coexistence of the native benthic amphipod diporeia spp. and exotic dreissenid mussels in the new...
TRANSCRIPT

Journal of Great Lakes Research 38 (2012) 226–235
Contents lists available at SciVerse ScienceDirect
Journal of Great Lakes Research
j ourna l homepage: www.e lsev ie r .com/ locate / jg l r
Coexistence of the native benthic amphipod Diporeia spp. and exotic dreissenidmussels in the New York Finger Lakes
James M. Watkins a,⁎, Lars G. Rudstam a,1, Edward L. Mills a,1, Mark A. Teece b,2
a Cornell Biological Field Station, 900 Shackleton Point Rd, Bridgeport, NY 13030, USAb State University of New York College of Environmental Science and Forestry, 1 Forestry Dr., Syracuse, NY 13210, USA
⁎ Corresponding author. Tel.: +1 315 633 9243.E-mail addresses: [email protected] (J.M. Watkin
(L.G. Rudstam), [email protected] (E.L. Mills), mteece@1 Tel.: +1 315 633 9243.2 Tel.: +1 315 470 4736.
0380-1330/$ – see front matter © 2012 International Adoi:10.1016/j.jglr.2012.02.001
a b s t r a c t
a r t i c l e i n f oArticle history:Received 24 March 2011Accepted 18 January 2012Available online 29 February 2012
Communicated by Christine Mayer
Index words:DiporeiaDreissenaFinger LakesFatty acids
Populations of the benthic amphipod Diporeia spp. have sharply declined since the early 1990s in all NorthAmerica's Great Lakes except Lake Superior. The onset and continued decline coincides with the invasion ofthese lakes by zebra (Dreissena polymorpha) and quagga (Dreissena rostriformis bugensis) mussels and thespread of quagga mussels to deep habitats. The six deepest Finger Lakes of central New York (Seneca,Cayuga, Skaneateles, Canandaigua, Keuka, and Owasco) have historically been Diporeia habitat and havehad dreissenids for more than a decade. These lakes represent a wide range of trophic state, maximumdepth, and dreissenid invasion history. We hypothesized that Diporeia abundance would be negatively im-pacted by dreissenid mussel expansion in the Finger Lakes. During 2006–2010, we sampled Diporeia andmussel populations in these six lakes. Diporeia was present in all six lakes, and was abundant (2000/m2)in Owasco Lake that has only zebra mussels and in Cayuga and Seneca Lakes that have had zebra and quag-ga mussels since 1994. Diporeia abundance was lowest (1000/m2) in Skaneateles, Canandaigua, and KeukaLakes where quagga mussels have recently expanded. Productivity indicators explained much of the vari-ability of Diporeia abundance. The persistence of Diporeia with quagga mussels in these lakes may be be-cause of available alternative food resources. Fatty acid tracers indicate that Diporeia from Owasco Lake,the lake without quagga mussels, utilize diatoms, but Diporeia from Cayuga Lake that coexist with abun-dant quagga mussels also use food resources associated with terrestrial detritus that cannot be interceptedby dreissenids.
© 2012 International Association for Great Lakes Research. Published by Elsevier B.V. All rights reserved.
Introduction
The benthic amphipod Diporeia (formerly known as Pontoporeia)has historically dominated soft substrate habitats of deep lakes ofNorth America, often comprising more than 60% of total benthic bio-mass in deep (>20 m) habitats (Sly and Christie, 1992). This burrowingamphipod feeds primarily on diatoms that sink to the sediment surfaceand it is a lipid-rich prey for many fish species including slimy sculpins(Cottus cognatus), lake whitefish (Coregonus clupeaformis) and deepwa-ter coregonids (C. hoyi and C. kiyi) (Owens and Dittman, 2003).Diporeiatherefore represents an important link between phytoplankton and fishproduction.Diporeia has long been recognized as an indicator organismfor assessing the condition of the offshore component of the Great Lakesecosystem (Neilson et al., 2003; Ryder and Edwards, 1985).
During the early 1990s, the disappearance of Diporeia was ob-served in shallow habitats in eastern Lake Erie (Dermott and Kerec,
s), [email protected] (M.A. Teece).
ssociation for Great Lakes Research.
1997), eastern Lake Ontario (Dermott, 2001), southern Lake Michigan(Nalepa et al., 1998), and Saginaw Bay of Lake Huron (Nalepa et al.,2003) soon after exotic zebra mussels (Dreissena polymorpha) andquagga mussels (Dreissena rostriformis bugensis) colonized theselakes. By the late 1990s, Diporeia at intermediate depths (30–90 m)in Lake Ontario were also in decline (Dermott and Geminiuc, 2003;Lozano et al., 2001; Watkins et al., 2007). This pattern of Diporeia de-cline associated with quagga mussel expansion into deeper habitatshas been repeated in lakes Michigan and Huron, and Diporeia has dis-appeared from all but the deepest (>100 m) habitats in these lakes(Nalepa et al., 2009). Diporeia populations have remained stable inLake Superior (Auer et al., 2009; Scharold et al., 2004), the onlyGreat Lake where offshore dreissenids are absent. Diporeia popula-tions have also persisted in many smaller lakes in Canada and thenortheastern United States without dreissenid mussels (Dermott etal., 2005). Together this suggests a coupling between Diporeia declineand the invasion of the dreissenid mussels, in particular the quaggamussel, possibly due to competition for the settling spring diatombloom or mussels acting as a vector for some pathogen.
Watkins et al. (2007) and Nalepa et al. (2009) noted that Diporeiadeclined in deeper waters in both lakes Ontario and Michigan beforequagga mussels were abundant at these depths. Spring diatom
Published by Elsevier B.V. All rights reserved.

Table 1Limnological characteristics of the six deepest Finger Lakes sampled in this study.Water quality parameters from Halfman and O'Neill (2009).
Lake Halfman andO'Neill (2009)values for 2005–2008
Maximum Average Surface TP chl a Secchi depth
Depth (m) Depth (m) Area (km2) μg/L μg/L m
Seneca 198.4 88.6 175.4 9.0 3.4 4.7Cayuga 132.6 54.5 172.1 9.7 3.2 4.1Skaneateles 90.5 43.5 35.9 3.7 0.7 7.6Canandaigua 83.5 38.8 42.3 7.8 1.7 6.5Keuka 55.8 30.5 47.0 6.3 2.0 5.3Owasco 54.0 29.3 26.7 7.5 2.8 4.1
227J.M. Watkins et al. / Journal of Great Lakes Research 38 (2012) 226–235
blooms in cold deep-water lakes occur when the water column is wellmixed, and quagga mussel biomass at intermediate depths in LakeMichigan has reached a level that likely exerts significant grazingpressure at that time (Vanderploeg et al., 2010) with possible nega-tive effects on Diporeia. This observation provides a plausible mecha-nism for deep mussel beds to intercept Diporeia's food supply.Counter to the food limitation hypothesis, Diporeia populations de-clined in Lake Michigan in the late 1990s despite sufficient flux of or-ganic matter reaching the sediments (Nalepa et al., 2006).
One notable exception to the possible coupling between musselincrease and Diporeia decline is that Diporeia has persisted in twolakes, Cayuga and Seneca Lakes of central New York, as recently as2001 despite the presence of dreissenid mussels for more than a de-cade (Dermott et al., 2005). There are six Finger Lakes (Seneca, Cayu-ga, Skaneateles, Canandaigua, Keuka, and Owasco) that are suitablydeep (>50 m maximum depth) for Diporeia habitat and have year-round oxygenated cold hypolimnia (Table 1, Callinan, 2001). Withina small geographic area, these lakes represent a broad range of tro-phic states ranging from oligotrophic Skaneateles Lake to mesotro-phic Owasco Lake (Table 1, Halfman and O'Neill, 2009). All six lakeshave dissolved calcium levels >28 mg/L and are therefore “highrisk” for invasion by dreissenid mussels (Callinan, 2001; Whittier etal., 2008). Zebra mussels were reported in the lakes at differenttimes, as early as 1991 in Cayuga Lake and as late as 1997 in OwascoLake (Table 2, USGS NAS Database). Two of the lakes, Cayuga and Sen-eca Lakes, have had quagga mussels since 1994 and quagga musselswere reported in Keuka Lake in 2008 (USGS NAS Database). Dreisse-nid establishment has been associated with increased water clarity inthe Finger Lakes but diatoms are still an important component of thephytoplankton community throughout the spring and summer(Hammers et al., 2007). The relative trophic status of the six lakesranging from oligotrophic to mesotrophic (Halfman and O'Neill,2009) remains similar to that reported in the 1970s prior to dreisse-nid introduction (Schaffner and Oglesby, 1978). Therefore, the sixlakes offer the opportunity to test several factors that regulate Dipor-eia abundance including trophic state, depth, and the abundance ofthe two dreissenid species.
Table 2Year of first reported and confirmed sighting of Dreissena polymorpha (zebra mussels)and Dreissena rostriformis bugensis (quagga mussels) in the New York Finger. Lakes(USGS NAS Database and this study).
Zebra Quagga
Seneca 1992 1994Cayuga 1991 1994Skaneateles 1995 2010Canandaigua 1994 2010Keuka 1994 2008Owasco 1997 Not present
In this paper we examine patterns of coexistence of the native am-phipod Diporeia and exotic dreissenid mussels in six Finger Lakes ofcentral New York State. We expected to see a negative impact ofdreissenid expansion on the abundance of Diporeia in these lakes,particularly for Cayuga and Seneca Lakes that have had quagga mus-sels for a longer time period. We also expected dreissenid impacts tobe higher for oligotrophic lakes with a lower supply of food and shal-low lakes with less suitable thermal habitat for Diporeia.
Methods
Benthic sampling
Our sampling of the profundal benthic communities of Cayuga Lakespanned from 2006 to 2009. In 2006, we conducted exploratory sam-pling (Fig. 1) at five sites near the northern end (Long Point StatePark) and 12 sites near the southern end (south of Myers Point) witha petite Ponar grab (area sampled per grab 0.023 m2). In 2007, webegan sampling a 100-m site west of Myers Point (43.372 N,79.353 W) that had been sampled monthly throughout 1964 (Green,1965). In 2008–2009, we collected triplicate grabs each month at twosites (40 and 80 m depths) near this location. The substrate at thethree sites is rich in coarse terrestrial organic matter such as leaf litterdue to a location near the inflow of Salmon Creek. In total 57 grabswere collected offshore of Myers Point from 2007 to 2009.
Our most recent benthic sampling in the other New York FingerLakes was during the summer and fall of 2010 using a petite Ponargrab. Triplicate samples were collected at three to six different depthsin Canandaigua, Keuka, Skaneateles, Owasco, and Seneca Lakes(Fig. 1).
Historical data for profundal benthic communities of Keuka, Ska-neateles, and Owasco Lakes are scarce. However, we can comparethe 2010 data to our previous sampling of Skaneateles Lake in 2006and Owasco Lake from 2006 to 2008. In 2006, we conducted explor-atory sampling of the two lakes using a petite Ponar grab, with atotal of 21 grabs taken in Skaneateles Lake and 14 grabs in OwascoLake. In 2007, we conducted two lake-wide benthic surveys ofOwasco Lake using a standard Ponar grab (area sampled per grab0.05 m2). Triplicate grabs were collected at 10 sites along a north–south transect. Sites were distributed every 10 m depth both northand south of the 50-m deep central basin. The transect was resampledin 2007 and 2008. In total, 124 grabs were collected from OwascoLake between 2006 and 2008 (Fig. 1).
Canandaigua, Cayuga, and Seneca Lakes have been sampled exten-sively in the past, starting with Birge and Juday's sampling of thethree lakes in 1918 with an Ekman dredge (Birge and Juday (1921,area sampled per grab 0.05 m2). Canandaigua Lake was the subjectof lake-wide benthic surveys (Ekman dredge) in 1927 and 1974(Eaton et al., 1975). Cayuga Lake was sampled in 1952–53 (orangepeel dredge, Henson, 1954), 1964 (Ekman dredge, Green, 1965),and 1994 and 2001 (Ekman dredge, Dermott et al., 2006). SenecaLake was sampled in 2001 (Ekman dredge, Dermott et al., 2006).
Sample processing
The sediment collected was sieved through a 500-μm sieve on theboat, and remaining material was preserved in 85% ethanol. All ben-thic organisms were sorted, identified, and enumerated in the lab.For the 2008 data at the 80-m site in Cayuga Lake, length of individualDiporeia and Dreissena were measured. For Diporeia, a PixeLINK cam-era (PL-B622CU) and the PixeLINK Capture SE Software was used torecord images through a Motic Series K Dissecting Scope (×6 or×12 magnification). A mm scale in the image background was usedfor calibration. Amphipod body length was measured by summingline segments along the curved path from the tip of the rostrum tothe base of the telson (Quigley and Lang, 1989). All Diporeia were
Fig. 1. Benthic sampling sites during 2006–2010 in the six deepest New York Finger Lakes.
228 J.M. Watkins et al. / Journal of Great Lakes Research 38 (2012) 226–235
measured from one of three triplicate grabs (each sample >150 indi-viduals) and binned at 1-mm size intervals. The 1964 data representall individuals from six Ekman grabs (0.05 m2) for each month thatwere measured with an ocular micrometer method and also binnedat 1-mm size intervals (Green, 1965). The histogram data presentedin this paper are standardized to numbers of individuals in each sizeinterval per petite Ponar grab (0.023 m2). For Dreissena in 2008, dig-ital photographs (300 dpi) of each sample including a cm scale wereprocessed using the freeware software ImageJ 1.43 (Image Processingand Analysis in Java, http://rsb.info.nih.gov/ij/). The program countsand measures the major axis of every object (traced as an ellipse) inan image in reference to a calibration scale. Shells were spread outon a white background for contrast. The size distribution data pre-sented were compiled from three images (triplicate grabs from onesite) for a total of 188 individual lengths binned at 0.25 cm intervalsthat were then normalized to a single grab. This automated imageanalysis method was comparable to manual caliper measurements.
Fatty acid analysis
We used fatty acid analysis for information on overall conditionand diet of Diporeia. Diporeia populations living in direct contactwith quagga mussels (Cayuga Lake) were compared with Diporeiapopulations with little direct contact with mussels (Owasco Lake).Seasonal differences were minimized by collecting both populationswithin a day of each other in June 2008 at 35-m depth sites. Samplingin June ensured that both populations had access to settling matterfrom phytoplankton blooms from the early spring. The temperaturerange is also small (5.7–7.2 °C) at this depth for both lakes duringthe summer (Watkins, unpublished). We collected Diporeia and sub-strate samples from each lake that were flash frozen soon after collec-tion and then freeze-dried.
It is important to note that the two sites selected to represent the twolakes had very different substrates. The Cayuga Lake site off of MyersPoint had abundant coarse (>500-μm) terrestrial organic matter (leaflitter), while benthic organisms were the only >500-μm component of
sediments at the Owasco Lake site. Future sampling will be necessaryto establish whether the detected differences in fatty acid compositionare lake-wide or due to local substrate conditions.
Concentrations of fatty acids in Diporeia and sedimentary sub-strates were measured according to methods of Teece et al. (2011).Eight individual adult Diporeia (7–8 mm in length) were selectedfrom each lake. Lyophilized aliquots were extracted three times in2:1 dichloromethane:methanol, combined, and dried under nitrogen.Extracts were esterified with methanolic hydrochloric acid (60 °C for60 min), and resulting fatty acid methyl esters extracted into hexanethree times and evaporated to dryness under a stream of nitrogen.The dry residue was silylated by the addition of bis(trimethylsilyl)-trifluoroacetamide (BSTFA, Sigma-Aldrich; 60 °C for 60 min) andthen dissolved in dichloromethane to ensure no residual BSTFA waspresent prior to gas chromatographic (GC) analysis. Samples weredissolved in 2,2,4-trimethylpentane and analyzed using a ShimadzuGC 17-A with a flame ionization detector (GC-FID). Fatty acids wereidentified using standard mixtures and concentrations (expressed as% total lipid) were determined relative to the internal standard (trico-sanoic acid). Control samples were treated in an identical fashion todetermine blank levels that were always below the fatty acid detec-tion limit of 1 ng mg−1, and the error in overall measurement was1–7%. Although a small sample size (n=8), the comparison is agood first step in testing whether the fatty acid signatures (and die-tary sources) of the two populations are unique.
Data analysis
We used stepwise multiple regression to model relationships be-tween lake-wide Diporeia abundance (integrated for ≥20 m depth)and several limnological parameters in the six lakes. We employedforward-selection in which models including each variable alone arecompared and the model with the smallest residual mean square isselected (Snedecor and Cochran, 1989). Then additional terms wereevaluated and retained if the explanatory power of the model in-creased over the single-variable model. The three trophic state

Fig. 2. (Print grayscale) Abundance of A) quaggamussels (Dreissena rostriformis bugensis),B) zebra mussels (Dreissena polymorpha) and C) the amphipod Diporeia in the six FingerLakes in 2010. Error bars represent one standard error. Symbols are Seneca (filled square),Cayuga (open square), Keuka (filled diamond), Owasco (open diamond), Canandaigua(filled triangle) and Skaneateles (open triangle).(Online color) Abundance of A) quaggamussels (Dreissena rostriformis bugensis), B) zebra mussels (Dreissena polymorpha) andC) the amphipod Diporeia in the six Finger Lakes in 2010. Error bars represent one stan-dard error. Symbols are Seneca (red square), Cayuga (orange square), Keuka (darkgreen diamond), Owasco (light green diamond), Canandaigua (dark blue triangle) andSkaneateles (light blue triangle).
Fig. 3. Abundance of Diporeia in Skaneateles in 2006 and 2010 (A) and Owasco Lake in2006, 2007, 2008, and 2010 (B). Error bars represent one standard error.
229J.M. Watkins et al. / Journal of Great Lakes Research 38 (2012) 226–235
parameters included surface values (averaged for 2005–2008) fortotal phosphorus (TP), chl a, and secchi depth (m) (Halfman andO'Neill, 2009). Diporeia abundance was also compared to the maxi-mum depth of each lake (Table 1) and integrated dreissenid abun-dance (average of all depths sampled). It is important to note thatthe trophic parameters are strongly correlated with each other butnot with dreissenid abundance. Although water clarity of all thelakes increased after dreissenid introduction, the relative differencesin the trophic state of the six lakes from oligotrophic to mesotrophichave remained consistent since the 1970s (Schaffner and Oglesby,1978).
Sites >20 m in the six lakes were selected to assess the relation-ship of mussel density and amphipod abundance in habitats because
they are in direct contact at this depth. Dreissenids are abundant atdepths b20 m, but Diporeia have historically been absent from thesesites because of high temperatures.
Fatty acid percentage data were arcsine squareroot transformed(Sokal and Rohlf, 1981), and statistical tests including t-tests, andone-way analysis of variance (ANOVA) followed by Tukey's HSDtest, were considered significant at α=0.05. Principal componentsanalysis (PCA) was run on arcsine squareroot transformed fatty acidproportion data using the software PC-ORD (McCune and Mefford,1999).
Results
Status of dreissenid mussels and Diporeia in the New York Finger Lakes
By 2010, quaggamussels were established in five out of the six deepFinger Lakes and outnumbered zebramussels. Cayuga and Seneca Lakeshad the highest abundances of quaggamussels, exceeding 10,000/m2 at20–30 m and 1000/m2 as deep as 60 m (Fig. 2A). In Canandaigua andSkaneateles Lakes, quagga mussels were also abundant at 20 mdepth (3980/m2 and 4928/m2) but decreased to 100/m2 by 40 m.In Keuka Lake, quagga mussels were abundant at 10 m but were rareat depths >15 m. Zebra mussels were rare in the five lakes in 2010(Fig. 2B).
Owasco Lake was the only study lake without quagga mussels. Zebramusselsweremost abundant at 10 mdepth averaging 5287/m2 (Fig. 2B).Zebra mussels were collected at low (b300/m2) abundances at deepersites, often clumped together as druzes on top of fine sediments.
Diporeia were not collected at depths b10 m in any of the six studylakes. The amphipod was collected from most sites deeper than 15 m,although it was notably rare from a 20-m site in Cayuga Lake and a30-m site in Seneca Lake (Fig. 2C). Abundance of Diporeia at 20-mdepth for the other four lakes was similar at 1000/m2. Diporeia abun-dance increased with depth in Cayuga, Seneca, and Owasco Lakes,
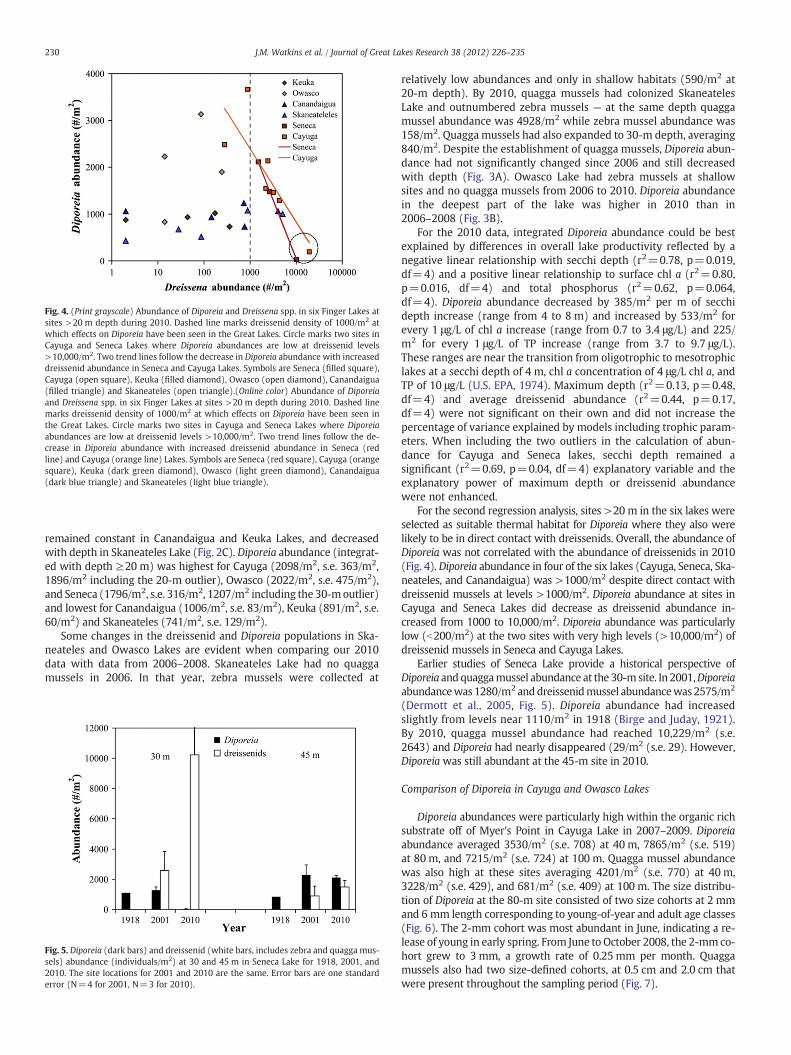
Fig. 4. (Print grayscale) Abundance of Diporeia and Dreissena spp. in six Finger Lakes atsites >20 m depth during 2010. Dashed line marks dreissenid density of 1000/m2 atwhich effects on Diporeia have been seen in the Great Lakes. Circle marks two sites inCayuga and Seneca Lakes where Diporeia abundances are low at dreissenid levels>10,000/m2. Two trend lines follow the decrease in Diporeia abundance with increaseddreissenid abundance in Seneca and Cayuga Lakes. Symbols are Seneca (filled square),Cayuga (open square), Keuka (filled diamond), Owasco (open diamond), Canandaigua(filled triangle) and Skaneateles (open triangle).(Online color) Abundance of Diporeiaand Dreissena spp. in six Finger Lakes at sites >20 m depth during 2010. Dashed linemarks dreissenid density of 1000/m2 at which effects on Diporeia have been seen inthe Great Lakes. Circle marks two sites in Cayuga and Seneca Lakes where Diporeiaabundances are low at dreissenid levels >10,000/m2. Two trend lines follow the de-crease in Diporeia abundance with increased dreissenid abundance in Seneca (redline) and Cayuga (orange line) Lakes. Symbols are Seneca (red square), Cayuga (orangesquare), Keuka (dark green diamond), Owasco (light green diamond), Canandaigua(dark blue triangle) and Skaneateles (light blue triangle).
230 J.M. Watkins et al. / Journal of Great Lakes Research 38 (2012) 226–235
remained constant in Canandaigua and Keuka Lakes, and decreasedwith depth in Skaneateles Lake (Fig. 2C). Diporeia abundance (integrat-ed with depth ≥20 m) was highest for Cayuga (2098/m2, s.e. 363/m2,1896/m2 including the 20-m outlier), Owasco (2022/m2, s.e. 475/m2),and Seneca (1796/m2, s.e. 316/m2, 1207/m2 including the 30-moutlier)and lowest for Canandaigua (1006/m2, s.e. 83/m2), Keuka (891/m2, s.e.60/m2) and Skaneateles (741/m2, s.e. 129/m2).
Some changes in the dreissenid and Diporeia populations in Ska-neateles and Owasco Lakes are evident when comparing our 2010data with data from 2006–2008. Skaneateles Lake had no quaggamussels in 2006. In that year, zebra mussels were collected at
Fig. 5. Diporeia (dark bars) and dreissenid (white bars, includes zebra and quagga mus-sels) abundance (individuals/m2) at 30 and 45 m in Seneca Lake for 1918, 2001, and2010. The site locations for 2001 and 2010 are the same. Error bars are one standarderror (N=4 for 2001, N=3 for 2010).
relatively low abundances and only in shallow habitats (590/m2 at20-m depth). By 2010, quagga mussels had colonized SkaneatelesLake and outnumbered zebra mussels — at the same depth quaggamussel abundance was 4928/m2 while zebra mussel abundance was158/m2. Quagga mussels had also expanded to 30-m depth, averaging840/m2. Despite the establishment of quagga mussels, Diporeia abun-dance had not significantly changed since 2006 and still decreasedwith depth (Fig. 3A). Owasco Lake had zebra mussels at shallowsites and no quagga mussels from 2006 to 2010. Diporeia abundancein the deepest part of the lake was higher in 2010 than in2006–2008 (Fig. 3B).
For the 2010 data, integrated Diporeia abundance could be bestexplained by differences in overall lake productivity reflected by anegative linear relationship with secchi depth (r2=0.78, p=0.019,df=4) and a positive linear relationship to surface chl a (r2=0.80,p=0.016, df=4) and total phosphorus (r2=0.62, p=0.064,df=4). Diporeia abundance decreased by 385/m2 per m of secchidepth increase (range from 4 to 8 m) and increased by 533/m2 forevery 1 μg/L of chl a increase (range from 0.7 to 3.4 μg/L) and 225/m2 for every 1 μg/L of TP increase (range from 3.7 to 9.7 μg/L).These ranges are near the transition from oligotrophic to mesotrophiclakes at a secchi depth of 4 m, chl a concentration of 4 μg/L chl a, andTP of 10 μg/L (U.S. EPA, 1974). Maximum depth (r2=0.13, p=0.48,df=4) and average dreissenid abundance (r2=0.44, p=0.17,df=4) were not significant on their own and did not increase thepercentage of variance explained by models including trophic param-eters. When including the two outliers in the calculation of abun-dance for Cayuga and Seneca lakes, secchi depth remained asignificant (r2=0.69, p=0.04, df=4) explanatory variable and theexplanatory power of maximum depth or dreissenid abundancewere not enhanced.
For the second regression analysis, sites >20 m in the six lakes wereselected as suitable thermal habitat for Diporeia where they also werelikely to be in direct contact with dreissenids. Overall, the abundance ofDiporeia was not correlated with the abundance of dreissenids in 2010(Fig. 4). Diporeia abundance in four of the six lakes (Cayuga, Seneca, Ska-neateles, and Canandaigua) was >1000/m2 despite direct contact withdreissenid mussels at levels >1000/m2. Diporeia abundance at sites inCayuga and Seneca Lakes did decrease as dreissenid abundance in-creased from 1000 to 10,000/m2. Diporeia abundance was particularlylow (b200/m2) at the two sites with very high levels (>10,000/m2) ofdreissenid mussels in Seneca and Cayuga Lakes.
Earlier studies of Seneca Lake provide a historical perspective ofDiporeia and quaggamussel abundance at the 30-m site. In 2001,Diporeiaabundancewas 1280/m2 anddreissenidmussel abundancewas 2575/m2
(Dermott et al., 2005, Fig. 5). Diporeia abundance had increasedslightly from levels near 1110/m2 in 1918 (Birge and Juday, 1921).By 2010, quagga mussel abundance had reached 10,229/m2 (s.e.2643) and Diporeia had nearly disappeared (29/m2 (s.e. 29). However,Diporeia was still abundant at the 45-m site in 2010.
Comparison of Diporeia in Cayuga and Owasco Lakes
Diporeia abundances were particularly high within the organic richsubstrate off of Myer's Point in Cayuga Lake in 2007–2009. Diporeiaabundance averaged 3530/m2 (s.e. 708) at 40 m, 7865/m2 (s.e. 519)at 80 m, and 7215/m2 (s.e. 724) at 100 m. Quagga mussel abundancewas also high at these sites averaging 4201/m2 (s.e. 770) at 40 m,3228/m2 (s.e. 429), and 681/m2 (s.e. 409) at 100 m. The size distribu-tion of Diporeia at the 80-m site consisted of two size cohorts at 2 mmand 6 mm length corresponding to young-of-year and adult age classes(Fig. 6). The 2-mm cohort was most abundant in June, indicating a re-lease of young in early spring. From June to October 2008, the 2-mmco-hort grew to 3 mm, a growth rate of 0.25 mm per month. Quaggamussels also had two size-defined cohorts, at 0.5 cm and 2.0 cm thatwere present throughout the sampling period (Fig. 7).

Fig. 6. Monthly size distribution of Diporeia at 80 m depth in Cayuga Lake off of Myer's Point in 1964 (Green, 1965) and 2008 (this study). The y-axis represents individuals perpetite Ponar (0.023 m2). The x-axis is length of individuals in mm.
231J.M. Watkins et al. / Journal of Great Lakes Research 38 (2012) 226–235
Individual adult Diporeia (7–8 mm in size) from Owasco Lake hadboth higher dry weight and lipid content than Diporeia from CayugaLake. The dry weight of 7–8 mm Diporeia from Owasco Lake was2.12 mg (s.e. 0.13) relative to 0.65 mg (s.e. 0.07) of Diporeia from Ca-yuga Lake (t-test, pb0.0001). Owasco Diporeia were also richer inlipids (51.6% of dry weight, s.e. 3.1%) than Cayuga Lake Diporeia(14.9% of dry weight, s.e. 3.3%) (t-test, pb0.0001). The specific lipidcomposition also differed for the two populations (Table 3). Saturatedfatty acids (SAFA), primarily palmitic acid (16:0), composed approx-imately 20% of the fatty acids of Diporeia in both lakes. Monounsatu-rated fatty acids (MUFAs) were 50.4% of the fatty acids of OwascoLake Diporeia and 41.7% of those from Cayuga Lake (t-test,p=0.0003). The common MUFA oleic acid (18:1w9) was a primaryfatty acid in both populations, approximately 20%. Owasco Diporeia
were particularly rich in the MUFA palmitoleic acid (16:1w7), 22.5%relative to Cayuga Diporeia at 7.0% (t-test, p=0.0001). Approximately5% of the fatty acids in Diporeia from both lakes were diunsaturatedfatty acids (DUFAs), particularly lineolic acid (18:2w6), a precursorto essential fatty acids. Diporeia from Cayuga Lake contained higher(32.6%) total polyunsaturated fatty acids (PUFAs) than those fromOwasco Lake (24.7%; t-test, p=0.051). The essential fatty acids eico-sapentaeonic acid (EPA, 20:5w3) and docosahexaenoic acid (DHA,22:6w3) were the two most prominent PUFAs in all Diporeia thatwe analyzed. Overall, Diporeia in Owasco Lake had a higher propor-tion of MUFAs (t-test, p=0.0003), whereas populations in CayugaLake contained higher proportions of PUFAs (t-test, p=0.051).
Principal components analysis identified two axes that explained64.4% of the variance (Table 4). The first axis explained 43.2% of the

Fig. 7. Size distribution of quagga mussels in Cayuga Lake at 80 m depth on September25, 2008. The y-axis represents individuals per petite Ponar (0.023 m2). The x-axis islength of individual mussels in cm.
Table 4Major axes from principal components analysis of arcsine squareroot transformed fattyacid data from Diporeia of Cayuga and Owasco Lakes. Positive and negative scores de-note association of each fatty acid with the specific axis. Percentage of variation in thedata explained by each axis noted at the bottom of the table.
Axis 1 Axis 2 Axis 3
14:0 0.254 −0.042 −0.403'16:0 0.336 −0.251 0.041'18:0 −0.311 −0.302 0.10814:1 0.083 0.237 0.475'16:1w7 0.396 0.096 0.073'16:1w9 0.014 −0.241 0.32218:1w7 0.021 −0.514 −0.26018:1w9 −0.140 0.419 0.21618:2w6 −0.186 −0.181 0.39418:3w3 −0.046 −0.363 0.39618:4w3 0.352 0.154 0.10620:4w6 ARA −0.390 0.019 −0.16020:5w3 EPA −0.361 −0.039 −0.09022:6w3 DHA −0.315 0.302 −0.139
Percent 43.2 21.2 16.5Variance 80.8Explained
232 J.M. Watkins et al. / Journal of Great Lakes Research 38 (2012) 226–235
variability in fatty acid composition and was characterized by a highproportion of 16:1w7 (palmitoleic acid) and a low relative contribu-tion of essential PUFAs. The second axis (explaining 21.2% of variabil-ity) is characterized by a high proportion of the PUFA DHA and lowrelative contribution of 18:1w7 (vaccenic acid) and 18:3w3. OwascoDiporeia cluster with high values for both axis 1 and 2, while CayugaDiporeia have a broad mixture of the two axes (Fig. 8).
Substrate analysis
Sediments at our Myer's Point sites in Cayuga Lake were rich incoarse (>500 μm) terrestrial organic matter, averaging 1452 g dryweight (DW)/m2 at 40 m and 862 g DW/m2 at 80 m during2008–2009. In terms of fatty acid content, the coarsematerial in Cayugasediments had a lipid content of 1.05 mg/g and was rich in the PUFAsEPA and DHA (Table 5). In comparison, Owasco substrates had littlecoarse organic matter other than macroinvertebrates and had a lowoverall lipid content (0.05 mg/g). Palmitoleic acid (16:1w7) was 7.5%of the overall lipids in Owasco sediments. Note that only one substratesample was analyzed for both lakes.
Table 3Fatty acid composition of Diporeia from Cayuga and Owasco Lakes in terms of percent-age of total lipid composition. Eight individual adult Diporeia from each lake were an-alyzed. Percentage data were arcsine squareroot transformed prior to significance test(exact p-values reported for alpha=0.05). Significantly different proportions aremarked by an asterisk.
Cayuga Owasco p-Value
Specific FAs14:0 0.1 1.1 0.1090'16:0 14.2 17.0 0.2160'18:0 3.8 1.1 0.0001*14:1 0.0 0.8 0.0642'16:1w7 7.0 22.5 0.0001*'16:1w9 0.5 0.4 0.528018:1w7 8.8 1.6 0.0260*18:1w9 18.0 23.9 0.425018:2w6 6.4 4.4 0.065018:3w3 2.6 2.2 0.138018:4w3 1.5 4.1 0.0001*20:4w6 ARA 4.6 1.7 0.0019*20:5w3 EPA 15.5 10.8 0.0016*22:6w3 DHA 7.4 5.7 0.3530
Lipid ClassesSAFA 18.5 20.3 0.4630MUFA 41.7 50.4 0.0003*DUFA 6.4 4.4 0.0660PUFA 32.6 24.7 0.0510
Discussion
Quagga mussels are replacing zebra mussels in both the GreatLakes and in their home range (Karatayev et al., 1998; Nalepa et al.,2009; Watkins et al., 2007). We show that this is also occurring inthe Finger Lakes of central New York. Both species feed on phyto-plankton, but the lower respiration rates of quagga mussels providethem with a competitive advantage when food resources are low(Baldwin et al., 2002; Stoeckmann, 2003). In Seneca and CayugaLakes, the two Finger Lakes that have had quagga mussels since1994, quagga mussels are abundant as deep as 60 m. This expansionof quagga mussels to deep habitats mirrors the trend seen in lakesOntario, Michigan, and Huron (Nalepa et al., 2009; Watkins et al.,2007). Quagga mussel expansion to deep habitats in many lakes ofNorth America reflects a tolerance of low water temperatures andsoft substrates.
The initial decline of the amphipod Diporeia in many of the GreatLakes occurred during the colonization of shallow habitats by bothdreissenid species (Dermott, 2001; Dermott and Kerec, 1997; Nalepaet al., 1998, 2003). However, the expansion of quagga mussels to soft
Fig. 8. Individual Diporeia from Cayuga (n=8) and Owasco (n=8) lakes plotted interms of their fatty acid composition described by two principal components axesexplaining 64% of the variability of arcsine squareroot transformed percentage data.Cayuga Diporeia are represented by filled circles and Owasco Diporeia are representedby open circles.

Table 5Fatty acid composition of substrate from Cayuga and Owasco Lakes in terms of percent-age of total lipid composition. Note that this is based on only one sample collected atthe same time as the Diporeia. Fatty acids were extracted from 344 mg of Owasco sur-ficial sediment and 69 mg of Cayuga coarse organic material.
Cayuga Owasco
Specific FAs14:0 0.6 0'16:0 19.0 16.0'18:0 13.0 25.914:1 0.0 0.8'16:1w7 2.8 7.5'16:1w9 10.1 8.818:1w7 3.7 4.318:1w9 5.7 7.518:2w6 3.6 2.218:3w3 5.1 4.418:4w3 1.3 2.320:4w6 ARA 4.8 5.020:5w3 EPA 17.2 4.222:6w3 DHA 13.0 11.8
Lipid classesSAFA 32.6 42.0MUFA 22.3 28.1DUFA 3.6 2.2PUFA 41.5 27.7
233J.M. Watkins et al. / Journal of Great Lakes Research 38 (2012) 226–235
substrates in deep habitats has considerably increased the areal cover-age of dreissenid beds and the overall filtering impact of dreissenidson phytoplankton populations (Vanderploeg et al., 2010). Therefore,the effect of quaggamussels onDiporeia is likely stronger than the effectof zebra mussels alone. The ranges of quagga mussels and Diporeia alsodirectly overlap.
Despite colonization by dreissenid mussels, populations of theamphipod Diporeia persist in the six deepest Finger Lakes. AlthoughOwasco Lake has only shallow zebra mussels and Keuka Lake has rel-atively low levels of quagga mussels, Diporeia was found in directcontact with quagga mussels at levels >1000/m2 for the four otherlakes. In contrast, Diporeia populations at intermediate depths in theLaurentian Great Lakes declined at a time when levels of dreissenidswere b1000/m2 (Nalepa et al., 2009). This difference is not likelydue to differences among Diporeia populations, as Finger Lake Dipor-eia are genetically similar to those of Lake Ontario, Michigan andHuron (although distinct from Diporeia of Lake Superior, Pilgrim etal., 2009). Therefore, other mechanisms are likely responsible forDiporeia populations in the New York Finger Lakes being able to toler-ate higher (up to 10,000/m2) levels of dreissenids than Great LakesDiporeia.
Lake average Diporeia abundance could be predicted by trophicstate parameters better than dreissenid abundance or lake morpholo-gy in the Finger Lakes. For example, the low abundance (b1000/m2)of Diporeia in Skaneateles Lake is consistent with the very low pro-ductivity of the lake. Skaneateles Lake has the lowest levels of average(May–October in the epilimnion) total phosphorus (TP, 3.7 μg/L) andchlorophyll (chl a, 0.7 μg/L) and the deepest secchi depth (7.6 m) ofall eleven Finger Lakes (Halfman and O'Neill, 2009, Table 1). In thelate 1970s the lake had higher values for average TP (6.1 μg/L) andchl a (1.95 μg/L), and a shallower secchi depth (6.6 m) (Callinan,2001). These parameters reflect a long-term oligotrophic state com-parable to the current trophic state of the offshore of the GreatLakes excluding Lake Erie. Diporeia abundance decreased with depthin both our samplings of Skaneateles Lake in 2006 and 2010, consis-tent with the decrease in Diporeia abundance with depth in Lake Su-perior (Auer et al., 2009).
Diporeia abundance was also relatively low in Canandaigua andKeuka Lakes, but did not decrease with depth. Canandaigua Lake isslightly more productive than Skaneateles Lake, with higher but stilllow values of TP (7.8 μg/L) and chl a (1.7 μg/L) and high transparency(6.5 m secchi depth, Halfman and O'Neill, 2009). The abundance of
Diporeia in Canandaigua Lake has been stable for nearly a century —
our measurements from 2010 (average 1006/m2) are comparable tothat measured in 1918 (911/m2, Birge and Juday, 1921), 1927(1066/m2, Eaton unpublished), and 1974 (1193/m2, Eaton et al.,1975). Keuka Lake is similarly oligotrophic with low TP (6.3 μg/L)and chl a (2.0 μg/L), and high transparency (5.3 m secchi depth)(Halfman and O'Neill, 2009).
The higher abundance (2000/m2) of Diporeia in Cayuga, Seneca,and Owasco lakes is consistent with the more mesotrophic state ofthe three lakes and associated higher algal production. The threelakes have higher average levels of chl a (3.2, 3.4, and 2.8 μg/L) aswell as lower transparency (4.1, 4.7, and 4.1 m secchi depth) thanSkaneateles, Canandaigua, and Keuka Lakes (Halfman and O'Neill,2009). A similar relationship of Diporeia abundance and productivityhas been observed within a single lake (New York State's Lake George)along a productivity gradient from the mesotrophic south end to themore oligotrophic north end (Siegfried, 1985). Prior to the recent de-cline, Diporeia abundance in the Great Lakes was also correlated withoverall productivity with Lake Superior having the fewest Diporeiaand LakeMichigan and LakeOntario havingmore abundant populations(Cook and Johnson, 1974). The relationship does not extend to eutro-phic lakes. Diporeia population declines in Round Lake (Ontario), LakeMemphremagog (Quebec) and Lakes Champlain and Winnipeg havebeen attributed to eutrophication rather than dreissenid introduction(Dermott et al., 2005).
Although persistence of Diporeia in most of the Finger Lakes couldbe due to low abundances of deep-water quagga mussels, Diporeia inCayuga and Seneca Lakes coexist with quaggamussels at abundancesof 1000–5000/m2. Decreased Diporeia populations were only seen atdreissenid levels exceeding 10,000/m2. This finding suggests thatdreissenids have less of an impact on Diporeia in the Finger Lakesthan the Great Lakes, perhaps due to higher levels of food or theavailability of alternative resources. In support of an alternativefood source, our fatty acid analysis indicates that Diporeia in CayugaLake are feeding to a greater extent on food resources other thandiatoms.
Diporeia in Owasco Lake were in better condition with a higherlipid content (51.6% relative to 14.9%) than those in Cayuga Lake.The higher lipid content of Diporeia in Owasco Lake suggests thatfood availability is high and perhaps benefits from lower lake-widelevels of dreissenids. The lipid content of Diporeia in Cayuga is lowerthan in Owasco Lake and healthy Diporeia populations from LakeMichigan collected in 1989 (21.7%, Cavaletto et al., 1996), but arecomparable to that of Diporeia from Charleston Lake, Ontario in2006 (13.7%, Kainz et al., 2010). Although lower than the 33.1% lipidcontent measured from populations in Cayuga Lake in June, 1964(Green, 1971), our 2008 size distribution data indicate that the popu-lation of Diporeia is still successfully reproducing. The growth rate ofDiporeia in 2008 calculated from our cohort analysis is also similarto the growth rate measured in 1964 of 0.33 mm per month (Fig. 6,Green, 1965). High lipid content does not necessarily mean high sur-vival for Diporeia. For example, Diporeia populations in Lake Michigandeclined in the late 1990s even as lipid content of adults increasedfrom 23.9% to 34.7% (Nalepa et al., 2006). In 30 day feeding experi-ments, fasting Diporeia had higher survival (70%) than Diporeia fed di-atoms (60%) despite having nearly half of the lipid content at the endof the feeding trial (Kainz et al., 2010).
High proportions of the essential fatty acids, particularly ARA, EPAand DHA, indicate that Diporeia in Owasco and Cayuga Lakes havehigh quality diets and an abundant supply of these fatty acids intheir diet. Diporeia in Cayuga Lake contained higher amounts of ARAthan those in Owasco Lake, a marker fatty acid that is preferentiallyretained by Diporeia even when its concentration in diet is low(Kainz et al., 2010). We measured higher amounts of ARA thanthose measured in Diporeia that were experimentally fed diatoms(Kainz et al., 2010) indicating that populations in Cayuga Lake have

234 J.M. Watkins et al. / Journal of Great Lakes Research 38 (2012) 226–235
access to high amounts of this required fatty acid. Levels of other fattyacids represent dietary tracers. Diporeia in Owasco Lake were partic-ularly rich in the MUFA palmitoleic acid (16:1w7), an indicator of di-atom consumption (Goedkoop et al., 2000). Clearly, the zebra musselsin Owasco Lake are not eliminating diatoms as a food source forDiporeia. The lower content of this diatom tracer fatty acid in Diporeiain Cayuga Lake suggests a greater reliance on detritus. Diatoms arealso a primary source of the PUFA EPA, so it is interesting that despitehaving low levels of the diatom tracer, the proportion of EPA in Dipor-eia in Cayuga Lake (15.5%) was significantly higher than that inOwasco Lake (10.8%) (t-test, p=0.0016). Coarse terrestrial organicmatter from Cayuga sediments was also high in EPA but low in the di-atom tracer (Table 5). Note that Owasco Diporeia had a 50% greaterabsolute content of EPA (mg/g DW) than Cayuga Diporeia. Both popu-lations had a DHA/EPA ratio near 0.50, where ratios much less thanone indicate a diatom-based diet (Budge and Parrish, 1998).
Other potential food web tracers were harder to interpret. Fattyacids with odd numbers of carbon in their chains (e.g. 17:0) and vac-cenic acid (18:1w7) have been used as markers for bacterial activity(Goedkoop et al., 2000; Parrish et al., 2000). No odd number carbonchains were detected, but Cayuga Diporeia had a significantly higherproportion of vaccenic acid than Owasco Diporeia (p=0.026). Levelsof the fatty acids 18:2w6 and 18:3w3, indicators of terrestrial plantmatter (Parrish et al., 2000), were higher for Cayuga Diporeia, butnot significantly (p=0.065 and 0.138). Even though the Cayuga sub-strate sample was visibly rich in terrestrial leaf litter, terrestrial indi-cators were small components of substrate fatty acids (Table 5) ineither lake and therefore not effective as dietary tracers. Therefore,the diatom tracer 16:1w7 and the bacterial tracer 18:1w7 emerge asthe most consistent indicators of different diets for Diporeia in thetwo lakes. The first two axes of the principal components analysisrepresent contributions from these tracers, with axis 1 associatedwith high proportion of the diatom tracer and axis 2 associatedwith a low proportion of the bacterial tracer. Individual Diporeia inOwasco Lake had a consistently high proportion of the diatom tracer,while individual Diporeia from Cayuga Lake indicated a mixture of di-atom and bacterial food sources.
The coexistence of Diporeia and dreissenid mussels suggests thatDiporeia in the Finger Lakes are much more resistant to dreissenid im-pacts than amphipods in the Great Lakes. When diatoms are availablein deeper water, as in Owasco Lake with only nearshore zebra mussels,Diporeia feed on settled diatoms and are in excellent condition. Howev-er, even when quagga mussels are abundant, Diporeia in the FingerLakes have access to an alternative food source in detritus and bacteria.These food sources should not be affected by mussel grazing.
Conclusions
Diporeia populations of the New York Finger Lakes in 2010 haveshown few negative impacts from the establishment of dreissenidmus-sels in the 1990s. Particularly interesting is Diporeia's resilience despiteliving in direct contactwith abundant (>1000/m2) quaggamussels thatwere present in four of the six lakes (Cayuga, Seneca, Skaneateles, andCanandaigua). Few examples of coexistence of the two organismsexist in the Great Lakes where Diporeia populations continue to decline.Potential reasons for the persistence of Diporeia in the Finger Lakes arethat alternative food sources exist in the Finger Lakes such as bacteriaassociated with terrestrial organic matter and/or that dreissenid filterfeeding in the deep Finger Lakes has not reduced the supply of diatomsto Diporeia. It is also possible that dreissenids colonizing the FingerLakes have not transported a pathogen lethal to Diporeia.
Acknowledgments
This researchwas funded by support fromEPA's Great LakesNationalProgram Office, New York State and the Finger Lakes Institute of Hobart
andWilliamSmith College for study of Owasco Lake, andCornell Univer-sity's IGERT program for Biogeochemistry and Environmental Biocom-plexity. We acknowledge the assistance of Ben Estes of SUNY-ESF inSyracuse, NY in the analysis of fatty acids. Dr. JohnHalfman of the FingerLakes Institute andDr. DawnDittman of USGS Tunison Aquatic ResearchCenter in Cortland, NY coordinated sampling of Owasco Lake in 2007.Undergraduate interns Thomas Stewart (Syracuse U.), Mark Leopold(Cornell U.), Dawn Moses (Cornell U.), and Dana Seidel (Cornell U.)provided assistance in the field and the laboratory. Dr. Tomas Hook,Dr. Marisol Sepulveda, Dan Ryan, Suman Maity, and Sunnie McCalla ofPurdue University collaborated in research of Diporeia in the FingerLakes. Dr. Erik Pilgrim of EPA's Ecological Exposure Research Divisionin Cincinnati, Ohio provided genetic information on Diporeia in theGreat Lakes and Finger Lakes. This is contribution #287 of the CornellBiological Field Station.
References
Auer, N.A., Cannon, B.A., Auer, M.T., 2009. Life history, distribution, and productionof Diporeia near the Keweenaw Peninsula Lake Superior. J. Great Lakes Res. 35,579–590.
Baldwin, B.S., Mayer, M.S., Dayton, J., Pau, N., Mendilla, J., Sullivan, M., Moore, A., Ma, A.,Mills, E.L., 2002. Comparative growth and feeding in zebra and quagga mussels(Dreissena polymorpha and Dreissena bugensis): implications for North Americanlakes. Can. J. Fish. Aquat. Sci. 59, 680–694.
Birge, E.A., Juday, C., 1921. Further limnological observations on the Finger Lakes ofNew York. Bull. U.S. Bur. Fish. 37, 210–252.
Budge, S.M., Parrish, C.C., 1998. Lipid biogeochemistry of plankton, settling matter andsediments in Trinity Bay, Newfoundland. II. Fatty acids. Org. Geochem. 29,1547–1559.
Callinan, C.W., 2001. Water quality study of the Finger Lakes. Publication of the NewYork Department of Conservation, Division of Water. 73 pp.
Cavaletto, J.F., Nalepa, T.F., Dermott, R., Gardner, W.S., Quigley, M.A., Lang, G.A., 1996.Seasonal variation of lipid composition, weight, and length in juvenile Diporeiaspp. (Amphipoda) from lakes Michigan and Ontario. Can. J. Fish. Aquat. Sci. 53,2044–2051.
Cook, D.G., Johnson, M.G., 1974. Benthic macroinvertebrates of the St. Lawrence GreatLakes. J. Fish. Res. Board Can. 31, 763–782.
Dermott, R., 2001. Sudden disappearance of the amphipod Diporeia from eastern LakeOntario. J. Great Lakes Res. 27, 423–433.
Dermott, R., Geminiuc, M., 2003. Changes in the benthic fauna of Lake Ontario1990–1995, with local trends after 1981. In: Munawar, M. (Ed.), State of LakeOntario (SOLO)-Past, Present, and Future. Ecovision World Monograph Series.Backhuys Publishers, Leiden, The Netherlands, pp. 323–345.
Dermott, R., Kerec, D., 1997. Changes to the deep-water benthos of eastern Lake Eriesince the invasion of Dreissena: 1979 to 1993. Can. J. Fish. Aquat. Sci. 54, 922–930.
Dermott, R., Bonnell, R., Jarvis, P., 2005. Population status of the amphipod Diporeia ineastern North American lakes with or without Dreissena. Verh. Int. Ver. Limnol. 29,880–886.
Dermott, R., Bonnell, R., Jarvis, P., 2006. Changes in abundance of deep water amphipodDiporeia (Pontoporeiidae) in eastern North American lakes with or without Dreis-sena mussels. Can. Tech. Rep. Fish. Aquat. Sci. 2636, 88p.
Eaton, S.W., Eaton, E.H., Weglinski, P., 1975. Benthos of the deep basin of CanandaiguaLake. Sci. Stud. 31, 27–33.
Goedkoop, W., Sonesten, L., Ahlgren, A., Boberg, M., 2000. Fatty acids in profundal ben-thic invertebrates and their major food resources in Lake Erken, Sweden: seasonalvariation and trophic indications. Can. J. Fish. Aquat. Sci. 57, 2267–2279.
Green, R.H. 1965. The population ecology of the glacial relict amphipod Pontoporeiaaffinis (Lindstrom) in Cayuga Lake, New York. PhD Thesis, Cornell University, Ithaca,NY.
Green, R.H., 1971. Lipid and caloric contents of the relict amphipod Pontoporeia affinisin Cayuga Lake, New York. J. Fish. Res. Board Can. 28, 776–777.
Halfman, J.D., O'Neill, K., 2009. Water quality of the Finger Lakes, New York:2005–2008. Finger Lakes Institute, Geneva, NY.
Hammers, B., Richardson, D., Pearsall, W., 2007. Ecological effects of zebra and quaggamussel (D. polymorpha and D. bugensis) invasion in the Western Finger Lakes Area.Final Report 1995–2004. New York Department of Conservation, Division of Fish,Wildlife and Marine Resources, Region 8. 551 pp.
Henson, E.B. 1954. The profundal bottom fauna of Cayuga Lake. PhD Thesis, CornellUniversity, Ithaca, NY.
Kainz, M.J., Johannsson, O.E., Arts, M.E., 2010. Diet effects on lipid composition, somaticgrowth potential, and survival of benthic amphipod Diporeia spp. J. Great LakesRes. 36, 351–356.
Karatayev, A.Y., Burlakova, L.E., Padilla, D.K., 1998. Physical factors that limit the distribu-tion and abundance of Dreissena polymorpha (Pall.). J. Shellfish. Res. 17, 1219–1235.
Lozano, S.J., Scharold, J.V., Nalepa, T.F., 2001. Recent declines in benthic macroinverte-brate densities in Lake Ontario. Can. J. Fish. Aquat. Sci. 58, 518–529.
McCune, B., Mefford, M.J., 1999. PC-ORD. Multivariate Analysis of Ecological Data,Version 4. MjM Software Design, Gleneden Beach, Oregon, USA.

235J.M. Watkins et al. / Journal of Great Lakes Research 38 (2012) 226–235
Nalepa, T.F., Hartson, D.J., Fanslow, D.L., Lang, G.A., Lozano, S.J., 1998. Declines in ben-thic macroinvertebrate populations in southern Lake Michigan, 1980–1993. Can.J. Fish. Aquat. Sci. 55, 2402–2413.
Nalepa, T.F., Fanslow, D.L., Lansing, M.B., Lang, G.A., 2003. Trends in the benthic macro-invertebrate community of Saginaw Bay, Lake Huron, 1987 to 1996: responses tophosphorus abatement and the zebra mussel Dreissena polymorpha. J. GreatLakes Res. 29, 14–33.
Nalepa, T.F., Fanslow, D.L., Foley, A.J., Lang, G.A., Eadie, B.J., Quigley, M.A., 2006. Continueddisappearance of the benthic amphipod Diporeia spp. in Lake Michigan: is thereevidence for food limitation? Can. J. Fish. Aquat. Sci. 63, 872–890.
Nalepa, T.F., Fanslow, D.L., Lang, G.A., 2009. Transformation of the offshore benthic com-munity in Lake Michigan: recent shift from the native amphipod Diporeia spp. tothe invasive mussel Dreissena rostriformis bugensis. Freshw. Biol. 54, 466–479.
Neilson, M.A., Painter, D.S., Warren, G., Hites, R.A., Basu, I., Weseloh, D.V., Whittle, D.M.,Christie, G., Barbiero, R., Tuchman, M., Johannsson, O.E., Nalepa, T.F., Edsall, T.A.,Fleischer, G., Bronte, C., Smith, S.B., Baumann, P.C., 2003. Ecological monitoringfor assessing the state of the nearshore and open waters of the Great Lakes. Envi-ron. Monit. Assess. 88, 103–117.
Owens, R.W., Dittman, D.E., 2003. Shifts in the diet of slimy sculpin (Cottus cognatus)and lake whitefish (Coregonus clupeaformis) in Lake Ontario following the collapseof the burrowing amphipod Diporeia. Aquat. Ecosyst. Heal. Manag. 6, 311–323.
Parrish, C.C., Abrajano, T.A., Budge, S.M., Helleur, R.J., Hudson, E.D., Pulchan, K., Ramos,C., 2000. Chapter 8 Lipid and phenolic biomarkers in marine ecosystems: analysisand applications. The Handbook of Environmental Chemistry, Vol. 5, Part D. MarineChemistry. Springer-Verlag, Berlin, p. 201.
Pilgrim, E.M., Scharold, J.V., Darling, J.A., Kelly, J.R., 2009. Genetic structure of the benthicamphipod Diporeia (Amphipoda: Pontoporeiidae) and its relationship to abundancein Lake Superior. Can. J. Fish. Aquat. Sci. 66, 1318–1327.
Quigley, M.A., Lang, G.A., 1989. Measurement of amphipod body length using a digitizer.Hydrobiologia 171, 255–258.
Ryder, R.A., Edwards, C.J. (Eds.), 1985. A conceptual approach for the application ofbiological indicators of ecosystem quality in the Great Lakes basin. Report to theGreat Lakes Science Advisory Board. International Joint Commission and Great LakesFishery Commission, Windsor, Ont.
Schaffner, W.R., Oglesby, R.T., 1978. Limnology of eight Finger Lakes: Hemlock, Canadice,Honeoye, Keuka, Seneca, Owasco, Skaneateles, and Otisco. In: Bloomfield, J.A. (Ed.),Lakes of New York State Volume 1 — Ecology of the Finger Lakes. Academic Press,London, pp. 313–470.
Scharold, J.V., Lozano, S.J., Corry, T.D., 2004. Status of the amphipod Diporeia spp. inLake Superior, 1994–2000. J. Great Lakes Res. 30 (Supp. 1), 360–368.
Siegfried, C.A., 1985. Life history, population dynamics, and production of Pontoporeiahoyi (Crustacea, Amphipoda) in relation to the trophic gradient of Lake George,New York. Hydrobiologia 122, 175–180.
Sly, P.G., Christie, W.J., 1992. Factors influencing densities and distributions of Ponto-poreia hoyi in Lake Ontario. Hydrobiologia 235 (236), 321–352.
Snedecor, G.W., Cochran, W.G., 1989. Statistical Methods, Eighth edition. Iowa StateUniversity Press, Ames.
Sokal, R.R., Rohlf, F.J., 1981. Biometrics, Second edition. Freeman, San Francisco.Stoeckmann, A., 2003. Physiological energetics of Lake Erie dreissenid mussels: a basis
for the displacement of Dreissena polymorpha by Dreissena bugensis. Can. J. Fish.Aquat. Sci. 60, 126–134.
Teece, M.A., Estes, B., Gelsleichter, A., Lirman, D., 2011. Heterotrophic and autotrophicassimilation of fatty acids by Scleractinian corals. Limnol. Oceanogr. 56, 1244–1255.
U.S. Environmental Protection Agency, 1974. An approach to a relative trophic indexsystem for classifying lakes and reservoirs. National Eutrophication Survey. WorkingPaper No. 24.
U.S. Geological Survey, a. Nonindigenous Aquatic Species (NAS) Database http://nas.er.usgs.gov/.
Vanderploeg, H.A., Liebig, J.R., Nalepa, T.F., Fahenstiel, G.L., Pothoven, S.A., 2010. Dreis-sena and the disappearance of the spring phytoplankton bloom in Lake Michigan.J. Great Lakes Res. 36, 50–59.
Watkins, J.M., Dermott, R., Lozano, S.J., Mills, E.L., Rudstam, L.G., Scharold, J.V., 2007.Evidence for remote effects of dreissenid mussels on the amphipod Diporeia:analysis of Lake Ontario benthic surveys, 1972–2003. J. Great Lakes Res. 33,642–657.
Whittier, T.R., Ringold, P.J., Herlihy, A.T., Pierson, S.M., 2008. A calcium-based invasionrisk assessment for zebra and quagga mussels (Dreissena spp.). Front. Ecol. Envi-ron. 6, 180–184.