commensalism: was it truly a natufian phenomenon? recent contributions from ethnoarchaeology and...
TRANSCRIPT

edited by
Ofer Bar-Yosef&
François R. Valla
InternatIonal Monographs In prehIstory
Archaeological Series 19
Natufian Foragers in the Levant
Terminal Pleistocene Social Changes in Western Asia

Library of Congress Cataloging-in-Publication Data
Natufian foragers in the Levant : terminal Pleistocene social changes in Western Asia / edited by Ofer Bar-Yosef & François Valla. pages cm. -- (Archaeological series / International Monographs in Prehistory ; 19) Papers from a symposium held in 2009. Includes bibliographical references. ISBN 978-1-879621-45-9 (paperback : acid-free paper) -- ISBN 978-1-879621-46-6 (hard cover : acid-free paper) 1. Natufian culture--Middle East--Congresses. 2. Hunting and gathering societies--Middle East--Congresses. 3. Pleistocene-Holocene boundary--Congresses. 4. Social archaeology--Middle East--Congresses. 5. Social change--Middle East--History--To 1500--Congresses. 6. Excavations (Archaeology)--Middle East--Congresses. 7. Middle East--Antiquities--Congresses. I. Bar-Yosef, Ofer. II. Valla, François Raymond. GN774.3.N38N28 2013 306.3›640956--dc23 2013035516
© 2013 by International Monographs in PrehistoryAll rights reserved
Printed in the United States of AmericaAll rights reserved
Paperback:ISBN 978-1-879621-45-9Hard Cover:ISBN 978-1-879621-46-6
This book is printed on acid-free paper. ∞
International Monographs in PrehistoryAnn Arbor, MichiganU.S.A.
Printed with the support of the American School of Prehistoric Research (Peabody Museum, Harvard University)

Table of Contents
List of Contributors ................................................................................................................... vii
Preface – The Natufian Culture in the Levant: Twenty Years LaterOfer Bar-Yosef and François R. Valla ...............................................................................xv
Acknowledgements ....................................................................................................................xix
Northern Levant
Natufian Lifeways in the Eastern Foothills of the Anti-Lebanon MountainsNicholas J. Conard, Knut Bretzke, Katleen Deckers, Andrew W. Kandel, Mohamed Masri, Hannes Napierala, Simone Riehl and Mareike Stahlschmidt ..............................1
The Natufian of Moghr el-Ahwal in the Qadisha Valley, Northern LebanonAndrew Garrard and Corine Yazbeck ..............................................................................17
The Natufian of Southwestern Syria Sites in the Damascus ProvinceKurt Felix Hillgruber ........................................................................................................28
The Natufian Occupations of Qarassa 3 (Sweida, Southern Syria)Xavier Terradas, Juan José Ibáñez, Franck Braemer, Lionel Gourichon and Luis C. Teira ...................................................................................................................................45
The Early Natufian Site of Jeftelik (Homs Gap, Syria)Amelia del Carmen Rodríguez Rodríguez, Maya Haïdar-Boustani, Jesús E.González Urquijo, Juan José Ibáñez, Michel Al-Maqdissi, Xavier Terradasand Lydia Zapata ..............................................................................................................61
Fish in the Desert? The Younger Dryas and its Influence on the Paleoenvironment at Baaz Rockshelter, Syria
Hannes Napierala .............................................................................................................73
Preliminary Results from Analyses of Charred Plant Remains from a Burnt Natufian Building at Dederiyeh Cave in Northwest Syria
Ken-ichi Tanno, George Willcox, Sultan Muhesen, Yoshihiro Nishiaki, YousefKanjo and Takeru Akazawa..............................................................................................83
Southern Levant
El-Wad
Spatial Organization of Natufian el-Wad through Time: Combining the Results of Past and Present Excavations
Mina Weinstein-Evron, Daniel Kaufman and Reuven Yeshurun ...................................88

iv
The Last Natufian Inhabitants of el-Wad TerraceNoga Bachrach, Israel Hershkovitz, Daniel Kaufman and MinaWeinstein-Evron..............................................................................................................107
Domestic Refuse Maintenance in the Natufian: Faunal Evidence from el-Wad Terrace, Mount Carmel
Reuven Yeshurun, Guy Bar-Oz, Daniel Kaufman and Mina Weinstein-Evron ...........118
Natufian Green Stone Pendants from el-Wad: Characteristics and Cultural ImplicationsDaniella E. Bar-Yosef Mayer, Naomi Porat and Mina Weinstein-Evron ......................139
Eynan
The Final Natufian Structure 215-228 at Mallaha (Eynan), Israel: an Attempt at Spatial Analysis
François R. Valla, Hamoudi Khalaily, Nicolas Samuelian, Anne Bridault, Rivka Rabinovich, Tal Simmons, Gaëlle Le Dosseur and Shoshana Ashkenazi ....................146
A Study of two Natufian Residential Complexes: Structures 200 and 203 at Eynan (Ain Mallaha), Israel
Nicolas Samuelian ..........................................................................................................172
Graves in Context: Field Anthropology and the Investigation of Interstratified Floors and Burials
Fanny Bocquentin, Teresa Cabellos and Nicolas Samuelian ........................................185
Obsidian in Natufian Context: the Case of Eynan (Ain Mallaha), IsraelHamoudi Khalaily and François R. Valla ......................................................................193
Flint Knapping and its Objectives in the Early Natufian. The Example of Eynan- Ain Mallaha (Israel)
Boris Valentin, François R. Valla and Hugues Plisson with the collaboration of Fanny Bocquentin ...........................................................................................................203
Searching for the Functions of Fire Structures in Eynan (Mallaha) and their Formation Processes: a Geochemical Approach
Ramiro J. March ..............................................................................................................227
Avifauna of the Final Natufian of EynanTal Simmons ....................................................................................................................284
Bone Ornamental Elements and Decorated Objects of the Natufian from MallahaGaëlle Le Dosseur and Claudine Maréchal ...................................................................293
Reconstruction of the Habitats in the Ecosystem of the Final Natufian Site of Ain Mallaha (Eynan)
Shoshana Ashkenazi .......................................................................................................312

v
Southern Levant - other sites
Wadi Hammeh 27: an open-air ‘base-camp’ on the Fringe of the Natufian ‘homeland’Phillip C. Edwards, Fanny Bocquentin, Sue Colledge, Yvonne Edwards, Gaëlle Le Dosseur, Louise Martin, Zvonkica Stanin and John Webb ...........................................319
Art Items from Wadi Hammeh 27Janine Major ...................................................................................................................349
The Final Epipaleolithic / PPNA site of Huzuq Musa (Jordan Valley)Dani Nadel and Danny Rosenberg .................................................................................382
Natufian Settlement in the Wadi al-Qusayr, West-Central JordanMichael Neeley ................................................................................................................397
The Steppic Early Natufian: Investigations in the Wadi al-Hasa, JordanDeborah I. Olszewski ......................................................................................................412
The Natufian of the Azraq Basin: An AppraisalTobias Richter and Lisa A. Maher ..................................................................................429
Chert Procurement Patterns And Exploitation Territory: Case Study From Late Natufian Hayonim Terrace (Western Galilee, Israel)
Christophe Delage ...........................................................................................................449
A Faunal Perspective on the Relationship between the Natufian Occupations of Hayonim Cave and Hayonim Terrace
Natalie D. Munro ............................................................................................................463
The Natufian at Raqefet CaveGyörgy Lengyel, Dani Nadel and Fanny Bocquentin ....................................................478
Hof Shahaf: A New Natufian Site on the Shore of Lake KinneretOfer Marder, Reuven Yeshurun, Howard Smithline, Oren Ackermann, Daniella E. Bar-Yosef Mayer, Anna Belfer-Cohen, Leore Grosman, Israel Hershkovitz, Noa Klein and Lior Weissbrod ...............................................................................................505
The Life History of Macrolithic Tools at Hilazon Tachtit CaveLaure Dubreuil and Leore Grosman ..............................................................................527
General Reviews, Climate and Interpretations
Breaking the Mould: Phases and Facies in the Natufian of the Mediterranean ZoneAnna Belfer-Cohen and A. Nigel Goring-Morris ...........................................................544
Ruminations on the Role of Periphery and Center in the NatufianA. Nigel Goring-Morris and Anna Belfer-Cohen ...........................................................562

vi
The Natufian and the Younger DryasDonald O. Henry .............................................................................................................584
Scaphopod Shells in the Natufian CultureAldona Kurzawska, Daniella E. Bar-Yosef Mayer and Henk K. Mienis ......................611
The Natufian Chronological Scheme – New Insights and their ImplicationsLeore Grosman ................................................................................................................622
Natufian Foragers and the ‘Monocot Revolution’: A Phytolith PerspectiveArlene M. Rosen ..............................................................................................................638
Lithic Technology in the Late Natufian – Technological Differences between ‘Core-area’ and ‘Periphery’
Hila Ashkenazy ...............................................................................................................649
Variability of Lunates and Changes in Projectile Weapons Technology during the NatufianAlla Yaroshevich, Daniel Kaufman, Dmitri Nuzhnyy, Ofer Bar-Yosef and Mina Weinstein-Evron..............................................................................................................671
Specialized Hunting of Gazelle in the Natufian: Cultural Cause or Climatic Effect?Guy Bar-Oz, Reuven Yeshurun and Mina Weinstein-Evron .........................................685
Commensalism: was it Truly a Natufian Phenomenon? Recent Cntributions from Ethnoarchaeology and Ecology
Lior Weissbrod, Daniel Kaufman, Dani Nadel, Reuven Yeshurun and Mina Weinstein-Evron..............................................................................................................699

699
Introduction
The commensalism model in archaeology, which was first proposed by Bar-Yosef and Tchernov (1966) links ecological consequences to social and cultural processes in human settlements. According to the model, important changes in the duration and inten-sity of human settlement occupation are expected to result in patterned and predictable changes in the populations and structure of communities of non-domesticated species of small animals (e.g. small rodents and birds) that have long coexisted with humans. Development of the model was tied to research on the Natufian culture of southwest Asia (ca. 16,000/15,000-11,200 cal BP). This is be-cause the Natufian period is widely believed to have involved a shift to sedentary lifeways and a signif-icant increase in the level of human occupation of settlements (e.g. Bar-Yosef and Belfer-Cohen 1989; Belfer-Cohen and Bar-Yosef 2000; Garrod 1957). Use of biological indicators from small animal remains to gauge changes in the level of human settlement occupation was also seen as a less biased method in comparison to other commonly employed indicators that are based on artifact and site characteristics (see Bar-Yosef and Belfer-Cohen 1989; Tchernov 1984). Early evidence for commensalism in the Natu-fian was based on abundant remains of house mice (Mus musculus domesticus) and house sparrows (Passer domesticus) in deposits of Hayonim Cave, northern Israel. These findings suggested that Natufian occupation of the site was long-term and intense based on the known association of such species with modern highly sedentary settlement environments (Bar-Yosef and Tchernov 1966). This inference was further supported by the finding that house mouse remains were rare in earlier deposits from the Final Pleistocene. In later publications, Tchernov elaborated on the theoretical underpin-
Commensalism: Was it Truly a Natufian Phenomenon?
Recent Contributions from Ethnoarcheology and Ecology
Lior Weissbrod, Daniel Kaufman, Dani Nadel, Reuven Yeshurun
and Mina Weinstein-Evron
nings of the commensalism model and together with colleagues provided additional supporting evidence for linking commensalism and Natufian sedentism (Auffray et al. 1988, 1990; Tchernov 1984, 1991a, b). This included evidence from both zooar-chaeological material and from ecological studies of commensalism in contemporary settings. Other authors have undermined the model, however, due to lack of empirical data on the range of settlement contexts wherein commensalism can be expected to develop, including sedentary or more mobile ones (Edwards 1989; Tangri and Wyncoll 1989). Here we assess the validity of the commen-salism-sedentism linkage and its relevance to the Natufian culture. We examine variability in Natufian micromammalian assemblages in light of empirical data from ethnoarcheological and ecological studies in contemporary settlement settings and other human modified environments. In this comparison we employ ecological data from present day settlements of seasonally mobile pastoralists (Weissbrod 2010a, b) and sedentary agriculturalists (Misonne 1963). We use data on living communities of small rodents and shrews (micromammals) associated with the settlements and consider patterns in the organization of these communities including the composition and relative frequencies of species and biological diversity, which may be represented by the number of species and heterogeneity in distribution of frequencies among different species. Data is also presented on assem-blages of micromammalian remains from a number of important Natufian sites and different phases of the Natufian (Fig. 1; Table 1). These samples were retrieved from different ecological and environmen-tal areas of the Mediterranean climate region and may represent different climatic fluctuations. We examine the variability among these samples and indications for commensalism by considering the recent information from ecology and ethnoarcheol-

700
Lior Weissbrod et al.
ogy and information on the archaeological context and taphonomy of the assemblages.
Factors affecting variability among Natufian micromammalian assemblages
The question of sedentarization in settlement occupation in the Natufian and in particular its pre-agricultural origins have been central to dis-cussions on the significance of the Natufian phe-nomenon in the cultural history of southwest Asia (Bar-Yosef 2002; Bar-Yosef and Belfer-Cohen 1989; Belfer-Cohen and Bar-Yosef 2000; Binford 1968; Flannery 1972; Henry 1985; Kaufman 1992; Perrot 1968). A wide range of drivers including climatic and social, limiting or enabling factors have been proposed to account for the appearance of sites with seemingly abrupt increase in size, density of artifacts, scale of construction, storage activities, burial, occurrence of heavy stone implements, and frequency and variety of material expressions of art and decoration. The majority of such Natufian sites with evidence for significant sedentism and all those contributing samples to the present study are con-fined to the Mediterranean climate region of Mount Carmel and the Galilee that are considered the core area of the Natufian culture (Fig. 1) (Bar-Yosef 1983). Evidence has also been considered, however, for significant variability in mode and intensity of settlement occupation both within and between different phases of the Natufian (Bar-Yosef 1983;
Byrd 1989; Valla 1998). Thus, whereas some sites that are located in prime resource locations, typ-ically in ecotonal zones, contain all the landmark characteristics of the Natufian culture and evoke significant levels of occupation (e.g. Bar-Yosef 1991; Valla et al. 2002; Weinstein-Evron 2009) others are of much smaller scale and suggest use on an ephemeral basis possibly for special purpose and/or seasonally scheduled activities such as use of seasonal resources, burial, and ritual/ceremonial practices (e.g. Bar-Yosef 1983; Grosman 2003; Marder et al. herein; Nadel et al. 2008; Valla 1998). Such differences in the function of sites and scale of occupation can also be identified within sites in relation to the different temporal phases of the Natufian and may be tied to either social or climatic developments or both. At the site of el-Wad Terrace on the western coastal aspect of Mount Carmel such a shift is recognized in the transition from the Early Natufian (EN) to the Late Natufian (LN) (Garrod 1957; Weinstein-Evron 2009; Weinstein-Evron et al. herein). Evidence for an extensive settlement in the EN phase includes a complex array of architectural components such as stone walls, slab pavements, and rock-cut ba-sins indicating residential activities in addition to organized burial in the form of concentrations of group and individual burials (Garrod and Bate 1937; Weinstein-Evron 1998, 2009; Weinstein-Evron et al. 2007). This can be contrasted with the LN phase of el-Wad Terrace when a part of the terrace area was utilized mainly as a burial ground and the spatial extent of occupation appears to have become more restricted (Weinstein-Evron et al. 2007). At the open-air Natufian site of Eynan in the Hula basin of the Jordan Valley (Valla and Khalaily 1997; Valla et al. 2002) there is some indication that the construction of structures became progressively less substantial from the EN phase through the LN and a final short phase of the Natufian (Final Natufian, FN). Other sites such as the caves of Raqefet in in-land Mount Carmel (Lengyel et al. 2005; Nadel et al. 2008, 2009) and Hilazon Tachtit in the Galilee Mountains (Grosman 2003; Grosman and Munro 2007), both are belonging to the LN phase, present a very different picture of mode of use and occupation. Both cave sites are situated on steep slopes and at relatively high elevations of 50 m and more above the valley bottom. The archaeological deposits of the two sites mainly occur within the confined space of the caves, which do not include extensive outside terraces as is the case at the larger-scale site of el-Wad, for example. At Raqefet Cave evidence was
Fig. 1. Map of northern Israel showing the location of the three Natufian sites supplying micro-mammalian assemblages to this study.

701
Commensalism: Was it Truly a Natufian Phenomenon?…
unearthed mainly of burial activities in addition to the hewing and utilization of a wide variety of bedrock mortars and cupmarks (Lengyel and Boc-quentin 2005; Nadel and Lengyel 2009). Similarly, a burial context was also uncovered at the cave of Hilazon Tachtit in conjunction with evidence for specialized ritual activities in the form of elaborate grave preparation and distinctive association of grave offerings (Grosman 2003; Grosman et al. 2008). Variability in environmental context and climat-ic fluctuations are additional factors to take into account in comparing micromammalian assemblag-es from different sites and temporal phases of the Natufian. The site of el-Wad Terrace is situated in what may be considered the quintessential ecotonal
zone at the contact of Mount Carmel and the coastal plain and at the opening of one of the Carmel’s sea-sonal stream valleys (Weinstein-Evron 1998). Data on the fauna of the site have shown that species fa-voring open and mainly grass-covered environments thriving on the coastal plain such as gazelle (Gazella gazella) and the social vole (Microtus guentheri) dominate the LN assemblage (Bar-Oz et al. 2004; Weinstein-Evron et al. 2007). A similar situation appears to have pertained throughout the Natufian sequence of el-Wad Terrace (Valla et al. 1986; see also Garrod and Bate 1937; Weinstein-Evron 1998). Species representing more forested environments occur as well but in relatively low frequencies (Valla et al. 1986; Weinstein-Evron et al. 2007; Yeshurun et al. herein).
Table 1. Description of samples of micromammalian assemblages from Natufian sites included in this study
Site/assem-blage
Temporal phase
Approxi-mate area of excavation
and sampling (m2)
No. samples Context description
el-Wad Terrace
EarlyNatufian
(EN)58/12 3
Samples collected from top of EN layer in associa-tion with a wall of a partially preserved structure (Str.II): 1) a living floor inside the structure (I/S Str.II) adjacent to the concave side of the wall, 2) above the structure (Abv. Str.II) in deposits overlying the wall, 3) outside the structure (O/S Str.II) adjacent to the convex side of the wall and situated between the wall and the bluff overhang-ing the terrace (Yeshurun et al. herein).
el-Wad Terrace
LateNatufian
(LN)58/1a 8
Seven of the samples (numbered I-VII) collected from a sediment column (50×50 cm2) in spatially undifferentiated deposits of LN layer, each sample representing a c.5 cm spit (Weissbrod et al. 2005). Additional sample collected from a LN burial de-posit (Bur.; Weinstein-Evron et al. 2007).
Raqefet Cave
LateNatufian
(LN)70/11 3
Two of the samples collected from loci with LN deposits containing multiple burials (Loc. I and Loc. III) and an additional sample from inside two human-made bedrock holes within the cave floor (HBH CI-II) (Nadel et al. 2008).
EynanFinal
Natufian (FN)
160/16 4
Samples collected from deposits inside well-de-fined structures associated with features in-cluding living floors, hearths, and burials: 235, 200/222/240b, 225, 228/242 (Valla et al. 2007).
aThe area of sampling in this case may be misleading in terms of the volume sampled because unlike all other samples in the study it represents a column with a substantial depth (c. 0.25 m).bSamples from several of the features were lumped for the purpose of this study due to their spatial association and relatively small sample sizes analyzed so far.

702
Lior Weissbrod et al.
The location of Raqefet Cave is further inland within an inner valley of southern Mount Carmel and at an elevation of 230 m above msl (Nadel et al. 2008, 2009). The cave is situated within a more mountainous topography but is also adjacent to the Menashe plateau and both gazelle and voles are highly abundant. Typical woodland species such as roe deer (Capreolus capreolus) and Persian squirrels (Sciurus anomalus) are represented at Raqefet Cave at somewhat greater frequencies than at the contemporaneous LN deposits of el-Wad Terrace on the western coastal side of the mountain (Nadel et al. 2008, 2009). The open-air site of Ey-nan lies at an elevation of 80 m above msl within the Jordan Valley and at the piedmont zone of the Galilee Mountains and the inundated basin of the Hula (Valla et al. 2007). Quantitative data on the micromammalian assemblage of the site have not yet been published in detail (Bouchud 1987; Valla et al. 1998). The Natufian inhabitants of Eynan would have had access to both open grassland and marshland environments of the valley floor and higher elevation woodlands of the Galilee Moun-tains (e.g. Leroi-Gourhan 1984). Recent high-resolution climate data from isotope records in cave speleothems in the region indicate that the Natufian represents a period of marked increase in rainfall and temperatures in comparison to the preceding relatively dry and cold period of the last glacial maximum (LGM) that peaked ca. 19,000 bp (Bar-Matthews et al. 2003). The LN phase is generally considered to coincide with a short-term retreat in the trend of warming and increased rainfall known as the Younger Dryas although the consequence of this oscillation for the environment of southwest Asia and for temporal developments within the Natufian culture has been widely debated (summarized in Byrd 2005; Weinstein-Evron 2009). Data on the geology and sedimentology of the Dead Sea in the Jordan Val-ley also reveal an increase rather than decrease in rainfall providing conflicting indications on the nature of Younger Dryas climate change (Stein et al. 2009). Cultural developments from the early to the later phase of the Natufian whether driven by external climatic events or by internal processes appear to have involved major changes and possibly increasing variability in the mode and intensity of use of sites within the Natufian core area (Bar-Yosef 2002; Weinstein-Evron 2009). A final set of considerations for inter-site/phase comparisons of micromammalian assemblages is related to the issue of taphonomy and factors which could have affected the manner in which
micromammalian remains accumulated at the sites. Taphonomic considerations that can further complicate such comparisons include differences in the mode of accumulation and preservation among sites in cave/terrace and open-air settings, the potential for direct contribution of humans to accumulation through small-game hunting, and within-site variability in assemblage composition that may be related to the spatial organization of human activities or to other processes affecting site formation such as site topography and modes of sedimentation. Previous study of the taphonomy of the LN assemblage of el-Wad Terrace revealed a signifi-cant contribution to accumulation at the site by nocturnal raptors such as owls (order Strigiforms) that could have been nesting or roosting within the cliff overhanging the site (Weissbrod et al. 2005). Certain species of owls that typically uti-lize cave or cliff settings where they accumulate the remains of their micromammalian prey are believed to be important taphonomic agents in such sites but less important in open-air sites (Tchernov 1984). Additional evidence from el-Wad Terrace showed that secondary contributions to the LN micromammal ian assem blage included human consumption of mole rats (Spalax ehren-bergi) and some in situ accumulation of mice of the genus Mus that may have lived at the settlement as commensals (Weissbrod et al. 2005). In situ or autochthonous accumulation may be an especially important process in the formation of assemblages of commensal animal remains but has received little attention in research (Evans 1978; Reitz and Wing 1999; Tchernov 1984; Weissbrod 2010a). Within-site spatial variability in taxonomic composition has been detected to some extent at el-Wad Terrace LN (Weinstein-Evron et al. 2007) and in a more pronounced manner at Raqefet Cave (Nadel et al. 2008), in both cases partly in relation to distinct activity contexts such as burials and bed-rock mortars versus less differentiated deposits. Conditions of preservation can also vary great-ly among cave, terrace, and open-air depositional environments. Previous analyses have shown that the el-Wad Terrace LN assemblage exhibits con-siderably greater rates of fragmentation than the LN Raqefet Cave assemblage in spite of the fact that both assemblages had high representation of elements from all parts of the skeleton (Nadel et al. 2008; Weissbrod et al. 2005). The high fragmentation at el-Wad Terrace was related to processes of tram-pling, exposure to aerial weathering, and transport from the location of initial deposition by raptors

703
Commensalism: Was it Truly a Natufian Phenomenon?…
in the cliff to the terrace (Weissbrod et al. 2005). A final important influence on observed variability in taxonomic composition of micromammalian assem-blages is the method of retrieval during excavation. Retrieval of the assemblages described here was conducted through fine wet-screening in mesh of 1-2 mm. It has been shown that some of the molar teeth of especially small species such as mice of the genus Mus may become lost even through 1 mm mesh screens (Weissbrod et al. 2005). It can therefore be expected that differences in the level of precision of retrieval will to some extent affect assemblage composition and comparability. The taphonomy of the assemblages included in our paper has so far been studied only in part. Nonetheless, in our preliminary consideration of variability among micromammalian assemblages from different sites and temporal phases of the Natufian we attempt to take into account a number of compounding factors and examine the potential for preservation of information on commensalism.
A brief review of the commensalism model and available evidence
The commensalism model posits that the dura-tion and intensity of human settlement occupation are major factors affecting the taxonomic composi-tion and organization of associated communities of micromammals. Tchernov invoked commensalism to account for the appearance or marked increase in abundance in the archaeological record of species that are well known commensals in present day settlement environments (Bar-Yosef and Tchernov 1966; Tchernov 1984, 1991a, b; see also Hesse 1979). He focused on the Natufian but also traced earlier Late Pleistocene fluctuations in the frequencies of commensal species including three rodents, house mouse, black rat (Rattus rattus), and spiny mouse (Acomys cahirinus), and one bird, house sparrow (Tchernov 1984). Tchernov linked early and rel-atively small-scale fluctuations in frequencies of commensal species to variability in duration and intensity of settlement occupation through time and emphasized a marked increase in frequencies in the Natufian and its relation to significant sedentariza-tion. In later publications (Tchernov 1991a, b) the emphasis shifted from evidence that was based on relative frequencies of species to establishing the appearance of a single key commensal species – the house mouse – in the Natufian. This approach was based on morphometric analysis of mouse cranial characteristics that revealed the occurrence in the record of two closely related species of common
mice of the genus Mus – one which occurred in the region at least since Middle Pleistocene times (Mus macedonicus) and another which appeared for the first time in Natufian deposits and is considered to be the true commensal house mouse (Auffray et al. 1988). In order to link the seemingly abrupt appear-ance of house mice in the Natufian to significant sed-entism, Tchernov also drew on ecological evidence on contemporary mouse species in Israel, which suggests their strong dependence on human set-tlement environments. A study on the distribution of the two extant species of common mouse showed that in drier regions of southern Israel where only house mice occur they occupy both settlements and non-settlement environments whereas in regions where the two species are overlapping (i.e. sympat-ric) within the Mediterranean climate zone, house mice are mainly restricted to settlements (Auffray et al. 1990). This indicates competitive exclusion between house and outdoor mice in their region of sympatry. The evidence for the exclusion of house mice from non-settlement environments by outdoor mice suggested to Tchernov (1991a, b) that only the creation of a new type of habitat in highly sedentary Natufian settlements could have opened up a niche to be filled by a specialized commensal species. Current evidence from DNA of the various members of the Mus group inhabiting regions of West and Central Asia and Europe show that the ancestor of the house mouse in southwest Asia originated in Central Asia during earlier periods of the Late Pleistocene and underwent a westerly expansion event which coincided with the post-gla-cial period of the Final Pleistocene (Boursot et al. 1993; Rajabi-Maham et al. 2008). It is conceivable that both climate change and the appearance of more sedentary human occupation sites could have contributed to the spread and establishment of the present day house mouse in regions which were for-merly inhabited by other, currently non-commensal species of common mice. It still remains unclear precisely what level of sedentism and related habitat alteration would have been necessary for the establishment of the commensal house mouse in Natufian settlements. A fundamental question that remains open is whether commensal species can also occur in the settlements of seasonally mobile communities (see Edwards 1989; Tangri and Wyncoll 1989). The ecological mechanism underlying the commensalism model is a universal one. In eco-logical theory, commensalism is a formal type of interspecies interaction involving a positive effect

704
Lior Weissbrod et al.
of one species on the equilibrium population size of another without immediate consequences for itself and is denoted a +/0 interaction (Dickman 1992, 2006). A key aspect of commensalism is that in con-trast to more direct ecological interactions such as predation (+/-) and competition (-/-) it often occurs indirectly through the contribution of one species to the suitability of the environment of another. Recent advances in ecological theory suggest that commensal interactions can be subsumed within a more generalized framework of indirect interac-tions involving environmental modification that is a fundamental and pervasive process in ecosystems worldwide (Dickman 2006). This process has been termed environmental engineering and is typical-ly identified in situations where the activities of environmentally more dominant organisms such as trees, elephants, and dam-building beavers alter physical conditions in the environment and indirectly affect the supply of critical resources to a host of other species (Jones et al. 1994, 1997; see also Smith 2007 on archaeological implications). In this framework, the creation and maintenance of settlements that is a basic characteristic of human culture may be seen as a specific form of environ-mental engineering. In his discussion of the development of commen-salism in the context of sedentarization in Natufian settlement occupation Tchernov (1991a) relates the activities of human settlement occupation to habitat alteration and associated ecological dynamics. By drawing on principles of mammalian behavioral ecology and optimal and central-place foraging, Tchernov ties human occupation of settlements to mobility, resource acquisition, and their ecological consequences (Tchernov 1984, 1991a, b). Here, increased permanency in use of settlements is pre-dicted to occur in conjunction with reduced mobility and enhanced and more spatially bounded effort of resource extraction from the environment surround-ing settlements. Such an effort of concentrated daily resource extraction activities that are sustained for long periods of time through intensified seasonal occupation and under conditions of growing human populations in settlements is expected to reduce the abundance of many species in the immediate environment and increase the abundance of a few others. This ecological process implies a reduction in biological diversity and a process of commen-salization (Tchernov 1991a: Fig. 2). According to Tchernov (1984, 1991a) the process of commensal-ization is predicted to accompany the development of sedentary settlements from less sedentary ones and should represent a threshold in the nature
and scale of ecological interactions among human inhabitants of settlements and coexisting species of small animals. The exact levels of commensalism or of human settlement occupation that define such a threshold remain largely unknown. Aspects of habitat engineering that could have impacted ecological interactions in Natufian set-tlements should include the extraction of subsis-tence resources from the immediate environment, collection of materials for construction and fuel, the erection of various structures (e.g. dwellings, installations, and fences or terrace walls), human traffic, and the accumulation of refuse and storage of foodstuffs. It can be envisaged that some of these activities constituting substantial disturbance of the natural environment of the settlements would have contributed to depressing the abundance of certain species whereas other activities could have at the same time enhanced the abundance of other species thereby leading to commensalism. Theoretically, factors that can contribute to for-mation of commensal bonds include the supply of food and shelter and reduction of pressures from competing species or predators (Dickman 1992). These improved habitat conditions will be indi-rectly provided by the unaffected partner to the commensal species. A range of ecological studies in present day settlement environments have generally shown that the conditions provided by human modified habitats can mitigate various limitations on population size that pertain in less modified habitats. Such research also indicates that only species with suitable pre-adaptations and a competitive advantage can benefit from the unique circumstances in such environments (Andrzejewski 1978; Berry 1981; Braithwaite 1980; Canova and Fasola 1994; Chernousova 2001, 2002; Courtney and Fenton 1976; Ganem 1991, 1993; Pocock et al. 2004). Tchernov (1991a) already observed that the bi-ological and ecological characteristics of commensal species may be equivalent to those of invasive or colonizing species and that an invasive or colonizing ability should represent a pre-adaptation to com-mensalism. A recent summary by Sakai et al. (2001) of the characteristics of invasive species includes: 1) tolerance of environmental stress, disturbance, and heterogeneity based on behavioral and/or phenotyp-ic plasticity, 2) high productivity and reproductive flexibility (i.e. r-selected life-history strategies), 3) competitiveness through aggressive behavior, 4) broad dietary preferences, and 5) wide distribution across varied habitats. It can be expected that such pre-adapted species occupying Natufian settlement

705
Commensalism: Was it Truly a Natufian Phenomenon?…
environments would have benefited from enhanced availability of food from organic refuse or stored supplies and of shelter from the construction of various structures. In addition, many competitors and predators of commensal species should have been excluded to some extent from Natufian set-tlement environments because they were either directly exploited by humans, depended on other exploited resources, or could otherwise not tolerate escalating levels of disturbance from human activ-ities. The exact manner in which different species may react to specific human engineering activities as well as to the overall impact of increasing levels of occupation in sedentarizing human settlements is little understood for most species of animals, however. Answers to some of the questions that remain open regarding the commensalism-sedentism link-age and the relevance of the commensalism model for identifying sedentism in the Natufian depend on the availability of additional empirical data. These data can only be obtained from observations on interactions between humans and micromam-mals in a wide range of contemporary settlement settings. Extrapolating ecological and cultural in-formation from contemporary settings back in time and across wide geographic regions is problematic. This endeavor may be useful, nonetheless, for es-tablishing generalized boundary conditions for the past such as the distinction between the ecological impact of seasonally mobile and sedentary human communities.
Ethnoarcheological and ecological contributions to the commensalism question
Direct ecological observations in settlements of complex hunter-gatherers equivalent to those of the Natufians would have been ideal for our purposes. Such societies do not exist in the modern world, however. Relevant data is presently available from only a few studies in other types of settlement contexts. Courtney and Fenton (1976) studied the influence of a seasonal holiday retreat in Canada on populations and community composition of micromammalian species. Their study focused on the garbage dump of the retreat which was used during May-October of each year by 4-5 families dumping >200 liter/week of garbage. Monitoring of micromammals in the dump and in an off-site control revealed greater overall abundances, levels of female fecundity, and juvenile survival in the dump than in the adjacent control site. These data
indicate increase in populations in the dump. In addition, it was shown that a local and high-ly abundant rodent species – white-footed mouse (Peromyscus leucopus) – was an early colonizer of the dump during the seasonal cycle but that it was eventually out-competed by introduced commensal house mice. These house mice were introduced sea-sonally with garbage trucks and died out following the termination of dumping each year. Courtney and Fenton (1976) hypothesized that white-footed mice initially benefited due to their omnivorous feeding habits but that house mice had the advantage of greater flexibility in their social structure and an ability to maintain denser populations. It was sug-gested that the seasonal and discontinuous use of the dump prevented establishment of stable house mouse populations and more broadly that the degree of human occupation influenced the structure and composition of the micromammalian community. This would appear to support the inference that high abundance of commensals in archaeological contexts will indicate more continuous human occupation (e.g. Hesse 1979; but see Tangri and Wyncoll 1989). The garbage dump situation in a seasonal holiday retreat in temperate Canada may not be readily comparable to that of Natufian settlements. Nonetheless, this case highlights the current lack of criteria for identifying the parameters of com-mensalism in human settlement environments and how varying levels of human occupation may affect the ecology of associated micromammalian communities. It may be asked, for example, whether the association of indigenous white-footed mice with the seasonal holiday retreat may in fact constitute a valid case of commensalism or whether year-round rather than seasonal occupation by only 4-5 families and anticipated permanent establishment of house mice would have. A more relevant approach may be based on comparison of ecological data on micromamma-lian communities of settlements from sedentary and seasonally occupied settlement environments among small-scale societies. Such a comparison allows us to better assess how varying signals of commensalism may relate to specific levels of settle-ment occupation along a sedentarization gradient. Available data are taken from a recent study by one of us (Weissbrod 2010a, b) on seasonal settlements of Maasai pastoralists in southern Kenya and from an earlier study (Misonne 1963) on agricultural villages in the northeastern Democratic Republic of Congo (DRC). The study of Maasai settlements was conducted in a semi-arid region where farming

706
Lior Weissbrod et al.
is not viable and mobile herding of cattle, sheep, and goats is the predominant economic pursuit. Settlements can remain in use for many years (> 40 years in some cases) but are occupied on a sea-sonal basis, a pattern that may be referred to as seasonal sedentism (Kent 1989). Typically, a part of or the entire population of Maasai settlements undertakes movements with herds between wet and dry seasons. Maasai settlements may be considered small, typically consisting of less than 10 houses and having a relatively small and stable human pop-ulation (Weissbrod 2010a, b). Important aspects of engineering of the settlements may include the construction and maintenance of houses and branch fences for livestock enclosures, considerable buildup of dung from the corralling of livestock, and grazing by livestock and collection of materials for house and fence construction and for fuel from surround-ing habitats. There is little or no storage of food or accumulation of organic food refuse. In contrast, the villages in Misonne’s (1963) study were located in one of the most productive agricultural regions of Africa within the Rwenzori mountain range and near Lakes Edward and Albert in northeastern DRC. It was reported that the studied villages consisted of between 10 and 100 houses and were adjacent to crop fields (Devignat 1946; Misonne 1963). In these circumstances we may expect sig-nificant sedentism in village occupation, relatively large and growing human populations, and storage of food and accumulation of organic refuse on a large scale, which constitute important aspects of engineering. The two studies of pastoralists and farmers vary greatly in terms of the environmental setting and cultural context but are comparable in terms of the characteristics of micromammalian communities of the settlements due to the way in which the studies were designed. In both cases ecological monitoring of micromammalian communities was done inside the settlements as well as in outside control sites al-lowing for a controlled comparison between the two contexts. Data on micromammalian communities were collected through systematic trapping in six settlements in the Kenya Maasai study (Weissbrod 2010a, b) and in 58 villages in the DRC study (Mi-sonne 1963). In addition, in the former case control sites were situated at a distance of 200-400 m from each of the settlements and in the latter a series of control sites were deployed along a transect of increasing distance from the villages beginning at 50 m in the adjacent crop fields and extending to less intensely utilized habitats beyond 200 m
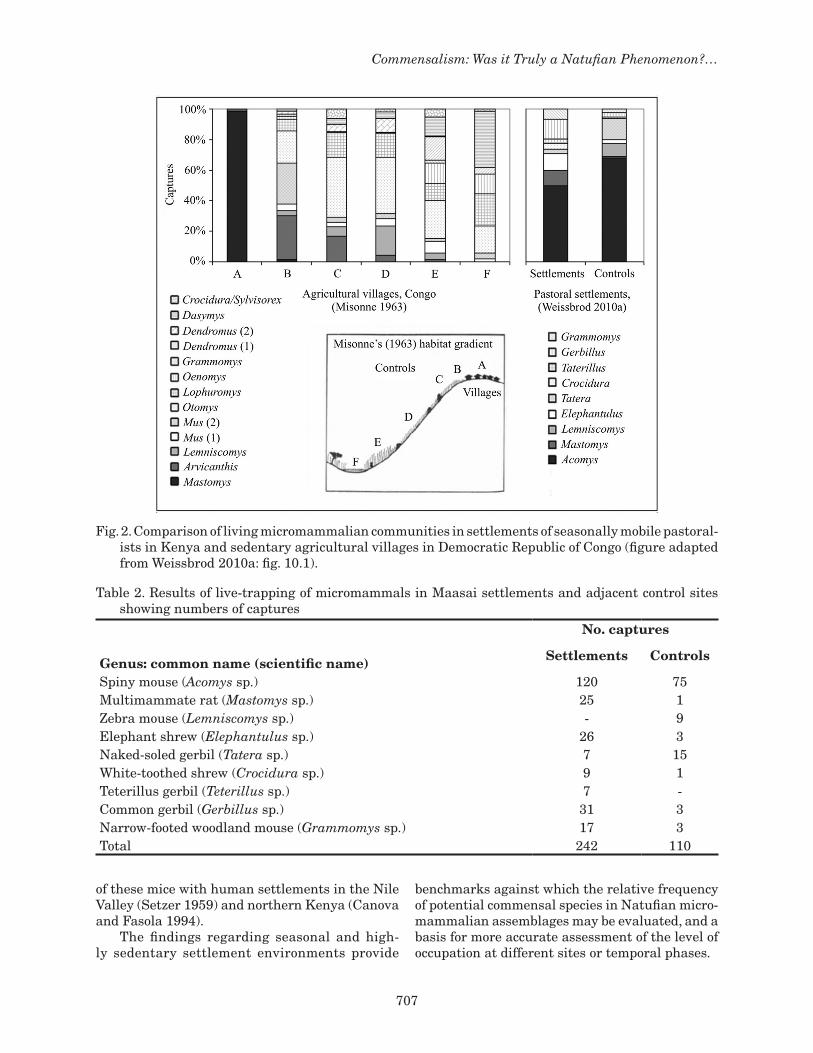
from the villages (see Fig. 2). The overall numbers of captures of micromammals in the two studies were 352 and 30,731 individuals, respectively. Figure 2 presents bar graphs that summarize the proportions of captures for each species of mi-cromammal across settlement and control sites for the agricultural villages and pastoral settlements. A stark difference can be observed when comparing the agricultural villages and pastoral settlements and their respective control sites. The agricultural villages exhibit dominance by a single species – multimammate rats (Mastomys sp.) – and are ex-ceedingly less heterogeneous than adjacent control habitats. Multimammate rats have a frequency of >98% inside the villages but drop to below 2% in the adjacent crop fields and are absent from habitats further away. The difference in absolute abundances between the villages and outside fields cannot be determined from Misonne’s (1963) published data. Nonetheless, it was stated by the author (Misonne 1963:106) that whereas multimammate rats far exceeded in abundance all other species within the villages they were practically absent in the outside fields. This pattern presents a clear case of commensalism in the villages and accords with the well known status of multimammate rats as one of the leading indigenous commensals in Africa (Kingdon 1974). On the other hand, the pastoral settlements have only as much as 50% of the most abundant species – spiny mice (Acomys sp.) – and show greater heterogeneity than in adjacent control sites (Fig. 2). In addition, the absolute abundances of most micromammalian species in the settlements exceeded those in the controls. Table 2 shows a ratio of 120:75 for spiny mice, 25:1 for multimam-mate rats, and intermediate ratios for four other species. Only Tatera gerbils (Tatera sp.) had fewer numbers in the settlements than in the controls. From a theoretical standpoint, this pattern supports a case of environmental engineering with overall positive effects for a wide variety of species that may involve commensal interactions (Weissbrod 2010a). The case of pronounced commensalism of multimammate rats in the sedentary agricultural villages (frequency of 98%) may represent an ex-treme along a continuum of commensalization. The association of spiny mice with Maasai settlements (frequency of 50%) may be considered a case of “subtle commensalism” that can be related to the seasonal use of the settlements and reduced mag-nitude of habitat modification. The inference on commensalism of spiny mice in Maasai settlements may be further supported by data on the association
707
Commensalism: Was it Truly a Natufian Phenomenon?…
of these mice with human settlements in the Nile Valley (Setzer 1959) and northern Kenya (Canova and Fasola 1994). The findings regarding seasonal and high-ly sedentary settlement environments provide
benchmarks against which the relative frequency of potential commensal species in Natufian micro-mammalian assemblages may be evaluated, and a basis for more accurate assessment of the level of occupation at different sites or temporal phases.
Fig. 2. Comparison of living micromammalian communities in settlements of seasonally mobile pastoral-ists in Kenya and sedentary agricultural villages in Democratic Republic of Congo (figure adapted from Weissbrod 2010a: fig. 10.1).
Table 2. Results of live-trapping of micromammals in Maasai settlements and adjacent control sites showing numbers of captures
No. captures Genus: common name (scientific name) Settlements Controls
Spiny mouse (Acomys sp.) 120 75Multimammate rat (Mastomys sp.) 25 1Zebra mouse (Lemniscomys sp.) - 9Elephant shrew (Elephantulus sp.) 26 3Naked-soled gerbil (Tatera sp.) 7 15White-toothed shrew (Crocidura sp.) 9 1Teterillus gerbil (Teterillus sp.) 7 -Common gerbil (Gerbillus sp.) 31 3Narrow-footed woodland mouse (Grammomys sp.) 17 3Total 242 110
708
Lior Weissbrod et al.
Variability in Natufian micromammalian assemblages
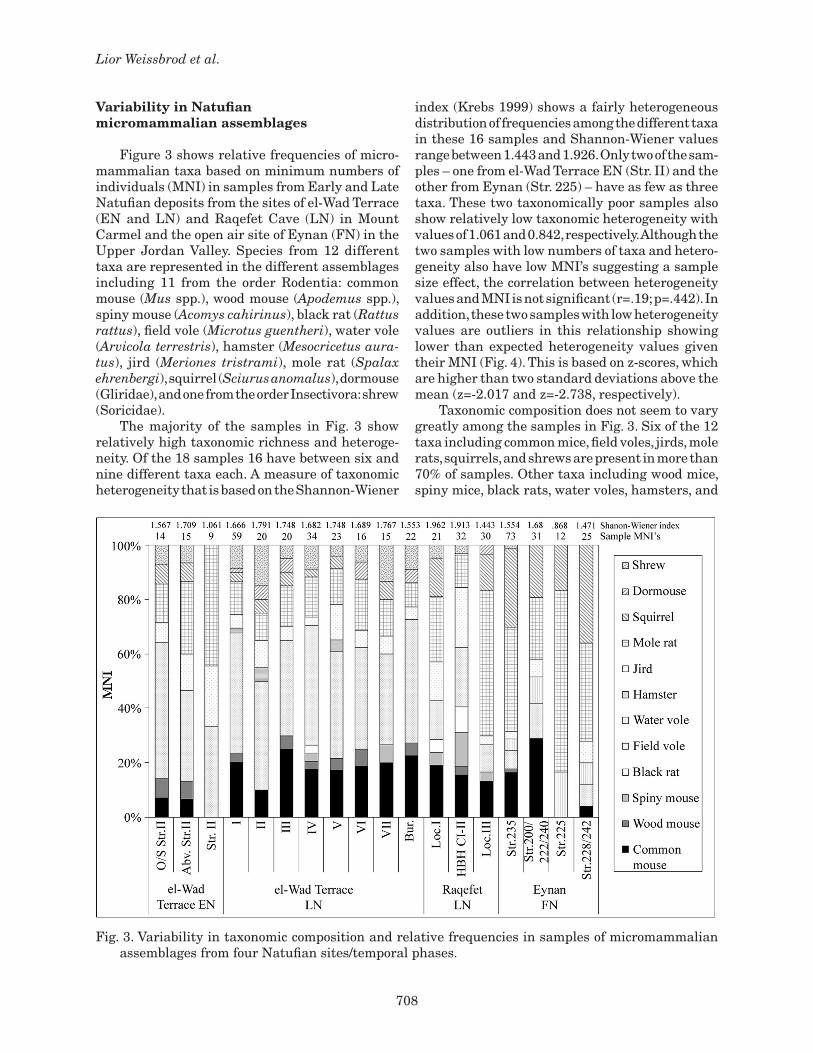
Figure 3 shows relative frequencies of micro-mammalian taxa based on minimum numbers of individuals (MNI) in samples from Early and Late Natufian deposits from the sites of el-Wad Terrace (EN and LN) and Raqefet Cave (LN) in Mount Carmel and the open air site of Eynan (FN) in the Upper Jordan Valley. Species from 12 different taxa are represented in the different assemblages including 11 from the order Rodentia: common mouse (Mus spp.), wood mouse (Apodemus spp.), spiny mouse (Acomys cahirinus), black rat (Rattus rattus), field vole (Microtus guentheri), water vole (Arvicola terrestris), hamster (Mesocricetus aura-tus), jird (Meriones tristrami), mole rat (Spalax ehrenbergi), squirrel (Sciurus anomalus), dormouse (Gliridae), and one from the order Insectivora: shrew (Soricidae). The majority of the samples in Fig. 3 show relatively high taxonomic richness and heteroge-neity. Of the 18 samples 16 have between six and nine different taxa each. A measure of taxonomic heterogeneity that is based on the Shannon-Wiener
index (Krebs 1999) shows a fairly heterogeneous distribution of frequencies among the different taxa in these 16 samples and Shannon-Wiener values range between 1.443 and 1.926. Only two of the sam-ples – one from el-Wad Terrace EN (Str. II) and the other from Eynan (Str. 225) – have as few as three taxa. These two taxonomically poor samples also show relatively low taxonomic heterogeneity with values of 1.061 and 0.842, respectively. Although the two samples with low numbers of taxa and hetero-geneity also have low MNI’s suggesting a sample size effect, the correlation between heterogeneity values and MNI is not significant (r=.19; p=.442). In addition, these two samples with low heterogeneity values are outliers in this relationship showing lower than expected heterogeneity values given their MNI (Fig. 4). This is based on z-scores, which are higher than two standard deviations above the mean (z=-2.017 and z=-2.738, respectively). Taxonomic composition does not seem to vary greatly among the samples in Fig. 3. Six of the 12 taxa including common mice, field voles, jirds, mole rats, squirrels, and shrews are present in more than 70% of samples. Other taxa including wood mice, spiny mice, black rats, water voles, hamsters, and
Fig. 3. Variability in taxonomic composition and relative frequencies in samples of micromammalian assemblages from four Natufian sites/temporal phases.
709
Commensalism: Was it Truly a Natufian Phenomenon?…
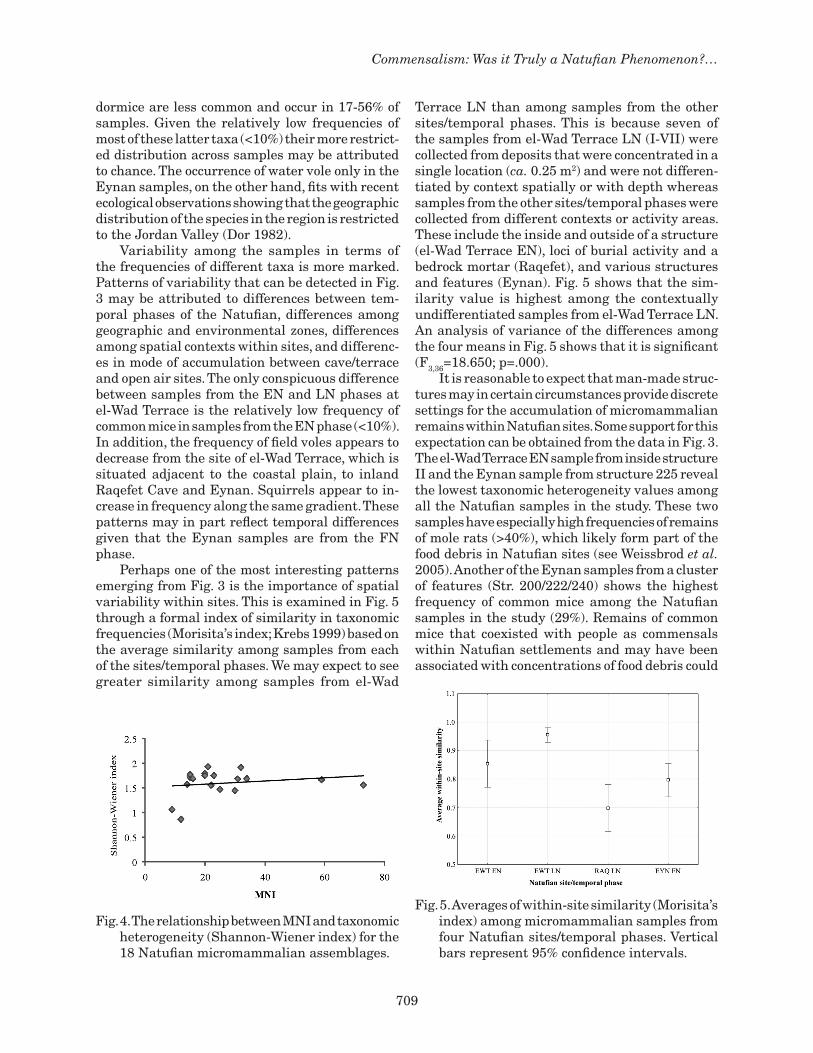
dormice are less common and occur in 17-56% of samples. Given the relatively low frequencies of most of these latter taxa (<10%) their more restrict-ed distribution across samples may be attributed to chance. The occurrence of water vole only in the Eynan samples, on the other hand, fits with recent ecological observations showing that the geographic distribution of the species in the region is restricted to the Jordan Valley (Dor 1982). Variability among the samples in terms of the frequencies of different taxa is more marked. Patterns of variability that can be detected in Fig. 3 may be attributed to differences between tem-poral phases of the Natufian, differences among geographic and environmental zones, differences among spatial contexts within sites, and differenc-es in mode of accumulation between cave/terrace and open air sites. The only conspicuous difference between samples from the EN and LN phases at el-Wad Terrace is the relatively low frequency of common mice in samples from the EN phase (<10%). In addition, the frequency of field voles appears to decrease from the site of el-Wad Terrace, which is situated adjacent to the coastal plain, to inland Raqefet Cave and Eynan. Squirrels appear to in-crease in frequency along the same gradient. These patterns may in part reflect temporal differences given that the Eynan samples are from the FN phase. Perhaps one of the most interesting patterns emerging from Fig. 3 is the importance of spatial variability within sites. This is examined in Fig. 5 through a formal index of similarity in taxonomic frequencies (Morisita’s index; Krebs 1999) based on the average similarity among samples from each of the sites/temporal phases. We may expect to see greater similarity among samples from el-Wad
Terrace LN than among samples from the other sites/temporal phases. This is because seven of the samples from el-Wad Terrace LN (I-VII) were collected from deposits that were concentrated in a single location (ca. 0.25 m2) and were not differen-tiated by context spatially or with depth whereas samples from the other sites/temporal phases were collected from different contexts or activity areas. These include the inside and outside of a structure (el-Wad Terrace EN), loci of burial activity and a bedrock mortar (Raqefet), and various structures and features (Eynan). Fig. 5 shows that the sim-ilarity value is highest among the contextually undifferentiated samples from el-Wad Terrace LN. An analysis of variance of the differences among the four means in Fig. 5 shows that it is significant (F3,36=18.650; p=.000). It is reasonable to expect that man-made struc-tures may in certain circumstances provide discrete settings for the accumulation of micromammalian remains within Natufian sites. Some support for this expectation can be obtained from the data in Fig. 3. The el-Wad Terrace EN sample from inside structure II and the Eynan sample from structure 225 reveal the lowest taxonomic heterogeneity values among all the Natufian samples in the study. These two samples have especially high frequencies of remains of mole rats (>40%), which likely form part of the food debris in Natufian sites (see Weissbrod et al. 2005). Another of the Eynan samples from a cluster of features (Str. 200/222/240) shows the highest frequency of common mice among the Natufian samples in the study (29%). Remains of common mice that coexisted with people as commensals within Natufian settlements and may have been associated with concentrations of food debris could
Fig. 4. The relationship between MNI and taxonomic heterogeneity (Shannon-Wiener index) for the 18 Natufian micromammalian assemblages.
Fig. 5. Averages of within-site similarity (Morisita’s index) among micromammalian samples from four Natufian sites/temporal phases. Vertical bars represent 95% confidence intervals.

710
Lior Weissbrod et al.
have accumulated in situ within structures (see also Yeshurun et al. herein). More detailed spatial and taphonomic analysis of larger samples from these sites would be required to assess the cultural significance of the observed patterns of variability, however. Varying modes of accumulation of micromam-malian remains in Natufian sites may also be re-lated to differences in the depositional environment between cave/terrace and open-air sites. In the former case we can expect a significant contribution of predators such as owls roosting in rock cavities (Weissbrod et al. 2005) whereas in open air sites predator activity is considered less likely and in situ accumulation may constitute a more significant contributor to the formation of micromammalian assemblages (Tchernov 1984). A comparison of samples from the open-air site of Eynan with those from Raqefet Cave and el-Wad Terrace EN and LN does not reveal marked differences that may specifically relate to site type and the depositional environment, however. Taxonomic heterogeneity (Shannon-Wiener index) of the Eynan samples ranging between 0.868 and 1.554 falls within the range of the other samples from cave/terrace sites. Moreover, intra-site differences in frequencies of specific taxa among the samples from Eynan seem just as marked as inter-site differences between samples from Eynan and the cave/terrace sites. For example, the frequency of common mice among all Natufian samples is highest in the Eynan sample from structures 220/222/240 whereas this taxon is absent or has an especially low frequency in two other Eynan samples (Str. 225 and 228/242, respectively). A fifth potential source of influence on variability among Natufian micromammalian assemblages is the ecological impact of Natufian settlements on their immediate surroundings and the formation of commensal micromammalian communities in association with the settlements. The Natufian micromammalian samples in Fig. 3 can be com-pared with living micromammalian communities from contemporary seasonal settlements of Maasai pastoralists and sedentary agricultural villages in Africa (see Fig. 2). The fossil and living communities may be compared in terms of the level of diversity and degree of dominance of expected commensal taxa. None of the fossil samples exhibit the extreme of depressed heterogeneity and dominance by a single taxon that characterizes micromammalian communities of the sedentary agricultural villages. House mice are considered a key commensal taxon of Natufian settlements. In Fig. 3, house
mice are represented by the taxonomic category of common mouse (genus Mus) including both the house mouse (M. musculus domesticus) and outdoor species (M. macedonicus). The maximum relative frequency of this taxon among the Natufian sam-ples is approximately 30%. This is markedly lower than both the frequency of spiny mice in seasonal Maasai settlements (50%) and of multimammate rats in sedentary agricultural villages (98%). Except for five samples with especially low frequencies or absence of common mice including the three sam-ples of el-Wad Terrace EN and two of the Eynan samples, all Natufian samples have frequencies of common mice ranging between 10 and 30%.
Discussion: An evaluation of the case for Natufian commensalism
In this section we assess the variability among micromammalian assemblages from Natufian sites in light of insights from ethnoarcheological and ecological data on commensalism. The data presented here on living micromammals of mobile and sedentary communities in Africa suggests that the frequency of a key commensal of southwest Asia – the house mouse – in a number of important Natufian sites is relatively low. This may be taken as an indication of relatively low duration and intensity of human occupation in the Natufian settlements. Given that the majority of the analyzed samples were collected from deposits of the late phase of the Natufian (LN) such an indication would correspond with expectations for increased mobility during this phase (Bar-Yosef 2002; Belfer-Cohen and Bar-Yosef 2000; Byrd 2005; but see Weinstein-Evron 2009). Several important factors may complicate the direct use of micromammalian assemblages from Natufian sites as indicators of commensalism and human level of occupation, however. It was shown that data on variability in micromammalian assem-blages from Natufian sites can contain information on geographic differences in environmental condi-tions that occur on a larger scale than the localized ecological impact of settlements on their immediate surroundings. Additional influencing factors include the accumulation of remains from food-related ac-tivities involving micromammals. This does not rule out the potential for preservation of information on ecological processes such as commensalism, which are expected to have occurred at the local scale of the Natufian settlement. It does imply, however, that micromammalian assemblages from Natu-fian sites represent palimpsests of remains that accumulated by different means and represent

711
Commensalism: Was it Truly a Natufian Phenomenon?…
environmental, ecological, and cultural processes occurring at varying spatial and temporal scales. An important fact to consider is that a substan-tial proportion of micromammalian assemblages from Natufian sites and particularly those situated in cave/terrace settings is likely to have been accu-mulated by predators such as owls (Weissbrod et al. 2005). It is unlikely that the hunting ranges of var-ious predators contributing micromammalian prey remains to Natufian deposits were confined to the settlements or their immediate surroundings where the ecological impact of the settlements would have been felt. A study by one of us (Weissbrod 2010a) on recent prey assemblages (owls and mongoose) in the environment of Maasai settlements in southern Kenya has shown marked differences in taxonomic composition between the prey assemblages and living micromammalian communities from inside the settlements. Several taxa that were frequent-ly recorded through trapping in the settlements were either absent or under-represented in the prey assemblages of predators, which occurred in proximity (< 1 km) to the settlements. This suggests that prey assemblages deposited within small-scale settlements or their vicinity will represent the wider environmental background of the settlements and the landscape rather than the localized ecological impact of the settlements (Weissbrod 2010a). Other such recent prey assemblages of owls from Israel provide a useful comparison to Natu-fian micromammalian assemblages. Dor (1982) analyzed 22 prey assemblages that were collected during the first half of the twentieth century from a wide range of geographic regions in the central and northern parts of the country. These assemblages show considerable variability in the frequency of common mice ranging between 0 and 75% (Dor 1982). Of the 22 assemblages only three had a fre-quency of common mice exceeding 30%, however, one from the Judean Mountains of central Israel (Har-Tov) and two from the Sea of Galilee in the north (Dgania and Tiberias). Additional assemblag-es collected more recently in settlement settings in Israel including a suburban site (Charter et al. 2007) and an agricultural village and crop fields (Charter et al. 2009) show a more restricted range of frequencies of 30-50%. This is decidedly greater than the frequencies of common mice in the Natufian samples (10-30%). Trapping studies have also shown that especially high frequencies of common mice of 50-90% can occur in secondary scrubland habitats (e.g. garigue), post-fire habitats, homogeneous tree plantations, and crop fields (Kahila Bar-Gal et al. 2000; Kupstein 2001; Lehmann and Perevolotsky
1992). Relatively intact scrub forest of various formations including oak (Quercus calliprinos), mock privet (Phillyrea media) or carob-pistacio (Ceratonia siliqua-Pistacia lentiscus) in the Mount Carmel region support intermediate frequencies of common mice of between 20 and 40% (Kupstein 2001; Lehmann and Perevolotsky 1992). If we view Natufian micromammalian assem-blages through the taphonomic lens of predator accumulation we may see evidence for environ-mental conditions at the broad spatial scale of the landscape. From this perspective, common mouse frequencies in Natufian contexts (10-30%) corre-spond to what we would expect in environments little disturbed by human activities (see also Lev-Ya-dun and Weinstein-Evron 2005). The EN samples included in the study from el-Wad Terrace with especially low frequencies of common mice may be too few and represent a still limited extent of the spatial variability in the excavated area to allow a reliable comparison with the LN and FN samples. In addition, it is important to keep in mind that remains of certain taxa in the assemblages may mainly represent food debris. Extracting remains of such taxa from the analysis will alter the rela-tive frequencies. For example, extracting mole rats and squirrels from the computations increases the frequencies of common mice to 40% in one of the samples from Raqefet Cave (Loc. III) and to 50% in two of the Eynan samples (Structures 235 and 200/222/240). In the remainder of the assemblag-es common mouse frequencies remain below or increase to only slightly above 30% in this case. Extracting certain taxa from frequency calculations should be based on more detailed taphonomic data from each of the assemblages, however. Tchernov (1984) reported on relatively high frequencies of common mice from two Natufian sites: Hayonim Cave – 48% and Eynan – 39%. In both instances the report does not differentiate chronologically the deposits containing the assem-blages and it is unclear whether they represent the Natufian as a whole or one or more of the separate phases. It is therefore difficult to compare these earlier data with our own or to interpret them in terms of temporal developments in levels of human settlement occupation within the Natufian. An additional taphonomic scenario for the for-mation of micromammalian assemblages in Natu-fian sites is the in situ accumulation of remains of taxa that coexisted with people within and in the immediate surroundings of Natufian settlements. Such in situ assemblages can be expected to reveal more direct information on the ecological impact of

712
Lior Weissbrod et al.
Natufian settlements at the local scale and in turn provide indications on commensalism and the level of human settlement occupation. Identification of in situ micromammalian assemblages in Natufian sites and their separation from predator accumulat-ed assemblages or human food debris will depend on a combination of taphonomic criteria and examina-tion of the spatial association of the remains with discrete contexts of accumulation such as structures or various installations where the introduction of predator derived material is less likely. Relevant taphonomic criteria can include the presence or absence of predator modifications or charring of skeletal elements due to food preparation activities on the one hand and a high level of completeness in representation of skeletal elements and/or high survivorship of individuals which may characterize in situ deposition of commensal animal remains, on the other (see Weissbrod et al. 2005). Data from our study showing considerable variability in mi-cromammalian assemblages within Natufian sites and especially in relation to structures supports such a focus on identification of in situ assemblages and on their implications for cultural and social activities.
Conclusions
We examined variability in micromammalian assemblages from Natufian sites to assess the potential for preservation of information on the ecological impact of Natufian settlements and the level of human occupation of the settlements. Our examination shows that such assemblages can represent highly complex palimpsests of data on multiple environmental and cultural processes that may be analyzed at varying spatial and temporal scales. The two main findings of this study include the indication for a low level of human disturbance of the environment at the landscape scale during the Late Natufian and the significance of spatial variability in taxonomic composition and hetero-geneity within Natufian sites for investigating the question of commensalism and human mobility versus sedentism. We suggest a framework for the interpretation of information on the ecological impact of Natufian settlements combining consid-erations of taxonomic diversity, taphonomy, and spatial association of the remains within sites. Thus, the combination of low taxonomic diversity, indication for in situ accumulation of the remains, and association of the remains with living floors inside structures, graves, bedrock mortars, hearths and possibly other engineered features may be used
to distinguish assemblages containing information on the ecological impact of settlements at a localized spatial scale from other assemblages which were accumulated by predators and represent wider environmental variability at the landscape scale. Continued research on the question of commen-salism and changing level of human settlement occupation in the Natufian should focus on identi-fication of in situ accumulated micromammalian assemblages and analysis of variability on a larger scale. This should encompass additional material from Early Natufian contexts and other contexts from periods both preceding and succeeding the Natufian. Such an approach is currently needed if we are to establish a more detailed understanding of both evolutionary developments and variability within Natufian settlement systems.
Acknowledgments
We wish to thank the editors, Ofer Bar-Yosef and François Valla, for inviting us to contribute to this important volume. We extend a special thanks to François Valla and Hamoudi Khalaily for providing micromammalian material from their excavations at the site of Eynan. The study of variability in micromammalian assemblages from Natufian sites was conducted during the tenure of L.W. at the Zinman Institute of Archaeology supported by a post-doctoral research fellowship from the University of Haifa. L.W. would also like to thank the Kenya government, Ministry of Science and Technology, the National Museums of Kenya, and Maasai residents of Eselenkei Group Ranch in Kajiado District, Kenya for allowing his study of the living micromammalian communities of Maasai settlements. His research in Kenya was supported in part by a grant from the National Science Foundation (no. BCS-0536507).
References Cited
Andrzejewski, R., Babińska-Werka, J., Gliwicz, J. and J. Goszczyński
1978 Synurbization processes in population of Apodemus agrarius I. Characteristics of populations in an urbanization gradient. Acta Theriologica 23:341-358.
Auffray, J-C., Tchernov, E., Bonhomme, F., Heth, G., Simson, S. and E. Nevo
1990 Presence and ecological distribution of Mus “spretoides” and Mus musculus do-

713
Commensalism: Was it Truly a Natufian Phenomenon?…
mesticus in Israel, circum-Mediterranean vicariance in the genus Mus. Zeitschrift für Säugetierkunde 55:1-10.
Auffray, J-C., Tchernov, E. and E. Nevo 1988 Origine du commensalisme de la souris
domestique (Mus musculus domesticus) vis-à-vis de l’homme. Comptes- Rendus de l’Académie des Sciences Paris 307:517-522.
Bar-Matthews, M., Ayalon, A., Gilmour, M., Matthews, A. and C. J. Hawkesworth
2003 Sea-land oxygen isotopic relationships from planktonic foraminifera and spe-leothems in the Eastern Mediterra-nean region and their implication for paleorainfall during interglacial inter-vals. Geochimica et Cosmochimica Acta 67:3181-3199.
Bar-Oz, G., Dayan, T., Kaufman, D. and M. Weinstein-Evron
2004 The Natufian economy at el-Wad Terrace with special reference to gazelle exploita-tion patterns. Journal of Archaeological Science 31:217-231.
Bar-Yosef, O. 1983 The Natufian in the southern Levant.
In The Hilly Flanks and Beyond: Essays on the Prehistory of Southwestern Asia, edited by T. C. Young, P. E. L. Smith and P. Mortensen, pp. 11-42. Oriental Institute, University of Chicago, Chicago.
1991 The archaeology of the Natufian layer at Hayonim Cave. In The Natufian Culture in the Levant, edited by O. Bar-Yosef and F. R. Valla, pp. 81-92. International Monographs in Prehistory, Ann Arbor.
2002 Natufian and the Early Neolithic: Social and economic trends in southwestern Asia. In Examining the Farming/Lan-guage Dispersal Hypothesis, edited by P. Bellwood and C. Renfrew, pp. 113-126. McDonald Institute for Archaeological Research, Cambridge.
Bar-Yosef, O. and A. Belfer-Cohen 1989 The origins of sedentism and farming
communities in the Levant. Journal of World Prehistory 3:447-498.
Bar-Yosef, O. and E. Tchernov 1966 Archaeological finds and the fossil
faunas of the Natufian and microlithic industries at Hayonim Cave (Western Galilee, Israel). Israel Journal of Zoology 15:121-140.
Belfer-Cohen, A. and O. Bar-Yosef 2000 Early sedentism in the Near East: A
bumpy road to village life. In Life in Neo lithic Farming Communities: Social Organization, Identity, and Differentiation, edited by I. Kujit, pp. 19-37. Kluwer Academic Publishers, New York.
Berry, R. J. 1981 Town mouse, country mouse: adaptation
and adaptability in Mus Domesticus (M. musculus domesticus). Mammal Review 11:92-130.
Binford, L. R. 1968 Post-Pleistocene adaptations. In New
Perspectives in Archaeology, edited by S. R. Binford and L. R. Binford, pp. 313-341. Aldine, Chicago.
Bouchud, J. 1987 La Fauna du Gesement Natufien de
Mallaha (Eynan), Israel. Mémoires et Travaux du Centre de Recherche Français de Jérusalem 4. Association Paléorient, Paris.
Boursot, P., Auffray, J-C., Britton-Davidian, J. and F. Bonhomme
1993 The evolution of house mice. Annual Review of Ecology and Systematics 24:119-152.
Braithwaite, R. W. 1980 The ecology of Rattus lutreolus III*. The
rise and fall of a commensal population. Australian Wildlife Research 7:199-215.
Byrd, B. F.1989 The Natufian: settlement variability and
economic adaptation in the Levant at the end of the Pleistocene. Journal of World Prehistory 3:159-197.
2005 Reassessing the emergence of village life in the Near East. Journal of Archaeolog-ical Research 13:231-290.
Canova, L. and M. Fasola 1994 Population density and diet of the Spiny
Mouse Acomys cf. cahirinus in a desertic area of northern Kenya. Revue d’Écologie 49:87-90.
Charter, M., Izhaki, I., Shapira, L. and Y. Leshem 2007 Diets of urban breeding barn owls (Tyto
alba) in Tel Aviv, Israel. The Wilson Journal of Ornithology 119:484-485.
Charter, M., Izhaki, I., Meyrom, K., Motro, Y. and Y. Leshem
2009 Diets of barn owls differ in the same agricultural region. The Wilson Journal of Ornithology 121:378-383.

714
Lior Weissbrod et al.
Chernousova, N. F. 2001 Specific features of the dynamics of
murine rodent communities under the effects of urbanization. 1. Dynamics of species composition and abundance. Russian Journal of Ecology 32:122-125.
2002 Specific features of the dynamics of murine rodent communities under the effects of urbanization: 2. Maintenance of population size. Russian Journal of Ecology 33:23-27.
Courtney, P. A. and M. B. Fenton 1976 The effects of a small rural garbage dump
on populations of Peromyscus Leucopus Rafinesque and other small mammals. Journal of Applied Biology 13:413-422.
Devignat, R. 1946 Aspects de l’ épidémiologie de la peste au
Lac Albert. Annales de la Société Belge de Médecine Tropicale 26:13-54.
Dickman, C. R. 1992 Commensal and mutualistic interactions
among terrestrial vertebrates. Trends in Ecology & Evolution (Amsterdam) 7:194.
2006 Species interactions: Indirect effects. In Ecology: An Australian Perspective, edited by P. Attiwill and B. A. Wilson, pp. 303-315. Oxford University Press, Oxford.
Dor, M. 1982 The barn owl as an aid to science. The
Torgos 5:29-35 (in Hebrew).Edwards, P. C.
1989 Problems of recognizing earliest seden-tism: The Natufian example. Journal of Mediterranean Archaeology 2:5-48.
Evans, J. G. 1978 An Introduction to Environmental Ar-
chaeology. Paul Elek, London.Flannery, K. V.
1972 The origins of the village as a settlement type in Mesoamerica and the Near East: A comparative study. In Man, Settlement, and Urbanism, edited by P. J. Ucko, R. Tringham and G. W. Dimbleby, pp. 23-53. Duckworth, London.
Ganem, G. 1991 A comparative study of different popula-
tions of Mus musculus: Emotivity as an index of adaptation to commensalism. Comparative Biochemistry and Physiol-ogy 99A:531-536.
1993 Ecological characteristics of Robertsonian populations of the house mouse. Is their
habitat relevant to their evolution? Mammalia 57:349-357.
Garrod, D. A. E. 1957 The Natufian culture: the life and
economy of a Mesolithic people in the Near East. Proceedings of the British Academy 43:211-227.
Garrod, D. A. E. and D. M. A. Bate 1937 The Stone Age of Mount Carmel. Claren-
don Press, Oxford.Grosman, L.
2003 Preserving cultural traditions in a period of instability: The Late Natufian of the Hilly Mediterranean Zone. Current Anthropology 4:571-580.
Grosman, L. and N. D. Munro 2007 The sacred and the mundane: Domestic
activities at a Late Natufian burial site in the Levant. Before Farming 4: article 4.
Grosman, L., Munro, N. D. and A. Belfer-Cohen 2008 12,000-year-old Shaman burial from the
southern Levant (Israel). Proceedings of the National Academy of Sciences 105:17665-17669.
Henry, D. O. 1985 Preagricultural sedentism: The Natu-
fian example. In Prehistoric Hunt-er-Gatherers: The Emergence of Cultural Complexity, edited by D. T. Price and J. A. Brown, pp. 365-384. Academic Press, San Diego.
Hesse, B. 1979 Rodent remains and sedentism in the
Neolithic: Evidence from Tepe Ganj Dareh, Western Iran. Journal of Mam-malogy 60:856-857.
Jones, C. G., Lawton, J. H. and M. Shachak 1994 Organisms as ecosystem engineers. Oikos
69:373-386. 1997 Positive and negative effects of organisms
as physical ecosystem engineers. Ecology 78:1946-1957.
Kahila Bar-Gal, G., Aviel, S., Aram, E., Ginsburg, Z. and E. Tchernov
2000 Introduction of barn owl (Tyto alba) as an effective biological control agent of rodent populations in Israel. Israel Journal of Zoology 46:373.
Kaufman, D. 1992 Hunter-gatherers of the Levantine Epi-
palaeolithic: The socioecological origins of sedentism. Journal of Mediterranean Archaeology 5:165-201.

715
Commensalism: Was it Truly a Natufian Phenomenon?…
Kent, S. (editor) 1989 Farmers as Hunters: The Implications of
Sedentism. Cambridge University Press, Cambridge.
Kingdon, J. 1974 East African Mammals: An Atlas of
Evolution in Africa. Vol II, Part B: Hares and Rodents. Academic Press, New York.
Krebs, C. J. 1999 Ecological Methodology. Benjamin/
Cummings, Menlo Park.Kupstein, H.
2001 The use of small-mammals community composition as a bio-indicator to the interference extent in ecological systems hosting outdoor recreation activities. M.A. thesis, University of Haifa.
Lehmann, T. and A. Perevolotsky 1992 Small mammals in coniferous plantations
and native environment in southern Mt. Carmel, Israel. Mammalia 56:575-585.
Lengyel, G. and F. Bocquentin 2005 Burials of Raqefet Cave in the context of
the Late Natufian. Journal of the Israel Prehistoric Society - Mitekufat Haeven 35:271-284.
Lengyel, G., Nadel, D., Tsatskin, A., Bar-Oz, G., Bar-Yosef Mayer, D. E., Be’eri, R. and I. Hershkovitz
2005 Back to Raqefet Cave, Mount Carmel, Israel. Journal of the Israel Prehistoric Society - Mitekufat Haeven 35:245-270.
Leroi-Gourhan, Arl. 1984 L’environnement de Mallaha (Eynan) au
Natoufien. Paléorient 10:103-105. Lev-Yadun, S. and M. Weinstein-Evron
2005 Modeling the influence of Natufian el-Wad on the forest of Mount Carmel. Journal of the Israel Prehistoric Society - Mitekufat Haeven 35:285-298.
Marder, O., Yeshurun, R., Smithline, H., Ackermann, O., Bar-Yosef Mayer, D. E., Belfer-Cohen, A., Grosman, L., Hershkovitz, I., Klein N. and L. Weissbrod
herein Hof Shahaf: A New Natufian Site on the Shore of Lake Kinnere, edited by O. Bar-Yosef and F. R. Valla, International Monographs in Prehistory, Ann Arbor.
Misonne, X. 1963 Les Rongeurs du Ruwenzori et des Régions
Voisines. Institut des Parcs Nationaux du Congo et du Rwanda, Brussels.
Nadel, D., Lengyel, G., Bocquentin, F., Tsatskin, A., Rosenberg, D., Yeshurun, R., Bar-Oz, G.,
Bar-Yosef Mayer, D.E., Beeri, R., Conyers, L., Filin, S., Hershkovitz, I., Kurzawska, A. and L. Weissbrod
2008 The Late Natufian at Raqefet Cave: The 2006 Excavation Season. Journal of the Israel Prehistoric Society - Mitekufat Haeven 38:59-131.
Nadel, D. and G. Lengyel 2009 Human-made bedrock holes (mortars
and cupmarks) as a Late Natufian social phenomenon. Archaeology, Ethnology and Anthropology of Eurasia 37:37-48.
Nadel, D., Lengyel, G., Cabellos, T., Bocquentin, F., Rosenberg, D., Yeshurun, R., Brown-Goodman, R., Tsatskin, A., Bar-Oz, G. and S. Filin
2009 The Raqefet Cave 2008 excavation season. Journal of the Israel Prehistoric Society - Mitekufat Haeven 39:21-62.
Perrot, J. 1968 Premiers villages de Syrie et de Pales-
tine. Comptes-Rendus des Séances de l’Académie des Inscriptions et Belles-Lettres 112:161-177.
Pocock, M. J. O., Searle, J. B. and P. C. L. White 2004 Adaptations of animals to commensal
habitats: population dynamics of house mice Mus musculus domesticus on farms. Journal of Animal Ecology 73:878-888.
Rajabi-Maham, H., Orth, A. and F. Bonhomme 2008 Phylogeography and postglacial expan-
sion of Mus musculus domesticus inferred from mitochondrial DNA coalescent, from Iran to Europe. Molecular Ecology 17:627-641.
Reitz, E. J. and E. S. Wing 1999 Zooarchaeology. Cambridge University
Press, Cambridge.Sakai, A. K., Allendorf, F. W., Holt, J. S., Lodge, D. M. M.,Jane, With, K. A., Baughman, S., Cabin, R. J., Cohen, J. E., Ellstrand, N. C., McCauley, D. E., O’Neil, P., Parker, I. M., Thompson, J. N. and S. G. Weller
2001 The population biology of invasive species. Annual Review of Ecology and Systematics 32:305-332.
Setzer, H. W. 1959 The spiny mice (Acomys) of Egypt.
Journal of the Egyptian Public Health Association 34:93-101.
Smith, B. D. 2007 Human niche construction and the be-
havioural context of plant and animal domestication. Evolutionary Anthropol-ogy 16:188-199.

716
Lior Weissbrod et al.
Stein, M., Torfstein, A., Gavrieli, I. and Y. Yechieli 2009 Abrupt aridities and salt deposition in
the post-glacial Dead Sea and their North Atlantic connection. Quaternary Science Reviews 29:567-575.
Tangri, D. and G. Wyncoll 1989 Of mice and men: Is the presence of
commensal animals in archaeological sites a positive correlate of sedentism? Paléorient 15/1:85-94.
Tchernov, E. 1984 Commensal animals and human seden-
tism in the Middle East. In Animals and Archaeology, 3: Early Herders and Their Flocks, edited by J. Clutton-Brock and C. Grigson, pp. 91-115. BAR International Series 202. Oxford.
1991a Biological evidence for human sedentism in Southwest Asia during the Natufian. In The Natufian Culture in the Levant, edited by O. Bar-Yosef and F. R. Valla, pp. 315-340. International Monographs in Prehistory. Ann Arbor.
1991b Of mice and men. Biological markers for long-term sedentism: A reply. Paléorient 17/1:153-160.
Valla, F. R. 1998 Natufian seasonality? A guess. In Season-
ality and Sedentism: Archaeological Per-spectives from Old and New World Sites, edited by T. R. Rocek and O. Bar-Yosef, pp. 93-108. Peabody Museum Bulletin 6. Peabody Museum of Archaeology and Ethnology, Harvard University, Cam-bridge.
Valla, F. R., Bar-Yosef, O., Smith, E., Tchernov, E. and J. Desse
1986 Un nouveau sondage sur la terrasse d’el Ouad, Israël. Paléorient 12/1:31-36.
Valla, F. R. and H. Khalaily 1997 The first sedentary peoples in Israel:
Mallaha (Eynan) 1996. Bulletin du Centre de Recherche Français de Jérusalem 1:72-82.
Valla, F. R., Khalaily, H., Samuelian, N., Bocquentin, F., Delage, C., Valentin, B., Plisson, H., Rabinovich, R. and A. Belfer-Cohen
1998 Le Natufien final et les nouvelles fouilles à Mallaha (Eynan), Israël, 1996-1997. Journal of the Israel Prehistoric Society - Mitekufat Haeven 28:105-176.
Valla, F. R., Khalaily, H., Samuelian, N. and F. Bocquentin
2002 From foraging to farming: The contribu-
tion of the Mallaha (Eynan) excavations, 1996–2001. Bulletin du Centre de Recher-che Français de Jérusalem 10:71-90.
Valla, F. R., Khalaily, H., Valladas, H., Kaltnecker, E., Bocquentin, F., Cabellos, T., Bar-Yosef Mayer, D. E., Le Dosseur, G., Regev, L., Chu, V., Weiner, S., Boaretto, E., Samuelian, N., Valentin, B., Delerue, S., Poupeau, G., Bridault, A., Rabinovich, R., Simmons, T., Zohar, I., Ashkenazi, S., Delgado Huertas, A., Spiro, B., Mienis, H., Rosen, A. M., Porat, N. and A. Belfer-Cohen
2007 Les fouilles de Ain Mallha (Eynan) de 2003 à 2005: Quatrième rapport prélim-inaire. Journal of the Israel Prehistoric Society - Mitekufat Haeven 37:135-383.
Weinstein-Evron, M. 1998 Early Natufian el-Wad Revisited. ERAUL
77, Liège.2009 Archaeology in the Archives: Unveiling
the Natufian Culture of Mount Carmel. Brill and American School of Prehistoric Research Monograph Series, Boston.
Weinstein-Evron, M., Kaufman, D., Bachrach, N., Bar-Oz, G., Bar-Yosef Mayer, D. E., Chaim, S., Druck, D., Groman-Yaroslavski, I., Hershkovitz, I., Liber, N., Rosenberg, D., Tsatskin, A. and L. Weissbrod
2007 After 70 years: new excavations at the el-Wad Terrace, Mount Carmel. Journal of the Israel Prehistoric Society - Mitekufat Haeven 37:37-134.
Weinstein-Evron, M., Kaufman, D. and R. Yeshurun
herein Spatial Organization of Natufian el-Wad through Time: Combining the Results of Past and Present Excavations. In The Natufian Culture in the Levant II, edited by O. Bar-Yosef and F. R. Valla, International Monographs in Prehistory, Ann Arbor.
Weissbrod, L. 2010a The small animals of Maasai settlements:
Ethnoarchaeoogical investigations of the commensalism model. Ph.D. dissertation, Washington University in Saint Louis, Saint Louis.
2010b Biological indicators of occupation inten-sity: An environmental ethnoarchaeology of Maasai settlements. In The Archaeology of Anthropogenic Environments, edited by R. M. Dean, pp. 295-320. Center for Archaeological Investigations, Southern Illinois University, Carbondale.

717
Commensalism: Was it Truly a Natufian Phenomenon?…
Weissbrod, L., Dayan, T., Kaufman, D. and M. Weinstein-Evron
2005 Micromammal taphonomy of el-Wad Ter-race, Mount Carmel, Israel: distinguish-ing natural from cultural depositional agents in the Late Natufian. Journal of Archaeological Science 32:1-17.
Yeshurun, R., Bar-Oz, G., Kaufman, D. and M. Weinstein-Evron
herein Domestic Refuse Maintenance in the Natufian: Faunal Evidence from el-Wad Terrace, Mount Carmel, edited by O. Bar-Yosef and F. R. Valla, International Monographs in Prehistory, Ann Arbor.