complex phytohormone responses during the cold acclimation of two wheat cultivars differing in cold...
TRANSCRIPT

Cc
KOA
a
b
c
d
e
a
ARR2A
KCDFPW
I
pasbbst
Cb
0d
Journal of Plant Physiology 169 (2012) 567– 576
Contents lists available at SciVerse ScienceDirect
Journal of Plant Physiology
jou rn al h o mepage: www.elsev ier .de / jp lph
omplex phytohormone responses during the cold acclimation of two wheatultivars differing in cold tolerance, winter Samanta and spring Sandra
lára Kosováa, Ilja Tom Prásil a, Pavel Vítámvása, Petre Dobrevb, Václav Motykab, Kristyna Flokovác,ndrej Novákc, Veronika Tureckovác, Jakub Rolcikc, Bedrich Pesekb, Alena Trávnickováb,lena Gaudinováb, Gabor Galibad,e, Tibor Jandad, Eva Vlasákováa, Pavla Prásilováa, Radomíra Vankováb,∗
Crop Research Institute, Drnovska Str. 507, Prague, CZ-161 06, Czech RepublicInstitute of Experimental Botany AS CR, Rozvojova 263, Prague 6, CZ-16502, Czech RepublicInstitute of Experimental Botany AS CR, Slechtitelu 11, Olomouc, CZ-783 71, Czech RepublicAgricultural Research Institute of the Hungarian Academy of Sciences, Martonvásár, Brunszvik Str. 2, H-2462, HungaryFaculty of Information Technology, Research Institute of Chemical and Process Engineering, University of Pannonia, Veszprém, Egyetem Str. 10, H-8200, Hungary
r t i c l e i n f o
rticle history:eceived 11 August 2011eceived in revised form9 November 2011ccepted 1 December 2011
eywords:old stressehydrinrost tolerance
a b s t r a c t
Hormonal changes accompanying the cold stress (4 ◦C) response that are related to the level of frosttolerance (FT; measured as LT50) and the content of the most abundant dehydrin, WCS120, were com-pared in the leaves and crowns of the winter wheat (Triticum aestivum L.) cv. Samanta and the springwheat cv. Sandra. The characteristic feature of the alarm phase (1 day) response was a rapid elevation ofabscisic acid (ABA) and an increase of protective proteins (dehydrin WCS120). This response was fasterand stronger in winter wheat, where it coincided with the downregulation of bioactive cytokinins andauxin as well as enhanced deactivation of gibberellins, indicating rapid suppression of growth. Next, theethylene precursor aminocyclopropane carboxylic acid was quickly upregulated. After 3–7 days of coldexposure, plant adaptation to the low temperature was correlated with a decrease in ABA and elevation of
hytohormonesheat
growth-promoting hormones (cytokinins, auxin and gibberellins). The content of other stress hormones,i.e., salicylic acid and jasmonic acid, also began to increase. After prolonged cold exposure (21 days), aresistance phase occurred. The winter cultivar exhibited substantially enhanced FT, which was associatedwith a decline in bioactive cytokinins and auxin. The inability of the spring cultivar to further increaseits FT was correlated with maintenance of a relatively higher cytokinin and auxin content, which wasachieved during the acclimation period.
© 2012 Elsevier GmbH. All rights reserved.
ntroduction
Low temperatures (cold and frost) represent a severe threat tolant survival and economic crop productivity in many regions. Thebility of plants to acclimate to low temperatures is crucial for theirurvival. Plant cold acclimation is a complex process accompaniedy profound changes in the expression of multiple genes regulated
y various phytohormones and resulting in the de novo biosynthe-is of several stress-protective compounds and an enhancement ofhe acquired frost tolerance (FT) level. With regard to the generalAbbreviations: ABA, abscisic acid; ACC, aminocyclopropane-1-carboxylic acid;K, cytokinin; CKX, cytokinin oxidase/dehydrogenase; FT, frost tolerance; GA, gib-erellin; IAA, indole-3-acetic acid; JA, jasmonic acid; SA, salicylic acid.∗ Corresponding author. Tel.: +420 225 106 427; fax: +420 225 106 456.
E-mail address: [email protected] (R. Vanková).
176-1617/$ – see front matter © 2012 Elsevier GmbH. All rights reserved.oi:10.1016/j.jplph.2011.12.013
dynamics of the plant stress response, several phases can be dis-tinguished: (a) a control, non-stressed phase; (b) an early alarmphase when the plant is not adapted and stress acts as a shockfactor; (c) an acclimation phase, when the plant actively redirectsits metabolism to deal with stress; (d) a resistance phase whenthe plant reaches its maximum stress tolerance level and a new,steady-state metabolic balance; and if the stress lasts too long (e)an exhaustion phase when the plant metabolism, adjusted to thestress conditions, collapses (Larcher, 2003). Each phase of the plantstress response is uniquely reflected at regulatory (phytohormone)as well as adaptive (water-relations, protein and metabolite) levels(Vankova, 2010; Kosova et al., 2011).
Changes in water relationships during plant cold stress response
have been described in detail in Larcher (2003) and Gusta et al.(2005, 2009). Upon exposure to cold, hydraulic conductivity of thecereal roots decreases, which results in a decrease of water poten-tial and induction of transient wilting of plant tissue, leading to
5 nt Ph
sttmodiPa
mrnamamApg2
pd(drWaisiaar
lb(t
rdohtiV
drclnaterCcabp
ha
68 K. Kosová et al. / Journal of Pla
tomata closure in the alarm phase. During the acclimation phase,he reduced water potential returns to levels similar to those beforehe cold exposure of plants due to the osmotic adjustment (accu-
ulation of osmotically active compounds). As a consequence ofsmotic adjustment, osmotic potential decreases in cereal organsuring cold acclimation, allowing stabilization of turgor and open-
ng of stomata for exchange of gases in leaves (Gusta et al., 2005;rasil et al., 2007). However, the growth and development of plantsre reduced (Larcher, 2003; Prasil et al., 2004).
Plant stress responses are regulated by a range of phytohor-ones, which may be divided into two groups: “positive growth
egulators” [auxin, cytokinins (CKs), gibberellins (GA) and brassi-osteroids] and “stress hormones” [abscisic acid (ABA), jasmoniccid (JA), salicylic acid (SA) and ethylene]. Due to intensive hor-one cross-talk (positive or negative), individual processes are
ffected by multiple hormones. Nevertheless, different stress hor-ones play a key role in the response to a particular stress. Thus,BA is a crucial hormone in the defense against abiotic stresses, SAlays a decisive role in the protection against biotrophs, while JAoverns the response to wounding and necrotroph attack (Vankova,010).
One of the major groups of stress-protective proteins is COR/LEAroteins. This group includes the LEA-II family of proteins (dehy-rins), which are elevated upon dehydration. In common wheatTriticum aestivum), an important group of cold-inducible dehy-rins is the WCS120 protein family (Sarhan et al., 1997), which isepresented by at least five members (WCS200, WCS180, WCS120,
CS66, and WCS40) that differ in their relative molecular weightss well as their abundance. The most abundant member of this fam-ly in cold-treated wheat samples is the WCS120 protein. Previoustudies (e.g., Vitamvas et al., 2007) have shown that differencesn the relative abundance of WCS120 and its homologues in cold-cclimated Gramineae species are correlated with the level ofcquired FT; i.e., the WCS120 protein (and its homologues) can beegarded as markers of acquired FT in cold-acclimated cereals.
Promoter analysis of the wheat Wcs120 gene, as well as its bar-ey homologue Dhn5, revealed that their expression is regulatedy both ABA-dependent and ABA-independent signalling pathwaysSarhan et al., 1997). The latter pathway involves C-repeat-bindingranscription factors (CBFs).
In addition to the stimulation of the expression of stress-elated genes, ABA exhibits other protective functions in cold stressefense, especially stabilization of membranes, protection againstxidative stress, improvement of water status by elevation of rootydraulic conductivity and stomata closure. The function of ABA inhe early stage of the cold stress response coincides with a transientncrease of ABA levels during chilling stress (Galiba et al., 1993;eisz et al., 1996).
Recent reports suggest that, apart from the stimulation ofefense mechanisms, an important component of the stressesponse is the modulation of plant growth and development. Theold-inducible CBF transcription factor was found to downregulateevels of bioactive GAs as well as to stabilize repressors of their sig-alling pathway, i.e., DELLA proteins (Achard et al., 2008). Much lessttention has been focused on CKs and auxins. Overexpression ofhe CK biosynthetic gene (ipt) was found to promote cold stress tol-rance in Arabidopsis (Guo et al., 2010). However, Jeon et al. (2010)eported cold-stimulated expression of negative regulators of theK signalling pathway (type-A response regulators), which indi-ated downregulation of the signal transduction of these hormones,t least during the initial phase of the cold stress response. A studyy Shibasaki et al. (2009) on cold-induced changes in auxin trans-
ort suggested the involvement of auxin in cold stress responses.The aim of the present study was to characterize individualormone responses associated with the reaction to cold stress in
cold-tolerant winter wheat, cv. Samanta, and a cold-sensitive
ysiology 169 (2012) 567– 576
spring wheat, cv. Sandra, during 21 days of cold treatment (4 ◦C)and their phytohormone relationships to the FT level (measuredas LT50) as well as changes in the content of the most abundantdehydrin, WCS120, in two different organs, leaves (green tissues)and crowns (non-green underground parts, not including roots).In addition to the main abiotic stress defense hormone ABA, thegrowth-promoting hormones CKs, auxin, GAs and their metabo-lites, as well as the stress hormones SA, JA and ethylene wereevaluated.
Materials and methods
Plant material and growth conditions
Seeds of two wheat (Triticum aestivum L.) cultivars, winter cv.Samanta and spring cv. Sandra, were obtained from the breedingcompany Selgen a.s. (Prague, Czech Republic). The seeds germi-nated in the dark on moist filter paper at 20 ◦C for 2 days. Theseedlings were planted in pots filled with soil (a mixture of field,soil and sand, 4:2:1) and grown at 18–20 ◦C; 12 h photoperiodand 350 �mol m−2 s−1 in growth chambers (Tyler, type T – 16/4,Budapest, Hungary) until the three-leaf stage. Next, the growthtemperature was decreased to 4 ◦C. During the entire experiment,the plants were watered regularly every 3rd day, with no fertilizerbeing supplied.
Leaf and crown tissues were sampled at 0 (control, non-treatedplants), 1, 3, 7 and 21 days of cold treatment. The 3rd leaf (theyoungest fully expanded leaf) was used for all analyses (frost tol-erance test, determination of WCS120 content, determination ofphytohormone levels). As crowns, non-green underground stemtissues not including roots were considered. During the entireexperiment, neither of the cultivars entered the reproductive stageof development (a double-ridge stage of shoot apex) (Prasil et al.,2004).
Frost tolerance tests
Leaf and crown segments (1 cm) were exposed to a set of freez-ing temperatures in the range of −1.5 ◦C to −30 ◦C according toPrasil and Zamecnik (1998). The plant materials (two replicatesper each frost temperature, consisting of 10 leaf segments or threecrowns) were inoculated with ice crystals at −1.5 ◦C and held at 12freezing temperatures for 20 min. The rate of cooling was 12 ◦C h−1,and the frozen samples were thawed in a water bath at 1 ◦C for 12 h.The extent of freezing damage was determined conductometri-cally according to Prasil and Zamecnik (1998). Lethal temperatures(LT50), i.e., the temperature causing 50% electrolyte leakage, werecalculated according to Janacek and Prasil (1991).
Determination of WCS120 content
The third young, fully expanded leaves were frozen in liquidnitrogen and stored at −80 ◦C until protein extraction and boiling.The dehydrin extraction, 1D SDS-PAGE and immunoblot analyseswere performed according to Vitamvas et al. (2007). The amountof WCS120 protein was analyzed densitometrically using QuantityOne software (Bio-Rad, version 4.6.2, Munchen, Germany).
Quantification of phytohormones (CK, IAA, ABA, GA, SA, ethyleneprecursor ACC and JA)
The endogenous phytohormone concentrations were deter-
mined in the leaves and crowns of both wheat cultivars. Allexperiments were repeated three times. For analyses of endoge-nous CKs, extraction and purification were performed accordingto Novak et al. (2003), and CK levels were quantified by ultra
nt Physiology 169 (2012) 567– 576 569
ps
iPU(iq
tsa
aHqAt
iwwMtS
dp5a2api
mJ
D
w(iopAnt(er
S
lb(ShSu
COLD TREATMENT (DAY)
20151050
WC
S120 R
EL
. A
CC
UM
UL
AT
ION
(%
)0
2
4
6
8
10
12
14
16
LT
50 (
°C
)
-16
-14
A
B
C
-12
-10
-8
-6
-4
-2
Samanta (leaf)
Samanta (crown)
Sandra (leaf)
Sandra (crown)
WCS120 REL. ACCUMULATION (%)
181614121086420
LT
50 (
°C
)
-18
-16
-14
-12
-10
-8
-6
-4
-2
Samanta (leaf)
Samanta (crown)
Sandra (leaf)
Sandra (crown)
Fig. 1. Survival of frost test, expressed as LT50 values (A) and (B) relative accumula-tion of WCS120 protein (B) in the leaves and crowns of the winter wheat cv. Samantaand the spring wheat cv. Sandra at 0, 1, 3, 7 and 21 days of cold treatment (4 ◦C).Bars represent standard error (SE; n = 6). (C) Correlation between relative WCS120accumulation and LT50 values for Sandra and Samanta leaves and crowns from (A)
K. Kosová et al. / Journal of Pla
erformance liquid chromatography–electrospray tandem masspectrometry (UPLC–MS/MS) (Novak et al., 2008).
Endogenous auxin was purified from leaf or crown tissues bymmunoaffinity chromatography and quantified as described byencik et al. (2009). Separation was performed using an AcquityPLC System (Waters) equipped with a Symmetry C18 column
5 �m, 2.1 mm × 150 mm; Waters), and the effluent was introducednto the electrospray ion source of a Quatro micro API tandemuadrupole mass spectrometer (Waters).
The ABA metabolites were determined using solid-phase extrac-ion on Oasis HLB cartridges (60 mg, 3 mL; Waters), a derivatizationtep, ABA-specific immunoaffinity extraction, and quantification by
UPLC–MS/MS system (Tureckova et al., 2009).Gibberellins were extracted and purified according to Dobrev
nd Kaminek (2002). Quantification was performed by LC/MS.PLC (Ultimate 3000, Dionex) coupled to a hybrid tripleuadrupole/linear ion trap mass spectrometer (3200 Q TRAP,pplied Biosystems) in negative ESI and selective reaction moni-
oring mode.SA was determined according to Ayaz et al. (2005). After grind-
ng in liquid nitrogen and extraction with 80% methanol, samplesere concentrated and pH adjusted to 2. SA was extracted 5 timesith diethyl ether and determined using HPLC linked to an Ion Trapass Spectrometer Finnigan MAT LCQ-MSn equipped with an elec-
rospray interface using an RP-C8 column (Merck; Supersphere RPelect B, 2 mm × 250 mm, 4 �m).
The aminocyclopropane-1-carboxylic acid (ACC) level wasetermined using overnight extraction with 80% (v/v) methanol,urification on a SPE C18 column (Strata C18-T, 55 �m, 140 A,00 mg/6 mL; Alltech, U.S.A.) and pH adjustment. The elu-te was applied to a SCX SPE column (Extract Clean, SCX00 mg; Grace, U.S.A.), cleaned with water and eluted with 4 Mmmonium hydroxide. After derivatization with 4 �L 2,3,4,5,6-entafluorobenzyl bromide for 15 min, the sample was injected
nto a GC–MS/MS (PolarisQ, Thermo, U.S.A.).JA was determined after overnight extraction with 80% (v/v)
ethanol and derivatization with 2 mL of diazomethane for 15 min.A was quantified with a GC–MS/MS (PolarisQ, Thermo, U.S.A.).
etermination of cytokinin oxidase/dehydrogenase activity
Cytokinin oxidase/dehydrogenase (CKX; EC 1.4.3.18/1.5.99.12)as extracted and partially purified according to Motyka et al.
2003). Due to a lack of material, CKX could not be determinedn crowns. CKX activity was determined by in vitro assays basedn the conversion of [2-3H]N6-(2-isopentenyl)adenine (iP; pre-ared by the Isotope Laboratory, Institute of Experimental BotanyS CR, Prague, Czech Republic) to [3H]adenine and expressed asmol adenine mg protein−1 h−1. CKX activity was determined inwo independent experiments, which showed the same tendencieswith different absolute values). The results of one representativexperiment (mean value of three parallel assays for each of threeeplicate protein preparations) are presented.
tatistical analyses
Data on the relative accumulation of WCS120 (n = 6; two bio-ogical replicates; three technical replicates corresponding to eachiological replicate) were analyzed using a multi-factorial ANOVAmultiple comparisons) with STATISTICA version 9 software (Stat-
oft Inc.). The significance of the differences among the measuredormone values was assessed using a t-test as implemented inigma Plot. Principal component analysis (PCA) was also performedsing STATISTICA version 9 software.and (B). Correlation coefficients (r) for leaf samples (r = 0.9303) and crown samples(r = 0.9797) revealing statistical significance at the 0.05 level (*) are indicated.
Results
Frost-tolerance tests
The winter wheat cultivar Samanta exhibited a slightly higherFT level (lower LT50 values) compared to the spring wheat cultivarSandra, even under control conditions (0 day of cold treatment;
Fig. 1A). Following the temperature decrease from 20 to 4 ◦C,the acquired FT level began to increase in both cultivars, morequickly and to higher extent in Samanta. In Sandra, the acquired
570 K. Kosová et al. / Journal of Plant Ph
Fig. 2. 1D SDS-PAGE gel stained with Coomassie Brilliant Blue (A) and a section ofa 1D immunoblot (B) showing the relative accumulation of WCS66, WCS120 andWCS40 in leaf (L) and crown (C) tissues from the winter wheat Samanta and thespring wheat Sandra at 0, 1, 3, 7 and 21 days of cold treatment (4 ◦C). Qualitativedta
F(icl
D
rwbfd(Wfd
itcatcThttiS
ifferences in the protein composition of leaf and crown tissues, as well as quanti-ative differences in WCS120 relative accumulation between Samanta and Sandrat later stages of cold treatment (3, 7, 21 days), are clearly visible in (A).
T level ceased to increase during the later stages of cold treatmentbetween 7 and 21 days). In contrast, the acquired FT rose steadilyn Samanta during the entire cold experiment (21 days). In bothultivars, the leaf segments displayed relatively higher acquired FTevels compared with the crown segments at all sampling dates.
ynamics of the relative accumulation of dehydrin (WCS120)
Immunoblot analysis of cold-treated leaf and crown samplesevealed five detectable dehydrin bands of different moleculareights: WCS200, WCS180, WCS120, WCS66 and WCS40. The
ands corresponding to the high molecular weight dehydrins wereaint and barely detectable, while the lower molecular weight dehy-rins, such as WCS66, WCS120 and WCS40, were more abundantFig. 2B). In all samples, the most abundant dehydrin protein was
CS120. Therefore, the bands corresponding to WCS120 were usedor densitometric analysis of changes in the relative dehydrin abun-ance during the cold treatment (Fig. 1B).
At 20 ◦C, a faint band corresponding to WCS120 was detectedn Samanta leaves. No such band was found in Sandra. Afterhe temperature decrease, a band corresponding to WCS120 waslearly detectable in all samples. A multivariate ANOVA for WCS120ccumulation indicated significant differences between all fac-ors studied: genotypes (Samanta, Sandra; P < 0.01), organs (leaves,rowns; P < 0.01) and treatments (0, 1, 3, 7, 21 days of cold; P < 0.01).hroughout the entire cold treatment, the WCS120 levels wereigher in Samanta compared to Sandra. In both cultivars, the crown
issues accumulated slightly higher amounts of WCS120 comparedo the leaf tissues. Both cultivars also exhibited different dynam-cs of relative WCS120 accumulation during the cold treatment. Inamanta, the WCS120 levels in both the leaf and crown samplesysiology 169 (2012) 567– 576
increased steadily during the entire experiment (up to 21 days),while in Sandra, the WCS120 levels in crowns stopped increasingbetween 7 and 21 days. Thus, relative WCS120 accumulation pre-sented dynamics analogous to those of acquired FT expressed asLT50 values during cold treatment.
WCS120 accumulation versus LT50
When the relative WCS120 accumulation values and LT50 valuesof the two cultivars were compared, a statistically significant cor-relation at the 0.05 level was found for both the leaves and crowns(Fig. 1C), indicating that WCS120 accumulation can be regardedas a reliable indicator of LT50 in wheat cultivars. Moreover, theregression lines were parallel but independent for the leaves andcrowns, which indicate that the correlations are valid for differentorgans. However, the relative WCS120/LT50 ratios were differentfor the leaves and crowns, which indicate that the same LT50 valuecorresponded to different WCS120 levels in the two organs.
Abscisic acid
Under control conditions, the ABA levels in Samanta leavesapproached approximately 8 pmol per g FW, while in Sandra theywere below the detection limit. After 1 day of cold, both cultivarsexhibited a sharp increase in ABA (Fig. 3A), approximately twiceas high in Samanta as in Sandra. A further slight increase of ABAwas detected after 3 days. Next, the ABA level gradually decreased,remaining well above the basal level in both cultivars until theend of the experiment (21 days). The levels of ABA metabolites areshown in Fig. 3B–D. The level of ABA glucosyl ester, the ABA stor-age form, which may be cleaved by specific �-glucosidases, wasrather low in the leaves of both cultivars. A moderately elevatedABA glucosyl ester content observed after 21 days was approxi-mately twice as high in Samanta compared to Sandra. Phaseic acid,an ABA catabolite with strongly reduced biological activity, exhib-ited its maximum level after 1 day in Samanta and after 3 daysin Sandra. The most abundant ABA catabolite, the physiologicallyinactive neophaseic acid, reached its maximum level after 3 days.The accumulation of ABA catabolites after 3 days of cold stress indi-cated intensive downregulation of free ABA levels after reaching themaximum in the early phase of the cold response. All ABA metabo-lites exhibited another peak after 21 days in Samanta, which maysuggest regulation of the levels of still actively biosynthesized ABA.In Sandra, ABA derivatives, with the exception of neophaseic acid,showed a decrease during this period.
In Samanta, the increase in ABA was more pronounced in crownscompared to leaves, especially at 1 day, while Sandra exhibiteda similar trend in crowns and leaves. After 21 days, the ABA lev-els were rather low in the crowns of both cultivars. Phaseic acidreached its maximum level after 3 days in Samanta. The neopha-seic acid was high in Samanta crowns and relatively high in Sandracrowns after 1 and 3 days.
Cytokinins
Upon exposure to cold, the levels of bioactive CKs (trans-zeatin,dihydrozeatin and isopentenyladenine and their correspondingribosides) decreased in Samanta leaves (Fig. 4A) with no suchdecline being observed in Sandra. After 3 and 7 days of cold, a tran-sient elevation of bioactive CKs was detected in Samanta. Prolongedexposure to 4 ◦C resulted in extensive downregulation of bioactiveCKs in Samanta to approximately same level as after 1 day of cold.
In Sandra, bioactive CK elevation was milder and shorter (maxi-mum after 3 days), but the bioactive CK content was maintainedafter 7 days at a relatively high level corresponding to the controlconditions.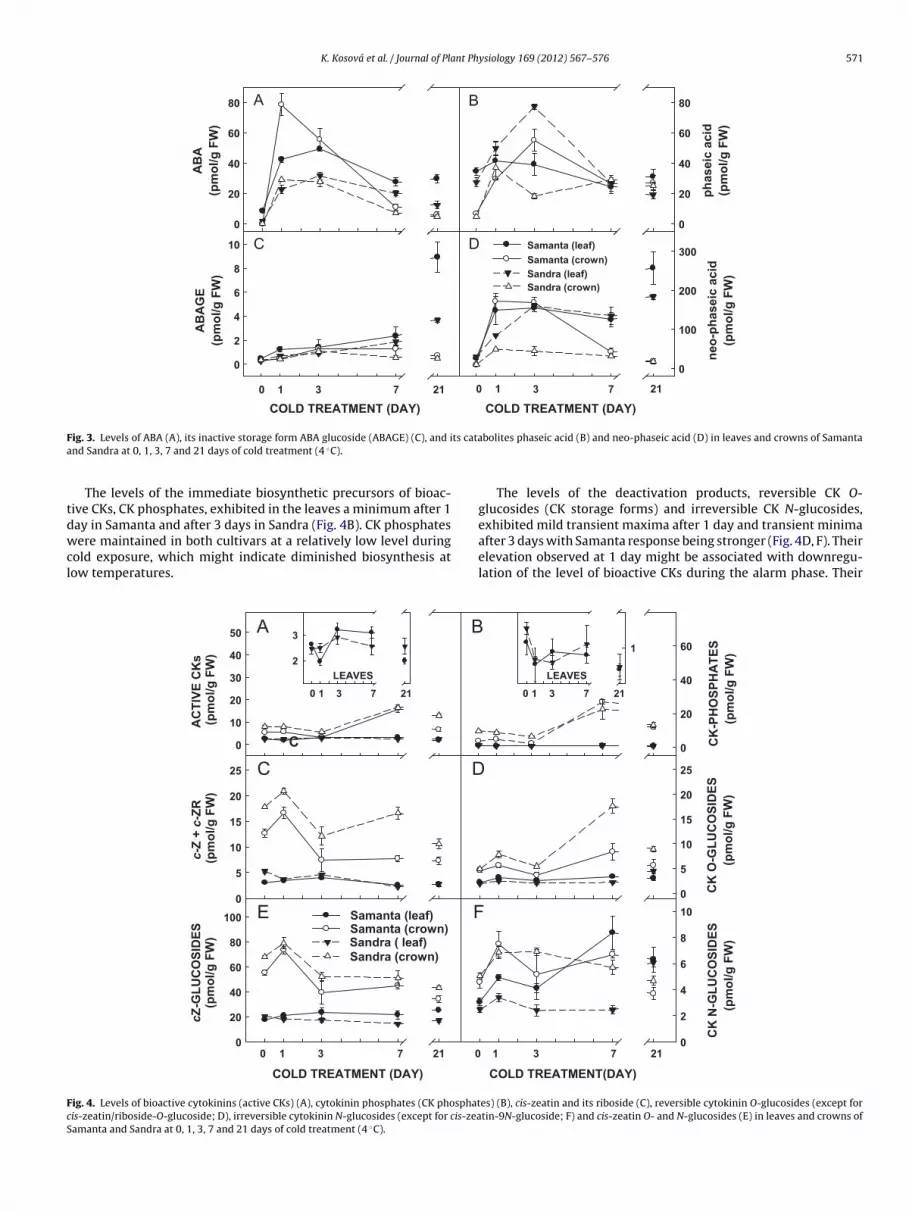
K. Kosová et al. / Journal of Plant Physiology 169 (2012) 567– 576 571
COLD TREATMENT (DAY)
ne
o-p
ha
se
ic a
cid
(pm
ol/
g F
W)
0
100
200
300
COLD TREATMENT (DAY)
AB
AG
E
(pm
ol/
g F
W)
0
2
4
6
8
10
AB
A
(pm
ol/
g F
W)
0
20
40
60
80
ph
as
eic
acid
(pm
ol/
g F
W)
0
20
40
60
80A
DC
B
10 73 21217310
Samanta (leaf)
Samanta (crown)
Sand ra (leaf)
Sand ra (crown)
F ts cataa
tdwcl
FcS
ig. 3. Levels of ABA (A), its inactive storage form ABA glucoside (ABAGE) (C), and ind Sandra at 0, 1, 3, 7 and 21 days of cold treatment (4 ◦C).
The levels of the immediate biosynthetic precursors of bioac-ive CKs, CK phosphates, exhibited in the leaves a minimum after 1
ay in Samanta and after 3 days in Sandra (Fig. 4B). CK phosphatesere maintained in both cultivars at a relatively low level duringold exposure, which might indicate diminished biosynthesis atow temperatures.
COLD TREATMENT (DAY)
0
20
40
60
80
100
cZ-G
LU
CO
SID
ES
(pm
ol/
g F
W)
c-Z
+ c-Z
R
(pm
ol/
g F
W)
0
5
10
15
20
25
0217310
AC
TIV
E C
Ks
(pm
ol/
g F
W)
0
10
20
30
40
50
2
3
LEAVES
0 1 3 7 21
Sa mant a (le af) Samanta (crown) Sandra ( le af)
Sand ra (crown)
A
DC
C
B
FE
ig. 4. Levels of bioactive cytokinins (active CKs) (A), cytokinin phosphates (CK phosphatis-zeatin/riboside-O-glucoside; D), irreversible cytokinin N-glucosides (except for cis-zeaamanta and Sandra at 0, 1, 3, 7 and 21 days of cold treatment (4 ◦C).
bolites phaseic acid (B) and neo-phaseic acid (D) in leaves and crowns of Samanta
The levels of the deactivation products, reversible CK O-glucosides (CK storage forms) and irreversible CK N-glucosides,
exhibited mild transient maxima after 1 day and transient minimaafter 3 days with Samanta response being stronger (Fig. 4D, F). Theirelevation observed at 1 day might be associated with downregu-lation of the level of bioactive CKs during the alarm phase. TheirCOLD TREATMENT(DAY)
CK
N-G
LU
CO
SID
ES
(p
mo
l/g
FW
)
0
2
4
6
8
10
CK
O-G
LU
CO
SID
ES
(p
mo
l/g
FW
)
0
5
10
15
20
25
CK
-PH
OS
PH
AT
ES
(pm
ol/
g F
W)
0
20
40
601
LEAVE S
0 1 3 7 21
21731
es) (B), cis-zeatin and its riboside (C), reversible cytokinin O-glucosides (except fortin-9N-glucoside; F) and cis-zeatin O- and N-glucosides (E) in leaves and crowns of

572 K. Kosová et al. / Journal of Plant Physiology 169 (2012) 567– 576
GA
19 (
pm
ol/
g F
W)
0
20
40
60
80
100
120
GA
8 (
pm
ol/
g F
W)
0
20
40
60
COLD TREATMENT (DAY)
GA
29 (
pm
ol/
g F
W)
0
50
100
150
200
COLD TREATMEN T (DAY)
GA
20 (
pm
ol/
g F
W)
0
5
10
15
20 FE
GA
1 (
pm
ol/
g F
W)
0
50
100
150
200
250
300A
DC
10 73 21217310
Samanta (leaf)
Samanta (cro wn) Sandra (leaf)
Sandra (cr own)
GA
4 (
pm
ol/
g F
W)
0
1
2
3
4
B
F rellins( d 21
miioc
i3sSfw(adaoc
G
cedop((do
ig. 5. Levels of the bioactive gibberellins GA1 (A) and GA4 (B) and inactive gibbedeactivation product, F) in leaves and crowns of Samanta and Sandra at 0, 1, 3, 7 an
inimum after 3 days preceded the elevation of bioactive CKs dur-ng the acclimation phase. Relatively high levels of cis-zeatin andts riboside were found in both cultivars with a mild maximumbserved after 3 days (Fig. 4C). The most abundant CK species wereis-zeatin glucosides (both O- and N-type, Fig. 4E).
In crowns, the content of bioactive CKs was maintained approx-mately at the control level after 1 day of cold, then decreased after
days and substantially increased after 7 days. This profile wasimilar for both cultivars with slightly higher values being seen inandra. A highly significant difference between the cultivars wasound after 21 days when a relatively low level of bioactive CKsas found in Samanta, while a high level was detected in Sandra
Fig. 4A). In both cultivars, the levels of CK phosphates followed similar trend to that of bioactive CKs. Transient maxima wereetected after 1 and 7 days for CK O- and N-glucosides in Samantand, in the case of O-glucosides, also for Sandra. Rather high levelsf cis-zeatin/riboside and their O- and N-glucosides were found inrowns, especially after 1 day of cold (Fig. 4B-F).
ibberellins
The level of bioactive GA4 was relatively low in the leaves androwns of both cultivars (Fig. 5B). The bioactive gibberellin GA1xhibited a similar time course in the leaves of Samanta and San-ra (Fig. 5A) with the exception of a minimum at 1 day that wasbserved only in the winter cultivar. The GA1 minimum was accom-anied by a transient elevation of the GA deactivation product GA29
Fig. 5F) and, to lesser extent, also of the deactivation product GA8Fig. 5D). The amount of GA29 was higher in Samanta than in San-ra (which did not exhibit a GA1 decrease). A gradual elevationf GA1 was observed between 1 and 7 days. The content of theGA19 (precursor, C), GA8 (deactivation product, D), GA20 (precursor, E) and GA29
days of cold treatment (4 ◦C).
deactivation product GA8 showed a maximum at 7 days, when thelevel of bioactive GA1 began to decrease in both cultivars. The timecourse of GA19, the precursor of GA1, slightly preceded that of thisbioactive GA (Fig. 5C). The product of GA19 oxidation, the imme-diate GA1 precursor GA20 (Fig. 5E), exhibited a similar profile toGA1.
In the crowns, transient GA1 maxima occurred at 3 and 7 daysin Samanta and Sandra, respectively. At the same time, the levelof GA29 increased, which might suggest that GA deactivation hadalready been induced during this period.
Indole-3-acetic acid
Both cultivars exhibited a similar time course of the auxin levelsin leaves (Fig. 6A). After 1 day, a mild decrease of IAA was observedin both genotypes. After a transient maximum at 3 days, IAA contentexhibited a mild, gradual decrease in Samanta, while a slight IAAelevation occurred in Sandra. A significant difference between thecultivars was detected in their crowns. After a mild increase at 3days, a steady IAA level was observed in Samanta. Sandra exhibiteda mild increase at 1 day followed by a gradual decrease until 7 days.Next, an increase in IAA was detected until 21 days.
Salicylic acid
After the transfer of plants to the cold, the SA level significantlydecreased in the Samanta leaves. A pronounced SA increase fol-
lowed, with a maximum after 7 days (Fig. 6B). With prolonged coldtreatment, a gradual decrease in SA occurred. In Sandra leaves, theSA profile followed basically the same trend; however, the regula-tion of SA levels was rather mild.
K. Kosová et al. / Journal of Plant Physiology 169 (2012) 567– 576 573
SA
(p
mo
l/g
FW
)
0
200
400
600
800
1000
COLD TREATMENT (DAY)
AC
C (
pm
ol /g
FW
)
0
50
100
150
COLD TREATMENT (DAY)
JA
(p
mo
l /g
FW
)
0
1000
2000
3000
4000
0
100
200
300C
B
D
10 73 21217310
Samanta (leaf)
Samanta (crown)
Sandra (leaf)
Sandra (crown)
217310
LEAVES
IAA
(p
mo
l/g
FW
)
0
20
40
60
80 A
F ic acidi ent (4
acccw
J
oowe
E
blSfS
C
lamice3s
P
i
the spring-type cereals (e.g., Limin and Fowler, 2002). Even underoptimal growth conditions, Samanta exhibited a higher basal FT, asindicated by a lower LT50, which coincided with detectable levels
CK
X A
CT
IVIT
Y
(nm
ol A
de/m
g p
rote
in.h
)
0.0
0.1
0.2
0.3
0.4
217310
Samanta (leaf)
Sandra (leaf)
ig. 6. Levels of auxin (indole-3-acetic acid, IAA) (A), salicylic acid (SA) (B), jasmonn leaves and crowns of Samanta and Sandra at 0, 1, 3, 7 and 21 days of cold treatm
A decrease in SA levels was observed in the Samanta crownsfter 1 day, followed by SA peak between 3 and 7 days. Prolongedold treatment was associated with a decrease of SA. In Sandrarowns, the downregulation of SA was much more pronouncedompared to Samanta, with a minimum being detected after 3 days,hen a gradual elevation of SA began.
asmonic acid
Cold exposure resulted in an early decrease in JA in the leavesf both cultivars (Fig. 6C). At 7 days, elevation of JA in leaves wasbserved. The elevated JA content was maintained in Samanta,hile another decrease was found in Sandra. In crowns, Samanta
xhibited lower JA levels compared to Sandra.
thylene (ACC)
The immediate precursor of ethylene, ACC, exhibited distinctiphasic behaviour with maxima after 1 and 7 days in Samanta
eaves. A similar but much more dramatic profile was detected inandra leaves (Fig. 6D). In crowns, a mild decrease after 3 days wasollowed by a gradual increase, which was significantly faster inamanta.
ytokinin oxidase/dehydrogenase activity
The time course of CKX activity exhibited similar profiles in theeaves of both cultivars (Fig. 7). Soon after exposure to the cold,
significant decrease in leaf CKX activity was observed with ainimum occurring after 3 days. Downregulation of CKX activity
ndicates a diminished necessity to deactivate bioactive CK underonditions of decreased CK biosynthesis (as reflected by low lev-ls of CK phosphates at 1 day) or during bioactive CK elevation (at
days). Prolonged cold treatment was associated with a gradualtimulation of CKX activity.
rincipal component analysis (PCA) of all data
When PCA was performed on all data obtained in the exper-ment, the most prominent differences were found between
(JA) (C) and the ethylene precursor aminocyclopropane carboxylic acid (ACC) (D)◦C).
different plant organs – leaves and crowns (Suppl. Fig. IA). Ofthe individual characteristics (LT50, WCS120, individual phytohor-mones), the largest differences were found in JA and SA levels withrespect to the remaining characteristics (LT50, WCS120, other phy-tohormones; Suppl. Fig. IB).
Discussion
Control conditions (day 0)
To characterize the changes in the levels of individual phyto-hormones accompanying the responses of wheat plants to coldtreatment, the time course of acclimation was compared in the highfrost tolerant winter cultivar Samanta and the low frost tolerantspring cultivar Sandra. The cultivars differ in the VRN1 gene, withthe winter cultivar Samanta possessing only recessive winter-typevrn1 alleles while spring cultivar Sandra possessing a dominantspring-type Vrn1 allele. It is well-known that winter-type cere-als can acclimate to lower LT50 under inducing conditions than
COLD TREATMENT (DAY)
Fig. 7. The activity of cytokinin oxidase/dehydrogenase (CKX) in leaves of Samantaand Sandra at 0, 1, 3, 7 and 21 days of cold treatment (4 ◦C).

5 nt Ph
o2geiVpc
A
wigrtimAps
suts2
tAmosctdrmwprards
aofeai
aSata
ognwo
74 K. Kosová et al. / Journal of Pla
f WCS120. Previous studies by our group (Vitamvas et al., 2007,010) have demonstrated that this dehydrin can be regarded as aood marker for cold stress tolerance. At 20 ◦C, the detectable lev-ls of WCS120 correlated with higher ABA levels in Samanta leavesn comparison with Sandra. This is in accordance with a report byeisz et al. (1996), who found higher ABA content in a winter com-ared to a spring cultivar. A higher ABA content in Samanta crownsoincided with lower levels of positive growth regulators CKs.
larm phase of the cold response (day 1)
This response phase can be characterized by changes associatedith drop in water potential and its component (turgor pressure),
nduction of a regulon of cold-regulated genes and suppression ofrowth (Gusta et al., 2009). This initial phase of the cold stressesponse coincided with a rapid increase in ABA. The time scale ofransient ABA elevation was similar in Samanta and Sandra, bothn leaves and crowns; however, the amplitude of the response was
uch higher in Samanta leaves and especially in Samanta crowns. transient increase in ABA upon cold stress appears to be a generalhenomenon as such a response has been reported in a number ofpecies (Gusta et al., 2005; Pociecha et al., 2009).
An important function of ABA during the early phase of coldtress is stabilization of water relationships, which involves the reg-lation of stomata aperture in leaves (to diminish the water loss ando allow re-establishment of turgor), and stimulation of the expres-ion of many genes associated with an increase in FT (Gusta et al.,005), e.g., the Wcs120 gene family.
It is interesting that genes for negative type-A response regula-ors of CK signalling (ARR5, ARR6 and ARR7) showed in cold-stressedrabidopsis similar expression profiles as CBF1 (a transient maxi-um between 2 and 4 h) (Jeon et al., 2010). The cold inducibility
f type-A ARRs indicates rapid downregulation (switch-off) of CKignal transduction upon cold exposure. Our data indicate that theold response of Samanta was associated with a decrease of bioac-ive CK content in leaves. In crowns, a decrease of bioactive CKs,elayed to 3 days, was observed in both cultivars. This delay mighteflect the fact that crowns are less exposed to adverse environ-ental conditions than leaves. Downregulation of bioactive CKs (asell as of CK signal transduction) may be associated with fast sup-ression of growth. In Samanta leaves, it may also contribute to theegulation of stomata aperture, as CKs are known to induce stom-ta opening (She and Song, 2006). This early phase of cold stressesponse was also reported to be associated with fast transientown-regulation of photosynthetic rate in cold resistant Arabidop-is accession (Nagele et al., 2011).
In A. thaliana, cold induction of CBF1 was reported to be associ-ted with the accumulation of DELLA proteins, which are repressorsf the GA signalling pathway (Achard et al., 2008). CBF1 wasound to stimulate the expression of genes for the GA-inactivatingnzymes (GA 2-oxidases). In accordance with this report, we found
mild decrease of GA1 accompanied by a dramatic elevation ofnactive hydroxylated forms (GA29 and GA8) in Samanta leaves.
Similarly, another growth-promoting hormone, IAA, showed decrease in the leaves of both genotypes. A recent report byhibasaki et al. (2009) indicates that cold stress (12 h) might alsoffect auxin transport by inhibiting the intracellular trafficking ofhe auxin efflux carriers PIN2 and PIN3. These authors did not findny change in auxin signalling.
The downregulation of the content and/or signal transductionf hormones associated with the stimulation of cell division and
rowth indicates that growth suppression is an important compo-ent of the early stress response, which appears to be associatedith the necessity of reallocating energy sources to the synthesisf numerous protective compounds under stress conditions.
ysiology 169 (2012) 567– 576
Considering other stress hormones, the ethylene precursor ACCexhibited rapid stimulation in the leaves and crowns of both geno-types. This quick reaction might be associated with ROS generation.The upregulation of ABA content coincided with rapid downregu-lation of SA and JA in the leaves and crowns of both genotypes.This finding is in accordance with a report by Pociecha et al. (2009),who found higher ABA levels together with lower SA content inmore frost resistant genotypes of Festulolium during the first 54 hof the cold stress response. ABA treatment also suppresses lowtemperature-induced SA production in maize leaves (Pal et al.,2011). These results reflect the antagonistic relationship amongthese three stress hormones. An antagonistic relationship was alsosuggested between ABA and ethylene (Sharp et al., 2000). Unfor-tunately, the time scale of our experiments has not allowed us toevaluate this hypothesis. However, as elevated ACC content wasfound apart of cold stress also at the beginning of drought stress,it seems to be more probable that ethylene could be generallyinvolved in the stress responses, irrespective of the type of stress.
Acclimation phase (days 3–7)
The characteristic features of this period are an increase inacquired FT (LT50) as well as a decrease in osmotic potential andfresh weight achieved by accumulation of sugars and other protec-tive compounds (e.g. osmoprotectants and antioxidants) (Larcher,2003; Prasil et al., 2004, 2007; Gusta et al., 2009). Our data on theacquired FT levels and WCS120 content show increase in both geno-types; the response of the winter cultivar being stronger than thespring one, which is in accordance with our previous results (Prasilet al., 2004; Vitamvas et al., 2010). Phytohormone changes, charac-teristic for this response phase, were downregulation of ABA levelsand an elevation of positive regulators of cell division and growth.At the beginning of this period, ABA levels were downregulatedsimilarly in both genotypes. The levels of the bioactive CKs, GA1and IAA exhibited in the leaves transient maxima between 3 and 7days. An increase of CK levels coincided with the minimal activityof the main CK-degrading enzyme, CKX, in leaves.
In crowns, the increase in CK and GA1 levels was delayed (to 7days). The meristematic character of these tissues coincided withmuch higher CK and GA contents in crowns compared to leaves.The increase in CKs, GAs and auxin (together with the accumula-tion of dehydrins, especially WCS120) indicates plant adaptation tolow temperature and a readjustment of metabolic activity to lessfavourable conditions. Winter cereals should not only be able tosurvive cold/frost periods, but they also need to grow and develop atlow temperature to achieve sufficient FT (Janda et al., 2007). Plantsmust maintain a sufficient efficiency of photosynthetic processesto provide energy for their growth as well as for the developmentof FT. As CKs exert a positive effect on plant photosynthesis, theirelevation during the adaptation phase might contribute to energyformation. This conclusion is in accordance with a report by Huet al. (2005), who found both improved maintenance of chlorophylllevels and enhanced cold tolerance in calluses of Festuca arund-inacea constitutively expressing the CK biosynthetic gene. Similarresults have been obtained by Guo et al. (2010) using ipt-expressingArabidopsis plants.
It is interesting that constitutive overexpression of ARR7 (a neg-ative type-A CK response regulator, the expression of which isstimulated during the initial phase of the cold response) resultsin longer-term in hypersensitivity to freezing temperatures (Jeonet al., 2010). In contrast, an arr7 knockout mutant was foundto exhibit increased freezing tolerance. These findings suggest
dynamic regulation of individual hormones during different phasesof the stress response.Similarly to CKs, GA1 content was enhanced during the adap-tation phase of the stress response. Apart from its positive effect

K. Kosová et al. / Journal of Plant Physiology 169 (2012) 567– 576 575
Table 1Dynamics of dehydrin and hormone levels during the cold stress progression in winter (W) and spring (S) wheat cultivar. The close and open triangles indicate increase anddecrease, respectively, of the corresponding parameter. Oblong indicates no change. The size of the symbol reflects the magnitude of the change. Localization of the symbolin the middle of the column indicates similar behaviour in both genotypes, while the use of two symbols reflects different behaviour of the winter and spring cultivar.
Stress duration (day)
Alarm phase Acclimation phase Resistance phase
1 3 7 21
W S W S W S W S
WCS120
ABA
Bioactive CKs
Bioactive GAs
IAA
JA
SA
oa
wAcraEtd
R
sseaicimghwechJistesomscp
ACC
CKX
n growth, GA1 might be involved in repression of the promoterctivity of GhDREB1 (Shan et al., 2007).
The decrease in ABA during the acclimation phase coincidedith the elevation of other stress hormones during this period. TheCC content exhibited a transient maximum at 7 days, which coin-ided with an elevation of SA. These data are in accordance with aeport by Janda et al. (2007), who found increased free and bound SAs well as its precursor O-hydroxycinnamic acid in winter wheat.levation of SA might be related to SA functioning in the regula-ion of ROS evolution, which is an important part of plant stressefenses. JA began to increase with some delay.
esistance phase (21 days)
LT50 analysis, as well as dehydrin WCS120 quantification,howed a highly significant difference between the winter andpring varieties after 21 days of cold. While Samanta was able tonhance its acquired FT during the entire cold treatment, Sandrachieved its maximal FT after 7 days and was not able to improvet subsequently. This difference in cold tolerance after 21 days wasorrelated with relatively higher concentrations of stress hormonesn the leaves of Samanta compared to Sandra. The ABA content was
aintained at a higher level in Samanta leaves. SA levels exhibited aradual decrease in Samanta leaves, but they were still significantlyigher compared to those in Sandra at the end of the experiment,hich is in accordance with the findings of other studies (e.g., Janda
t al., 2007) that reported elevated SA content, especially of its glu-osylated form, in cold-tolerant cultivars. The JA content was alsoigher in Samanta leaves, increasing between 7 and 21 days. As
A inhibits cell division and growth in a similar manner to ABA,t is possible that JA may replace ABA during cold stress progres-ion to suppress growth. Elevated JA levels correspond well withhe results of Gaudet et al. (2011), who found upregulation of thexpression of the JA synthetic gene, which encodes allene oxideynthase, between 21 and 49 days of cold exposure. Simultane-usly, they found increased resistance of winter wheat to snow
ould, powdery mildew and stripe rust pathogens. These findingsuggest that tolerance to biotic stresses is also promoted duringold hardening. ACC levels were significantly higher in crowns com-ared to leaves, but they did not differ between the genotypes.
The observed ACC elevation in crowns is in accordance with thereported upregulation of ethylene responsive factors during laterstages of cold hardening (Gaudet et al., 2011).
Strongly enhanced FT in Samanta coincided with lower levels ofgrowth-promoting hormones CKs, GAs and IAA in both the leavesand crowns. The significant downregulation of CK content foundin Samanta leaves and crowns differed remarkably from what wasseen in Sandra, which reached its maximal acclimation on 7 daysand maintained relatively high CK levels from that time until theend of experiment. Thus, the maintenance of CK levels and a mildelevation of IAA levels coincided with a constant level of acquiredFT in Sandra. The achieved results were summarized in Table 1.
Conclusion
Dehydrin and hormone analyses allowed distinction of threephases of the cold stress response: the alarm, acclimation andresistance phases. After cold exposure, both the winter and springcultivars began to increase their endogenous ABA levels, whichcontributed to elevation of the acquired FT and WCS120 pro-tein levels. The winter cultivar presented a simultaneous decreasein the levels of growth-promoting hormones bioactive CKs andauxin and enhanced GA deactivation through GA 2-oxidase. Alongwith cold treatment progression (3–7 days), a significant accu-mulation of WCS120 protein was observed in both cultivars,which was considerably higher in the winter genotype. The lev-els of growth-promoting hormones (IAA, bioactive CKs, bioactiveGAs – GA1, GA4) began to rise, which might indicate an adjust-ment of plant metabolism to growth and development under lowtemperatures. In the spring cultivar, restoration of the levels ofgrowth-promoting hormones was accompanied by cessation of theincrease of acquired FT and WCS120 accumulation. In contrast, thewinter cultivar continued to increase its acquired FT and WCS120protein content until the end of experiment (21 days of cold treat-ment), which was associated with a significant decrease in thecontent of growth-promoting hormones. The stress hormones SA
and JA exhibited mild decreases at the beginning of cold expo-sure, which coincided with an increase in ABA. During prolongedcold treatment, SA and JA contents were gradually elevated, whichmight enhance plant tolerance to multiple stresses.
5 nt Ph
A
pb
A
t
R
A
A
D
G
G
G
G
G
H
J
J
J
K
L
L
76 K. Kosová et al. / Journal of Pla
cknowledgements
This work was supported by the Czech Science Foundation,roject nos. 522/09/2058 and P501/11/P637, and by the MEYS CR,ilateral project nos. MEB040713 and MEB040924.
ppendix A. Supplementary data
Supplementary data associated with this article can be found inhe online version, at doi:10.1016/j.jplph.2011.12.013.
eferences
chard P, Gong F, Cheminant S, Alioua M, Hedden P, Genschik P. The cold-inducibleCBF1 factor-dependent signaling pathway modulates the accumulation of thegrowth-repressing DELLA proteins via its effect on gibberellin metabolism. PlantCell 2008;20:2117–29.
yaz FA, Hayirlioglu-Ayaz S, Gruz J, Novak O, Strnad M. Separation, characteriza-tion, and quantitation of phenolic acids in a little-known blueberry (Vacciniumarctostaphylos L.) fruit by HPLC–MS. J Agric Food Chem 2005;53:8116–22.
obrev P, Kaminek M. Fast and efficient separation of cytokinins from auxin andabscisic acid and their purification using mixed-mode solid-phase extraction. JChromatogr A 2002;950:21–9.
aliba G, Tuberosa R, Kocsy G, Sutka J. Involvement of chromosome 5A and 5D incold-induced abscisic acid accumulation in and frost tolerance of wheat calli.Plant Breeding 1993;110:237–42.
audet DA, Wang Y, Frick M, Puchalski B, Penniket C, Quellet T, et al. Low tempera-ture induced defence gene expression in winter wheat in relation to resistanceto snow moulds and other wheat disease. Plant Sci 2011;180:99–110.
uo JC, Duan RJ, Hu XW, Li KM, Fu SP. Isopentenyl transferase gene (ipt) downstreamtranscriptionally fused with gene expression improves the growth of transgenicplants. Transgenic Res 2010;18:197–209.
usta LV, Trischuk R, Weiser CJ. Plant cold acclimation: the role of abscisic acid. JPlant Growth Regul 2005;24:308–18.
usta LV, Wisniewski ME, Trischuk RG. Patterns of freezing in plants: the influenceof species, environment and experimental procedures. In: Gusta LV, WisniewskiME, Tanino KK, editors. Plant cold hardiness. From the laboratory to the field.CAB International; 2009. pp. 214–25.
u YL, Jia WL, Wang JD, Zhang YQ, Yang LL, Lin ZP. Transgenic tall fescue containingthe Agrobacterium tumefaciens ipt gene shows enhanced cold tolerance. PlantCell Rep 2005;23:705–9.
anda T, Szalai G, Leskó K, Yordanova R, Apostol S, Popova LP. Factors contributingto enhanced freezing tolerance in wheat during frost hardening in the light.Phytochemistry 2007;68:1674–82.
anacek J, Prasil IT. Quantification of plant frost injury by nonlinear fitting of anS-shaped function. CryoLetters 1991;12:47–52.
eon J, Kim NY, Kim S, Kang NY, Novak O, Ku SJ, et al. A subset of cytokinin two-component signaling system plays a role in cold temperature stress response inArabidopsis. J Biol Chem 2010;285:23369–84.
osova K, Vitamvas P, Prasil I, Renaut J. Plant proteome changes under abiotic stress– contribution of proteomics studies to understanding plant stress response. JProteomics 2011;74:1301–22.
archer W. Physiological plant ecology. 4th ed. Berlin, Heidelberg: Springer Verlag;2003.
imin AE, Fowler DB. Developmental traits affecting low-temperature toleranceresponse in near-isogenic lines for the vernalization locus Vrn-A1 in wheat(Triticum aestivum L. em Thell). Ann Bot 2002;89:579–85.
ysiology 169 (2012) 567– 576
Motyka V, Vankova R, Capkova V, Petrasek J, Kaminek M, Schmulling T. Cytokinin-induced upregulation of cytokinin oxidase activity in tobacco includeschanges in enzyme glycosylation and secretion. Physiol Plant 2003;117:11–21.
Nagele T, Kandel BA, Frana S, Meissner M, Heyer AG. A systems biology approach forthe analysis of carbohydrate dynamics during acclimation to low temperaturein Arabidopsis thaliana. FEBS J 2011;278:506–18.
Novak O, Tarkowski P, Tarkowska D, Dolezal K, Lenobel R, Strnad M. Quantitativeanalysis of cytokinins in plants by liquid chromatography/single-quadrupolemass spectrometry. Anal Chim Acta 2003;480:207–18.
Novak O, Hauserová E, Amakorova P, Dolezal K, Strnad M. Cytokinin profiling in planttissues using ultra-performance liquid chromatography–electrospray tandemmass spectrometry. Phytochemistry 2008;69:2214–24.
Pal M, Janda T, Szalai G. Abscisic acid may alter the salicylic acid-related abiotic stressresponse in maize. J Agron Crop Sci 2011., doi:10.1111/j.1439-037X. 00474.x.
Pencik A, Rolcik J, Novak O, Magnus V, Bartak P, Buchtik R, et al. Isolation ofnovel indole-3-acetic acid conjugates by immunoaffinity extraction. Talanta2009;80:651–5.
Pociecha E, Plazek A, Janowiak F, Waligorski P, Zwierzykowski Z. Changes in abscisicacid, salicylic acid and phenylpropanoid concentrations during cold acclimationof androgenic forms of Festulolium (Festuca pratensis × Lolium multiflorum) inrelation to resistance to pink snow mould (Microdochium nivale). Plant Breeding2009;128:397–403.
Prasil IT, Prasilova P, Marik P. Comparative study of direct and indirect evaluationsof frost tolerance in barley. Field Crop Res 2007;102:1–8.
Prasil IT, Prasilova P, Pankova K. Relationships among vernalization, shoot apexdevelopment and frost tolerance in wheat. Ann Bot 2004;94:413–8.
Prasil IT, Zamecnik J. The use of a conductivity measurement method for assessingfreezing injury. I. Influence of leakage time, segment number, size and shapein a sample on evaluation of the degree of injury. Environ Exp Bot 1998;40:1–10.
Sarhan F, Ouellet F, Vazquez-Tello A. The wheat wcs120 gene family. A useful modelto understand the molecular genetics of freezing tolerance in cereals. PhysiolPlant 1997;101:439–45.
Shan DP, Huang JG, Yang YT, Guo YH, Wu CA, Yang GD, et al. Cotton GhDREB1increases plant tolerance to low temperature and is negatively regulated bygibberellic acid. New Phytol 2007;176:70–81.
Sharp RE, LeNole ME, Else MA, Thorne ET, Gherardi F. Endogenous ABA maintainsshoot growth in tomato independently of effects on plant water balance: evi-dence for an interaction with ethylene. J Exp Bot 2000;51:1575–84.
She XP, Song XG. Cytokinin- and auxin-induced stomatal opening is related tothe change of nitric oxide levels in guard cells in broad bean. Physiol Plant2006;128:569–79.
Shibasaki K, Uemura M, Tsurumi S, Rahman A. Auxin response in Arabidop-sis under cold stress: underlying molecular mechanisms. Plant Cell 2009;21:3823–38.
Tureckova V, Novak O, Strnad M. Profiling ABA metabolites in Nicotiana tabacum L.leaves by ultra-performance liquid chromatography–electrospray tandem massspectrometry. Talanta 2009;80:390–9.
Vankova R. Plant hormone functions in abiotic and biotic stress responses. In: Pes-sarakli M, editor. Handbook of plant and crop stress. 3rd ed. Boca Raton, FL, USA:CRC Press, Taylor and Francis; 2010. p. 191–211.
Veisz O, Galiba G, Sutka J. Effect of abscisic acid on the cold hardiness of wheatseedlings. J Plant Physiol 1996;149:439–43.
Vitamvas P, Kosova K, Prasilova P, Prasil IT. Accumulation of WCS120 protein in
wheat cultivars grown at 9 ◦C or 17 ◦C in relation to their winter survival. PlantBreeding 2010;129:611–6.Vitamvas P, Saalbach G, Prasil IT, Capkova V, Opatrna J, Jahoor A. WCS120 proteinfamily and proteins soluble upon boiling in cold-acclimated winter wheat. J PlantPhysiol 2007;164:1197–207.