conformational stability of human erythrocyte transglutaminase
TRANSCRIPT

Eur. J. Biochem. 266, 575±582 (1999) q FEBS 1999
Conformational stability of human erythrocyte transglutaminasePatterns of thermal unfolding at acid and alkaline pH
Carlo M. Bergamini1, Mariangela Dean1, Gabriella Matteucci1, Stefania Hanau1, Fabio Tanfani2, Carlo Ferrari2,Marisa Boggian3 and Angelo Scatturin3
1Department of Biochemistry and Molecular Biology, University of Ferrara, Italy; 2Institute of Biochemistry, University of Ancona, Italy;3Department of Pharmaceutical Sciences, University of Ferrara, Italy
Tissue-type transglutaminase is irreversibly inactivated during heat treatment. The rate of inactivation is low at
pH 7.5; it increases slightly at acid pH (6.1) but much more at alkaline pH (9.0±9.5), suggesting that specific
effects take place in the alkaline range, possibly in relation to decreased stability of the transition-state
intermediate as pH is raised above 9.0. Differential scanning calorimetry experiments indicate that thermal
unfolding of the protein occurs with two separate transitions, involving independent regions of the enzyme. They
are assigned to domains 1 and 2 and domains 3 and 4, respectively, by a combination of calorimetric and
spectroscopic techniques. When considering the effects of pH, we noted that transglutaminase was unfolded via
different pathways at the different pH values considered. At acid pH, the whole structure of the protein was lost
irreversibly, with massive aggregation. At neutral and, even more so, at alkaline pH, aggregation was absent (or
very limited at high protein concentration) and the loss of secondary structure was dependent on the ionization
state of crucial lysine residues. Unfolding at pH 9.5 apparently chiefly involved the N-terminal region, as testified
by changes in protein intrinsic fluorescence. In addition, the C-terminal region was destabilized at each pH value
tested during thermal unfolding, as shown by digestion with V8 proteinase, which is inactive on the native
protein. Evidence was obtained that the N-terminal and C-terminal regions interact with each other in
determining the structure of the native protein.
Keywords: conformational stability; heat treatment; pH; thermal unfolding; transglutaminase.
Tranglutaminases catalyze calcium-dependent acyltransferreactions between glutaminyl residues in peptide chains andprimary amines by formation of glutamyl isopeptide bonds witheither low-molecular-mass free amines or amino groups oflysines in peptide chains [1]. They are involved in severalimportant processes, including wound healing and bloodclotting, terminal differentiation of cells [2] and programmedcell death [3]. They are also finding increasing applications asuseful tools for preparing protein gels of high resistance totensile strength and proteolytic breakdown, for instance in foodtechnology [4].
The structure of the main transglutaminase isoform, thetissue-type isoenzyme, was elucidated recently by a combina-tion of protein modeling and biochemical techniques. Thesestudies demonstrated that, beside its transamidating activity,tissue-type transglutaminase also has a calcium-independentGTPase activity, acting as a G-like protein in signal trans-duction [5]. This bifunctional role of the protein is related to aconformational shift between calcium-stabilized and GTP-stabilized states. Protein modeling and biochemical studieshave revealed, in great detail, the structure of tissue
transglutaminase, indicating that the GTP-stabilized state isquite similar to the ligand-free enzyme [6,7]. This wealth ofinformation on the structure of transglutaminase is removedfrom that on protein folding and stability to denaturation [8,9],with the exception of factor XIII, the structural perturbations ofwhich during guanidine [10] and thermal treatment have beenexplored [11,12]. This information on tissue transglutaminasecould be used in studies on protein folding and for itsapplication to biotechnological projects.
The importance of this issue is underlined by recent studieson the pathogenesis of celiac disease [13], an autoimmunedisease that leads to severe atrophy of the intestinal mucosa.The pathogenetic picture assumes a double role for the enzyme,acting initially as a glutaminase on selected glutamine residuesof the alimentary gluten protein gliadin and later cross-linkingitself to the modified gliadin, forming protein complexes,recognized as non-self by the immune system. This raises asevere IgA-mediated autoimmune reaction against the intestinalmucosa, the intensity of which correlates clinically with thetiter of the specific anti-transglutaminase antibodies [14]. Theimmune process does not appreciably affect tissues other thanthe intestinal mucosa, suggesting direct involvement of themodified, possibly unfolded, transglutaminase.
A basic knowledge of the pattern of folding±unfolding oftransglutaminase would help us to understand its involvementin this autoimmune pathology as well as elucidate mechanismsfor its biotechnological application. For these reasons, we haveinvestigated the unfolding of transglutaminase, evaluating thecombined effects of pH and heat treatment on the confor-mational stability of the enzyme in solution.
Correspondence to C. M. Bergamini, Department of Biochemistry
and Molecular Biology, University of Ferrara, Via L. Borsari 46,
44100 Ferrara, Italy. Fax: + 39 0532 202723, Tel.: + 39 0532 291425,
E-mail: [email protected]
Abbreviations: DSC, differential scanning calorimetry; FT-IR, Fourier
transform infrared.
Enzyme: transglutaminase or protein-glutamine g-glutamyltransferase
(EC 2.3.2.13).
(Received 1 June 1999, accepted 27 September 1999)

576 C. M. Bergamini et al. (Eur. J. Biochem. 266) q FEBS 1999
M A T E R I A L S A N D M E T H O D S
Type 2 transglutaminase was purified from human erythrocytesas previously reported [7]. The purified protein displayed highspecific activity (5 mmol´min21´mg21) and was devoid ofminor contaminants that are occasionally present in the enzymepurified by the original procedure. Activity was measured asthe calcium-dependent incorporation of radioactive putrescineinto dimethylcasein at 30 8C and pH 7.5, by the filter paperassay of Lorand, as previously reported [15].
Heat inactivation was carried out by incubating the enzyme(0.1±0.2 mg´mL21) at fixed temperatures and removingsamples at timed intervals for activity assay. During the heattreatment, the enzyme diluents used to obtain the final pHreported in the text were acetate, Tris or borate buffer (each at80 mm concentration). Differential scanning calorimetry (DSC)experiments, at defined pH values, were performed with acomputer-controlled Microcal VP-DSC microcalorimeter at ascanning rate of 1±1.5 8C´min21, from 20 to 85 8C. Themeasuring cell contained the enzyme at a concentration of0.2±0.4 mg´mL21 at the required pH. Further details areavailable in the figure legends. The experimental data weredeconvoluted with the Origin software, provided by theproducer.
Fluorescence measurements were carried out on a Perkin-Elmer MPF3L spectrofluorimeter connected to an externalrecirculatory bath. Samples of enzyme (0.05±0.15 mg´mL21)and solvent were placed in the fluorimetric cuvettes, with athermocouple placed inside the reference cell to record thetemperature within the compartment. Tryptophan emission wasrecorded continuously, in the ratio mode, at 330 and 350 nmupon excitation at 295 nm. The ratio of emission intensities atthese wavelengths was calculated from the recorder traces afterbaseline subtraction. For turbidometric experiments, wave-lengths of excitation and emission were both fixed at 420 nm.
Fourier transform infrared (FT-IR) spectra were recordedusing a Perkin-Elmer 1760-x FT-IR spectrometer; details oninstrumentation, sample preparation, measurements, watersubtraction, spectra deconvolution and analyses are availablein previous studies from our laboratories [16,17]. Thedenaturation of the soluble enzyme in 80 mm sodium acetateor 80 mm sodium borate in deuterium oxide, buffered at pD 6.1or 9.5, respectively, was followed by recording FT-IR spectra at5 8C intervals, from 20 through to 90 8C, while heating insidethe spectrometer cell. The concentration of the enzyme was30±40 mg´mL21. We also analyzed the transglutaminaseprecipitates formed during heating at 50 or 40 8C, at pH 6.1or 9.5, respectively. The precipitates were washed several timeswith 50 mm Tris/HCl, pD 7.0, and analyzed at 20 8C.
Far-UV CD spectra were recorded at different temperaturesusing a Jasco model 500A automatic recording circulardichrograph, interfaced (IF-500 II) with an IBM AT computerequipped with an external water-recirculating bath and athermocouple to record the temperature in the assay compart-ment. A cylindrical fused quartz cell of 0.02 cm pathlengthwas used for CD measurements. The usual precautions weretaken to avoid artefacts. Spectra, corrected by subtraction ofbuffer baseline, are given in mean residue ellipticity(degrees´cm2´dmol21). Measurements were performed at aprotein concentration of 0.3 mg´mL21 after prolonged dialysisagainst buffer prepared with Fluka Microselect salts.
In proteolysis studies, the protein was incubated at therequired temperature and pH for increasing periods of timein the presence of V8 proteinase, at a substrate/proteinaseratio of 100 : 1 (w/w). Samples were removed and subjected
to SDS/PAGE [18] (after quenching the proteinase activity with2 mm phenylmethanesulfonyl fluoride), with protein staining.Protein bands were also blotted on to poly(vinylidenedifluoride) membranes with Caps buffer, pH 10, and sequencedusing an Applied Biosystems 470 sequencer, operated accord-ing to the supplier's instructions.
R E S U LT S
Inactivation of transglutaminase during heat treatment atdifferent pH values
The activity of transglutaminase (0.1±0.2 mg´mL21) was lostin a time-dependent way, with apparent first-order kinetics,when the enzyme was subjected to heat treatment even atrelatively mild temperatures (30±40 8C). The rate of inacti-vation at 37 8C (Fig. 1) was low at pH 7.5 (0.006 min21),increased slightly at acid pH 6.1±6.5 (0.008 min21) butincreased very sharply at pH 9.2±9.5 (0.092 min21), at whichpH complete inactivation was observed within a few minutes at40 8C. All attempts to recover activity by incubating theenzyme at 4 8C with appropriate ligands for increasing periodsof time before assay, or to reverse fluorescence signals bydecreasing temperature (see below), were unsuccessful, sug-gesting irreversible thermal inactivation. In the Arrhenius plot(Fig. 2), the data at pH 6.1 and 7.5 yielded two near-parallelstraight lines, from the slopes of which we calculated anenthalpy value of 170 kJ´mol21. In contrast, data at pH 9.5fitted a biphasic plot with a break around 32 8C; the slope atlow temperature was identical with that at pH 6.1 and 7.5,while it was higher at temperatures above 32 8C, with acalculated enthalpy value of about 126 kJ´mol21, suggestinginvolvement of intermediates of different stability in thethermal inactivation at pH 9.5 at temperatures above 32 8C.
Differential scanning calorimetry
These effects were further investigated by DSC. Progressiveheating at pH 7.5 brought about protein unfolding with twodistinct transitions, with tm centred at about 51 and 60 8C,respectively, suggesting that two distinct regions of the proteinunderwent separate unfolding with potentially different effects.During the second transition, at the protein concentrationsemployed for DSC (but even more markedly at the con-centrations employed in FT-IR spectroscopy), the enzyme
Fig. 1. Semilogarithmic plot of time course of inactivation of erythro-
cyte transglutaminase (0.16 mg´mL21) during heating at different
temperatures at pH 6.1, 7.5 and 9.5 (A, B and C, respectively). Numbers
close to the experimental lines denote temperatures of preincubation.

q FEBS 1999 Thermal unfolding of tissue transglutaminase (Eur. J. Biochem. 266) 577
underwent rapid aggregation and appreciable precipitation, asmanifested by a large exothermic heat exchange. The markedinsolubilization of the intact protein during heating madethermodynamic treatment of the data and calculation ofenthalpy and entropy values questionable, also because ofuncertainty about the baseline assignment. This last point wastentatively solved by running fusion experiments at pH 3.5 in aglycine buffer, conditions under which the protein does notaggregate, assuming that the DCp values for the native/denatured transition were independent of the experimentalconditions as usual. With these assumptions, we were able toperform curve deconvolution (Fig. 3). The middle trace inFig. 3 refers to DSC at pH 7.5, with the distinct transitionsreferred to above. We assigned them, respectively, to theN-terminal domains 1 and 2 and to the C-terminal domains 3and 4 by comparing DSC melting temperatures with thosederived from tryptophan intrinsic fluorescence measurements(see below). When we considered the effects of pH on thethermal fusion of transglutaminase by DSC, we noted that thefirst transition was very sensitive to alkaline pH, as recognizedby the downshift in the fusion temperature, which wasrecognizable despite the large decline in the heat capacity ofunfolding. At pH 9.5 (Fig. 3, lower trace), the tm of the firsttransition decreased to 41 8C while it was essentiallyunmodified in the acid pH range. Conversely, we observedmajor differences in the second transition at acid pH. In fact, atpH 6.1, the second transition was shifted to lower temperatures,with tm at 52 8C, resulting in it almost being fused with the firsttransition. Under these conditions, marked precipitationoccurred, even at a protein concentration as low as0.04 mg´mL21, and was detectable by light-scattering measure-ments, continuously recording turbidity during the heating of
protein samples within a fluorimeter cell, while it was notdetected at pH 7.5 or 9.5 (Fig. 4). The tm value at pH 6.1obtained by light scattering was 54 8C, i.e. about 4 8C higherthan that of the first transition and close to the calorimetricvalue of the tm for the second transition. The calculatedcalorimetric DH values were < 850, 880 and 280 for the firsttransition and 290, 150 and 140 kJ´mol21, for the secondtransition, at pH 6.1, 7.5 and 9.5, respectively.
Fluorescence studies of protein unfolding
The tryptophan intrinsic fluorescence spectrum of nativetransglutaminase is highly blue-shifted, indicating an apolarenvironment of tryptophan residues, with emission maximum at333 nm and a ratio of fluorescence intensities at 350 and330 nm of 0.76. Upon thermal denaturation at pH 7.5 (byheating up to 65 8C at a constant rate of 1 8C´min21), the ratioof fluorescence emission at these wavelengths increasedprogressively to reach a value of 0.95 (Fig. 5), generatingmelting profiles characterized by co-operative transitionsbetween signals of native and unfolded protein. Concomitantly,the maximal emission was shifted only slightly to a wavelengthof 335 nm. The approximate tm for this transition, whichdepends on the spectral properties of tryptophan residues, i.e. ofdomains 1 and 2 [7], was 50 8C, in agreement with the value ofthe first DSC transition. During heating, the enzyme also lostactivity, with an inactivation profile that could be superimposed
Fig. 2. Arrhenius plot of ln(apparent rate constant) for inactivation
against the reciprocal absolute temperature. Values for apparent rate
constants of inactivation were calculated from semilogarithmic plots of the
time course of inactivation as in Fig. 1.
Fig. 3. DSC of transglutaminase. The protein was scanned between 20
and 90 8C at a rate of 1 8C´min21 and a protein concentration of
0.25 mg´mL21. The traces have been shifted on the vertical axis for
clarity: top, pH 6.1; middle, pH 7.5; bottom, pH 9.5. The data are presented
after baseline subtraction and normalization for protein concentration. Note
that at each pH considered, thicker traces identify the normalized scans and
that thin lines indicate deconvoluted peaks. The vertical ruler close to the
ordinate axis indicates DCp of 41.9 kJ´mol21. For further details see the
text.

578 C. M. Bergamini et al. (Eur. J. Biochem. 266) q FEBS 1999
perfectly on that derived from the fluorescence measurements(data not shown), suggesting a close correlation betweeninactivation and unfolding of the N-terminal region, containingdomains 1 and 2 and all the tryptophan fluorophores of theprotein.
The calculated tm values of the fluorescence titration weredependent on pH, decreasing from 50 8C at pH between 6.1 and8.0, down to 41 8C, as pH was increased to over 9.0 (Fig. 6).The apparent pK was 9.0, suggesting a relevant role for lysineresidues in the pH-dependence of the thermal effects. Accord-ing to the spectral properties, the unfolded protein assumeddifferent conformations after heating at pH 6.1 or at pH 9.5, asthe emission spectrum (lmax 333 nm in the native protein) wasvirtually unaltered in the enzyme heated at pH 6.1 and 7.5(lmax 334 and 335 nm, respectively), but was definitely red-shifted on heating at pH 9.5 (lmax 340 nm), indicating that
transglutaminase, denatured at alkaline pH, was characterizedby a more open conformation and prominent unfolding oftryptophan-containing regions. Accordingly, values of the ratioof fluorescence emission intensity at 350 and 330 nm (0.76 inthe native protein) increased up to 0.87 and 0.92 in the enzymeheated at pH 6.1 and 9.5, confirming the lower stability tounfolding at pH 9.5 of the N-terminal region.
Changes in secondary structure during unfolding
The secondary structure of native and heat-denatured trans-glutaminase was determined by both FT-IR and CD analysis onsamples unfolded at alkaline pH 7.5 and 9.5, but only by FT-IRat pH 6.1, as the extensive development of turbidity, even atlow protein concentration, prevented CD measurement atpH 6.1. FT-IR spectra were recorded at a much higher proteinconcentration (30 mg´mL21) than CD (0.3 mg´mL21), but thegeneral agreement of the data allows a combined description.
In Fig. 7 we present the second-derivative deconvolutedFT-IR spectra, recorded on equilibration at different tempera-tures, at pD 6.1 and 9.2. Data on thermal fusion at pD 7.5 wereidentical with those previously reported [17]. At the initialtemperature (20 8C) the spectrum was dominated at everypD by signals of b structures, as previously reported, indicatingno, or small, differences in transglutaminase secondarystructure. As the temperature was raised, the spectrum wasmodified, with the progressive disappearance of the specificbands of the helical structures and b structures, leading tototal loss of protein secondary structure. At pD 6.1, thisphenomenon was observed at 65 8C, because at 60 8C thespectrum showed a residual a-helix band. At pD 9.5 there wascomplete loss of secondary structure at 55 8C, indicating alower thermal stability at this pD. It must be underlined thatthese temperatures are not the tm but the temperatures at whichthe protein was completely unfolded, as judged by the FT-IRspectra. Approximate tm values were estimated by monitoringthe amide I 0 bandwidth as a function of temperature [17,19].
Fig. 4. Turbidometric changes during unfolding of erythrocyte trans-
glutaminase by progressive heating in 0.08 mm acetate buffer, pH 6.1, and
in 0.1 mm borate buffer, pH 9.5. L.S., light scattering.
Fig. 5. Changes in fluorescence emission ratio during unfolding of
erythrocyte transglutaminase by progressive heating at pH 6.1, 7.5 and
9.5.
Fig. 6. Changes in tm, calculated from the midpoint of emission ratio
melting profiles, during progressive heating of transglutaminase in
acetate, Tris and borate buffers, at pH 6.1, 7.5 and 9.5, respectively.
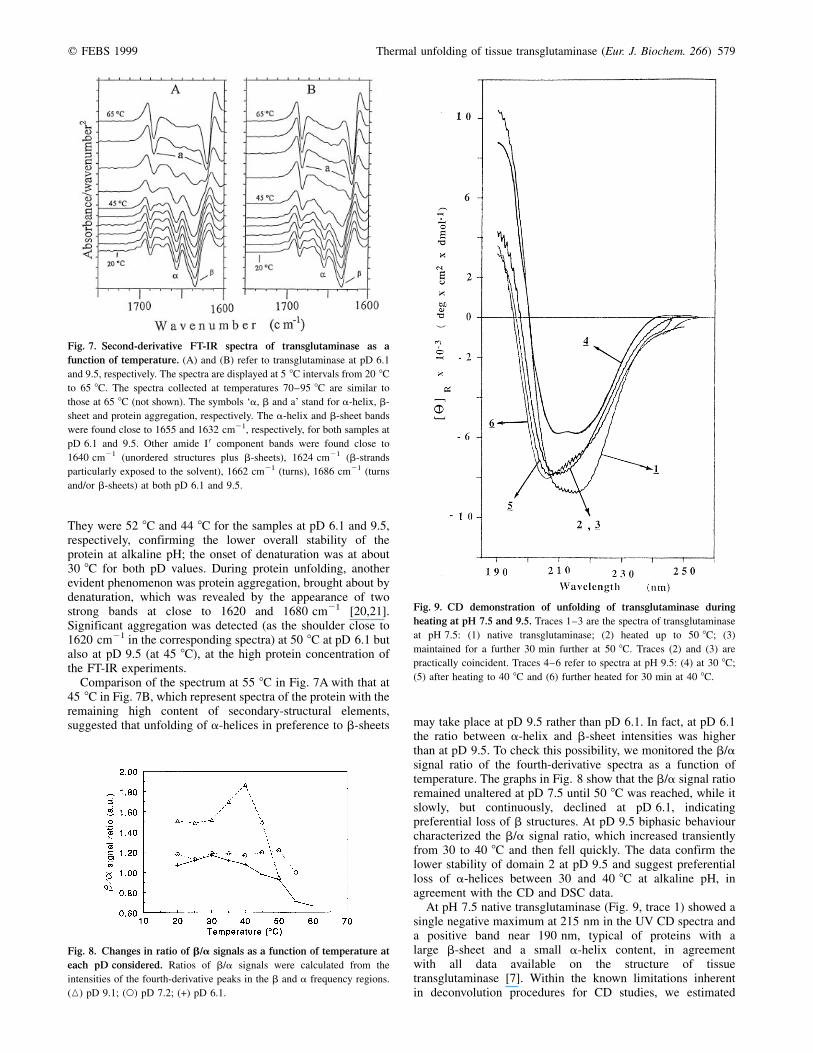
q FEBS 1999 Thermal unfolding of tissue transglutaminase (Eur. J. Biochem. 266) 579
They were 52 8C and 44 8C for the samples at pD 6.1 and 9.5,respectively, confirming the lower overall stability of theprotein at alkaline pH; the onset of denaturation was at about30 8C for both pD values. During protein unfolding, anotherevident phenomenon was protein aggregation, brought about bydenaturation, which was revealed by the appearance of twostrong bands at close to 1620 and 1680 cm21 [20,21].Significant aggregation was detected (as the shoulder close to1620 cm21 in the corresponding spectra) at 50 8C at pD 6.1 butalso at pD 9.5 (at 45 8C), at the high protein concentration ofthe FT-IR experiments.
Comparison of the spectrum at 55 8C in Fig. 7A with that at45 8C in Fig. 7B, which represent spectra of the protein with theremaining high content of secondary-structural elements,suggested that unfolding of a-helices in preference to b-sheets may take place at pD 9.5 rather than pD 6.1. In fact, at pD 6.1
the ratio between a-helix and b-sheet intensities was higherthan at pD 9.5. To check this possibility, we monitored the b/asignal ratio of the fourth-derivative spectra as a function oftemperature. The graphs in Fig. 8 show that the b/a signal ratioremained unaltered at pD 7.5 until 50 8C was reached, while itslowly, but continuously, declined at pD 6.1, indicatingpreferential loss of b structures. At pD 9.5 biphasic behaviourcharacterized the b/a signal ratio, which increased transientlyfrom 30 to 40 8C and then fell quickly. The data confirm thelower stability of domain 2 at pD 9.5 and suggest preferentialloss of a-helices between 30 and 40 8C at alkaline pH, inagreement with the CD and DSC data.
At pH 7.5 native transglutaminase (Fig. 9, trace 1) showed asingle negative maximum at 215 nm in the UV CD spectra anda positive band near 190 nm, typical of proteins with alarge b-sheet and a small a-helix content, in agreementwith all data available on the structure of tissuetransglutaminase [7]. Within the known limitations inherentin deconvolution procedures for CD studies, we estimated
Fig. 7. Second-derivative FT-IR spectra of transglutaminase as a
function of temperature. (A) and (B) refer to transglutaminase at pD 6.1
and 9.5, respectively. The spectra are displayed at 5 8C intervals from 20 8C
to 65 8C. The spectra collected at temperatures 70±95 8C are similar to
those at 65 8C (not shown). The symbols `a, b and a' stand for a-helix, b-
sheet and protein aggregation, respectively. The a-helix and b-sheet bands
were found close to 1655 and 1632 cm21, respectively, for both samples at
pD 6.1 and 9.5. Other amide I 0 component bands were found close to
1640 cm21 (unordered structures plus b-sheets), 1624 cm21 (b-strands
particularly exposed to the solvent), 1662 cm21 (turns), 1686 cm21 (turns
and/or b-sheets) at both pD 6.1 and 9.5.
Fig. 8. Changes in ratio of b/a signals as a function of temperature at
each pD considered. Ratios of b/a signals were calculated from the
intensities of the fourth-derivative peaks in the b and a frequency regions.
(K) pD 9.1; (W) pD 7.2; (+) pD 6.1.
Fig. 9. CD demonstration of unfolding of transglutaminase during
heating at pH 7.5 and 9.5. Traces 1±3 are the spectra of transglutaminase
at pH 7.5: (1) native transglutaminase; (2) heated up to 50 8C; (3)
maintained for a further 30 min further at 50 8C. Traces (2) and (3) are
practically coincident. Traces 4±6 refer to spectra at pH 9.5: (4) at 30 8C;
(5) after heating to 40 8C and (6) further heated for 30 min at 40 8C.

580 C. M. Bergamini et al. (Eur. J. Biochem. 266) q FEBS 1999
a-helix and b-structure contents of 18 and 38%, in acceptableagreement with other estimations [17]. During heating to 50 8C,the negative maximum was blue-shifted at 210 nm and becameless intense (trace 2), indicating a conformational change, withappreciable contribution from random-coil conformation,which remained unaltered during further heating at constanttemperature (trace 3). Adjustment of the pH to 9.5 at 30 8C wassufficient to induce an appreciable conformational change(trace 4), more easily detected by CD than by FT-IRspectroscopy. During subsequent heating to 38 8C, a confor-mational transition from ordered to random-coil structure wasobserved, with a progressive decrease in the a-helical signals(traces 5 and 6 in Fig. 9).
Limited proteolysis by V8 proteinase
To characterize heat-denatured transglutaminase further, weexplored its sensitivity to proteinases. The native protein is
quite sensitive to proteinases, which invariably cleave it at theloop connecting domains 2 and 3 [7]. Proteinase V8 is anexception, transglutaminase being resistant to it at lowtemperature but not if it is heated. In experiments carried outat different pH values (6.1, 7.5 and 9.5), we noted commence-ment of cleavage at temperatures roughly corresponding to tmvalues determined by inactivation and fluorescence measure-ments, as exemplified in Fig. 10, always yielding similarpatterns of cleavage, with main bands of 72, 58, 39, 34, 19 and17 kDa. Automated Edman degradation of the major peptidesfailed to give a detectable N-terminal sequence, indicating thatthe peptides identified start with a blocked N-terminus in theintact protein, and therefore cleavage progressively shortenedthe peptide chain starting from the C-terminal side of theprotein. Thus, irrespective of the pH at which unfolding wascarried out, the C-terminal region becomes the preferential siteof progressive proteolysis.
D I S C U S S I O N
Thermal denaturation analysis is a favored technique in thestructural dynamic studies of proteins, by DSC [11,22] or acombination of heat treatment with other techniques [23]. Herewe describe the combined effects of temperature and pH ontissue-type transglutaminase, utilizing activity measurementsand assessment of the stability of selected protein regions byDSC. Results were also validated by spectroscopic techniquesand measurement of sensitivity to proteolysis.
Our present data indicate that different regions of trans-glutaminase unfold irreversibly with distinct thermal profiles,generating highly co-operative transitions typical of stronglyinteracting protein regions. We resolved two transitions relatedto the unfolding of the N-terminal (domains 1 and 2) and theC-terminal region (domain 3 and 4), respectively, byanalysing calorimetric and fluorimetric data. From a functionalpoint of view, denaturation of the N-terminal region correlateswith loss of catalytic activity, as expected from the availablestructural data, which indicate that the active centre is withindomain 2, while denaturation of domains 3 and 4 is involved inthe solubility of the protein. When considering the effects ofpH, we noted that the enzyme stability is highly pH-dependent,in agreement with previous reports [24], and that unfoldingfollows different pathways at different pH values: at alkalinepH, the region of the protein that displays lower stability is thatrepresented by the N-terminal region (domains 1 and 2),possibly in relation to the ionization state of a lysine residue;under these conditions the protein is unfolded with loss of thehelical structures from domain 2 and considerable increase insolvent accessibility of the tryptophan residues. In contrast,denaturation at acid pH heavily involves domains 3 and 4, witha decrease in b secondary structure consistent with unfolding ofthe two C-terminal barrels. This perturbation exposes buriedprotein surfaces which mediate massive protein aggregation (atpH between 4 and 6), rendering the inactivation processcompletely irreversible. In other words, the denaturationpathway at alkaline pH proceeds through selective unfoldingof the N-terminal region, while at acid pH the C-terminalregion is the main target of inactivation. In both instances, theC-terminal moiety of the protein is made accessible to a pro-teinase that is inactive on the native enzyme. We hypothesizethat the susceptibility to proteolysis of the C-terminal regiondepends on different mechanisms, i.e. its primary unfoldingat acid pH, and, at alkaline pH, the loss of the protectionexerted in the native protein by the folded domains 1 and 2,the prevailing unfolding of which allows the proteolytic
Fig. 10. Effects of temperature on the time course of cleavage of
transglutaminase during incubation with V8 proteinase (1 : 100, w/w) at
pH 6.1, 7.5 and 9.5. After timed intervals of heating in the presence of V8,
samples were removed and analyzed by SDS/PAGE as described in
Materials and methods section. Gel A (pH 6.1): lane 1, unproteolyzed
control; lanes 2±4, transglutaminase treated for 5, 15 and 30 min at 35 8C;
lanes 5±7, treatment for 5, 15 and 30 min at 40 8C; lanes 8±10, treatment
for 5, 15 and 30 min at 45 8C. Gel B (pH 7.5): lane 1, unproteolyzed
control; lanes 2±4, treatment for 5, 15 and 30 min at 40 8C; lanes 5±7,
treatment for 5, 15 and 30 min at 45 8C; lanes 8±10, treatment for 5, 15 and
30 min at 50 8C. Gel C (pH 9.5): lane 1, unproteolyzed control; lane 2,
transglutaminase treated with V8 proteinase for 15 min at 20 8C; lanes 3±5,
treatment for 5, 10 and 15 min at 38 8C; lanes 6±8, treatment for 5, 10 and
15 min at 40 8C; lanes 9±11, treatment for 5, 10 and 15 min at 45 8C.

q FEBS 1999 Thermal unfolding of tissue transglutaminase (Eur. J. Biochem. 266) 581
degradation to take place. It is interesting to note that recentstudies have clearly documented the existence of proteolyzedforms of transglutaminase in the mammalian intestinalmucosa, which might eventually exhibit even calcium-independent, i.e. unregulated, activity [25]. The N-terminaland C-terminal regions in transglutaminases are now knownto perform different physiological roles, in relation to thecontrol of catalytic activity (including inhibition by theC-terminal moiety in the homologous transglutaminase factorXIIIa [26]), anchorage to extracellular matrix proteins [27]and interaction with phospholipases [28]; their selectivedegradation on unfolding might contribute to the patho-genesis of a number of important diseases, includingneurodegenerative syndromes [29]. All these aspectsrequire further investigation.
When comparing the present findings with those available inthe literature, we must underline that unfolding studies ontransglutaminases have largely been carried out on factor XIII[11,12], a dimeric transglutaminase with a considerable degreeof homology with the tissue isoenzyme [30,31]. DSC studies atlow (2.9±3.6) and high (8.4±11.0) pH, and in the presence ofguanidine, have identified several changes in the structure offactor XIII, including subunit dissociation and separateunfolding of domains 1 and 2 and domains 3 and 4 [11].Analysis was complex because of the interrelationship betweendissociation and unfolding, but the data clearly indicatethat the N-terminal region (domains 1 and 2) was moreheat labile (tm about 65±70 8C) than the C-terminaldomains 3 and 4 (tm over 90 8C). Dong et al. [12]analyzed changes in secondary structure during heatingand detected a marked tendency of factor XIII tointermolecular aggregation, on heating to 80 8C, throughinteracting b-strands, probably involving the more heat-stable C-terminal region.
Our data on tissue-type transglutaminase do not yet achievecomplete molecular understanding, although they clearlyprovide evidence for the alkaline thermolability of the protein,which is evident at temperatures as low as 30 8C. They furtherindicate a considerable overlap in the unfolding mechanism oftransglutaminase and factor XIII, although the intrinsic proteinstability is higher for factor XIII than for transglutaminase. Webelieve that a thorough analysis of the interdomain interactionsin transglutaminase and factor XIII will eventually explainthese differences, which will be able to be proved experi-mentally once the isolated domains become available in largeamounts for use in physicochemical analysis in the isolated andcombined state. Nevertheless, the present data begin to shedlight on the dynamics of folding of tissue-type transglutaminaseand might possibly be useful in future studies to explore its useas a tool to modify proteins for investigative [32] or applicativepurposes, using soluble or immobilized transglutaminase[33,34] as well as for a molecular understanding of thepathogenesis of celiac disease and eventually of otherautoimmune conditions.
A C K N O W L E D G E M E N T S
The authors are indebted to Professor Franco Dallocchio for helpful
discussions and to Dr Patrizia Polverino de Laureto for determination of the
N-terminal sequence of V8-generated peptides. The authors are also
grateful to Mr Marco Signorini, now retired, and Miss Monica Squerzanti
for technical and secretarial assistance. This work was supported by grants
from the Ministry of the University (MURST ex 60%) and the Italian CNR
to C. M. B.
R E F E R E N C E S
1. Folk, J.E. & Fynnlayson, S.S. (1977) The 1-(g-glutamyl) lysine
crosslink and the catalytic activity of transglutaminases. Adv. Protein
Chem. 31, 1±133.
2. Greenberg, C.S., Birckbichler, P.J. & Rice, R.H. (1991) Transgluta-
minases: multifunctional crosslinking enzymes that stabilize tissues.
FASEB J. 5, 3071±3077.
3. Fesus, L., Davies, P.J.A. & Piacentini, M. (1992) Apoptosis: molecular
mechanisms in programmed cell death. Eur. J. Cell Biol. 56,
170±177.
4. Nielsen, P.M. (1995) Reactions and potential industrial applications of
transglutaminases. Review of literature and patents. Food Biotechnol.
9, 119±156.
5. Iismaa, S.E., Chung, L., Wu, M.-J., Teller, D.C., Yee, V.C. & Graham,
R.M. (1997) The core domain of the tissue transglutaminase Gh
hydrolyzes GTP and ATP. Biochemistry 36, 11655±11664.
6. Monsonego, A., Friedmann, I., Shani, Y., Eisenstein, M. & Schwartz,
M. (1998) GTP dependent conformational changes associated with
the functional switch between Galpha and crosslinking activities of
brain-derived tissue transglutaminase. J. Mol. Biol. 282, 713±720.
7. Casadio, R., Polverini, E., Mariani, P., Spinozzi, F., Carsughi, F.,
Fontana, A., de Polverino Laureto, P., Matteucci, G. & Bergamini,
C.M. (1999) The structural basis for regulation of tissular
transglutaminase by calcium ions. Eur. J. Biochem. 262, 672±679.
8. Nury, S., Meunier, J.C. & Mouranche, A. (1989) The kinetics of the
thermal deactivation of transglutaminase from guinea-pig liver.
Eur. J. Biochem. 180, 161±166.
9. Nury, S. & Meunier, J.C. (1990) Molecular mechanism of the
irreversible thermal denaturation of guinea-pig liver transglutami-
nase. Biochem. J. 266, 487±490.
10. Rinas, U., Risse, B., Jaenicke, R. & Abel, K.-J. (1990) Denaturation-
renaturation of the fibrin-stabilizing factor XIII a-chain isolated from
human placenta. Biol. Chem. Hoppe Seyler 371, 49±56.
11. Kurochkin, I.V., Procyk, R., Bishop, P.D., Yee, V.C., Teller, D.C.,
Ingham, K.C. & Medved, L.V. (1995) Domain structure, stability and
domain±domain interactions in recombinant factor XIII. J. Mol. Biol.
248, 414±430.
12. Dong, A., Kendrick, B., Kreilgard, L., Matsuura, J., Manning, M.C. &
Carpenter, J.F. (1997) Spectroscopic study of secondary structure and
thermal denaturation of recombinant factor XIII in aqueous solutions.
Arch. Biochem. Biophys. 347, 213±220.
13. Schuppan, D., Dieterich, W., Ehnis, T., Bauer, M., Donner, P., Volta, U.
& Riecken, E.O. (1998) Identification of the autoantigen of the celiac
disease. Ann. N.Y. Acad. Sci. 859, 121±126.
14. Halttunen, T. & Maki, M. (1999) Serum immunoglobulin A from
patients with celiac disease inhibits human T84 intestinal crypt
epithelial cell differentiation. Gastroenterolology 116, 566±572.
15. Bergamini, C.M. & Signorini, M. (1993) Studies on tissular
transglutaminases: interaction of erythrocyte type 2 transglutaminase
with GTP. Biochem. J. 291, 37±39.
16. Banecki, B., Zylicz, M., Bertoli, E. & Tanfani, F. (1992) Structural and
functional relationships in Dnak and Dnak 756 heat-shock proteins
from Escherichia coli. J. Biol. Chem. 267, 25051±25058.
17. Tanfani, F., Bertoli, E., Signorini, M. & Bergamini, C.M. (1993)
Structural investigation of transglutaminase by Fourier transform
infrared spectroscopy. Eur. J. Biochem. 218, 499±505.
18. Laemmli, U.K. (1970) Cleavage of structural proteins during the
assembly of the head of bacteriophage T4. Nature (London) 227,
680±685.
19. Lippe, G., Di Pancrazio, F., Dabbeni-Sala, F., Bertoli, E. & Tanfani, F.
(1995) Influence of ADP, AMP-PNP and of depletion of nucleotides
on the structural properties of F1 ATPase: a Fourier transform
infrared study. FEBS Lett. 373, 141±145.
20. Jackson, M. & Mantsch, H.H. (1991) Beware of proteins in DMSO.
Biochim. Biophys. Acta 1078, 231±235.
21. Jackson, M. & Mantsch, H.H. (1992) Halogenated alcohols as solvents
for proteins. FTIR spectroscopy studies. Biochim. Biophys. Acta
1118, 139±143.

582 C. M. Bergamini et al. (Eur. J. Biochem. 266) q FEBS 1999
22. Privalov, P.L. (1989) Thermodynamic problems of protein structure.
Annu. Rev. Biophys. Biophys. Chem. 18, 47±69.
23. Arnold, U., Rucknagel, K.P., Schierhorn, A. & Ulbrich-Hofmann, K.
(1996) Thermal unfolding and proteolytic susceptibility of ribo-
nuclease A. Eur. J. Biochem. 237, 862±869.
24. Lichti, U., Benn, T. & Yuspa, S.H. (1985) Retinoic-acid induced
transglutaminase in mouse epidermal cells is distinct from epidermal
transglutaminase. J. Biol. Chem. 260, 1422±1426.
25. Tsai, Y.H., Lai, W.T.F., Chen, S.H. & Johnson, L.R. (1998) A novel
calcium independent enzyme capable of incorporating putrescine into
proteins. Biochem. Biophys. Res. Commun. 244, 1261±1267.
26. Mitkevich, O.V., Shainoff, J.R., Di Bello, P.M., Yee, V.C., Teller, D.C.,
Smeikal, G.B., Bishop, P.D., Kolotushkina, I.S., Fickenscher, K. &
Samokhin, G.P. (1998) Coagulation Factor XIII a undergoes a
conformational change evoked by glutamine substrates. Studies of
kinetcs of inhibition and binding of factor XIIIa by crossreacting
antifibrinogen antibody. J. Biol. Chem. 273, 14387±14391.
27. Jeong, J.M., Murthy, S.N.P., Radek, J.T. & Lorand, L. (1995) The
fibronectin binding domain of transglutaminase. J. Biol. Chem. 270,
5654±5658.
28. Hwang, K.-C., Gray, C.D., Sivasubramanian, N. & Im, J.-M. (1995)
Interaction site of GTP binding Gh (transglutaminase II) with phos-
pholipase C. J. Biol. Chem. 270, 27058±27061.
29. Lorand, L. (1996) Neurodegenerative diseases and transglutaminase.
Proc. Natl Acad. Sci. USA 93, 14310±14313.
30. Nakanishi, K., Nara, K., Hagiwara, H., Aoyama, Y., Ueno, H. &
Hirose, S. (1991) Cloning and sequence analysis of cDNA clones
from bovine aortic-endothelial cells transglutaminase. Eur. J.
Biochem. 202, 15±21.
31. Yee, V.C., Pedersen, L.C., Le Trong, I., Bishop, P.D., Sternkamp, R.E.
& Teller, D.C. (1994) Three dimensional structure of a trans-
glutaminase: human blood coagulation factor XIII. Proc. Natl Acad.
Sci. USA 91, 7296±7300.
32. Matsumura, Y., Chanyongvorakul, Y., Kumazawa, Y., Ohtsuka, T. &
Mori, T. (1996) Enhanced susceptibility to transglutaminase of
alpha-lactalbumin in the molten globule state. Biochim. Biophys. Acta
1296, 69±76.
33. Tanimoto, S.-Y. & Kinsella, J.E. (1988) Enzymatic modification of
proteins: effects of transglutaminase crosslinking on some physical
properties of beta-lactoglobulin. J. Agric. Food Chem. 36, 281±285.
34. Huang, X.L., Catignani, G.L. & Swaisgood, H.E. (1995) Immobi-
lization of biotinylated transglutaminase by bioselective adsorption to
immobilized avidin and characterization of the immobilized activity.
J. Agric. Food Chem. 43, 895±901.