coenzyme q10 production in recombinant escherichia coli strains engineered with a heterologous...
TRANSCRIPT

ARTICLE IN PRESS
1096-7176/$ - se
doi:10.1016/j.ym
�CorrespondGyeongsang Na
E-mail addr
Metabolic Engineering 8 (2006) 406–416
www.elsevier.com/locate/ymben
Coenzyme Q10 production in recombinant Escherichia coli strainsengineered with a heterologous decaprenyl diphosphate synthase gene
and foreign mevalonate pathway
Hossein Shahbani Zahiria,b, Sang Hwal Yoonc, Jay. D. Keaslingd, Si Hyoung Leee,Seon Won Kimb,c, Sung Chul Yoonb, Yong Chul Shina,e,�
aDepartment of Microbiology, Gyeongsang National University, Gazwa-dong 900, Jinju 660-701, KoreabDivision of Applied Life Science (BK21), Gyeongsang National University, Jinju 660-701, Korea
cEnvironmental Biotechnology Research Center, Gyeongsang National University, Jinju 660-701, KoreadDepartment of Chemical Engineering, 201 Gilman Hall, University of California, Berkeley, CA 94720-1462, USA
eAmicogen, Inc., Jinju 660-852, Korea
Received 17 November 2005; received in revised form 5 May 2006; accepted 8 May 2006
Available online 20 May 2006
Abstract
In the present work, Escherichia coli DH5a was metabolically engineered for CoQ10 production by the introduction of decaprenyl
diphosphate synthase gene (ddsA) from Agrobacterium tumefaciens. Grown in 2YTG medium (1.6% tryptone, 1% yeast extract, 0.5%
NaCl, and 0.5% glycerol) with an initial pH of 7, the recombinant E. coli was capable of CoQ10 production up to 470mg/gDCW (dry cell
weight). This value could be further elevated to 900 mg/gDCW simply by increasing the initial culture pH from 7 to 9. Supplementation of
4-hydroxy benzoate did not improve the productivity any further. However, engineering of a lower mevalonate semi-pathway so as to
increase the isopentenyl diphosphate (IPP) supply of the recombinant strain using exogenous mevalonate efficiently increased the CoQ10
production. Lower mevalonate semi-pathways of Staphylococcus aureus, Streptococcus pyogenes, Streptococcus pneumoniae,
Enterococcus faecalis, and Saccharomyces cerevisiae were tested. Among these, the pathway of Streptococcus pneumoniae proved to
be superior, yielding CoQ10 production of 27007115mg/gDCW when supplemented with exogenous mevalonate of 3mM. In order to
construct a complete mevalonate pathway, the upper semi-pathway of the same bacterium, Streptococcus pneumoniae, was recruited. In a
recombinant E. coli DH5a harboring three plasmids encoding for upper and lower mevalonate semi-pathways as well as DdsA enzyme,
the heterologous mevalonate pathway could convert endogenous acetyl-CoA to IPP, resulting in CoQ10 production of up to
2428775mg/gDCW, without mevalonate supplementation. In contrast, a whole mevalonate pathway constructed in a single operon was
found to be less efficient. However, it provided CoQ10 production of up to 1706786mg/gDCW, which was roughly 1.9 times higher than
that obtained by ddsA alone.
r 2006 Elsevier Inc. All rights reserved.
Keywords: Coenzyme Q10; ddsA gene; Mevalonate pathway; Metabolic engineering; IPP; Escherichia coli
1. Introduction
Ubiquinones, including coenzyme Q10 (CoQ10), areobligatory cofactors in aerobic respiratory electron transferfor energy generation. As membrane-bound redox-activemolecules, they also participate in several other cellular
e front matter r 2006 Elsevier Inc. All rights reserved.
ben.2006.05.002
ing author. Department of Microbiology, Gyeongsang
tional University, Gazwa-dong 900, Jinju 660-701, Korea.
ess: [email protected] (Y.C. Shin).
functions such as formation of disulfide bonds in proteins,detoxification of harmful reactive oxygen species, controlof cellular redox status, generation of cell signals, and geneexpression (Battino et al., 1990; Seballe and Poole, 1999;Bader et al., 1999; Kawamukai, 2002; Turunen et al., 2004;Crane, 2001; Georgellis et al., 2001; Groneberg et al.,2005).The administration of CoQ10 has proved promising in
the prevention and treatment of cardiovascular, neurode-generative, and mitochondrial diseases (Matthews et al.,

ARTICLE IN PRESSH.S. Zahiri et al. / Metabolic Engineering 8 (2006) 406–416 407
1998; Sharma et al., 2004; Tran et al., 2001; Bhagavan andChopra, 2005; Rosenfeldt et al., 2005; Somayajulu et al.,2005). Accordingly, in recent years, there has been growinginterest in CoQ10 as a medicine or health-promotingadditive to foods and cosmetics. In line with a globalincrease in CoQ10 demand, attempts at its production bymeans of synthetic (Negishi et al., 2002), semi-synthetic(Lipshutz et al., 2002), and biotechnological (Park et al.,2004; Yoshida et al., 1998) procedures have been made.Coenzyme Q is composed of a benzoquinone ring that isprenylated with an isoprenoid chain of various lengths(Meganathan, 2001) (Fig. 1). The benzoquinone ring issynthesized from 4-hydroxybenzoate (4HB), whereas theisoprenoid chain is made up of isopentenyl diphosphate(IPP) units. The formation of a side chain is catalyzed bypolyprenyl diphosphate synthase, which combines the IPPunits up to a certain length depending on the organism(Okada et al., 1996). Polyprenyl diphosphate synthasesfrom human and some other organisms including Agro-
bacterium tumefaciens combine 10 IPP units to formdecaprenyl diphosphate.
Several studies have shown that metabolic engineering ofIPP biosynthesis by overexpression of rate-limiting genes ofthe non-mevalonate pathway (Kim and Keasling, 2001;Matthews and Wurtzel, 2000; Albrecht et al., 1999) orthrough the use of an additional foreign mevalonatepathway (Martin et al., 2003) results in a significantincrease in isoprenoids biosynthesis in Escherichia coli. Inthis report, we describe CoQ10 production in a recombi-nant E. coli that was engineered with a decaprenyldiphosphate synthase gene from A. tumefaciens. The effectof culture pH and increased IPP supply, achieved byengineering of a foreign mevalonate pathway, were studiedin terms of CoQ10 production in various recombinantE. coli strains.
2. Materials and methods
2.1. Bacterial strains and plasmids
E. coli DH5a was used for plasmid construction andCoQ10 production (Table 2). A. tumefaciens (ATCC33970), Staphylococcus aureus (ATCC 35556), Streptococ-
cus pyogenes (ATCC 12344), Streptococcus pneumoniae
(ATCC 6314), Enterococcus faecalis (ATCC 700802),Saccharomyces cerevisiae (ATCC 47058), Ralstonia eutro-
pha (ATCC 43123), and E. coli W3110 (ATCC 27325) wereused as gene donors.
The vectors pSTV28, pTrc99A, and pBBR1MCS2 wereused for cloning and expression of genes and operons(Table 2). Restriction enzymes were purchased from RocheApplied Science and New England Biolabs. T4 DNA ligaseand DNA polymerase (Klenow fragment) were purchasedfrom New England Biolabs. Pfu DNA polymerase waspurchased from Promega. Plasmid preparation, gel extrac-tion, and PCR purification kits were of Qiagen products.
2.2. Media and culture conditions
Luria–Bertani (LB) medium was used for routinecultivation of E. coli strains. For CoQ10 production, 5mlof 2YT medium (1.6% tryptone, 1% yeast extract, 0.5%NaCl) was used for overnight precultivation of E. coli in atest tube at 37 1C. Main cultures were conducted by usingthe same medium supplemented with 0.5% glycerol and0.01% 4-hydroxy benzoic acid (4HB, sodium salt, Sigma),either in 25ml tubes containing 7ml medium or 250mlflasks containing 25ml medium. Where necessary, meva-lonate was included in the culture medium. Mevalonatewas prepared by alkaline hydrolysis of mevalonolactone(Sigma) according to a method reported previously (Kimet al., 1992). Appropriate antibiotics (ampicillin 100 mg/ml,chloramphenicol 50 mg/ml, kanamycin 30 mg/ml, and fos-midomycin 1mM) were added to the culture media asrequired. Main cultures were inoculated to a startingOD600 of 0.1 and incubated at 30 1C for 24 h in a rotaryshaking incubator at 200 rpm. Cell growth was measured at600 nm and converted to dry cell weight (gDCW/l) by usinga standard curve.
2.3. Plasmid construction
The ddsA gene, coding for decaprenyl diphosphatesynthase, was isolated from genomic DNA of A. tumefa-
ciens by PCR using pfu DNA polymerase. The PCRproduct was cut by EcoRI and BamHI, and introducedinto pTrc99A vector, resulting in pTatdds (Fig. 2a).The idi gene and the genes of the mevalonate pathway
were cloned from genomic DNA of corresponding yeast orbacterial strains by PCR using pfu DNA polymerase, asgiven in Tables 1 and 2. The mevalonate pathway wasdivided into lower and upper semi-pathways (Fig. 1), whichwere constructed in two separate artificial operons. Inorder to construct the operons, each open reading framefollowing a Shine–Dalgarno ribosomal binding sequenceand a spacer was cloned into the multiple cloning site of agiven plasmid. The lower mevalonate semi-pathwayencoding for mevalonate kinase (mvaK1), phosphomeva-lonate kinase (mvaK2), and mevalonate diphosphatedecarboxylase (mvaD) was constructed together with theidi gene in a single operon using pSTV28 as a vector(Fig. 2b). A functional lower mevalonate semi-pathwayconverts mevalonate to IPP.The operon coding for the upper mevalonate semi-
pathway containing acetoacetyl-CoA thiolase (phbA), 3-hydroxy-3-methylglutaryl-CoA synthase (mvaS), and 3-hydroxy-3-methylglutaryl-CoA reductase (mvaA) fromStreptococcus pneumoniae was constructed inpBBR1MCS2, resulting in pBSNmvaASP (Fig. 2c). Thissemi-pathway produces mevalonate from acetyl-CoA andmay feed to the lower mevalonate semi-pathway for IPPbiosynthesis.In order to construct the whole mevalonate pathway in a
single operon, the upper mevalonate semi-pathway was
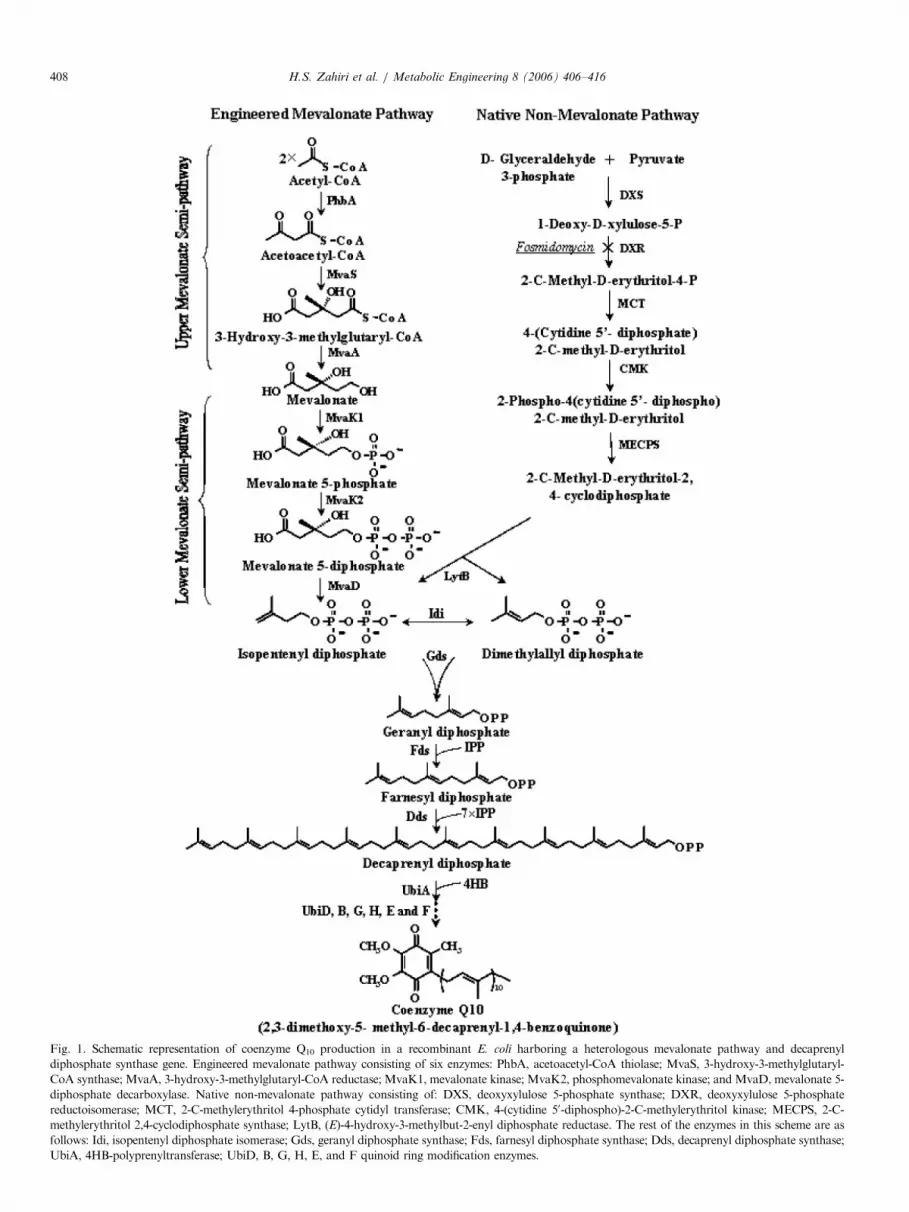
ARTICLE IN PRESS
Fig. 1. Schematic representation of coenzyme Q10 production in a recombinant E. coli harboring a heterologous mevalonate pathway and decaprenyl
diphosphate synthase gene. Engineered mevalonate pathway consisting of six enzymes: PhbA, acetoacetyl-CoA thiolase; MvaS, 3-hydroxy-3-methylglutaryl-
CoA synthase; MvaA, 3-hydroxy-3-methylglutaryl-CoA reductase; MvaK1, mevalonate kinase; MvaK2, phosphomevalonate kinase; and MvaD, mevalonate 5-
diphosphate decarboxylase. Native non-mevalonate pathway consisting of: DXS, deoxyxylulose 5-phosphate synthase; DXR, deoxyxylulose 5-phosphate
reductoisomerase; MCT, 2-C-methylerythritol 4-phosphate cytidyl transferase; CMK, 4-(cytidine 50-diphospho)-2-C-methylerythritol kinase; MECPS, 2-C-
methylerythritol 2,4-cyclodiphosphate synthase; LytB, (E)-4-hydroxy-3-methylbut-2-enyl diphosphate reductase. The rest of the enzymes in this scheme are as
follows: Idi, isopentenyl diphosphate isomerase; Gds, geranyl diphosphate synthase; Fds, farnesyl diphosphate synthase; Dds, decaprenyl diphosphate synthase;
UbiA, 4HB-polyprenyltransferase; UbiD, B, G, H, E, and F quinoid ring modification enzymes.
H.S. Zahiri et al. / Metabolic Engineering 8 (2006) 406–416408

ARTICLE IN PRESS
Fig. 2. Main plasmid constructs utilized for metabolic engineering of E. coli for CoQ10 production. (a) pTrc99A with ddsA gene (decaprenyl diphosphate
synthase) from A. tumefaciens. (b) pSTV28 vector with lower mevalonate semi-pathway operon consisting of mvaK1 (mevalonate kinase), mvaD
(mevalonate diphosphate decarboxylase), and mvaK2 (phosphomevalonate kinase) as well as idi (isopentenyl diphosphate isomerase). (c) pBBR1MCS2
vector with upper mevalonate semi-pathway operon consisting of mvaA (3-hydroxy-3-methylglutaryl-CoA reductase), mvaS (3-hydroxy-3-methylglutaryl-
CoA synthase), and phbA (acetoacetyl-CoA thiolase). (d) pSTV28 with whole mevalonate pathway as well as idi.
H.S. Zahiri et al. / Metabolic Engineering 8 (2006) 406–416 409
obtained by PCR amplification from pBSNmvaASP usingPfu DNA polymerase and oligonucleotides UMP-1 andUMP-2 (Table 1). The amplified fragment was cut withEcoRI and inserted into pSSN12Didi containing the lowermevalonate semi-pathway of Streptococcus pneumoniae
(Fig. 2d). The resulting plasmid, pSSNSN12Didi, con-tained a whole mevalonate pathway capable of convertingacetyl-CoA to IPP.
Fosmidomycin (sodium salt, Molecular Probes), aspecific inhibitor of the non-mevalonate pathway(Kuzuyama et al., 1998), was used to a final concentrationof 1mM for selection of recombinant clones carrying afunctional mevalonate pathway and a lower semi-pathwayfed with exogenous mevalonate (Kuzuyama and Seto,2003). The authenticity of the constructed plasmids wasfurther verified by a restriction map analysis and DNAsequencing. The molecular biology techniques were carriedout as described by Sambrook and Rusell (2001). The
plasmid constructs were used for the transformation ofE. coli by using an electroporator (Bio Rad, Gene Pulser).
2.4. Thin layer chromatography
Functional expression of the lower mevalonate semi-pathway and Dds was studied by using cell-free homo-genates of E. coli PTBS. In the case of lower mevalonatesemi-pathway enzymes, the cell-free homogenate was usedfor in vitro conversion of [14C] mevalonolactone toradiolabeled intermediate metabolites including mevalo-nate phosphate and mevalonate diphosphate. Two hun-dred microliters of potassium phosphate buffer (pH 8)containing 30 mM ATP, 90 mM KH2PO4, 5mM MgCl2,50mM KCl, 60 mM [14C] mevalonolactone (Amersham),and 300 mg of cytosolic proteins was incubated at 30 1C for2 h. The reaction products were spotted on a silica thinlayer chromatography (TLC) plate (Sigma) and developed

ARTICLE IN PRESS
Table 1
Primers used for the construction of plasmids
Designation Sequence
Agrobacterium tumefaciens; ddsA gene
atdds-1 50-GGAATTCTTGCCGCACAAGGCATCAG-30 (EcoRI)
atdds-2 50-CGGGATCCGCGTTAGTTGAGACGCTCGATG-30 (BamHI)
atdds-3 50-TCCCCCCGGGCACAGGAAACAGACCATGGAGTTCTTG-30 (XmaI)
atdds-4 50-TCCCCCCGGGTTAGTTGAGACGCTCGATGCAGAAG-30 (XmaI)
Saccharomyces cerevisiae; lower mevalonate semi-pathway genes
PMK-5 (ERG8) 50-CCCATGGAAGAGTTGAGAGCCTTCAGTG-30 (NcoI)
PMK3-3 (ERG8) 50-CGAATTCTGA GTTACATCAG CCATTGAACC-30 (EcoRI)
MPDC5-1 (ERG19) 50-GGAATTCAGGAGACAACACAATG-30 (EcoRI)
MPDC3-2 (ERG19) 50-GGAGCTCTTA TTCCTTTGG-30(SacI)
MK5-1 (ERG12) 50-CGAGCTC AGGAG ATATCAAT ATG-30 (SacI)
MK3-2 (ERG12) 50-GCCCGGGTTA TGAAGTCC-30 (SmaI)
Ralstonia eutropha; phbA
phbA-51 50-CGGGATCCAGGACTACACAATGACTGAC-30 (BamHI)
phbA-31 50-CGGGATCCTTATTTGCGCTCGACTGCC-30 (BamHI)
Streptococcus pneumoniae; mvaK1, mvaD and mvaK2
SN12D-5 50-CGAATTCAATGACAAAAAAAGTTGGTGTCGG-30 (EcoRI)
SN12D-3 50-TCCCGGGTTA CGATTTGTCG TCATGTCC-30 (XmaI)
Streptococcus pyogenes; mvaK1, mvaD and mvaK2
SY12D-5 50-CGGTACCAATGAACGAAAACATTGGATATGG-30(KpnI)
SY12D-3 50-TCCCGGGTTAGTCATTCTCT CCCCACC-30 (XmaI)
Staphylococcus aureus; mvaK1, mvaD and mvaK2
SA12D-5 50-AGAATTCACATGACAAGAAAAGGATATGGGG-30 (EcoRI )
SA12D-3 50-TCCCGGGCCC TATCAAATCA TTTATTGCCC-30 (XmaI)
Enterococcus faecalis; mvaK1, mvaD and mvaK2
EF12D-5 50-AGGTACCAAATGAATATAAAAAAACAAGGCC-30 (KpnI)
EF12D-3 50-GCCCGGGTCA TCTTTTCGAT TCATGCTTC-30 (XmaI)
Streptococcus pneumonia; mvaA and mvaS
SNSA-5 50-GGAATTCACATGAATGATAAAACAGAGG-30 (EcoRI)
SNSA-3 50-GGGATCCGATTCCTCTGAGTTTTTATGATC-30 (BamHI)
Escherichia coli; idi
idi-5 50-CCCCGGGAGGAGAGAAATTATGCAAACGGAAC30 (XmaI)
Idi-3 50-TGTCGAC TTATTTAAGCTGGGTAAATG-30 (SalI)
Amplification of upper mevalonate pathway using pBSNmvaASP as template
UMP-1 50-TCGAATTCAGGAGGATTCATAATGAAGATAAGTTG-30 (EcoRI)
UMP-2 50-TCGAATTCCTCCTTTATTTGCGCTCGACTGCCAG-30 (EcoRI)
H.S. Zahiri et al. / Metabolic Engineering 8 (2006) 406–416410
in ethanol, ammonia, and water (80:10:17). The relativepositions of intermediate metabolites of the lower meva-lonate semi-pathway were recognized by using [14C] IPP(Amersham) as standard.
To show the functional expression of Dds, the samereaction mixture was used with the exception that ATP andmevalonolactone were replaced with 0.05% Triton X-100,60 mM [14C] IPP, and 100 mM farnesyl diphosphate(Sigma). Polyprenyl diphosphates in the reaction mixturewere extracted with butanol, dephosphorylated, andspotted on a reverse-phase C18 TLC plate (Koyamaet al., 1985). The TLC plate was developed using acetone/water (19:1) as a mobile phase. Plates with radiolabeledproducts were exposed to an X-ray Fuji film and theradioactivity distribution was determined by autoradiogra-
phy. The relative position of decaprenol on the TLC platewas recognized using solanesol (Sigma) as a standard.
2.5. Determination of mevalonate
The mevalonate concentration in the culture mediumwas determined as mevalonolactone by gas chromatogra-phy (GC) analysis (Nozaki et al., 1996). The culturemedium was centrifuged at 12,000 rpm for 10min toseparate the cells. The supernatant was acidified by theaddition of HCl (1M) to pH 2, and ethyl acetate andanhydrous sodium sulfate were added. After completemixing, the solution was left standing overnight to ensurecomplete lactonization. The ethyl acetate extract was theninjected without derivatization into a GC apparatus

ARTICLE IN PRESS
Table 2
Strains and plasmids used in this study for CoQ10 production
Strain and plasmid Description Source
Strains
E. coli DH5a [F0/endA1 hsdR17 (rK� mK
+) glnV44 thi-1 recA1 gyrA Gibco BRL
(Nalr) relA1 D(lacIZYA-argF)U169 deoR 5(f80dlacD(lacZ)M15)]
E. coli PT E. coli DH5a harboring pTatdds This study
E. coli PTA E. coli PT harboring pSSA12Didi This study
E. coli PTC E. coli PT harboring pSSC12Didi This study
E. coli PTF E. coli PT harboring pSEF12Didi This study
E. coli PTN E. coli PT harboring pSSN12Didi This study
E. coli PTY E. coli PT harboring pSSY12Didi This study
E. coli PB E. coli DH5a harboring pBSNmvaASP This study
E. coli PBS E. coli PB harboring pSSN12Didi This study
E. coli PTBS E. coli PTN harboring pBSNmvaASP This study
E. coli PTSN E. coli PT harboring pSSNSN12Didi This study
Plasmids
pTrc99A Ptrc expression vector, pBR322 origin, lacIq, Ampr, high copy Amersham Biosci.
pSTV28 Plac cloning vector, pACYC184 origin, LacZa, Cmr, low copy Takara Shuzo Co., Ltd.
pBBR1MCS2 Plac cloning vector, pBBR1 origin, LacZa, broad-host-range, Kmr Kovach et al. (1995)
pTatdds pTrc99A with ddsA gene from A. tumefaciens This study
pSEF12Didi pSTV28 with mvaK1, mvaD and mvaK2 genes from E. faecalis and idi from E. coli This study
pSSA12Didi pSTV28 with mvaK1, mvaD and mvaK2 genes from S. aureus and idi from E. coli This study
pSSC12Didi pSTV28 with mvaK1, mvaD and mvaK2 genes from S. cerevisiae and idi from E. coli This study
pSSN12Didi pSTV28 with mvaK1, mvaD and mvaK2 genes from S. pneumoniae and idi from E. coli This study
pSSY12Didi pSTV28 with mvaK1, mvaD and mvaK2 genes from S. pyogenes and idi from E. coli This study
pSSNSN12Didi pSTV28 with mvaA, mvaS, mvaK1, mvaD and mvaK2 This study
genes from S. pneumoniae; phbA from R. eutropha and idi from E. coli
pBSNmvaASP pBBR1MCS2 with mvaA and mvaS from S. pneumoniae and phbA from R. eutropha This study
H.S. Zahiri et al. / Metabolic Engineering 8 (2006) 406–416 411
equipped with a DB-1 column (0.53mm ID� 30m, 5 mmfilm; J&W Scientific). The temperatures at the injectionport and the detector were 240 and 250 1C, respectively.The column temperature was maintained at 60 1C for thefirst 2min after the injection of a sample and then raised to190 1C at a constant rate of 8 1C/min. Authentic mevalo-nolactone (Sigma) was used as an internal standard.
2.6. Coenzyme Q10 assay
The amounts of CoQ10 produced by various recombi-nant E. coli strains were determined via high-performanceliquid chromatography (HPLC). CoQ10 was extracted fromE. coli cells with hexane as follows. A 500 ml sample of eachculture was centrifuged at 14,000 rpm using a bench-topcentrifuge. The pellet was washed once with 1ml ofdistilled water and then with 1ml of 20mM Tris-HCl, pH7.6. The washed cells were lysed by adding 450 ml of CellLytic B solution (Sigma) and 50 ml of egg white lysozymestock solution (10mg/ml, Amresco). After incubation at37 1C for 30min, CoQ10 was extracted from the cell lysatewith 900 ml of a hexane/propanol (5:3) mixture and thenwith 500 ml of hexane. Following each extraction, thehexane organic phase containing CoQ10 was separatedfrom the aqueous phase by centrifugation at 14,000 rpmand collected into a new tube. The hexane extract wasevaporated to dryness by overnight incubation in a vacuumchamber at room temperature. The remaining pellet was
dissolved in 500 ml of absolute ethanol (Fisher Scientific,HPLC grade) from which 10 ml was injected into a HPLCmachine (Shimadzu 10A system) equipped with a Symme-trys C18 column (Waters). The chromatography was runat room temperature using an isocratic solvent mixture ofethanol and methanol (70:30 v/v, Fisher Scientific, HPLCgrade) as a mobile phase at a flow rate of 1ml/min. A UVdetector was used at 275 nm for the detection andquantification of CoQ10. Authentic standards, CoQ9 andCoQ10 (Sigma), were used to distinguish the correspondingpeaks in the HPLC chromatograms of the experimentalsamples. A standard curve was made for quantification ofCoQ10 by making serial dilutions of a CoQ10 standardstock solution (1mg/ml). There was a reproducible highcorrelation (r240:999) between CoQ10 concentrations andpeak areas.
3. Results
3.1. Coenzyme Q10 production by recombinant E. coli
harboring pTatdds
E. coli DH5a was transformed into a CoQ10-producingorganism (E. coli PT) by the introduction of pTatdds(Fig. 2a). Recombinant E. coli PT was cultured in 2YTmedium supplemented with 0.5% glycerol (2YTG medium)at 30 1C and 200 rpm shaking for 24 h. As a result of theddsA gene expression, E. coli PT began to produce CoQ10
ARTICLE IN PRESS
0
500
1000
1500
2000
2500
3000
Mevalonate concentration(mM)
0 1
Cel
lmas
s(g/
l)C
oQ10
con
tent
(µg
/gD
CW
)
0
1
2
3
4
5
2 3 4
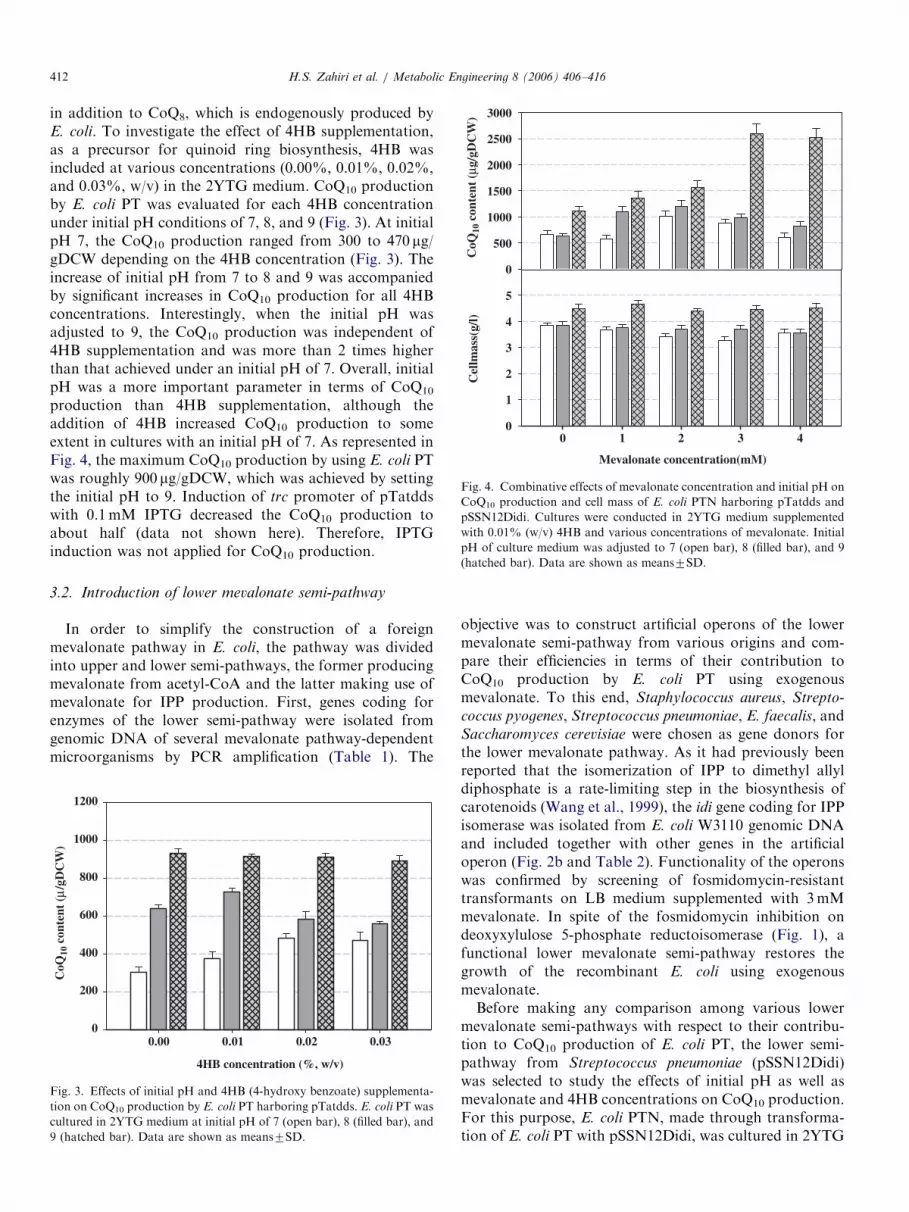
Fig. 4. Combinative effects of mevalonate concentration and initial pH on
CoQ10 production and cell mass of E. coli PTN harboring pTatdds and
pSSN12Didi. Cultures were conducted in 2YTG medium supplemented
with 0.01% (w/v) 4HB and various concentrations of mevalonate. Initial
pH of culture medium was adjusted to 7 (open bar), 8 (filled bar), and 9
(hatched bar). Data are shown as means7SD.
H.S. Zahiri et al. / Metabolic Engineering 8 (2006) 406–416412
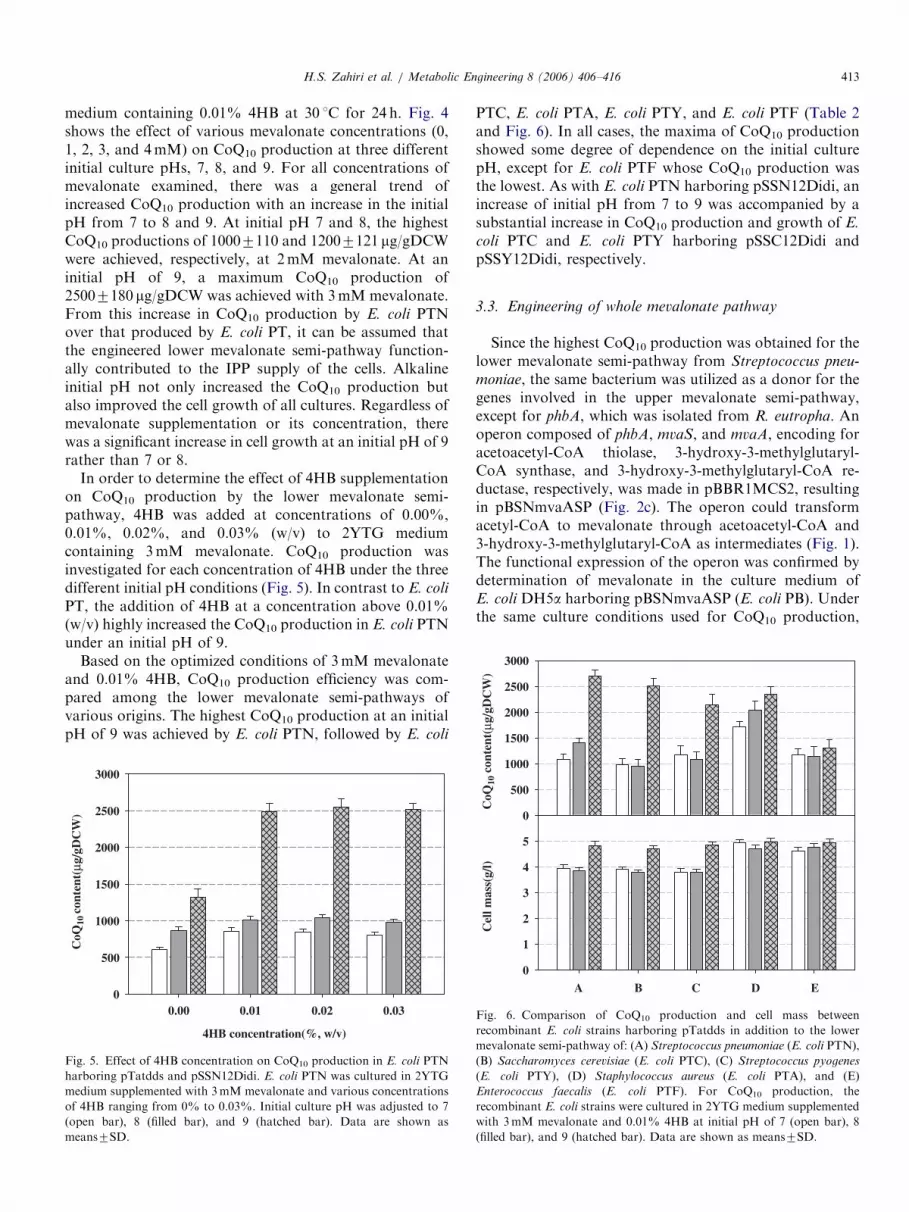
in addition to CoQ8, which is endogenously produced byE. coli. To investigate the effect of 4HB supplementation,as a precursor for quinoid ring biosynthesis, 4HB wasincluded at various concentrations (0.00%, 0.01%, 0.02%,and 0.03%, w/v) in the 2YTG medium. CoQ10 productionby E. coli PT was evaluated for each 4HB concentrationunder initial pH conditions of 7, 8, and 9 (Fig. 3). At initialpH 7, the CoQ10 production ranged from 300 to 470 mg/gDCW depending on the 4HB concentration (Fig. 3). Theincrease of initial pH from 7 to 8 and 9 was accompaniedby significant increases in CoQ10 production for all 4HBconcentrations. Interestingly, when the initial pH wasadjusted to 9, the CoQ10 production was independent of4HB supplementation and was more than 2 times higherthan that achieved under an initial pH of 7. Overall, initialpH was a more important parameter in terms of CoQ10
production than 4HB supplementation, although theaddition of 4HB increased CoQ10 production to someextent in cultures with an initial pH of 7. As represented inFig. 4, the maximum CoQ10 production by using E. coli PTwas roughly 900 mg/gDCW, which was achieved by settingthe initial pH to 9. Induction of trc promoter of pTatddswith 0.1mM IPTG decreased the CoQ10 production toabout half (data not shown here). Therefore, IPTGinduction was not applied for CoQ10 production.
3.2. Introduction of lower mevalonate semi-pathway
In order to simplify the construction of a foreignmevalonate pathway in E. coli, the pathway was dividedinto upper and lower semi-pathways, the former producingmevalonate from acetyl-CoA and the latter making use ofmevalonate for IPP production. First, genes coding forenzymes of the lower semi-pathway were isolated fromgenomic DNA of several mevalonate pathway-dependentmicroorganisms by PCR amplification (Table 1). The
4HB concentration (%, w/v)
0.00 0.01 0.02 0.03
CoQ
10 c
onte
nt (
µ/g
DC
W)
0
200
400
600
800
1000
1200
Fig. 3. Effects of initial pH and 4HB (4-hydroxy benzoate) supplementa-
tion on CoQ10 production by E. coli PT harboring pTatdds. E. coli PT was
cultured in 2YTG medium at initial pH of 7 (open bar), 8 (filled bar), and
9 (hatched bar). Data are shown as means7SD.
objective was to construct artificial operons of the lowermevalonate semi-pathway from various origins and com-pare their efficiencies in terms of their contribution toCoQ10 production by E. coli PT using exogenousmevalonate. To this end, Staphylococcus aureus, Strepto-
coccus pyogenes, Streptococcus pneumoniae, E. faecalis, andSaccharomyces cerevisiae were chosen as gene donors forthe lower mevalonate pathway. As it had previously beenreported that the isomerization of IPP to dimethyl allyldiphosphate is a rate-limiting step in the biosynthesis ofcarotenoids (Wang et al., 1999), the idi gene coding for IPPisomerase was isolated from E. coli W3110 genomic DNAand included together with other genes in the artificialoperon (Fig. 2b and Table 2). Functionality of the operonswas confirmed by screening of fosmidomycin-resistanttransformants on LB medium supplemented with 3mMmevalonate. In spite of the fosmidomycin inhibition ondeoxyxylulose 5-phosphate reductoisomerase (Fig. 1), afunctional lower mevalonate semi-pathway restores thegrowth of the recombinant E. coli using exogenousmevalonate.Before making any comparison among various lower
mevalonate semi-pathways with respect to their contribu-tion to CoQ10 production of E. coli PT, the lower semi-pathway from Streptococcus pneumoniae (pSSN12Didi)was selected to study the effects of initial pH as well asmevalonate and 4HB concentrations on CoQ10 production.For this purpose, E. coli PTN, made through transforma-tion of E. coli PT with pSSN12Didi, was cultured in 2YTG
ARTICLE IN PRESS
nt(µ
g/gD
CW
)
1500
2000
2500
3000
H.S. Zahiri et al. / Metabolic Engineering 8 (2006) 406–416 413
medium containing 0.01% 4HB at 30 1C for 24 h. Fig. 4shows the effect of various mevalonate concentrations (0,1, 2, 3, and 4mM) on CoQ10 production at three differentinitial culture pHs, 7, 8, and 9. For all concentrations ofmevalonate examined, there was a general trend ofincreased CoQ10 production with an increase in the initialpH from 7 to 8 and 9. At initial pH 7 and 8, the highestCoQ10 productions of 10007110 and 12007121 mg/gDCWwere achieved, respectively, at 2mM mevalonate. At aninitial pH of 9, a maximum CoQ10 production of25007180 mg/gDCW was achieved with 3mM mevalonate.From this increase in CoQ10 production by E. coli PTNover that produced by E. coli PT, it can be assumed thatthe engineered lower mevalonate semi-pathway function-ally contributed to the IPP supply of the cells. Alkalineinitial pH not only increased the CoQ10 production butalso improved the cell growth of all cultures. Regardless ofmevalonate supplementation or its concentration, therewas a significant increase in cell growth at an initial pH of 9rather than 7 or 8.
In order to determine the effect of 4HB supplementationon CoQ10 production by the lower mevalonate semi-pathway, 4HB was added at concentrations of 0.00%,0.01%, 0.02%, and 0.03% (w/v) to 2YTG mediumcontaining 3mM mevalonate. CoQ10 production wasinvestigated for each concentration of 4HB under the threedifferent initial pH conditions (Fig. 5). In contrast to E. coli
PT, the addition of 4HB at a concentration above 0.01%(w/v) highly increased the CoQ10 production in E. coli PTNunder an initial pH of 9.
Based on the optimized conditions of 3mM mevalonateand 0.01% 4HB, CoQ10 production efficiency was com-pared among the lower mevalonate semi-pathways ofvarious origins. The highest CoQ10 production at an initialpH of 9 was achieved by E. coli PTN, followed by E. coli
4HB concentration(%, w/v)
0.00 0.01 0.02 0.03
CoQ
10 c
onte
nt(µ
g/gD
CW
)
0
500
1000
1500
2000
2500
3000
Fig. 5. Effect of 4HB concentration on CoQ10 production in E. coli PTN
harboring pTatdds and pSSN12Didi. E. coli PTN was cultured in 2YTG
medium supplemented with 3mM mevalonate and various concentrations
of 4HB ranging from 0% to 0.03%. Initial culture pH was adjusted to 7
(open bar), 8 (filled bar), and 9 (hatched bar). Data are shown as
means7SD.
PTC, E. coli PTA, E. coli PTY, and E. coli PTF (Table 2and Fig. 6). In all cases, the maxima of CoQ10 productionshowed some degree of dependence on the initial culturepH, except for E. coli PTF whose CoQ10 production wasthe lowest. As with E. coli PTN harboring pSSN12Didi, anincrease of initial pH from 7 to 9 was accompanied by asubstantial increase in CoQ10 production and growth of E.
coli PTC and E. coli PTY harboring pSSC12Didi andpSSY12Didi, respectively.
3.3. Engineering of whole mevalonate pathway
Since the highest CoQ10 production was obtained for thelower mevalonate semi-pathway from Streptococcus pneu-
moniae, the same bacterium was utilized as a donor for thegenes involved in the upper mevalonate semi-pathway,except for phbA, which was isolated from R. eutropha. Anoperon composed of phbA, mvaS, and mvaA, encoding foracetoacetyl-CoA thiolase, 3-hydroxy-3-methylglutaryl-CoA synthase, and 3-hydroxy-3-methylglutaryl-CoA re-ductase, respectively, was made in pBBR1MCS2, resultingin pBSNmvaASP (Fig. 2c). The operon could transformacetyl-CoA to mevalonate through acetoacetyl-CoA and3-hydroxy-3-methylglutaryl-CoA as intermediates (Fig. 1).The functional expression of the operon was confirmed bydetermination of mevalonate in the culture medium ofE. coli DH5a harboring pBSNmvaASP (E. coli PB). Underthe same culture conditions used for CoQ10 production,
CoQ
10 c
onte
0
500
1000
A C E
Cel
l mas
s(g/
l)
0
1
2
3
4
5
B D
Fig. 6. Comparison of CoQ10 production and cell mass between
recombinant E. coli strains harboring pTatdds in addition to the lower
mevalonate semi-pathway of: (A) Streptococcus pneumoniae (E. coli PTN),
(B) Saccharomyces cerevisiae (E. coli PTC), (C) Streptococcus pyogenes
(E. coli PTY), (D) Staphylococcus aureus (E. coli PTA), and (E)
Enterococcus faecalis (E. coli PTF). For CoQ10 production, the
recombinant E. coli strains were cultured in 2YTG medium supplemented
with 3mM mevalonate and 0.01% 4HB at initial pH of 7 (open bar), 8
(filled bar), and 9 (hatched bar). Data are shown as means7SD.
ARTICLE IN PRESS
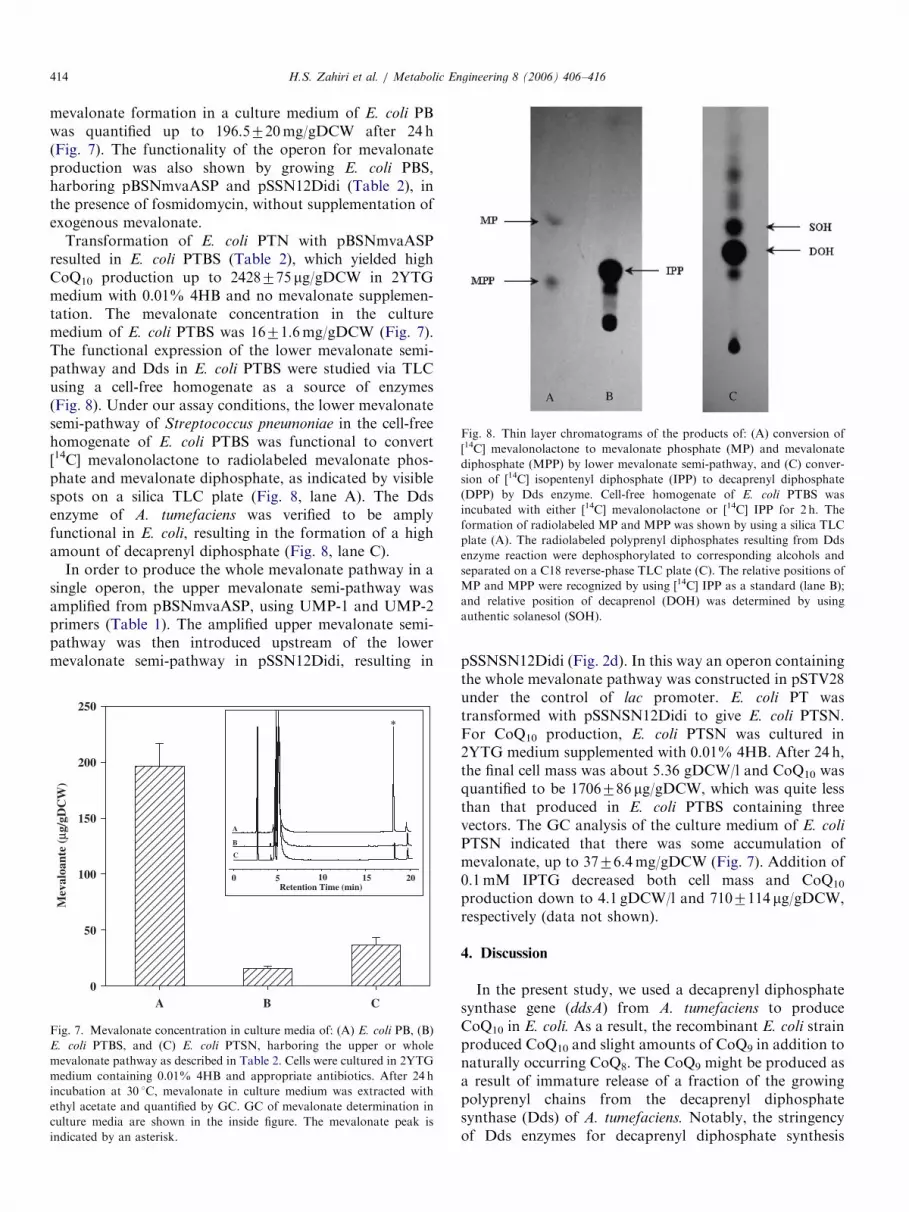
Fig. 8. Thin layer chromatograms of the products of: (A) conversion of
[14C] mevalonolactone to mevalonate phosphate (MP) and mevalonate
diphosphate (MPP) by lower mevalonate semi-pathway, and (C) conver-
sion of [14C] isopentenyl diphosphate (IPP) to decaprenyl diphosphate
(DPP) by Dds enzyme. Cell-free homogenate of E. coli PTBS was
incubated with either [14C] mevalonolactone or [14C] IPP for 2 h. The
formation of radiolabeled MP and MPP was shown by using a silica TLC
plate (A). The radiolabeled polyprenyl diphosphates resulting from Dds
enzyme reaction were dephosphorylated to corresponding alcohols and
separated on a C18 reverse-phase TLC plate (C). The relative positions of
MP and MPP were recognized by using [14C] IPP as a standard (lane B);
and relative position of decaprenol (DOH) was determined by using
authentic solanesol (SOH).
H.S. Zahiri et al. / Metabolic Engineering 8 (2006) 406–416414
mevalonate formation in a culture medium of E. coli PBwas quantified up to 196.5720mg/gDCW after 24 h(Fig. 7). The functionality of the operon for mevalonateproduction was also shown by growing E. coli PBS,harboring pBSNmvaASP and pSSN12Didi (Table 2), inthe presence of fosmidomycin, without supplementation ofexogenous mevalonate.
Transformation of E. coli PTN with pBSNmvaASPresulted in E. coli PTBS (Table 2), which yielded highCoQ10 production up to 2428775 mg/gDCW in 2YTGmedium with 0.01% 4HB and no mevalonate supplemen-tation. The mevalonate concentration in the culturemedium of E. coli PTBS was 1671.6mg/gDCW (Fig. 7).The functional expression of the lower mevalonate semi-pathway and Dds in E. coli PTBS were studied via TLCusing a cell-free homogenate as a source of enzymes(Fig. 8). Under our assay conditions, the lower mevalonatesemi-pathway of Streptococcus pneumoniae in the cell-freehomogenate of E. coli PTBS was functional to convert[14C] mevalonolactone to radiolabeled mevalonate phos-phate and mevalonate diphosphate, as indicated by visiblespots on a silica TLC plate (Fig. 8, lane A). The Ddsenzyme of A. tumefaciens was verified to be amplyfunctional in E. coli, resulting in the formation of a highamount of decaprenyl diphosphate (Fig. 8, lane C).
In order to produce the whole mevalonate pathway in asingle operon, the upper mevalonate semi-pathway wasamplified from pBSNmvaASP, using UMP-1 and UMP-2primers (Table 1). The amplified upper mevalonate semi-pathway was then introduced upstream of the lowermevalonate semi-pathway in pSSN12Didi, resulting in
A B C
Mev
aloa
nte
(µg/
gDC
W)
0
50
100
150
200
250
20151050Retention Time (min)
A
B
C
Fig. 7. Mevalonate concentration in culture media of: (A) E. coli PB, (B)
E. coli PTBS, and (C) E. coli PTSN, harboring the upper or whole
mevalonate pathway as described in Table 2. Cells were cultured in 2YTG
medium containing 0.01% 4HB and appropriate antibiotics. After 24 h
incubation at 30 1C, mevalonate in culture medium was extracted with
ethyl acetate and quantified by GC. GC of mevalonate determination in
culture media are shown in the inside figure. The mevalonate peak is
indicated by an asterisk.
pSSNSN12Didi (Fig. 2d). In this way an operon containingthe whole mevalonate pathway was constructed in pSTV28under the control of lac promoter. E. coli PT wastransformed with pSSNSN12Didi to give E. coli PTSN.For CoQ10 production, E. coli PTSN was cultured in2YTG medium supplemented with 0.01% 4HB. After 24 h,the final cell mass was about 5.36 gDCW/l and CoQ10 wasquantified to be 1706786 mg/gDCW, which was quite lessthan that produced in E. coli PTBS containing threevectors. The GC analysis of the culture medium of E. coli
PTSN indicated that there was some accumulation ofmevalonate, up to 3776.4mg/gDCW (Fig. 7). Addition of0.1mM IPTG decreased both cell mass and CoQ10
production down to 4.1 gDCW/l and 7107114 mg/gDCW,respectively (data not shown).
4. Discussion
In the present study, we used a decaprenyl diphosphatesynthase gene (ddsA) from A. tumefaciens to produceCoQ10 in E. coli. As a result, the recombinant E. coli strainproduced CoQ10 and slight amounts of CoQ9 in addition tonaturally occurring CoQ8. The CoQ9 might be produced asa result of immature release of a fraction of the growingpolyprenyl chains from the decaprenyl diphosphatesynthase (Dds) of A. tumefaciens. Notably, the stringencyof Dds enzymes for decaprenyl diphosphate synthesis

ARTICLE IN PRESSH.S. Zahiri et al. / Metabolic Engineering 8 (2006) 406–416 415
varies among enzymes isolated from various organisms(Saiki et al., 2003; Takahashi et al., 2003).
In our study, pH was found to play a major role withrespect to obtaining high CoQ10 production. The use ofinitial alkaline pH for cultures greatly improved CoQ10
production and the growth of recombinant E. coli strains.Alkaline pH improved CoQ10 production in E. coli PTharboring only ddsA gene as well as recombinant E. coli
strains harboring a heterologous mevalonate pathway inaddition to ddsA. This indicates that the effect of alkalinepH on CoQ10 production primarily has no relation with theheterologous mevalonate pathway. As ubiquinones aremolecules of redox capacity, it is expected that theiraccumulation in biological membranes will affect thephysiology of cells. It appears that the role of alkalinepH is related to the functions of CoQ10 in membrane.However, further study is necessary to reveal the exact roleof pH in CoQ10 production.
It has been shown that an insufficient supply of IPP bythe non-mevalonate pathway limits the biosynthesis ofIPP-derived products in E. coli (Sandmann, 2002; Huanget al., 2001). There have been several reports on increasedcarotenoid production in E. coli through metabolicengineering of non-mevalonate pathways (Matthews andWurtzel, 2000; Harker and Bramley, 1999; Farmer andLiao, 2000; Kim and Keasling, 2001; Alper et al., 2005).Instead of metabolic engineering of a non-mevalonatepathway, we utilized a foreign mevalonate pathway toincrease the IPP supply in the CoQ10-producing E. coli
strains. Mevalonate is a key intermediate of the mevalonatepathway and is not produced or consumed by wild-typeE. coli (Tabata and Hashimoto, 2004). Therefore, thefunctional expression of the foreign mevalonate pathwaywas confirmed by determination of mevalonate in culturemedia of several recombinant E. coli strains. It was shownthat E. coli PB, harboring the upper mevalonate semi-pathway, produced a high amount of mevalonate (Fig. 7).Interestingly, in E. coli PTBS containing the lowermevalonate semi-pathway and Dds enzyme in addition tothe upper mevalonate semi-pathway, mevalonate produc-tion severely decreased (Fig. 7). It seems reasonable topresume that the mevalonate production by the uppermevalonate semi-pathway is efficiently channeled towardCoQ10 production in E. coli PTBS. This assumption issupported by the observation of elevated CoQ10 produc-tion in the strain compared with E. coli PT. In the case ofE. coli PTSN, which contains the whole mevalonatepathway under the control of a lac promoter in a singleplasmid, the rate of mevalonate production appears to behigher than its consumption for IPP biosynthesis (Fig. 7).As a result, higher amounts of mevalonate were detected inthe culture medium and net CoQ10 production was less thanthat produced in E. coli PTBS. Using TLC, it was confirmedthat the lower mevalonate semi-pathway of Streptococcus
pneumoniae was functionally expressed in E. coli.
To the best of our knowledge, this is the first report onmetabolic engineering of a foreign mevalonate pathway for
enhancing CoQ10 production in E. coli. We demonstratedthat the IPP shortage is a primary drawback in CoQ10
production by E. coli, and this problem could besignificantly diminished via an additional IPP providingpathway. Through the improvement of IPP supply andoptimization of pH, CoQ10 production increased consider-ably to approximately 2.4mg/gDCW, a value unprece-dented in E. coli. This level of CoQ10 production, which is2.7 times higher than obtained with ddsA gene alone,resulted from leaky expression of the mevalonate pathwaywithout any induction. It should be taken into account thataccumulation of IPP or intermediate metabolites of theforeign mevalonate pathway, which may arise uponinduction, potentially inhibits general metabolism as wellas the pathway itself (Andreassi et al., 2004; Martin et al.,2003).However, the efficiency of the mevalonate pathway in
improving the biotechnological production of CoQ10 andother isoprenoid compounds appears to be significantlyaffected by the extent of the physiological impact arisingfrom accumulation of the product. CoQ10 directly affectsthe metabolism and survival of the host cells due to its rolein the respiratory chain and other redox reactions. There-fore, the host cell reaction towards CoQ10 accumulation isexpected to be vastly different from that to other IPP-derived products.
Acknowledgments
This work was supported by 21C Frontier MicrobialGenomics and Applications Program of the KoreanMinistry of Science and Technology. H.S. Zahiri is gratefulfor a scholarship received from KOSEF.
References
Albrecht, M., Misawa, N., Sandmann, G., 1999. Metabolic engineering of
the terpenoid biosynthetic pathway of E. coli for production of the
carotenoids b-carotene and zeaxanthin. Biotechnol. Lett. 21, 791–795.
Alper, H., Miyaoku, K., Stephanopoulos, G., 2005. Construction of
lycopene-overproducing E. coli strains by combining systematic and
combinatorial gene knockout targets. Nat. Biotechnol. 23, 612–616.
Andreassi, J.L., Dabovic, K., Leyh, T.S., 2004. Streptococcus pneumoniae
isoprenoid biosynthesis is downregulated by diphosphomevalonate: an
antimicrobial target. Biochemistry 43, 16461–16466.
Bader, M., Muse, W., Ballou, D.P., Gassner, C., Bardwell, J.C.A., 1999.
Oxidative protein folding is driven by the electron transport system.
Cell 98, 217–227.
Battino, M., Ferri, E., Gorini, A., Federico, V.R., Rodriguez, H.J.F.,
Fiorella, P., Genova, M.L., Lenaz, G., Marchetti, M., 1990. Natural
distribution and occurrence of coenzyme Q. Membr. Biochem. 9,
179–190.
Bhagavan, H.N., Chopra, R.K., 2005. Potential role of ubiquinone
(coenzyme Q10) in pediatric cardiomyopathy. Clin. Nutr. 24, 331–338.
Crane, F.L., 2001. Biochemical functions of coenzyme Q10. J. Am. Coll.
Nutr. 20, 591–598.
Farmer, W.R., Liao, J.C., 2000. Improving lycopene production in E. coli
by engineering metabolic control. Nat. Biotechnol. 18, 533–537.
Georgellis, D., Kwon, O., Lin, E.C., 2001. Quinones as the redox signal
for the arc two-component system of bacteria. Science 292, 2314–2316.

ARTICLE IN PRESSH.S. Zahiri et al. / Metabolic Engineering 8 (2006) 406–416416
Groneberg, D.A., Kindermann, B., Althammer, M., Klapper, M., Vormann,
J., Littarrue, G.P., DOring, F., 2005. Coenzyme Q10 affects expression of
genes involved in cell signaling, metabolism and transport in human
CaCo-2 cells. Int. J. Biochem. Cell. Biol. 37, 1208–1218.
Harker, M., Bramley, P.M., 1999. Expression of prokaryotic 1-deoxy-D-
xylulose-5-phosphatases in E. coli increases carotenoid and ubiquinone
biosynthesis. FEBS Lett. 448, 115–119.
Huang, Q.L., Roessner, C.A., Croteau, R., Scotta, A.I., 2001. Engineering
E. coli for the synthesis of taxadiene, a key intermediate in the
biosynthesis of taxol. Bioorgan. Med. Chem. 9, 2237–2242.
Kawamukai, M., 2002. Biosynthesis, bioproduction and novel roles of
ubiquinone. J. Biosci. Bioeng. 94, 511–517.
Kim, C.M., Goldstein, J.L., Brown, M.S., 1992. cDNA cloning of MEV, a
mutant protein that facilitates cellular uptake of mevalonate, and
identification of the point mutation responsible for its gain of function.
J. Biol. Chem. 297, 23113–23121.
Kim, S.W., Keasling, J.D., 2001. Metabolic engineering of non-
mevalonate isopentenyl diphosphate synthesis pathway in E. coli
enhances lycopene production. Biotechnol. Bioeng. 72, 408–415.
Kovach, M.E., Elzer, P.H., Hill, D.S., Robertson, G.T., Farris, M.A.,
Roop, R.M., Peterson, K.M., 1995. Four new derivatives of the broad-
host-range cloning vector pBBR1MCS, carrying different antibiotic-
resistance cassettes. Gene 166, 175–176.
Koyama, T., Fujii, H., Ogura, K., 1985. Enzymatic hydrolysis of
polyprenyl pyrophosphates. Methods Enzymol. 110, 153–155.
Kuzuyama, T., Seto, H., 2003. Diversity of the biosynthesis of the
isoprene units. Nat. Prod. Rep. 20, 171–183.
Kuzuyama, T., Shimizu, T., Takahashi, S., Seto, H., 1998. Fosmidomycin,
a specific inhibitor of 1-deoxy-D-xylulose 5-phosphate reductoisome-
rase in the nonmevalonate pathway for terpenoid biosynthesis.
Tetrahedron Lett. 39, 7913–7916.
Lipshutz, B.H., Mollard, P., Pfeiffer, S.S., Chrisman, W., 2002. A short, highly
efficient synthesis of coenzyme Q10. J. Am. Chem. Soc. 124, 14282–14283.
Martin, V.J.J., Pitera, D.J., Withers, S.T., Newman, J.D., Keasling, J.D.,
2003. Engineering a mevalonate pathway in E. coli for production of
terpenoids. Nat. Biotechnol. 21, 796–802.
Matthews, P.D., Wurtzel, E.T., 2000. Metabolic engineering of carotenoid
accumulation in E. coli by modulation of the isoprenoid precursor
pool with expression of deoxyxylulose phosphate synthase. Appl.
Microbiol. Biotechnol. 53, 396–400.
Matthews, R.T., Yang, L., Browne, S., Baik, M., Beal, M.F., 1998.
Coenzyme Q10 administration increases brain mitochondrial concen-
trations and exerts neuroprotective effects. Proc. Natl. Acad. Sci. 95,
8892–8897.
Meganathan, R., 2001. Ubiquinone biosynthesis in microorganisms.
FEMS Microbiol. Lett. 203, 131–139.
Negishi, E., Liou, S.Y., Xu, C., Huo, S., 2002. A novel, highly selective,
and general methodology for the synthesis of 1,5-diene-containing
oligoisoprenoids of all possible geometrical combinations exemplified
by an iterative and convergent synthesis of coenzyme Q10. Org. Lett. 4,
261–264.
Nozaki, S., Nakagawa, T., Nakata, A., Yamashita, S., Kameda-
Takemura, K., Nakamura, T., Keno, Y., Tokunaga, K., Matsuzawa,
Y., 1996. Effects of pravastatin on plasma and urinary mevalonate
concentrations in subjects with familial hypercholesterolaemia: a
comparison of morning and evening administration. Eur. J. Clin.
Pharmacol. 49, 361–364.
Okada, K., Suzuki, K., Kamiya, Y., Zhu, X., Fujisaki, S., Nishimura, Y.,
Nishino, T., Nakagawa, T., Kawamukai, M., Matsuda, H., 1996.
Polyprenyl diphosphate synthase essentially defines the length of the
side chain of ubiquinone. Biochim. Biophys. Acta 1302, 217–223.
Park, Y.C., Kim, S.J., Choi, J.H., Lee, W.H., Park, K.M., Kawamukai,
M., Ryu, Y.W., Seo, J.H., 2004. Batch and fed-batch production of
coenzyme Q10 in recombinant E. coli containing the decaprenyl
diphosphate synthase gene from Gluconobacter suboxyduns. Appl.
Microbiol. Biotechnol. 67, 192–196.
Rosenfeldt, F., Marasco, S., Lyon, W., Wowk, M., Sheeran, F., Bailey,
M., Esmore, D., Davis, B., Pick, A., Rabinov, A., Smith, J., Nagley,
P., Pepe, S., 2005. Coenzyme Q10 therapy before cardiac surgery
improves mitochondrial function and in vitro contractility of
myocardial tissue. J. Thorac. Cardiovasc. Surg. 129, 25–32.
Saiki, R., Nagata, A., Uchida, N., Kainow, T., Matsuda, H., Kawamukai,
M., 2003. Fission yeast decaprenyl diphosphate synthase consists of
Dps1 and the newly characterized Dlp1 protein in a novel hetero-
tetrameric structure. Eur. J. Biochem. 270, 4113–4121.
Sambrook, J., Rusell, D.W., 2001. Molecular Cloning: A Laboratory
Manual, third ed. Cold Spring Harbor Laboratory Press,, Cold Spring
Harbor, New York, USA.
Sandmann, G., 2002. Combinatorial biosynthesis of carotenoids in a
heterologous host: a powerful approach for the biosynthesis of novel
structures. ChemBioChem 3, 629–635.
Seballe, B., Poole, R.K., 1999. Ubiquinone limits oxidative stress in
E. coli. Microbiology 145, 1817–1830.
Sharma, S., Kheradpezhou, M., Shavali, S., El Refaey, H., Eken, J.,
Hagen, C., Ebadi, M., 2004. Neuroprotective actions of coenzyme Q10
in Parkinson’s disease. Methods Enzymol. 382, 488–509.
Somayajulu, M., McCarthy, S., Hung, M., Sikorska, M., Borowy-
Borowski, H., Pandeya, S., 2005. Role of mitochondria in neuronal
cell death induced by oxidative stress; neuroprotection by coenzyme
Q10. Neurobiol. Dis. 18, 618–627.
Tabata, K., Hashimoto, S.I., 2004. Production of mevalonate by a
metabolically-engineered E. coli. Biotechnol. Lett. 26, 1487–1491.
Takahashi, S., Nishino, T., Koyama, T., 2003. Isolation and expression of
Paracoccus denitrificans decaprenyl diphosphate synthase gene for
production of ubiquinone-10 in E. coli. Biochem. Eng. J. 16,
183–190.
Tran, M.T., Pharm, D., Mitchell, T.M., Kennedy, D.T., Giles, J.T., 2001.
Role of coenzyme Q10 in chronic heart failure, angina, and
hypertension. Pharmacotherapy 21, 797–806.
Turunen, M., Olsson, J., Dallner, G., 2004. Metabolism and function of
coenzyme Q. Biochim. Biophys. Acta 1660, 171–199.
Wang, C.W., Oh, M.K., Liao, J.C., 1999. Engineered isoprenoid pathway
enhances astaxanthin production in E. coli. Biotechnol. Bioeng. 62,
235–241.
Yoshida, H., Katani, Y., Ochiai, K., Araki, K., 1998. Production of
ubiquinone-10 using bacteria. J. Gen. Appl. Microbiol. 44, 19–26.