chemical dynamics in slash-and-burn agriculture
TRANSCRIPT

ELSEVIER Agriculture, Ecosystems and Environment 58 (1996) 49-60
Agriculture Ecosystems & Environment
Chemical dynamics in slash-and-burn agriculture
Anthony S.R. Juo *, Andrew Manu Department of Soil and Crop Sciences, Texas A and M University, College Station, TX 77843-2474, USA
Abstract
The stability of the slash-and-burn system is more a function of the total nutrient stock of the entire ecosystem than the net gain of the soil after slash-and-burn. During the first cycle of land exploitation, slash-and-burn releases a large portion of nutrients stored in the above-ground biomass into the soil. Therefore, the gain of nutrient stock in the soil is at the expense of the plant biomass. Published data on slash-and-burn systems are mostly concerned with the changes in soil nutrient status during cropping. Studies dealing with the dynamics of total nutrient stock in the primary forest ecosystem and the subsequent cropping and fallow cycles are scarce.
Conservation of the total mineral nutrient stock of the slash-and-burn system, starting from the clearing of a primary forest followed by burning, cropping and secondary forest fallow, is possible only in an ideal case of shifting cultivation - the equilibrium model. A significant portion of mineral nutrients released from burning may be lost either through erosion and runoff or through leaching (i.e. K, Mg, Ca, nitrate, and sulfate). A portion of the plant nutrients will be removed in harvested crops. Thus, the total mineral nutrient stock in the whole ecosystem gradually declines during the subsequent cycles of fallow and cropping - the depletion model.
For various social and economic reasons, slash-and-burn agriculture is being rapidly replaced by new or modified crop production systems throughout the tropics. In strongly weathered soils, external inputs of mineral nutrients are required, particularly, those aiming at sustaining high levels of crop or pasture productivity. Although nitrogen can be replenished through biological N-fixation, other nutrients, especially P and K, must be supplied by external sources. Therefore, a sustainable alternative to the slash-and-bum system should be judged not only by the stability of crop yield but also by the efficiency of the agroecosystem to recycle mineral nutrients as a function of time.
Keywords: Nutrient cycling; Forest ecosystems; Managed ecosystems
1. Introduction
Although slash-and-bum agriculture remains a dominant system of food production in the humid and subhumid tropics, it has also become a major
* Corresponding author. Tel: + 1 409 845 8841; Fax: + 1 409 845 0456.
cause of deforestation and land degradation (Nye and Greenland, 1960; Sanchez and Buol, 1975; Barrow, 1991). Ideally, slash-and-burn agriculture is ecologi- cally stable under very low density of human popula- tion. However, such practice rarely exists today ex- cept in some remote regions of the Amazon and Congo basins. In a greater part of the humid and subhumid tropics, the fallow period has been reduced and the areas cleared for cultivation have become
0167-8809/96/$15.00 © 1996 Elsevier Science B.V. All rights reserved SSDI 01 67-8809(95)00656-7

50 A.S.R. Juo, A. Manu / Agriculture, Ecosystem and Environment 58 (1996) 49-60
larger. These modifications have led to an eventual breakdown of the slash-and-burn system in the long run because loss of mineral nutrients during the cultivation phase (i.e. through runoff, erosion, leach- ing, and crop removal) can no longer be restored by short periods of bush fallow.
The search for workable alternatives to slash- and-burn agriculture has been a central focus of tropical agricultural research in the past decades. Some researchers have attempted to replace it with modern systems and practices including continuous cropping with chemical fertilization. Others have promoted various systems of tree-crop mixtures such as alley cropping and multistory home gardens (Sanchez et al., 1982; Nicholaides et al., 1985; Juo, 1989; Juo and Kang, 1989). While the adoption of new or modified soil management systems by small-scale farmers depends largely on the social, economic, and cultural environments, the central problem may still rest on the ability of the modified systems to recycle and conserve mineral nutrients.
In this paper, we attempt to conceptualize, from an ecosystem perspective, mechanisms and processes that influence the dynamics of mineral nutrients (i.e. Ca, Mg, K, P) in the slash-and-burn and modified cropping systems. The dynamics of organic matter and nitrogen cycling are discussed by Palm et al. (1996), in this volume.
2. Nutrients in primary forest ecosystems
Nutrient cycling and conservation mechanisms in tropical forest ecosystems have been reviewed by Jordan (1985) and Proctor (1989). Tropical rain- forests thrive on both fertile and infertile soils, but the total amount of nutrient accumulation in the plant biomass in a given forest ecosystem depends on the
inherent fertility of the soil. Vitousek and Sanford (1986) observed that the most productive forests thrive on relatively fertile soils. They recycle and store large quantities of mineral nutrients. On the other hand, forests growing on infertile soils are less productive and recycle smaller amounts of nutrients. This relationship is shown by Furch and Klinge (1989), who measured the annual litter fall and nutrient concentrations of forests growing on soils with different levels of inherent fertility in the Ama- zonian flood plains near Manaus, Brazil (Table 1).
In primary forest ecosystems, two nutrient cycling mechanisms have been identified. The first involves internal cycling of nutrients released from the de- composing litter and captured by the root mat on the forest floor. The other involves a combination of recovery of mineral nutrients from the subsoils and saprolites and internal cycling (Jordan et al., 1972; Herrera et al., 1978; Baillie, 1989).
Primary forests established on strongly acidic and leached soils, such as the rainforests in the Amazon and Congo basins, depend on internal cycling to meet their mineral requirements. Losses are offset solely by atmospheric inputs, and the nutrients ab- sorbed by the forest vegetation are derived from litter decomposition and infiltration of surface water. The thick layer of root mat on the forest floor retains large amounts of water and available nutrients and facilitates nutrient uptake by the plant. In such forest ecosystems, over 80% of the N and P is held by the total plant biomass, including leaves, branches, trunk, and roots (Scott, 1978 as cited by Jordan, 1985; Jordan, 1985). On the other hand, the bulk of the Ca, Mg, and K is held in the soil (Sanchez, 1979). Primary forests established on high base status soils benefit from the supply of additional mineral nutri- ents in deeper layers of the soil and saprolites. These are Alfisols and Andisols in the volcanic and lime-
Table l Leaf litter fall mass and concentration of nutrient elements from Varzea and Igapo primary forests near Manaus, Brazil (Furch and Klinge, 1989)
Forest Soil Litter f a l l Concentration of nutrient elements (g m - 2 year- ~ ) (tha-I year-I) N P K Ca Mg
Varzea Loamy alluvial deposits 10.6 15.5 1.0 7.2 31.4 6.1 lgapo Leached sand on kaolinitic clay 5.0 8.0 0.1 1.5 2.8 0.7

A.S.R. Juo, A. Manu /Agriculture, Ecosystem and Environment 58 (1996) 49-60 51
stone regions of Central America and the Alfisols derived from the basement complex in West Africa.
An important implication of such unbalanced nu- trient distribution between biomass and the soil is that a substantial amount of the mineral nutrients released during the slash-and-bum operation may be lost through leaching and runoff if the soil system is unable to retain them. Nevertheless, in the ideal slash-and-bum system, nutrients added to the small patch of manually cleared field are captured either by the surrounding forest vegetation or by the living forest root mat remaining in cropped field. As pointed out by Brinkman (1983), the Amazonian rainforest ecosystems on the tertiary formations display a dy- namic steady state system with tightly closed biogeo- chemical cycles for Ca, Mg, P, and N. If these forests are transformed into human-managed systems on a large scale, the nutrient pool, primarily the organic components, becomes drastically reduced.
From a pedological viewpoint, two properties of strongly weathered soils influence the processes of the internal cycling of mineral nutrients in the natu- ral forest ecosystem: (1) the two-phased water flow systems and (2) the positive charge characteristics of the subsoil. First, availability of soil nutrients to the plant is dependent on the flow of water through the soil system. Experimental evidence shows that two water transport systems may occur in strongly weathered and highly permeable soils under forests. One is determined by macropores through which water drains rapidly during rainstorms. The other is determined by micropores through which water flow is slower (Pleysier and Juo, 1981; Grimme and Juo, 1985; Nortcliff and Thome, 1989). Soluble nutrients released from decomposing litter are mainly retained in micropores that permit sufficient residence time for their uptake by plants. This direct nutrient supply system can be determined only by examining the concentrations of soluble nutrients in micropores, mesopores, and macropores. Thus, analysis of the nutrient concentrations in stream and subsurface flow gives an unsatisfactory assessment of the nutrient dynamics of the rainforest ecosystem (Jordan et al., 1972; Nortcliff and Thome, 1989). Secondly, the presence of positive charges in the subsurface layers of many oxidic Oxisols and Ultisols may result in a slower rate of downward movement of anions such as nitrate and sulfates (Wong et al., 1987). The
retarded leaching process also increases the resi- dence time of nutrients, which in turn favors nutrient uptake by plants.
3. Nutrient accumulation during the fallow period
Accumulation of above-ground biomass and min- eral nutrients during the fallow phase after shifting cultivation has been reviewed by Szott and Palm (1986). They concluded that soil fertility, forest type, and frequency and intensity of disturbance may all affect the rate of forest recovery. From an ecosystem perspective, the main function of the fallow phase is essentially the transfer of mineral nutrients from the soil back into forest biomass. Because of the in- evitable loss of mineral nutrients during the cropping phase, recovery of the total nutrient stock in the entire ecosystem during the subsequent fallow cycles is expected to be progressively smaller than that of the original primary forest ecosystem. This is also a function of mining of mineral nutrients from the subsoil and saprolites
Published data show that total biomass accumula- tion during the first 10 years of fallow varies consid- erably, ranging from 48 to 160 t ha -~. Data from the Congo basin (Bartholomew et al., 1953) and the Rocky Mountain region of the United States (Scott, 1977) show that biomass accumulation reached a steady state after 15 and 7 years of fallow, respec- tively. However, the total amount of biomass reached only 35 and 45%, respectively, of the biomass of the primary forest. These data suggest that the total amount of biomass of the secondary forest may take hundreds of years to reach the level comparable with that of the primary forest once the level of the total nutrient stock has been significantly reduced and the nutrient cycling and conservation mechanisms are disrupted by repeated cycles of slash-and-burn culti- vation.
Mature secondary forests (15 years or older) in the humid and subhumid tropics may produce 300 to 450 t ha-~ of above-ground biomass. They store approximately 1 to 2 t ha- ~ of N, 0.7 to 2.5 t ha- of K, 2 to 3 t ha- i of Ca, 300 to 800 kg ha- l of Mg, and 100 to 250 kg ha-~ of P (Bartholomew et al., 1953; Ogawa et al., 1965; Klinge et al., 1975; Ad- edeji, 1984; Kyuma et al., 1985; Saldarriaga et al.,
52 A.S.R. Juo, A. Manu / Agriculture, Ecosystem and Environment 58 (1996) 49-60
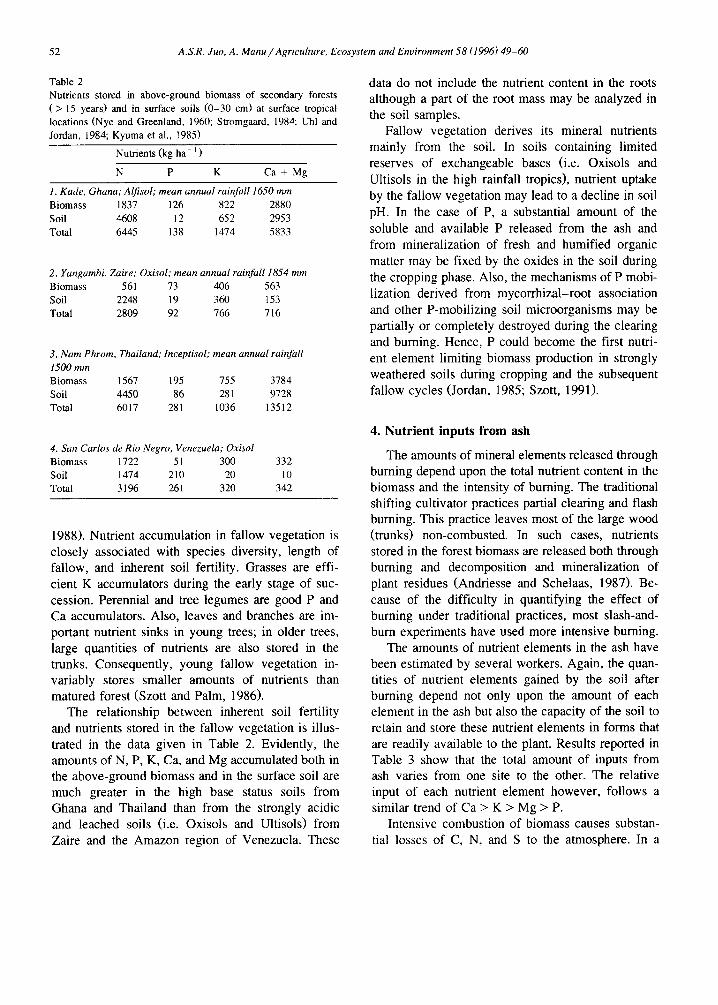
Table 2 Nutrients stored in above-ground biomass of secondary forests ( > 15 years) and in surface soils (0-30 cm) at surface tropical locations (Nye and Greenland, 1960; Stromgaard, 1984; Uhl and Jordan, 1984; Kyuma et al., 1985)
Nutrients (kg ha - i )
N P K Ca + Mg
1. Kade, Ghana;Alfisol; mean annualrainJal11650 mm Biomass 1837 126 822 2880 Soil 4608 12 652 2953 To~l 6445 138 1474 5833
2. Yangambi, Zaire; Oxisol; mean annualrainfall1854 mm Biomass 561 73 406 563 Soil 2248 19 360 153 Total 2809 92 766 716
3. Nam Phrom, Thailand; lnceptisol; mean annual rainJ~tll 1500 mm Biomass 1567 195 755 3784 Soil 4450 86 281 9728 Total 6017 281 1036 13512
4. San Carlos de Rio Negro, Venezuela; Oxisol Biomass 1722 51 300 332 Soil 1474 210 20 10 Total 3196 261 320 342
1988). Nutrient accumulation in fallow vegetation is closely associated with species diversity, length of fallow, and inherent soil fertility. Grasses are effi- cient K accumulators during the early stage of suc- cession. Perennial and tree legumes are good P and Ca accumulators. Also, leaves and branches are im- portant nutrient sinks in young trees; in older trees, large quantities of nutrients are also stored in the trunks. Consequently, young fallow vegetation in- variably stores smaller amounts of nutrients than matured forest (Szott and Palm, 1986).
The relationship between inherent soil fertility and nutrients stored in the fallow vegetation is illus- trated in the data given in Table 2. Evidently, the amounts of N, P, K, Ca, and Mg accumulated both in the above-ground biomass and in the surface soil are much greater in the high base status soils from Ghana and Thailand than from the strongly acidic and leached soils (i.e. Oxisols and Ultisols) from Zaire and the Amazon region of Venezuela. These
data do not include the nutrient content in the roots although a part of the root mass may be analyzed in the soil samples.
Fallow vegetation derives its mineral nutrients mainly from the soil. In soils containing limited reserves of exchangeable bases (i.e. Oxisols and Ultisols in the high rainfall tropics), nutrient uptake by the fallow vegetation may lead to a decline in soil pH. In the case of P, a substantial amount of the soluble and available P released from the ash and from mineralization of fresh and humified organic matter may be fixed by the oxides in the soil during the cropping phase. Also, the mechanisms of P mobi- lization derived from mycorrhizal-root association and other P-mobilizing soil microorganisms may be partially or completely destroyed during the clearing and burning. Hence, P could become the first nutri- ent element limiting biomass production in strongly weathered soils during cropping and the subsequent fallow cycles (Jordan, 1985; Szott, 1991).
4. Nutrient inputs from ash
The amounts of mineral elements released through burning depend upon the total nutrient content in the biomass and the intensity of burning. The traditional shifting cultivator practices partial clearing and flash burning. This practice leaves most of the large wood (trunks) non-combusted. In such cases, nutrients stored in the forest biomass are released both through burning and decomposition and mineralization of plant residues (Andriesse and Schelaas, 1987). Be- cause of the difficulty in quantifying the effect of burning under traditional practices, most slash-and- burn experiments have used more intensive burning.
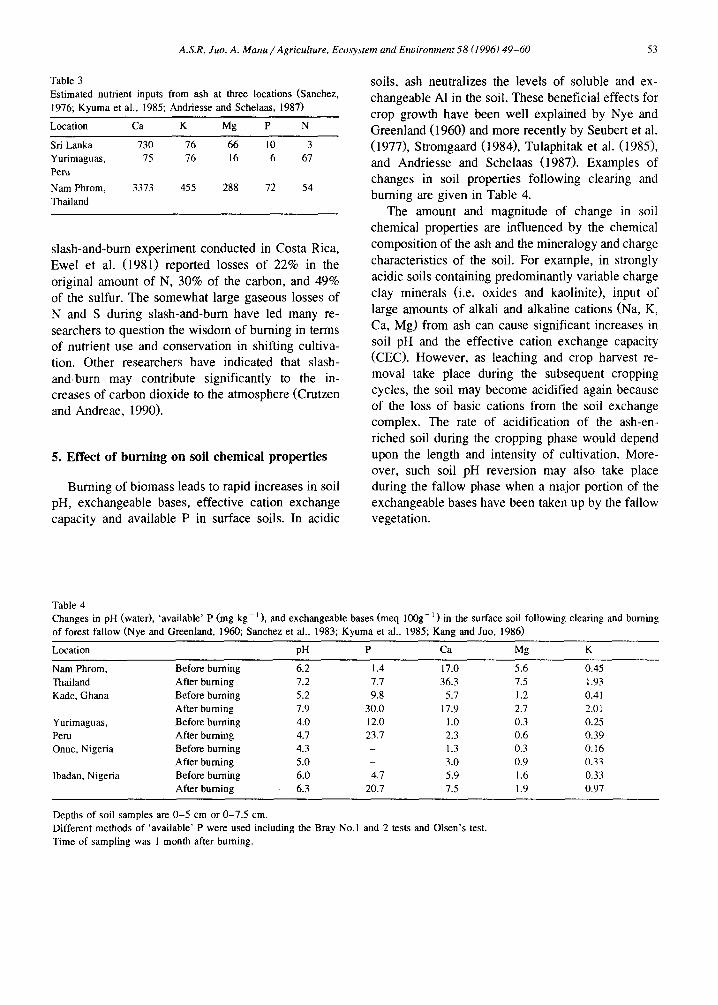
The amounts of nutrient elements in the ash have been estimated by several workers. Again, the quan- tities of nutrient elements gained by the soil after burning depend not only upon the amount of each element in the ash but also the capacity of the soil to retain and store these nutrient elements in forms that are readily available to the plant. Results reported in Table 3 show that the total amount of inputs from ash varies from one site to the other. The relative input of each nutrient element however, follows a similar trend of Ca > K > Mg > P.
Intensive combustion of biomass causes substan- tial losses of C, N, and S to the atmosphere. In a
A.S.R. Juo, A. Manu /Agriculture, Ecosystem and Environment 58 (1996) 49-60 53
Table 3 Estimated nutrient inputs from ash at three locations (Sanchez, 1976; Kyuma et al., 1985; Andriesse and Schelaas, 1987)
Location Ca K Mg P N
Sri Lanka 730 76 66 10 3 Yurimaguas, 75 76 16 6 67 Peru
Nam Phrom, 3373 455 288 72 54 Thailand
slash-and-burn experiment conducted in Costa Rica, Ewel et al. (1981) reported losses of 22% in the original amount of N, 30% of the carbon, and 49% of the sulfur. The somewhat large gaseous losses of N and S during slash-and-burn have led many re- searchers to question the wisdom of burning in terms of nutrient use and conservation in shifting cultiva- tion. Other researchers have indicated that slash- and-burn may contribute significantly to the in- creases of carbon dioxide to the atmosphere (Crutzen and Andreae, 1990).
5. Effect of burning on soil chemical properties
Burning of biomass leads to rapid increases in soil pH, exchangeable bases, effective cation exchange capacity and available P in surface soils. In acidic
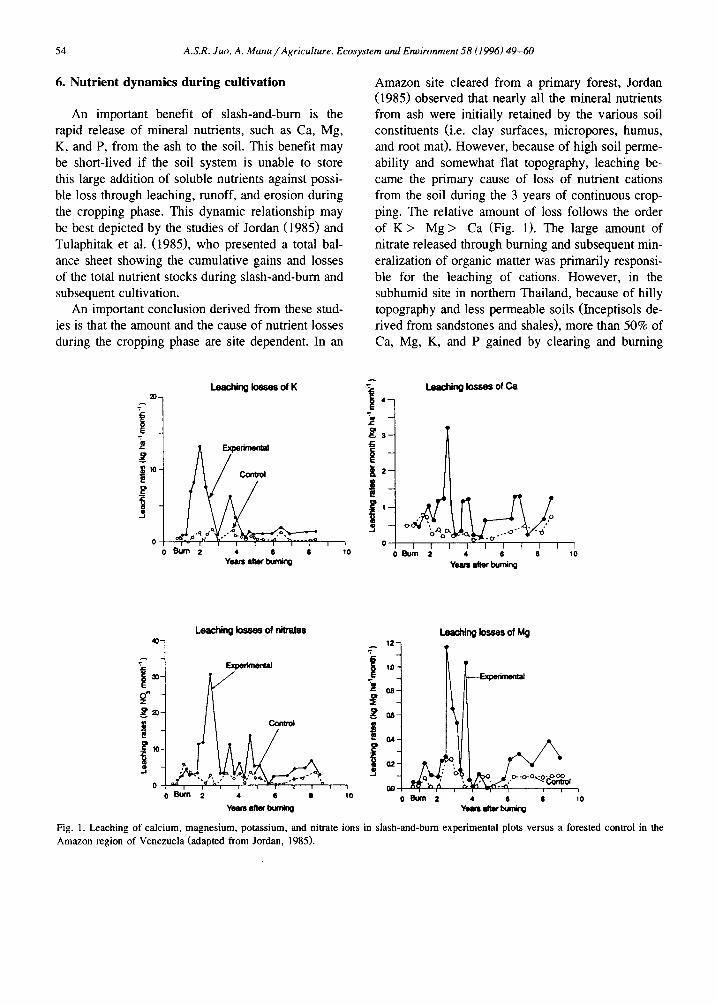
soils, ash neutralizes the levels of soluble and ex- changeable A1 in the soil. These beneficial effects for crop growth have been well explained by Nye and Greenland (1960) and more recently by Seubert et al. (1977), Stromgaard (1984), Tulaphitak et al. (1985), and Andriesse and Schelaas (1987). Examples of changes in soil properties following clearing and buming are given in Table 4.
The amount and magnitude of change in soil chemical properties are influenced by the chemical composition of the ash and the mineralogy and charge characteristics of the soil. For example, in strongly acidic soils containing predominantly variable charge clay minerals (i.e. oxides and kaolinite), input of large amounts of alkali and alkaline cations (Na, K, Ca, Mg) from ash can cause significant increases in soil pH and the effective cation exchange capacity (CEC). However, as leaching and crop harvest re- moval take place during the subsequent cropping cycles, the soil may become acidified again because of the loss of basic cations from the soil exchange complex. The rate of acidification of the ash-en- riched soil during the cropping phase would depend upon the length and intensity of cultivation. More- over, such soil pH reversion may also take place during the fallow phase when a major portion of the exchangeable bases have been taken up by the fallow vegetation.
Table 4 Changes in pH (water), 'available' P (rag kg - i ), and exchangeable bases (meq 100g- 1) in the surface soil following clearing and burning of forest fallow (Nye and Greenland, 1960; Sanchez et al., 1983; Kyuma et al., 1985; Kang and Juo, 1986)
Location pH P Ca Mg K
Nam Phrom, Before burning 6.2 1.4 17.0 5.6 0.45 Thailand After burning 7.2 7.7 36.3 7.5 1.93 Kade, Ghana Before burning 5.2 9.8 5.7 1.2 0.41
After burning 7.9 30.0 17.9 2.7 2.01 Yurimaguas, Before burning 4.0 12.0 1.0 0.3 0.25 Peru After burning 4.7 23.7 2.3 0.6 0.39 Onne, Nigeria Before burning 4.3 - 1.3 0.3 0.16
After burning 5.0 - 3.0 0.9 0.33 lbadan, Nigeria Before burning 6.0 4.7 5.9 1.6 0.33
After burning 6.3 20.7 7.5 1.9 0.97
Depths of soil samples are 0 -5 cm or 0-7.5 cm. Different methods of 'available' P were used including the Time of sampling was 1 month after burning.
Bray No.1 and 2 tests and Olsen's test.
54 A.S.R. Jut,, A. Manu / Agriculture, Ecosystem and Environment 58 (1996)49-60
6. Nutrient dynamics during cult ivation
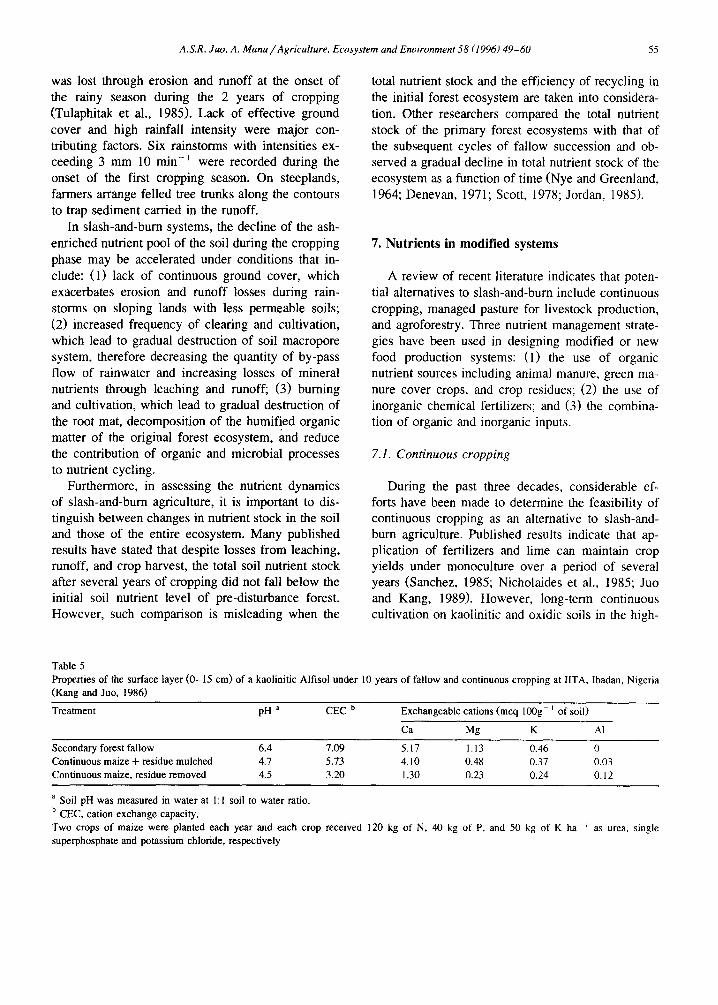
An important benefit of slash-and-burn is the rapid release of mineral nutrients, such as Ca, Mg, K, and P, from the ash to the soil. This benefit may be short-lived if the soil system is unable to store this large addition of soluble nutrients against possi- ble loss through leaching, runoff, and erosion during the cropping phase. This dynamic relationship may be best depicted by the studies of Jordan (1985) and Tulaphitak et al. (1985), who presented a total bal- ance =sheet showing the cumulative gains and losses of the total nutrient stocks during slash-and-burn and subsequent cultivation.
An important conclusion derived from these stud- ies is that the amount and the cause of nutrient losses during the cropping phase are site dependent. In an
Amazon site cleared from a primary forest, Jordan (1985) observed that nearly all the mineral nutrients from ash were initially retained by the various soil constituents (i.e. clay surfaces, micropores, humus, and root mat). However, because of high soil perme- ability and somewhat fiat topography, leaching be- came the primary cause of loss of nutrient cations from the soil during the 3 years of continuous crop- ping. The relative amount of loss follows the order of K > M g > Ca (Fig. 1). The large amount of nitrate released through burning and subsequent min- eralization of organic matter was primarily responsi- ble for the leaching of cations. However, in the subhumid site in northern Thailand, because of hilly topography and less permeable soils (Inceptisols de- rived from sandstones and shales), more than 50% of Ca, Mg, K, and P gained by clearing and burning
2D-
T"
E
4=
Leaching losses of K
e
i i B u m 2 4 6 8 1 0
leaching losses of Ca 4 - -
3 - -
2 - -
0 I I ~ m 2
I I I 1 I 4 6 8
a~ - bun~ng
I I 10
4 0 - 1 2 -
3 0 - .
g o - ~ ll -
-
~ - ~ i , ~ , - _ 0 ~ ~ ~
B l x n 2 4 6 i i 10
Yearn alt~ burnl,',g
L_ _,_,_,_,_,_,_,_,_~__ ing bs.~s of Mg
f
i ~ i -i i i i i
$ $ i
10
Fig. 1. Leaching of calcium, magnesium, potassium, and nitrate ions in slash-and-bum experimental plots versus a forested control in the Amazon region of Venezuela (adapted from Jordan, 1985).
A.S.R. Jut, A. Manu / Agriculture, Ecosystem and Environment 58 (1996) 49-60 55
was lost through erosion and runoff at the onset of the rainy season during the 2 years of cropping (Tulaphitak et al., 1985). Lack of effective ground cover and high rainfall intensity were major con- tributing factors. Six rainstorms with intensities ex- ceeding 3 mm 10 min-~ were recorded during the onset of the first cropping season. On steeplands, farmers arrange felled tree trunks along the contours to trap sediment carried in the runoff.
In slash-and-burn systems, the decline of the ash- enriched nutrient pool of the soil during the cropping phase may be accelerated under conditions that in- clude: (1) lack of continuous ground cover, which exacerbates erosion and runoff losses during rain- storms on sloping lands with less permeable soils; (2) increased frequency of clearing and cultivation, which lead to gradual destruction of soil macropore system, therefore decreasing the quantity of by-pass flow of rainwater and increasing losses of mineral nutrients through leaching and runoff; (3) burning and cultivation, which lead to gradual destruction of the root mat, decomposition of the humified organic matter of the original forest ecosystem, and reduce the contribution of organic and microbial processes to nutrient cycling,
Furthermore, in assessing the nutrient dynamics of slash-and-burn agriculture, it is important to dis- tinguish between changes in nutrient stock in the soil and those of the entire ecosystem. Many published results have stated that despite losses from leaching, runoff, and crop harvest, the total soil nutrient stock after several years of cropping did not fall below the initial soil nutrient level of pre-disturbance forest. However, such comparison is misleading when the
total nutrient stock and the efficiency of recycling in the initial forest ecosystem are taken into considera- tion. Other researchers compared the total nutrient stock of the primary forest ecosystems with that of the subsequent cycles of fallow succession and ob- served a gradual decline in total nutrient stock of the ecosystem as a function of time (Nye and Greenland, 1964; Denevan, 1971; Scott, 1978; Jordan, 1985).
7. Nutrients in modified systems
A review of recent literature indicates that poten- tial alternatives to slash-and-burn include continuous cropping, managed pasture for livestock production, and agroforestry. Three nutrient management strate- gies have been used in designing modified or new food production systems: (1) the use of organic nutrient sources including animal manure, green ma- nure cover crops, and crop residues; (2) the use of inorganic chemical fertilizers; and (3) the combina- tion of organic and inorganic inputs.
7.1. Continuous cropping
During the past three decades, considerable ef- forts have been made to determine the feasibility of continuous cropping as an alternative to slash-and- burn agriculture. Published results indicate that ap- plication of fertilizers and lime can maintain crop yields under monoculture over a period of several years (Sanchez, 1985; Nicholaides et al., 1985; Jut and Kang, 1989). However, long-term continuous cultivation on kaolinitic and oxidic soils in the high-
Table 5 Properties of the surface layer (0-15 cm) of a kaolinitic Alfisol under l0 years of fallow and continuous cropping at IITA, lbadan, Nigeria (Kang and Jut, 1986)
Treatment pH a CEC b Exchangeable cations (meq 100g-i of soil)
Ca Mg K A1
Secondary forest fallow 6.4 7.09 5.17 1.13 0.46 0 Continuous maize + residue mulched 4.7 5.73 4.10 0.48 0.37 0.03 Continuous maize, residue removed 4.5 3.20 1.30 0.23 0.24 0.12
a Soil pH was measured in water at 1:1 soil to water ratio. b CEC, cation exchange capacity. Two crops of maize were planted each year and each crop received 120 kg of N, 40 kg of P, and 50 kg of K ha L superphosphate and potassium chloride, respectively
as urea, single
56 A.S.R. Juo, A. Manu // Agriculture, Ecosystem and Enoironment 58 (1996) 49-60
rainfall tropics often resulted in a rapid decline in soil pH and a corresponding increase in soluble and exchangeable AI in the soil (Friessen et al., 1981; Sanchez et al., 1983; Kang and Juo, 1986). This effect is further accentuated when heavy or moderate doses of acidifying fertilizers were used (Table 5). The use of lime to ameliorate soil acidity on kaolinitic Ultisols under continuous cultivation had only a short residual effect (Friessen et al., 1981; Sanchez et al., 1983).
It has been shown that high yields of annual crops over a period of 10 years or more could be obtained using modern soil management practices including conservation tillage, crop rotation and judicious use of fertilizers and lime (i.e. small dosages and fre- quent applications). Nevertheless, these results were
mostly obtained on small experimental plots. The transfer of these findings to larger fields, particularly when tractors are used for land preparation, has led to deterioration of physical soil properties of many kaolinitic and oxidic soils. Under large-scale crop- ping, soil compaction and surface sealing may be- come a more serious yield-limiting factor than nutri- ent and lime inputs (Lal et al., 1986). Leguminous cover crops such as Puereria phaseolus (kudzu) and Mucuna utilis have been used as cover crops in rubber and oil palm plantations during early stages of tree crop establishment. Such practices not only help to reduce soil erosion and loss of nutrients through runoff (Juo et al., 1994) but they also supply N to the ecosystem through biological N-fixation (Williams et al., 1980; Nair, 1990).
0 " r N S-
B u m /
i 2 3 4 s 8 ? ii • 10 !112 13
Years under pamm
J
, , P , , , , , , , , , , , , , i
I 2 3 4 S 6 7 8 9 10 11 s2 t3
IS"
.L.~ 6
£
Years tma~r pasture
2 "
g
E ~'
+2 I <" W
Bum
1 . . . . ; ~ i i ' s k ~ i 11~1i li is
YearsunderpasS.m 1 2 3 4 S 6 7
Yemnl undm" pasture
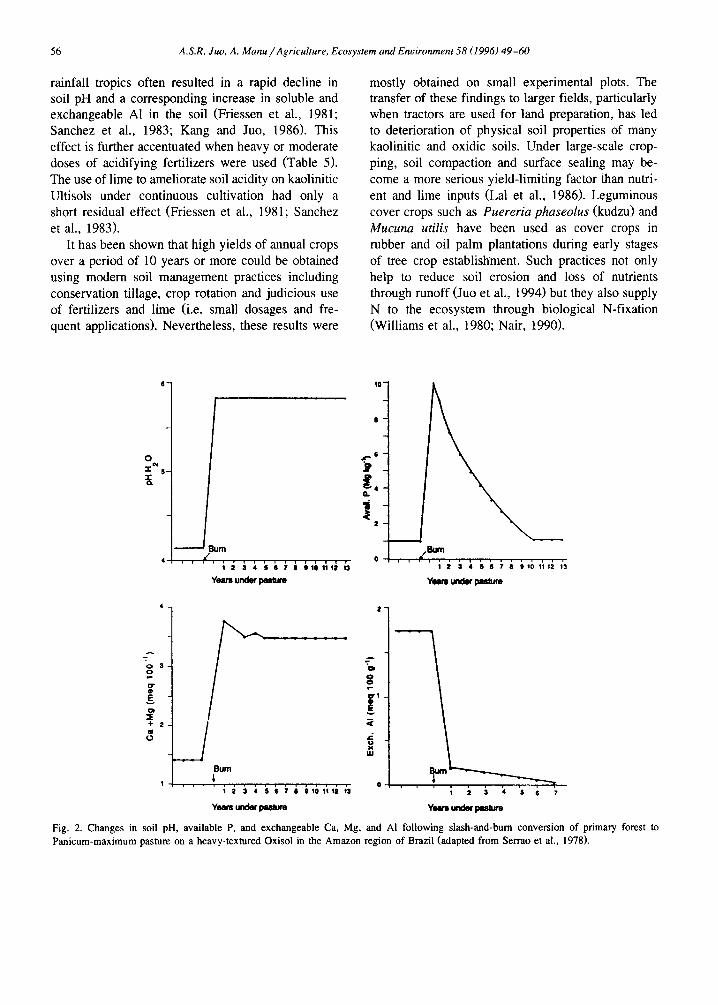
Fig. 2. Changes in soil pH, available P, and exchangeable Ca, Mg, and A1 following slash-and-burn conversion of primary forest to Panicum-maximum pasture on a heavy-textured Oxisol in the Amazon region of Brazil (adapted from Serrao et al., 1978).

A.S.R. Juo, A. Manu /Agriculture, Ecosystem and Environment 58 (1996) 49-60 57
7.2. Pasture and livestock production
Long-term studies conducted on the infertile Ox- isols and Ultisols in the Amazon region of Brazil showed that when grass pasture (mainly Panicum maximum) was established soon after burning of the forest biomass, the raised levels of soil pH, CEC, and exchangeable Ca, Mg, and K as a result of burning were maintained throughout the experimen- tal period of 13 years (Fig. 2). But P deficiency became a limiting factor, resulting in significant decline of pasture productivity after the 5th year. Pasture productivity was then improved through fer- tilization with P, establishing a legume-grass mix- ture, and maintaining an optimum level of grazing (Serrao et al., 1978).
From both ecological and economic viewpoints, conversion of tropical rainforests to livestock pro- duction is among the least desirable alternatives to shifting cultivation. In humid tropical environments, livestock is exposed to severe pests and disease problems, and the infertile soils impose nutritional limits to pasture production (Goodland, 1980; Manet, 1982). Overgrazing can result in severe soil erosion and compaction and prevent pasture regrowth (De- nevan, 1981 as cited by Jordan, 1985). Moreover, to meet the food and nutrition needs of the increasing population in the humid tropics, food legumes are far more efficient than cattle as protein converters per unit area of land.
7.3. Crop-tree mixed systems
Interplanting fast-growing tree legumes such as Leucaena leucocephala with annual crops (i.e. alley cropping) has also been tried on high base status Alfisols and Entisols for more than 10 years at the International Institute of Tropical Agriculture (IITA) near Ibadan, Nigeria (Kang et al., 1981; Kang et al., 1984). The system mimics the bush fallow system while allowing continuous cropping. The tree legume fixes nitrogen from the atmosphere and recycles mineral nutrients from the subsoil. The hedgerows are pruned periodically to provide green manure and mulch for the accompanying food crop. For example, the total annual production of green manure from hedgerows growing on an Alfisol near Ibadan, Nige- ria, can be as much as 7 t ha-~ (dry weight). The
green manure contains 250 kg N, 20 kg P, 185 kg K, 100 kg Ca, and 15 kg Mg (Juo and Kang, 1989). Another advantage of alley cropping is that it pro- vides continuous ground cover, hence minimizing nutrient losses through leaching and runoff.
Because of large annual additions of Leucaena green manure, the alley system is prone to losses of nitrate and nutrient cations through leaching. This often results in decreases in pH of the surface soil. Long-term field trials in Nigeria have shown signifi- cant decreases in pH and exchangeable K and Mg in the soil under alley cropping (Kang et al., 1981; Yamoah et al., 1986). However, what proportions of the leached cations and nitrate are recycled by the deep-rooting tree legume and what proportion are lost to seepage and ground water remain to be determined.
Performance of alley cropping on low base infer- tile soils was evaluated in the lowland rainforest zone of the Amazon basin of Peru (Szott et al., 1991). Prunnings of Inga edulis, Erythrina sp., Cas- sia reticulata and Gliricidia sepium were applied. The average dry matter yield was 2.5 t ha - 1. The average quantity of nutrients per hectare contained in the hedgerows were 66 kg N, 6 kg P, 33 kg K, 20 kg Ca, and 6 kg Mg. Over time, additions of prunnings resulted in increased soil acidity as decreased pH and increased exchangeable A1. Levels of exchangeable nutrient cations declined to levels similar to systems which received neither organic nor inorganic inputs. Thus, alley cropping system on strongly acidic and infertile soils is only sustainable if food crops are interplanted with deep-rooting, acid tolerant woody species such as Dactyldenia barteri (syn. Acioa barteri), Flemingia macrophylla and Calliandra calothyrsus. Under these circumstances, external in- puts of nutrients and lime may be required as initial investment to sustain crop yield and to facilitate nutrient cycling (Everson, 1989; Juo and Kang, 1989; Wilson and Swennen, 1989; Kang et al., 1989; Ruhigwa et al., 1992; Juo et al., 1994).
8. Conclusions
In terms of nutrient dynamics, the stability of the slash-and-burn system is more a function of the total nutrient stock of the entire ecosystem than the net

58 A.S.R. Juo. A. Manu / Agriculture, Ecosystem and Environment 58 (1996) 49-60
gain of the soil after slash-and-bum. During the first cycle of land exploitation, slash-and-bum releases a large portion of nutrients stored in the above-ground biomass into the soil. Therefore, the gain of nutrient stock in the soil is at the expense of the plant biomass. Published data on slash-and-bum systems are mostly concerned with the changes in soil nutri- ent status during cropping. Studies dealing with the dynamics of total nutrient stock in the primary forest ecosystem and the subsequent cropping and fallow cycles are scarce. Based on the review of published work, we are tempted to make the following observa- tions: • Conservation of the total mineral nutrient stock of
the slash-and-burn system, starting from the clear- ing of a primary forest followed by burning, cropping and secondary forest fallow, is possible only in an ideal case of shifting cultivation - the equilibrium model. Such a system may be estab- lished under the following conditions: (1) the size of the cropped plot is sufficiently small that it is surrounded by vast areas of primary forest vegeta- tion; (2) the length of cultivation is kept below 2 years; and (3) the length of fallow is kept suffi- ciently long.
• In terms of nutrient cycling, most slash-and-bum systems are no longer in equilibrium as cropped fields become larger and length of fallow be- comes shorter. A significant portion of mineral nutrients released from burning may be lost either through erosion and runoff or through leaching (i.e. K, Mg, Ca, nitrate, and sulfate). A portion of the plant nutrients will be removed in harvested crops. Thus, the total mineral nutrient stock in the whole ecosystem gradually declines during the subsequent cycles of fallow and cropping - the depletion model (Fig. 3). The rate of decline of the total nutrient stock depends upon the size of the cropped field, the length and intensity of cultivation, and the length of fallow.
• Major causes for the depletion of the total nutri- ent stock in the slash-and-bum system are soil erosion, runoff, leaching, nutrient removal through harvested crops, destruction of forest root mat, and decline of the content of organic matter in the surface soil.
• For various social and economic reasons, slash- and-burn agriculture is being rapidly replaced by
J
Q T . Q6 + OB
Bum
Twao
Fig. 3. A conceptual model depicting the changes of nutrient stock in slash-and-burn agriculture as a function of time. QT = total nutrient stock of the ecosystem, QB = nutrient stock in plant biomass, and QS = nutrient stock in the soil.
new or modified crop production systems throughout the tropics. In strongly weathered soils, external inputs of mineral nutrients are required, particularly those aiming at sustaining high levels of crop or pasture productivity. Although nitrogen can be replenished through biological N-fixation, other nutrients, especially P and K, must be supplied by external sources. Therefore, a sustainable alternative to the slash- and-bum system should be judged not only by the stability of crop yield but also by the efficiency of the agroecosystem to recycle mineral nutrients as a function of time.
References
Adedeji, F.O., 1984. Nutrient cycles and successional changes following shifting cultivation practice in moist semideciduous forests in Nigeria. For. Ecol. Manage., 9: 87-99.
Andriesse, J.P. and Schelaas, R.M., 1987. A monitoring study on nutrient cycles in soils under shifting cultivation under various climatic conditions in tropical Asia. 3: The effect of land clearing through burning on fertility level. Agric. Ecosyst. Environ., 19:311-332.
Baillie, I.C., 1989. Soil characteristics and classification in rela- tion to the mineral nutrition of tropical wooded ecosystems. In: J. Proctor (Editor), Mineral Nutrients in Tropical Forest and Savanna Ecosystems. Blackwell Scientific Publications, Oxford, pp. 15-26.

A.S.R. Juo, A. Manu / Agriculture, Ecosystem and Environment 58 (1996) 49-60 59
Barrow, C.J., 1991. Land Degradation. Cambridge University Press, Cambridge/London/New York.
Bartholomew, W.V., Meyer, J. and Laudelout, H., 1953. Mineral nutrient immobilization under forest and grass fallow in the Yangambi region. Serie Scientifique No. 57, Institute National pour l'Etude Agronomique du Congo, Brussels.
Brinkman, W.L.F., 1983. Nutrient balances of a central Amazo- nian rainforest: comparison of natural and man-managed sys- tems. In: R. Keller (Editor), Hydrology of Humid Tropical Regions. International Association of Scientific Hydrologists, Publication No. 140, pp. 153-163.
Crutzen, P.J. and Andreae, M.O., 1990. Biomass burning in the tropics: impact on atmospheric chemistry and biogeochemical cycles. Science, 250: 1669-1678.
Denevan, W.M., 1971. Campa subsistence in the Gran Pajomal, eastern Peru. Geogr. Rev., 61: 496-518.
Denevan, W.M., 1981. Swiddens and cattle versus forests: the imminent demise of the Amazon forest reexamined. In: V.H. Sutlive. N. Altshuler and M.D. Zamora (Editors), Deforesta- tion in the Third World. Publication No. 13, Studies in Third World Societies, College of William and Mary, Williamsburg, Virginia, pp. 25-44.
Everson, C.L.I., 1989. Alley cropping and green manuring for upland crop production in West Sumatra. Ph.D dissertation, University of Hawaii, Honolulu, Hawaii.
Ewel, J., Berish, C., Brown, B., Price, N. and Raich, J. 1981. Slash-and-bum impacts on a Costa Rican wet forest site. Ecology, 62: 816-829.
Friessen, D.K., Juo, A.S.R. and Miller, M.H., 1981. Residual effects of lime and leaching of calcium in a kaolinitic Ultisol in the high rainfall tropics. Soil Sci. Soc. Am. J., 46:1184- 1189.
Furch, K. and Klinge, H., 1989. Chemical relationships between vegetation, soil and water in contrasting inundation areas of Amazonia. In: J. Proctor (Editor), Mineral Nutrients in Tropi- cal Forest and Savanna Ecosystems. Blackwell Science Publi- cations, Oxford, pp. 189-204.
Goodland, R.J., 1980. Environmental ranking of Amazonian de- velopment projects in Brazil. Environ. Conserv., 7: 9-26.
Grimme, H. and Juo, A.S.R., 1985. Inorganic nitrogen losses through leaching and denitrification in soils of the humid tropics. In: B.T. Kang and J. van der Heide (Editors), Nitrogen Management in Fanning Systems in the Humid and Subhumid Tropics. Institute of Soil Fertility, Haren, Netherlands, pp. 57-72.
Herrera, R.R., Jordan, F.C., Kline, H. and Medina, E., 1978. Amazon ecosystems: their structure and function with special emphasis on nutrients. Interciencia, 3: 223-232.
Jordan, C.F., 1985. Nutrient Cycling in Tropical Forest Ecosys- tems. Wiley, New York.
Jordan, C.F., Kline, J.R. and Sasscer, D.S., 1972. Relative stabil- ity of mineral cycles in forest ecosystems. Am. Nat., 106: 237-253.
Juo, A.S.R., 1989. New farming system development in the wetter tropics. Exp. Agric. (Camb.), 25: 145-163.
Juo, A.S.R. and Kang, B.T. 1989. Nutrient effects of modification of shifting cultivation in West Africa. In: J. Proctor (Editor),
Mineral Nutrients in Tropical Forest and Savanna Ecosystems. Blackwell Science Publications, Oxford, pp. 289-300.
Juo, A.S.R., Caldwell, J.O. and Kang, B.T., 1994. Place of alley cropping in sustainable agriculture in humid tropics. In: S.M. Virmani (Editor), Trans. 15th World Congress of Soil Science, Vol. 7a, International Soil Science Society, pp. 98-109.
Kang, B.T. and Juo, A.S.R., 1986. Effect of forest clearing on soil chemical properties and crop performance. In: R. Lal, P.A. Sanchez and R.W. Cummings Jr. (Editors), Land Clearing and Development in the Tropics. A.A. Balkema, Rotterdam/Bos- ton, pp. 383-394.
Kang, B.T., Wilson, G.F. and Sipkens, L., 1981. Alley cropping maize and Leucaena leucocephala in southern Nigeria. Plant Soil, 63: 165-t79.
Kang, B.T., Wilson, G.F. and Lawson, T.L., 1984. Alley Crop- ping: A Stable Alternative to Shifting Cultivation. IITA, Ibadan, Nigeria.
Kang, B.T., van der Kruijs, A.C. and Couper, D.C., 1989. Alley cropping for food crop production. In: B.T. Kang and L. Reynolds (Editors), Alley Farming in the Humid and Subhu- mid Tropics. IDRC, Ottawa, Canada, pp. 16-26.
Klinge, H., Rodriquez, W.A., Brunig, E. and Fittkau, E.J., 1975. Biomass and structure in a central Amazon rain forest. In: F.B. Golley and E. Medina (Editors), Tropical Ecological Systems. Springer-Verlag, New York, pp. 115-122.
Kyuma, K., Tulaphitak, T. and Pairintra, C., 1985. Changes in soil fertility and tilth under shifting cultivation: 1: General descrip- tion of soils and effect of burning on soil characteristics. Soil Sci. Plant Nutr. (Japan), 31: 227-238.
Lal, R., Sanchez, P.A. and Cummings, R.W., Jr. (Editors), 1986. Land Clearing and Development in the Tropics. A.A. Balkema, Rotterdam/Boston.
Manet, J.L., 1982. Nutritional limits to animal production from tropical pastures. In: J.B. Hacker (Editors), Problems of Ani- mal Production from Tropical Pastures. Commonwealth Agri- cultural Bureau, Slough, UK.
Nair, P.K.R., 1990. The prospects of agroforestry in the tropics. World Bank technical paper no. ISSN 0253-7494, no. 131.
Nicholaides, J.J., Bandy, D.E., Sanchez, P.A., Benites, J.R., Vil- lachica, J.H., Coutou, A.J. and Valverde, C.S.. 1985. Agricul- tural alternatives for the Amazon Basin. Bioscience, 35: 279- 285.
Nortcliff, S. and Thome, J.B., 1989. Variation in soil nutrients in relation to soil moisture status in a tropical forest ecosystems. In: J. Proctor (Editor), Mineral Nutrients in Tropical Forest and Savanna Ecosystems. Blackwell Science Publications, Ox- ford, pp. 43-54.
Nye, P.H. and Greenland, D.J., 1960. The soils under shifting cultivation. Commonwealth Bureau Soils, Tech. Comm. No. 51, Harpenden, UK.
Nye, P.H. and Greenland, D.J., 1964. Changes in soil properties after clearing a tropical forest. Plant Soil, 21:101-112.
Ogawa, H., Yoda, K., Ogino, K. and Kira, T., 1965. Comparative ecological studies on three main types of forest vegetation in Thailand. 2: Plant biomass. Nat. Life Southeast Asia, 4: 49- 80.
Palm, C.A., Swift, M.J. and Woomer, P.L., 1996. Soil biological

60 A.S.R. Juo, A. Manu / Agriculture, Ecosystem and Environment 58 (1996) 49-60
dynamics in slash-and-burn agriculture. Agric Ecosyst. Envi- ron., (this issue)
Pleysier, J.L. and Juo, A.S.R., 1981. Leaching of nutrient ions in an Ultisol from the high rainfall tropics: leaching through undisturbed soil columns. Soil Sci. Soc. Am. J., 45: 754-760.
Proctor, J. (Editor), 1989. Mineral Nutrients in Tropical Forest and Savanna Ecosystems. Blackwell Science Publications, Ox- ford.
Ruhigwa, B.A., Gichuru, M.P., Mambani, B. and Tariah, N.M., 1992. Root distribution of Acioa barteri, Alchornea cordifo- lia, Cassia siamea, and Gmelina arborea in acid Ultisol. Agrofor. Syst., 19: 67-78.
Saldarriaga, J.G., West, J.G., Tharp, M.L. and Uhl, C., 1988. Long term chronosequence of forest succession in the upper Rio Negro of Colombia and Venezuela. J. Ecol., 76: 938-956.
Sanchez, P.A., 1976. Properties and Management of Soils in the Tropics. John and Wiley and Sons, New York.
Sanchez, P.A., 1979. Soil fertility and conservation considerations for agroforestry systems in the humid tropics of Latin Amer- ica. In: H.O. Mongi and P.A. Huxley (Editors), Soil Research in Agroforestry. International Council for Research in Agro- forestry, Nairobi, Kenya, pp. 79-124.
Sanchez, P,A., 1985. Fertilizers make continuous cropping possi- ble in the Amazon. Better Crops Int., l: 12-15.
Sanchez, P.A. and Buol, S.W., 1975. Soils in the tropics and world food crisis. Science, 188: 598-603.
Sanchez, P.A., Bandy, D.E., Villachica, J.H. and Nicholaides, J.J., 1982. Amazon basin soils: Management for continuous crop- ping. Science, 216: 821-827.
Sanchez, P.A., Villachica, J.H. and Bandy, D.E., 1983. Soil fertility dynamics after cleating a tropical rainforest in Peru. Soil Sci. Soc. Am. J., 47: 1171-1178.
Scott, G., 1977. The importance of old-field succession biomass increments to shifting cultivation. Great Plains-Rocky Mt. Geogr. J., 6: 318-327.
Scott, G.A.J., 1978. Grassland development in the Gran Pajonal of eastern Peru. Ph.D. dissertation, Geography Department, Uni- versity of Hawaii, Honolulu.
Serrao, E.A., Falesi, I.C., Bastos de Veiga, J. and Teixeira-Neto, J.F., 1978. Productivity of cultivated pastures on low fertility soils in the Amazon of Brazil. In: P.A. Sanchez and L.E.
Tergas (Editors), Pasture Production in Acid Soils of the Tropics. Centro Intemacional de Agricultura Tropical (CIAT), Cali, Colombia, pp. 195-225.
Seubert, C.E., Sanchez, P.A. and Valverde, C., 1977. Effect of land cleating on crop performance in an UItisol of the Amazon jungle of Peru. Trop. Agric. (Trinidad), 54: 307-321.
Stromgaard, P., 1984. The immediate effect of burning and ash- fertilization. Plant Soil, 80: 307-320.
Szott, L.T., 1991. Phosphorus cycling in humid tropical succes- sional forest. In: D. Tiessen, D. Lopez-Hernandez and I.H. Salcedo (Editors), Phosphorus Cycles in Terrestrial and Aquatic Ecosystems. Institute of Pedology, Saskatoon, Canada, pp. 121-133.
Szott, L.T. and Palm, C.A., 1986. Soil and vegetation dynamics in shifting cultivation fallows. In: Simposio do Tropico Umido, Vol. l, EMBRAPA, Belem, Brazil, pp. 360-379.
Szott, L.T., Palm, C.A. and Sanchez, P.A., 1991. Agroforestry in acid soils of the humid tropics. Adv. Agron., 45: 275-301.
Tulaphitak, T., Pairintra, C. and Kyuma, K., 1985. Changes in soil fertility and soil tilth under shifting cultivation. 2: Changes in soil nutrient status. Plant Soil, 3 l: 239-249.
Uhl, C. and Jordan, C.F., 1984. Succession and nutrient dynamics following forest cutting and burning in the Amazon. Ecology, 65: 1476-1490.
Vitousek, P.M. and Sanford, R.L., 1986. Nutrient cycling in moist tropical forest. Ann. Rev. Ecol. Syst., 17: 137-167.
Williams, C.N., Chen, W.Y. and Rajartuam, J.H., 1980. Tree and Field Crops of the Wetter Regions of the Tropics. Longman, London.
Wilson, G.F. and Swennen, R., 1989. Alley cropping: potential for plantain and banana production. In: B.T. Kang and L. Reynolds (Editors), Alley Fanning in the Humid and Subhumid Tropics. IDRC, Ottawa, Canada, pp. 37-41.
Wong, M.T.F., Wild, A. and Juo, A.S.R., 1987. Retarded leaching of nitrate measured in monolith lysimeters in south-east Nige- ria. J. Soil Sci., 38: 511-518.
Yamoah, C.F., Agboola, A.A., Wilson, G.F. and Mulongoy, K., 1986. Soil properties as affected by use of leguminous shrubs for alley cropping with maize. Agric. Ecosyst. Environ., 18: 167-177.