chromosomal translocation mechanisms at intronic alu elements in mammalian cells
TRANSCRIPT

Molecular Cell, Vol. 17, 885–894, March 18, 2005, Copyright ©2005 by Elsevier Inc. DOI 10.1016/j.molcel.2005.02.028
TechniqueChromosomal TranslocationMechanisms at Intronic AluElements in Mammalian Cells
Beth Elliott,1 Christine Richardson,1,2
and Maria Jasin1,*1Molecular Biology ProgramMemorial Sloan-Kettering Cancer Center1275 York AvenueNew York, New York 10021
Summary
Repetitive elements comprise nearly half of the hu-man genome. Chromosomal rearrangements involv-ing these elements occur in somatic and germlinecells and are causative for many diseases. To beginto understand the molecular mechanisms leading tothese rearrangements in mammalian cells, we devel-oped an intron-based system to specifically inducechromosomal translocations at Alu elements, themost numerous family of repetitive elements in hu-mans. With this system, we found that when double-strand breaks (DSBs) were introduced adjacent toidentical Alu elements, translocations occurred athigh frequency and predominantly arose from repairby the single-strand annealing (SSA) pathway (85%).With diverged Alu elements, translocation frequencywas unaltered, yet pathway usage shifted such thatnonhomologous end joining (NHEJ) predominated asthe translocation pathway (93%). These results em-phasize the fluidity of mammalian DSB repair path-way usage. The intron-based system is highly adapt-able to addressing a number of issues regardingmolecular mechanisms of genomic rearrangementsin mammalian cells.
Introduction
Genomic rearrangements are characteristic of tumorcells, and specific genomic rearrangements are respon-sible for many inherited diseases, yet genetic systemsto study their etiology at the molecular level have beenlimited. Repetitive elements, which comprise at least45% the human genome (Lander et al., 2001), presentample opportunity for genomic rearrangements (Dei-ninger et al., 2003). Alu elements make up the largestfamily of repetitive elements, numbering approximatelyone million copies and comprising an estimated 11%of the genome (Lander et al., 2001).
In germline cells, Alu-Alu intrachromosomal recombi-nation has been implicated in the etiology of severalinherited diseases, including some cancers (Deiningerand Batzer, 1999; Kolomietz et al., 2002). In somaticcells, recombination between Alu elements has alsobeen documented, with the most intensively studiedexample involving the MLL gene (Hess, 2004). Recom-bination between intronic Alu elements, leading to par-
*Correspondence: [email protected]
2 Present address: Institute for Cancer Genetics, Columbia Univer-sity, 1150 St. Nicholas Ave., New York, New York 10032.tial duplication of MLL, has been demonstrated incases of acute myeloid leukemia (AML) in which pa-tients lack cytogenetic defects (or have trisomy 11)(Caligiuri et al., 1994; Schichman et al., 1994; So et al.,1997; Strout et al., 1998). The involvement of Alu ele-ments in MLL partial duplications is not limited to Alu-Alu recombination, as breakpoint junctions between anAlu element and non-Alu sequences have also beenidentified (Strout et al., 1998). Alu-Alu recombinationhas also been reported in the generation of a reciprocaltranslocation present in tumor DNA (Onno et al., 1992).However, in most reciprocal translocations, Alu ele-ments are either joined to non-Alu sequences or fusedto other Alu elements or simply occur within the vicinityof breakpoint junctions (Kolomietz et al., 2002).
Genomic rearrangements arise from breakage andmisrepair, especially of DSBs. Multiple pathways of re-pair have been demonstrated in mammalian cells forthe repair of a single chromosomal DSB (Liang et al.,1998). The two primary pathways are NHEJ, involvinglittle or no homology, and conservative homologous re-combination (HR) (van Gent et al., 2001). A third path-way, SSA, can also occur at homologous sequencesnear a DSB. Unlike HR, which involves strand invasion,SSA involves the annealing of DNA strands formed af-ter resection at the DSB (Pâques and Haber, 1999).
We set out to investigate the involvement of DSB re-pair pathways in genomic rearrangements in mamma-lian cells, specifically chromosomal translocations. Pre-viously, we showed that translocations could be formedduring the repair of DSBs in mouse ES cells (Richard-son and Jasin, 2000). Translocations were a fraction ofthe recovered repair products in this system, and eachtranslocation had the same overall structure due toconstraints in the substrate design. We devised, there-fore, a strategy that would allow us to specifically se-lect translocations in order to determine which mecha-nism of DSB repair predominates as the translocationpathway. This strategy involves an intron-based sub-strate design that is adaptable to addressing a numberof issues regarding mechanisms of genomic rearrange-ments in mammalian cells.
Results
Translocation Substrates: Breakpoint Junctionswithin an IntronKey to our translocation substrate design is an intron-containing neomycin phosphotransferase gene (neo):the DSBs occur within a split intron of the neo gene toallow the selection of a variety of breakpoint junctionsin the formation of the derivative (der) chromosomes(Figure 1A). For example, translocations involving NHEJand/or SSA can be selected. Another important aspectof the design is that an Alu element is incorporated ad-jacent to each of the DSB sites. A second set of re-peats, derived from the puromycin resistance gene(puro), is also incorporated adjacent to the DSB sitesbut on the opposite side from the Alu elements (brack-ets, Figure 1A).

Molecular Cell886
Figure 1. Translocation Substrates and Pos-sible Reciprocal Translocation Outcomes
(A) Translocation substrates on chromo-somes (chrs.) 17 and 14 are based on a splitneo intron design with an Alu element fromthe MLL gene (blue box) inserted 3# and 5#of the splice donor and splice acceptor por-tions of the neo gene (neoSD and SAneo,respectively). DSBs generated by the I-SceIendonuclease followed by interchromosomalrepair can potentially generate translocationchromosomes. Repair by SSA at the iden-tical Alu elements or puro sequences (brack-ets) will delete one of the repeats, whereasNHEJ could fuse the Alu elements or purosequences, as shown. A neo+ gene is formedon der(17) by either pathway, whereas onder(14), a puro+ gene is formed by SSA anda puro− gene is formed by NHEJ. By usingSSA and/or NHEJ, four combinations of re-ciprocal translocations are possible. The to-tal size of the intron after precise NHEJ isapproximately 1 kb.(B) Gene targeting of the chrs. 17 and 14 atthe Pim1 and Rb loci to create targeted al-leles with the translocation substrates (p5and pF, respectively). The chr. 17 substratewas targeted first for the creation of the p5cell line. The chr. 14 substrate was subse-quently targeted to the Rb locus in the p5cell line to create the p5pF cell lines #5, #6,and #18. Vertical black bars are exons 1–4for the Pim1 locus and exon 20 for the Rblocus. Abbreviations: HII, HincII and Pst,PstI.
linked HPRT mini-gene, the pF translocation substrate
cation assays.
Table 1. DSB-Induced Translocation Frequencya
neo+ Colonies (×10−5)b
Cell Lines No DNA DSB-Induced (pCBASce)
p5pF <0.01 2.6 ± 1.9Hom Alu <0.01 5.0 ± 4.0Het Alu <0.01 2.7 ± 1.4
a For the p5pF and Het Alu cell lines, three independent clones wereexamined for each. For the Hom Alu cell lines, two independentclones were examined. Two experiments were done on each cellline, except for one cell line of p5pF, Hom Alu, and Het Alu.b The frequency of neo+ colonies was determined by dividing thenumber of neo+ colonies by the number of electroporated cells (2 ×
The two translocation substrates were targeted to loci wion chromosomes 17 and 14 in mouse ES cells (Figure 1).
The chromosome 17 substrate, termed p5, consists of pathe following four components: a 5# neo fragment with a
splice donor site (neoSD), intronic sequences including tan Alu element, an I-SceI endonuclease cleavage sitefor DSB formation, and a 3# puro fragment (3#puro)(Figure 1A). The Alu element, derived from intron 1 ofthe MLL gene, has been demonstrated to participate insome of the partial tandem duplications of MLL foundin patients with AML (Hess, 2004). By using a linkedhygromycin resistance gene (hyg), we targeted thistranslocation substrate to the Pim1 locus on chromo-some 17 in ES cells to create the p5 allele (Figure 1B).The chromosome 14 substrate, termed pF, consists ofa 5# puro fragment (5#puro), an I-SceI site, the sameMLL intron 1 Alu element followed by additional intronicsequences, and a 3# neo fragment with a splice accep-tor site (SAneo) (Figure 1A). The two Alu elements share290 bp of identity, whereas the 5#puro and 3#puro share265 bp of identity (brackets, Figure 1A). By using a
as introduced into the p5 cell line by targeting to anntron in the Rb locus on chromosome 14 to create theF allele (Figure 1B). HPRT+ colonies were selected,nd three independent clones that were correctlyargeted (Figure 1B) were chosen for use in the translo-
107), correcting for 50% viability after electroporation.
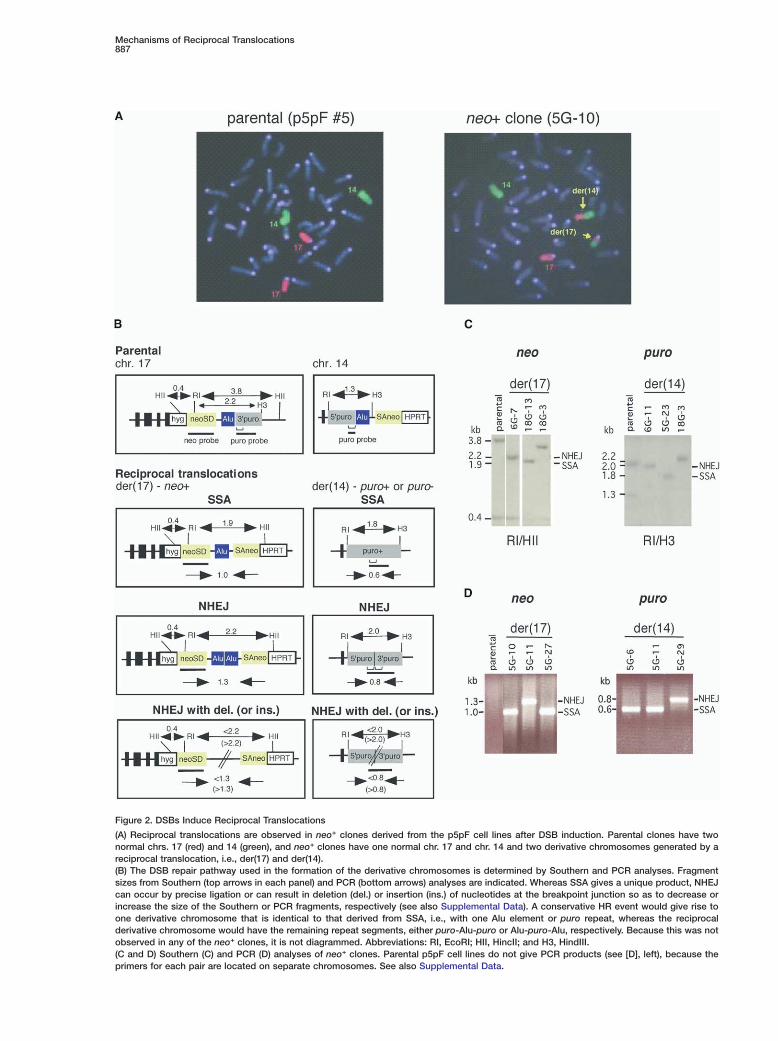
Mechanisms of Reciprocal Translocations887
Figure 2. DSBs Induce Reciprocal Translocations
(A) Reciprocal translocations are observed in neo+ clones derived from the p5pF cell lines after DSB induction. Parental clones have twonormal chrs. 17 (red) and 14 (green), and neo+ clones have one normal chr. 17 and chr. 14 and two derivative chromosomes generated by areciprocal translocation, i.e., der(17) and der(14).(B) The DSB repair pathway used in the formation of the derivative chromosomes is determined by Southern and PCR analyses. Fragmentsizes from Southern (top arrows in each panel) and PCR (bottom arrows) analyses are indicated. Whereas SSA gives a unique product, NHEJcan occur by precise ligation or can result in deletion (del.) or insertion (ins.) of nucleotides at the breakpoint junction so as to decrease orincrease the size of the Southern or PCR fragments, respectively (see also Supplemental Data). A conservative HR event would give rise toone derivative chromosome that is identical to that derived from SSA, i.e., with one Alu element or puro repeat, whereas the reciprocalderivative chromosome would have the remaining repeat segments, either puro-Alu-puro or Alu-puro-Alu, respectively. Because this was notobserved in any of the neo+ clones, it is not diagrammed. Abbreviations: RI, EcoRI; HII, HincII; and H3, HindIII.(C and D) Southern (C) and PCR (D) analyses of neo+ clones. Parental p5pF cell lines do not give PCR products (see [D], left), because theprimers for each pair are located on separate chromosomes. See also Supplemental Data.

Molecular Cell888
Figure 3. Translocations by an SSA Mechanism Predominate at Identical Alu Elements
(A) Classes of reciprocal translocation outcomes obtained in neo+ clones from the p5pF and Rev48 cell lines after I-SceI expression. Thesetwo cell lines differ in that the p5pF cell line has the I-SceI sites in the same relative orientation on chrs. 17 and 14, whereas for the Rev48cell line, the I-SceI sites are in opposite orientation. Thus, in the formation of translocation chromosomes, intact I-SceI overhangs at the DNAends have the potential to be precisely ligated in the p5pF cell line, but not in the Rev48 cell line.(B) Sequence analysis of breakpoint junctions from der(17) and der(14) from p5pF neo+ clones. The boxed nucleotides show the breakpointsequences on both strands after I-SceI cleavage. Breakpoint junction sequences were obtained for each of the derivative chromosomesexcept in four cases, which are indicated in parenthesis. The number of nucleotides deleted (del.) from the top strand of each end areindicated, as well as the number or sequence of nucleotides inserted (ins.). Nucleotides that could have been derived from the I-SceI overhangon the bottom strand are in bold. The four base I-SceI overhangs are underlined with thin lines in the three junctions derived from preciseligation (asterisks); microhomologies which occur at three other junctions are underlined with thick lines.(C) Local derivation of the insertion on der(14) of clone 18G-3. The insertion is derived from sequences 5# and 3# of the chr. 17 DSB (thickand thin black bars, respectively). This includes 173 bp of the Alu element, partial I-SceI sequences, and 3#puro sequence, in addition to 9bp of a (TAn)2 insertion. Spaces between thick bars represent sequences from chr. 17 that were not contiguous. The number of insertednucleotides is indicated in bp below each insertion. The dotted line indicates 31 bp of originally noncontiguous sequence that was repeatedin the junction.

Mechanisms of Reciprocal Translocations889
To induce translocations, the three p5pF cell lineswere electroporated with the I-SceI endonuclease ex-pression vector, and neo+ colonies were selected. Eachof the p5pF cell lines gave similar numbers of neo+ col-onies after I-SceI expression, resulting in an averagefrequency of 2.6 ± 1.9 × 10−5 (Table 1). In the absence ofI-SceI expression, spontaneously arising neo+ colonieswere not detected (<10−7), indicating that neo+ coloniesarose from DSB repair. Fluorescence in situ hybridiza-tion (FISH) demonstrated that the neo+ clones carriedthe two derivative chromosomes, i.e., der(14) and der(17),expected from a reciprocal translocation (Figure 2A).The neo+ clones also had one intact chromosome 14and chromosome 17, as expected, as the translocationsubstrates are present on only one chromosome 14 andchromosome 17 in the parental p5pF cell lines.
SSA Predominates as the Translocation Pathwaywith Identical Alu ElementsTo identify the translocation pathway(s), 47 neo+ clonesfrom the p5pF cell lines were analyzed by Southernblotting and PCR (Figures 2B–2D; see SupplementalData available online with this article). The predominantclass of clones was found to have both derivativeschromosomes formed by SSA (Class 1, 81%; Figure3A). Considering the individual derivative chromo-somes, 40 (85%) of the der(17) chromosomes were de-rived from SSA of the identical Alu elements, with theremaining seven (15%) derived from NHEJ (Figure 3A).Similarly, 39 (83%) of the der(14) chromosomes werederived from SSA at the puro repeat, whereas onlyeight (17%) were derived from NHEJ. Thus, SSA is thepreferred translocation pathway for generating eitherderivative chromosome in the p5pF cell lines.
Translocations associated with deletions that extendbeyond the intronic sequences into the neo coding se-quences would preclude the formation of an intact neogene. We therefore took advantage of the ability to se-lect der(14) translocations with puromycin in order todetermine if translocations selected in this way wouldarise in a similar manner as when neo+ clones wereselected. Of 54 puro+ clones, we found that 52 clones(96%) were also neo+ (data not shown). Thus, neo+ se-lection appears to capture most of the translocationsassociated with DSB induction on chromosomes 14and 17. As before, der(17) primarily arose by SSA of theAlu elements (data not shown).
Because the 18 bp I-SceI sites in the translocationsubstrates are in the same relative orientation on bothchromosomes 14 and 17, it is formally possible that aderivative chromosome could be formed by precise li-gation of the I-SceI cohesive overhangs but then un-dergo another DSB that is repaired by intrachromoso-mal SSA. To verify that SSA is the initial repair eventleading to the translocations, we constructed anothercell line, termed Rev48, in which the I-SceI sites are inopposite relative orientations on chromosomes 14 and17. In this cell line, the p5 allele (Figure 1B) is presentas before on chromosome 17, but the pF allele on chro-mosome 14 was modified to contain the I-SceI site inreverse orientation (see Supplemental Data). I-SceI en-donuclease was expressed in the Rev48 cell line and20 neo+ clones were analyzed at the molecular level to
determine the translocation pathway. From theseclones, 18 (80%) had der(17) chromosomes generatedby SSA and all 20 had der(14) generated by SSA (Figure3A). Thus, the bias toward SSA at the translocationbreakpoint junctions is not the result of a secondaryDSB repair event.
Translocation Breakpoint Junctions Derivedfrom NHEJA total of 15 of the 47 translocation breakpoint junc-tions from the p5pF neo+ clones were formed by NHEJ.These junctions were analyzed in more detail. Three ofthe breakpoint junctions had a restored I-SceI site (as-terisk, Figure 3B) from precise ligation of the I-SceIoverhangs (thin underline, Figure 3B). Small deletionsand/or insertions, i.e., involving <22 bp, were found inseven junctions. In one junction, a larger insertion (224bp) was found that had a complex origin (Figure 3C).The insertion was “locally derived,” i.e., derived fromnearby sequences, as has been seen in translocationbreakpoint junctions in a number of cancers (Zucman-Rossi et al., 1998). Larger insertions of approximately0.9 kb, and 0.7 kb occurred at two der(17) junctions,as determined by Southern analysis. For one der(14)junction, no signal was obtained by Southern blotanalysis, indicating that the puro probe sequence wasdeleted (data not shown). For another der(14) junction,a more complex Southern hybridzation pattern was ob-served, indicating that the puro gene sequences wererearranged and/or deleted. We also sequenced five ofthe SSA products (as indicated in Figure 3B) and veri-fied that no mutations were introduced into the repeatsduring SSA.
In summary, from 47 neo+ clones, 15 of the 94 break-point junctions were derived from NHEJ, the majority(10 of 15 junctions) of which had little, if any, degrada-tion or other alteration to the ends (0–21 bp); the re-maining breakpoint junctions involved more extensiveend modifications.
Heterology between the Alu Elements Has LittleEffect on Translocation FrequencyAlu elements in human genomes are frequently quite di-vergent from each other, with a range of divergence fromthe consensus Alu element of between 2% and 30%(Smit, 1996). We therefore asked if the substitution of aheterologous Alu element in one of the translocationsubstrates would affect the translocation frequencyand/or translocation pathway. For this, we substitutedthe MLL intron 1 Alu element in the chromosome 14translocation substrate with an Alu element from intron6 of the MLL gene, which is within the major breakpointcluster region (see Figure S1). The two Alu elements,which are both from the Alu Sx subfamily, are 20% di-vergent from each other, although there are regions ofup to 25 bp of complete identity. We paired these par-ticular Alu elements because somatic recombinationevents between them have been found in patients withAML (So et al., 1997; Strout et al., 1998) (Figure S1).
To construct cell lines containing the heterologousAlu element, termed the Het Alu cell lines, the chromo-some 14 targeting vector (Figure 1B, Figure S1A) wasmodified to contain the intron 6 Alu element and then
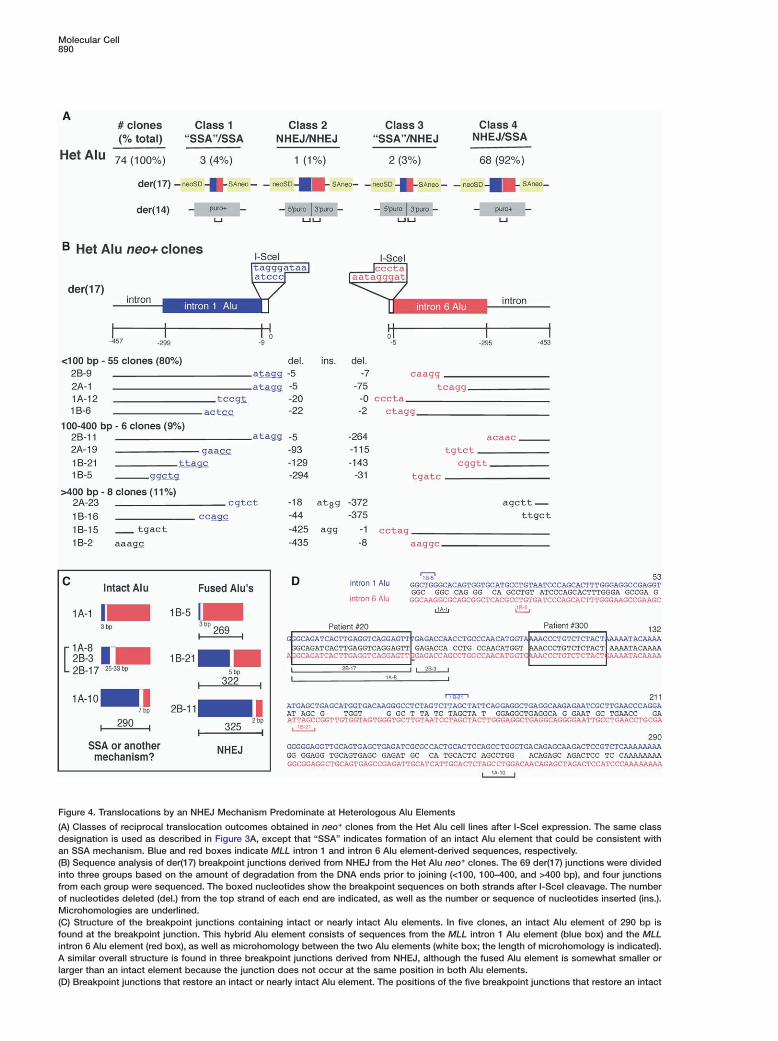
Molecular Cell890
Figure 4. Translocations by an NHEJ Mechanism Predominate at Heterologous Alu Elements
(A) Classes of reciprocal translocation outcomes obtained in neo+ clones from the Het Alu cell lines after I-SceI expression. The same classdesignation is used as described in Figure 3A, except that “SSA” indicates formation of an intact Alu element that could be consistent withan SSA mechanism. Blue and red boxes indicate MLL intron 1 and intron 6 Alu element-derived sequences, respectively.(B) Sequence analysis of der(17) breakpoint junctions derived from NHEJ from the Het Alu neo+ clones. The 69 der(17) junctions were dividedinto three groups based on the amount of degradation from the DNA ends prior to joining (<100, 100–400, and >400 bp), and four junctionsfrom each group were sequenced. The boxed nucleotides show the breakpoint sequences on both strands after I-SceI cleavage. The numberof nucleotides deleted (del.) from the top strand of each end are indicated, as well as the number or sequence of nucleotides inserted (ins.).Microhomologies are underlined.(C) Structure of the breakpoint junctions containing intact or nearly intact Alu elements. In five clones, an intact Alu element of 290 bp isfound at the breakpoint junction. This hybrid Alu element consists of sequences from the MLL intron 1 Alu element (blue box) and the MLLintron 6 Alu element (red box), as well as microhomology between the two Alu elements (white box; the length of microhomology is indicated).A similar overall structure is found in three breakpoint junctions derived from NHEJ, although the fused Alu element is somewhat smaller orlarger than an intact element because the junction does not occur at the same position in both Alu elements.(D) Breakpoint junctions that restore an intact or nearly intact Alu element. The positions of the five breakpoint junctions that restore an intact

Mechanisms of Reciprocal Translocations891
pathway, such that 85% of the derivative chromosomeseither in their entirety or between portions of the ele-
hybrid Alu element are shown in brackets below the aligned sequences. Three of these junctions occur at a position where 32 of 33 bp areidentical between the two Alu elements and at which the breakpoint junction (boxed sequence) occurs for AML patient 20 (So et al., 1997).The breakpoint junction for patient 300 (Strout et al., 1998) is also boxed. The position of two junctions from clones with fused Alu elementsthat are not in register are also indicated; the point of fusion with respect to the intron 1 Alu and intron 6 Alu is shown above and below theline, respectively.
targeted to chromosome 14 in the p5 cell line. Threeclones that were correctly targeted were used in thesubsequent analysis. Because this modification led tosome sequence changes near the I-SceI site, we re-cloned the intron 1 Alu element into the chromosome14 targeting vector in an identical manner to create two“Hom Alu” cell lines in which the intron 1 Alu elementis on both chromosomes 17 and 14 (Figure S1A). In theHom Alu and Het Alu parental cell lines, the I-SceI siteon chromosome 14 is in the opposite orientation rela-tive to the I-SceI site on chromosome 17, as with theRev48 cell line.
The I-SceI expression vector was electroporated intothe Hom Alu and Het Alu cell lines, and neo+ colonieswere selected. The Hom Alu cell lines had an averagefrequency of 5.0 ± 4.0 × 10−5 of neo+ colonies, and theHet Alu cell lines had an average frequency of neo+ col-onies of 2.7 ± 1.4 × 10−5 (p = 0.27; Table 1). By usingsimilar analyses as described for the p5pF cell lines,the neo+ clones from the Hom Alu and Het Alu cell lineswere found to contain reciprocal chromosomal translo-cations (see below; data not shown). Thus, the substi-tution of a highly diverged Alu element has little or noeffect on the overall translocation frequency.
NHEJ Predominates as the Translocation PathwayWhen Heterology Is Present at the DNA EndsWe analyzed a number of neo+ clones from the HomAlu cell lines and found that, as with the p5pF andRev48 cell lines, SSA predominated as the transloca-tion pathway for both derivative chromosomes (datanot shown). We next characterized 74 neo+ clones de-rived from the Het Alu cell lines. In contrast to cell lineswith identical Alu elements in which >80% of neo+
clones were in Class 1 (SSA/SSA), 92% of neo+ clonesderived from the Het Alu cell lines fell into Class 4(NHEJ/SSA) (Figure 4A). Thus, although heterology atthe chromosome ends did not substantially affect thetranslocation frequency, it dramatically shifted translo-cation pathway usage from SSA to NHEJ in the forma-tion of the der(17) chromosome. Formation of the recip-rocal der(14) chromosome by SSA was not affected bythis shift in pathway usage.
To further characterize the der(17) breakpoint junc-tions derived from NHEJ, we analyzed PCR productsfrom the neo+ gene. The 69 clones with breakpointjunctions derived from NHEJ were classified into threegroups depending on the estimated amount of degra-dation from the DNA ends prior to NHEJ (Figure 4B) asdeduced from the size of the neo PCR products (datanot shown). The majority of neo+ clones (55 clones;80%) had less than 100 bp deleted at one or bothDSBs; an additional six clones (9%) had between 100and 400 bp deleted. Thus, in most of the clones, NHEJled to fusion of the two heterologous Alu elements,
ments. In some cases, the fusion formed a nearly unit-length Alu element (Figure 4C). The remaining eightclones (11%) had greater than 400 bp deleted, leadingto complete loss of one of the Alu elements (Figure 4B).In these cases, the deletion approached the intron/exon border of the neo gene on one side.
Junctions with various deletion lengths were se-quenced (Figure 4B). In some junctions, the deletionswere nearly symmetrical around the DSBs, as in clone1B-21 in which 129 and 143 bp were deleted from thechromosome 17 and chromosome 14 ends, respec-tively. In several other junctions, the deletions werehighly asymmetrical, as in clone 1B-15 in which 425and 1 bp were deleted from the chromosome 17 andchromosome 14 ends, respectively. Microhomology of1–5 bp was present at all of the junctions that did notcontain an insertion. Positions of microhomology areindicated for two of the fused Alu elements that formeda nearly unit-length Alu element (clones 1B-5 and 1B-21; Figures 4C and 4D).
In addition to the clones with bona fide NHEJ junc-tions, five clones had events that led to the formationof a single, intact Alu element at the breakpoint junc-tion, which would be consistent with an SSA event(hence, “SSA” in Figure 4A). In these intact Alu ele-ments, the 5# portion was derived from the intron 1 Aluelement, and the 3# portion was derived from the intron6 Alu element (Figure 4C). This was very similar to thefused Alu elements arising from NHEJ, except thatthese five junctions were “in register,” i.e, at the sameposition in both Alu elements, rather than being offset(Figure 4D). Microhomology was present at each of thefive junctions. In two clones, the microhomology wasshort, i.e., 3 and 7 bp (clones 1A-1 and 1A-10, respec-tively; Figures 4C and 4D). In the other three clones(clones 1A-8, 2B-3, and 2B-17), the junction occurredwithin the longest stretch of near identity between thetwo Alu elements at the position of the breakpoint junc-tion found in an AML patient (patient 20; Figure 4C).Within this stretch, 32 of 33 nucleotides are identical:the junctions of clones 2B-17 and 2B-3 occurred 5# and3# of the single nucleotide polymorphism that exists be-tween the two Alu elements in this stretch, respectively,whereas the junction for clone 1A-8 contained a G/T toA mutation at this polymorphic site.
Discussion
In this report, we investigated translocation pathwaychoice in mammalian cells. In particular, we examinedthe role of Alu elements, because these repetitive ele-ments comprise a large portion of the human genome.By using an intron-based translocation system, wefound that when identical Alu elements are present atDNA ends, SSA predominated as the translocation

Molecular Cell892
had a single Alu element at the breakpoint junction. tOWhen diverged Alu elements are present at DNA ends,
translocation frequency was not substantially altered; e(however, there was a dramatic shift to the NHEJ path-
way, such that 93% of the derivative chromosomes had aobreakpoint junctions involving little or no sequence
identity. eOur translocation system provides a good model for
oncogenic translocations, which frequently have break- Aspoint junctions within intronic sequences and often in
the vicinity of Alu elements (Deininger and Batzer, 1999; slGreaves and Wiemels, 2003; Kolomietz et al., 2002). A
yeast intron-based translocation system has also been egrecently developed (Yu and Gabriel, 2004), which will
be important for phylogenetic comparisons. Although tfintron-based systems impose some constraints, our re-
sults suggest that the majority of translocation chromo- tnsomes are recovered with neo+ gene selection in our
system. The intron-based system, therefore, provides Methe flexibility necessary to recover a variety of break-
point junctions. fvdTranslocation Pathway ChoicemOur results provide an explanation for why most onco-Sgenic translocation junctions rarely involve recombina-gtion between repetitive elements: sequence divergenceais sufficient to shift homology-based repair events tobnonhomologous repair events. It seems remarkable
that the translocation frequency in our experiments wastnot substantially altered in cell lines containing heterol-rogous Alu elements compared with cell lines containingtidentical elements, even though the DSB repair path-mway dramatically shifted. We do not expect the SSAyevents on der(14) to somehow “drive” the recovery ofmder(17) events by NHEJ, because we find that translo-tcations can occur in the absence of nearby homologyvat a similar frequency to the events described here (D.pWeinstock, B.E., and M.J., unpublished data).wA priori it could have been predicted that cellularifactors would control repair pathway usage, as hashbeen seen in yeast (Frank-Vaillant and Marcand, 2002;sKarathanasis and Wilson, 2002). Our results, therefore,khighlight the potential for overlapping use of repairspathways at a DSB in mammalian cells and emphasizelhow readily cells can shift from one pathway to another,cindicating the “fluidity” of DSB repair. Competition andtcollaboration of DSB repair pathways have been notedepreviously in other studies in vertebrate cells; however,athese other studies have utilized DSB repair mutantsiwhich are defective for one or more pathways of repairt(e.g., Couedel et al. [2004], Mills et al. [2004], Stark etsal. [2004], and Takata et al. [1998]).
Factors that are thought to influence the choice ofDSB repair pathway include cell cycle stage, cell type, A
Ogrowth conditions, and age of cells. This control of DSBrepair pathway usage would have made it seem likely n
ethat cells would “choose” a pathway prior to sensingthe degree of sequence homology at the DNA ends and w
athen begin processing the DNA ends in a manner ap-propriate to the chosen pathway. Based on studies in p
ayeast, the initial processing steps at a DSB differ forSSA and NHEJ. In SSA, DNA ends are resected in a 5# w
pto 3# direction to produce 3# single-stranded tails that
hen anneal to each other (Pâques and Haber, 1999).n the other hand, NHEJ efficiently utilizes intact DNAnds and is suppressed by 5# to 3# end resection
Frank-Vaillant and Marcand, 2002; Ira et al., 2004). Asresult, when SSA is impaired in yeast by the removal
f repeat sequences or by rad52 mutation, NHEJ is notnhanced (Karathanasis and Wilson, 2002).For SSA events in our cell lines containing identical
lu elements, w290 nucleotides would need to be re-ected from each end in order to reveal complementarytrands for annealing. By contrast, for NHEJ in our cellines containing heterologous Alu elements, a need forxtensive end modification would seem to be abro-ated for most events, given the limited end modifica-ions observed at the breakpoint junctions. A major dif-erence between NHEJ in yeast and mammalian cells ishe ability of mammalian cells to efficiently join aumber of DNA end structures (Smith et al., 2001).ammalian cells may be able to either fill in resected
nds or, alternatively, make use of resected DNA endsor NHEJ in a “micro-SSA” type of NHEJ reaction in-olving only a few bps of sequence identity. However,istinct repair factors are expected to be recruited foricro-SSA events involving a few bps and for “macro-SA” events involving 290 bp, because in yeast, theenetic requirement for these events differ (Kramer etl., 1994). Therefore, these events are not expected toe equivalent.Given the major class of reciprocal translocations in
he Het Alu cell lines (i.e., class 4), as well as previousesults (Richardson and Jasin, 2000), our results implyhat NHEJ and SSA are available to repair broken chro-osome ends at the same stage(s) of the cell cycle. In
east, strand resection is cell cycle regulated, beinguch reduced in G1 cells (Ira et al., 2004); presumably,
herefore, SSA would also be reduced in G1 cells. Inertebrate cells, NHEJ is considered to be the preferredathway in the G1 phase of the cell cycle, whereas HR,hich like SSA requires strand resection, is most active
n late S/G2 (Takata et al., 1998). Recent work, however,as emphasized that NHEJ is not restricted to the G1tage (Couedel et al., 2004; Mills et al., 2004; Roth-amm et al., 2003). Thus, both NHEJ and SSA are pre-umably active in late S/G2, suggesting that our trans-ocations may be occurring at this point in the cellycle. It should be noted that translocations arehought to be only a small portion of total DSB repairvents in our cell lines. Translocations are recoveredt a frequency of approximately 3–5 × 10−5, whereas
ntrachromosomal NHEJ and HR events are presumedo occur at >10−2, based on results from other I-SceIubstrates in ES cells (Moynahan et al., 2001).
lu Elements and Genomic Rearrangementsur experiments raise the question as to the mecha-ism of Alu-Alu recombination that gives rise to dis-ase-causing alleles. At least in somatic cells, HRould not appear to be a favored mechanism to gener-te chromosomal rearrangements, given previouslyublished reports (Richardson and Jasin, 2000; Rich-rdson et al., 1998). Moreover, conservative HR eventsould be dependent on RAD51, yet we find that ex-ression of a dominant negative RAD51 protein (Stark

Mechanisms of Reciprocal Translocations893
et al., 2004) does not reduce the recovery of transloca-tions (B.E. and M.J., unpublished observations).
The five Het Alu clones in which intact, unit-lengthAlu elements were generated by translocation (Figure4C) are similar to the SSA products from cell lines con-taining identical elements. However, it is not certainthat these junctions are derived from SSA. Annealingof the strands from the two heterologous Alu elementswould produce multiple mismatches along the lengthsof the Alu elements and would have to escape hetero-duplex rejection. Presumably, such an annealed pro-duct would be a substrate for mismatch repair (Suga-wara et al., 2004), which might be expected to producean Alu element containing patches from each Alu ele-ment, rather than the hybrid element we observed witha single crossover position. It is notable, however, thatwe obtained fused Alu elements from NHEJ that werenot in register but which came close in size to restoringan intact Alu element (Figure 4C). In each case, thebreakpoint junction for the fused elements occurred ata region of microhomology, such that the structure ofthese fused elements is strikingly similar to the intactAlu elements.
These observations suggest the possibility that ap-parent Alu-Alu recombination events between heterolo-gous elements may arise in some instances from micro-homology-mediated NHEJ in which the two Aluelements are in register with each other. This is espe-cially attractive given that NHEJ is very efficient inmammalian cells and that sequence divergence signifi-cantly suppresses SSA and HR. Microhomology-medi-ated NHEJ has the potential to give rise to a wide rangeof events—deletions, duplications (if between sisterchromatids), and translocations. The intron-based sys-tem we have developed in this report will allow us tofurther explore the genetic requirements of theseevents.
Experimental Procedures
DNA Manipulations and Cell Line ConstructionsThe first intron from the adenovirus (Ad1) major late transcriptionunit was previously used in a mouse lacZ recombination reporter(Moynahan et al., 1996) and was adapted for use here in the neogene of pMC1neo. The resulting construct was verified to give riseto G418R colonies upon transfection into mammalian cells. Thisp-i-neo construct was used in subsequent steps to derive neoSDand SAneo. The Pim1 and Rb targeting constructs have been de-scribed previously (Richardson et al., 1998). For details on cloning,see Supplemental Data.
Translocation AnalysisFor translocation experiments, 2 × 107 cells in 1 ml of phosphate-buffered saline were electroporated with 20–25 µg of the I-SceI ex-pression vector pCBASce (Richardson et al., 1998) in 0.4 cmelectrode-gap cuvette (250 V, 960 µF). Electroporated cells werealiquoted into 2–5 10 cm diameter dishes. Colonies were selectedin ES media with a drug 20–24 hr after electroporation and weregrown in selection media for 10–14 days before colony counts (orafter colony expansion): G418 (200 �g/ml), hygromycin (150 �g/ml),puromycin (1.6 �g/ml), or HAT (1× Sigma H0262). For analyses ofneo+ clones, see Supplemental Data.
Supplemental DataSupplemental Data include Supplemental Experimental Procedures,Supplemental References, and one figure and are available on-
line with this article at http://www.molecule.org/cgi/content/full/17/6/885/DC1/.
Acknowledgments
We thank Margaret Leversha at the Molecular Cytogenetics CoreFacility (MSKCC) for performing FISH experiments and MichaelBacklund and David Weinstock for assistance and discussions.This project was supported by the Dorothy Rodbell Cohen Founda-tion (B.E.), National Science Foundation 0346354, and National In-stitutes of Health GM54668 (M.J.).
Received: December 4, 2004Revised: February 4, 2005Accepted: February 24, 2005Published: March 17, 2005
References
Caligiuri, M.A., Schichman, S.A., Strout, M.P., Mrozek, K., Baer,M.R., Frankel, S.R., Barcos, M., Herzig, G.P., Croce, C.M., andBloomfield, C.D. (1994). Molecular rearrangement of the ALL-1gene in acute myeloid leukemia without cytogenetic evidence of11q23 chromosomal translocations. Cancer Res. 54, 370–373.
Couedel, C., Mills, K.D., Barchi, M., Shen, L., Olshen, A., Johnson,R.D., Nussenzweig, A., Essers, J., Kanaar, R., Li, G.C., et al. (2004).Collaboration of homologous recombination and nonhomologousend-joining factors for the survival and integrity of mice and cells.Genes Dev. 18, 1293–1304.
Deininger, P.L., and Batzer, M.A. (1999). Alu repeats and human dis-ease. Mol. Genet. Metab. 67, 183–193.
Deininger, P.L., Moran, J.V., Batzer, M.A., and Kazazian, H.H., Jr.(2003). Mobile elements and mammalian genome evolution. Curr.Opin. Genet. Dev. 13, 651–658.
Frank-Vaillant, M., and Marcand, S. (2002). Transient stability ofDNA ends allows nonhomologous end joining to precede homolo-gous recombination. Mol. Cell 10, 1189–1199.
Greaves, M.F., and Wiemels, J. (2003). Origins of chromosometranslocations in childhood leukaemia. Nat. Rev. Cancer 3, 639–649.
Hess, J.L. (2004). MLL: a histone methyltransferase disrupted inleukemia. Trends Mol. Med. 10, 500–507.
Ira, G., Pellicioli, A., Balijja, A., Wang, X., Fiorani, S., Carotenuto,W., Liberi, G., Bressan, D., Wan, L., Hollingsworth, N.M., et al.(2004). DNA end resection, homologous recombination and DNAdamage checkpoint activation require CDK1. Nature 431, 1011–1017.
Karathanasis, E., and Wilson, T.E. (2002). Enhancement of Saccha-romyces cerevisiae end-joining efficiency by cell growth stage butnot by impairment of recombination. Genetics 161, 1015–1027.
Kolomietz, E., Meyn, M.S., Pandita, A., and Squire, J.A. (2002). Therole of Alu repeat clusters as mediators of recurrent chromosomalaberrations in tumors. Genes Chromosomes Cancer 35, 97–112.
Kramer, K.M., Brock, J.A., Bloom, K., Moore, J.K., and Haber, J.E.(1994). Two different types of double-strand breaks in Saccharo-myces cerevisiae are repaired by similar RAD52-independent, non-homologous recombination events. Mol. Cell. Biol. 14, 1293–1301.
Lander, E.S., Linton, L.M., Birren, B., Nusbaum, C., Zody, M.C.,Baldwin, J., Devon, K., Dewar, K., Doyle, M., FitzHugh, W., et al.(2001). Initial sequencing and analysis of the human genome. Na-ture 409, 860–921.
Liang, F., Han, M., Romanienko, P.J., and Jasin, M. (1998). Homol-ogy-directed repair is a major double-strand break repair pathwayin mammalian cells. Proc. Natl. Acad. Sci. USA 95, 5172–5177.
Mills, K.D., Ferguson, D.O., Essers, J., Eckersdorff, M., Kanaar, R.,and Alt, F.W. (2004). Rad54 and DNA Ligase IV cooperate to main-tain mammalian chromatid stability. Genes Dev. 18, 1283–1292.
Moynahan, M.E., Akgun, E., and Jasin, M. (1996). A model for test-

Molecular Cell894
ing recombinogenic sequences in the mouse germline. Hum. Mol.Genet. 5, 875–886.
Moynahan, M.E., Pierce, A.J., and Jasin, M. (2001). BRCA2 is re-quired for homology-directed repair of chromosomal breaks. Mol.Cell 7, 263–272.
Onno, M., Nakamura, T., Hillova, J., and Hill, M. (1992). Rearrange-ment of the human tre oncogene by homologous recombinationbetween Alu repeats of nucleotide sequences from two differentchromosomes. Oncogene 7, 2519–2523.
Pâques, F., and Haber, J.E. (1999). Multiple pathways of recombina-tion induced by double-strand breaks in Saccharomyces cerevis-iae. Microbiol. Mol. Biol. Rev. 63, 349–404.
Richardson, C., and Jasin, M. (2000). Frequent chromosomal trans-locations induced by DNA double-strand breaks. Nature 405, 697–700.
Richardson, C., Moynahan, M.E., and Jasin, M. (1998). Double-strand break repair by interchromosomal recombination: suppres-sion of chromosomal translocations. Genes Dev. 12, 3831–3842.
Rothkamm, K., Kruger, I., Thompson, L.H., and Lobrich, M. (2003).Pathways of DNA double-strand break repair during the mamma-lian cell cycle. Mol. Cell. Biol. 23, 5706–5715.
Schichman, S.A., Caligiuri, M.A., Strout, M.P., Carter, S.L., Gu, Y.,Canaani, E., Bloomfield, C.D., and Croce, C.M. (1994). ALL-1 tan-dem duplication in acute myeloid leukemia with a normal karyotypeinvolves homologous recombination between Alu elements. CancerRes. 54, 4277–4280.
Smit, A.F. (1996). The origin of interspersed repeats in the humangenome. Curr. Opin. Genet. Dev. 6, 743–748.
Smith, J., Baldeyron, C., De Oliveira, I., Sala-Trepat, M., and Papa-dopoulo, D. (2001). The influence of DNA double-strand breakstructure on end-joining in human cells. Nucleic Acids Res. 29,4783–4792.
So, C.W., Ma, Z.G., Price, C.M., Dong, S., Chen, S.J., Gu, L.J., So,C.K., Wiedemann, L.M., and Chan, L.C. (1997). MLL self fusion me-diated by Alu repeat homologous recombination and prognosis ofAML-M4/M5 subtypes. Cancer Res. 57, 117–122.
Stark, J.M., Pierce, A.J., Oh, J., Pastink, A., and Jasin, M. (2004).Genetic steps of mammalian homologous repair with distinct muta-genic consequences. Mol. Cell. Biol. 24, 9305–9316.
Strout, M.P., Marcucci, G., Bloomfield, C.D., and Caligiuri, M.A.(1998). The partial tandem duplication of ALL1 (MLL) is consistentlygenerated by Alu-mediated homologous recombination in acutemyeloid leukemia. Proc. Natl. Acad. Sci. USA 95, 2390–2395.
Sugawara, N., Goldfarb, T., Studamire, B., Alani, E., and Haber, J.E.(2004). Heteroduplex rejection during single-strand annealing re-quires Sgs1 helicase and mismatch repair proteins Msh2 and Msh6but not Pms1. Proc. Natl. Acad. Sci. USA 101, 9315–9320.
Takata, M., Sasaki, M.S., Sonoda, E., Morrison, C., Hashimoto, M.,Utsumi, H., Yamaguchi-Iwai, Y., Shinohara, A., and Takeda, S.(1998). Homologous recombination and non-homologous end-join-ing pathways of DNA double-strand break repair have overlappingroles in the maintenance of chromosomal integrity in vertebratecells. EMBO J. 17, 5497–5508.
van Gent, D.C., Hoeijmakers, J.H., and Kanaar, R. (2001). Chromo-somal stability and the DNA double-stranded break connection.Nat. Rev. Genet. 2, 196–206.
Yu, X., and Gabriel, A. (2004). Reciprocal translocations in Sacchar-omyces cerevisiae formed by nonhomologous end joining. Genet-ics 166, 741–751.
Zucman-Rossi, J., Legoix, P., Victor, J.M., Lopez, B., and Thomas,G. (1998). Chromosome translocation based on illegitimate recom-bination in human tumors. Proc. Natl. Acad. Sci. USA 95, 11786–11791.