how long, Suleiman said yes. I am also indebted to
Mohammed Salah, Hamdee Mubarak, Salim and Yusuf for showing me Bedouin culture and
making me feel at home. I am honoured to have met Nasr Mansour, whose knowledge of the
flora and fauna of St Katherine and his desire to show them to me contributed significantly
to my research.
Féaron Cassidy kept me sane for yet another thesis, and visited me in Sinai. My brother Brian
and my friend Peter also visited for a week, unwittingly becoming my field assistants, helping
me measure 200 trees in two days. I would also like to thank Seán Kelly for all his advice.
Finally I would like to thank my family for their support throughout the year.

iii
Contents Abstract ....................................................................................................................................... i
Acknowledgements .....................................................................................................................ii
1 Introduction ............................................................................................................................ 1
2 Materials and Methods ........................................................................................................... 8
Study area .............................................................................................................................. 8
Distribution and population estimates .................................................................................. 8
Larval food plant features .................................................................................................... 13
Behaviour ............................................................................................................................. 14
Statistical analysis ................................................................................................................ 14
3 Results ................................................................................................................................... 15
Local knowledge ................................................................................................................... 15
Rhamnus dispermus ............................................................................................................. 15
Distribution ...................................................................................................................... 15
Phenology of Rhamnus dispermus ................................................................................... 19
Sinai Hairstreak .................................................................................................................... 22
Distribution ...................................................................................................................... 22
Butterfly movements ....................................................................................................... 22
Population estimates ....................................................................................................... 30
Activity times .................................................................................................................... 31
Behaviour and ecology ..................................................................................................... 33
Predators and parasites ................................................................................................... 33
Statistical analysis ............................................................................................................ 34
4 Discussion .............................................................................................................................. 34
Rhamnus dispermus ............................................................................................................. 34
Sinai Hairstreak .................................................................................................................... 35
Habitat requirements ........................................................................................................... 35
Dispersal ability .................................................................................................................... 36
Conservation ........................................................................................................................ 39
References ............................................................................................................................... 41
Appendix I - Odonata of St Katherine ...................................................................................... 45
Appendix II - Birds of St Katherine ........................................................................................... 47

1
1 Introduction
One of the most important necessities for the conservation of a rare endemic species is
information on their distribution (De Groot et al., 2009). However, this is often unavailable in
countries where there is less emphasis on conservation and in the case of recently
discovered species. An analysis of the ecological characteristics of threatened and non-
threatened butterflies revealed that threatened butterflies are characterised by narrow
niche breadth, restricted resource distribution, poor dispersal ability and short flight period
(Kotiaho et al., 2005). The two major threats facing endemic species are climate change and
habitat destruction.
The effect of climate change is often exacerbated in mountain regions. Global
warming tends to push species poleward or to higher altitudes where the climate is cooler
(Wilson et al., 2007). An analysis of butterfly surveys dating from 1992 to 2010 in North
America showed a sharp increase in sub-tropical species that have expanded northward
from warmer climates, indicating how butterfly ranges can change in response to global
warming but also how quickly climate change is affecting species distribution (Breed et al.,
2012). Species pushed to higher altitudes can become isolated in high-altitude ‘islands’.
Migration out of these areas is difficult as there is no suitable habitat nearby and many
mountain endemics have low powers of dispersal (Thomas et al., 2004). As global warming
increases, species are pushed higher, restricting further their available habitat. Mountain
endemics face a heightened risk of extinction because of these factors (Thomas et al., 2004).
Habitat destruction is a significant threat to butterfly species, especially to species
with limited ranges. The Xerces Blue Glaucopsyche xerces was an American butterfly species
found on coastal sand dunes that became extinct as a result of habitat destruction caused by
urban development (Emmel and Emmel, 1993). The amount of available habitat is significant
for explaining the occurrence of a species. Habitat destruction changes the spatial structure
and the metapopulation dynamics of a species, and understanding the dynamics is crucial for
a conservation strategy to be successful. Habitat destruction decreases habitat area as well
as the connectivity between suitable habitats, making it more difficult to move between sites
and colonise suitable areas of habitat. This is frequently observed in butterflies because their
distribution is dependent on the presence of their larval food plant or a specific habitat type;
for example the Marsh Fritillary Euphdryas aurinia can live in a variety of habitats but only
where its larval food plant, the Devil’s-bit scabious Succisa pratensis, is found (Wahlberg et
al., 2002). The Marsh Fritillary therefore occurs in isolated patches which contain suitable
larval food plants. The species has undergone a severe decline in Britain and Ireland in the
last century as habitat destruction and human activities have altered the landscape, isolating
breeding habitats and hence increasing the likelihood of local extinction (Schtickzelle et al.,
2005).
The connectivity of suitable habitat is vital for the long-term survival of many species
because it determines the extent to which metapopulation dynamics apply (Joyce and Pullin,
2003). The mere existence of fragmented habitat is a necessary but not a sufficient condition

2
to establish that a species occurs in a metapopulation, a term now applied much more
broadly than it should be (Fronhofer et al., 2012). The extent to which habitats are patchy to
a butterfly depends on the typical distances it disperses, and there is a trade-off between
dispersal distance and population growth rate: poor dispersers have greater potential
population growth rate, but suffer more from stochasticity of their local environments
(Baguette and Schtickzelle, 2006). This may be especially important in arid environments,
which are patchy and fragmented due to the limited and patchy natural resources available,
especially water.
The population structure of a species is dependent on its distribution, the availability
of suitable habitat and dispersal ability. Some migratory butterflies have incredible powers of
dispersal and can travel large distances; Eastern North American Monarch butterflies Danaus
plexippus can travel distances of up to 4000 km on migration (Zhu et al., 2009). More typical
non-migratory butterflies cannot travel great distances, such as the Chalk-hill Blue
Polyommatus coridon, which can travel a maximum distance of 3.7 km but typically moves
far less (Schmitt et al., 2006). Some butterflies have very poor powers of dispersal, such as
the Sinai Baton Blue Pseudophilotes sinaicus which rarely moves further than 100 m (James,
2006a, James, 2006c, Gilbert et al., 2010). Butterflies are model organisms for the study of
population structure, impact of habitat loss and fragmentation as well as climate change
(Butterfly Conservation Europe, 2008).
The spatial structure of a species can be highly variable and a variety of models exist.
These models are based on dispersal ability and connectivity of suitable habitat. Some
species are separated into isolated populations that do not interact at all as dispersal is
limited. These populations may be at increased risk of extinction as they are unlikely to be
recolonised (Frankham, 2005). Isolated populations may differentiate genetically over time
due to insufficient gene flow from neighbouring populations (Bouzat et al., 2009). Isolation
can have detrimental effects on a population as genetic diversity decreases with inbreeding
(Bouzat et al., 2009). A study of the Golden-brown mouse lemur Microcebus ravelobensis
showed that there was significant genetic differentiation between populations (Guschanski
et al., 2007). These populations have become isolated over time from a combination of
natural barriers, habitat fragmentation and destruction as a result of human intervention
(Guschanski et al., 2007). A species can also persist as a metapopulation where populations
are connected by limited dispersal. A true metapopulation contains discrete habitat patches
that are incapable of containing self-supporting populations without constant exchange of
individuals through migration; the populations of these patches fluctuate randomly between
extinction and recolonisation events (Wheat et al., 2011). A source-sink model describes how
individuals may move from areas of high population density (source), where habitat
conditions are favourable, to areas of low population density (sink) where conditions may be
less favourable. The sink would not be able to support a population on its own and for
persistence relies on excess individuals from the source (Boughton, 2000). This is similar to
the mainland-island model which describes how one large population may act as source to
smaller populations. However, these smaller populations can persist on their own over short
periods of time, but are at a greater risk of extinction because of their small size (Fronhofer
et al., 2012). In some cases multiple patches of suitable habitat may be closely linked
through very high powers of dispersal. This is known as a patchy population, and local

3
extinction does not occur as result of the high powers of dispersal between the habitat
patches. In this scenario a species exists as a single panmictic population (Fronhofer et al.,
2012). In these populations no genetic structure can be found as populations do not become
isolated. A genetic study of Moose Alces alces in Canada showed no genetic structure,
suggesting that they exist as a single panmictic population (Finnegan et al., 2012). It is very
important to assign the correct population structure to a species when creating a
conservation strategy (Guiney et al., 2010). Basing a conservation strategy on the wrong
population structure may waste valuable resources and may not benefit a species (Fronhofer
et al., 2012).
This study concerns the Sinai Hairstreak Satyrium jebelia (Nakamura, 1975), now
Egypt’s only known endemic butterfly species after the recent discovery of the Sinai Baton
Blue in Saudi Arabia (Dr Samy Zalat, pers. comm.). It is known to occur in three small sites in
the mountains (2000 m altitude) around the town of St Katherine in the St Katherine
Protectorate (South Sinai). It was discovered in 1974 and all the holotypes and paratypes
collected from just two isolated sites; Jebel Bab and Jebel Ahmar (Nakamura, 1975). The
species was named after the Jebelia, the Bedouin tribe that live in the area (Nakamura,
1975). Another specimen was observed and collected at a new, third site in 1993 at Jebel
Katherine (Weidenhoffer et al., 2004). These represent the only three localities where this
species is known to be found, all lying within a ten-kilometre radius of the city of St
Katherine. It is highlighted as a species urgently needing reliable assessment and although
likely to be Critically Endangered, there are few data to back this up. As of 2007 there have
been fourteen known records of the Sinai Hairstreak with the latest being from 2001 (Gilbert
and Zalat, 2007).
The genus Satyrium belongs to the family Lycaenidae, the second largest butterfly
family, which also includes the Sinai Baton Blue (Nakamura 1975). The classification of
Holarctic hairstreaks has long been a contentious issue (Clench, 1978) and it remains so. The
nomenclatural history of the genus Satyrium (Scudder 1876) is complex with repeated
alterations. The Sinai Hairstreak currently belongs to the subgenus Superflua (Strand, 1910),
characterised by a strongly marked underside compared with the rest of the genus and the
female usually having well pronounced submarginal spots on the underside (Weidenhoffer
et al., 2004). However, the Sinai Hairstreak is probably most closely related to the Blue Spot
Hairstreak Satyrium spini (Schiffermüller) of the subgenus Satyrium (Nakamura, 1975), found
throughout western Europe as far east as Iran (Tshikolovets, 2011). There have not been any
recent studies on the phylogeny of Satyrium and the attribution of the Sinai Hairstreak to the
subgenus Superflua is not well supported (Gian Christophoro Bozano, pers. comm.). The Blue
Spot Hairstreak has a similar wing pattern and egg structure, as well as being relatively close
geographically. In the mountains of the West Bank (Palestine), it uses the same larval food
plant as the Sinai Hairstreak (Nakamura, 1975). As well as differences in wing morphology,
the size of the male genitalia of the Sinai Hairstreak is different from that of the Blue Spot
Hairstreak. The most striking characteristic of this distinct species, that it does not share with
any other members of the genus, is the continuous, smoothly curved postmedian band on
the underside of the hind wing (Nakamura, 1975) (Plate 1.1).

4
The genus Satyrium is found across the Holarctic, while the Sinai Hairstreak is
distributed in a small area of the Sinai on the southern edge of this zone (Weidenhoffer et
al., 2004). South Sinai is an arid landscape with little rainfall, not characteristic of the habitat
used by other species of the genus. The Blue Spot Hairstreak is found at both low and high
altitude in areas of high plant diversity such as scrubland, grassland and woodland edges and
clearings (Settle et al., 2010). The Sinai and Middle East region once had considerably more
rainfall and more extensive arboreal vegetation during the glacial periods of the Quaternary
(Issar et al., 2011). This habitat would have extended north through Israel into Europe,
allowing faunal exchange between the two continents, so the Sinai Hairstreak is likely to
have had a larger range before the desiccation processes that occurred after the Atlantic
period 5,000 years ago (Nakamura, 1975). Many other relict species from cooler climates can
be found in the Sinai such as Phoenicean Juniper, or Arar, Juniperus phoenicea which is
normally found in much wetter areas of the Mediterranean basin (El-Bana et al., 2010).
The Sinai Hairstreak is on the wing between late May and early July and is said to be
not uncommon (Nakamura, 1975) in areas where its larval food plant occurs, usually in wadis
(valleys) and along rocky slopes. This species is univoltine and lays its eggs at the base of the
thorns, always on the underside of the twig, or in positions facing away from the sun
(Nakamura, 1975). Like the Blue Spot Hairstreak, the eggs are said to go through a diapause
with a delayed emergence until the following spring. The Blue Spot Hairstreak caterpillars
hatch in spring and feed on emerging flower buds and young leaves of the larval food plant;
its caterpillars secrete a sugary substance and are tended to by ants (Settle et al., 2010).
Myrmecophilous relationships are common in Lycaenidae butterflies with about 75% of
species having an association with ants (Pierce et al., 2002). However, the genus Satyrium
has fewer species with ant associations than other genera (Fiedler, 1991). Attendant ants
may provide protection to the caterpillars from parasites or predators but the benefits of a
myrmecophilous relationship are likely to be species-specific (Pierce et al., 2002). It is
unknown if the larvae of the Sinai Hairstreak are tended by ants. Unlike the Sinai Baton Blue,
the Sinai Hairstreak has received little attention since its discovery and very little is known
about its biology and ecology. What little is known comes from the original description by
(Nakamura, 1975).
The larval food plant of the Sinai Hairstreak is the Sinai Buckthorn Rhamnus
dispermus, found in Egypt, South Jordan, Palestine, Israel and Lebanon (Boulos, 2000) (Plate
1.2). Rhamnus dispermus trees were searched for the presence of adult Sinai Hairstreaks, or
a related species, in South Jordan in 1998 but none were found (Benyamini, 2000). Rhamnus
dispermus is a dioecious, polygamous shrub (i.e. it has both hermaphroditic and unisexual
flowers on the same tree or on separate trees). It has small green leaves and an intricately
branched stem, with a woody spine at the end of each twig. Both sexes produce flowers
while the females also produce seeds encased in a yellowish drupe. Mature trees have
previously been recorded as being between 0.5 and 1.8 m in height (Boulos, 2000). Several
features of Rhamnus dispermus and the quality of habitat will be assessed in this study to
describe as accurately as possible the specific requirements of the Sinai Hairstreak. Factors
other than the presence of the larval food plant may be important for the Sinai Hairstreak
such as the size or location of the larval food plant, the presence of a specific food plant or
the quality of habitat. Understanding the specific habitat requirements for a species is very

5
important for its conservation and developing management plans if necessary (Sutherland,
2009). For example, the Woodland Brown butterfly Lopinga achine is a Palaearctic species
found in woodland that has been in decline across its range in recent decades. The specific
habitat requirements for the Woodland Brown butterfly differ across its range and
understanding these differences is essential for conservation planning (Kodandaramaiah et
al., 2012).
It is unknown whether Rhamnus dispermus is under threat in the St Katherine
Protectorate. The Bedouin collect many plants in the mountains and wadis for medicinal
purposes. Sinai Thyme, the larval food plant for the Sinai Baton Blue, is collected extensively
by Bedouin herbalists, and might be one of the main threats facing the Sinai Baton Blue
(Hoyle and James, 2005). Historically the thorns of Rhamnus dispermus trees have been
occasionally used by Bedouin to construct traps for Nubian Ibex Capra nubiana (Hobbs,
1992). However, these were only used as a substitute if Palm tree leaves or Acacia raddiana
thorns were unavailable, and it is likely that only a small part of the tree was used (Hobbs,
1992). The hunting of Ibex has been illegal in the protectorate since 1996, and so this is very
unlikely to represent a real threat to Rhamnus dispermus. It has not previously been
established if Rhamnus dispermus is used in any other way by local people. Over-grazing of a
habitat can have negative implications for a butterfly species. The abundance of the Alpine
endemic species Erebia calcaria was found to be significantly affected by grazing intensity
(De Groot et al., 2009). The Bedouin keep herds of goats and allow them to graze in the
wadis, and thus theoretically many plant species may be under threat, although there is little
or no evidence for this (Gilbert, 2011). The three known sites for the Sinai Hairstreak are
relatively remote and at high altitude, so a priori it seems unlikely the trees are at serious
risk from grazing in these areas.
Global warming and habitat fragmentation pose a significant threat to biodiversity
(Opdam and Wascher, 2004). The Sinai Hairstreak could be at great risk of extinction from
global warming. In the last one hundred years, the average temperature in the Middle East
has risen by 0.7° Celsius (Brohan et al., 2006). A study of European butterflies showed that
the ranges of many southern species have retracted northward (Reemer et al., 2003). As the
genus Satyrium is Holarctic and the Sinai Hairstreak is at the southernmost limit of the
distribution, wedged between Europe, Africa and the Middle East, this could be a significant
threat to its existence. Projections suggest that Egypt will have decreased precipitation and
in the Middle East generally an increase in temperature of between 3.5° and 5°C has been
predicted (Met Office, 2011). Thus, the habitat available to the Sinai Hairstreak may become
further restricted in the future.
It is reasonable to assume the Sinai Hairstreak lives in discrete patches like the Sinai
Baton Blue due to the arid environment and distribution of its larval food plant (James,
2006b). Further knowledge of the distribution of these patches will provide background to
the spatial dynamics of this butterfly. The landscape of the Sinai is harsh and fragmented,
and nothing is known about the dispersal ability of the Sinai Hairstreak. Nakamura suggests
that the Sinai Hairstreak is a good flier, but he gave no data (Nakamura, 1975). By
quantifying the amount of suitable habitat, the connectivity of the habitats and the dispersal

6
ability of the Sinai Hairstreak, we can understand more about the conservation requirements
of this unique butterfly.
Thus the aims of this study are to describe the distribution of both the Sinai
Hairstreak and its larval food plant Rhamnus dispermus in the St Katherine Protectorate, as
well as to provide the first population estimate for the Sinai Hairstreak. The general ecology,
behaviour and habitat/conservation requirements of the Sinai Hairstreak will also be
examined. The Sinai Hairstreak is a flagship species of the area, yet next to nothing is known
about its conservation requirements. Considering the extreme nature of the habitat of South
Sinai and the small size of the sites where the butterfly is found, the Sinai Hairstreak may be
at considerable risk of extinction.

7
.
Plate 1.1: Two adult Sinai Hairstreaks Satyrium jebelia on a Rhamnus dispermus branch in
Wadi El-Freya, Saint Katherine Protectorate, South Sinai. Photo by Andrew Power (2012)
Plate 1.2: A typical Rhamnus dispermus tree in Wadi Ahmar, Saint Katherine Protectorate,
South Sinai. Photo by Andrew Power (2012)

8
2 Materials and Methods
Study area
The St Katherine Protectorate is one of Egypt’s largest National Parks and is situated in the
South Sinai, spanning an area of 4,350 km² (Grainger and Gilbert, 2008). Known as the ‘Roof
of Egypt’ the area is predominantly mountainous with deep extensive dry valleys ‘wadis’ cut
throughout the landscape. The igneous Pre-Cambrian Ring Dyke encompassing 640 km² lying
at the centre of the National Park was classified by UNESCO as a World Heritage Site in 2002
for its cultural and religious importance. The massif is 600 million years old and is comprised
mainly of red granite (UNESCO, 2002). At the heart of the Ring Dyke lies the city of St
Katherine, situated at the foot of Mount Sinai, where according to Abrahamic religions
Moses received the ten commandments, and Jebel Katherine, Egypt’s highest mountain at
2,641 m above sea level. It is also home to the Monastery of St Katherine, the oldest
continuously working Christian monastery in the world, whence the area derives its name
(Grainger and Gilbert, 2008). The study sites are found around the high mountain peaks in
the heart of the massif.
The mountain massif is a local hotspot of biodiversity. The Sinai Peninsula is wedged
between Africa and Asia and is very close to Europe; the area has an unusual blend of flora
and fauna from these continents. South Sinai has a Sahara-Mediterranean climate with hot
summers (up to 36°C) and cold winters, and is the coolest area in Egypt (UNESCO, 2002). On
average the area receives 62 mm of rainfall a year (Zahran and Willis, 2009). The high
mountains are sometimes covered in snow during the winter, and snow melt provides the
area with more water throughout the rest of the year (Grainger, 2003). The relatively high
amount of water available compared to the rest of Egypt has resulted in a high diversity of
plants and animals. There are almost 500 species of plant in the area, many of which are
endemic (Ayyad et al., 2000). The IUCN have declared the area as one of the most important
centres for plant diversity in the Middle East (Grainger and Gilbert, 2008). The area is also
important for insects; two-thirds of the butterfly species found in Egypt have been recorded
in the St Katherine Protectorate (Gilbert and Zalat, 2007). Important numbers of resident
and migratory birds utilise the area and it was designated as an important bird area (IBA) by
Birdlife International (BirdLife International, 2012).
Distribution and population estimates
The mountains and wadis around the city of St Katherine were searched for the presence of
Rhamnus dispermus in April and May before the main flight period of the Sinai Hairstreak.
The location of each tree discovered was recorded with a portable Global Positioning
Satellite receiver (GPS) (Garmin, eTrex Venture HC, accurate to ± 5 m) and a map of all the
sites produced. Each tree was assigned an individual number and yellow tags were used to
distinguish trees in the field more easily. All the sites were revisited during the main flight
period to search for the presence of adult Sinai Hairstreaks. In addition to providing
information on the distribution of the larval food plant the local Bedouin were asked if the

9
trees have any medicinal, cultural or practical importance to their tribe. All fieldwork in St
Katherine took place under the supervision of an experienced Bedouin guide.
Population estimates for adult Sinai Hairstreaks were carried out at each site in June
and July. A five-day Mark-Release-Recapture (MRR) experiment was carried out at each of
the four sites. Every tree at each site was visited multiple times to search for butterflies
following a rough transect. Butterflies were caught with a large butterfly net from 08:00 to
16:30 (local time, GMT+2) for five consecutive days. Each butterfly captured was given an
individual mark with a felt-tip pen through the net and released as quickly as possible. An
example of a marked individual is represented in Plate 2.1. The location, behaviour and time
were recorded for each capture or subsequent recapture. Population sizes were estimated
using Eberhardt’s geometric model (Pollard, 1977). The first MRR took place between the 2nd
and 6th of June in Jebel Katherine (Fig 2.1). Abu Druce was surveyed between the 17th and
21st of June (Fig 2.2), Abu Towaita between the 24th and 28th of June (Fig 2.3) and the final
MRR was conducted in the Jebel Bab region between the 7th and 11th of July (Fig 2.4). Maps
were created using Google Earth showing the movements of captured Sinai Hairstreaks, the
distribution of Rhamnus dispermus and the Sinai Hairstreak.
Given the large size of the Wadi Ahmar region and the apparent high population of
Sinai Hairstreaks at the site, a different method was used here. A pre-determined transect
route through Ahmar was walked on the 8th of June encompassing as many of the larval food
plants as possible (Fig 2.5). Adults were caught and given a site-specific mark and released
quickly. The site was revisited on the 12th of June giving the marked individuals time to
redistribute into the population at random. The same transect route was used, unmarked
individuals were given a different mark and recaptured individuals were re-marked to avoid
double counting. Marking took place between 08:00 and 16:00 (local time). The population
size was estimated using the Lincoln index.
Plate 2.1: A marked Sinai Hairstreak Satyrium jebelia on a Rhamnus dispermus tree in Abu
Druce, Saint Katherine Protectorate, South Sinai. Photo by Andrew Power (2012)

10
Figure 2.1: Map of Jebel Katherine showing transect line used for MRR and the distribution
of Rhamnus dispermus (Google Earth, 2012) (white symbols show Rhamnus trees where no
Sinai Hairstreaks were recorded during the study, red symbols show Rhamnus trees where
Sinai Hairstreaks were present during study)
Figure 2.2: Map of Abu Druce showing transect line used for MRR and the distribution of
Rhamnus dispermus (Google Earth, 2012) (white symbols show Rhamnus trees where no
Sinai Hairstreaks were recorded during the study, red symbols show Rhamnus trees where
Sinai Hairstreaks were present during study)
11
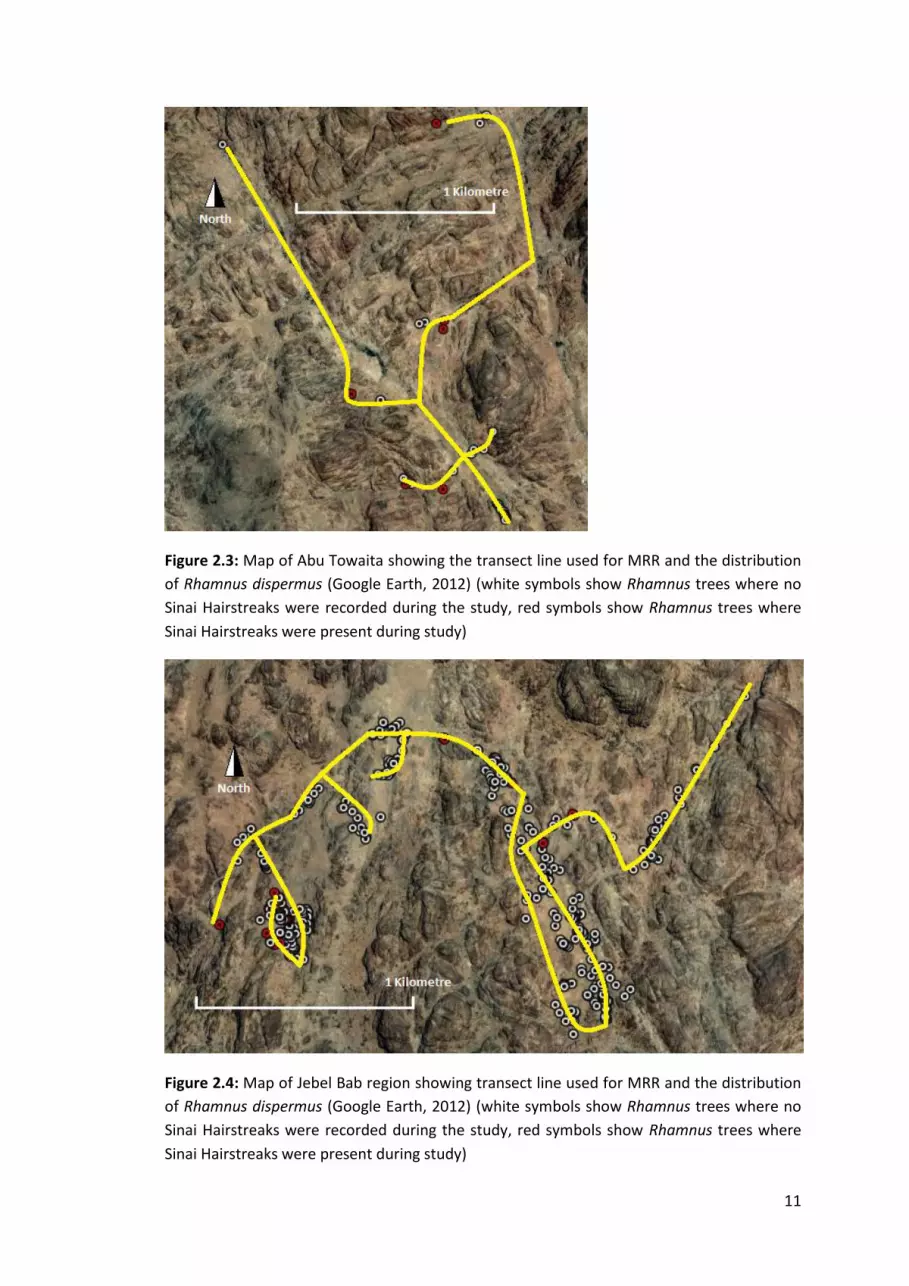
Figure 2.3: Map of Abu Towaita showing the transect line used for MRR and the distribution
of Rhamnus dispermus (Google Earth, 2012) (white symbols show Rhamnus trees where no
Sinai Hairstreaks were recorded during the study, red symbols show Rhamnus trees where
Sinai Hairstreaks were present during study)
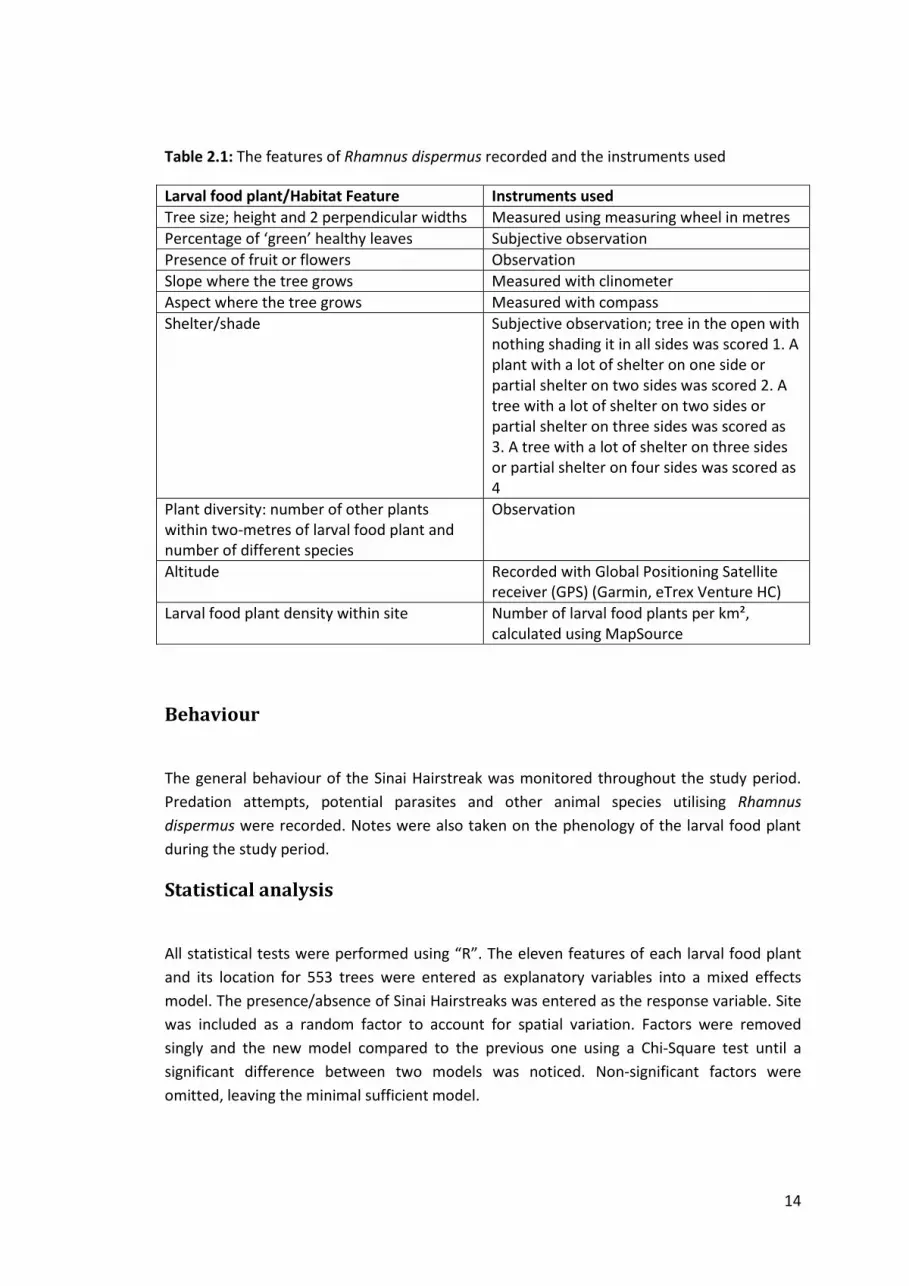
Figure 2.4: Map of Jebel Bab region showing transect line used for MRR and the distribution
of Rhamnus dispermus (Google Earth, 2012) (white symbols show Rhamnus trees where no
Sinai Hairstreaks were recorded during the study, red symbols show Rhamnus trees where
Sinai Hairstreaks were present during study)

12
Figure 2.5: Map of Wadi Ahmar region showing transect line used for MRR and the
distribution of Rhamnus dispermus (Google Earth, 2012) (white symbols show Rhamnus trees
where no Sinai Hairstreaks were recorded during the study, red symbols show Rhamnus
trees where Sinai Hairstreaks were present during study)
Wadi Ahmar was visited throughout the study period to identify the flight time and peak
emergence of the Sinai Hairstreak. A fixed transect route was used to census the butterflies
(Fig 2.6). Starting at 10:00 (local time), a 720 m transect was walked, encompassing as many
trees in the wadi as possible. Transects were walked at a slow, steady pace counting all
butterflies seen within a fixed distance of 2.5 m on either side and five metres ahead. The
total number of adult Sinai Hairstreaks observed was noted. The transect method used was
based on standardised ‘Pollard Walks’ frequently adopted in butterfly monitoring schemes in
Great Britain (Pellet, 2008). The survey was conducted on the 29th of May, 11th of June, 2nd of
July, 14th of July, 17th of July and on the 27th of July.

13
Figure 2.6: Map of Wadi Ahmar showing transect line used and the distribution of Rhamnus
dispermus (Google Earth, 2012) (white symbols show Rhamnus trees where no Sinai
Hairstreaks were recorded during the study, red symbols show Rhamnus trees where Sinai
Hairstreaks were present during study)
Larval food plant features
To assess the habitat and larval food plant requirements, eleven features of the
larval food plant and its location were recorded (Table 2.1). The height and two
perpendicular widths of each tree were measured using a measuring wheel. The volume was
then calculated assuming the tree is semi-ellipsoidal in shape. The percentage of the tree
covered in healthy green leaves was noted subjectively. Each tree was checked for the
presence of fruit or flowers. The slope of the area where each tree was growing was
measured in degrees using a clinometer. The aspect was measured using a compass (North,
South, East, and West). The amount of shelter/shade covering a tree was also noted
subjectively. The number of other plants within a two-metre radius of each tree was
recorded, as well as the number of different species, to enable the calculation of biodiversity
around each tree - recorded as simple species richness. An estimation of diversity at a site
level was calculated by selecting 25 random Rhamnus trees from each site (excluding Wadi
El-Freya) and measuring diversity around all 25 trees using Hill’s numbers 1D (i.e.
exponential Shannon)(Tuomisto, 2010). The altitude was assessed using a portable GPS. The
total number of larval food plants at a given site was recorded as well as the density of larval
food plants in the site. Site size was calculated using MapSource (mapping software provided
by Garmin).
14
Table 2.1: The features of Rhamnus dispermus recorded and the instruments used
Larval food plant/Habitat Feature Instruments used
Tree size; height and 2 perpendicular widths Measured using measuring wheel in metres
Percentage of ‘green’ healthy leaves Subjective observation
Presence of fruit or flowers Observation
Slope where the tree grows Measured with clinometer
Aspect where the tree grows Measured with compass
Shelter/shade Subjective observation; tree in the open with nothing shading it in all sides was scored 1. A plant with a lot of shelter on one side or partial shelter on two sides was scored 2. A tree with a lot of shelter on two sides or partial shelter on three sides was scored as 3. A tree with a lot of shelter on three sides or partial shelter on four sides was scored as 4
Plant diversity: number of other plants within two-metres of larval food plant and number of different species
Observation
Altitude Recorded with Global Positioning Satellite receiver (GPS) (Garmin, eTrex Venture HC)
Larval food plant density within site Number of larval food plants per km², calculated using MapSource
Behaviour
The general behaviour of the Sinai Hairstreak was monitored throughout the study period.
Predation attempts, potential parasites and other animal species utilising Rhamnus
dispermus were recorded. Notes were also taken on the phenology of the larval food plant
during the study period.
Statistical analysis
All statistical tests were performed using “R”. The eleven features of each larval food plant
and its location for 553 trees were entered as explanatory variables into a mixed effects
model. The presence/absence of Sinai Hairstreaks was entered as the response variable. Site
was included as a random factor to account for spatial variation. Factors were removed
singly and the new model compared to the previous one using a Chi-Square test until a
significant difference between two models was noticed. Non-significant factors were
omitted, leaving the minimal sufficient model.

15
3 Results
Local knowledge
The Sinai Buckthorn Rhamnus dispermus, known locally as ‘Orontol’, did not appear to be an
important tree to the Bedouin people. Many Bedouin are unfamiliar with this species and do
not distinguish it from other thorn bushes in the area, such as the Sinai Hawthorn Crataegus
x sinaica, known locally as ‘Zah-rur’. Crataegus x sinaica is more widespread than Rhamnus
dispermus and has a place in Bedouin folklore, with most local people being aware of it. A
large partially damaged Rhamnus dispermus tree was observed in Abu Towaita on the 21st of
May where branches had been removed to block off a deep unused well next to the tree.
This was the only observed use of the tree. As to the butterfly, while some Bedouin were
familiar with the Sinai Baton Blue (due to our previous research in the area, rather than it
being an animal known in Bedouin folklore), they were unfamiliar with the Sinai Hairstreak.
Very few insects are distinguished by species (Zalat et al., 2008).
Rhamnus dispermus
Distribution
590 Rhamnus dispermus trees were discovered in nine discrete ‘sites’ (Fig 3.1): site
boundaries were assigned using the natural topography of the landscape. A large site (0.7
km²) of 145 trees, referred to in this study as Ahmar, was found in Jebel Ahmar, spreading
into Wadi Ahmar and partially into Wadi Matter (Fig 3.2). Most trees were found along the
wadi with many on the steep slopes on either side. A total of 33 Rhamnus dispermus trees
were discovered in Jebel Katherine (0.2 km²) along a small wadi near Ain Shinar, adjacent to
the main path to the summit of Jebel Katherine (Fig 3.3). Most trees were on the rocky
slopes rather than in the valley itself. A compact site of 45 trees (0.1 km²) was found in Abu
Druce with one outlier tree (not shown in Fig 3.4). Most trees were found along the bottom
of the wadi. A large (2.3 km²) patchy site was discovered in Abu Towaita with 27 trees (Fig
3.5). Six trees were discovered in Wadi El-Freya (2.1 km²), all of them high up on the
mountain slopes (Fig 3.6). The largest site discovered (2.2 km²), referred to in this study as
Bab, stretched from the top of Nagba Harear into Wadi Zatar and Ain Nigela to the foot of
Jebel Bab. A total of 334 trees were found in this site (Fig 3.7). Single trees were discovered
high up near Abu Himmen, in a tributary valley off Wadi Jebel and in Abu Teenya. The size of
each site and densities of Rhamnus dispermus trees are shown in Table 3.1.

16
Figure 3.1: Map of St Katherine Protectorate showing distribution of Rhamnus dispermus and
Sinai Hairstreak (white symbols show Rhamnus trees where no Sinai Hairstreak were
recorded during the study, red symbols show Rhamnus trees where Sinai Hairstreaks were
present during study)
Figure 3.2: Map of Ahmar region showing distribution of Rhamnus dispermus and Sinai
Hairstreak (white symbols show Rhamnus trees where no Sinai Hairstreak were recorded
during the study, red symbols show Rhamnus trees where Sinai Hairstreaks were present
during study

17
Figure 3.3: Map of Jebel Katherine showing distribution of Rhamnus dispermus and Sinai
Hairstreak (white symbols show Rhamnus trees where no Sinai Hairstreak were recorded
during the study, red symbols show Rhamnus trees where Sinai Hairstreaks were present
during study)
Figure 3.4: Map of Abu Druce showing distribution of Rhamnus dispermus and Sinai
Hairstreak (white symbols show Rhamnus trees where no Sinai Hairstreak were recorded
during the study, red symbols show Rhamnus trees where Sinai Hairstreaks were present
during study)

18
Figure 3.5: Map of Abu Towaita showing distribution of Rhamnus dispermus and Sinai
Hairstreak (white symbols show Rhamnus trees where no Sinai Hairstreak were recorded
during the study, red symbols show Rhamnus trees where Sinai Hairstreaks were present
during study)
Figure 3.6: Map of Wadi El-Freya showing distribution of Rhamnus dispermus and Sinai
Hairstreak (white symbols show Rhamnus trees where no Sinai Hairstreak were recorded
during the study, red symbols show Rhamnus trees where Sinai Hairstreaks were present
during study)

19
Figure 3.7: Map of Bab region showing distribution of Rhamnus dispermus and Sinai
Hairstreak (white symbols show Rhamnus trees where no Sinai Hairstreak were recorded
during the study, red symbols show Rhamnus trees where Sinai Hairstreaks were present
during study
Table 3.1: The number and density of Rhamnus dispermus trees in each site
Site Area km² No. of trees Trees per km²
Jebel Katherine 0.2 km² 33 165
Abu Druce 0.1 km² 42 420
Abu Towaita 2.3 km² 27 11.8
Wadi El-Freya 1.3 km² 6 4.6
Bab 2.2 km² 330 150
Ahmar 0.7 km² 145 207
Phenology of Rhamnus dispermus
On the 22nd of April in Jebel Ahmar the trees looked in excellent condition with bright green
leaves and many buds, some flowering. All flowers observed were light green/yellow.
Similarly on the 25th of April the trees in Abu Druce appeared to be in very good condition
with more flowers than emerging buds. A red colouration at the centre of some flowers was
noted. Fruit (red and green) and withered flowers were first noticed in Wadi El-Freya on the

20
10th of May. The leaves also appeared more ‘tatty’. From the 2nd of June to the 6th of June in
Jebel Katherine, 11 out of 33 trees had flowers while 17 out of 33 had fruit. Flowers were last
recorded between the 17th and 21st of June in Abu Druce where only two out of 43 trees had
any flowers. The leaves on all trees in Wadi El-Freya had almost completely withered by the
3rd of July with fruit present on two out of six trees. From the 7th to the 11th of July, clusters
of trees with withered leaves were noticed in Bab. Several trees showed signs of withering in
Ahmar on the 17th of July but generally trees were still in good condition. Withering was
more evident in Ahmar by the 6th of August and in Bab on the 9th of August. Fruit was still
present in both sites at this time with 35 out of 108 trees in Jebel Ahmar having fruit and 114
out of 300 in Jebel Bab. Tree size varied highly with height ranging from 0.02 m to 5.90 m
with the average being 1.29 m (Fig 3.8). Rhamnus dispermus trees were found at altitudes of
between 1584 m and 2368 m above sea level. Fruit was recorded on 187 trees in total out of
553 (Fig 3.9). Most trees had a healthy percentage of green leaves (Fig 3.10).
Figure 3.8: Frequency distribution of tree heights
0
20
40
60
80
100
120
140
160
No
. of
tre
es
Metres

21
Figure 3.9: The number of Rhamnus dispermus trees with or without fruit
Figure 3.10: The percentage of healthy green leaves on all Rhamnus dispermus trees
0
50
100
150
200
250
300
350
400
Fruit No Fruit
No
. of
tre
es
0
20
40
60
80
100
120
10 20 30 40 50 60 70 80 90 100
No
. of
tre
es
% Green

22
Sinai Hairstreak
Distribution
Sinai Hairstreaks were recorded on a total of 130 out of the 590 trees across all sites.
Butterflies were absent from only three sites; however, only one tree was present in each of
these sites. Sinai Hairstreaks were recorded on 62 trees in Ahmar, nine trees in Jebel
Katherine, 24 trees in Abu Druce, seven trees in Abu Towaita, three trees in Wadi El-Freya
and observed on 25 trees in Bab, mostly located in the west near the foot of Jebel Bab. Single
trees were discovered high up near Abu Himmen, in a tributary valley off Wadi Jebel and in
Abu Teenya. Sinai Hairstreaks were not seen at any of these locations. The dates each site
was visited for MRR can be seen in Table 3.2.
Table 3.2: The dates each site was visited for MRR
Site visited Date
Jebel Katherine 2-6/06/2012
Ahmar 8 and 12/06/2012
Abu Druce 17-21/06/2012
Abu Towaita 24-28/06/2012
Wadi El-Freya 3/07/2012
Bab 7-11/07/2012
Butterfly movements
The trees utilised by Sinai Hairstreaks and the movements of recaptured individuals can be
seen in Figures 3.11-3.14. The average distance travelled by butterflies recaptured was 104
m. The maximum dispersal distance recorded was 938 m by butterfly b1 in Bab (Fig 3.14 (a)).
A frequency distribution of distances travelled by recaptured butterflies can be seen in
Figure 3.15.

23
Figure 3.11: Map of Jebel Katherine showing movements of Sinai Hairstreaks and distribution
of Rhamnus dispermus (white symbols show Rhamnus trees where no Sinai Hairstreaks were
recorded during the study, red symbols show Rhamnus trees where Sinai Hairstreaks were
present during study)
Table 3.3: Recaptured Sinai Hairstreak movements in Jebel Katherine showing Rhamnus
dispermus tree number where captured (unless otherwise stated), time of capture and total
distance travelled by each butterfly
Butterfly No. Day 1 Day 2 Day 3
Day 4 Day 5 Total distance travelled (metres)
(b) 1 14 (11.26) 1 (15:19)
227.83
(b) 4 16 (12:30) 13 (16:11) 10 (12:11)
60.87
(b) 5 Shohat (8:35)
Shohat (8:48) 23 (13:50)
9
(b) 10 19 (10:34) 16 (12:51)
31.73
(b) 11 16 (11:07) 19 (9:57)
31.73
(b) 12 10 (11:40) 16 (12:59)
59.38
(b) 16 13 (12:42) 16 (12:50)
12.42

24
Figure 3.12 (a): Map Abu Druce showing movements of Sinai Hairstreaks and distribution of
Rhamnus dispermus (white symbols show Rhamnus trees where no Sinai Hairstreaks were
recorded during the study, red symbols show Rhamnus trees where Sinai Hairstreaks were
present during study)
Table 3.4 (a): Recaptured Sinai Hairstreak movements in Abu Druce showing Rhamnus
dispermus tree number where captured (unless otherwise stated), time of capture and total
distance travelled by each butterfly
Butterfly No. Day 1 Day 2 Day 3 Day 4 Day 5 Total distance travelled (metres)
(b) 8
5 (13:29)
10 (15:26) 12
(b) 9
5 (12:33)
9 (13:48) 11 (13:56)
2 (11:36) 2 (11:37) 7 (11:53)
128.36
(b) 10 7 (13:39)
2 (11:31) 26 (15:30)
149.28
(b) 26 29 (14:29) 32 (15:44) 12.15
(b) 27 14 (13:57) 10 (13:48) 15.24
(b) 38 7 (13:02) 33 (13:00) 79
(b) 56
17 (13:45) 8 (14:32)
24.34

25
Figure 3.12 (b): Map Abu Druce showing movements of Sinai Hairstreaks and distribution of
Rhamnus dispermus (white symbols show Rhamnus trees where no Sinai Hairstreaks were
recorded during the study, red symbols show Rhamnus trees where Sinai Hairstreaks were
present during study)
Table 3.4 (b): Recaptured Sinai Hairstreak movements in Abu Druce showing Rhamnus
dispermus tree number where captured (unless otherwise stated), time of capture and total
distance travelled by each butterfly
Butterfly no. Day 1 Day 2 Day 3 Day 4 Day 5 Total distance travelled (metres)
(b) 12 10 (13:51)
5 (12:17) 11 (13:52)
9 (11:55)
9 (11:56)
53.50
(b) 15 14 (14:09)
6 (15:16)
22.19
(b) 37 2 (12:06)
23 (13:28)
71.53
(b) 43 22 (15:09)
Crataegus x sinaica (10:58)
63.80
(b) 51 7 (12:00)
29 (13:57)
108.53

26
Figure 3.12 (c): Map Abu Druce showing movements of Sinai Hairstreaks and distribution of
Rhamnus dispermus (white symbols show Rhamnus trees where no Sinai Hairstreaks were
recorded during the study, red symbols show Rhamnus trees where Sinai Hairstreaks were
present during study)
Table 3.4 (c): Recaptured Sinai Hairstreak movements in Abu Druce showing Rhamnus
dispermus tree number where captured (unless otherwise stated), time of capture and total
distance travelled by each butterfly
Butterfly no. Day 1 Day 2 Day 3 Day 4 Day 5 Total distance travelled (metres)
(b) 16 14 (14:14) 23 (14:20)
7 (12:03)
38.43
(b) 41 11 (14:23) 9 (16:09)
9 (11:55) 22 (12:27)
7 (11:57)
6.21
(b) 56 17 (13:45) 8 (14:32)
24.34
(b) 63 14 (12:24) At water source near tree 7 (13:45)
17

27
Figure 3.13: Map Abu Towaita showing movements of Sinai Hairstreaks and distribution of
Rhamnus dispermus (white symbols show Rhamnus trees where no Sinai Hairstreaks were
recorded during the study, red symbols show Rhamnus trees where Sinai Hairstreaks were
present during study)
Table 3.5: Recaptured Sinai Hairstreak movements in Abu Towaita showing Rhamnus
dispermus tree number where captured (unless otherwise stated), time of capture and total
distance travelled by each butterfly
Butterfly no. Day 1 Day 2 Day 3 Day 4 Total distance travelled (metres)
(b) 9 Crataegus x sinaica (12:45)
9 (13:11)
424.76
(b) 11 13 (11:30)
13 (11:24) 12 (14:40)
12 (11:35)
37.16

28
Figure 3.14 (a): Map of Jebel Bab region showing movements of Sinai Hairstreaks and
distribution of Rhamnus dispermus (white symbols show Rhamnus trees where no Sinai
Hairstreaks were recorded during study, red symbols show Rhamnus trees where Sinai
Hairstreaks were present during study)
Table 3.6 (a): Recaptured Sinai Hairstreak movements in the Jebel Bab region showing
Rhamnus dispermus tree number where captured (unless otherwise stated), time of capture
and total distance travelled by each butterfly.
Butterfly no. Day 1 Day 2 Day 3 Day 4 Day 5 Total distance travelled (metres)
(b) 1 186 (9:40)
C Crataegus x sinaica 6 (14:14)
938
(b) 11 Near Crataegus x sinaica (10:14)
Crataegus x sinaica 7a (9:34)
150.43
(b) 12 Crataegus x sinaica 4 (10:32) Crataegus x sinaica 7b (15:40)
24
(b) 21 Plant X (16:04)
182 (10:00) 24.81

29
Figure 3.14 (b): Map of Jebel Bab region showing movements of Sinai Hairstreaks and
distribution of Rhamnus dispermus (white symbols show Rhamnus trees where no Sinai
Hairstreaks were recorded during the study, red symbols show Rhamnus trees where Sinai
Hairstreaks were present during study)
Table 3.6 (b): Recaptured Sinai Hairstreak movements in the Jebel Bab region showing
Rhamnus dispermus tree number where captured (unless otherwise stated), time of capture
and total distance travelled by each butterfly
Butterfly no. Day 1 Day 2 Day 3 Day 4 Day 5 Total distance travelled (metres)
(b) 14 236 (12:39) 268 (13:59) 293 (14:06)
140
(b) 17 268 (13:54) 235 (12:00)
87.67
(b) 31 291 (13:52) Crataegus x sinaica 6 (13:49)
254 (11:11) 293 (11:47)
359.65
(b) 32
221 (14:20) 235 (12:00) 75
(b) 33 224 (14:35) Near 235 (12:01)
59
(b) 34 Crataegus x sinaica 8 (14:44)
237 (12:39) 243 (14:21)
68.56

30
Fig 3.15: Frequency distribution of total distances travelled by all butterflies recpatured
Population estimates
54 Sinai Hairstreaks were marked in Ahmar on the 8th of June. 11 previously marked
individuals and 82 newly marked individuals were observed in Ahmar on the 12th of June.
The number of captures and recaptures during the five-day MRR in Jebel Katherine, Abu
Druce, Abu Towaita and Bab can be seen in Table 3.7. An MRR was attempted in Wadi El-
Freya on the 3rd of July; however, there were no butterflies present. Rhamnus dispermus
trees were in bad condition in comparison to previous visits. A minimum of six individuals
were recorded there previously on the 10th of May. Population estimates for each site can be
seen in Table 3.8. The Lincoln index was used in Ahmar because sampling took place on just
two days. Eberhardt’s geometric model was used where sampling took place over five
consecutive days.
Table 3.7: The number of butterflies caught at each site and the number of times butterflies
were recaptured
Site Total no. of butterflies caught
No. of butterflies caught once
No. of butterflies recaught once
No. of butterflies recaught twice
No. of butterflies recaught 3 times
Jebel Katherine
19 11 7 1 0
Abu Druce 66 45 15 4 2
Abu Towaita 25 20 3 2 0
Bab 45 35 9 1 0
0
5
10
15
20
25
30
<100 <200 <300 <400 <500 <600 <700 <800 <900 <1000
No
. of
bu
tte
rflie
s
Distance travelled (metres)

31
Table 3.8: Population estimates for each site
Site Method Population Estimate
Ahmar Lincoln index 403 Jebel Katherine
Eberhardt’s geometric model 57
Abu Druce Eberhardt’s geometric model 214
Abu Towaita Eberhardt’s geometric model 111
Bab Eberhardt’s geometric model 225
Wadi El-Freya
Transect count 6
Total 1,010 95% confidence interval 745-1,275
Activity times
Adult Sinai Hairstreaks were first observed on the 10th of May in Wadi El-Freya (six
individuals), and the last individual on the 27th of July in Wadi Ahmar. Regular transect
counts did not start in Wadi Ahmar until the 29th of May as potential new sites were being
searched for the presence of Rhamnus dispermus. No butterflies were sighted between the
initial discovery on the 10th of May and the start of transect counts on the 29th. However, the
areas visited in this period did not contain Rhamnus dispermus trees and were not suitable
as habitat for Sinai Hairstreaks. According to the transect results the main flight period was
between late May to mid-July (Fig 3.16). The numbers of individuals observed on each
transect is represented in Table 3.9. The time of each butterfly capture during MRR showed a
single peak activity between 11:00 and 14:00 (local time), with the majority of the captures
taking place between 10:00 and 15:00 (Fig 3.17).

32
Figure 3.16: Number of Sinai Hairstreaks observed on transect walks through Wadi Ahmar,
the first Sinai Hairstreaks recorded in Wadi El-Freya are shown on day one
Table 3.9: Number of Sinai Hairstreaks observed on transect walks through Wadi Ahmar
Day number Date No. of butterflies recorded
20 29th May 48
33 11th June 46
54 2nd July 35
66 14th July 9
69 17th July 1
79 27th July 0

33
Figure 3.17: The time of day that Sinai Hairstreaks were captured during the study period
Behaviour and ecology
Based on observations the Sinai Hairstreak is a highly territorial and aggressive species. Long-
tailed Blues Lampides boeticus were commonly observed being chased off by the Sinai
Hairstreak. Painted Lady Vanessa cardui, Pomegranate Playboy Deudorix livia, Clouded
Yellow Colias croceus and an unidentified red parasitic wasp (about 5 cm in length) were also
chased off. Butterflies were almost always seen on or in the vicinity of their larval food plant.
They were also recorded landing on Oreganum syriacum, Mentha longifolia, Teucrium
polium and Crataegus x sinaica. Although age was not specifically analysed in this study, a
Sinai Hairstreak was observed on the 2nd of July in Ahmar displaying a mark from the MRR in
the same location on the 8th of June. At 25 days, this individual was the oldest Sinai
Hairstreak recorded.
Predators and parasites
A Rock Martin Ptyonoprogne fuligula predation attempt on an adult Sinai Hairstreak was
witnessed in Wadi El-Freya on the 10th of May and in Jebel Ahmar on the 29th of May. Rock
Martins are common and actively hunt in all sites. Scrub Warblers Scotocera inquieta were
recorded in every site and regularly frequented Rhamnus dispermus trees in search of food,
often hunting methodically in pairs or small groups. A Scrub Warbler nest was found on
Rhamnus dispermus trees in Jebel Katherine and Wadi Ahmar. A female Blackcap Sylvia
atricapilla and a female Redstart Phoenicurus phoenicurus were observed foraging in a
Rhamnus dispermus tree in Jebel Katherine on the 4th of June. A praying mantid species was
found in a Rhamnus dispermus tree in Wadi El-Freya on the 31st May. A White-Crowned
0
10
20
30
40
50
60

34
Black Wheatear Oenanthe leucopyga was observed trying to pull off a yellow plant tag on
Rhamnus dispermus trees in Jebel Katherine on the 6th of June and in Abu Towaita on the
27th of June. A relatively large unidentified red parasitic wasp was commonly seen patrolling
around Rhamnus dispermus in Jebel Katherine, Ahmar and Abu Druce in June.
Statistical analysis
When all the habitat/tree variables were entered into a mixed effects model to explain the
occurrence of Sinai Hairstreaks, tree size, species richness, presence of fruit and the
percentage green were the only characteristics to contribute significantly to the final model
(Table 3.10). The variance component of the random factor (site) is 0.597. Slope, aspect,
altitude, shelter, tree density were ejected from the model as non-significant.
Table 3.10: Results from the mixed effects model
Factor Chi-squared value
Degrees of freedom (df)
P
% green 4.0741 6 0.04354
Fruit 19.755 5 8.805e-06 Local plant richness
29.914 5 4.517e-08
Tree size 74.324 3 2.2e-16
4 Discussion
Rhamnus dispermus
The morphology of Rhamnus dispermus was highly variable. Most trees were between 0.5 m
and 1.5 m high, as stated in Boulus (2000), but many grew much larger. The highest tree
recorded was 5.9 m. The phenology of Rhamnus dispermus was variable between different
areas. Most trees were in healthy condition at the start of the field season in mid-April with
healthy green leaves, fruit and flower buds. The main flowering period was between late-
April and early June. By mid-June most flowers had withered. Tree leaves started to
degenerate (lost green colouration, turning brown and starting to wither) mainly in late July
and August. Not all trees followed this pattern with some withering much earlier; all trees in
Wadi El-Freya had severely withered by the 3rd of July. During the same field season it was
observed that the phenology of Sinai Thyme was several weeks further advanced than in
previous years (Katy Thompson, pers. comm.). This study has provided the first detailed

35
account of the phenology of Rhamnus dispermus so it is possible that its phenology was also
further advanced than usual. The phenology is similar to other Rhamnus species such as
Rhamnus alpinus found in southern-Europe and North Africa. The flower buds of Rhamnus
alpinus open at the end of April/early May and flower for 2-3 weeks. Leaves begin to wither
in August and trees lose their leaves completely during late-September (Bañuelos and
Obeso, 2004). No Rhamnus dispermus were observed having completely withered. However,
the study period ended in mid-August.
Sinai Hairstreak
The main flight period of the Sinai Hairstreak during this study appeared to be from late May
to early-mid July (Fig 3.16), longer than the earlier estimation of its flight period of late May
to early June (Nakamura, 1975). The life-cycle closely mirrored that of Rhamnus dispermus.
The young larvae presumably fed on the emerging young buds and leaves in April
(Nakamura, 1975) and adults emerged in May when the main flowering period begun. The
eggs of the Blue Spot Hairstreak do not hatch until the following Spring, when the young
larvae feed on young buds and leaves of its larval food plant (Settle et al., 2010). The peak
emergence of adult butterflies in Sinai extended from May to mid-June and the butterfly
season declined and ended as Rhamnus dispermus trees began to wither in late-July and
August. As expected, the Sinai Hairstreak was most active between 10:00 and 15:00 (Fig
3.17), the hottest part of the day. Butterflies are heliotherms that derive much of their heat
from the sun (Vande Velde et al., 2011).
Habitat requirements
Understanding what factors determine the distribution and survival of a species is
fundamental to its conservation. Rhamnus dispermus trees were found in nine discrete sites
in the St Katherine Protectorate. The Sinai Hairstreak was discovered in six of these sites.
However, the sites where the butterfly was not recorded contained only one tree of a very
small size and cannot be considered to be suitable for Sinai Hairstreaks. Within the six main
sites, there were many trees where the Sinai Hairstreak was not observed. Different sites
contained very different numbers of butterflies, and site was the most important predictor
of the presence of butterflies on single trees, explaining almost 60% of variation in the data.
Until now, the only known habitat criterion for the Sinai Hairstreak was the presence
of the larval food plant. Tree size, the presence of fruit, the percentage of healthy green
leaves and high plant diversity were all important factors in explaining the presence or
absence of the Sinai Hairstreak on individual trees within each site. All of these factors are a
function of habitat quality. Good quality habitat with nutrient-rich soil and higher water
availability are probably the most important elements for the growth of large healthy flora
and high floral diversity. Fruit is only present on female trees, and this may also relate to
good quality habitat as female trees require more nutrients than males to grow both fruit
and flowers (Yu and Lu, 2011). In an arid environment the availability of good habitat is

36
presumably very important for the persistence of a butterfly species. The Sinai Hairstreak
was observed feeding on several species of flower and areas with high species abundance
would provide higher food availability. Tree selection by Sinai Hairstreaks for oviposition was
not examined during this study, but it seems likely that butterflies would be found near the
vicinity of a suitable larval food plant. It has been well established that many butterflies
select green leaves for oviposition (Prokopy and Owens, 1983). A large amount of recent
research has concentrated on larval food plant selection and larval food plant specificity and
its implications for butterfly conservation (Strausz et al., 2012). Large trees provide a large
resource area for Sinai Hairstreak to lay eggs on and large trees are also likely to be healthier
providing larvae with a good quality food source.
It is possible that male Sinai Hairstreaks use large trees as outposts to perch and
patrol for females or to defend a territory from other males. Male Grey Hairstreaks Strymon
melinus pick a prominent perch tree where they wait for females, and will defend the perch
from intruding males (Alcock and Oneill, 1986). Sinai Hairstreaks are highly aggressive and
were frequently observed fighting with many butterfly species. On some occasions they
were observed alighting on Crataegus x sinaica. They may be using it as a perching station
but it has been suggested that this may be a second larval food plant (Dr Sabreen Rashad,
pers. comm.). No evidence for this was found in this study, and the issue requires further
investigation. Crataegus x sinaica is common in areas where Rhamnus dispermus occurs, and
Sinai Hairstreaks may simply be using them as temporary resting stations because they are
so prominent in the landscape. Further investigation into the larval food plant preference of
the Sinai Hairstreak is required.
Dispersal ability
Sinai Hairstreaks are very strong fliers. They can be very shy and difficult to approach, with
the slightest disturbance causing the butterfly to fly quickly out of sight. Catching could be
very difficult and butterflies were rarely caught away from the larval food plant. 26% of
butterflies marked were recaptured; 77% of these were recaptured only once. A longer more
intense study would yield more information regarding butterfly movements. Despite this,
however, the data show the Sinai Hairstreak has a high propensity to move and therefore a
strong ability to disperse. The average distance covered by recaptured Hairstreaks was 104
m. This is in sharp contrast to the Sinai Baton Blue which rarely moves further than 100 m
(James, 2006e).
Some Sinai Hairstreaks covered impressive distances, such as butterfly b1 in Jebel
Katherine, which travelled a total distance of 227 m (Fig 3.11). Butterfly b11 in Abu Towaita
moved 425 m (Fig 3.13) while butterfly b31 in Bab travelled 359 m (Fig 3.14 (b)). Butterfly b1
in the Bab region was originally caught on the first day of the MRR at 09:40, and was
subsequently recaptured on the second day 938 m away as the crow flies (Fig 3.14 (a)). The
landscape between these two locations is difficult to navigate, showing that the Sinai
Hairstreak is more than capable of travelling large distances quickly, and certainly has the
ability to move between sites. The Ahmar region and Jebel Katherine are separated by

37
roughly 934 m, while Abu Towaita and Abu Druce are separated by just over a kilometre, all
well within reach of a dispersing Sinai Hairstreak.
Not only has the Sinai Hairstreak been recorded travelling large distances along a
continuous network of larval food plants, as in Bab, it has also been recorded travelling large
distances across terrain bereft of its larval food plant. Assuming that butterfly b11 in Abu
Towaita used the wadi to move between trees, it would have travelled a distance of 425 m
without encountering any Rhamnus dispermus trees (Fig 3.13). A series of interconnecting
wadis in the St Katherine Protectorate link most areas together and a dispersing individual
can use these wadis as corridors where there is less wind resistance and where food plants
are more likely to grow. Some male butterflies can sense female pheromones from great
distances (Nieberding et al., 2008) which may encourage movement between sites, and
butterflies may also learn where larval food plants are. A study of the Meadow Brown
Maniola jurtina butterfly revealed that they do not seek habitat by random flight and can
return to suitable habitat they have previously visited (Conradt et al., 2000).
No butterflies were recaptured in a site different from where it was originally
marked. The emphasis of this study was to assess the population size, but the study was
designed to maximise the chances of recording between-site movements; after an MRR was
completed in one site, the site closest to it was sampled next. Further studies concentrating
on fewer sites need to be carried to confirm whether butterflies move between sites. The
Sinai Hairstreak can live up to at least 25 days, which is a long time for a butterfly: male
Glanville Fritillaries Melitaea cinxia have an average lifespan of 14 days and a maximum of 36
days (Saastamoinen et al., 2009). The Sinai Baton Blue has a short lifespan of between three
and six days on average, while the maximum lifespan for a male recorded was 24 and 18 for
the female (James, 2006d).
The entire population of Sinai Hairstreaks appears to be split across a fragmented
landscape into six smaller populations or habitat patches. These smaller populations almost
certainly interact with each other at some level. The long lifespan of the Sinai Hairstreak
coupled with its ability to travel relatively large distances quickly suggests that it has high
powers of dispersal and could discover and colonise new areas of potential habitat. All six
main sites were occupied by Sinai Hairstreaks but, within these sites there were suitable
trees apparently unoccupied, or at least unvisited. For example a large, healthy tree bearing
fruit was discovered in Abu Towaita, 1 km away from the main site to the northwest (Fig
3.7). No butterflies were discovered on the tree but visitors may have been there on other
days. Conversely, the Rhamnus dispermus trees in Abu Druce were all found in a compact
cluster with the exception of one outlier 1.4 km away to the south (Fig 2.2). This tree was
isolated and yet supported several adult Sinai Hairstreaks. During the MRR butterflies were
observed flying quickly through the wadi between the isolated tree and the main site; in the
event that butterflies failed to lay eggs on the isolated tree in any one year, it is likely that it
would be used again in the future from dispersing butterflies from the main site.
Without further research it is difficult to assign a population structure to the Sinai
Hairstreak. The evidence in this study suggests that the Sinai Hairstreak does not fit the
metapopulation model. A true metapopulation is defined by a number of criteria (Hanski et

38
al., 1995). All habitat patches must be able to support a breeding population. These
populations must be vulnerable to extinction and persist in a stochastic balance between
local extinction and re-colonisation. The dynamics of these populations are asynchronous
which prevents the entire metapopulation becoming extinct. A limited amount of dispersal is
required between populations (Harrison, 1991). There are few empirical examples of true
metapopulations or classic metapopulations (CM) in the field (Fronhofer et al., 2012). A
species with a metapopulation structure is likely to be at the brink of extinction where
habitat fragmentation has isolated sites (Fronhofer et al., 2012). If the sites discovered in this
study represent separate populations, it is unlikely that the smaller sites are incapable of
supporting their own breeding populations (the larger ones almost certainly are capable) and
the risk of extinction between sites is likely to be highly variable. Dispersal between sites is
likely to be high, which is also not consistent with the CM model. Abu Druce, Ahmar and Bab
have the largest populations, presumably with the lowest risk of extinction. The size of Bab
and Ahmar in particular suggest these populations could survive without dispersal from
other sites. Wadi El-Freya, Jebel Katherine and Abu Towaita have much smaller populations,
and if independent would therefore presumably suffer a greater risk of extinction. However,
these sites may be ‘island’ sites that require dispersing individuals from the larger ‘mainland’
sites for continued persistence. Mainland sites are usually much larger than source
populations, but often habitat quality is the same (Harrison, 1991). Ahmar contains 145
trees, while Jebel Katherine, the closest site to it, contains only 33 trees. It is possible that
Ahmar is a source population to Jebel Katherine. A study of butterflies in Venezuela
indicated that some tropical butterfly species exist as a mainland-island population where
small habitat fragments are continuously recolonised from a larger source population
(Shahabuddin and Terborgh, 1999). Similarly a source-sink model may be applicable to
certain sites. A source-sink model is different from that of a mainland-island because the
smaller sites are not of equal quality and are unable to persist on their own except over the
very short term (Nystrand et al., 2009). Wadi El-Freya has the lowest density and number of
trees of any site, and has the smallest butterfly population. This population could be
supplemented by excess of individuals from Abu Druce, which has a far greater population
and a large number of trees.
However, the most likely scenario, based on this evidence, is that the Sinai
Hairstreak does not persist in separate independent populations (as assumed in the previous
paragraph) but as one patchy population. A patchy population is described as a set of habitat
patches linked together by high levels of dispersal that prevent local extinction (Harrison,
1991); we assume such a population is panmictic. Habitat patches are prevented from
extinction by a large rescue effect (Fronhofer et al., 2012). A study of the Karner blue
butterfly Lycaeides melissa samuelis in the United States showed that it occupied all patches
in a network of habitat and was living as a panmictic population (Guiney et al., 2010). The
Sinai Hairstreak is a powerful flier and all the sites are in close proximity to one another. It
could not be established definitively if dispersal is possible between all sites: Bab is four km
from the nearest site. As the most isolated site, it may have low immigration rates compared
to other sites. However, it is likely to be well within reach of the Sinai Hairstreak and thus a
part of a single panmictic population. Incorrect assumptions on the population structure may
lead to the implementation of unnecessary and wasteful conservation strategies so great

39
care must be taken when assigning a population structure to a species (Fronhofer et al.,
2012). Therefore, it is imperative that future studies investigate further the population
structure of the Sinai Hairstreak.
Conservation
The population of Sinai Hairstreaks was estimated to be at least 1,010 individuals spread
across six sites covering an area of roughly 83 km². This is likely to represent a large
proportion, if not all, of its worldwide distribution, since it is as yet unrecorded from any
other location. As with the Sinai Baton Blue, it is possible that populations exist in
northwestern Saudi Arabia. Due to its small population size and the small size of available
habitat, the Sinai Hairstreak is at considerable risk of extinction, and according to IUCN
criteria should be classed as Critically Endangered (CR) purely on this basis.
Short term threats to the Sinai Hairstreak are less evident than those of the Sinai
Baton Blue. The larval food plant is not collected by the Bedouin. All the sites where
Rhamnus dispermus was found growing are isolated and remote. The city of St Katherine is
expanding, but according to local information the number of people living near Rhamnus
dispermus sites is decreasing, and was in any case initially very low. Given the remoteness of
some of the sites there is a very low risk of habitat destruction from human development.
Rhamnus dispermus trees are longer-lived and larger (>2 metres) compared to most plants in
the region and are less likely to be affected by grazing. Few signs of grazing animals were
discovered in the study sites. On a couple of occasions small herds of feral goat (<20
individuals) were observed near Jebel Ahmar and between Abu Druce and Abu Towaita. The
feral goats near Jebel Ahmar were observed feeding on Crataegus x sinaica, a similar-sized
thorny bush often found in the same areas as Rhamnus dispermus. Wild grazing species
appeared to exist in very low numbers. Remains of Rock Hyrax Procavia capensis were
discovered on a couple of occasions in the Ahmar region, and Nubian Ibex Capra nubiana
were observed near Jebel Bab. Their grazing impact on Rhamnus dispermus trees is likely to
be negligible. Adult Sinai Hairstreaks are probably most predated by passerine bird species.
The White-crowned Black Wheatear was observed in Rhamnus dispermus trees frequently
and on a couple of occasions were seen pulling the yellow tags off the tree. The larvae of the
Sinai Hairstreak are green with yellow stripes (Nakamura, 1975). It is possible, therefore, that
the White-crowned Black Wheatear searches for and feeds on Sinai Hairstreak larvae.
Over 80% of the Sinai Hairstreak population was concentrated in the three largest
sites (in terms of population size and the number of trees); Bab, Ahmar and Abu Druce. The
large number of healthy adult trees in close proximity decreases the likelihood of extinction
in these sites, if these represent separate butterfly populations. Sinai Hairstreaks are at
highest risk of extinction in Wadi El-Freya and Abu Towaita, simply because the density of
trees is much lower. Over 50% of the butterflies caught in Abu Towaita were found on only
three trees. The destruction of a small number of trees in these areas could cause a great
reduction in the already meagre populations of Sinai Hairstreaks there, if these are really
separate populations. However this study suggests the Sinai Hairstreak is more likely to exist

40
as a single, panmictic population. The loss of trees in these habitat patches could further
restrict the population size and home range of the Sinai Hairstreak and increase its
vulnerability to extinction.
The most significant threat facing the Sinai Hairstreak is likely to be global warming.
The climate of the South Sinai has been predicted to become hotter and drier (Hulme et al.,
2001), and thus has the potential to disrupt the interaction between the Sinai Hairstreak and
its larval food plant and possibly adult food plants. The existence of a butterfly species is
dependent on the synchronisation of its life-cycle with its larval food plant. For example,
Brown Hairstreak Thecla betulae larvae feed on the developing leaves of its larval food plant
Prunus spinosa. The leaves are only suitable for a short period time and if the larva fails to
hatch at the correct moment they will have insufficient food and die (De Vries et al., 2011).
This study has shown that larval food plant condition is an important determining factor for
the presence of Sinai Hairstreaks. Water is already a limited resource in the St Katherine
Protectorate, and a drier climate would reduce the natural resources available for Rhamnus
dispermus and other plants in the area. The distribution and health of Rhamnus dispermus
trees may decline if global warming does indeed cause the predicted changes in climate. The
number of flowering plants available for adult Sinai Hairstreaks may also be at risk; during a
dry year the number of flowering plants can be reduced by 40% (James, 2006f). As a result of
this decline the availability of suitable habitat for Sinai Hairstreaks would be reduced.
Considering the small amount of potential habitat available to the Sinai Hairstreak, global
warming poses a significant threat to the long-term survival of this species.
The danger posed by global warming could be accelerated by natural water sources
decreasing as a result of increased human demand in St Katherine (Abdulla et al., 2003).
Rapid population increase in St Katherine and the higher levels of tourism in the Sinai in
recent years (Hilmi et al., 2012) may be constraining the water available to the local flora.
This study has confirmed that the Sinai Hairstreak has an extremely small population
size and a greatly restricted home range; it is therefore highly susceptible to extinction. More
information about its biology is clearly needed, and the populations of both Sinai Hairstreak
and Rhamnus dispermus need to be monitored in the future so that we can understand
population trends and changes in habitat. In this study, a large amount of time was invested
in finding Rhamnus dispermus trees and estimating the population size. Future studies
should concentrate on the behaviour and life-cycle of the Sinai Hairstreak. It is imperative
that the small number of Rhamnus dispermus trees remain protected to ensure the survival
of this extremely rare and beautiful butterfly.

41
References
Abdulla, H., Ghodeif, K., El-Shatoury, S. & Dewedar, A. 2003. Potential contamination of groundwater in the World Heritage Site of the St. Katherine Protectorate, Egypt. Egyptian Journal of Biology, 5, 1-9.
Alcock, J. & Oneill, K. M. 1986. Density dependent mating tactics in the Grey hairstreak, Strymon melinus (Lepidoptera: Lycaenidae). Journal of Zoology, 209, 105-113.
Ayyad, M. A., Fakhry, A. M. & Moustafa, A. R. A. 2000. Plant biodiversity in the Saint Catherine area of the Sinai Peninsula, Egypt. Biodiversity and Conservation, 9, 265-281.
Baguette, M. & Schtickzelle, N. 2006. Negative relationship between dispersal distance and demography in butterfly metapopulations. Ecology, 87, 648-654.
Bañuelos, M. J. & Obeso, J. R. 2004. Resource allocation in the dioecious shrub Rhamnus alpinus: the hidden costs of reproduction. Evolutionary Ecology Research, 6, 397-413.
Benyamini, D. 2000. Pseudophilotes jordanicus a new relict species of the SE. Mediterranean (Lepidoptera : Lycaenidae). Linneana Belgica, 17, 359-370.
Birdlife International. 2012. Important Bird Areas factsheet: St Katherine Protectorate [Online]. Available: http://www.birdlife.org.
Boughton, D. A. 2000. The dispersal system of a butterfly: a test of source‐sink theory suggests the intermediate‐scale hypothesis. The American Naturalist, 156, 131-144.
Boulos, L. 2000. Flora of Egypt, vol. 2. Al Ghoneimy, Cairo. Bouzat, J. L., Johnson, J. A., Toepfer, J. E., Simpson, S. A., Esker, T. L. & Westemeier, R. L.
2009. Beyond the beneficial effects of translocations as an effective tool for the genetic restoration of isolated populations. Conservation Genetics, 10, 191-201.
Breed, G. A., Stichter, S. & Crone, E. E. 2012. Climate-driven changes in northeastern US butterfly communities. Nature Climate Change.
Brohan, P., Kennedy, J. J., Harris, I., Tett, S. F. B. & Jones, P. D. 2006. Uncertainty estimates in regional and global observed temperature changes: A new data set from 1850. Journal of Geophysical Research, 111, D12106.
Butterfly Conservation Europe. 2008. Available: http://www.bc-europe.org. Clench, H. K. 1978. The names of certain Holarctic hairstreak genera (Lycaenidae). J. Lepid.
Soc, 32, 277-281. Conradt, L., Bodsworth, E. J., Roper, T. J. & Thomas, C. D. 2000. Non-random dispersal in the
butterfly Maniola jurtina: implications for metapopulation models. Proceedings of the Royal Society of London. Series B: Biological Sciences, 267, 1505-1510.
De Groot, M., Rebeusek, F., Grobelnik, V., Govedic, M., Salamun, A. & Verovnik, R. 2009. Distribution modelling as an approach to the conservation of a threatened alpine endemic butterfly (Lepidoptera: Satyridae). European Journal of Entomology, 106, 77-84.
De Vries, H. H., Ens, S. H., De Graaf, G., Teunissen, L., Van Der Velde, R., Vogelaar, L., Winterink, A. & Visser, M. E. 2011. Synchronisation of egg hatching of brown hairstreak (Thecla betulae) and budburst of blackthorn (Prunus spinosa) in a warmer future. Journal of insect conservation, 15, 311-319.
El-Bana, M., Shaltout, K., Khalafallah, A. & Mosallam, H. 2010. Ecological status of the Mediterranean Juniperus phoenicea L. Relicts in the desert mountains of North Sinai, Egypt. Flora-Morphology, Distribution, Functional Ecology of Plants, 205, 171-178.
Emmel, Т. & Emmel, J. F. 1993. The Xerces Blue, Glaucopsyche xerces (Boisduval). Conservation biology of Lycaenidae (Butterflies). IUCN Species Survival Commission Occ. Pap, 8, 137-138.

42
Fiedler, K. 1991. European and North West African Lycaenidae (Lepidoptera) and their associations with ants. J. Res. Lepid, 28, 57.
Finnegan, L. A., Wilson, P. J., Price, G. N., Lowe, S. J., Patterson, B. R., Fortin, M. J. & Murray, D. L. 2012. The complimentary role of genetic and ecological data in understanding population structure: a case study using moose (Alces alces). European Journal of Wildlife Research, 1-9.
Frankham, R. 2005. Genetics and extinction. Biological Conservation, 126, 131-140. Fronhofer, E. A., Kubisch, A., Hilker, F. M., Hovestadt, T. & Poethke, H. J. 2012. Why are
metapopulations so rare? Ecology, 93, 1967-1978. Gilbert, F., Rashad, S., Kamel, M., Ismail, A. E. D., James, M. & Zalat, S. 2010. Monitoring of
the endemic Sinai Baton Blue butterfly Pseudophilotes sinaicus in the St Katherine Protectorate, South Sinai. Egyptian Journal of Biology, 12.
Gilbert, F. & Zalat, S. 2007. Butterflies of Egypt: atlas, red data listing & conservation, Arab Republic of Egypt.
Gilbert, H. 2011. Environmental Narratives and Conservation Politics: challenging ideas of pastoral destruction in South Sinai.
Grainger, J. 2003. ‘People are living in the park'. Linking biodiversity conservation to community development in the Middle East region: a case study from the Saint Katherine Protectorate, Southern Sinai. Journal of arid environments, 54, 29-38.
Grainger, J. & Gilbert, F. 2008. Around the sacred mountain: the St Katherine Protectorate in South Sinai, Egypt. Protected Landscapes and Cultural and Spiritual Values. Volume 2 in the series Values of Protected Lands-capes and Seascapes, IUCN, GTZ and Obra Social de Caixa Catalunya, 21.
Guiney, M. S., Andow, D. A. & Wilder, T. T. 2010. Metapopulation structure and dynamics of an endangered butterfly. Basic and Applied Ecology, 11, 354-362.
Guschanski, K., Olivieri, G., Funk, S. M. & Radespiel, U. 2007. MtDNA reveals strong genetic differentiation among geographically isolated populations of the golden brown mouse lemur, Microcebus ravelobensis. Conservation Genetics, 8, 809-821.
Hanski, I., Pakkala, T., Kuussaari, M. & Lei, G. 1995. Metapopulation persistence of an endangered butterfly in a fragmented landscape. Oikos, 21-28.
Harrison, S. 1991. Local extinction in a metapopulation context: an empirical evaluation. Biological journal of the Linnean Society, 42, 73-88.
Hilmi, N., Safa, A., Reynaud, S. & Allemand, D. 2012. Coral Reefs and Tourism in Egypt’s Red Sea. Topics in Middle Eastern and African Economies, 14, 416.
Hobbs, J. J. 1992. Bedouin life in the Egyptian wilderness, University of Texas Press. Hoyle, M. & James, M. 2005. Global warming, human population pressure, and viability of
the world's smallest butterfly. Conservation Biology, 19, 1113-1124. Hulme, M., Doherty, R., Ngara, T., New, M. & Lister, D. 2001. African climate change: 1900-
2100. Climate research, 17, 145-168. Issar, A. S., Ginat, H. & Zohar, M. 2011. Shifts from deserted to inhabited terrain in the arid
part of the Middle East, a function of climate changes. Journal of arid environments. James, M. 2006a. Metapopulations and the Sinai Baton Blue (Pseudophilotes sinaicus
Nakamura): an introduction. Egyptian Journal of Biology 8, 7-16. James, M. 2006b. Interactions among species in a tri-trophic system: the influence of ants on
the distribution of the Sinai Baton Blue butterfly Egyptian Journal of Biology, 8, 17-26.
James, M. 2006c. Demographic processes in a local population: seasonal dynamics of the Sinai Baton Blue butterfly. Egyptian Journal of Biology, 8.
James, M. 2006d. Immigration and emigration in the Sinai Baton Blue butterfly: estimates from a single patch. Egyptian Journal of Biology, 8, 39-50.
James, M. 2006e. Intra-patch movement in the Sinai Baton Blue butterfly: influence of micro-habitat and individual characteristics. Egyptian Journal of Biology, 8, 51-66.

43
James, M. 2006f. The natural history of the Sinai Baton Blue: the smallest butterfly in the world. Egyptian Journal of Biology, 8, 67-83.
Joyce, D. A. & Pullin, A. S. 2003. Conservation implications of the distribution of genetic diversity at different scales: a case study using the marsh fritillary butterfly (Euphydryas aurinia). Biological Conservation, 114, 453-461.
Kodandaramaiah, U., Konvicka, M., Tammaru, T., Wahlberg, N. & Gotthard, K. 2012. Phylogeography of the threatened butterfly, the woodland brown Lopinga achine (Nymphalidae: Satyrinae): implications for conservation. Journal of insect conservation, 1-9.
Kotiaho, J. S., Kaitala, V., Komonen, A. & Päivinen, J. 2005. Predicting the risk of extinction from shared ecological characteristics. Proceedings of the National Academy of Sciences of the United States of America, 102, 1963-1967.
Met Office 2011. Climate: Observations, projections and impacts. Nakamura, I. 1975. Descriptions of two new species of butterflies (Lepidoptera, Lycaenidae)
from the South Sinai. Journal of Entomology Series B-Taxonomy & Systematics, 44, 283-295.
Nieberding, C. M., De Vos, H., Schneider, M. V., Lassance, J. M., Estramil, N., Andersson, J., Bång, J., Hedenström, E., Löfstedt, C. & Brakefield, P. M. 2008. The male sex pheromone of the butterfly Bicyclus anynana: towards an evolutionary analysis. PloS one, 3, e2751.
Nystrand, M., Griesser, M., Eggers, S. & Ekman, J. 2009. Habitat‐specific demography and source–sink dynamics in a population of Siberian jays. Journal of Animal Ecology, 79, 266-274.
Opdam, P. & Wascher, D. 2004. Climate change meets habitat fragmentation: linking landscape and biogeographical scale levels in research and conservation. Biological Conservation, 117, 285-297.
Pellet, J. 2008. Seasonal variation in detectability of butterflies surveyed with Pollard walks. Journal of insect conservation, 12, 155-162.
Pierce, N. E., Braby, M. F., Heath, A., Lohman, D. J., Mathew, J., Rand, D. B. & Travassos, M. A. 2002. The ecology and evolution of ant association in the Lycaenidae (Lepidoptera). Annual review of entomology, 47, 733-771.
Pollard, E. 1977. Method for assesing changes in abundance of butterflies. Biological Conservation, 12, 115-134.
Prokopy, R. J. & Owens, E. D. 1983. Visual detection of plants by herbivorous insects. Annual review of entomology, 28, 337-364.
Reemer, M., Van Helsdingen, P. J. & Kleukers, R. M. J. C. 2003. Changes in ranges. Saastamoinen, M., Ikonen, S. & Hanski, I. 2009. Significant effects of Pgi genotype and body
reserves on lifespan in the Glanville fritillary butterfly. Proceedings of the Royal Society B: Biological Sciences, 276, 1313-1322.
Schmitt, T., Habel, J. C., Besold, J., Becker, T., Johnen, L., Knolle, M., Rzepecki, A., Schultze, J. & Zapp, A. 2006. The Chalk-hill Blue Polyommatus coridon (Lycaenidae, Lepidoptera) in a highly fragmented landscape: How sedentary is a sedentary butterfly? Journal of insect conservation, 10, 311-316.
Schtickzelle, N., Choutt, J., Goffart, P., Fichefet, V. & Baguette, M. 2005. Metapopulation dynamics and conservation of the marsh fritillary butterfly: population viability analysis and management options for a critically endangered species in Western Europe. Biological Conservation, 126, 569-581.
Settle, J., Kudrna, O., Harpke, A., Kühn, I., Van Swaay, C., Verovnik, R., Warren, M., Wiemers, M., Hanspach, J. & Hickler, T. 2010. Climatic risk atlas of European butterflies.
Shahabuddin, G. & Terborgh, J. W. 1999. Frugivorous butterflies in Venezuelan forest fragments: abundance, diversity and the effects of isolation. Journal of Tropical Ecology, 15, 703-722.

44
Strausz, M., Fiedler, K., Franzén, M. & Wiemers, M. 2012. Habitat and host plant use of the Large Copper Butterfly Lycaena dispar in an urban environment. Journal of insect conservation, 1-13.
Sutherland, W. J. 2009. Managing habitats and species. Conservation science and action, 202-219.
Thomas, C. D., Cameron, A., Green, R. E., Bakkenes, M., Beaumont, L. J., Collingham, Y. C., Erasmus, B. F. N., De Siqueira, M. F., Grainger, A. & Hannah, L. 2004. Extinction risk from climate change. Nature, 427, 145-148.
Tshikolovets, V. V. 2011. Butterflies of Europe & the Mediterranean area, Pardubice, Czech Republic, Tshikolovets Publications.
Tuomisto, H. 2010. A consistent terminology for quantifying species diversity? Yes, it does exist. Oecologia, 164, 853-860.
Unesco. 2002. Nomination of the St Catherine area, Sinai, Egypt as a mixed world cultural and natural heritage site [Online]. Available: http://whc.unesco.org/en/list/954/documents/.
Vande Velde, L., Turlure, C. & Van Dyck, H. 2011. Body temperature and territory selection by males of the speckled wood butterfly (Pararge aegeria): what makes a forest sunlit patch a rendezvous site? Ecological Entomology, 36, 161-169.
Wahlberg, N., Klemetti, T. & Hanski, I. 2002. Dynamic populations in a dynamic landscape: the metapopulation structure of the marsh fritillary butterfly. Ecography, 25, 224-232.
Weidenhoffer, Z., Bozano, G. C. & Churkin, S. 2004. Guide to the Butterflies of the Palearctic Region. Lycaenidae part II, Omnes Artes, 94p.
Wheat, C. W., Fescemyer, H. W., Kvist, J., Tas, E. V. A., Vera, J. C., Frilander, M. J., Hanski, I. & Marden, J. H. 2011. Functional genomics of life history variation in a butterfly metapopulation. Molecular ecology, 20, 1813-1828.
Wilson, R. J., Gutierrez, D., Gutierrez, J. & Monserrat, V. J. 2007. An elevational shift in butterfly species richness and composition accompanying recent climate change. Global Change Biology, 13, 1873-1887.
Yu, L. & Lu, J. 2011. Does landscape fragmentation influence sex ratio of dioecious plants? A case study of Pistacia chinensis in the Thousand-Island lake region of China. PloS one, 6, e22903.
Zahran, M. A. & Willis, A. J. 2009. The Sinai Peninsula. The Vegetation of Egypt, 213-249. Zalat, S., Gilbert, F. & Gheith, A. 2008. Gardens of a sacred landscape: Bedouin heritage and
natural history in the high mountains of Sinai, American University in Cairo Press. Zhu, H., Gegear, R. J., Casselman, A., Kanginakudru, S. & Reppert, S. M. 2009. Defining
behavioral and molecular differences between summer and migratory monarch butterflies. BMC biology, 7, 14.

45
Appendix I - Odonata of St Katherine
Wandering Glider Pantala flavescens
Fox Camp, St Katherine 25/07/2012; male Photograph – Andrew Power
Broad Scarlet Crocothemis erythraea
Wadi Matter 14/07/2012; male Wadi Ahmar; 14/07/2012; female Photograph 1 – Andrew Power,
male
Photograph 2 – Andrew Power,
female
Vagrant Emperor Anax ephippiger
Wadi Ahmar 14/07/2012 Photograph – Andrew Power
Lesser Emperor Anax parthenope
Fox Camp St Katherine 23/06/2012 Photograph – Andrew Power

46
Red-veined Dropwing Trithemis arteriosa
Wadi Jebel 18/04/2012; male Abu Teenya 25/06/2012; female Photograph 1 – Andrew Power,
male
Photograph 2 – female, Andrew
Power, female
Red-veined Darter Sympetrum fonscolombei
Jebel Bab 9/08/2012, female Photograph – Andrew Power
Southern Skimmer Orthetrum brunneum
Wadi Matter 14/07/2012 Photograph – Andrew Power
Only 2 previous records from Sinai - Dumont, H. J. 1991. Odonata of the Levant, Israel academy of sciences and humanities.
Desert Skimmer, Ransonnet's Skimmer Orthetrum ransonnetii
Fox Camp, St Katherine 2/08/2012; female Photograph – Andrew Power
Evan’s Bluetail Ischnura evansi
Wadi Ahmar 14/07/2012; male and female Photograph 1 – Andrew Power,
male

47
Appendix II - Birds of St Katherine
Chukar Alectoris chukar
Farsh Elia 21/04/2012; 1
Between Wadi Albein & Jebel Ahmar 22/04/2012
Jebel Ahmar 8/05/2012; 20 plus
Jebel Katherine 5/06/2012; female
Wadi Ahmar 12/06/2012
Abu Druce 17/06/2012
Abu Druce 18/06/2012
Abu Druce 19/06/2012
Abu Druce 20/06/2012
Sagare 27/06/2012
Sagare 28/06/2012
Ain Nigela 7/07/2012
Wadi Ahmar 27/07/2012
Wadi Ahmar 28/07/2012
Wadi Ahmar 29/07/2012
Wadi Ahmar 6/08/2012 Photograph – Andrew Power, Farsh Elia
Sand Partridge Ammoperdix heyi Wadi El-Freya 10/05/2012; 3
Wadi Jebel 13/05/2012; 4
Abu Druce 20/06/2012
Ain Nigela 8/07/2012; 2
Ain Nigela 10/07/2012
Sparrowhawk Accipter nisus
On route to Wadi Jebel, Wadi Jebel 18/04/2012; both females
Fox Camp 30/04/2012
Wadi Jebel 16/05/2012
Abu Druce 18/06/2012 Photograph – Andrew Power, Wadi Jebel
Kestrel Falco tinnunculus
Fox Camp 12/05/2012; hunting
Foot of Jebel Katherine/Wadi Albein 23/05/2012; attacking Buzzard species
Fox Camp 27/05/2012
Abu Druce 17/06/2012
Wadi Ahmar 29/07/2012 Photograph – Andrew Power, Fox Camp

48
Bonelli’s Eagle Aquila fasciata
Fox Camp 17/06/2012 Photograph – Andrew Power
Long-legged Buzzard Buteo rufinus
Wadi Jebel 18/04/2012 Photograph – Andrew Power Part of migration of 60+ Buzzards in Wadi Jebel including Long-legged Buzzard Buteo rufinus, Steppe Buzzard Buteo buteo vulpinus & Honey Buzzard Pernis apivorus
Steppe Buzzard Buteo buteo vulpinus
Wadi Jebel 18/04/2012
Wadi Jebel 16/05/2012
Photograph – Andrew Power
Part of migration of 60+ Buzzards in Wadi Jebel including Long-legged Buzzard Buteo rufinus, Steppe Buzzard Buteo buteo vulpinus & Honey Buzzard Pernis apivorus
Honey Buzzard Pernis apivorus
Wadi Jebel 18/04/2012 Photograph – Andrew Power
Part of migration of 60+ Buzzards in Wadi Jebel including Long-legged Buzzard Buteo rufinus, Steppe Buzzard Buteo buteo vulpinus & Honey Buzzard Pernis apivorus
Laughing Dove Streptopelia senegalensis
Fox Camp 17/04/2012
Very common in city of St Katherine
Fox Camp 7/06/2012; fledgling
Photograph – Andrew Power; Fox camp

49
Rock Dove Columba livia
St Katherine; common - Feral pigeon;
Fox Camp 17/04/2012 (present everyday)
Photograph – Andrew Power; Fox Camp
Collared Dove Streptopelia decaocto
Wadi Albein 6/06/2012
St Katherine Monastery 9/06/2012
Turtle Dove Streptopelia turtur Fox Camp 7/06/2012; 3
Fox Camp 16/06/2012
St Katherine Monastery 26/07/2012
Hoopoe Upupa epops St Katherine 18/04/2012; 1 in flight near petrol station
Bee-eater Merops apiaster
Fox camp 20/04/2012; 5 flying over
Wadi Cla 23& 25/04/2012
Wadi El-Freya 10 /05/2012
Wadi El-Freya 15/05/2012
Wadi Jebel 16/05/2012
Photograph- Andrew Power, Fox Camp
Golden Oriole Oriolus oriolus
Fox Camp 27/05/2012
Fox Camp 28/05/2012
Fox Camp 29/05/2012
Fox Camp 30/05/2012
Wadi El-Freya 31/05/2012
Photograph – Andrew Power
White-spectacled Bulbul Pycnonotus xanthopygos
Abu Towaita 25/06/2012
Abu Towaita 26/06/2012
Sagare 27/06/2012
Sagare near garden of Shop 28/06/2012
Wadi Cla 28/06/2012; 2
Photograph 1 – Andrew Power, Sagare

50
Palestine Sunbird Cinnyris osea
Fox Camp 17 & 20/04/2012; male & female
On way to Wadi Ahmar 8/05/2012; 2 males
Wadi Jebel 16/05/2012
Abu Towaita 21/05/2012; 4
Jebel Katherine 24/05/2012
Fox Camp 1/06/2012
Jebel Katherine 2/06/2012
Wadi Ahmar 12/06/2012
Abu Teenya 17/06/2012
Abu Teenya 19/06/2012
Abu Teenya 20/06/2012
Wadi Jebel 24/06/2012
Abu Towaita 26/06/2012
Sagare 28/06/2012
Ain Nigela 7/07/2012
Ain Nigela 8/07/2012
Wadi Ahmar 14/07/2012; 5 plus
Wadi Ahmar 27/07/2012
Wadi Ahmar 28/07/2012
Wadi Ahmar 29/07/2012
Wadi Ahmar 6/08/2012 Photograph 1 – Brian Power; female in Wadi Ahmar Photograph 2 – Andrew Power; male in Abu Towaita
Tristram’s Starling Onychognathus tristramii
Fox Camp 17/04/2012; 15-20
On route to Wadi Jebel 18/04/2012; 20-30 high on mountain
Fox Camp 1/05/2012; 20 plus
Fox Camp 11/05/2012; 2
Fox Camp 14/05/2012
Wadi El-Freya 15/05/2012
Wadi Jebel 16/05/2012
Fox Camp 28/05/2012; 15 plus
Jebel Ahmar 29/05/2012
Jebel Katherine 4/05/2012; 2
Wadi Albein 8/06/2012
Wadi Ahmar 12/06/2012
Abu Druce 19/06/2012; 5 plus
Abu Druce 20/06/2012
St Katherine Monastery 26/07/2012 Photograph 1 – Andrew Power; Fox Camp Photograph 2 – Andrew Power, Fox Camp

51
Wheatear Oenanthe oenanthe
Fox Camp 20/04/2010; male Photograph 1 – Andrew Power, Fox Camp Photograph 2 – Andrew Power, near Hotel Raha St Katherine
White-crowned Black Wheatear Oenanthe leucopyga
Fox Camp, St Katherine 17/04/2012
- very common in St Katherine Photograph – Andrew Power; adult and chick in Abu Towaita
Hooded Wheatear Oenanthe monacha
Jebel Katherine 3/06/2012
Jebel Katherine 4/06/2012
Wadi Ahmar 8/06/2012; male
Wadi Ahmar 12/06/2012
Abu Druce 20/06/2012; female
Abu Towaita 27/06/2012; male
Wadi Jebel near Hagbut Nada 7/07/2012; male
Ain Nigela 7/07/2012; male and female
Ain Nigela 8/07/2012
Ain Nigela 10/07/2012
Ain Nigela 11/07/2012
Wadi Ahmar 27/07/2012; 2 male and 1 female
Wadi Ahmar 28/07/2012
Wadi Ahmar 29/07/2012
Wadi Ahmar 6/08/2012 Photograph 1 – Andrew Power, male in Abu Towaita Photograph 2 – Andrew Power, female in Jebel Katherine
Rock Thrush Monticola saxatilis
St Katherine 17/04/2012; male
Between Fox camp and St Katherine 18/04/2012; male
Photograph – Andrew Power, near Hotel Raha St Katherine

52
Whinchat Saxicola rubetra
Wadi El-Freya 10/05/2012 Photograph – Andrew Power
House Martin Delichon urbicum 19/04/2012; 3 flying with Rock Martins
Fox Camp 1/05/2012
Wadi Jebel 16/05/2012
Rock Martin Ptyonoprogne fuligula
Very common in St Katherine Photograph 1 – Andrew Power, St Katherine Photograph 2 – Brian Power, Wadi Ahmar
Swallow Hirundo rustica
Dahab 27/04/2012
Dump outside St Katherine 11/05/2012
Fox Camp 12/05/2012
Wadi Jebel 16/05/2012 Photograph – Olivia Norfolk, injured Swallow in Wadi Jebel
Red-rumped Swallow Cecropis daurica Fox Camp 3/05/2012; juvenile
Swift Apus apus Petrol Station St Katherine 6/05/2012
Abu Druce 18/06/2012; 10 plus Wood Warbler Phylloscopus sibilatrix
Fox Camp 20/04/2012
Fox Camp 3/05/2012
Abu Teenya 17/06/2012 Photograph – Andrew Power, Fox Camp
Garden Warbler Sylvia boring
Fox Camp 27/05/2012
Fox Camp 28/05/2012
Fox Camp 7/06/2012; feeding on apricot
Fox Camp 9/06/2012 Photograph – Andrew Power

53
Blackcap Sylvia atricapilla
Fox Camp 12/05/2102
Fox Camp 27/05/2012; male and female
Fox Camp 28/05/2012; male and female
Jebel Ahmar 29/05/2012; female
Fox Camp; female
Photograph 1 – Andrew Power, male feeding on apricot tree in Fox Camp garden Photograph 2 – Andrew Power, female in Fox Camp garden
Eastern Olivaceous Warbler Iduna pallida
Fox Camp 20/04/2012
Photograph – Andrew Power
Eastern Bonelli’s Warbler Phylloscopus orientalis
Wadi Matter 28/07.2012
Photograph – Brian Power, Wadi Matter
Eastern Orphean Warbler Sylvia crassirostris
Wadi Matter 28/07/2012; up to 5 individuals Photograph – Brian Power, Wadi Matter
Lesser Whitethroat Sylvia curraca
Fox Camp 27/05/2012 Photograph – Andrew Power

54
Scrub Warbler Scotocerca inquieta
Wadi Sherage 21/04/2012
Abu Druce 25/04/2012
Wadi Albein, Abu Himmen 12/05/2012
Wadi El-Freya 31/05/2012, 10 plus
Abu Teenya 17/06/2012
Abu Druce 19/06/2012; 5 plus
Abu Druce 20/06/2012
Abu Towaita 26/06/2012
Sagare 27/06/2012; 10 plus
Abu Towaita 27/06/2012 Photograph 1 – Andrew Power, Abu Druce Photograph 2 – Andrew Power, Abu Towaita
Semi-Collared Flycatcher Ficedula semitorquata
Fox Camp 20/04/2010; pair
Fox Camp 28/04/2012
Fox Camp 7/04/2012
Fox Camp 14/05/2012 Photograph 1; Andrew Power; Fox Camp, male Photograph 1; Andrew Power; Fox Camp, female
Spotted Flycatcher Muscicapa striata
Fox Camp 3/05/2012
Fox Camp 11/05/2012
Fox Camp 12/05/2012
Fox Camp 14/05/2012
Fox Camp 15/05/2012
Fox Camp 18/05/2012
Fox Camp 19/05/2012
Fox Camp 20/05/2012 Photograph – Andrew Power
Masked Shrike Lanius nubicus
Petrol Station St Katherine 6/05/2012
Fox Camp 27/05/2012;2
Fox Camp 28/05/2012
Fox Camp 29/05/2012
Fox Camp 30/05/2012 Photograph – Andrew Power, Fox Camp

55
Isabelline Shrike Lanius isabellinus
Fox Camp 7/05/2012
Fox Camp 8/05/2012 Photograph – Andrew Power
Ortolan Bunting Emberiza hortulana Wadi Ahmar 8/06/2012; 2
Black-headed Bunting Emberiza melanocephala
Abu Towaita 21/05/2012; male and female
Photograph 1 – Andrew Power, male Photograph 2 – Andrew Power, female
Yellow Wagtail Motacilla flava
Wadi El-Freya 10/05/2012; feeding around opium garden
Photograph – Andrew Power
House Sparrow Passer domesticus
Dump outside St Katherine 11/05/2012; probably nesting in adjacent building; 5
Fox Camp 6/06/2012; 5
Fox Camp 7/06/2012; 4
Fox Camp 12/07/2012
Fox Camp 13/07/2012 Photograph – Andrew Power, dump outside St Katherine
Redstart Phoenicurus phoenicurus
Fox Camp 20/04/2012; male
Fox Camp 2/05/2012; female
Fox Camp 3/05/2012; female
Fox Camp 12/05/2012; male and female
Fox Camp 15/05/2012; female Photograph – Andrew Power

56
Sinai Rosefinch Carpodacus synoicus
Between Fox camp and St Katherine 18/04/2012; 2 pairs
Mount Sinai 21/04/2012
Between Wadi Albein & Jebel Ahmar 22/04/2012
Abu Druce 25/04/2012
Hotel Raha 30/04/2012; 1 male
Wadi Jebel 16/05/2012; 5 drinking
Abu Towaita 21/05/2012
Wadi Albein 29/05/2012; 4
Jebel Ahmar 29/05/2012; 2
Jebel Katherine 3/06/2012
Jebel Katherine 5/06/2012; 5
Wadi Albein 8/06/2012
Wadi Ahmar; 12/05/2012
Abu Druce 17/06/2012
Abu Teenya 17/06/2012
Abu Druce 18/06/2012
Abu Druce 19/06/2012; 5 plus
Abu Druce 20/06/2012; 10 plus
Abu Towaita 28/06/2012
Ain Nigela 7/07/2012
St Katherine Monastery 26/07/2012
Wadi Ahmar 6/08/2012 Photograph 1 – Andrew Power, St Katherine Photograph 2 – Andrew Power, Wadi Albein
Desert Lark Ammonmanes deserti
Wadi Jebel 18/04/2012; 2 pairs
Between Fox Camp & St Katherine 18 & 20/04/2012
Elia 21/04/2012
Wadi Raha 24/04/2012
Wadi El-Freya near opium field 10/05/2012
Abu Towaita 21/05/2012
Jebel Katherine 23/05/2012
Sagare 27/06/2012 Photograph – Andrew Power, Abu Towaita
Tree Pipit Anthus trivialis
St Katherine 19/04/2012; trapped in greenhouse
Fox camp garden 20/04/2010 Photograph 1 - Katy Thompson, St Katherine Photograph 1 - Andrew Power, Fox Camp

57