characterization of the genetic lineages responsible for pneumococcal invasive disease in portugal
TRANSCRIPT

10.1128/JCM.43.4.1706-1715.2005.
2005, 43(4):1706. DOI:J. Clin. Microbiol. I. Serrano, J. Melo-Cristino, J. A. Carriço and M. Ramirez Disease in PortugalResponsible for Pneumococcal Invasive Characterization of the Genetic Lineages
http://jcm.asm.org/content/43/4/1706Updated information and services can be found at:
These include:
REFERENCEShttp://jcm.asm.org/content/43/4/1706#ref-list-1at:
This article cites 17 articles, 12 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on July 24, 2014 by guesthttp://jcm
.asm.org/
Dow
nloaded from
on July 24, 2014 by guesthttp://jcm
.asm.org/
Dow
nloaded from

JOURNAL OF CLINICAL MICROBIOLOGY, Apr. 2005, p. 1706–1715 Vol. 43, No. 40095-1137/05/$08.00�0 doi:10.1128/JCM.43.4.1706–1715.2005Copyright © 2005, American Society for Microbiology. All Rights Reserved.
Characterization of the Genetic Lineages Responsible forPneumococcal Invasive Disease in PortugalI. Serrano,1 J. Melo-Cristino,1 J. A. Carrico,2 and M. Ramirez1*
Institute of Molecular Medicine, Lisbon Faculty of Medicine, Lisbon,1 and Biomathematics Group,Instituto de Tecnologia Quımica e Biologica, Oeiras,2 Portugal
Received 17 August 2004/Returned for modification 24 September 2004/Accepted 6 December 2004
The availability of a conjugate vaccine has the potential to reduce the disease burden of pneumococci andto alter the serotype frequency in the disease-causing population through immunoselection. These changes willprobably be reflected in the distributions of individual genetic lineages within the population. We present acharacterization of a collection of recent (1999 to 2002) invasive isolates from Portugal (n � 465) by macro-restriction profiling with pulsed-field gel electrophoresis (PFGE) and multilocus sequence typing. During thistime, serotypes 14, 1, 3, 4, 8, 9V, 23F, 7F, 19A, and 12B were the 10 most prevalent overall by decreasing rankorder. By combining the PFGE data with the sequence types (STs) of 104 isolates, we were able to identify thegenetic lineages of the majority of the isolates. We found 66 STs, including 20 novel STs, corresponding to 47different lineages by e-BURST analysis. We found in our collection a number of previously identified inter-nationally disseminated lineages, especially among macrolide-resistant and penicillin-resistant isolates, andthese accounted for most of the isolates. Most of the major lineages (17 of 25) were identified in all years ofthe study, suggesting that the pneumococcal population associated with invasive disease was stable. This studyprovides a characterization of the pneumococcal population associated with invasive disease that will be usefulfor detecting potential selective effects of the novel conjugate vaccine.
The recent introduction of a seven-valent conjugate vaccinehas the potential to alter the genetic structure of disease-causing pneumococci by exerting a negative selective pressureagainst clones expressing vaccine serotypes. Changes in theprevalence of various serotypes causing invasive disease bothin adults and in children were already noted in a recent studyin the United States (17). However, little is known about thegenetic relationship of these emerging serotypes to previouslyestablished clones and whether the genetic structure of theclones expressing vaccine serotypes is changing. In order tocharacterize the expected changes, a detailed knowledge of thedisease-causing population prior to widespread vaccine use isneeded.
Pulsed-field gel electrophoresis (PFGE) of chromosomal mac-rorestriction digests and multilocus sequence typing (MLST)are two complementary techniques widely used to characterizethe relationships among pneumococcal isolates. PFGE allowsrapid and cost-effective clustering of genetically related iso-lates, and MLST then can be used to assign easily comparableand portable clone identifiers to clusters of similar isolates.These sequence types (STs) can be compared to those in apublic database to identify relationships to other isolates re-covered in various geographic areas.
Although there is abundant information on the geneticstructure of the population of pneumococci asymptomaticallycarried by children attending day care centers in Portugal (4),there is no current information on the molecular epidemiologyof invasive disease. We have used PFGE and MLST to char-acterize a recently described collection of isolates recovered in
Portugal during the years 1999 to 2002 (13) in order to identifythe clones responsible for invasive disease.
MATERIALS AND METHODS
Isolates. A collection of 465 sterile site isolates of Streptococcus pneumoniae(pneumococci) recovered during the years 1999 to 2002 from invasive disease inPortugal were analyzed. The collection included 88 isolates from 1999, 112isolates from 2000, 147 isolates from 2001, and 118 isolates from 2002. Theisolates were provided by 30 laboratories, geographically distributed throughoutthe country, that were asked to submit all nonduplicate invasive isolates. Theisolates expressed over 20 different serotypes; 12% were recovered from children�2 years old, and 18% were recovered from children �6 years old (Table 1). Theresults of serotyping and antimicrobial susceptibility testing of these isolates werereported previously (13).
Twenty-six strains representative of internationally disseminated clones (10)recognized by the pneumococcal molecular epidemiology network (PMEN) werekindly provided by Hermınia de Lencastre and were compared to all isolatesexpressing the same serotype or other serotypes also previously found associatedwith these clones.
PFGE and MLST. PFGE and MLST were performed as described previously(6, 14). Bionumerics software (Applied-Maths, Sint-Martens-Latem, Belgium)was used to make UPGMA (unweighted-pair group method using average link-ages) dendrograms of fragment patterns. The Dice similarity coefficient was usedwith optimization and position tolerance settings of 1.0 and 1.5%, respectively, asdescribed previously (8). PFGE-based clusters were defined as isolates with�80% relatedness on the dendrogram. DNA fragments smaller than 19 kb werenot considered for dendrogram construction.
PFGE profiling was done for 457 isolates; for the remaining 8 isolates, noPFGE pattern could be obtained in spite of multiple attempts. A major lineagewas defined as �4 isolates whose PFGE patterns clustered with a UPGMA Dicecoefficient of �80%. MLST analysis was performed for at least one isolate ineach major lineage. For large PFGE clusters, we chose a variable number ofisolates to represent the diversity observed. All non-penicillin-susceptible isolatesand isolates recovered from the cerebrospinal fluid (CSF) with PFGE profilesnot identical to those of other isolates for which MLST profiles were determinedwere also subjected to an independent MLST analysis. A subset of 104 isolateswere characterized by MLST.
MLST alleles and STs were identified by searching the pneumococcal MLSTdatabase (spneumoniae.mlst.net). Whenever new alleles were identified, se-quence traces of both strands were submitted electronically to the pneumococcal
* Corresponding author. Mailing address: Laboratory of Microbiol-ogy, Lisbon Faculty of Medicine, Av. Prof. Egas Moniz PT 1649-028,Lisbon, Portugal. Phone: 351-21 799 9460. Fax: 351-21 799 9464. E-mail: [email protected].
1706
on July 24, 2014 by guesthttp://jcm
.asm.org/
Dow
nloaded from
database curator for approval. Lineage assignment was done by use of e-BURSTanalysis (7) with default parameters and the complete S. pneumoniae database(spneumoniae.mlst.net).
Statistical tests were performed by use of the SPSS 10 software package, andP values of �0.05 were considered significant.
RESULTS
New MLST alleles and allelic profiles. We identified sevennovel alleles among the genes used in MLST, one in gdh (89),one in gki (98), one in recP (66), one in spi (97), one in xpt
(144), and two in ddl (44 and 45). New allele combinationsproducing novel STs were also noted and were numbered 790,1220 to 1226, 1230 to 1233, and 1365 to 1372; these weredistributed in 18 different groups by e-BURST analysis. Boththe novel alleles and the novel STs were submitted to theS. pneumoniae MLST database (spneumoniae.mlst.net).
Lineage distribution within individual serotypes. Below wedescribe in detail the clones found among the 10 most frequentserotypes in our collection as well as the 10 most frequently
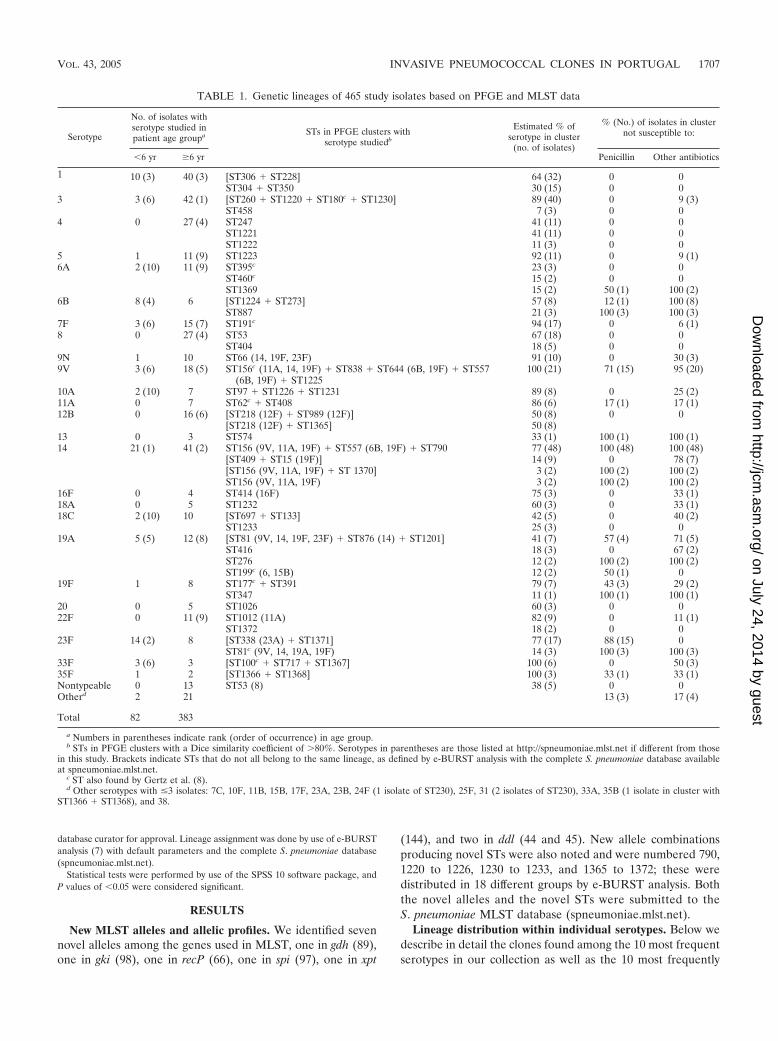
TABLE 1. Genetic lineages of 465 study isolates based on PFGE and MLST data
Serotype
No. of isolates withserotype studied inpatient age groupa STs in PFGE clusters with
serotype studiedb
Estimated % ofserotype in cluster
(no. of isolates)
% (No.) of isolates in clusternot susceptible to:
�6 yr �6 yr Penicillin Other antibiotics
1 10 (3) 40 (3) [ST306 � ST228] 64 (32) 0 0ST304 � ST350 30 (15) 0 0
3 3 (6) 42 (1) [ST260 � ST1220 � ST180c � ST1230] 89 (40) 0 9 (3)ST458 7 (3) 0 0
4 0 27 (4) ST247 41 (11) 0 0ST1221 41 (11) 0 0ST1222 11 (3) 0 0
5 1 11 (9) ST1223 92 (11) 0 9 (1)6A 2 (10) 11 (9) ST395c 23 (3) 0 0
ST460c 15 (2) 0 0ST1369 15 (2) 50 (1) 100 (2)
6B 8 (4) 6 [ST1224 � ST273] 57 (8) 12 (1) 100 (8)ST887 21 (3) 100 (3) 100 (3)
7F 3 (6) 15 (7) ST191c 94 (17) 0 6 (1)8 0 27 (4) ST53 67 (18) 0 0
ST404 18 (5) 0 09N 1 10 ST66 (14, 19F, 23F) 91 (10) 0 30 (3)9V 3 (6) 18 (5) ST156c (11A, 14, 19F) � ST838 � ST644 (6B, 19F) � ST557
(6B, 19F) � ST1225100 (21) 71 (15) 95 (20)
10A 2 (10) 7 ST97 � ST1226 � ST1231 89 (8) 0 25 (2)11A 0 7 ST62c � ST408 86 (6) 17 (1) 17 (1)12B 0 16 (6) [ST218 (12F) � ST989 (12F)] 50 (8) 0 0
[ST218 (12F) � ST1365] 50 (8)13 0 3 ST574 33 (1) 100 (1) 100 (1)14 21 (1) 41 (2) ST156 (9V, 11A, 19F) � ST557 (6B, 19F) � ST790 77 (48) 100 (48) 100 (48)
[ST409 � ST15 (19F)] 14 (9) 0 78 (7)[ST156 (9V, 11A, 19F) � ST 1370] 3 (2) 100 (2) 100 (2)ST156 (9V, 11A, 19F) 3 (2) 100 (2) 100 (2)
16F 0 4 ST414 (16F) 75 (3) 0 33 (1)18A 0 5 ST1232 60 (3) 0 33 (1)18C 2 (10) 10 [ST697 � ST133] 42 (5) 0 40 (2)
ST1233 25 (3) 0 019A 5 (5) 12 (8) [ST81 (9V, 14, 19F, 23F) � ST876 (14) � ST1201] 41 (7) 57 (4) 71 (5)
ST416 18 (3) 0 67 (2)ST276 12 (2) 100 (2) 100 (2)ST199c (6, 15B) 12 (2) 50 (1) 0
19F 1 8 ST177c � ST391 79 (7) 43 (3) 29 (2)ST347 11 (1) 100 (1) 100 (1)
20 0 5 ST1026 60 (3) 0 022F 0 11 (9) ST1012 (11A) 82 (9) 0 11 (1)
ST1372 18 (2) 0 023F 14 (2) 8 [ST338 (23A) � ST1371] 77 (17) 88 (15) 0
ST81c (9V, 14, 19A, 19F) 14 (3) 100 (3) 100 (3)33F 3 (6) 3 [ST100c � ST717 � ST1367] 100 (6) 0 50 (3)35F 1 2 [ST1366 � ST1368] 100 (3) 33 (1) 33 (1)Nontypeable 0 13 ST53 (8) 38 (5) 0 0Otherd 2 21 13 (3) 17 (4)
Total 82 383
a Numbers in parentheses indicate rank (order of occurrence) in age group.b STs in PFGE clusters with a Dice similarity coefficient of �80%. Serotypes in parentheses are those listed at http://spneumoniae.mlst.net if different from those
in this study. Brackets indicate STs that do not all belong to the same lineage, as defined by e-BURST analysis with the complete S. pneumoniae database availableat spneumoniae.mlst.net.
c ST also found by Gertz et al. (8).d Other serotypes with �3 isolates: 7C, 10F, 11B, 15B, 17F, 23A, 23B, 24F (1 isolate of ST230), 25F, 31 (2 isolates of ST230), 33A, 35B (1 isolate in cluster with
ST1366 � ST1368), and 38.
VOL. 43, 2005 INVASIVE PNEUMOCOCCAL CLONES IN PORTUGAL 1707
on July 24, 2014 by guesthttp://jcm
.asm.org/
Dow
nloaded from

found among isolates recovered from children �6 years old(Table 1). The importance of this age group relates to currentrecommendations suggesting vaccination of children who are�6 years old and who attend day care centers (3), since mostchildren in Portugal are in this category. The data are summa-rized in Table 1, which also shows clones also found in a surveyof U.S. invasive isolates carried out by Gertz et al. (8).
(i) Serotype 1. Two main lineages could be seen in serotype1, each represented by two STs—a cluster including the ma-jority of isolates representing ST306 and ST228 and anotherincluding ST304 and ST350, both double-locus variants of eachother (Fig. 1). These four STs were previously shown to bemembers of different branches of a lineage found associatedwith invasive disease exclusively in Europe and North Ameri-ca—lineage A (2).
(ii) Serotype 3. Although two main PFGE clusters could beidentified in serotype 3, the cluster containing the majority ofthe isolates was clearly divided into two unrelated lineages bye-BURST analysis, suggesting that isolates expressing this se-rotype belong to three lineages (Fig. 1). The lineage represent-ing more isolates (n � 25) contained two single-locus vari-ants—ST260, which was previously found to be an importantclone in Spain (5), and the novel ST1220. In the same PFGEcluster was another lineage (n � 15), differing at all seven lociused in MLST from ST260 and ST1220 and also including twosingle-locus variants, ST180 and ST1230, representing a glo-bally distributed lineage previously found to be associated withinvasive disease in Europe, Taiwan, Canada, and the UnitedStates (6, 8). The third lineage included three isolates repre-senting ST458, which was previously found to be associatedwith invasive disease in England (spneumoniae.mlst.net).
(iii) Serotype 4. Equal numbers of isolates (n � 11) weredistributed in two different lineages, ST247 and ST1221 (Fig.1). ST247 was found to be associated with invasive disease in anumber of European countries (6) and belongs to the lineagewith the largest number of related STs expressing serotype 4presently in the database. ST1221 is a novel ST that belongs tothe same lineage as ST993, which was previously found in anisolate from invasive disease in Scotland. Three isolates belong
to a third lineage, ST1222, a novel ST belonging to the samelineage as ST800 and ST801, which were previously found ininvasive isolates in the Czech Republic.
(iv) Serotype 6A. Only three small clusters of related PFGEprofiles were observed in serotype 6A (Fig. 1). A cluster ofthree isolates included an isolate recovered from the CSF andrepresenting ST395, which was previously found to be associ-ated only with carriage in England (1). Two other clusterscontained a non-penicillin-susceptible isolate with the novelST1369 and an isolate recovered from the CSF and represent-ing ST460, which was previously found to be associated with ameningitis case in England (spneumoniae.mlst.net). The twoisolates from children were found in the ST460 cluster and hada PFGE profile unrelated to any other in this serotype.
(v) Serotype 6B. Two PFGE clusters contained the majorityof the isolates expressing serotype 6B. The larger PFGE clus-ter (n � 8) included isolates related (ST1224 and ST273) tothe internationally disseminated clone Greece6B-22 (www.sph.emory.edu/PMEN/pmen_clone_collection.html), with the ex-ception of a non-penicillin-susceptible isolate belonging to theSpain6B-2 lineage (10) (Fig. 2). The smaller PFGE cluster (n �3) included only non-penicillin-susceptible isolates related(ST887) to clone Poland6B-20 (www.sph.emory.edu/PMEN/pmen_clone_collection.html). Three isolates were not groupedin these clusters.
(vi) Serotype 7F. With the exception of a single isolate, allother isolates (n � 17) were grouped in a single PFGE clus-ter representing ST191 (Fig. 3), which is found worldwide inpneumococci expressing serotype 7F (spneumoniae.mlst.net).A dominance of this clone in invasive isolates expressing sero-type 7F was documented in the United States (8).
(vii) Serotype 8. The majority of the isolates (n � 18) belongto a cluster representing ST53 (Fig. 3), a clone previouslyfound to be associated with invasive disease in various Euro-pean countries and Brazil (spneumoniae.mlst.net). Anothercluster (n � 5) included isolates of ST404, which was previ-ously found to be associated uniquely with carriage in childrenin the United Kingdom (1). This constitutes the first report of
FIG. 2. Serotype 6B. For details, see the legend to Fig. 1.
1708 SERRANO ET AL. J. CLIN. MICROBIOL.
on July 24, 2014 by guesthttp://jcm
.asm.org/
Dow
nloaded from
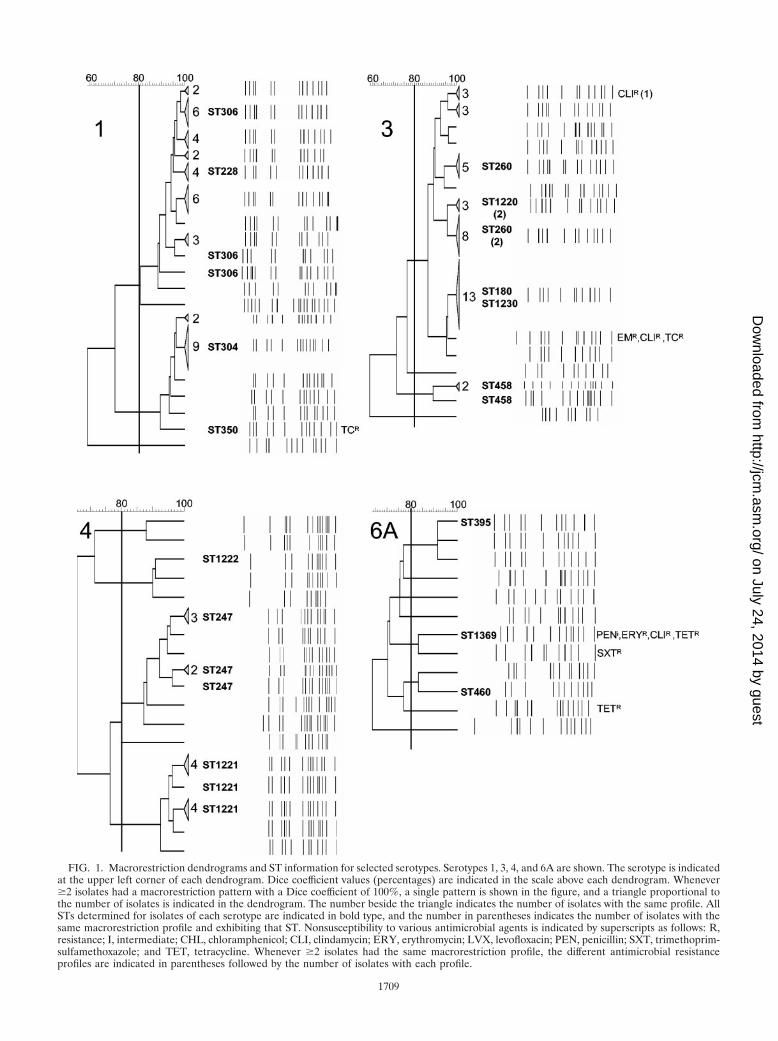
FIG. 1. Macrorestriction dendrograms and ST information for selected serotypes. Serotypes 1, 3, 4, and 6A are shown. The serotype is indicatedat the upper left corner of each dendrogram. Dice coefficient values (percentages) are indicated in the scale above each dendrogram. Whenever�2 isolates had a macrorestriction pattern with a Dice coefficient of 100%, a single pattern is shown in the figure, and a triangle proportional tothe number of isolates is indicated in the dendrogram. The number beside the triangle indicates the number of isolates with the same profile. AllSTs determined for isolates of each serotype are indicated in bold type, and the number in parentheses indicates the number of isolates with thesame macrorestriction profile and exhibiting that ST. Nonsusceptibility to various antimicrobial agents is indicated by superscripts as follows: R,resistance; I, intermediate; CHL, chloramphenicol; CLI, clindamycin; ERY, erythromycin; LVX, levofloxacin; PEN, penicillin; SXT, trimethoprim-sulfamethoxazole; and TET, tetracycline. Whenever �2 isolates had the same macrorestriction profile, the different antimicrobial resistanceprofiles are indicated in parentheses followed by the number of isolates with each profile.
1709
on July 24, 2014 by guesthttp://jcm
.asm.org/
Dow
nloaded from

an association of this lineage with invasive disease, including acase of meningitis.
(viii) Serotype 9V. Serotype 9V included a large number ofnon-penicillin-susceptible isolates. All isolates were grouped ina single PFGE cluster related to the internationally dissemi-nated clone Spain9V-3 (10), in spite of having STs that differedin up to three loci—ST156, ST557, ST644, ST838, and ST1225(Fig. 4). e-BURST analysis of the MLST profiles confirmedthat these isolates belong to a single lineage, with the ST ofclone Spain9V-3 being the predicted founder.
(ix) Serotype 10A. A single large PFGE cluster (n � 8)included all but one of the isolates expressing this serotype(Fig. 3). In spite of the different STs found in isolates of thiscluster—ST97, ST1226, and ST1231—they were found bye-BURST analysis to represent the same lineage, with ST97being the predicted founder.
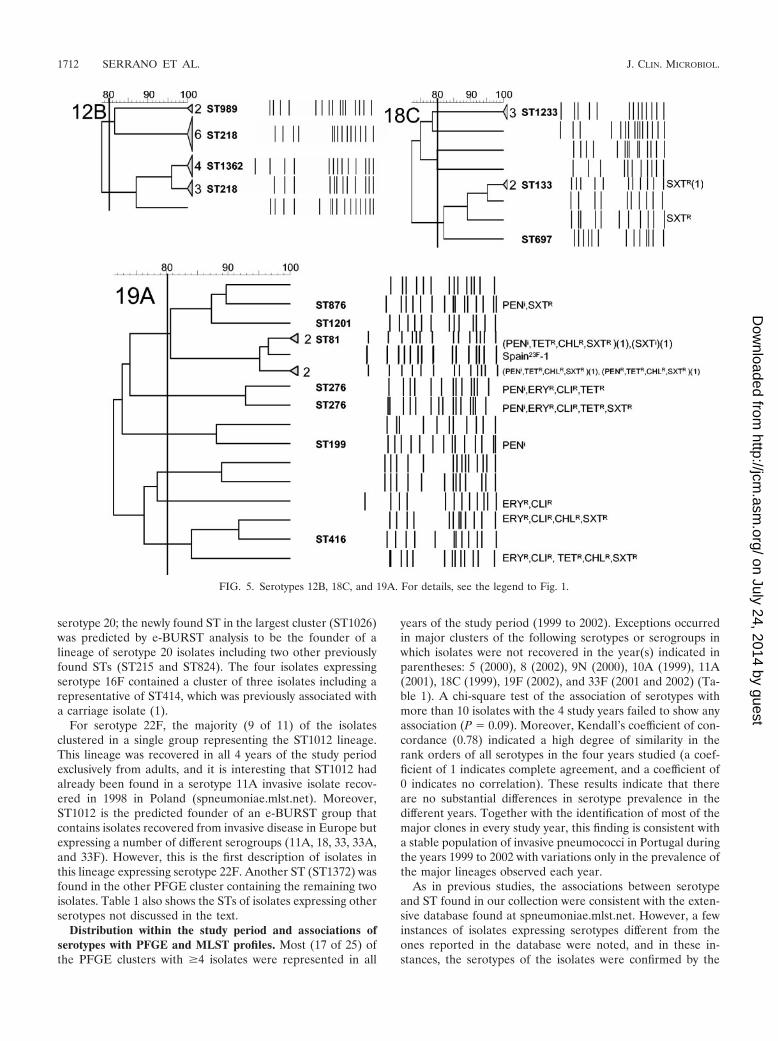
(x) Serotype 12B. Although serotype 12B isolates weregrouped in two PFGE clusters (Fig. 5), both included repre-sentatives of ST218, which was previously found to be associ-ated exclusively with isolates expressing serotype 12F and re-covered from patients with invasive disease in Europe, NorthAmerica, and South America (6, 8). Both clusters containedeight isolates; one contained a representative of ST989 (whichwas previously found to be associated with isolates expressingserotype 6B and recovered from the nasopharynx), and theother contained the novel ST1365, a singleton (does not groupwith any other ST), as determined by e-BURST analysis. TheMLST data suggested that these lineages constitute separatebranches and that the observed PFGE clustering does notreflect true genetic relationships. Also of note is the fact thatall isolates in this collection with ST218 express serotype 12B
and not serotype 12F, as was found previously (8). Althoughthis finding could be attributable to capsular switching (11), itis also possible that interconversion between these serotypesfollows a route similar to the one recently described for sero-types 15B and 15C (16).
(xi) Serotype 14. Most isolates (84%) expressing serotype 14were non-penicillin-susceptible. A single cluster comprisingonly non-penicillin-susceptible isolates included most of theisolates (Fig. 6). This cluster was found to be related to cloneSpain9V-3 (10) both by PFGE and by e-BURST analysis andincluded representatives of ST156, ST557, and the novelST790. In two other clusters of non-penicillin-susceptible iso-lates, three isolates were representatives of ST156 as well,raising the proportion of isolates belonging to this lineage to98% of all non-penicillin-susceptible isolates expressing thisserotype. The second largest cluster (n � 10) was composed ofisolates related to clone England14-9 (10), including a repre-sentative of ST15, all fully susceptible to penicillin.
(xii) Serotype 18C. The largest cluster contained five iso-lates, but the two MLST profiles determined shared only twoalleles, suggesting that this cluster represents diverse lineages(Fig. 5). Both ST133 and ST697 were previously identified inserotype 18C isolates recovered from CSF in Spain and En-gland, respectively (spneumoniae.mlst.net). The second largestcluster contained three isolates, including the isolates fromchildren �6 years old, and represented the novel ST1233,which was not found to be related to any other lineage bye-BURST analysis.
(xiii) Serotype 19A. The largest cluster (n � 8) containedtwo distinct branches—a set of closely associated isolatesof ST81 (n � 5), belonging to the same lineage as clone
FIG. 3. Serotypes 7F, 8, and 10A. For details, see the legend to Fig. 1.
1710 SERRANO ET AL. J. CLIN. MICROBIOL.
on July 24, 2014 by guesthttp://jcm
.asm.org/
Dow
nloaded from

Spain23F-1 (10), and another group (n � 3) with representa-tives of two unrelated lineages, ST876 and ST1201 (Fig. 5).Both of the latter STs had been reported only once previously(spneumoniae.mlst.net) and were found by e-BURST analysisto belong to groups with a large number of STs and differentserotypes. This cluster contains 57% (n � 4) of the non-peni-cillin-susceptible isolates expressing serotype 19A, three relat-ed to Spain23F-1 and the other belonging to ST876; both ofthese lineages had already been associated with penicillin non-susceptibility (spneumoniae.mlst.net). The remaining three non-penicillin-susceptible isolates were found in clusters of twoisolates of ST276 and one isolate of ST199. Non-penicillin-sus-ceptible serotype 19 isolates had been identified previously inboth STs (6). The remaining five isolates included a cluster of twoisolates; a cluster of three isolates representing lineage ST416,which was previously found in serotype 19A isolates recoveredfrom blood and associated with carriage (1, 9); and an isolatewith a PFGE profile unrelated to any other in this serotype.
(xiv) Serotype 23F. The majority of the isolates were groupedin a single cluster (n � 17) containing mostly non-penicillin-susceptible isolates of ST338, the same lineage as the widelydistributed clone Colombia23F-26 (Fig. 7). We also observed asmaller cluster (n � 3) that also included non-penicillin-sus-ceptible isolates related to clone Spain23F-1 (ST81).
(xv) Serotype 33F. Although a single PFGE cluster wasidentified, it contained representatives of at least three inde-pendent lineages, ST1367, ST100, and ST717 (Fig. 7). ST717was found exclusively in the three isolates recovered fromchildren �6 years old; these isolates formed a cluster withidentical PFGE profiles.
Other serotypes. We analyzed by MLST representatives of anumber of other serotypes expressed by significant numbers of
isolates in our collection; the data are summarized in Table 1.Among serotype 5 isolates, the majority (11 of 12) belong tothe novel ST1223, are included in a single cluster, and belongto the same lineage as the internationally disseminated cloneColombia5-19.
A single cluster contained all but one of the isolates express-ing serotype 9N (n � 10). Isolates in this cluster belong toST66, similar to what was recently found for isolates recoveredfrom the nasopharynges of healthy children in the UnitedKingdom (1) but in contrast to the results of a study of invasiveisolates in the United States, where ST632 dominated (8).
The majority (six of seven) of the isolates expressing sero-type 11A were contained in a single PFGE cluster includingmembers of ST62 and ST408, both of which were found bye-BURST analysis to belong to the same lineage. This lineagewas also the most prevalent in both carriage isolates from theUnited Kingdom and invasive isolates from the United States(2, 8), suggesting that it is widely disseminated.
Although serotype 19F was only the 18th overall in ourcollection and a single isolate was recovered from children �6years old, 44% (four of nine) of the isolates expressing thisserotype were non-penicillin-susceptible. The largest cluster(n � 7) was found to be related to clone Portugal19F-21 both byPFGE and by MLST (ST177 and ST391); this clone was foundfrequently in isolates resistant to at least one antimicrobialagent and recovered from the nasopharynges of healthy chil-dren in Portugal (12).
Among the five isolates expressing serotype 18A, three wererecovered in 2002 and were found to be identical by PFGE;they included a member of the novel ST1232. The other twoisolates were grouped in another cluster. A similar distributionin two clusters was observed for the five isolates expressing
FIG. 4. Serotype 9V. For details, see the legend to Fig. 1.
VOL. 43, 2005 INVASIVE PNEUMOCOCCAL CLONES IN PORTUGAL 1711
on July 24, 2014 by guesthttp://jcm
.asm.org/
Dow
nloaded from
serotype 20; the newly found ST in the largest cluster (ST1026)was predicted by e-BURST analysis to be the founder of alineage of serotype 20 isolates including two other previouslyfound STs (ST215 and ST824). The four isolates expressingserotype 16F contained a cluster of three isolates including arepresentative of ST414, which was previously associated witha carriage isolate (1).
For serotype 22F, the majority (9 of 11) of the isolatesclustered in a single group representing the ST1012 lineage.This lineage was recovered in all 4 years of the study periodexclusively from adults, and it is interesting that ST1012 hadalready been found in a serotype 11A invasive isolate recov-ered in 1998 in Poland (spneumoniae.mlst.net). Moreover,ST1012 is the predicted founder of an e-BURST group thatcontains isolates recovered from invasive disease in Europe butexpressing a number of different serogroups (11A, 18, 33, 33A,and 33F). However, this is the first description of isolates inthis lineage expressing serotype 22F. Another ST (ST1372) wasfound in the other PFGE cluster containing the remaining twoisolates. Table 1 also shows the STs of isolates expressing otherserotypes not discussed in the text.
Distribution within the study period and associations ofserotypes with PFGE and MLST profiles. Most (17 of 25) ofthe PFGE clusters with �4 isolates were represented in all
years of the study period (1999 to 2002). Exceptions occurredin major clusters of the following serotypes or serogroups inwhich isolates were not recovered in the year(s) indicated inparentheses: 5 (2000), 8 (2002), 9N (2000), 10A (1999), 11A(2001), 18C (1999), 19F (2002), and 33F (2001 and 2002) (Ta-ble 1). A chi-square test of the association of serotypes withmore than 10 isolates with the 4 study years failed to show anyassociation (P � 0.09). Moreover, Kendall’s coefficient of con-cordance (0.78) indicated a high degree of similarity in therank orders of all serotypes in the four years studied (a coef-ficient of 1 indicates complete agreement, and a coefficient of0 indicates no correlation). These results indicate that thereare no substantial differences in serotype prevalence in thedifferent years. Together with the identification of most of themajor clones in every study year, this finding is consistent witha stable population of invasive pneumococci in Portugal duringthe years 1999 to 2002 with variations only in the prevalence ofthe major lineages observed each year.
As in previous studies, the associations between serotypeand ST found in our collection were consistent with the exten-sive database found at spneumoniae.mlst.net. However, a fewinstances of isolates expressing serotypes different from theones reported in the database were noted, and in these in-stances, the serotypes of the isolates were confirmed by the
FIG. 5. Serotypes 12B, 18C, and 19A. For details, see the legend to Fig. 1.
1712 SERRANO ET AL. J. CLIN. MICROBIOL.
on July 24, 2014 by guesthttp://jcm
.asm.org/
Dow
nloaded from
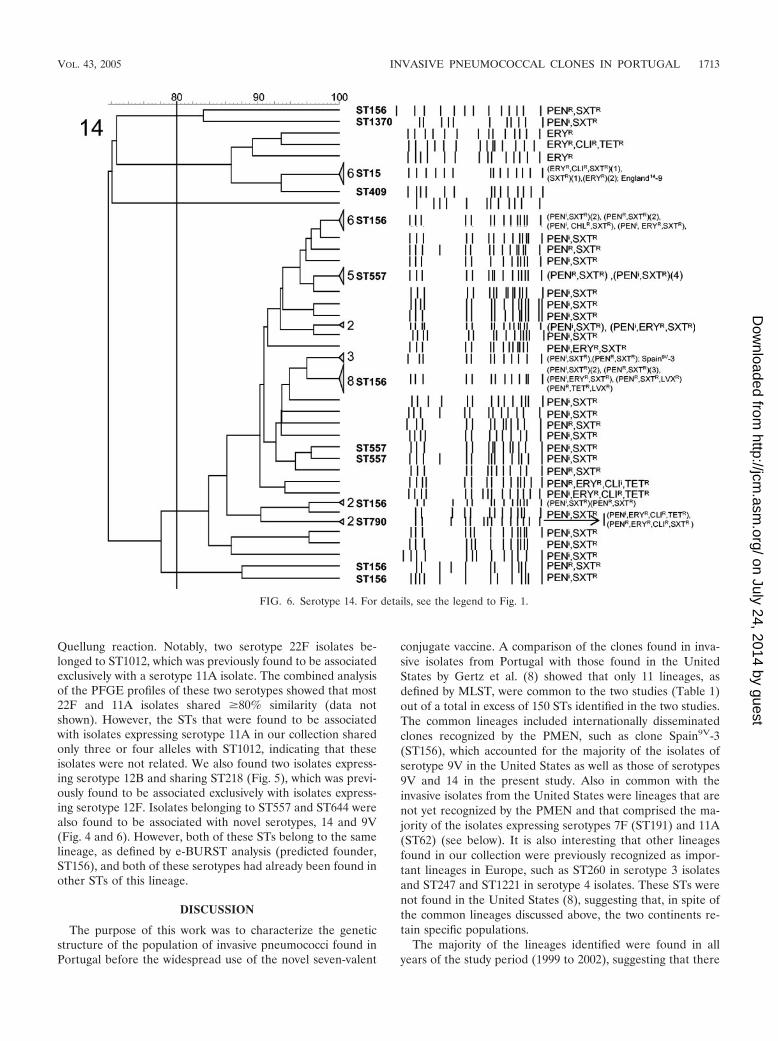
Quellung reaction. Notably, two serotype 22F isolates be-longed to ST1012, which was previously found to be associatedexclusively with a serotype 11A isolate. The combined analysisof the PFGE profiles of these two serotypes showed that most22F and 11A isolates shared �80% similarity (data notshown). However, the STs that were found to be associatedwith isolates expressing serotype 11A in our collection sharedonly three or four alleles with ST1012, indicating that theseisolates were not related. We also found two isolates express-ing serotype 12B and sharing ST218 (Fig. 5), which was previ-ously found to be associated exclusively with isolates express-ing serotype 12F. Isolates belonging to ST557 and ST644 werealso found to be associated with novel serotypes, 14 and 9V(Fig. 4 and 6). However, both of these STs belong to the samelineage, as defined by e-BURST analysis (predicted founder,ST156), and both of these serotypes had already been found inother STs of this lineage.
DISCUSSION
The purpose of this work was to characterize the geneticstructure of the population of invasive pneumococci found inPortugal before the widespread use of the novel seven-valent
conjugate vaccine. A comparison of the clones found in inva-sive isolates from Portugal with those found in the UnitedStates by Gertz et al. (8) showed that only 11 lineages, asdefined by MLST, were common to the two studies (Table 1)out of a total in excess of 150 STs identified in the two studies.The common lineages included internationally disseminatedclones recognized by the PMEN, such as clone Spain9V-3(ST156), which accounted for the majority of the isolates ofserotype 9V in the United States as well as those of serotypes9V and 14 in the present study. Also in common with theinvasive isolates from the United States were lineages that arenot yet recognized by the PMEN and that comprised the ma-jority of the isolates expressing serotypes 7F (ST191) and 11A(ST62) (see below). It is also interesting that other lineagesfound in our collection were previously recognized as impor-tant lineages in Europe, such as ST260 in serotype 3 isolatesand ST247 and ST1221 in serotype 4 isolates. These STs werenot found in the United States (8), suggesting that, in spite ofthe common lineages discussed above, the two continents re-tain specific populations.
The majority of the lineages identified were found in allyears of the study period (1999 to 2002), suggesting that there
FIG. 6. Serotype 14. For details, see the legend to Fig. 1.
VOL. 43, 2005 INVASIVE PNEUMOCOCCAL CLONES IN PORTUGAL 1713
on July 24, 2014 by guesthttp://jcm
.asm.org/
Dow
nloaded from

were no major changes in overall diversity during this time.Widespread vaccine usage has the potential to change theserotype prevalence in the pneumococcal population causinginvasive disease, as was recently demonstrated in the UnitedStates (17). This change in serotype composition will probablybe paralleled by an alteration in the clonal composition of thepopulation, but it is also possible that transformation to non-vaccine serotypes of the presently circulating clones will playan important role (11).
It is interesting that although large clusters of related iso-lates were frequently observed, the overall diversity withinserotypes varied greatly (Fig. 1 to 6). Serotypes including al-most exclusively a single lineage (7F) (Fig. 3) contrasted withothers in which small unrelated clusters were found (6A) (Fig.1). Even though it was previously suggested that isolates sus-ceptible to antimicrobial agents are more diverse than penicil-lin-resistant ones (15), the intrinsic properties of each serotypeclearly play an important role, since both serotype 6A andserotype 7F included mostly isolates susceptible to all antimi-crobial agents tested. Moreover, the diversity found in serotype1 isolates was less than that found in serotype 14 isolates inspite of the fact that the latter serotype included mostly isolatesresistant to at least one of the antimicrobial agents tested andserotype 1 included overwhelmingly fully susceptible isolates.
These observations could be a direct consequence of the di-versity of the global pneumococcal population expressing eachserotype, perhaps due to recent evolutionary history, or couldbe related to the selection of successful invasive clones inPortugal. Knowledge of the population of asymptomaticallycarried pneumococci is necessary to begin to address thesequestions. Although detailed studies of the pneumococci car-ried by children attending day care centers in Portugal havebeen published, they have so far been restricted to resistantisolates (4, 12), preventing a direct comparison of the twopopulations. In spite of these limitations, it is already clear thata lineage found frequently in our collection, Spain9V-3 (see be-low), was also frequently found colonizing the nasopharyngesof children attending day care centers in the Lisbon area (4).
A number of internationally disseminated clones (www.sph.emory.edu/PMEN/pmen_clone_collection.html) were foundin our collection. Isolates related to clone Spain9V-3 (express-ing either serotype 9V or 14) and clone Colombia23F-26 ac-counted for 72% of non-penicillin-susceptible isolates. Most ofthe remaining non-penicillin-susceptible isolates were also re-lated to other clones recognized by the PMEN (10). The dom-inance of internationally disseminated clones, including thetwo penicillin-susceptible clones Greece6B-22 and England14-9,was also found in erythromycin-resistant isolates; isolates be-
FIG. 7. Serotypes 23F and 33F. For details, see the legend to Fig. 1.
1714 SERRANO ET AL. J. CLIN. MICROBIOL.
on July 24, 2014 by guesthttp://jcm
.asm.org/
Dow
nloaded from

longing to clusters including clones recognized by the PMENaccounted for 59% of resistant isolates (27 of 46) (Fig. 2 and6). Notwithstanding these data, other large clusters of relatedisolates that accounted for large proportions of the isolatesexpressing each serotype and that were not related to clonesalready accepted by the PMEN were also identified (e.g., theST191 lineage in serotype 7F or the ST180 lineage in serotype3). However, most of these isolates were fully susceptible to allantimicrobial agents tested and would therefore fall outsidethe stated aim of PMEN of identifying antibiotic-resistantclones (10). It will be interesting to monitor the expectedchanges in serotype prevalence due to vaccine use and todetermine whether these will lead to an overall reduction inresistance rates due to negative selection for these clones ex-pressing vaccine serotypes or whether capsular transformationevents will allow the persistence of these genotypes. Also note-worthy is the absence of PMEN clones that were found in theUnited States by Gertz et al. (8), namely, those that comprisedsignificant portions of the total population of their respectiveserotypes, specifically clones Taiwan19F-14 and Taiwan23F-16.
Gertz et al. (8) showed, using the same analysis parametersand methods that we applied, that most of the isolates belong-ing to clusters differing in individual PFGE patterns by aUPGMA Dice coefficient of �20% had �5 shared alleles intheir MLST profiles (8). However, these authors also describeda number of exceptions; e.g., isolates differing at more than twoloci had PFGE profiles with �80% relatedness. An analysis ofour own collection largely confirms the correlation observed byGertz et al. (8), but exceptions were also noted. In serotype 9V,ST838 and ST1225 are triple-locus variants of each other inspite of the fact that their PFGE profiles differ by �20% (Fig.4). Similar situations were noted with ST790 and ST156 inserotype 14 (Fig. 6) and ST717 and ST100 in serotype 33F (Fig.7). In serotype 19A and 12B isolates, two PFGE clusters with�80% relatedness contained STs sharing only a single allele—ST876 and ST81 and ST989 and ST218 (Fig. 5), respectively. Inserotype 3, ST260 and ST180 did not share any of the allelesat the seven MLST loci, but their PFGE profiles differed bya UPGMA Dice coefficient of �20%. Notwithstanding thesedifferences in MLST profiles, the STs differing at three loci inserotypes 9V and 14 belonged to the same lineage as thatdefined by e-BURST analysis, suggesting that the PFGE datareflect true relatedness. By combining PFGE and MLST data,we were able to infer the STs of the majority of isolates ana-lyzed (Table 1).
In conclusion, this study provides a characterization of thepneumococcal population causing invasive disease in Portugalbefore widespread vaccine use. This information will allow in-vestigators to address important issues, such as the impact ofvaccination on the genetic structure of the population causinginvasive disease and, together with studies of the carriage pop-ulation, the relative weights of serotypes and other clonal prop-erties in determining invasiveness.
ACKNOWLEDGMENTS
This work was partly supported by Fundacao Calouste Gulbenkian,contract LSHM-CT-2003-503413 (Pneumococcal Resistance Epide-micity and Virulence—an International Study) (PREVIS), from the
European Commission and Fundacao para a Ciencia e Tecnologia(POCTI/ESP/47914). I.S. was the recipient of a fellowship from Fun-dacao para a Ciencia e Tecnologia (SFRH/BD/14158/2003), and J.A.C.was the recipient of a similar fellowship (SFRH/BD/3123/2000). Wethank Francisco Pinto for help with the statistical tests and Hermıniade Lencastre for providing the PMEN reference strains.
We acknowledge the use of the pneumococcal MLST database,which is located at Imperial College London and is funded by theWellcome Trust
REFERENCES
1. Brueggemann, A. B., D. T. Griffiths, E. Meats, T. Peto, D. W. Crook, andB. G. Spratt. 2003. Clonal relationships between invasive and carriage Strep-tococcus pneumoniae and serotype- and clone-specific differences in invasivedisease potential. J. Infect. Dis. 187:1424–1432.
2. Brueggemann, A. B., and B. G. Spratt. 2003. Geographic distribution andclonal diversity of Streptococcus pneumoniae serotype 1 isolates. J. Clin.Microbiol. 41:4966–4970.
3. Centers for Disease Control and Prevention. 2000. Preventing pneumococcaldisease among infants and young children: recommendations of the AdvisoryCommittee on Immunization Practices (ACIP). Morb. Mortal. Wkly. Rep.49:1–29.
4. de Lencastre, H., and A. Tomasz. 2002. From ecological reservoir to disease:the nasopharynx, day-care centres and drug-resistant clones of Streptococcuspneumoniae. J. Antimicrob. Chemother. 50(Suppl. S2):75–81.
5. Enright, M. C., A. Fenoll, D. Griffiths, and B. G. Spratt. 1999. The threemajor Spanish clones of penicillin-resistant Streptococcus pneumoniae are themost common clones recovered in recent cases of meningitis in Spain.J. Clin. Microbiol. 37:3210–3216.
6. Enright, M. C., and B. G. Spratt. 1998. A multilocus sequence typing schemefor Streptococcus pneumoniae: identification of clones associated with seriousinvasive disease. Microbiology 144:3049–3060.
7. Feil, E. J., B. C. Li, D. M. Aanensen, W. P. Hanage, and B. G. Spratt. 2004.eBURST: inferring patterns of evolutionary descent among clusters of re-lated bacterial genotypes from multilocus sequence typing data. J. Bacteriol.186:1518–1530.
8. Gertz, R. E., Jr., M. C. McEllistrem, D. J. Boxrud, Z. Li, V. Sakota, T. A.Thompson, R. R. Facklam, J. M. Besser, L. H. Harrison, C. G. Whitney, andB. Beall. 2003. Clonal distribution of invasive pneumococcal isolates fromchildren and selected adults in the United States prior to seven-valent con-jugate vaccine introduction. J. Clin. Microbiol. 41:4194–4216.
9. Johnson, A. P., C. L. Sheppard, S. J. Harnett, A. Birtles, T. Harrison, N. P.Brenwald, M. J. Gill, R. A. Walker, D. M. Livermore, and R. C. George. 2003.Emergence of a fluoroquinolone-resistant strain of Streptococcus pneu-moniae in England. J. Antimicrob. Chemother. 52:953–960.
10. McGee, L., L. McDougal, J. Zhou, B. G. Spratt, F. C. Tenover, R. George, R.Hakenbeck, W. Hryniewicz, J. C. Lefevre, A. Tomasz, and K. P. Klugman.2001. Nomenclature of major antimicrobial-resistant clones of Streptococcuspneumoniae defined by the pneumococcal molecular epidemiology network.J. Clin. Microbiol. 39:2565–2571.
11. Nesin, M., M. Ramirez, and A. Tomasz. 1998. Capsular transformation of amultidrug-resistant Streptococcus pneumoniae in vivo. J. Infect. Dis. 177:707–713.
12. Sa-Leao, R., A. Tomasz, and H. de Lencastre. 2001. Multilocus sequencetyping of Streptococcus pneumoniae clones with unusual drug resistancepatterns: genetic backgrounds and relatedness to other epidemic clones.J. Infect. Dis. 184:1206–1210.
13. Serrano, I., M. Ramirez, J. Melo-Cristino, et al. 2004. Invasive Streptococcuspneumoniae from Portugal: implications for vaccination and antimicrobialtherapy. Clin. Microbiol. Infect. 10:652–656.
14. Soares, S., K. G. Kristinsson, J. M. Musser, and A. Tomasz. 1993. Evidencefor the introduction of a multiresistant clone of serotype 6B Streptococcuspneumoniae from Spain to Iceland in the late 1980s. J. Infect. Dis. 168:158–163.
15. Tomasz, A., A. Corso, E. P. Severina, G. Echaniz-Aviles, M. C. Brandileone,T. Camou, E. Castaneda, O. Figueroa, A. Rossi, and J. L. Di Fabio. 1998.Molecular epidemiologic characterization of penicillin-resistant Streptococ-cus pneumoniae invasive pediatric isolates recovered in six Latin-Americancountries: an overview. Microb. Drug Resist. 4:195–207.
16. van Selm, S., L. M. van Cann, M. A. B. Kolkman, B. A. M. van der Zeijst, andJ. P. M. van Putten. 2003. Genetic basis for the structural difference betweenStreptococcus pneumoniae serotype 15B and 15C capsular polysaccharides.Infect. Immun. 71:6192–6198.
17. Whitney, C. G., M. M. Farley, J. Hadler, L. H. Harrison, N. M. Bennett, R.Lynfield, A. Reingold, P. R. Cieslak, T. Pilishvili, D. Jackson, R. R. Facklam,J. H. Jorgensen, and A. Schuchat. 2003. Decline in invasive pneumococcaldisease after the introduction of protein-polysaccharide conjugate vaccine.N. Engl. J. Med. 348:1737–1746.
VOL. 43, 2005 INVASIVE PNEUMOCOCCAL CLONES IN PORTUGAL 1715
on July 24, 2014 by guesthttp://jcm
.asm.org/
Dow
nloaded from