external morphology of limb development in the amphipod orchestia cavimana (crustacea, malacostraca,...
TRANSCRIPT

ORIGINAL ARTICLE
Petra Ungerer Æ Carsten Wolff
External morphology of limb development in the amphipod Orchestiacavimana (Crustacea, Malacostraca, Peracarida)
Received: 23 April 2004 / Accepted: 1 December 2004 / Published online: 6 April 2005� Springer-Verlag 2005
Abstract The external morphology of limb developmentin Orchestia cavimana is examined by scanning electronmicroscopy and fluorescence staining from the appear-ance of the first limb buds until hatching. As otheramphipods, O. cavimana undergoes direct developmentand the degree of segmental differentiation shows a moreor less continual decrease in anteroposterior direction.Limbs form ventrally as small buds, which elongate anddivide into podomeres early in development. This earlysubdivision largely corresponds with the limb segmen-tation of the hatchling. When the post-naupliar limbsstart to develop, the germ band begins to split into twohalves along the midline, so that the trunk limbs tran-siently occupy a very dorsolateral position. After thegerm band has closed again, the differentiation into thecharacteristic amphipodan tagmata (cephalothorax,pereon, pleon) takes place and the limb podomeres losetheir round-shape. The late embryo is covered by a so-called intermediate cuticle, which is formed after anembryonic moult and shed after hatching. The earlydevelopment of O. cavimana reveals the Anlage of avestigial seventh pleonic segment that is assumed tobelong to the ground pattern of malacostracans, but isretained as a free, limbless segment only in adult Lep-tostraca. A transient subdivision of the proximal seg-ment of the pleopods suggests the occurrence of a coxaand a basis in these limbs. The mandible attains its up-right, adult position via a characteristic bending processthat is strikingly similar to that in Archaeognatha (In-secta).
Keywords Scanning electron microscopy ÆSEM Æ Mouthparts Æ Direct development ÆEmbryology Æ Ontogeny
Introduction
The amphipod Orchestia cavimana Heller, 1865,exhibits a complex invariant cell division pattern duringthe formation and differentiation of the post-naupliargerm band that is typical for malacostracan develop-ment (Dohle and Scholtz 1988, 1997; Scholtz 1990;Scholtz and Dohle 1996; Dohle et al. 2004). At a stageof about four hundred cells, transverse rows of cells arearranged in a grid-like pattern in the ectoderm of thepost-naupliar germ band. They subsequently undergostereotyped cell division patterns that allow us to tracethe fate of individual cells. Because of its relatively flatearly germ band, that is restricted to the ventral side ofthe egg, Orchestia is a suitable object for cell lineagestudies (Dohle and Scholtz 1988; Scholtz 2000) thatrange from cleavage, gastrulation, and germ disc for-mation (Scholtz and Wolff 2002; Wolff and Scholtz2002) to the formation of segmental structures (char-acterisation of the median neuroblast: Gerberding andScholtz 1999, 2001). Detailed analyses of the geneexpression patterns of engrailed (e.g. Scholtz and Dohle1996) and Distal-less (Scholtz et al. 1998; Hejnol andScholtz 2004) were undertaken in the germ band ofOrchestia, while limb development and aspects ofneurogenesis are currently being investigated on thecellular level. To provide a basis for this research, adetailed description of the morphology of the devel-opmental stages through which the embryo passes isindispensable. Besides its usefulness as a basis forexperimental studies, an investigation of the morpho-genesis is also well placed to reveal, and further discuss,morphological aspects. Using scanning electronmicroscopy and Hoechst fluorescence staining, we de-scribe the external features of the head and trunk limb
P. Ungerer Æ C. Wolff (&)Humboldt Universitat zu Berlin, Vergleichende Zoologie,Institut fur Biologie, Philippstr. 13, 10115 Berlin, GermanyE-mail: [email protected].: +49-30-20936006Fax: +49-30-20936002E-mail: [email protected].: +49-30-20936007Fax: +49-30-20936002
Zoomorphology (2005) 124: 89–99DOI 10.1007/s00435-005-0114-2

ontogeny of O. cavimana from the formation of thelimb buds until hatching.
Materials and methods
Specimens of Orchestia cavimana (Crustacea, Malacost-raca, Peracarida, Amphipoda, Talitridae) were collectedat the lakefront of the Tegeler See, Berlin, and cultured inthe laboratory. The eggs and embryos (10–20 per female,300–600 lm in diameter) were isolated from the ventralbrood pouch (marsupium) of the females using a glasspipette. As is typical for amphipods, O. cavimana under-goes direct development (Scholtz 2000) and, therefore,passes through all the developmental stages within the eggmembranes. Prior to fixation, the egg membranes wereremoved with insect pins and tweezers.
For scanning electron microscopy (SEM), the em-bryos were fixed in Bouin’s fluid (75% saturated aque-ous picric acid solution, 20% saturated formaldehyde,5% glacial acetic acid) between 10 min and 60 min,dehydrated in a graded ethyl alcohol series and criticalpoint dried with a CPD BALTEC 030 following stan-dard procedures. The mounted specimens were sputteredwith gold using a SCD BALTEC 005, and a Leo 1450VPwas used to take SEM photographs.
The material for nuclear fluorescent dye staining wasfixed in 4% formaldehyde in PBS (0,01 mol/l Na2HPO4;0,01 mol/l NaH2PO4; 0,15 mol/l NaCl in aqua dest.;pH 7,4) washed for 10 minutes in PBS and treated witha 0,01% Hoechst stain solution (Bisbenzimide H 33258)for 10 minutes. The dissected embryos were washed in
PBS for 10 minutes and mounted in glycerol on glassslides. They were analysed and photographed using aZeiss Axiophot fluorescence microscope.
Results
While studying the development of Orchestia cavimana,we found considerable differences in the developmentalrate among the eggs of different broods, and even withinsingle broods. Consequently, the most common way ofstaging, namely staging by age, seemed inappropriate,and we chose to stage the developmental processes byevents.
We describe the morphogenesis of O. cavimana fromthe point when the first transversal rows of ectodermalcells (Fig. 1a) have begun their stereotyped cell divisionpattern, providing the ectoderm for the formation oflimbs, ganglia, etc. Starting in the anterior post-naupliarregion of the germ band, each of the ectodermal rows,which are composed of an unpaired midline and pairedlateral cells (Fig. 1a–c), undergo two regular mitoticwaves with longitudinally orientated spindle axes lead-ing to an elongation of the germ band. The orientationof the following differential cleavages is different foreach cell, causing the grid-like pattern to be dissolvedalong with the beginning neurogenesis and limb devel-opment (Fig. 1c). In the naupliar region, such a stereo-typed cell lineage pattern cannot be identified (Fig. 1a,b) (see Scholtz and Dohle 1996).
Limbs form ventrally as small round or elongatebuds. The degree of limb differentiation shows a more
Fig. 1 Ventral view of earlyembryos of Orchestia cavimana.Anterior is up. Midlineindicated by arrows. a Germband stained with a nuclearfluorescent dye (Bisbenzimide)showing the regular grid-likearrangement of ectoderm cellsin the post-naupliar region. Thedotted line marks the borderbetween the naupliar (upper)and the post-naupliar (lower)region. b Formation of the limbbuds in the naupliar region.c Post-naupliar region of thesame embryo as inb. Intersegmental furrowsindicated by white arrow heads,Anlagen of ganglia indicated byblack arrow heads. A1 firstantenna, A2 second antenna,Md mandible, Mgg prospectivemidgut gland, Mx1 firstmaxilla, Mx2 second maxilla,Mxp maxilliped, Pe1-Pe5pereomeres 1–5, yc yolky cells
90

or less continual decrease in anteroposterior direction,except for the two maxillary segments that show slightretardation in development. The two body halves of theembryo do not develop completely synchronously—oneof the two sides is always slightly advanced.
An amphipod body is typically divided into threemajor tagmata. The cephalothorax bears two pairs ofantennae, a mandible, a first and a second maxilla, andthe first thoracopod (called maxilliped). The pereonbears seven pairs of uniramous thoracopods (pereopods),of which the anterior-most two pairs are specialised asgnathopods. The pleon bears six biramous limbs: thepleopods. The first three pairs are differentiated intoswimmerets, while the last three pairs, the uropods, makeup the short urosome, with the telson at its terminal end.
Formation of the naupliar limb buds
The limb buds of the first and second antenna and themandible are the first to develop (Fig. 1b). At this point,the intersegmental furrows and theAnlagen of the gangliaappear close to the midline within the segments of secondmaxilla and maxilliped and in the anterior region of theprospective pereon, while more posteriorly, the regular,grid-like pattern of ectodermal rows is still visible(Fig. 1c). During this stage of development, the dorsalhalf of the embryo consists entirely of a thin layer of large,yolky cells known as extra-embryonic ectoderm (Fig. 1c).
Formation of the post-naupliar limb buds and initialgerm band split
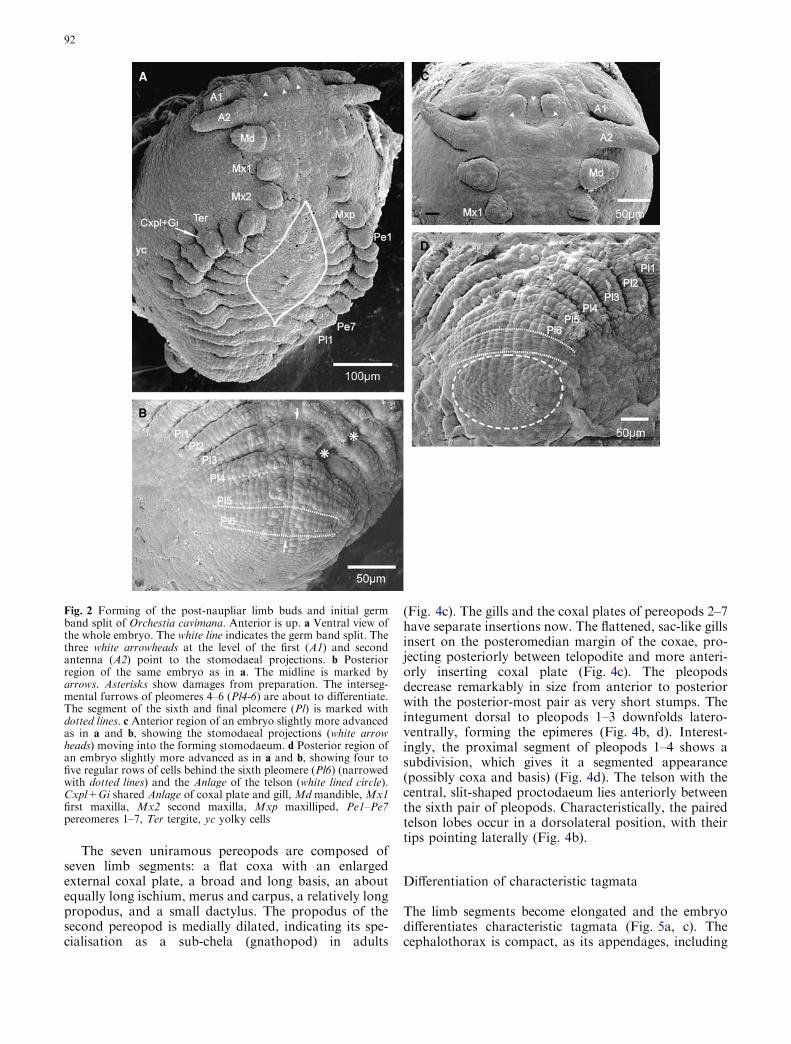
Starting anteriorly with the segment of the maxilliped,the germ band begins to separate into two halves alongthe median line between the right and left Anlagen of theganglia (Fig. 2a) (see also Gerberding and Scholtz 1999).Three stomodaeal projections grow out at the level ofthe first antennal buds. They lie more or less in a hori-zontal line and subsequently move into the stomodaeumas it forms (Fig. 2a, c, and 3a). The undivided buds ofthe first and second antenna are the first to elongate,with their tips projecting laterally. The mandible attainsa characteristic triangular form, while the buds of thefirst and second maxilla and the maxilliped appear assmall, more or less round, undivided lobes (Fig. 2a, c).
The broader buds of the pereopods lying most to theanterior already show proximolaterally the sharedAnlagen of the coxal plates and gills. Also, their pro-spective tergites are visible adjacent to the large dorsalyolky cells (Fig. 2a). The anterior-most pleopod budsstart to develop, while the intersegmental furrows of thefollowing pleomeres are about to differentiate (Fig. 2a,b, d). The Anlage of the telson appears as a broad, moreor less round field of cells, with paired anterior lobes.Posteromedially to the paired lobes, there are a fewsmall cells indicating the point of invagination of theforming proctodaeum (Fig. 2d). At the same time, four
to five regular rows of cells become visible between thesixth pleomere and the Anlage of the telson, similar to a‘normal’ segment Anlage, but without limb buds orprimordia of ganglia (Fig. 2d).
Elongation and initial segmentation of limbs
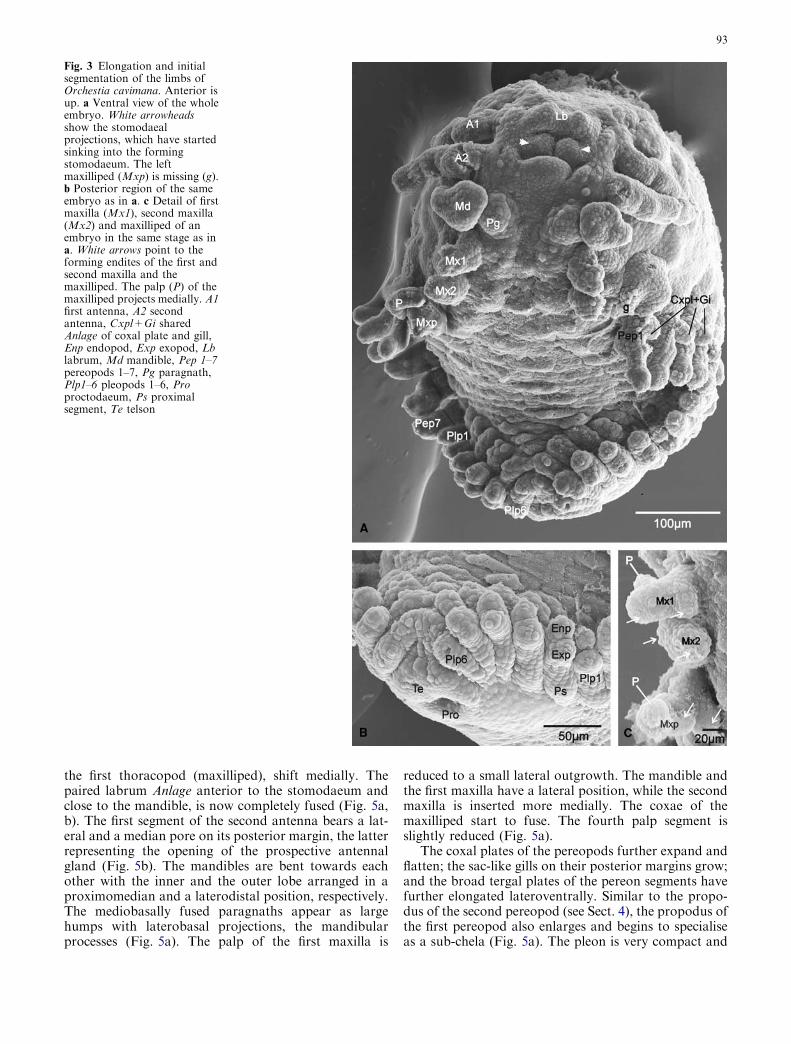
The stomodaeum, with the sunken stomodaeal projec-tions still visible, is forming at the level between the firstand second antenna. The Anlage of the labrum appearsas paired epidermal protuberances in front of the sto-modaeum (Fig. 3a). The first and second antenna be-come elongated further and start to divide into theirprospective segments. The triangular mandible increasesin size and develops a central longitudinal depressionthat gives it a bilobate shape. The paragnaths grow outof the mandibular sternal area, close to the posterome-dian base of the mandible. The first maxilla developsthree lobes: an external palp, a median endite and alateral endite. The second maxilla develops two lobateendites, and the elongated maxilliped is now composedof two basal segments (coxa and basis) that bear enditesand an elongated palp (Fig. 3a, c).
The split of the germ band further advances, so thatthe trunk limbs insert in a very dorsolateral position.Like the antennae, the uniramous pereopods 1–7 elon-gate and start to divide into podomeres. The Anlage ofthe coxal plate in pereopod 1 and the shared Anlagen ofthe coxal plates and gills in pereopods 2–7 are enlarged(Fig. 3a). Six pairs of pleopods have developed, showinga differentiation into a proximal segment and two lobaterami that represent the endo- and exopods (Figs. 3a, b).The telson bears the forming proctodaeum (Fig. 3b).
Closure of the germ band
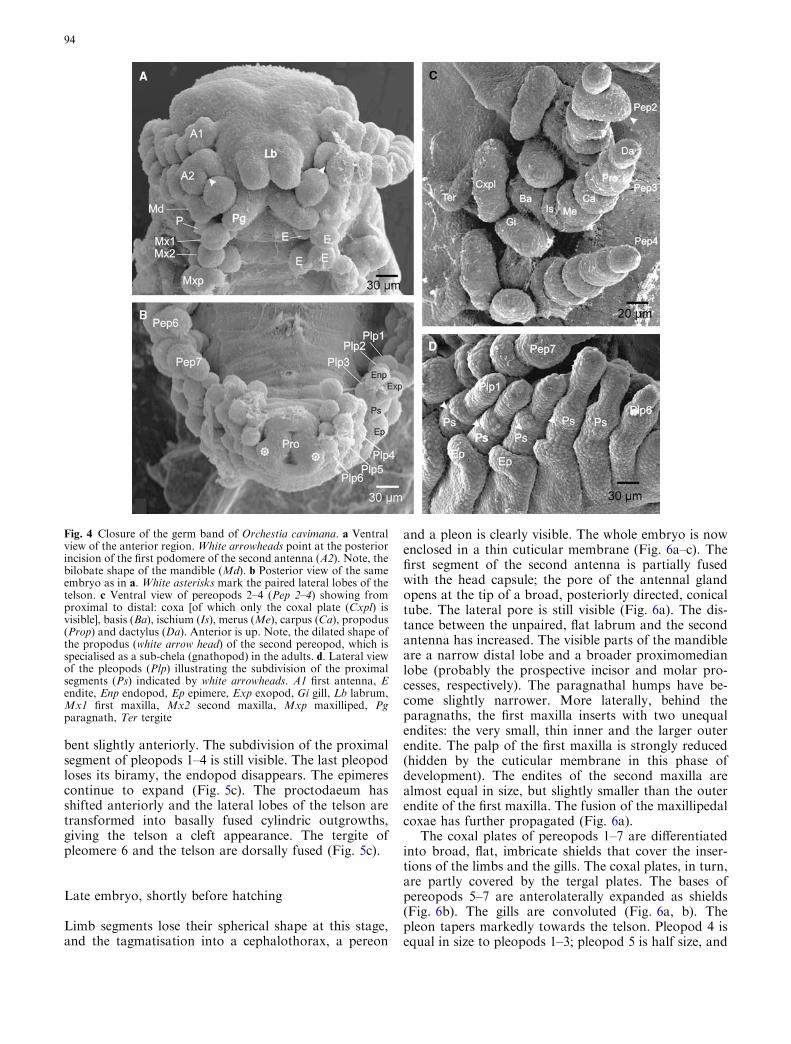
The limb segments attain a rounded shape (Fig. 4a–d).As the germ band halves are about to fuse again, thebody becomes curved with concave side ventral (Fig. 4a,b). In the course of the initial tagmatisation process, theAnlage of the head capsule differentiates in front of theanterolaterally pointing first and second antenna. Thepaired labrum Anlage starts to fuse basally and is di-rected posteriorly, concealing the stomodaeum (Fig. 4a).The first podomeres of the antennae are larger thanthose that follow and that of the second antenna has aconspicuous posterior incision. The incipiently bilobatemandible differentiates into a broad, rounded medianlobe and a lateral lobe. The bases of the elongatedparagnaths join proximomedially, while their tips pro-ject laterally between the mandible and the first maxilla.The two endites of the first maxilla differ significantly insize. The outer endite is large, while the inner, smallendite is about the same size as the external palp. Therounded inner and outer endite of the second maxilla areof about the same size. The endites of coxa and basis ofthe maxilliped have grown, and the elongate palp issubdivided into four segments (Fig. 4a).
91
The seven uniramous pereopods are composed ofseven limb segments: a flat coxa with an enlargedexternal coxal plate, a broad and long basis, an aboutequally long ischium, merus and carpus, a relatively longpropodus, and a small dactylus. The propodus of thesecond pereopod is medially dilated, indicating its spe-cialisation as a sub-chela (gnathopod) in adults
(Fig. 4c). The gills and the coxal plates of pereopods 2–7have separate insertions now. The flattened, sac-like gillsinsert on the posteromedian margin of the coxae, pro-jecting posteriorly between telopodite and more anteri-orly inserting coxal plate (Fig. 4c). The pleopodsdecrease remarkably in size from anterior to posteriorwith the posterior-most pair as very short stumps. Theintegument dorsal to pleopods 1–3 downfolds latero-ventrally, forming the epimeres (Fig. 4b, d). Interest-ingly, the proximal segment of pleopods 1–4 shows asubdivision, which gives it a segmented appearance(possibly coxa and basis) (Fig. 4d). The telson with thecentral, slit-shaped proctodaeum lies anteriorly betweenthe sixth pair of pleopods. Characteristically, the pairedtelson lobes occur in a dorsolateral position, with theirtips pointing laterally (Fig. 4b).
Differentiation of characteristic tagmata
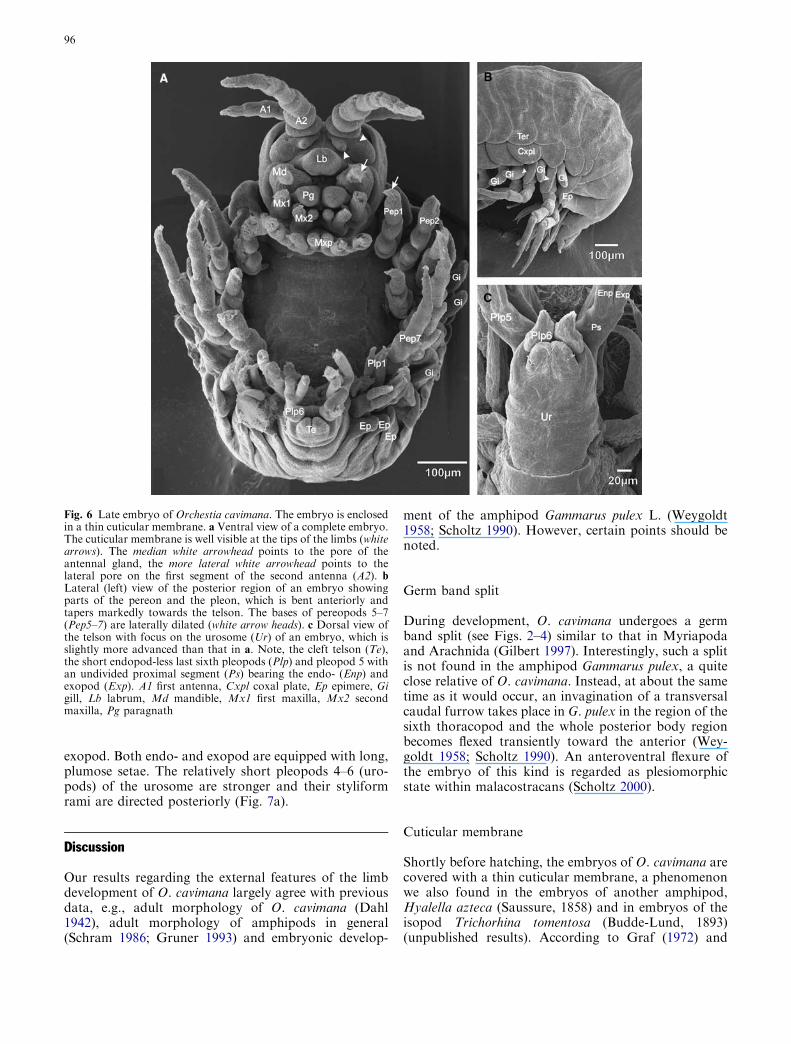
The limb segments become elongated and the embryodifferentiates characteristic tagmata (Fig. 5a, c). Thecephalothorax is compact, as its appendages, including
Fig. 2 Forming of the post-naupliar limb buds and initial germband split of Orchestia cavimana. Anterior is up. a Ventral view ofthe whole embryo. The white line indicates the germ band split. Thethree white arrowheads at the level of the first (A1) and secondantenna (A2) point to the stomodaeal projections. b Posteriorregion of the same embryo as in a. The midline is marked byarrows. Asterisks show damages from preparation. The interseg-mental furrows of pleomeres 4–6 (Pl4-6) are about to differentiate.The segment of the sixth and final pleomere (Pl) is marked withdotted lines. c Anterior region of an embryo slightly more advancedas in a and b, showing the stomodaeal projections (white arrowheads) moving into the forming stomodaeum. d Posterior region ofan embryo slightly more advanced as in a and b, showing four tofive regular rows of cells behind the sixth pleomere (Pl6) (narrowedwith dotted lines) and the Anlage of the telson (white lined circle).Cxpl+Gi shared Anlage of coxal plate and gill, Md mandible, Mx1first maxilla, Mx2 second maxilla, Mxp maxilliped, Pe1–Pe7pereomeres 1–7, Ter tergite, yc yolky cells
92
the first thoracopod (maxilliped), shift medially. Thepaired labrum Anlage anterior to the stomodaeum andclose to the mandible, is now completely fused (Fig. 5a,b). The first segment of the second antenna bears a lat-eral and a median pore on its posterior margin, the latterrepresenting the opening of the prospective antennalgland (Fig. 5b). The mandibles are bent towards eachother with the inner and the outer lobe arranged in aproximomedian and a laterodistal position, respectively.The mediobasally fused paragnaths appear as largehumps with laterobasal projections, the mandibularprocesses (Fig. 5a). The palp of the first maxilla is
reduced to a small lateral outgrowth. The mandible andthe first maxilla have a lateral position, while the secondmaxilla is inserted more medially. The coxae of themaxilliped start to fuse. The fourth palp segment isslightly reduced (Fig. 5a).
The coxal plates of the pereopods further expand andflatten; the sac-like gills on their posterior margins grow;and the broad tergal plates of the pereon segments havefurther elongated lateroventrally. Similar to the propo-dus of the second pereopod (see Sect. 4), the propodus ofthe first pereopod also enlarges and begins to specialiseas a sub-chela (Fig. 5a). The pleon is very compact and
Fig. 3 Elongation and initialsegmentation of the limbs ofOrchestia cavimana. Anterior isup. a Ventral view of the wholeembryo. White arrowheadsshow the stomodaealprojections, which have startedsinking into the formingstomodaeum. The leftmaxilliped (Mxp) is missing (g).b Posterior region of the sameembryo as in a. c Detail of firstmaxilla (Mx1), second maxilla(Mx2) and maxilliped of anembryo in the same stage as ina. White arrows point to theforming endites of the first andsecond maxilla and themaxilliped. The palp (P) of themaxilliped projects medially. A1first antenna, A2 secondantenna, Cxpl+Gi sharedAnlage of coxal plate and gill,Enp endopod, Exp exopod, Lblabrum, Md mandible, Pep 1–7pereopods 1–7, Pg paragnath,Plp1–6 pleopods 1–6, Proproctodaeum, Ps proximalsegment, Te telson
93
bent slightly anteriorly. The subdivision of the proximalsegment of pleopods 1–4 is still visible. The last pleopodloses its biramy, the endopod disappears. The epimerescontinue to expand (Fig. 5c). The proctodaeum hasshifted anteriorly and the lateral lobes of the telson aretransformed into basally fused cylindric outgrowths,giving the telson a cleft appearance. The tergite ofpleomere 6 and the telson are dorsally fused (Fig. 5c).
Late embryo, shortly before hatching
Limb segments lose their spherical shape at this stage,and the tagmatisation into a cephalothorax, a pereon
and a pleon is clearly visible. The whole embryo is nowenclosed in a thin cuticular membrane (Fig. 6a–c). Thefirst segment of the second antenna is partially fusedwith the head capsule; the pore of the antennal glandopens at the tip of a broad, posteriorly directed, conicaltube. The lateral pore is still visible (Fig. 6a). The dis-tance between the unpaired, flat labrum and the secondantenna has increased. The visible parts of the mandibleare a narrow distal lobe and a broader proximomedianlobe (probably the prospective incisor and molar pro-cesses, respectively). The paragnathal humps have be-come slightly narrower. More laterally, behind theparagnaths, the first maxilla inserts with two unequalendites: the very small, thin inner and the larger outerendite. The palp of the first maxilla is strongly reduced(hidden by the cuticular membrane in this phase ofdevelopment). The endites of the second maxilla arealmost equal in size, but slightly smaller than the outerendite of the first maxilla. The fusion of the maxillipedalcoxae has further propagated (Fig. 6a).
The coxal plates of pereopods 1–7 are differentiatedinto broad, flat, imbricate shields that cover the inser-tions of the limbs and the gills. The coxal plates, in turn,are partly covered by the tergal plates. The bases ofpereopods 5–7 are anterolaterally expanded as shields(Fig. 6b). The gills are convoluted (Fig. 6a, b). Thepleon tapers markedly towards the telson. Pleopod 4 isequal in size to pleopods 1–3; pleopod 5 is half size, and
Fig. 4 Closure of the germ band of Orchestia cavimana. a Ventralview of the anterior region. White arrowheads point at the posteriorincision of the first podomere of the second antenna (A2). Note, thebilobate shape of the mandible (Md). b Posterior view of the sameembryo as in a. White asterisks mark the paired lateral lobes of thetelson. c Ventral view of pereopods 2–4 (Pep 2–4) showing fromproximal to distal: coxa [of which only the coxal plate (Cxpl) isvisible], basis (Ba), ischium (Is), merus (Me), carpus (Ca), propodus(Prop) and dactylus (Da). Anterior is up. Note, the dilated shape ofthe propodus (white arrow head) of the second pereopod, which isspecialised as a sub-chela (gnathopod) in the adults. d. Lateral viewof the pleopods (Plp) illustrating the subdivision of the proximalsegments (Ps) indicated by white arrowheads. A1 first antenna, Eendite, Enp endopod, Ep epimere, Exp exopod, Gi gill, Lb labrum,Mx1 first maxilla, Mx2 second maxilla, Mxp maxilliped, Pgparagnath, Ter tergite
94

the endopod-less pleopod 6 is a very short stump. Thesubdivision of the proximal segment of pleopods 1–4 isno longer visible. The epimeres are further enlarged,tapering ventrally and hiding the insertions of pleopods1–3. Pleomeres 4–6 and the fused, cleft telson form theurosome (Fig. 6a–c).
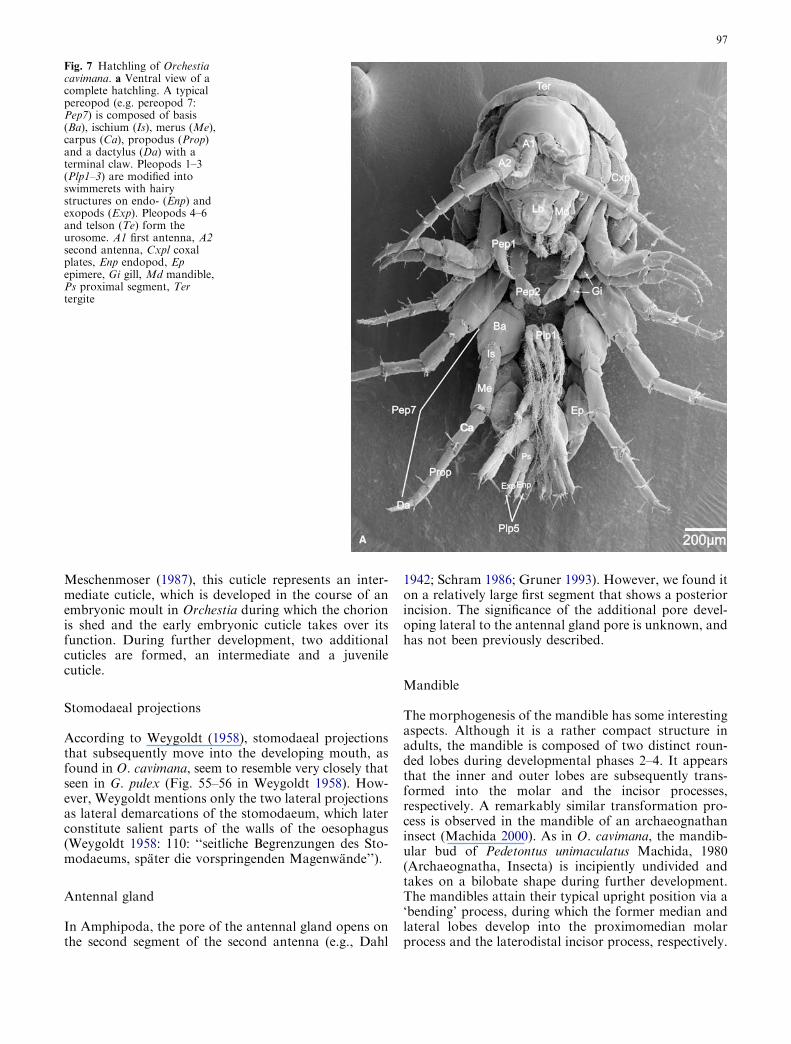
Hatchling
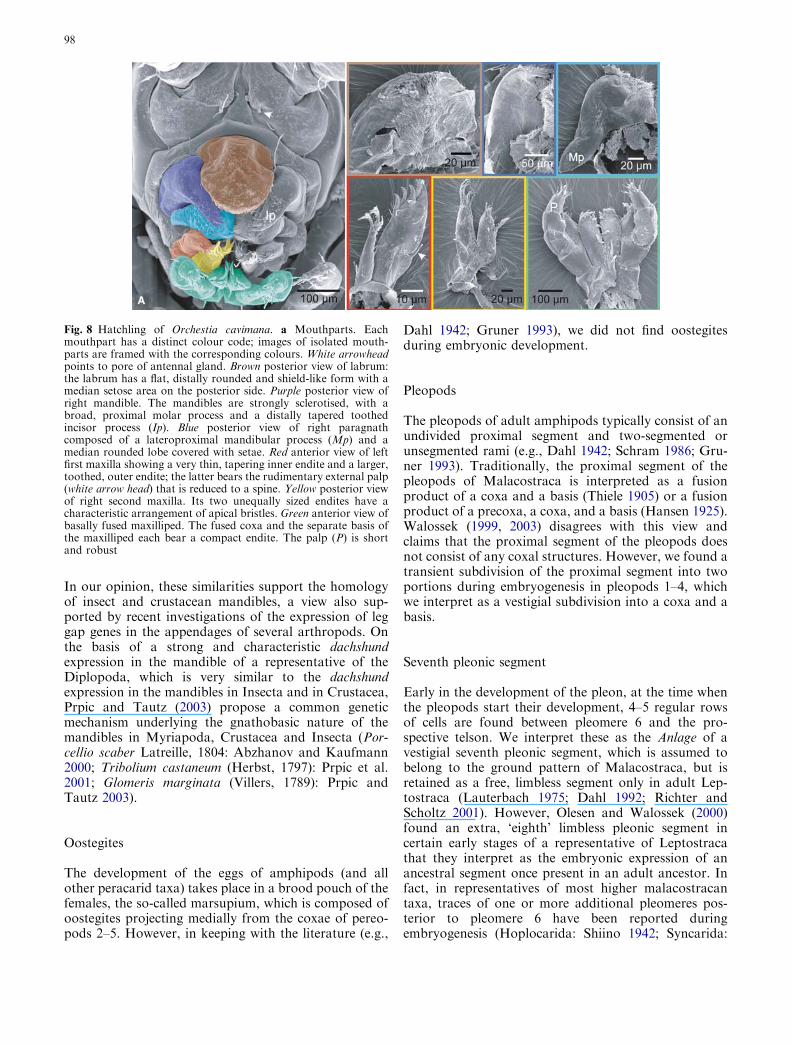
The body of the hatchling is laterally compressed andstrongly curved. The fused cephalothoracic segmentsform a compact cuticular head capsule, with the twopairs of uniramous antennae pointing anteroventrally.The first antenna is one-third length of the second an-tenna (Fig. 7a). The opening of the antennal gland isvisible as a short, narrow tube posteromedially to thehind margin of the first antennal segment; but the sec-ond, more lateral pore has disappeared, and is probablyhidden in the fold between the antennal base and headcapsule (Fig. 8a).
More ventrally, the shield-like labrum inserts. Itsposterior side bears a median setose area and borders theanterior margin of the oral opening, partly covering theventrally directed mouthparts. Laterally, the mouth isdemarcated by the strongly sclerotised mandible, withthe proximomedian broad molar process and the distallytapered toothed incisor process. Posteromedian to themandible are the medially fused paragnaths, whichconstitute the posterior border of the oral opening. Theparagnaths are composed of rounded, setose medianhooks and proximolateral mandibular processes. Thefirst maxilla is composed of a very thin, tapering innerendite and a larger, toothed, outer endite. The outerendite bears the rudimentary external palp, which isreduced to a spine. The second maxilla is composed oftwo short endites with setose tips, the outer endite isslightly longer than the inner endite. The fused coxa andseparate basis of the maxilliped each bear a short endite.The palp has four segments (of which the most distalsegment is strongly reduced) and is short and robust(Fig. 8a).
The tergal and coxal plates of the pereon and theepimeres of the pleon appear bloated. The slender andvery short sub-chelate gnathopods 1–2 are bent antero-medially. The propodus of gnathopod 1 is relativelylarge, with its dactylus recurved back along it. The shortdactylus of gnathopod 2 is closed against a distallyrounded, elongate and setose part of the propodus(Fig. 7a). Pereopods 3–4 are directed anterolaterally,with dactyli pointing posteriorly. Pereopods 5–7 are bentposteriorly, with anteriorly directed dactyli. Their basesare long and broad compared to the relatively slenderbases of pereopods 3–4. Pereopods 6–7 are significantlylonger than pereopods 3–5. The short convoluted gillsnow project medially (Fig. 7a). The three anterior-most,medially shifted pleopods are differentiated into swim-merets. They consist of a proximal segment bearing athin, undivided endopod, and a thin two-segmented
Fig. 5 Differentiation of characteristic tagmata in Orchestia cavi-mana. Anterior is up. a Ventral view of anterior body regionshowing the endites (E) of the first (Mx1) and second maxilla(Mx2), the external palp (P) of the first maxilla and the paragnaths(Pg) with laterobasal mandibular processes (Mp). Note, thecharacteristic shape of the propodi (white arrow heads) ofpereopods 1–2 (Pep 1–2), indicating a beginning specialisation asgnathopods in the adults: the propodus of the first pereopod isbroader and longer than the propodi of pereopods 3–7 and thepropodus of the second pereopod has a broad median extension.The left side of the embryo was damaged during preparation(asterisks). b Detail of the proximal region of the second antenna(A2) and the labrum (Lb), showing the pore of the prospectiveantennal gland (white arrow head) and a more lateral situated pore(black arrow head) on the first segment of the second antenna. Leftsecond antenna is broken off (white asterisk). c Posterior view ofpleon. The subdivision of the proximal segment (Ps) of pleopods 1–4 (Plp 1–4) is still visible. Pleopod 6 (Plp6) has lost its endopod(Enp). A1 first antenna, Cxpl coxal plate, Ep epimere, Exp exopod,Gi gill, Md mandible, Mxp maxilliped, Te telson
95
exopod. Both endo- and exopod are equipped with long,plumose setae. The relatively short pleopods 4–6 (uro-pods) of the urosome are stronger and their styliformrami are directed posteriorly (Fig. 7a).
Discussion
Our results regarding the external features of the limbdevelopment of O. cavimana largely agree with previousdata, e.g., adult morphology of O. cavimana (Dahl1942), adult morphology of amphipods in general(Schram 1986; Gruner 1993) and embryonic develop-
ment of the amphipod Gammarus pulex L. (Weygoldt1958; Scholtz 1990). However, certain points should benoted.
Germ band split
During development, O. cavimana undergoes a germband split (see Figs. 2–4) similar to that in Myriapodaand Arachnida (Gilbert 1997). Interestingly, such a splitis not found in the amphipod Gammarus pulex, a quiteclose relative of O. cavimana. Instead, at about the sametime as it would occur, an invagination of a transversalcaudal furrow takes place in G. pulex in the region of thesixth thoracopod and the whole posterior body regionbecomes flexed transiently toward the anterior (Wey-goldt 1958; Scholtz 1990). An anteroventral flexure ofthe embryo of this kind is regarded as plesiomorphicstate within malacostracans (Scholtz 2000).
Cuticular membrane
Shortly before hatching, the embryos of O. cavimana arecovered with a thin cuticular membrane, a phenomenonwe also found in the embryos of another amphipod,Hyalella azteca (Saussure, 1858) and in embryos of theisopod Trichorhina tomentosa (Budde-Lund, 1893)(unpublished results). According to Graf (1972) and
Fig. 6 Late embryo of Orchestia cavimana. The embryo is enclosedin a thin cuticular membrane. a Ventral view of a complete embryo.The cuticular membrane is well visible at the tips of the limbs (whitearrows). The median white arrowhead points to the pore of theantennal gland, the more lateral white arrowhead points to thelateral pore on the first segment of the second antenna (A2). bLateral (left) view of the posterior region of an embryo showingparts of the pereon and the pleon, which is bent anteriorly andtapers markedly towards the telson. The bases of pereopods 5–7(Pep5–7) are laterally dilated (white arrow heads). c Dorsal view ofthe telson with focus on the urosome (Ur) of an embryo, which isslightly more advanced than that in a. Note, the cleft telson (Te),the short endopod-less last sixth pleopods (Plp) and pleopod 5 withan undivided proximal segment (Ps) bearing the endo- (Enp) andexopod (Exp). A1 first antenna, Cxpl coxal plate, Ep epimere, Gigill, Lb labrum, Md mandible, Mx1 first maxilla, Mx2 secondmaxilla, Pg paragnath
96
Meschenmoser (1987), this cuticle represents an inter-mediate cuticle, which is developed in the course of anembryonic moult in Orchestia during which the chorionis shed and the early embryonic cuticle takes over itsfunction. During further development, two additionalcuticles are formed, an intermediate and a juvenilecuticle.
Stomodaeal projections
According to Weygoldt (1958), stomodaeal projectionsthat subsequently move into the developing mouth, asfound in O. cavimana, seem to resemble very closely thatseen in G. pulex (Fig. 55–56 in Weygoldt 1958). How-ever, Weygoldt mentions only the two lateral projectionsas lateral demarcations of the stomodaeum, which laterconstitute salient parts of the walls of the oesophagus(Weygoldt 1958: 110: ‘‘seitliche Begrenzungen des Sto-modaeums, spater die vorspringenden Magenwande’’).
Antennal gland
In Amphipoda, the pore of the antennal gland opens onthe second segment of the second antenna (e.g., Dahl
1942; Schram 1986; Gruner 1993). However, we found iton a relatively large first segment that shows a posteriorincision. The significance of the additional pore devel-oping lateral to the antennal gland pore is unknown, andhas not been previously described.
Mandible
The morphogenesis of the mandible has some interestingaspects. Although it is a rather compact structure inadults, the mandible is composed of two distinct roun-ded lobes during developmental phases 2–4. It appearsthat the inner and outer lobes are subsequently trans-formed into the molar and the incisor processes,respectively. A remarkably similar transformation pro-cess is observed in the mandible of an archaeognathaninsect (Machida 2000). As in O. cavimana, the mandib-ular bud of Pedetontus unimaculatus Machida, 1980(Archaeognatha, Insecta) is incipiently undivided andtakes on a bilobate shape during further development.The mandibles attain their typical upright position via a‘bending’ process, during which the former median andlateral lobes develop into the proximomedian molarprocess and the laterodistal incisor process, respectively.
Fig. 7 Hatchling of Orchestiacavimana. a Ventral view of acomplete hatchling. A typicalpereopod (e.g. pereopod 7:Pep7) is composed of basis(Ba), ischium (Is), merus (Me),carpus (Ca), propodus (Prop)and a dactylus (Da) with aterminal claw. Pleopods 1–3(Plp1–3) are modified intoswimmerets with hairystructures on endo- (Enp) andexopods (Exp). Pleopods 4–6and telson (Te) form theurosome. A1 first antenna, A2second antenna, Cxpl coxalplates, Enp endopod, Epepimere, Gi gill, Md mandible,Ps proximal segment, Tertergite
97
In our opinion, these similarities support the homologyof insect and crustacean mandibles, a view also sup-ported by recent investigations of the expression of leggap genes in the appendages of several arthropods. Onthe basis of a strong and characteristic dachshundexpression in the mandible of a representative of theDiplopoda, which is very similar to the dachshundexpression in the mandibles in Insecta and in Crustacea,Prpic and Tautz (2003) propose a common geneticmechanism underlying the gnathobasic nature of themandibles in Myriapoda, Crustacea and Insecta (Por-cellio scaber Latreille, 1804: Abzhanov and Kaufmann2000; Tribolium castaneum (Herbst, 1797): Prpic et al.2001; Glomeris marginata (Villers, 1789): Prpic andTautz 2003).
Oostegites
The development of the eggs of amphipods (and allother peracarid taxa) takes place in a brood pouch of thefemales, the so-called marsupium, which is composed ofoostegites projecting medially from the coxae of pereo-pods 2–5. However, in keeping with the literature (e.g.,
Dahl 1942; Gruner 1993), we did not find oostegitesduring embryonic development.
Pleopods
The pleopods of adult amphipods typically consist of anundivided proximal segment and two-segmented orunsegmented rami (e.g., Dahl 1942; Schram 1986; Gru-ner 1993). Traditionally, the proximal segment of thepleopods of Malacostraca is interpreted as a fusionproduct of a coxa and a basis (Thiele 1905) or a fusionproduct of a precoxa, a coxa, and a basis (Hansen 1925).Walossek (1999, 2003) disagrees with this view andclaims that the proximal segment of the pleopods doesnot consist of any coxal structures. However, we found atransient subdivision of the proximal segment into twoportions during embryogenesis in pleopods 1–4, whichwe interpret as a vestigial subdivision into a coxa and abasis.
Seventh pleonic segment
Early in the development of the pleon, at the time whenthe pleopods start their development, 4–5 regular rowsof cells are found between pleomere 6 and the pro-spective telson. We interpret these as the Anlage of avestigial seventh pleonic segment, which is assumed tobelong to the ground pattern of Malacostraca, but isretained as a free, limbless segment only in adult Lep-tostraca (Lauterbach 1975; Dahl 1992; Richter andScholtz 2001). However, Olesen and Walossek (2000)found an extra, ‘eighth’ limbless pleonic segment incertain early stages of a representative of Leptostracathat they interpret as the embryonic expression of anancestral segment once present in an adult ancestor. Infact, in representatives of most higher malacostracantaxa, traces of one or more additional pleomeres pos-terior to pleomere 6 have been reported duringembryogenesis (Hoplocarida: Shiino 1942; Syncarida:
Fig. 8 Hatchling of Orchestia cavimana. a Mouthparts. Eachmouthpart has a distinct colour code; images of isolated mouth-parts are framed with the corresponding colours. White arrowheadpoints to pore of antennal gland. Brown posterior view of labrum:the labrum has a flat, distally rounded and shield-like form with amedian setose area on the posterior side. Purple posterior view ofright mandible. The mandibles are strongly sclerotised, with abroad, proximal molar process and a distally tapered toothedincisor process (Ip). Blue posterior view of right paragnathcomposed of a lateroproximal mandibular process (Mp) and amedian rounded lobe covered with setae. Red anterior view of leftfirst maxilla showing a very thin, tapering inner endite and a larger,toothed, outer endite; the latter bears the rudimentary external palp(white arrow head) that is reduced to a spine. Yellow posterior viewof right second maxilla. Its two unequally sized endites have acharacteristic arrangement of apical bristles. Green anterior view ofbasally fused maxilliped. The fused coxa and the separate basis ofthe maxilliped each bear a compact endite. The palp (P) is shortand robust
98

Hickman 1937; Mysidacea: Manton 1928; Tanaidacea:Scholl 1963; Isopoda: Stromberg 1967; Decapoda:Scholtz 1995). Weygoldt (1958) did not find a seventhpleonic segment in his investigation of the developmentof the amphipod Gammarus pulex. However, he reportsthat the Anlage of the sixth pleonic ganglion is subdi-vided into two distinct, adjacent areas. Weygoldtinterprets this subdivision as a vestigial formation of aseventh pleonic ganglion.
Acknowledgements We are very grateful to G. Drescher of theInstitute of Systematic Zoology of the Natural History Museum,Berlin, for support using the SEM. Prof. Gerhard Scholtz spentvaluable time going through the manuscript, which is gratefullyacknowledged. We thank S. Richter for helpful discussions. Manythanks also to P. T. Graham for improving the English.
References
Abzhanov A, Kaufman TC (2000) Homologs of Drosophilaappendage genes in the patterning of arthropod limbs. Dev Biol227:673–689
Dahl F (1942) Krebstiere oder Crustacea, IV: Flohkrebse oderAmphipoda. In: Schellenberg A (ed) Die Tierwelt Deutsch-lands, vol 40. Gustav Fischer Verlag, Jena, pp 1–252
Dahl E (1992) Aspects of malacostracan evolution. Acta Zool73:339–346
Dohle W, Scholtz G (1988) Clonal analysis of the crustacean seg-ment: the discordance between genealogical and segmentalborders. Dev Suppl 104:147–160
Dohle W, Scholtz G (1997) How far does cell lineage influence cellfate specification in crustacean embryos? Cell Dev Biol 8:379–390
Dohle W, Gerberding M, Hejnol A, Scholtz G (2004) Cell lineage,segment differentiation, and gene expression in crustaceans.In: Scholtz G (ed) Crustacean issues 15, evolutionary develop-mental biology of Crustacea. Balkema, Lisse, pp 95–133
Gerberding M, Scholtz G (1999) Cell lineage of the midline cells inthe amphipod crustacean Orchestia cavimana (Crustacea, Mal-acostraca) during formation and separation of the germ band.Dev Genes Evol 209:91–102
Gerberding M, Scholtz G (2001) Neurons and glia in the midline ofthe higher crustacean Orchestia cavimana are generated via aninvariant cell lineage that comprises a median neuroblast andglial progenitors. Dev Biol 235:397–409
Gilbert SF (1997) Chapter 13. Arthropods: the crustaceans, spi-ders, and myriapods. In: Gilbert SF, Raunio AM (eds)Embryology. Sinauer Associates, Sunderland, pp 237–258
Graf F, note transmise par Grasse P-P (1972) Stockage de calciumet formation des soies chez l’embryon d’ Orchestia (Crustace,Amphipode, Talitride). Notion d’intermue embryonnaire. C RAcad Sc Paris 275:1669–1672
Gruner HE (1993) 1. Klasse Crustacea. In: Gruner HE (ed) Lehr-buch der speziellen Zoologie. Band 1: Wirbellose Tiere. 4. Teil:Arthropoda (ohne Insecta). Gustav Fischer Verlag, Jena, pp448–1030
Hansen HJ (1925) Studies on Arthropoda II. On the comparativemorphology of the appendages in the Arthropoda. A Crusta-cea. Copenhagen Gyldendalske Boghandel. Kopenhagen,London, Berlin, pp 1–176
Hejnol A, Scholtz G (2004) Clonal analysis of Distal-less and en-grailed expression patterns during early morphogenesis ofuniramous and biramous crustacean limbs. Dev Genes Evol214(10):473-485
Hickman VV (1937) The embryology of the syncarid crustacean,Anaspides tasmaniae. Proc Roy Soc Tasmania 1–35
Lauterbach K-E (1975) Uber die Herkunft der Malacostraca(Crustacea). Zool Anz 194:165–179
Machida R (2000) Serial homology of the mandible and maxilla inthe jumping bristletail Pedetontus unimaculatus Machida, basedon external embryology (Hexapoda: Archaeognatha, Machili-dae). J Morphol 245:19–28
Manton SM (1928) On the embryology of a mysid crustacean,Hemimysis lamornae. Philos Trans R Soc Lond (B) 216:363–463
Meschenmoser M (1987) Zur Embryonalentwicklung von Orches-tia cavimana Heller 1865 (Crustacea, Amphipoda) unter be-sonderer Berucksichtigung des Dorsalorgans. Diplomarbeit,Westfalische-Wilhelms-Universitat Munster
Olesen J, Walossek D (2000) Limb ontogeny and trunk segmen-tation in Nebalia species (Crustacea, Malacostraca, Leptostra-ca). Zoomorphology 120:47–64
Prpic N-M, Tautz D (2003) The expression of the proximodistalaxis patterning genes Distal-less and dachshund in the append-ages of Glomeris marginata (Myriapoda: Diplopoda) suggests aspecial role of these genes in patterning the head appendages.Dev Biol 260:97–112
Prpic N-M, Wigand B, Damen WGM, Klingler M (2001)Expression of dachshund in wild-type and Distal-less mutantTribolium corroborates serial homologies in insect appendages.Dev Genes Evol 211:467–477
Richter S, Scholtz G (2001) Phylogenetic analysis of the Mala-costraca (Crustacea). J Zool Syst Evol Res 39:113–136
Scholl G (1963) Embryologische Untersuchungen an Tanaidaceen(Heterotanais oerstedi Kroyer). Zool Jb Anat 80:500–545
Scholtz G (1990) The formation, differentiation and segmentationof the post-naupliar germ band of the amphipod Gammaruspulex L. (Crustacea, Malacostraca, Peracarida). Proc R SocLond B 239:163–211
Scholtz G (1995) Expression of the Engrailed gene reveals nineputative segment-anlagen in the embryonic pleon of the fresh-water crayfish Cherax destructor (Crustacea, Malacostraca,Decapoda). Biol Bull 188:157–165
Scholtz G (2000) Evolution of the nauplius stage in malacostracancrustaceans. J Zool Syst Evol Res 38: 175–187
Scholtz G, Dohle W (1996) Cell lineage and cell fate in crustaceanembryos - a comparative approach. Int J Dev Biol 40:211–220
Scholtz G, Wolff C (2002) Cleavage, gastrulation, and germ discformation of the amphipod Orchestia cavimana (Crustacea,Malacostraca, Peracarida). Contrib Zool 71:9–28
Scholtz G, Mittmann B, Gerberding M (1998) The pattern ofDistal-less expression in the mouthparts of crustaceans, myr-iapods and insects: new evidence for a gnathobasic mandibleand the common origin of Mandibulata. Dev Biol 42:801–810
Schram FR (1986) Crustacea. Oxford Unviersity Press, New York,Oxford, pp 1–606
Shiino SM (1942) Studies on the embryology of Squilla oratoria deHaan. Mem Coll Sci Kyoto Imp Univ B 17:77–174
Stromberg JO (1967) Segmentation and organogenesis in Limnorialignorum (Rathke) (Isopoda). Ark Zool 20:145–179
Thiele J (1905) Betrachtungen uber die Phylogenie der Crusta-ceenbeine. Z Wiss Zool 82:445–471
Walossek D (1999) On the Cambrian diversity of Crustacea. In:Schram FR, von Vaupel Klein JC (eds) Crustaceans and thebiodiversity crisis. In: Proceedings of the 4th InternationalCrustacean Congress, Amsterdam, The Netherlands, July 20–24, 1998, vol. 1, Brill Academic Publishers, Leiden, pp 3–27
Walossek D (2003) Cambrian ‘Orsten’-type arthropods and thephylogeny of Crustacea. In: Legakis A et al eds. The newpanorama of animal evolution. In: Proceedings of the 18thinternational congress of Zoology, Athens, Pensoft Publishers,Sofia, Moscow, pp 67–84
Weygoldt P (1958) Die Embryonalentwicklung des AmphipodenGammarus pulex pulex (L.). Zool Jb Anat 77:51–110
Wolff C, Scholtz G (2002) Cell lineage, axis formation, and theorigin of germ layers in the amphipod crustacean Orchestiacavimana. Dev Biol 250:44–58
99