complementary stimulation of hepatobiliary transport and detoxification systems by rifampicin and...
TRANSCRIPT

C
CDi
HULC*a§
U
BcmlmmtRwcitmutptdtUacohMUutsteTtc
GASTROENTEROLOGY 2005;129:476–485
LINICAL–LIVER, PANCREAS, AND BILIARY TRACT
omplementary Stimulation of Hepatobiliary Transport andetoxification Systems by Rifampicin and Ursodeoxycholic Acid
n Humans
ANNS–ULRICH MARSCHALL,* MARTIN WAGNER,‡ GERNOT ZOLLNER,‡ PETER FICKERT,‡
LF DICZFALUSY,§ JUDITH GUMHOLD,‡ DAGMAR SILBERT,‡ ANDREA FUCHSBICHLER,�
ISBET BENTHIN,* ROSITA GRUNDSTRÖM,¶ ULF GUSTAFSSON,¶ STAFFAN SAHLIN,¶
URT EINARSSON,* and MICHAEL TRAUNER‡
Department of Medicine, Karolinska Institutet, Karolinska University Hospital Huddinge, Stockholm, Sweden; ‡Laboratory of Experimentalnd Molecular Hepatology, Division of Gastroenterology and Hepatology, Department of Medicine, Medical University, Graz, Austria;Laboratory Medicine, Karolinska Institutet, Karolinska University Hospital Huddinge, Stockholm, Sweden; �Institute of Pathology, Medical
niversity, Graz, Austria; and ¶Department of Surgery, Danderyds Hospital, Stockholm, SwedenUlfilbcittfipt
waPtpmb
B7DCtOct
ackground & Aims: Rifampicin (RIFA) and ursodeoxy-holic acid (UDCA) improve symptoms and biochemicalarkers of liver injury in cholestatic liver diseases by
argely unknown mechanisms. We aimed to study theolecular mechanisms of action of these drugs in hu-ans. Methods: Thirty otherwise healthy gallstone pa-
ients scheduled for cholestectomy were randomized toIFA (600 mg/day for 1 week) or UDCA (1 g/day for 3eeks) or no medication before surgery. Routine bio-hemistry, lipids, and surrogate markers for P450 activ-ty (4�-hydroxy cholesterol, 4�-OH-C) and bile acid syn-hesis (7�-hydroxy-4-cholesten-3-one, C-4) wereeasured in serum. Bile acids were analyzed in serum,
rine, and bile. A wedge liver biopsy specimen wasaken to study expression of hepatobiliary ABC trans-orters as well as detoxification enzymes and regulatoryranscription factors. Results: RIFA enhanced bile acidetoxification as well as bilirubin conjugation and excre-ion as reflected by enhanced expression of CYP3A4,GT1A1, and MRP2. These molecular effects were par-lleled by decreased bilirubin and deoxycholic acid con-entrations in serum and decreased lithocholic and de-xycholic acid concentrations in bile. UDCA on the otherand stimulated the expression of BSEP, MDR3, andRP4. UDCA became the predominant bile acid afterDCA treatment and lowered the biliary cholesterol sat-ration index. Conclusions: RIFA enhances bile acid de-oxification as well as bilirubin conjugation and exportystems, whereas UDCA stimulates the expression ofransporters for canalicular and basolateral bile acidxport as well as the canalicular phospholipid flippase.hese independent but complementary effects may jus-ify a combination of both agents for the treatment of
holestatic liver diseases.rsodeoxycholic acid (UDCA) improves clinical andbiochemical serum parameters in a variety of cho-
estatic liver diseases.1 UDCA is nowadays considered therst-line treatment for patients with chronic cholestaticiver diseases such as primary biliary cirrhosis (PBC)ecause a combined analysis of the 3 largest randomizedlinical trials of UDCA in PBC indicates that UDCAmproves survival free of liver transplantation.2 However,he efficacy of UDCA treatment has been debated,3,4 andhe mechanism(s) of action in humans are still not de-ned.1 Suggested mechanisms of UDCA include im-roved bile acid transport and/or detoxification, cytopro-ection, and antiapoptotic effects.1
Before the era of UDCA, there have been small trialsith rifampicin (RIFA) in cholestatic liver disease, which
ll showed a marked improvement of pruritus.5–11 InBC, RIFA also significantly decreased serum levels ofransaminases,7 �-glutamyltransferase,7,10 alkaline phos-hatase,7 and total bile salts.5,7 Nowadays, RIFA isainly used for treatment of pruritus not responding to
ile acid sequestrates.12 A proposed molecular mecha-
Abbreviations used in this paper: 4�-OH-C, 4�-hydroxy cholesterol;AAT, bile acid amino transferase; BSEP, bile salt export pump; C4,�-hydroxy-4-cholesten-3-one; CA, cholic acid; CYP, cytochrome P450;CA, deoxycholic acid; HCA, hyocholic acid; LCA, lithocholic acid;DCA, chenodeoxycholic acid; HDCA, hyodeoxycholic acid; MDR, mul-idrug resistance protein; MRP, multidrug resistance-related protein;ATP, organic anion transporting polypeptide; PBC, primary biliaryirrhosis; RIFA, rifampicin; UGT, uridine diphosphate glucuronosyl-ransferase; UDCA, ursodeoxycholic acid.
© 2005 by the American Gastroenterological Association0016-5085/05/$30.00
doi:10.1053/j.gastro.2005.05.009

npavmzrmm
tcegtd
ctubiispnwae
t(wgS(pIw
paSCtsc
pwcwtTem
ddfdi(bw
gRmbCSPSaAuprafiascaoparfgtic
t
August 2005 EFFECTS OF RIFAMPICIN AND URSODEOXYCHOLIC ACID 477
ism of action of RIFA is the hydroxylation of hydro-hobic bile acids, thereby decreasing their cytotoxicity,n effect mediated by stimulation of CYP3A expressionia activation of the nuclear receptor SXR/PXR inice.13,14 Moreover, decreased expression of the key en-
yme of bile acid synthesis, Cyp7a1, was observed inodents.13 Again, the anticholestatic and antipruriticode of action of RIFA in humans remains to be deter-ined.To identify potential molecular mechanism(s) of ac-
ion of UDCA and RIFA in humans, we performed aomprehensive biochemical and molecular study of theffects of these agents on bile acids, biliary lipids, andenes involved in bile acid/bilirubin metabolism15 andransport16 in otherwise healthy patients with gallstoneisease.
Patients and Methods
Patients
Otherwise healthy patients scheduled for laparascopicholestectomy for symptomatic gallstone disease were invitedo participate in a clinical study of the metabolic and molec-lar effects of UDCA and RIFA. This approach was takenecause such a detailed analysis of liver, bile, serum, and urines not feasible in patients not scheduled for a surgical abdom-nal procedure. Potential candidates completed a clinical re-earch file consisting of a detailed questionnaire about theatient’s history and lifestyle. Previous or ongoing liver, kid-ey, intestinal, or metabolic diseases were exclusion criteria asell as the use of medications known to affect liver function
nd metabolism, eg, lipid or glucose lowering drugs, P450nzyme inducers.
After giving a written informed consent, 30 patients be-ween 47 and 58 years of age were randomized to RIFARimactan, Sandoz, Basel, Switzerland, 600 mg/day for 1eek) or UDCA (Ursofalk, Dr Falk, Freiburg, Germany, 1/day for 3 weeks) or no medication (controls) before surgery.tudy drugs were administered open-labeled in 1 (RIFA) or 2UDCA) daily doses until the day before surgery. The studyrotocol was approved by the ethics committee at Karolinskanstitutet. The prevalence of cholesterol or pigment gallstonesas judged visually (by C.E.).
Clinical Chemical and Bile Acid Analyses inSerum and Urine
Routine parameters including serum lipids and li-oproteins as well as liver and kidney function tests werenalyzed using standard clinical biochemical procedures.erum 4�-hydroxycholesterol (4�-OH-C), a marker ofYP3A4,17 7�-hydroxy-4-cholesten-3-one (C-4), a sensi-
ive marker of endogenous bile acid synthesis,18 as well aserum bile acids were analyzed by isotope-dilution gas
hromatography-mass spectrometry (GCMS) as described ureviously.18 –20 From morning urine samples, bile acidsere isolated using solid-phase extraction and screened for
onjugates using electrospray mass spectrometry (ESMS),hich is a highly sensitive method for detection of different
ypes of conjugated bile acids, as previously described.21
hese analyses were performed at the beginning and at thend of the treatment periods in patients randomized toedication.
Biliary Lipid Analyses
During surgery, gallbladder bile was sampled for theetermination of cholesterol, phospholipids, and bile acids asescribed previously.22 In addition, bile aliquots were screenedor bile acid conjugates other than glycine- or taurine-ami-ates. For this purpose, bile acids were extracted after disrupt-ng protein adsorption by incubating with 1 mL 0.5 mol/LNHEt4)2SO4 at 64°C for 30 minutes and analyzed by ESMSoth before and after cleavage of glycine or taurine conjugatesith cholylglycine hydrolase.21
mRNA and Protein Expression Analyses
A wedge liver biopsy specimen was taken during sur-ery and immediately frozen in liquid nitrogen. MessengerNA (mRNA) and protein were then prepared for the deter-ination of mRNA expression levels of enzymes involved in
ile acid synthesis and metabolism15 (BAAT; CYP3A4,YP7A1, CYP27, CYP8B1, UGT1A1, UGT2B4, UGT2B7,ULT2A1), regulatory nuclear transcription factors (HNF-4�,XR/NR1I2, RXR/NR1B1), and mRNA levels (OATP1B1/LC21A1) and protein as well as mRNA levels of hepatobili-ry ABC transporters16 (ABCG8, BSEP/ABCB11, MDR3/BCB4, MRP2/ABCC2, MRP3/ABCC3, MRP4/ABCC4)sing recently in-detail-described methodology.23–25 Humanrimer and probe sets used for mRNA estimations by TaqManeal-time PCR and competitive RT-PCR, respectively, arevailable on request. Liver biopsy specimen material was suf-cient to perform the complete set of mRNA expressionnalyses from all 30 patients included. For Western analyses,ufficient protein was available from all patients serving asontrols but only from 9 and 8 patients randomized to RIFAnd UDCA, respectively. Protein expression levels from RIFAr UDCA treated patients were estimated in comparison withroteins prepared from controls, with a sample from a patientssigned to RIFA or UDCA treatment groups serving aseference (R in Figures 4 and 5). To verify that antibodies usedor Western analyses estimated expression of membrane-tar-eted ABC transport proteins, tissue distribution of canalicularransporters BSEP, MDR3, and MRP2 was investigated bymmunofluorescence labeling and confocal laser scanning mi-roscopy as described.26
Statistical Analysis
Data are expressed as mean values � standard devia-ion (SD). Differences within patient groups were analyzed
sing Student paired t test and, among patient groups, un-
p(c
U(2wsg7s
pksitRs
UeRwaw(Om
tiocbctU
pcc1304
T
SAB
T
TLHAALTBAA�ACUAT
Na
478 MARSCHALL ET AL GASTROENTEROLOGY Vol. 129, No. 2
aired Student t test using the SigmaStat statistic programJandel Scientific, San Rafael, CA). A P value of �.05 wasonsidered significant.
Results
Patients’ Demographics, RoutineBiochemistry and Type of Gallstones
The patients randomized to control, RIFA, orDCA groups did not differ significantly in mean age
range, 29–77 years of age) or body mass index (range,4.4–35.9 kg/m2). Females outnumbered males (in lineith the target population in cholestatic liver diseases
uch as PBC), and there were more women in the UDCAroup (n � 9) than in the RIFA (n � 6) or control (n �) groups (Table 1). All patients had cholesterol gall-tones.
No adverse effects from study medications were re-orted. Serum lipids and lipoproteins as well as liver andidney function tests remained in the normal range in alltudy patients and did not change significantly from thenitial value (Table 2). Of interest, serum bilirubin washe only marker that decreased in each patient in theIFA group. For the entire RIFA group, this change was
tatistically significant (P � .05).
able 1. Characteristics of Patients Studied
Control RIFA UDCA
ex 7 F, 3 M 6 F, 4 M 9 F, 1 Mge (y) 58.5 � 7.2 48.6 � 9.8 47.2 � 10.0MI (kg/m2) 26.5 � 2.2 25.1 � 3.0 26.0 � 3.5
able 2. Serum Biochemistry of Patients Studied
Control Be
otal cholesterol (�5.2 mmol/L) 5.8 � 0.8 5.6DL cholesterol (�3.4 mmol/L) 3.4 � 0.8 3.3DL cholesterol (�1.4 mmol/L) 1.5 � 0.3 1.7PO A1 (�1.4 mmol/L) 1.5 � 0.2 1.5PO B (�0.8 mmol/L) 1.1 � 0.1 1.0p(a) (�0.3 mmol/L) 0.2 � 0.2 0.3riglycerides (�2.0 mmol/L) 1.5 � 0.4 1.2ilirubin (�26 �mol/L) 13.0 � 2.3 12.3LAT (�36 U/L) 21.6 � 5.4 22.8SAT (�36 U/L) 25.2 � 5.4 25.2GT (�48 U/L) 37.2 � 28.8 49.2LP (�270 U/L) 178.8 � 43.2 174.0reatinine (45–84 mmol/L) 77.8 � 6.4 83.7rea (3–10 mmol/L) 4.4 � 0.6 5.1lbumin (34–47 mmol/L) 41.1 � 1.5 42.7otal bile acids (�5 �mol/L) 1.1 � 0.4 0.9
OTE. Normal range in parentheses.
P � .05, Student paired t test.Compliance
Compliance of all patients randomized to RIFA orDCA was confirmed by ESMS analyses of urine sample
xtracts taken at the ends of the study periods. EitherIFA (molecular weight, 822 g/mol) or its metabolitesere found with quasimolecular ions between m/z 821
nd 682 or N-acetylglucosamine conjugates of UDCA,27
ith or without additional glycine- or taurine-amidationsee below). In addition, increased formation of 4�-H-C was found in any patient randomized to studyedication17 (see below and Figure 1).
Effects of RIFA and UDCA on Biliary Lipidsand Bile Acids
Both RIFA and UDCA significantly increasedotal lipid concentrations in gallbladder bile because ofncreased bile acid concentrations (Table 3). However,nly UDCA patients had a significantly lower biliaryholesterol saturation index as compared with controlsecause patients randomized to RIFA also had increasedholesterol concentrations in bile. Phospholipid concen-rations did not differ between control, RIFA, andDCA groups (Table 3).RIFA significantly lowered secondary but increased
rimary bile acids in bile, as shown in Figure 2B. Litho-holic acid (LCA), deoxycholic acid (DCA), chenodeoxy-holic acid (CDCA), and cholic acid (CA) accounted for.4% � 0.9%, 32.8% � 9.1%, 32.5% � 6.8%, and3.0% � 8.0%, respectively, in bile of controls and.6% � 0.2%, 5.8% � 4.6%, 44.8% � 7.1%, and8.3% � 8.1%, respectively, in patients administered
RIFA UDCA
After Before After
9 5.7 � 0.9 5.7 � 1.1 5.8 � 1.07 3.4 � 0.9 3.1 � 1.1 3.5 � 1.02 1.8 � 0.3 1.7 � 0.4 1.8 � 0.41 1.6 � 0.2 1.5 � 0.3 1.6 � 0.32 1.0 � 0.2 1.0 � 0.2 1.0 � 0.33 0.4 � 0.3 0.2 � 0.2 0.2 � 0.24 1.1 � 0.3 1.4 � 0.7 1.4 � 0.69 7.9 � 1.9a 11.1 � 5.1 13.7 � 7.12 28.2 � 5.4 26.4 � 10.2 36.6 � 20.44 28.8 � 3.0 27.0 � 4.8 26.4 � 4.8.8 45.6 � 21.0 27.6 � 15.6 54.0 � 57.6.4 184.2 � 43.2 179.4 � 30.0 198.6 � 37.8.0 42.7 � 2.2 77.4 � 8.8 79.1 � 1.9
3 5.3 � 0.7 4.8 � 0.9 4.3 � 0.52 43.2 � 1.8 40.7 � 1.9 4.1 � 1.53 0.8 � 0.2 0.8 � 0.3 2.1 � 0.7a
fore
� 0.� 0.� 0.� 0.� 0.� 0.� 0.� 4.� 7.� 5.� 37� 41� 13� 7.� 2.� 0.

R9cpCstapH(b
goM
abeoc
tfctatsUb
esashparcao
s(ili1wl
FacTctb
T
TCPBC
a
August 2005 EFFECTS OF RIFAMPICIN AND URSODEOXYCHOLIC ACID 479
IFA. In UDCA patients, UDCA contributed 47.4% �.6% of total bile acids compared with 0.8% � 0.9% inontrols and 0.6% � 0.6% in RIFA patients. In UDCAatients, the relative amounts of DCA (12.2% � 9.6%),DCA (17.0% � 9.6%), and CA (22.1% � 8.0%) were
ignificantly lower as compared with controls, whereashe relative amount of LCA (1.3% � 0.7%) was the sames in controls, very similar to the bile acid composition inatients with PBC after 2-year treatment with UDCA.28
owever, only RIFA changed concentrations of primaryhigher) and secondary (lower) bile acids in gallbladderile, as shown in Figure 2.
In native bile extracts from control, RIFA, and UDCAroups, ESMS only showed anions indicative of glycine-r taurine-conjugated di- or trihydroxylated bile acids.inor anions at m/z 455 and 471 indicating small
igure 1. Effects of RIFA and UDCA on cytochrome P450 (A) and bilecid synthesis (B). The serum marker of CYP3A4 induction 4�-hydroxy-holesterol increased in each patient treated with RIFA or UDCA (A).he serum marker for endogenous bile acid synthesis 7�-hydroxy-4-holestene-3-one increased significantly only in the group of patientsreated with RIFA (B). P � .05: significantly different compared withaseline. RIFA, solid line; UDCA, broken line.
able 3. Biliary Lipid Composition After Treatment With Rifamin 3 Weeks
Control
otal lipids (mmol/L) 95.1 � 26.2holesterol (mmol/L) 9.3 � 3.5hospholipids (mmol/L) 28.0 � 10.5ile acids (mmol/L) 58.9 � 26.9holesterol Saturation Index (%) 127.1 � 40.3
P � .05, treatment vs controls, Student paired t test.
mounts (�5%) of sulfated mono- and dihydroxylatedile acids were first seen after treatment of bile acidxtracts with cholylglycine hydrolysis. Anions indicativef glycosidic bile acid conjugates, ie, glucuronides, glu-osides, or N-acetylglucosaminides were not recorded.
Effects of RIFA and UDCA on Serum andUrine Bile Acids
In contrast to bile, total serum bile acid concen-rations in patients administered RIFA did not differrom controls (Table 2). However, as in bile, these non-holestatic patients had significantly lower DCA concen-rations (Figure 2), whereas the concentrations of CDCAnd CA were the same in all study groups. Patientsreated with UDCA had 2-fold higher (P � .05) totalerum bile acids (Table 2) because of the abundance ofDCA (58.4% � 12.4%). Serum LCA was not evaluatedecause of levels �.05 �mol/L.Screening for bile acid metabolites in native urine
xtracts using ESMS indicated the renal excretion ofimilar RIFA metabolites as found in bile (see above). Inddition, anions at m/z 567, 583, 624, and 640 repre-enting nonamidated and glycine-amidated di- and tri-ydroxy bile acid glucuronides emerged.29 In urine fromatients administered UDCA, anions at m/z 594, 651,nd 701 representing nonamidated and glycine- or tau-ine-amidated dihydroxy bile acid N-acetylglucosamineonjugates21,24 were recorded (see above). No ions indic-tive of any bile acid metabolite were recorded in urinef patients before treatment or controls.
Effects of RIFA and UDCA on MetabolicGenes
RIFA as the prototype cytochrome P450 inducerignificantly increased hepatic CYP3A4 gene expressionFigure 3). This was also reflected by the highly signif-cant increase (246.5% � 65.4% compared with base-ine) in the formation of 4�-OH-C (Figure 1). UDCAncreased the formation of this metabolite as well (Figure) but to a much lesser extent (37.5% � 13.0%), whichas not reflected in increased CYP3A4 gene expression
evels (Figure 3). RIFA patients expressed significantly
n, 600 mg/day in 1 Week, or Ursodeoxycholic Acid, 1 g/day
RIFA UDCA
180.2 � 62.1a 215.6 � 51.7a
17.7 � 7.9a 7.0 � 2.031.5 � 16.8 45.2 � 16.2
130.6 � 47.7a 163.4 � 37.8a
172.8 � 61.7 48.4 � 5.2a
pici

lurCpa(pa(
afUegR
Mas
btMp
ochpcstuwu
maBclf
FgaFtaot
Fnbfoa
480 MARSCHALL ET AL GASTROENTEROLOGY Vol. 129, No. 2
ower RXR mRNA levels compared with controls (Fig-re 3). Genes of HNF-4� and PXR, nuclear factorsegulation CYP3A4 expression, and genes of CYP7A1,YP27, and CYP8B1, enzymes regulating alternativeathways of bile acid synthesis in humans were expressedt similar levels in control, RIFA, and UDCA groupsFigure 3). However, as estimated by serum bile acidrecursor C-4, a small but significantly increased bilecid synthesis rate was found in RIFA patientsFigure 1).
Expression levels of genes encoding enzymes for bilecid conjugation with glycine- or taurine (BAAT), sul-ate (SULT2A1) or glucuronidate (UGT2B4 andGT2B7) did not differ between study groups. How-
ver, expression levels of UGT1A1 coding for bilirubinlucuronosyltransferase were significantly higher inIFA patients (Figure 3).
Effects of RIFA and UDCA on HepatobiliaryABC Transporter Gene and ProteinExpression
RIFA patients expressed significantly higherRP2 mRNA levels compared with controls (Figure 3)
nd conversely showed significantly more protein expres-
igure 2. Effects of RIFA and UDCA on bile acids in serum (A) andallbladder (B) bile. Serum (A) and biliary (B) bile acid concentrationst the end of treatment with RIFA or UDCA, compared with controls.or easier comparison, baseline serum data that did not differ be-ween groups are not shown. LCA, lithocholic acid; DCA, deoxycholiccid; CDCA, chenodeoxycholic acid; CA, cholic acid; UDCA, ursode-xycholic acids. P � .05: significantly different compared with con-rols.
ion (Figure 4) of MRP2, responsible for the excretion of .
ilirubin glucuronides. Expression levels of other hepa-obiliary ABC transporters (ABCG8, BSEP, MDR3,RP3, MRP4, and OATP1B1) did not differ in RIFA
atients from controls.In UDCA patients, mRNA expression levels of none
f the hepatobiliary ABC transporter genes differed fromontrols. However, in UDCA patients, significantlyigher protein levels of dedicated hepatobiliary exportroteins were found by Western blotting. These were theanalicular BSEP and MDR3 proteins, exporting bilealts and phospholipids, and the basolateral MRP4 pro-ein, exporting bile salt conjugates (Figure 5). Canalic-lar membrane localization of BSEP, MDR3, and MRP2as confirmed by immunofluorescence microscopy (Fig-re 6).
Discussion
The present study was performed to define theolecular mechanisms of action of 2 drugs—RIFA5,7,8
nd UDCA (recently reviewed by Paumgartner andeuers1)—that were shown to improve serum liver bio-hemistry and symptoms in patients with cholestaticiver diseases such as primary biliary cirrhosis (PBC). Weound that RIFA and UDCA both enhance expression of
igure 3. Effects of RIFA and UDCA on hepatic mRNA expression ofuclear receptors and hepatobiliary transport systems (A) and biliru-in and bile acid metabolizing enzymes (B). Total RNA was isolatedrom liver biopsy specimens of patients randomized to control, RIFA,r UDCA and analyzed by real-time PCR as described in the Materialsnd Methods section. Data are expressed as percentage � SD. P �
05: significantly different compared with controls.
dmi
nobctspofwsfbrp
ahvsm
mtRcnlcbLtac
FaUMic
Fbotl
August 2005 EFFECTS OF RIFAMPICIN AND URSODEOXYCHOLIC ACID 481
istinct but complementary sets of bile acid/bilirubinetabolizing/detoxifying enzymes and their correspond-
ng hepatobiliary ABC transporter proteins.
RIFA Lowers Serum Bilirubin Levels andAlters Bile Acid Metabolism
We provide evidence that RIFA improves elimi-ation of bilirubin by coordinately enhancing expressionf the bilirubin-conjugating enzyme UGT1A1 and theilirubin diglucuronide excreting MRP2 protein at theanalicular membrane. In fact, the only biochemical rou-ine parameter that significantly changed during ourtudy was serum bilirubin, a finding that may be ex-lained by these molecular RIFA effects. Our findings intherwise healthy gallstones patients are in line with datarom a long-term study with RIFA in PBC patients inhich a trend toward lower bilirubin levels was de-
cribed8 and by the observation of a complete remissionrom cholestasis with very high conjugated hyperbiliru-inemia in 3 women on RIFA, 2 of them with benignecurrent intrahepatic cholestasis.30 However, we herein
igure 4. Effects of RIFA on hepatic protein expression of hepatobili-ry transport proteins. Proteins were isolated from controls, RIFA, orDCA patients and analyzed by Western blotting as described in theaterials and Methods section. Protein expression levels calculated
n relation to indicated (R) reference. P � .05: significantly differentompared with controls.
rovide for the first time the potential molecular mech- d
nism underlying this important clinical effect. The en-anced hepatobiliary excretion of conjugated bilirubinia MRP2 may be further assisted by enhanced expres-ion of this protein in luminal membrane of intestinalucosa.31
We also found RIFA to alter significantly bile acidetabolism. Lowering of systemic and presumably in-
rahepatic levels of bile acids during treatment withIFA was seen as an explanation for the relief from
holestatic pruritus5,8 that was consistently observed in aumber of trials with RIFA.5–8,11 However, the under-ying molecular mechanisms have so far remained un-lear. We found a significant reduction of the secondaryile acid DCA in serum, confirming a recent study byütjohann et al,32 and, in addition, a significant reduc-ion of biliary levels of both hydrophobic secondary bilecids, ie, lithocholic acid (LCA) and DCA. However, theoncentrations of primary bile acids, CA and CDCA, and
igure 5. Effects of UDCA on hepatic protein expression of hepato-iliary transport proteins. Proteins were isolated from controls, RIFA,r UDCA patients and analyzed by Western blotting as described inhe Materials and Methods section. Protein expression levels calcu-ated in relation to indicated (R) reference. P � .05: significantly
ifferent compared with controls.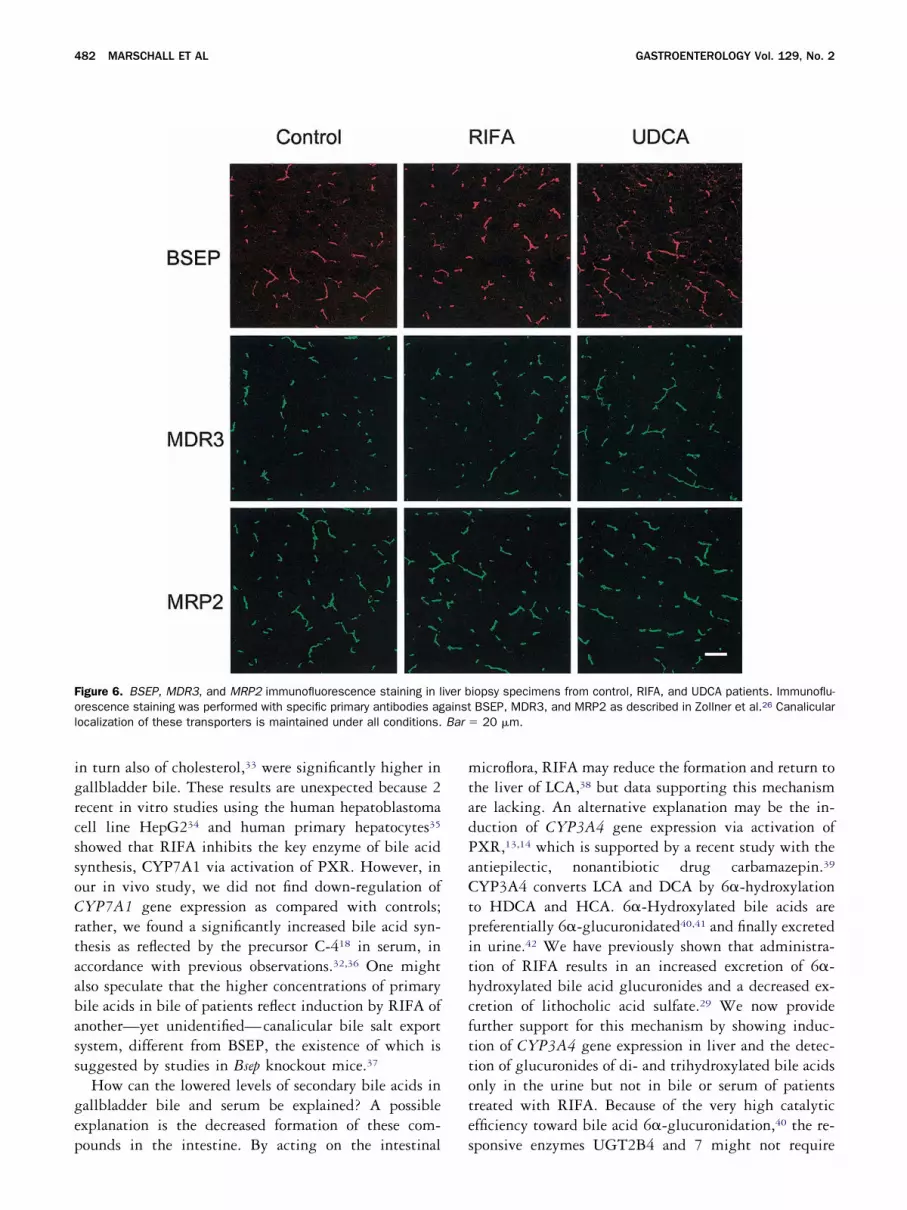
igrcssoCrtaabass
gep
mtadPaCtpithcfttote
Fol Bar �
482 MARSCHALL ET AL GASTROENTEROLOGY Vol. 129, No. 2
n turn also of cholesterol,33 were significantly higher inallbladder bile. These results are unexpected because 2ecent in vitro studies using the human hepatoblastomaell line HepG234 and human primary hepatocytes35
howed that RIFA inhibits the key enzyme of bile acidynthesis, CYP7A1 via activation of PXR. However, inur in vivo study, we did not find down-regulation ofYP7A1 gene expression as compared with controls;
ather, we found a significantly increased bile acid syn-hesis as reflected by the precursor C-418 in serum, inccordance with previous observations.32,36 One mightlso speculate that the higher concentrations of primaryile acids in bile of patients reflect induction by RIFA ofnother—yet unidentified—canalicular bile salt exportystem, different from BSEP, the existence of which isuggested by studies in Bsep knockout mice.37
How can the lowered levels of secondary bile acids inallbladder bile and serum be explained? A possiblexplanation is the decreased formation of these com-
igure 6. BSEP, MDR3, and MRP2 immunofluorescence staining in lrescence staining was performed with specific primary antibodies agocalization of these transporters is maintained under all conditions.
ounds in the intestine. By acting on the intestinal s
icroflora, RIFA may reduce the formation and return tohe liver of LCA,38 but data supporting this mechanismre lacking. An alternative explanation may be the in-uction of CYP3A4 gene expression via activation ofXR,13,14 which is supported by a recent study with thentiepilectic, nonantibiotic drug carbamazepin.39
YP3A4 converts LCA and DCA by 6�-hydroxylationo HDCA and HCA. 6�-Hydroxylated bile acids arereferentially 6�-glucuronidated40,41 and finally excretedn urine.42 We have previously shown that administra-ion of RIFA results in an increased excretion of 6�-ydroxylated bile acid glucuronides and a decreased ex-retion of lithocholic acid sulfate.29 We now provideurther support for this mechanism by showing induc-ion of CYP3A4 gene expression in liver and the detec-ion of glucuronides of di- and trihydroxylated bile acidsnly in the urine but not in bile or serum of patientsreated with RIFA. Because of the very high catalyticfficiency toward bile acid 6�-glucuronidation,40 the re-
iopsy specimens from control, RIFA, and UDCA patients. Immunoflu-BSEP, MDR3, and MRP2 as described in Zollner et al.26 Canalicular
20 �m.
iver bainst
ponsive enzymes UGT2B4 and 7 might not require

scMinsdIgt
iHtcoohUma1tm
otcahihpBcbdpcsMlaOhswf
UhMppaibt
etaRriamtwdspse
FUmittabopettaci
August 2005 EFFECTS OF RIFAMPICIN AND URSODEOXYCHOLIC ACID 483
timulation of expression by RIFA. These bile acid glu-uronides are most likely excreted from the liver viaRP3.43,44 Of note, MRP3 was found to be up-regulated
n PBC.23 However, the antipruritic effect of RIFA can-ot be attributed to CYP3A4 induction alone because aimilar increase in CYP3A4 activity by phenobarbitoneid not result in a comparable clinical improvement.7
mproved excretion of other still unidentified prurito-enic compounds via induced MRP2 could further con-ribute to this effect.
UDCA Induces Expression of Phase IMetabolic Enzymes and Transporters inHuman Liver
UDCA was also found to enhance CYP3A4 activ-ty as reflected by increased serum 4�-OH-C levels.17
owever, compared with RIFA, formation of this me-abolite by UDCA was minor and not reflected byhanges in CYP3A4 gene expression levels or reductionsf secondary bile acids. Thus, PXR-dependent inductionf CYP3A4 by UDCA as shown in primary humanepatocytes45 seems to be a minor mechanism of action ofDCA.1 As to bile acid metabolism, we found enrich-ent of UDCA by approximately 50% in bile, leading tosignificantly lower lithogenic index (48%; in controls,27%), and by approximately 70% in serum, as well ashe abundance of N-acetylglucosamine conjugates as theajor metabolites of UDCA in urine.21,27,41
Most important, however, were the effects of UDCAn the expression of hepatobiliary ABC transporter pro-eins. Because of marked species differences in bile acidomposition and transporter regulation between humansnd rodents, it was essential to study UDCA effects inumans. We now directly demonstrate for the first timen humans that UDCA enhances protein expression ofepatobiliary membrane transport proteins: the phos-holipid transporter MDR3 and the bile salt transporterSEP at the canalicular site and the alternative bile saltonjugate export protein MRP4 at the basolateral mem-rane. The changes in MDR3 and BSEP are in accor-ance with the enhanced excretion of bile acid andhospholipids in patients with primary sclerosingholangitis during UDCA treatment.46 From animaltudies, it was known that UDCA enhances Bsep andrp2 protein expression47 and insertion into the cana-
icular membrane.48,49 Functionally, Bsep insertion wasccompanied by increased excretion of taurocholic acid.48
f note, our previous study in mice did not find en-anced gene expression levels of Bsep indicating posttran-criptional regulation of Bsep protein expression,47
hich we now confirm even in humans. In mice, UDCA
eeding induced hepatic MRP4 expression.50 This aDCA effect is also confirmed in our present study inumans. In contrast, UDCA feeding induced Mrp2 andrp3 only in mice51 but not in man as now shown in the
resent study. The induction of key hepatobiliary trans-ort proteins by UDCA in humans may contribute to itsnticholestatic and hepatoprotective effects by stimulat-ng both biliary/canalicular excretory and alternative/asolateral excretory routes for bile acids and other po-entially toxic biliary constituents.
Which Conclusions Can Be Drawn FromOur Data for Clinical Practice?
UDCA is the first-line treatment of PBC, but itsffect on pruritus is notoriously poor. Bile acid seques-rates are often added under this condition, but thesegents may interfere with the resorption of UDCA. Thus,IFA seems to be more suitable, but clinicians are
eluctant to prescribe RIFA because of the risk of drug-nduced hepatitis, which has been reported to occur inpproximately 10% of patients during long-term treat-ent.8,52 This, of course, warrants careful monitoring of
ransaminases. Induction of CYP3A4 may also interfereith the metabolism of other prescribed drugs. However,eterioration of cholestasis as suggested from in vitrotudies showing RIFA to inhibit the bile salt exportump53,54 is unlikely to occur from our data that actuallyhowed higher biliary bile salt concentrations. In fact,levations of serum bile acids have only been observed
igure 7. Schematic representation of observed effects of RIFA andDCA on hepatobiliary transport systems and bile acid and bilirubinetabolizing enzymes. RIFA enhances excretion of bilirubin by (1)
nducing the bilirubin glucuronyl transferase UGT1A1 and (2) inducinghe bilirubin-glucuronide transport protein MDR2; RIFA as the proto-ype inducer of CYP3A4 facilitates conversion of hydrophobic bilecids (LCA, CDCA) into hydrophilic 6�-hydroxylated compounds thatecome glucuronidated and most likely are excreted via MRP3, thenly transport system known so far to be intrinsically up-regulated inrimary biliary cirrhosis. UDCA on the other hand induces proteinxpression of various transport systems including BSEP for the excre-ion of bile salts and MDR3 for the excretion of phospholipids, both athe canalicular pole, and MRP4 for the excretion of bile salts/organicnion conjugates at the basolateral side of the hepatocyte. Theseomplementary effects may help to reduce cholestatic liver injurynduced by bile acids and other potentially toxic biliary constituents.
fter single high-dose RIFA55 treatment but not in long-

ta(cuttaecpc
ortsepjdef
1
1
1
1
1
1
1
1
1
1
2
2
2
2
2
2
2
2
2
484 MARSCHALL ET AL GASTROENTEROLOGY Vol. 129, No. 2
erm treatment.5,7,8 From our data showing independentnd mutual supportive effects on the detoxificationCYP3A4, UGT1A1) and elimination of hepatotoxicompounds (BSEP, MRP2, MRP4) (Figure 7), we spec-late that, at least in advanced PBC, a combinationherapy with RIFA and UDCA might be more effectivehan single standard UDCA treatment. Provided thebsence of obstructive cholestasis where enhanced biliaryxcretion of bile acids via BSEP might be deleterious,56
ombination therapy might be even more important inrimary sclerosing cholangitis, which often is compli-ated by severe pruritus.57
In summary, RIFA was found to enhance eliminationf bilirubin by increasing UGT1A1 expression and up-egulation of MRP2 in addition to induce phase I de-oxification of bile acids by increased CYP3A4 expres-ion. UDCA is a weak CYP3A4 inducer but significantlynhances expression of hepatobiliary bile salt (BSEP),hospholipid (MDR3), and organic anion-bile salt con-ugate/glutathione (MRP4) transporters. These indepen-ent but complementary hepatobiliary transporter andnzyme effects support the combined use of both agentsor the treatment of advanced cholestatic liver disease.
References1. Paumgartner G, Beuers U. Mechanisms of action and therapeutic
efficacy of ursodeoxycholic acid in cholestatic liver disease. ClinLiver Dis 2004;8:67–81.
2. Poupon RE, Lindor KD, Cauch-Dudek K, Dickson ER, Poupon R,Heathcote EJ. Combined analysis of randomized controlled trialsof ursodeoxycholic acid in primary biliary cirrhosis. Gastroenter-ology 1997;113:884–890.
3. Goulis J, Leandro G, Burroughs AK. Randomised controlled trialsof ursodeoxycholic-acid therapy for primary biliary cirrhosis: ameta-analysis. Lancet 1999;354:1053–1060.
4. Poupon RE. Ursodeoxycholic acid for primary biliary cirrhosis:lessons from the past—issues for the future. J Hepatol 2000;32:685–688.
5. Hoensch HP, Balzer K, Dylewizc P, Kirch W, Goebell H, OhnhausEE. Effect of rifampicin treatment on hepatic drug metabolismand serum bile acids in patients with primary biliary cirrhosis. EurJ Clin Pharmacol 1985;28:475–477.
6. Ghent CN, Carruthers SG. Treatment of pruritus in primary biliarycirrhosis with rifampin. Results of a double-blind, crossover, ran-domized trial. Gastroenterology 1988;94:488–493.
7. Bachs L, Pares A, Elena M, Piera C, Rodes J. Comparison ofrifampicin with phenobarbitone for treatment of pruritus in biliarycirrhosis. Lancet 1989;1:574–576.
8. Bachs L, Pares A, Elena M, Piera C, Rodes J. Effects of long-termrifampicin administration in primary biliary cirrhosis. Gastroenter-ology 1992;102:2077–2080.
9. Podesta A, Lopez P, Terg R, Villamil F, Flores D, Mastai R,Udaondo CB, Companc JP. Treatment of pruritus of primary biliarycirrhosis with rifampin. Dig Dis Sci 1991;36:216–220.
0. Yerushalmi B, Sokol RJ, Narkewicz MR, Smith D, Karrer FM. Useof rifampin for severe pruritus in children with chronic cholesta-sis. J Pediatr Gastroenterol Nutr 1999;29:442–447.
1. Cynamon HA, Andres JM, Iafrate RP. Rifampin relieves pruritus inchildren with cholestatic liver disease. Gastroenterology 1990;
98:1013–1016.2. Hofmann AF. Rifampicin and treatment of cholestatic pruritus.Gut 2002;51:756–757.
3. Staudinger JL, Goodwin B, Jones SA, Hawkins-Brown D, MacKen-zie KI, LaTour A, Liu Y, Klaassen CD, Brown KK, Reinhard J,Willson TM, Koller BH, Kliewer SA. The nuclear receptor PXR is alithocholic acid sensor that protects against liver toxicity. ProcNatl Acad Sci U S A 2001;98:3369–3374.
4. Xie W, Radominska-Pandya A, Shi Y, Simon CM, Nelson MC, OngES, Waxman DJ, Evans RM. An essential role for nuclear recep-tors SXR/PXR in detoxification of cholestatic bile acids. Proc NatlAcad Sci U S A 2001;98:3375–3380.
5. Chiang JY. Regulation of bile acid synthesis: pathways, nuclearreceptors, and mechanisms. J Hepatol 2004;40:539–551.
6. Trauner M, Boyer JL. Bile salt transporters: molecular character-ization, function, and regulation. Physiol Rev 2003;83:633–671.
7. Bodin K, Andersson U, Rystedt E, Ellis E, Norlin M, Pikuleva I,Eggertsen G, Bjorkhem I, Diczfalusy U. Metabolism of 4 �-hy-droxycholesterol in humans. J Biol Chem 2002;277:31534–31540.
8. Axelson M, Bjorkhem I, Reihner E, Einarsson K. The plasma levelof 7 �-hydroxy-4-cholesten-3-one reflects the activity of hepaticcholesterol 7 �-hydroxylase in man. FEBS Lett 1991;284:216–218.
9. Bodin K, Bretillon L, Aden Y, Bertilsson L, Broome U, EinarssonC, Diczfalusy U. Antiepileptic drugs increase plasma levels of4�-hydroxycholesterol in humans: evidence for involvement ofcytochrome p450 3A4. J Biol Chem 2001;276:38685–38689.
0. Ewerth S, Angelin B, Einarsson K, Nilsell K, Bjorkhem I. Serumconcentrations of ursodeoxycholic acid in portal venous and sys-temic venous blood of fasting humans as determined by isotopedilution-mass spectrometry. Gastroenterology 1985;88:126–133.
1. Marschall HU, Broome U, Einarsson C, Alvelius G, Thomas HG,Matern S. Isoursodeoxycholic acid: metabolism and therapeuticeffects in primary biliary cirrhosis. J Lipid Res 2001;42:735–742.
2. Gustafsson U, Sahlin S, Einarsson C. Biliary lipid composition inpatients with cholesterol and pigment gallstones and gallstone-free subjects: deoxycholic acid does not contribute to formationof cholesterol gallstones. Eur J Clin Invest 2000;30:1099–1106.
3. Zollner G, Fickert P, Silbert D, Fuchsbichler A, Marschall HU,Zatloukal K, Denk H, Trauner M. Adaptive changes in hepatobili-ary transporter expression in primary biliary cirrhosis. J Hepatol2003;38:717–727.
4. Wagner M, Fickert P, Zollner G, Fuchsbichler A, Silbert D,Tsybrovskyy O, Zatloukal K, Guo GL, Schuetz JD, Gonzalez FJ,Marschall HU, Denk H, Trauner M. Role of farnesoid X receptor indetermining hepatic ABC transporter expression and liver injury inbile duct-ligated mice. Gastroenterology 2003;125:825–838.
5. Fickert P, Fuchsbichler A, Wagner M, Zollner G, Kaser A, Tilg H,Krause R, Lammert F, Langner C, Zatloukal K, Marschall HU,Denk H, Trauner M. Regurgitation of bile acids from leaky bileducts causes sclerosing cholangitis in Mdr2 (Abcb4) knockoutmice. Gastroenterology 2004;127:261–274.
6. Zollner G, Fickert P, Zenz R, Fuchsbichler A, Stumptner C, KennerL, Ferenci P, Stauber RE, Krejs GJ, Denk H, Zatloukal K, TraunerM. Hepatobiliary transporter expression in percutaneous liverbiopsies of patients with cholestatic liver diseases. Hepatology2001;33:633–646.
7. Marschall HU, Griffiths WJ, Gotze U, Zhang J, Wietholtz H, BuschN, Sjovall J, Matern S. The major metabolites of ursodeoxycholicacid in human urine are conjugated with N-acetylglucosamine.Hepatology 1994;20:845–853.
8. Combes B, Carithers RL Jr, Maddrey WC, Munoz S, Garcia-TsaoG, Bonner GF, Boyer JL, Luketic VA, Shiffman ML, Peters MG,White H, Zetterman RK, Risser R, Rossi SS, Hofmann AF. Biliarybile acids in primary biliary cirrhosis: effect of ursodeoxycholic
acid. Hepatology 1999;29:1649–1654.
2
3
3
3
3
3
3
3
3
3
3
4
4
4
4
4
4
4
4
4
4
5
5
5
5
5
5
5
5
KtSm
JgA
August 2005 EFFECTS OF RIFAMPICIN AND URSODEOXYCHOLIC ACID 485
9. Wietholtz H, Marschall HU, Sjovall J, Matern S. Stimulation of bileacid 6 �-hydroxylation by rifampin. J Hepatol 1996;24:713–718.
0. Cancado EL, Leitao RM, Carrilho FJ, Laudanna AA. Unexpectedclinical remission of cholestasis after rifampicin therapy in pa-tients with normal or slightly increased levels of gamma-glutamyltranspeptidase. Am J Gastroenterol 1998;93:1510–1517.
1. Fromm MF, Kauffmann HM, Fritz P, Burk O, Kroemer HK, WarzokRW, Eichelbaum M, Siegmund W, Schrenk D. The effect of ri-fampin treatment on intestinal expression of human MRP trans-porters. Am J Pathol 2000;157:1575–1580.
2. Lutjohann D, Hahn C, Prange W, Sudhop T, Axelson M, Sauer-bruch T, von Bergmann K, Reichel C. Influence of rifampin onserum markers of cholesterol and bile acid synthesis in men. IntJ Clin Pharmacol Ther 2004;42:307–313.
3. Northfield TC, Hofmann AF. Biliary lipid output during three mealsand an overnight fast. I. Relationship to bile acid pool size andcholesterol saturation of bile in gallstone and control subjects.Gut 1975;16:1–11.
4. Bhalla S, Ozalp C, Fang S, Xiang L, Kemper JK. Ligand-activatedpregnane X receptor interferes with HNF-4 signaling by targetinga common coactivator PGC-1�: functional implications in hepaticcholesterol and glucose metabolism. J Biol Chem 2004;279:45139–45147.
5. Li T, Chiang JY. Mechanism of rifampicin and pregnane X receptor(PXR) inhibition of human cholesterol 7�-hydroxylase gene(CYP7A1) transcription. Am J Physiol Gastrointest Liver Physiol2005;288:G74–G84.
6. von Bergmann K, Fierer J, Mok HY, Grundy SM. Effects of rifampinon biliary lipids in humans. Antimicrob Agents Chemother 1981;19:342–345.
7. Wang R, Lam P, Liu L, Forrest D, Yousef IM, Mignault D, PhillipsMJ, Ling V. Severe cholestasis induced by cholic acid feeding inknockout mice of sister of P-glycoprotein. Hepatology 2003;38:1489–1499.
8. Setchell KDR, Street MS, Sjovall J. Fecal bile acids. In: SetchellKDR, Kritchevsky D, Nair PP, eds. The bile acids. Vol. 4. Methodsand applications. New York: Plenum Press, 1988:441–570.
9. Bodin K, Lindbom U, Diczfalusy U. Novel pathways of bile acidmetabolism involving CYP3A4. Biochim Biophys Acta 2005;1687:84–93.
0. Marschall HU, Matern H, Egestad B, Matern S, Sjovall S. 6�-Glucuronidation of hyodeoxycholic acid by human liver, kidneyand small bowel microsomes. Biochim Biophys Acta 1987;921:392–397.
1. Marschall HU, Matern H, Wietholtz H, Egestad B, Matern S,Sjovall J. Bile acid N-acetylglucosaminidation. In vivo and in vitroevidence for a selective conjugation reaction of 7 �-hydroxylatedbile acids in humans. J Clin Invest 1992;89:1981–1987.
2. Alme B, Sjovall J. Analysis of bile acid glucuronides in urine.Identification of 3 �, 6 �, 12 �-trihydroxy-5 �-cholanoic acid. JSteroid Biochem 1980;13:907–916.
3. Zelcer N, Saeki T, Reid G, Beijnen JH, Borst P. Characterization ofdrug transport by the human multidrug resistance protein 3(ABCC3). J Biol Chem 2001;276:46400–46407.
4. Zelcer N, Reid G, Borst P, Marschall HU. Basolateral hepaticexport of bile acid glucuronides via mrp3: an important route forbile acid detoxification? Gastroenterology 2003;124(Suppl 1):113.
5. Schuetz EG, Strom S, Yasuda K, Lecureur V, Assem M, Brimer C,Lamba J, Kim RB, Ramachandran V, Komoroski BJ, Venkatara-manan R, Cai H, Sinal CJ, Gonzalez FJ, Schuetz JD. Disrupted bileacid homeostasis reveals an unexpected interaction among nu-clear hormone receptors, transporters, and cytochrome P450.
J Biol Chem 2001;276:39411–39418.6. Stiehl A, Rudolph G, Sauer P, Theilmann L. Biliary secretion ofbile acids and lipids in primary sclerosing cholangitis. Influence ofcholestasis and effect of ursodeoxycholic acid treatment. J Hepa-tol 1995;23:283–289.
7. Fickert P, Zollner G, Fuchsbichler A, Stumptner C, Pojer C, Zenz R,Lammert F, Stieger B, Meier PJ, Zatloukal K, Denk H, Trauner M.Effects of ursodeoxycholic and cholic acid feeding on hepatocel-lular transporter expression in mouse liver. Gastroenterology2001;121:170–183.
8. Kurz AK, Graf D, Schmitt M, Vom DS, Haussinger D. Taurour-sodesoxycholate-induced choleresis involves p38(MAPK) activa-tion and translocation of the bile salt export pump in rats. Gas-troenterology 2001;121:407–419.
9. Beuers U, Bilzer M, Chittattu A, Kullak-Ublick GA, Keppler D,Paumgartner G, Dombrowski F. Tauroursodeoxycholic acid in-serts the apical conjugate export pump, Mrp2, into canalicularmembranes and stimulates organic anion secretion by proteinkinase C-dependent mechanisms in cholestatic rat liver. Hepa-tology 2001;33:1206–1216.
0. Zollner G, Halilbasic E, Wagner M, Fickert P, Silbert D, GumholdJ, Fuchsbichler A, Zatlaukal K, Marschall HU, Denk H, Trauner M.Cholic acid (CA) and ursodeoxycholic acid (UDCA) exert differenteffects on hepatic and renal expression of Mrp4 and Mrp6, twonovel ABC transporters. Gastroenterology 2004;126(Suppl 2):115.
1. Zollner G, Fickert P, Fuchsbichler A, Silbert D, Wagner M, ArbeiterS, Gonzalez FJ, Marschall HU, Zatloukal K, Denk H, Trauner M.Role of nuclear bile acid receptor, FXR, in adaptive ABC trans-porter regulation by cholic and ursodeoxycholic acid in mouseliver, kidney and intestine. J Hepatol 2003;39:480–488.
2. Prince MI, Burt AD, Jones DE. Hepatitis and liver dysfunction withrifampicin therapy for pruritus in primary biliary cirrhosis. Gut2002;50:436–439.
3. Stieger B, Fattinger K, Madon J, Kullak-Ublick GA, Meier PJ. Drug-and estrogen-induced cholestasis through inhibition of the hep-atocellular bile salt export pump (Bsep) of rat liver. Gastroenter-ology 2000;118:422–430.
4. Byrne JA, Strautnieks SS, Mieli-Vergani G, Higgins CF, Linton KJ,Thompson RJ. The human bile salt export pump: characterizationof substrate specificity and identification of inhibitors. Gastroen-terology 2002;123:1649–1658.
5. Galeazzi R, Lorenzini I, Orlandi F. Rifampicin-induced elevation ofserum bile acids in man. Dig Dis Sci 1980;25:108–112.
6. Fickert P, Zollner G, Fuchsbichler A, Stumptner C, Weiglein AH,Lammert F, Marschall HU, Tsybrovskyy O, Zatloukal K, Denk H,Trauner M. Ursodeoxycholic acid aggravates bile infarcts in bileduct-ligated and Mdr2 knockout mice via disruption of cholangi-oles. Gastroenterology 2002;123:1238–1251.
7. Mendes FD, Lindor KD. Primary sclerosing cholangitis. Clin LiverDis 2004;8:195–211.
Received January 25, 2005. Accepted April 20, 2005.Address requests for reprints to: Hanns-Ulrich Marschall, MD, PhD,
arolinska Institutet, Department of Medicine, Division of Gastroen-erology and Hepatology, Karolinska University Hospital Huddinge K63,-141 86 Stockholm, Sweden. e-mail: [email protected]; fax: (46) 8-58582335.Supported by grants from Karolinska Institutet, Ruth och Richard
ulins Fond, and the Swedish Medical Association (to H.U.M.) and byrant 10266 from the Austrian National Bank and P15502 from theustrian Science Foundation (to M.T.).
The authors thank all patients for their participation in this study.