chromium (d-phenylalanine)3 improves obesity-induced cardiac contractile defect in ob/ob mice
TRANSCRIPT

Pharmacology and Therapeutics
Chromium (D-Phenylalanine)3 ImprovesObesity-Induced Cardiac Contractile Defect inob/ob MiceFeng Dong,* Xiaoping Yang,* Nair Sreejayan, and Jun Ren
AbstractDONG, FENG, XIAOPING YANG, NAIR SREEJAYAN,AND JUN REN. Chromium (D-phenylalanine)3 improvesobesity-induced cardiac contractile defect in ob/ob mice.Obesity. 2007;15:2699–2711.Objective: Low-molecular weight chromium compounds,such as chromium picolinate [Cr(pic)3], improve insulinsensitivity, although toxicity is a concern. We synthesized anovel chromium complex, chromium (D-phenylalanine)3
[Cr(D-phe)3], in an attempt to improve insulin sensitivitywith reduced toxicity. The aim of this study was to comparethe two chromium compounds on cardiac contractile func-tion in ob/ob obese mice.Research Methods and Procedures: C57BL lean and ob/obobese mice were randomly divided into three groups: H2O,Cr(D-phe)3, or Cr(pic)3 (45 �g/kg per day orally for 6 months).Results: The glucose tolerance test displayed improvedglucose clearance by Cr(D-phe)3 but not Cr(pic)3. Myocytesfrom ob/ob mice exhibited depressed peak shortening (PS)and maximal velocity of shortening/relengthening (�dL/dt), prolonged time-to-PS and time-to-90% relengthening(TR90), reduced electrically stimulated rise in intracellularCa2� (�fura-2 fluorescence intensity), and slowed intracel-lular Ca2� decay. Although a 3-month Cr(D-phe)3 treatmentfor a separate group of ob/ob and lean 2-month-old miceonly rectified reduced �dL/dt in ob/ob mice, all mechanicaland intracellular Ca2� abnormalities were significantly at-tenuated or ablated by 6 months of Cr(D-phe)3 but not
Cr(pic)3 treatment (except TR90). Sarco(endo)plasmic re-ticulum Ca2� ATPase activity and Na�-Ca2� exchangerexpression were depressed in ob/ob mice, which were re-versed by both Cr(D-phe)3 and Cr(pic)3, with a more pro-nounced effect from Cr(D-phe)3. Cr(D-phe)3 corrected re-duced insulin-stimulated glucose uptake and improvedbasal phosphorylation of Akt and insulin receptor, as wellas insulin-stimulated phosphorylation of Akt and insulinreceptor in ob/ob myocytes. Heart homogenates from ob/obmice had enhanced oxidative stress and protein carbonylformation compared with the lean group, which were atten-uated by both Cr(D-phe)3 and Cr(pic)3.Discussion: Our data suggest that the new Cr(D-phe)3 com-pound possesses better cardio-protective and insulin-sensi-tizing properties against obesity.
Key words: cell signaling, obesity treatment, heart, diet
IntroductionAn ample amount of clinical and experimental evidence
has shown that uncorrected obesity leads to insulin resis-tance, cardiac hypertrophy, compromised myocardial func-tion, and energy metabolism, which contributes to enhancedcardiovascular morbidity and mortality and risk of develop-ing type 2 diabetes (1–3). A number of theories, includingoxidative stress, dyslipidemia, and hyperleptinemia, havebeen implicated to attribute to obesity-associated cardiacmorphological and functional defects (2,4–7). Nonetheless,proper management of obesity and insulin resistance has notbeen radically achieved so as to reduce the risk for cardio-vascular diseases in overweight or obese populations. Cur-rent therapeutic regimens including pharmacological treat-ment (such as biguanides and thiazolidinedione), caloricrestriction, and exercise have proven to be fairly effective,although none is deemed ideal. Search for effective, orallyactive drugs that can mimic or enhance the property ofinsulin is in high demand for the long-term management ofobesity and insulin resistance. One of the compounds draw-ing some attention is the mineral chromium, which is
Received for review December 2, 2006.Accepted in final form February 26, 2007.The costs of publication of this article were defrayed, in part, by the payment of pagecharges. This article must, therefore, be hereby marked “advertisement” in accordance with18 U.S.C. Section 1734 solely to indicate this fact.Center for Cardiovascular Research and Alternative Medicine and Division of Pharmaceu-tical Sciences, University of Wyoming, Laramie, Wyoming.* These authors contributed equally to the paper.Address correspondence to Jun Ren, Division of Pharmaceutical Sciences and Center forCardiovascular Research and Alternative Medicine, University of Wyoming, Laramie, WY82071.E-mail: [email protected] © 2007 NAASO
OBESITY Vol. 15 No. 11 November 2007 2699

thought to play a key role in carbohydrate metabolism bypotentiating the action of insulin, leading to increased insu-lin sensitivity in type 2 diabetes and obesity (8). Dietarydeficiency in chromium has been shown to be positivelyassociated with the risk of diabetes (9). Consistently, re-duced levels of chromium have been documented in dia-betic subjects with existing cardiovascular diseases (10,11).Clinical trials have indicated that supplementation withchromium chloride or low-molecular weight complexes ofchromium can lower blood glucose levels in the diabeticpatient (12). Recent evidence suggests that low-molecularweight organic chromium complexes possess better bio-availability compared with chromium salts (2% to 5% vs.0.5% to 2%), leading to the development of low-molecularweight organic complexes of chromium as therapeuticagents to counter the diminished insulin effect (13). Chro-mium picolinate [Cr(pic)3],1 the most popularly used dietarysupplement, has been shown to modulate glucose metabo-lism and improve dyslipidemia associated with insulin re-sistance in several animal and human studies (14,15). How-ever, Cr(pic)3 has been recently shown to cause deleteriousDNA mutation potentially by its prooxidant property (16).The prooxidant nature of Cr(pic)3 has greatly limited itstherapeutic application and prompted us to seek safer syn-thetic chromium complexes with a natural amino acid li-gand (16). Our laboratory has recently synthesized a low-molecular weight complex, chromium (D-phenylalanine)3
[Cr(D-phe)3], which enhances insulin signaling and glucoseuptake in adipocytes and improves glucose clearance in anob/ob obese model of insulin resistance with lesser cytotox-icity compared with Cr(pic)3 (13,17). Nonetheless, the ef-fect of Cr(D-phe)3 on heart function under obesity andinsulin resistance has not been elucidated. To assess theoverall therapeutic efficacy of the newly synthesized Cr(D-phe)3, the current study was designed to evaluate the effectof chronic treatment of Cr(D-phe)3 on cardiomyocyte con-traction, intracellular Ca2� ([Ca2�]i), oxidative stress, pro-tein damage, glucose uptake, and the insulin signaling cas-cade with an emphasis on insulin receptor tyrosinephosphorylation and post-receptor Akt in ob/ob obese insu-lin-resistant mice. For comparison, the effect of the classicchromium compound Cr(pic)3 was also examined in parallelto that of Cr(D-phe)3.
Research Methods and ProceduresSynthesis and Characterization of Cr(D-phe)3
Aqueous solutions of CrCl3.6H2O (2.6 grams; 10 mM in50 mL H2O) and D-phenylalanine (4.8 grams, 30 mM in 50mL H2O) were mixed at 80 °C and refluxed for 4 hours. Thehomogeneous green reaction mixture was freeze-dried. Thegreenish-violet solid obtained was washed with acetone anddried in an air oven. The yield was 81%, and the meltingpoint was �300 °C; we found C, 47.84; H, 5.60; and N,5.92. The stoichiometry Cr(C27H30N3O6).3HCl.2H2O re-quires C, 47.00; H, 5.40; and N, 6.09. The electrospray massspectrometry of the complex in methanolic solution regis-ters signals at 545.1 and 165.9, representing the Tris chelateand the deprotonated ligand, respectively. Formation of thecomplex was associated with �C � O (1563 cm�1) and �N-H
(3535 cm�1) shifts in the infrared spectrum by �40 and 30cm�1, respectively. The broadening of the moderately sharpabsorption band in the free ligand (2900 to 3100 cm�1) to�600 cm�1 may be attributed to the reorganization inintramolecular hydrogen bonding after chelation. New ab-sorption bands in the far infrared region, �370 and 310cm�1, can be assigned to the Cr-O and Cr-N bonds. Theultraviolet-visible spectrum of the methanolic solution ofthe complex registered bands at 15,673 (�1) and 22,075 (�2)cm�1. The complex being green in color, the above twobands are due to the absorption in yellow and blue parts ofthe spectrum. These absorptions are due to the spin-allowedtransitions: 4T2 grams 4
4A2 grams (�1) and 4T1 grams(F)4 4A2 grams (�2). The third band, �3, overlaps with ultra-violet absorption of the ligand. These observations suggesta hexa-coordinate environment around chromium (III). ThepH of the aqueous solution of the complex is 4.7, and thepresence of chloride demonstrates the presence of HCl inthe lattice. Based on the stoichiometry, elemental analysis,and spectral studies, the product obtained is a complexcontaining a 1:3 ratio of chromium to phenylalanine (13).
Experimental Animals and Plasma MeasurementThe experimental procedures described in this study were
approved by the University of Wyoming (Laramie, WY)Animal Use and Care Committee. In brief, homozygousB6.V-lep�ob�/J male mice were obtained from the Jack-son Laboratory (Bar Harbor, ME) at 5 weeks of age andwere housed within the School of Pharmacy Animal Facilityat the University of Wyoming with free access to food andwater. The ob/ob obese (leptin-deficient) mice and age-matched wild-type C57BL/6J mice were each randomlydivided into three groups: control (H2O), Cr(D-phe)3, andCr(pic)3. The drugs were provided in drinking water at adose of 45 �g/kg per day for 6 months starting at 2 monthsof age for the mice. For the 3-month Cr(D-phe)3 treatment,a high dose at 150 �g/kg per day was employed using aseparate group of lean and ob/ob mice (2-month-old). Bloodglucose and plasma leptin levels were measured by an
1 Nonstandard abbreviations: Cr(pic)3, chromium picolinate; Cr(D-phe)3, chromium (D-phenylalanine)3; [Ca2�]i, intracellular Ca2�; IPGTT, intraperitoneal glucose tolerance test;BW, body weight; KHB, Krebs-Henseleit bicarbonate; PS, peak shortening; TPS, time-to-PS; TR90, time-to-90% relengthening; �dL/dt, maximal velocity of shortening; �dL/dt,maximal velocity of relengthening; FFI, fura-2 fluorescence intensity; NCX, Na�-Ca2�
exchanger; SDS, sodium dodecyl sulfate; SERCA, sarco(endo)plasmic reticulum Ca2�
ATPase; GSH, glutathione; GSSG, glutathione disulfide; SE, standard error; HW, heartweight.
A Novel Chromium Complex and Cardiac Function, Dong et al.
2700 OBESITY Vol. 15 No. 11 November 2007
Accu-Chek glucometer (Boehringer Mannheim Diagnos-tics, Indianapolis, IN) and radioimmunoassay (Linco Re-search, St. Charles, MO), respectively. Blood insulin levelswere measured using a mouse insulin enzyme-linked im-munosorbent assay kit (LINCO Research).
Intraperitoneal Glucose Tolerance Test (IPGTT)Two days before sacrifice, mice fasted for 12 hours were
given an intraperitoneal challenge of glucose [2 g/kg bodyweight (BW)]. Blood samples were drawn from the tailveins immediately before the glucose challenge and 15, 60,and 120 minutes thereafter. Blood glucose levels were de-termined using a glucometer. Area underneath the IPGTTcurve was determined using the GraphPad Prism Software(GraphPad Software, Inc., San Diego, CA).
Isolation of Mouse CardiomyocytesSingle murine cardiomyocytes were enzymatically iso-
lated as described (18). Briefly, hearts were removed andperfused (at 37 °C) with oxygenated (5% CO2-95% O2)Krebs-Henseleit bicarbonate (KHB) buffer containing 118mM NaCl, 4.7 mM KCl, 1.25 mM CaCl2, 1.2 mM MgSO4,1.2 mM KH2PO4, 25 mM NaHCO3, 10 mM HEPES, and11.1 mM glucose. Hearts were subsequently perfused witha Ca2�-free KHB buffer containing 223 U/mL collagenaseD (Worthington Biochemical, Freehold, NJ) for 20 minutes.After perfusion, left ventricles were removed and minced todisperse individual cardiomyocytes in Ca2�-free KHBbuffer. Extracellular Ca2� was added incrementally back toa final concentration of 1.25 mM. Myocytes with obvious
sarcolemmal blebs or spontaneous contractions were notused. Only rod-shaped myocytes with clear edges wereselected for recording of mechanical and [Ca2�]i measure-ments. Neither obesity nor chromium treatment [Cr(D-phe)3
or Cr(pic)3] significantly affected the yield of murine car-diomyocytes.
Cell Shortening/RelengtheningMechanical properties of cardiomyocytes were assessed
using an IonOptix MyoCam system (IonOptix Corp., Mil-ton, MA) (18). In brief, myocytes were placed in a chambermounted on the stage of an inverted microscope and super-fused with a buffer containing 131 mM NaCl, 4 mM KCl, 1mM CaCl2, 1 mM MgCl2, 10 mM glucose, and 10 mMHEPES (pH 7.4). The cells were field stimulated withsuprathreshold voltage and at a frequency of 0.5 Hz (3-msduration) with the use of a pair of platinum wires placed onopposite sides of the chamber connected to an FHC stimu-lator (Brunswick, NE). The polarity of stimulatory elec-trodes was reversed frequently to avoid possible build up ofthe electrolyte byproducts. The myocyte being studied wasdisplayed on the computer monitor with the use of anIonOptix MyoCam camera, which rapidly scans the imagearea at every 8.3 ms such that the amplitude and velocity ofshortening/relengthening are recorded with good fidelity.SoftEdge software (IonOptix) was used to capture changesin cell length during shortening and relengthening. Cellshortening and relengthening were assessed using the fol-lowing indices: peak shortening (PS), time-to-PS (TPS),
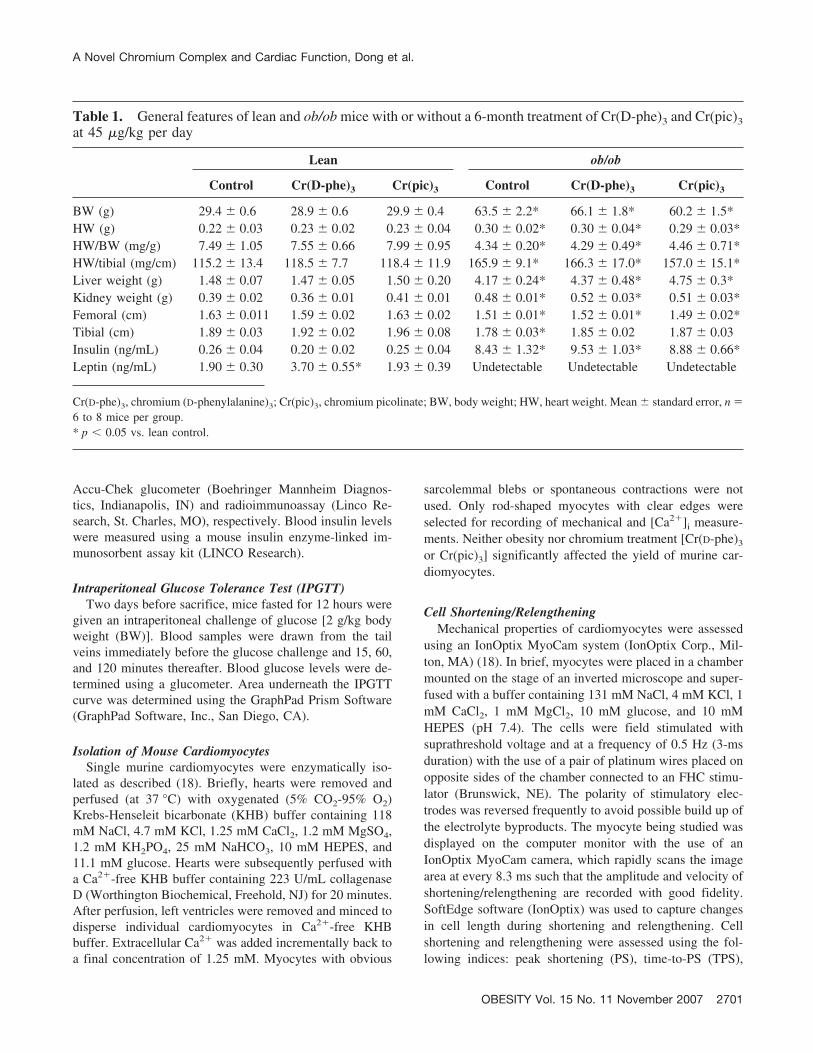
Table 1. General features of lean and ob/ob mice with or without a 6-month treatment of Cr(D-phe)3 and Cr(pic)3at 45 �g/kg per day
Lean ob/ob
Control Cr(D-phe)3 Cr(pic)3 Control Cr(D-phe)3 Cr(pic)3
BW (g) 29.4 � 0.6 28.9 � 0.6 29.9 � 0.4 63.5 � 2.2* 66.1 � 1.8* 60.2 � 1.5*HW (g) 0.22 � 0.03 0.23 � 0.02 0.23 � 0.04 0.30 � 0.02* 0.30 � 0.04* 0.29 � 0.03*HW/BW (mg/g) 7.49 � 1.05 7.55 � 0.66 7.99 � 0.95 4.34 � 0.20* 4.29 � 0.49* 4.46 � 0.71*HW/tibial (mg/cm) 115.2 � 13.4 118.5 � 7.7 118.4 � 11.9 165.9 � 9.1* 166.3 � 17.0* 157.0 � 15.1*Liver weight (g) 1.48 � 0.07 1.47 � 0.05 1.50 � 0.20 4.17 � 0.24* 4.37 � 0.48* 4.75 � 0.3*Kidney weight (g) 0.39 � 0.02 0.36 � 0.01 0.41 � 0.01 0.48 � 0.01* 0.52 � 0.03* 0.51 � 0.03*Femoral (cm) 1.63 � 0.011 1.59 � 0.02 1.63 � 0.02 1.51 � 0.01* 1.52 � 0.01* 1.49 � 0.02*Tibial (cm) 1.89 � 0.03 1.92 � 0.02 1.96 � 0.08 1.78 � 0.03* 1.85 � 0.02 1.87 � 0.03Insulin (ng/mL) 0.26 � 0.04 0.20 � 0.02 0.25 � 0.04 8.43 � 1.32* 9.53 � 1.03* 8.88 � 0.66*Leptin (ng/mL) 1.90 � 0.30 3.70 � 0.55* 1.93 � 0.39 Undetectable Undetectable Undetectable
Cr(D-phe)3, chromium (D-phenylalanine)3; Cr(pic)3, chromium picolinate; BW, body weight; HW, heart weight. Mean � standard error, n �6 to 8 mice per group.* p � 0.05 vs. lean control.
A Novel Chromium Complex and Cardiac Function, Dong et al.
OBESITY Vol. 15 No. 11 November 2007 2701

time-to-90% relengthening (TR90), maximal velocity ofshortening (�dL/dt), and maximal velocity of relengthening(�dL/dt).
[Ca2�]i Fluorescence MeasurementA separate cohort of myocytes was loaded with fura-
2/AM (0.5 �M) for 15 minutes, and fluorescence intensitywas measured with a dual-excitation fluorescence photo-multiplier system (Ionoptix) (18). Myocytes were placed onan inverted Olympus microscope and imaged through aFluor 40 oil objective. Cells were exposed to light emittedby a 75-W mercury lamp and passed through either a 360-or a 380-nm filter. The myocytes were stimulated to con-tract at 0.5 Hz. Fluorescence emissions were detected be-tween 480 and 520 nm by a photomultiplier tube after cellswere first illuminated at 360 nm for 0.5 seconds and then at380 nm for the duration of the recording protocol (333-Hzsampling rate). The 360-nm excitation scan was repeated atthe end of the protocol, and qualitative change in fura-2fluorescence intensity (FFI) was inferred from the ratio offluorescence intensity at the two wavelengths (360/380).Fluorescence decay time was calculated as an indicator of[Ca2�]i clearing.
Glucose Uptake MeasurementCardiomyocytes were washed three times with Krebs-
Ringer-HEPES buffer containing 136 mM NaCl, 4.7 mMKCl, 1.25 mM CaCl2, 1.25 mM MgSO4, and 10 mMHEPES (pH 7.4) and were incubated at 37 °C for 30 min-utes. Some cells were exposed to insulin (100 nM) for 10minutes. Glucose uptake was initiated by the addition of 0.1mL of KRH buffer and 2-deoxy-D-[3H] glucose (0.2 �Ci/mL) and 5 mM glucose. Glucose uptake was terminated 30minutes later by washing the cells three times with coldphosphate-buffered saline. The cells were lysed overnightwith 0.5 mL of 0.5 M NaOH and 0.1% (w/v) sodiumdodecyl sulfate (SDS). The radioactivity retained by celllysates was determined by a scintillation counter (BeckmanLC 6000IC; Beckman Coulter, Fullerton, CA) and normal-ized to protein content (18).
Sarco(endo)plasmic Reticulum Ca2� ATPase (SERCA)Activity Measured by 45Ca2� Uptake
Cardiomyocytes were sonicated and solubilized in a Tris-sucrose homogenization buffer consisting of 30 mM Tris-HCl, 8% sucrose, 1 mM phenylmethylsulfonylfluoride, and2 mM dithiothreitol (pH 7.1). To determine SERCA-depen-dent Ca2� uptake, myocytes were treated with and withoutthe SERCA inhibitor thapsigargin (10 �M) for 15 minutes.The difference between the two readings was deemed thethapsigargin-sensitive SERCA uptake. Uptake was initiatedby the addition of an aliquot of supernatant to a solutionconsisting of 100 mM KCl, 5 mM NaN3, 6 mM MgCl2, 0.15mM EGTA, 0.12 mM CaCl2, 30 mM Tris-HCl (pH 7.0), 10
mM oxalate, 2 mM ATP, and 1 mM �Ci 45CaCl2 at 37 °C.Aliquots of samples were injected onto glass filters on asuction manifold and washed three times. Filters were thenremoved from the manifold, placed in scintillation fluid andcounted. SERCA activity was expressed as cpm/mg protein(19).
Glutathione (GSH) and Glutathione Disulfide (GSSG)Assay
GSH levels were determined in cardiac tissue as anindication of oxidative stress. For measurement of GSH,pellets of cardiomyocytes were homogenized in 4 volumes(w/v) of 1% picric acid. Acid homogenates were centrifugedat 16,000g (30 minutes), and supernatant fractions werecollected. Supernatant fractions were assayed for total GSHand GSSG by the standard recycling method, and GSHcontent was determined using a standard curve generatedfrom known concentrations of GSH. The procedure con-sisted of using one-half of each sample for GSSG determi-nation and the other one-half for GSH. Samples for GSSGdetermination were incubated at 4 °C with 2 �L of 4-vinylpyridine per 100 �L sample for 1 hour after vigorousvortexing. Incubation with 4-vinyl pyridine conjugates anyGSH present in the sample so that only GSSG is recycled toGSH without interference by GSH. The GSSG (as GSHx2)was then subtracted from the total GSH to determine actualGSH level (20).
Protein Carbonyl AssayProtein carbonyl content of the total protein lysate was
determined as described (18). In brief, protein was precip-itated by adding an equal volume of 20% trichloroaceticacid and centrifuged for 1 minute. The sample resuspendedin 10 mM 2,4-dinitrophenylhydrazine solution for 15 to 30minutes at room temperature before 20% trichloroaceticacid was added, and samples were centrifuged for 3 min-utes. The precipitate was resuspended in 6 M guanidinesolution. The maximum absorbance (360 to 390 nm) wasread against appropriate blanks (2 M HCl), and carbonylcontent was calculated using the formula: absorption at 360nm 45.45 nM/protein content (milligrams).
Immunoblot of Akt, pAkt, Insulin Receptor TyrosinePhosphorylation, and Na�-Ca2� Exchanger (NCX)
Left ventricles were rapidly removed and homogenized ina lysis buffer containing 20 mM Tris (pH 7.4), 150 mMNaCl, 1 mM EDTA, 1 mM EGTA, 1% Triton, 0.1% SDS,and 1% protease inhibitor cocktail. Samples were then son-icated for 15 seconds and centrifuged at 12,000g for 20minutes at 4 °C. The protein concentration of the superna-tant was evaluated using the Protein Assay Reagent (Bio-Rad Laboratories, Inc., Hercules, CA). Equal amounts (50�g protein/lane) of protein and prestained molecular weightmarker (Gibco-BRL, Gaithersburg, MD) were loaded onto
A Novel Chromium Complex and Cardiac Function, Dong et al.
2702 OBESITY Vol. 15 No. 11 November 2007

7% SDS-polyacrylamide gels in a minigel apparatus (Mini-PROTEAN II; Bio-Rad) before being separated and trans-ferred to nitrocellulose membranes (0.2-�m pore size; Bio-Rad). Membranes were incubated for 1 hour in a blockingsolution containing 5% non-fat milk in Tris-buffered saline,washed in Tris-buffered saline, and incubated overnight at4 °C with an anti-Akt (1:1000), anti-pAkt (1:1000), anti-NCX (1:1000), anti-phosphoinsulin receptor �/insulin-likegrowth factor receptor (1:1000), or �-actin (1:5000, loadingcontrol) antibody. Rabbit polyclonal anti-NCX antibodywas purchased from Swant (Bellinzona, Switzerland), andall other antibodies were obtained from Cell Signaling(Beverly, MA). After incubation with the primary antibody,blots were incubated with an anti-mouse IgG horseradishperoxidase-linked antibody at a dilution of 1:5000 for 1hour at room temperature. Immunoreactive bands were de-tected using the Super Signal West Dura Extended DurationSubstrate (Pierce, Milwaukee, WI). The intensity of bandswas measured with a scanning densitometer (Model GS-800; Bio-Rad) coupled with Bio-Rad PC analysis software(18).
Statistical AnalysesData are presented as mean � standard error (SE). Sta-
tistical significance (p � 0.05) for each variable was eval-uated by ANOVA followed by a Dunnett’s post hoc anal-ysis.
ResultsGeneral Features of Lean and ob/ob Mice Treated withCr(D-phe)3 and Cr(pic)3
As expected, ob/ob mice displayed a significantly greaterBW, heart weight (HW), liver weight, and kidney weightcompared with age-matched lean controls. Femoral andtibial lengths were slightly, but significantly, shorter in theob/ob group. Although the heart-to-BW ratio was signifi-cantly reduced in the ob/ob group, which is likely due tooverweight, the HW-to-tibial length was significantly in-creased, indicating cardiac hypertrophy. The plasma leptinlevels were undetectable, whereas blood insulin levels weresignificantly elevated in ob/ob mice. Chronic treatment withCr(D-phe)3 or Cr(pic)3 did not affect body and organ weightand length of femoral and tibial bones in both lean andob/ob groups. Interestingly, Cr(D-phe)3 but not Cr(pic)3
significantly elevated plasma leptin levels in lean mice.Neither chromium complex affected leptin levels in ob/obmice. Blood insulin levels were not affected by either chro-mium complex. After acute intraperitoneal glucose chal-lenge, the blood glucose levels in lean mice started to dropafter peaking at 15 minutes and returned to nearly baselineafter 120 minutes. However, the post-challenge glucoselevels maintained at high levels from 15 to 120 minutes inob/ob mice, indicative of glucose intolerance and insulin
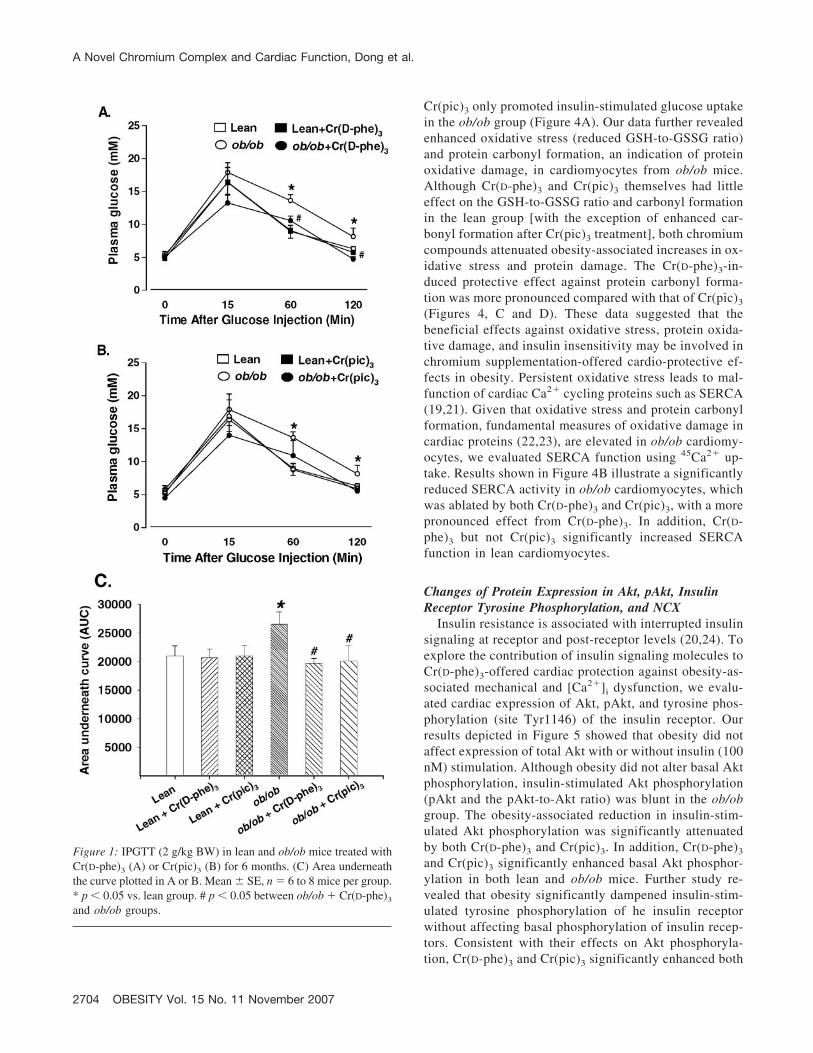
resistance. Although neither chromium compound affectedthe glucose clearance rate in lean mice, Cr(D-phe)3 signifi-cantly facilitated the glucose clearance in ob/ob mice withlower plasma glucose levels at 60 and 120 minutes com-pared with ob/ob mice without chromium treatment.Cr(pic)3 exhibited a tendency of reducing plasma glucoselevels in ob/ob mice, although no statistical significancewas reached. However, calculation of the area underneaththe glucose clearance curve displayed that both Cr(D-phe)3
and Cr(pic)3 significantly alleviated obesity-induced in-crease in area underneath the curve, indicating beneficialeffects of both chromium complexes against glucose intol-erance. There was no difference in basal fasting glucoselevels among all six mouse groups, excluding the presenceof full-blown diabetes (Figure 1).
Mechanical and Fluorescent Properties of Myocytesfrom Lean and ob/ob Mice
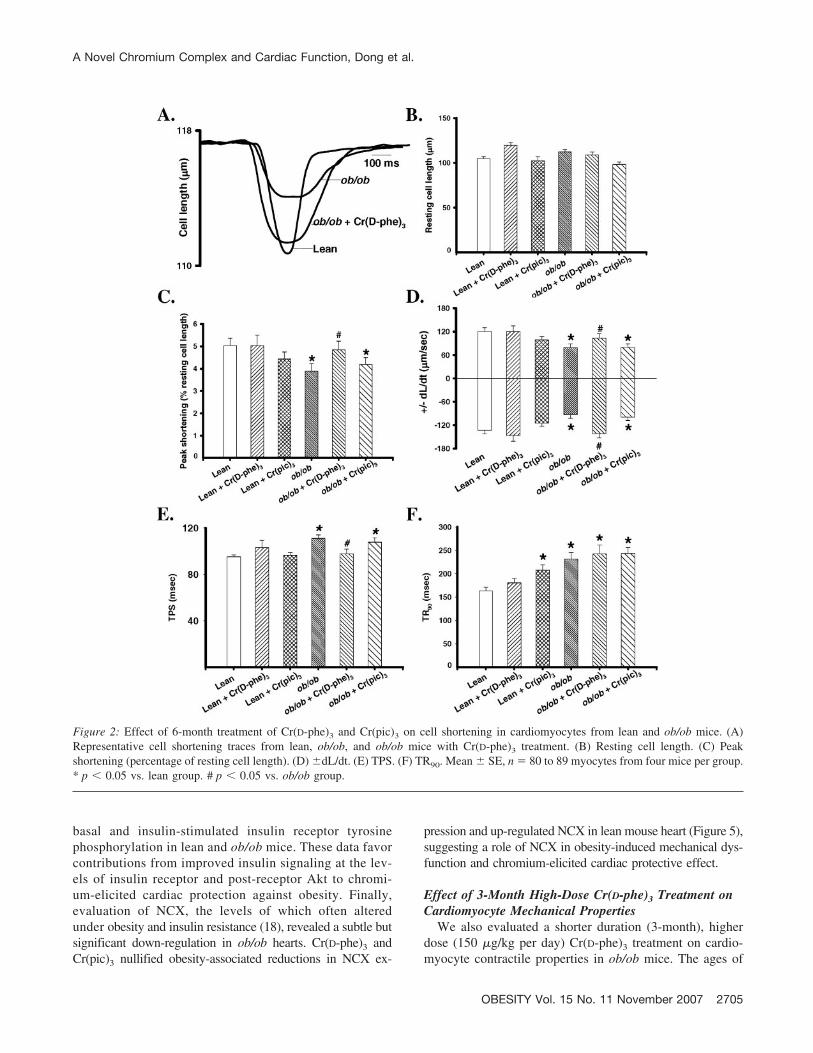
Neither obesity nor chromium treatment significantly af-fected the resting cell length of cardiomyocytes. Cardiomy-ocytes from ob/ob mice displayed significantly depressedPS amplitude and �dL/dt and prolonged TPS and TR90
compared with those from lean controls. Interestingly, Cr(D-phe)3 but not Cr(pic)3 treatment effectively corrected ab-normal PS, �dL/dt, and TPS without affecting prolongedTR90 in cardiomyocyte from ob/ob mice. Neither chromiumcompound significantly affected mechanical indices in leancardiomyocytes, with the exception of a prolonged TR90
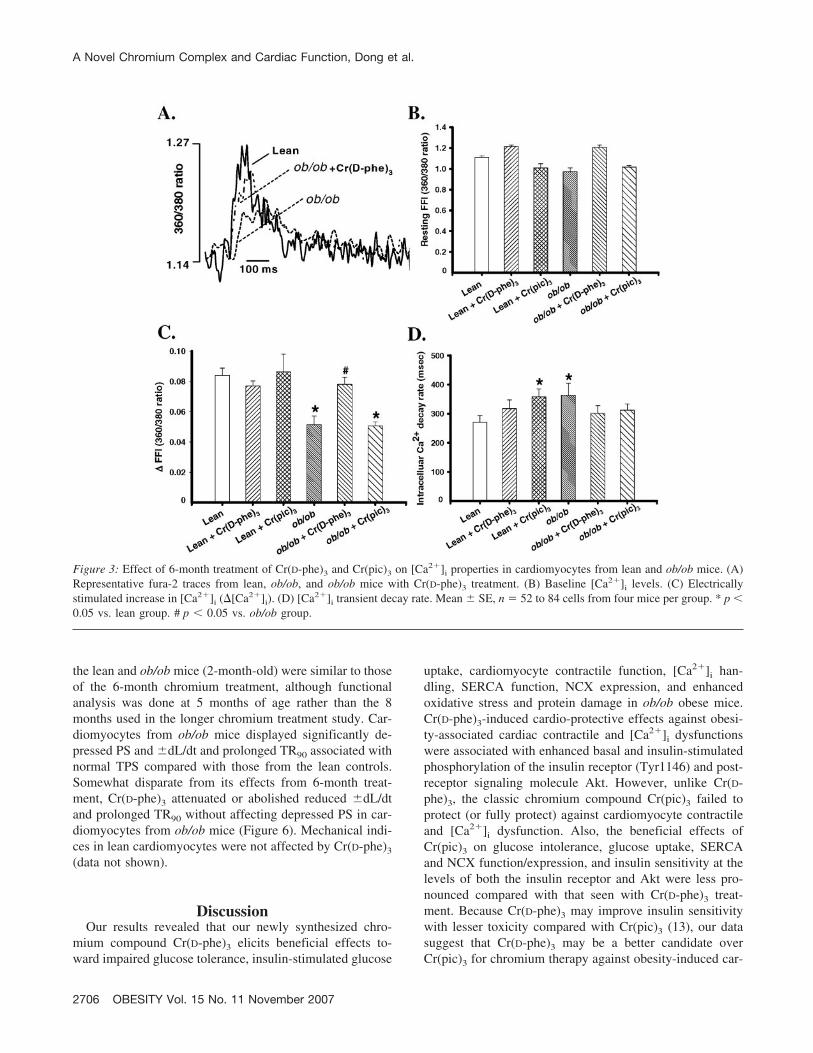
after Cr(pic)3 treatment (Figure 2). Our fura-2 study re-vealed depressed electrically stimulated �FFI and slowed[Ca2�]i clearing rate associated with normal resting FFI inmyocytes from ob/ob mice, consistent with reduced PS andprolonged TR90 in these myocytes. Somewhat similar totheir effects on cell shortening, Cr(D-phe)3 but not Cr(pic)3
reconciled the obesity-induced decrease in �FFI, whereasboth compounds negated prolonged [Ca2�]i clearing inob/ob cardiomyocytes. Neither chromium compound af-fected the resting [Ca2�]i levels (resting FFI), �FFI, and[Ca2�]i decay in lean cardiomyocytes with the exception ofdelayed [Ca2�]i decay after Cr(pic)3 treatment (Figure 3).
Effect of Chromium Supplementation on GlucoseUptake, SERCA Activity, Oxidative Stress, and ProteinCarbonyl Formation
To determine whether insulin sensitivity is involved inchromium supplementation-induced cardiomyocyte con-tractile and [Ca2�]i responses, basal and insulin-stimulatedglucose uptake was determined. Basal glucose uptake,which was significantly reduced by obesity, was dramati-cally improved by both Cr(D-phe)3 and Cr(pic)3 in both leanand ob/ob groups. Insulin (100 nM) significantly stimu-lated glucose uptake in lean but not ob/ob cardiomyo-cytes. Cr(D-phe)3 enhanced insulin-stimulated glucoseuptake in both lean and ob/ob cardiomyocytes, whereas
A Novel Chromium Complex and Cardiac Function, Dong et al.
OBESITY Vol. 15 No. 11 November 2007 2703
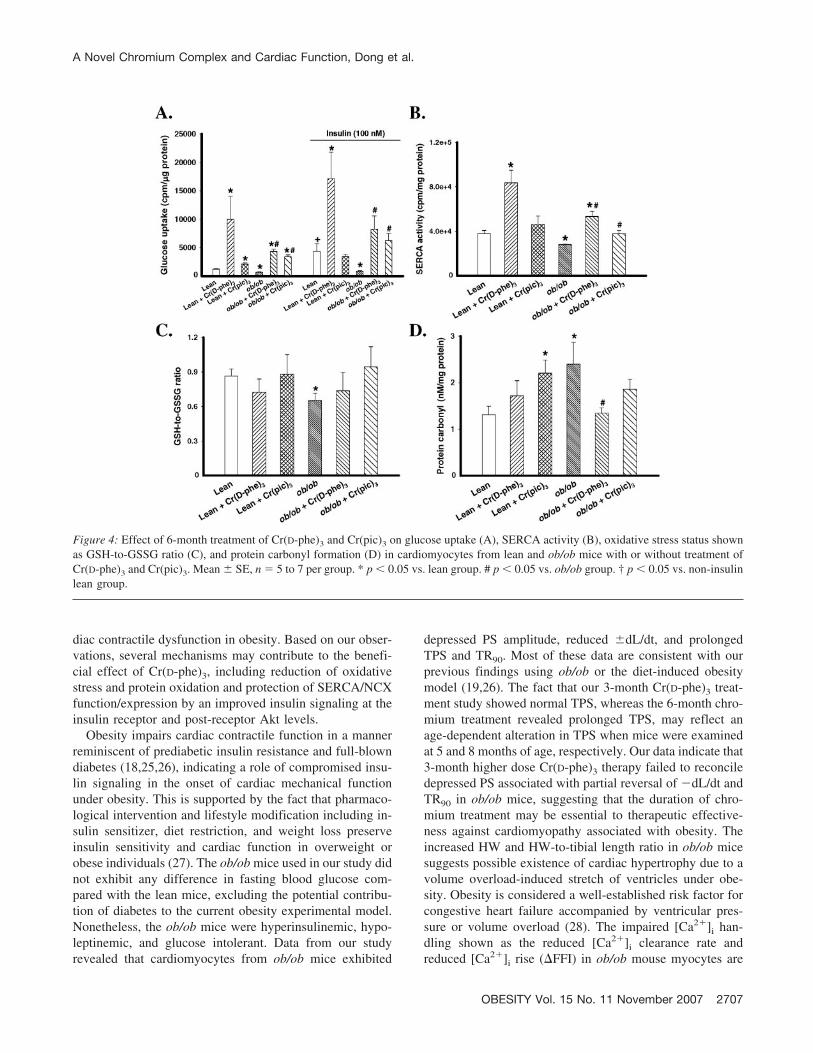
Cr(pic)3 only promoted insulin-stimulated glucose uptakein the ob/ob group (Figure 4A). Our data further revealedenhanced oxidative stress (reduced GSH-to-GSSG ratio)and protein carbonyl formation, an indication of proteinoxidative damage, in cardiomyocytes from ob/ob mice.Although Cr(D-phe)3 and Cr(pic)3 themselves had littleeffect on the GSH-to-GSSG ratio and carbonyl formationin the lean group [with the exception of enhanced car-bonyl formation after Cr(pic)3 treatment], both chromiumcompounds attenuated obesity-associated increases in ox-idative stress and protein damage. The Cr(D-phe)3-in-duced protective effect against protein carbonyl forma-tion was more pronounced compared with that of Cr(pic)3
(Figures 4, C and D). These data suggested that thebeneficial effects against oxidative stress, protein oxida-tive damage, and insulin insensitivity may be involved inchromium supplementation-offered cardio-protective ef-fects in obesity. Persistent oxidative stress leads to mal-function of cardiac Ca2� cycling proteins such as SERCA(19,21). Given that oxidative stress and protein carbonylformation, fundamental measures of oxidative damage incardiac proteins (22,23), are elevated in ob/ob cardiomy-ocytes, we evaluated SERCA function using 45Ca2� up-take. Results shown in Figure 4B illustrate a significantlyreduced SERCA activity in ob/ob cardiomyocytes, whichwas ablated by both Cr(D-phe)3 and Cr(pic)3, with a morepronounced effect from Cr(D-phe)3. In addition, Cr(D-phe)3 but not Cr(pic)3 significantly increased SERCAfunction in lean cardiomyocytes.
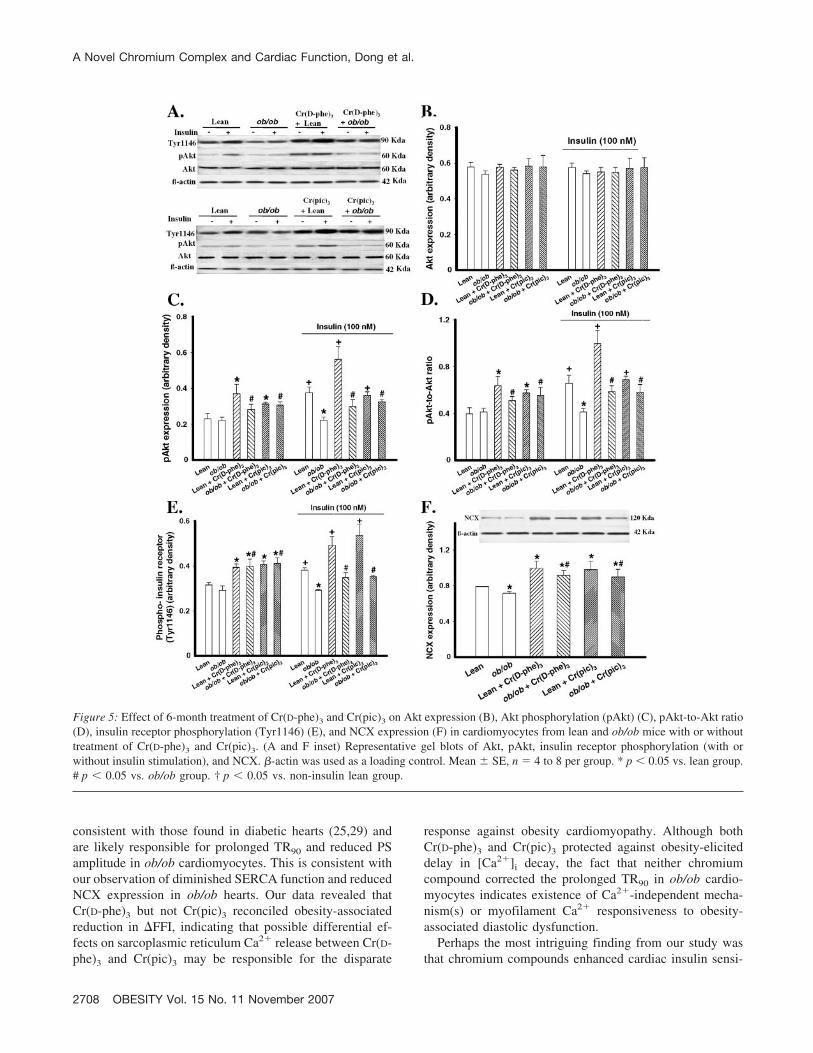
Changes of Protein Expression in Akt, pAkt, InsulinReceptor Tyrosine Phosphorylation, and NCX
Insulin resistance is associated with interrupted insulinsignaling at receptor and post-receptor levels (20,24). Toexplore the contribution of insulin signaling molecules toCr(D-phe)3-offered cardiac protection against obesity-as-sociated mechanical and [Ca2�]i dysfunction, we evalu-ated cardiac expression of Akt, pAkt, and tyrosine phos-phorylation (site Tyr1146) of the insulin receptor. Ourresults depicted in Figure 5 showed that obesity did notaffect expression of total Akt with or without insulin (100nM) stimulation. Although obesity did not alter basal Aktphosphorylation, insulin-stimulated Akt phosphorylation(pAkt and the pAkt-to-Akt ratio) was blunt in the ob/obgroup. The obesity-associated reduction in insulin-stim-ulated Akt phosphorylation was significantly attenuatedby both Cr(D-phe)3 and Cr(pic)3. In addition, Cr(D-phe)3
and Cr(pic)3 significantly enhanced basal Akt phosphor-ylation in both lean and ob/ob mice. Further study re-vealed that obesity significantly dampened insulin-stim-ulated tyrosine phosphorylation of he insulin receptorwithout affecting basal phosphorylation of insulin recep-tors. Consistent with their effects on Akt phosphoryla-tion, Cr(D-phe)3 and Cr(pic)3 significantly enhanced both
Figure 1: IPGTT (2 g/kg BW) in lean and ob/ob mice treated withCr(D-phe)3 (A) or Cr(pic)3 (B) for 6 months. (C) Area underneaththe curve plotted in A or B. Mean � SE, n � 6 to 8 mice per group.* p � 0.05 vs. lean group. # p � 0.05 between ob/ob � Cr(D-phe)3
and ob/ob groups.
A Novel Chromium Complex and Cardiac Function, Dong et al.
2704 OBESITY Vol. 15 No. 11 November 2007
basal and insulin-stimulated insulin receptor tyrosinephosphorylation in lean and ob/ob mice. These data favorcontributions from improved insulin signaling at the lev-els of insulin receptor and post-receptor Akt to chromi-um-elicited cardiac protection against obesity. Finally,evaluation of NCX, the levels of which often alteredunder obesity and insulin resistance (18), revealed a subtle butsignificant down-regulation in ob/ob hearts. Cr(D-phe)3 andCr(pic)3 nullified obesity-associated reductions in NCX ex-
pression and up-regulated NCX in lean mouse heart (Figure 5),suggesting a role of NCX in obesity-induced mechanical dys-function and chromium-elicited cardiac protective effect.
Effect of 3-Month High-Dose Cr(D-phe)3 Treatment onCardiomyocyte Mechanical Properties
We also evaluated a shorter duration (3-month), higherdose (150 �g/kg per day) Cr(D-phe)3 treatment on cardio-myocyte contractile properties in ob/ob mice. The ages of
Figure 2: Effect of 6-month treatment of Cr(D-phe)3 and Cr(pic)3 on cell shortening in cardiomyocytes from lean and ob/ob mice. (A)Representative cell shortening traces from lean, ob/ob, and ob/ob mice with Cr(D-phe)3 treatment. (B) Resting cell length. (C) Peakshortening (percentage of resting cell length). (D) �dL/dt. (E) TPS. (F) TR90. Mean � SE, n � 80 to 89 myocytes from four mice per group.* p � 0.05 vs. lean group. # p � 0.05 vs. ob/ob group.
A Novel Chromium Complex and Cardiac Function, Dong et al.
OBESITY Vol. 15 No. 11 November 2007 2705
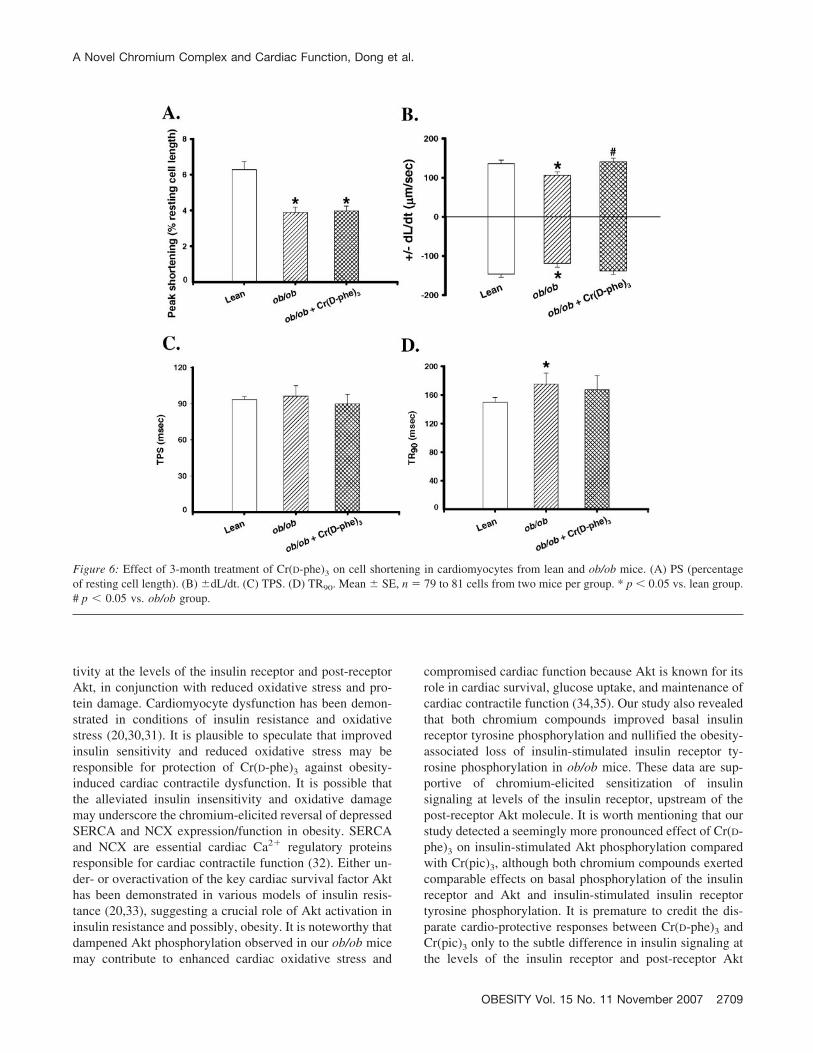
the lean and ob/ob mice (2-month-old) were similar to thoseof the 6-month chromium treatment, although functionalanalysis was done at 5 months of age rather than the 8months used in the longer chromium treatment study. Car-diomyocytes from ob/ob mice displayed significantly de-pressed PS and �dL/dt and prolonged TR90 associated withnormal TPS compared with those from the lean controls.Somewhat disparate from its effects from 6-month treat-ment, Cr(D-phe)3 attenuated or abolished reduced �dL/dtand prolonged TR90 without affecting depressed PS in car-diomyocytes from ob/ob mice (Figure 6). Mechanical indi-ces in lean cardiomyocytes were not affected by Cr(D-phe)3
(data not shown).
DiscussionOur results revealed that our newly synthesized chro-
mium compound Cr(D-phe)3 elicits beneficial effects to-ward impaired glucose tolerance, insulin-stimulated glucose
uptake, cardiomyocyte contractile function, [Ca2�]i han-dling, SERCA function, NCX expression, and enhancedoxidative stress and protein damage in ob/ob obese mice.Cr(D-phe)3-induced cardio-protective effects against obesi-ty-associated cardiac contractile and [Ca2�]i dysfunctionswere associated with enhanced basal and insulin-stimulatedphosphorylation of the insulin receptor (Tyr1146) and post-receptor signaling molecule Akt. However, unlike Cr(D-phe)3, the classic chromium compound Cr(pic)3 failed toprotect (or fully protect) against cardiomyocyte contractileand [Ca2�]i dysfunction. Also, the beneficial effects ofCr(pic)3 on glucose intolerance, glucose uptake, SERCAand NCX function/expression, and insulin sensitivity at thelevels of both the insulin receptor and Akt were less pro-nounced compared with that seen with Cr(D-phe)3 treat-ment. Because Cr(D-phe)3 may improve insulin sensitivitywith lesser toxicity compared with Cr(pic)3 (13), our datasuggest that Cr(D-phe)3 may be a better candidate overCr(pic)3 for chromium therapy against obesity-induced car-
Figure 3: Effect of 6-month treatment of Cr(D-phe)3 and Cr(pic)3 on [Ca2�]i properties in cardiomyocytes from lean and ob/ob mice. (A)Representative fura-2 traces from lean, ob/ob, and ob/ob mice with Cr(D-phe)3 treatment. (B) Baseline [Ca2�]i levels. (C) Electricallystimulated increase in [Ca2�]i (�[Ca2�]i). (D) [Ca2�]i transient decay rate. Mean � SE, n � 52 to 84 cells from four mice per group. * p �0.05 vs. lean group. # p � 0.05 vs. ob/ob group.
A Novel Chromium Complex and Cardiac Function, Dong et al.
2706 OBESITY Vol. 15 No. 11 November 2007
diac contractile dysfunction in obesity. Based on our obser-vations, several mechanisms may contribute to the benefi-cial effect of Cr(D-phe)3, including reduction of oxidativestress and protein oxidation and protection of SERCA/NCXfunction/expression by an improved insulin signaling at theinsulin receptor and post-receptor Akt levels.
Obesity impairs cardiac contractile function in a mannerreminiscent of prediabetic insulin resistance and full-blowndiabetes (18,25,26), indicating a role of compromised insu-lin signaling in the onset of cardiac mechanical functionunder obesity. This is supported by the fact that pharmaco-logical intervention and lifestyle modification including in-sulin sensitizer, diet restriction, and weight loss preserveinsulin sensitivity and cardiac function in overweight orobese individuals (27). The ob/ob mice used in our study didnot exhibit any difference in fasting blood glucose com-pared with the lean mice, excluding the potential contribu-tion of diabetes to the current obesity experimental model.Nonetheless, the ob/ob mice were hyperinsulinemic, hypo-leptinemic, and glucose intolerant. Data from our studyrevealed that cardiomyocytes from ob/ob mice exhibited
depressed PS amplitude, reduced �dL/dt, and prolongedTPS and TR90. Most of these data are consistent with ourprevious findings using ob/ob or the diet-induced obesitymodel (19,26). The fact that our 3-month Cr(D-phe)3 treat-ment study showed normal TPS, whereas the 6-month chro-mium treatment revealed prolonged TPS, may reflect anage-dependent alteration in TPS when mice were examinedat 5 and 8 months of age, respectively. Our data indicate that3-month higher dose Cr(D-phe)3 therapy failed to reconciledepressed PS associated with partial reversal of �dL/dt andTR90 in ob/ob mice, suggesting that the duration of chro-mium treatment may be essential to therapeutic effective-ness against cardiomyopathy associated with obesity. Theincreased HW and HW-to-tibial length ratio in ob/ob micesuggests possible existence of cardiac hypertrophy due to avolume overload-induced stretch of ventricles under obe-sity. Obesity is considered a well-established risk factor forcongestive heart failure accompanied by ventricular pres-sure or volume overload (28). The impaired [Ca2�]i han-dling shown as the reduced [Ca2�]i clearance rate andreduced [Ca2�]i rise (�FFI) in ob/ob mouse myocytes are
Figure 4: Effect of 6-month treatment of Cr(D-phe)3 and Cr(pic)3 on glucose uptake (A), SERCA activity (B), oxidative stress status shownas GSH-to-GSSG ratio (C), and protein carbonyl formation (D) in cardiomyocytes from lean and ob/ob mice with or without treatment ofCr(D-phe)3 and Cr(pic)3. Mean � SE, n � 5 to 7 per group. * p � 0.05 vs. lean group. # p � 0.05 vs. ob/ob group. † p � 0.05 vs. non-insulinlean group.
A Novel Chromium Complex and Cardiac Function, Dong et al.
OBESITY Vol. 15 No. 11 November 2007 2707
consistent with those found in diabetic hearts (25,29) andare likely responsible for prolonged TR90 and reduced PSamplitude in ob/ob cardiomyocytes. This is consistent withour observation of diminished SERCA function and reducedNCX expression in ob/ob hearts. Our data revealed thatCr(D-phe)3 but not Cr(pic)3 reconciled obesity-associatedreduction in �FFI, indicating that possible differential ef-fects on sarcoplasmic reticulum Ca2� release between Cr(D-phe)3 and Cr(pic)3 may be responsible for the disparate
response against obesity cardiomyopathy. Although bothCr(D-phe)3 and Cr(pic)3 protected against obesity-eliciteddelay in [Ca2�]i decay, the fact that neither chromiumcompound corrected the prolonged TR90 in ob/ob cardio-myocytes indicates existence of Ca2�-independent mecha-nism(s) or myofilament Ca2� responsiveness to obesity-associated diastolic dysfunction.
Perhaps the most intriguing finding from our study wasthat chromium compounds enhanced cardiac insulin sensi-
Figure 5: Effect of 6-month treatment of Cr(D-phe)3 and Cr(pic)3 on Akt expression (B), Akt phosphorylation (pAkt) (C), pAkt-to-Akt ratio(D), insulin receptor phosphorylation (Tyr1146) (E), and NCX expression (F) in cardiomyocytes from lean and ob/ob mice with or withouttreatment of Cr(D-phe)3 and Cr(pic)3. (A and F inset) Representative gel blots of Akt, pAkt, insulin receptor phosphorylation (with orwithout insulin stimulation), and NCX. �-actin was used as a loading control. Mean � SE, n � 4 to 8 per group. * p � 0.05 vs. lean group.# p � 0.05 vs. ob/ob group. † p � 0.05 vs. non-insulin lean group.
A Novel Chromium Complex and Cardiac Function, Dong et al.
2708 OBESITY Vol. 15 No. 11 November 2007
tivity at the levels of the insulin receptor and post-receptorAkt, in conjunction with reduced oxidative stress and pro-tein damage. Cardiomyocyte dysfunction has been demon-strated in conditions of insulin resistance and oxidativestress (20,30,31). It is plausible to speculate that improvedinsulin sensitivity and reduced oxidative stress may beresponsible for protection of Cr(D-phe)3 against obesity-induced cardiac contractile dysfunction. It is possible thatthe alleviated insulin insensitivity and oxidative damagemay underscore the chromium-elicited reversal of depressedSERCA and NCX expression/function in obesity. SERCAand NCX are essential cardiac Ca2� regulatory proteinsresponsible for cardiac contractile function (32). Either un-der- or overactivation of the key cardiac survival factor Akthas been demonstrated in various models of insulin resis-tance (20,33), suggesting a crucial role of Akt activation ininsulin resistance and possibly, obesity. It is noteworthy thatdampened Akt phosphorylation observed in our ob/ob micemay contribute to enhanced cardiac oxidative stress and
compromised cardiac function because Akt is known for itsrole in cardiac survival, glucose uptake, and maintenance ofcardiac contractile function (34,35). Our study also revealedthat both chromium compounds improved basal insulinreceptor tyrosine phosphorylation and nullified the obesity-associated loss of insulin-stimulated insulin receptor ty-rosine phosphorylation in ob/ob mice. These data are sup-portive of chromium-elicited sensitization of insulinsignaling at levels of the insulin receptor, upstream of thepost-receptor Akt molecule. It is worth mentioning that ourstudy detected a seemingly more pronounced effect of Cr(D-phe)3 on insulin-stimulated Akt phosphorylation comparedwith Cr(pic)3, although both chromium compounds exertedcomparable effects on basal phosphorylation of the insulinreceptor and Akt and insulin-stimulated insulin receptortyrosine phosphorylation. It is premature to credit the dis-parate cardio-protective responses between Cr(D-phe)3 andCr(pic)3 only to the subtle difference in insulin signaling atthe levels of the insulin receptor and post-receptor Akt
Figure 6: Effect of 3-month treatment of Cr(D-phe)3 on cell shortening in cardiomyocytes from lean and ob/ob mice. (A) PS (percentageof resting cell length). (B) �dL/dt. (C) TPS. (D) TR90. Mean � SE, n � 79 to 81 cells from two mice per group. * p � 0.05 vs. lean group.# p � 0.05 vs. ob/ob group.
A Novel Chromium Complex and Cardiac Function, Dong et al.
OBESITY Vol. 15 No. 11 November 2007 2709

levels. Further study is warranted to examine other check-points in the insulin signaling cascade, especially thosedirectly associated with [Ca2�]i homeostasis and cardiaccontractile function.
Our current knowledge of the clinical relevance of chro-mium is derived from the early reports on chromium intakeand risk of diabetes or insulin resistance. Patients receivingtotal parenteral nutrition devoid of chromium display signsof insulin resistance, which can be reversed by chromiumsupplementation, suggesting the role of chromium as anessential trace metal for carbohydrate and lipid metabolism(36). Further experimental and clinical studies have consol-idated the efficacy of chromium in the management of type2 diabetes and associated comorbidities such as insulinresistance and obesity (37,38). In a recent clinical trial,effects of chromium on cardiac function were assessed indiabetic patients with a focus on Q-T interval prolongation,a parameter associated with high fasting glucose levels, highplasma insulin levels, and increased fasting C-peptide levelsin diabetes. Chromium treatment was found to shorten theQ-T interval, with a more prominent effect in patients witha higher BMI (39). These observations justified the use ofchromium therapy to improve cardiovascular function inobese patients with severe insulin resistance. Nonetheless,poor bioavailability of inorganic chromium and the discov-ery that chromium exists as a complex with polypeptideslead to the synthesis and use of low-molecular weightchromium complexes (12,40). Nonetheless, the toxicity at-tributed to the ligand in Cr(pic)3, the most popular chro-mium supplement (16), has prompted the use of a non-toxicamino acid, D-phenylalanine, as the ligand to chelate chro-mium. The phenylalanine ligand displays a better solubilityat physiological pH while serving as a scavenger of hy-droxyl radicals (41). In fact, derivatives of D-phenylalanine,such as nateglinide, are also shown to be beneficial in type2 diabetes (42). Therefore, it is rational to use Cr(D-phe)3 asa highly bioavailable, less toxic, and more effective biomi-metic complex of chromium.
Although chromium has been found to reduce levels ofcholesterol and low-density lipoprotein (17,43), the lipid-lowering effect does not lead to alteration in total bodymass. Despite the potential clinical implication of chro-mium complexes in obesity and insulin-resistant conditions,the benefit vs. risk has not been carefully weighed forlong-term chromium treatment. Anderson et al. (14) re-vealed that chromium ingestion in both short (2 months) andlonger (4 months) terms displays beneficial effects onplasma glucose, insulin, and hemoglobin A1c concentra-tions. In another study, Cr(pic)3 supplementation signifi-cantly improved insulin sensitivity and glucose control andattenuated BW gain and visceral fat in type 2 diabeticpatients taking sulfonylurea (38). Further study is warrantedto scrutinize the longitudinal effect of chromium treatmentin diabetes, obesity, and insulin resistance.
In summary, our present study sheds some light on theeffect of a newly synthesized chromium compound Cr(D-phe)3 on cardiac contractile defects, [Ca2�]i mishandling,oxidative stress, protein damage, glucose intolerance, andcompromised insulin signaling in ob/ob obesity. Our datadepicted an advantage of Cr(D-phe)3 over the classic chro-mium compound Cr(pic)3 in terms of cardio-protective ef-fectiveness against obesity cardiomyopathy. Given that theprecise pathogenesis of aberrant cardiac contractile func-tion, [Ca2�]i homeostasis, cardiac Ca2� regulatory proteinfunction, oxidative stress status, and protein oxidative dam-age under obesity is far from clear, it is imperative for us todelineate the role of obesity and potential protective mech-anism(s) of chromium for a better usage of chromium inobesity and other insulin-resistant states.
AcknowledgmentsThis work was supported, in part, by the American Heart
Association Pacific Mountain Affiliate (to J.R.) and by theAmerican Diabetes Association (Junior Faculty Award toN.S.). There is no conflict of interest for any of the authors.
References1. Eckel RH, Barouch WW, Ershow AG. Report of the Na-
tional Heart, Lung, and Blood Institute-National Institute ofDiabetes and Digestive and Kidney Diseases Working Groupon the pathophysiology of obesity-associated cardiovasculardisease. Circulation. 2002;105:2923–8.
2. Sowers JR. Obesity as a cardiovascular risk factor. Am J Med.2003;115(Suppl 8A):37–41S.
3. Ren J, Sowers JR, Walsh MF, Brown RA. Reduced con-tractile response to insulin and IGF-I in ventricular myocytesfrom genetically obese Zucker rats. Am J Physiol Heart CircPhysiol. 2000;279:H1708–14.
4. Ren J. Leptin and hyperleptinemia: from friend to foe forcardiovascular function. J Endocrinol. 2004;181:1–10.
5. Unger RH, Orci L. Lipotoxic diseases of nonadipose tissuesin obesity. Int J Obes Relat Metab Disord. 2000;24 Suppl4:S28–32.
6. Wold LE, Relling DP, Duan J, Norby FL, Ren J. Abrogatedleptin-induced cardiac contractile response in ventricular myo-cytes under spontaneous hypertension: role of Jak/STAT path-way. Hypertension. 2002;39:69–74.
7. Zhou YT, Grayburn P, Karim A, et al. Lipotoxic heartdisease in obese rats: implications for human obesity. ProcNatl Acad Sci U S A. 2000;97:1784–9.
8. Anderson RA. Chromium in the prevention and control ofdiabetes. Diabetes Metab. 2000;26:22–7.
9. Jeejeebhoy KN, Chu RC, Marliss EB, Greenberg GR,Bruce-Robertson A. Chromium deficiency, glucose intoler-ance, and neuropathy reversed by chromium supplementation,in a patient receiving long-term total parenteral nutrition. Am JClin Nutr. 1977;30:531–8.
10. Anderson RA. Chromium and diabetes. Nutrition. 1999;15:720–2.
11. Cefalu WT, Hu FB. Role of chromium in human health andin diabetes. Diabetes Care. 2004;27:2741–51.
A Novel Chromium Complex and Cardiac Function, Dong et al.
2710 OBESITY Vol. 15 No. 11 November 2007

12. Morris BW, Kouta S, Robinson R, MacNeil S, Heller S.Chromium supplementation improves insulin resistance inpatients with type 2 diabetes mellitus. Diabet Med. 2000;17:684–5.
13. Yang X, Palanichamy K, Ontko AC, et al. A newly syn-thetic chromium complex-chromium(phenylalanine)3 im-proves insulin responsiveness and reduces whole body glucosetolerance. FEBS Lett. 2005;579:1458–64.
14. Anderson RA, Cheng N, Bryden NA, et al. Elevated intakesof supplemental chromium improve glucose and insulin vari-ables in individuals with type 2 diabetes. Diabetes. 1997;46:1786–91.
15. Lee NA, Reasner CA. Beneficial effect of chromium supple-mentation on serum triglyceride levels in NIDDM. DiabetesCare. 1994;17:1449–52.
16. Bagchi D, Stohs SJ, Downs BW, Bagchi M, Preuss HG.Cytotoxicity and oxidative mechanisms of different forms ofchromium. Toxicology. 2002;180:5–22.
17. Yang X, Li SY, Dong F, Ren J, Sreejayan N. Insulin-sensitizing and cholesterol-lowering effects of chromium (D-phenylalanine)3. J Inorg Biochem. 2006;100:1187–93.
18. Dong F, Fang CX, Yang X, Zhang X, Lopez FL, Ren J.Cardiac overexpression of catalase rescues cardiac contractiledysfunction induced by insulin resistance: role of oxidativestress, protein carbonyl formation and insulin sensitivity. Dia-betologia. 2006;49:1421–33.
19. Li SY, Yang X, Ceylan-Isik AF, Du M, Sreejayan N, RenJ. Cardiac contractile dysfunction in Lep/Lep obesity is ac-companied by NADPH oxidase activation, oxidative modifi-cation of sarco(endo)plasmic reticulum Ca(2�)-ATPase andmyosin heavy chain isozyme switch. Diabetologia. 2006;49:1434–46.
20. Fang CX, Dong F, Ren BH, Epstein PN, Ren J. Metallo-thionein alleviates cardiac contractile dysfunction induced byinsulin resistance: role of Akt phosphorylation, PTB1B,PPARgamma and c-Jun. Diabetologia. 2005;48:2412–21.
21. Adachi T, Weisbrod RM, Pimentel DR, et al.S-Glutathiolation by peroxynitrite activates SERCA duringarterial relaxation by nitric oxide. Nat Med. 2004;10:1200–7.
22. Pantke U, Volk T, Schmutzler M, Kox WJ, Sitte N, GruneT. Oxidized proteins as a marker of oxidative stress duringcoronary heart surgery. Free Radic Biol Med. 1999;27:1080–6.
23. Li SY, Du M, Dolence EK, et al. Aging induces cardiacdiastolic dysfunction, oxidative stress, accumulation of ad-vanced glycation endproducts and protein modification. AgingCell. 2005;4:57–64.
24. Ceriello A, Motz E. Is oxidative stress the pathogenic mech-anism underlying insulin resistance, diabetes, and cardiovas-cular disease? The common soil hypothesis revisited. Arterio-scler Thromb Vasc Biol. 2004;24:816–23.
25. Duan J, Zhang HY, Adkins SD, et al. Impaired cardiacfunction and IGF-I response in myocytes from calmodulin-diabetic mice: role of Akt and RhoA. Am J Physiol EndocrinolMetab. 2003;284:E366–76.
26. Relling DP, Esberg LB, Fang CX, et al. High-fat diet-induced juvenile obesity leads to cardiomyocyte dysfunctionand upregulation of Foxo3a transcription factor independentof lipotoxicity and apoptosis. J Hypertens. 2006;24:549–61.
27. Matthaei S, Stumvoll M, Kellerer M, Haring HU. Patho-physiology and pharmacological treatment of insulin resis-tance. Endocr Rev. 2000;21:585–618.
28. Di B, V, Santini F, Di Cori A, et al. Obesity cardiomyopathy:is it a reality? An ultrasonic tissue characterization study. J AmSoc Echocardiogr. 2006;19:1063–71.
29. Ren J, Ceylan-Isik AF. Diabetic cardiomyopathy: do womendiffer from men? Endocrine. 2004;25:73–83.
30. Hintz KK, Ren J. Prediabetic insulin resistance is not per-missive to the development of cardiac resistance to insulin-like growth factor I in ventricular myocytes. Diabetes Res ClinPract. 2002;55:89–98.
31. Goldhaber JI, Qayyum MS. Oxygen free radicals andexcitation-contraction coupling. Antioxid Redox Signal. 2000;2:55–64.
32. Bers DM. Cardiac excitation-contraction coupling. Nature.2002;415:198–205.
33. Tomas E, Lin YS, Dagher Z, et al. Hyperglycemia andinsulin resistance: possible mechanisms. Ann N Y Acad Sci.2002;967:43–51.
34. Matsui T, Nagoshi T, Rosenzweig A. Akt and PI 3-kinasesignaling in cardiomyocyte hypertrophy and survival. CellCycle. 2003;2:220–3.
35. Matsui T, Tao J, del Monte F, et al. Akt activation preservescardiac function and prevents injury after transient cardiacischemia in vivo. Circulation. 2001;104:330–5.
36. Freund H, Atamian S, Fischer JE. Chromium deficiencyduring total parenteral nutrition. JAMA. 1979;241:496–8.
37. Broadhurst CL, Domenico P. Clinical studies on chromiumpicolinate supplementation in diabetes mellitus: a review. Di-abetes Technol Ther. 2006;8:677–87.
38. Martin J, Wang ZQ, Zhang XH, et al. Chromium picolinatesupplementation attenuates body weight gain and increasesinsulin sensitivity in subjects with type 2 diabetes. DiabetesCare. 2006;29:1826–32.
39. Vrtovec M, Vrtovec B, Briski A, Kocijancic A, AndersonRA, Radovancevic B. Chromium supplementation shortensQTc interval duration in patients with type 2 diabetes mellitus.Am Heart J. 2005;149:632–6.
40. Vincent JB. Elucidating a biological role for chromium at amolecular level. Acc Chem Res. 2000;33:503–10.
41. Nukuna BN, Goshe MB, Anderson VE. Sites of hydroxylradical reaction with amino acids identified by (2)H NMRdetection of induced (1)H/(2)H exchange. J Am Chem Soc.2001;123:1208–14.
42. Phillips LS, Dunning BE. Nateglinide (Starlix): update on anew antidiabetic agent. Int J Clin Pract. 2003;57:535–41.
43. Press RI, Geller J, Evans GW. The effect of chromiumpicolinate on serum cholesterol and apolipoprotein fractions inhuman subjects. West J Med. 1990;152:41–5.
A Novel Chromium Complex and Cardiac Function, Dong et al.
OBESITY Vol. 15 No. 11 November 2007 2711