characterization of cpp32like protease activity following apoptotic challenge in sh-sy5y...
TRANSCRIPT

IOU/flu1 of Nc‘uro«heinisIr~
Lippincott—Raven Publishers, Philadelphia© 1997 International Society for Neurochemistry
Characterization of CPP32-Like Protease Activity FollowingApoptotic Challenge in SH-SY5Y Neuroblastoma Cells
*tRand Posmantur, tKim McGinnis, tRavi Nadimpalli, *Rjchard B. Gilbertsen,
and lHKevin K. W. Wang
Departments of *Immunopathologv and ~Neuroscience Therapeutics, Parke-Davis Pharmaceutical Research,Warner-Lanibert Company, Ann Arbor, Michigan, U.S.A
Abstract: We characterized the activation of interleukin-1/3-converting enzyme (ICE)-like proteases(caspases) inhuman neuroblastoma cells (SH-SY5Y) following chal-lenge with staurosporine, an established agent known toinduce apoptosis. Time course analyses of lactate dehy-drogenase release detected a significant increase in celldeath as early as 6 h that continued at least until 24 hfol lowing staurosporine treatment. Western blot analysesusing anti-poly(ADP-ribose) polymerase (anti-PARP) andanti-CPP32 antibodies revealed proteolytic processing ofCPP32 (an ICE homologue) as well as fragmentation ofPAAP as early as 3 h following staurosporine challenge.Furthermore, the hydrolysis of the CPP32 substrate ace-tyl-DEVD-7-amido-4-methylcoumarin was detected asearly as 3 h and became maximal at 6 h after stauro-sporine challenge, suggesting a delayed and sustainedperiod of CPP32-like activation. In addition, we used thefirst immunohistochemical examination of CPP32 andPARP in cells following an apoptotic challenge. The local-ization of CPP32 in untreated SH-SY5Y cells was exclu-sively restricted to the cytoplasm. Following stauro-sporine challenge there was a condensing of CPP32 im-munofluorescence from the cytoplasm to a regionadjacent to the plasma membrane. In contrast, PAAPimmunofluorescence was evenly distributed in the nu-cleus in untreated SH-SY5Y cells and on staurosporinechallenge was found to be associated with condensedchromatin. lt is important that a pan ICE inhibitor [carbo-benzoxy-Asp-CH
2OC(O)-2,6-dichlorobenzene] was ableto attenuate lactate dehydrogenase release and PARPand CPP32 cleavage and altered immunohistochemicalstaining patterns for PARP and CPP32 following stauro-sporine challenge. Key Words: Interleukin-1/3-convert-ing enzyme-like proteases—Human SH-SY5Y neu-roblastoma cells—Apoptosis—CPP32—Poly(ADP-ribose) polymerase.J. Neurochem. 68, 2328—2337 (1997).
Apoptosis (programmed cell death) is a physiologi-cal process involved in the elimination of excess neu-rons during nervous system development and matura-tion (Homma et al., 1994; Zamenhof and Guthrie,1995; Yaginuma et al., 1996). Apoptotic cell deathhas
been associated with several hallmark morphologicalchanges, including condensation of the nucleus, frag-mentation of chromatin, membrane blebbing, and theformation of apoptotic bodies (Kerr and Harmon,1991; Gavrieli et al., 1992; Brown et al., 1993; Martinet al., 1995). Recently, the involvement of unregulatedapoptosis has been implicated in several neurologicaldisorders, including cerebral ischemia (Linnik et al.,1993; MacManus et al., 1993; Hill et al., 1995; Kohet al., 1995), Huntington‘s disease (Portera-Cailliauet al., 1995; Zeitlin et al., 1995; Goldberg et al., 1996),Alzheimer‘s disease (Nixon et al., 1994; Satou et al.,1995; Gschwind et al., 1996; Vito et al.. 1996; Yamat-suji et al., 1996), amyotrophic lateral sclerosis (Troostet al., 1995; Marx et al., 1996), and Parkinson‘s dis-ease (Ziv et al., 1994; Walkinshaw and Waters, 1995).
A new class of proteases has been implicated inapoptotic cell death belonging to the interleukin-l/3-converting enzyme (ICE)/CED-3 or caspase family ofcysteine proteases. These consist of at least 10 differenthomologues, including ICE, CPP32, lch-l (Nedd2),lch-2, lCE~111, Mch-2, Mch-3, Mch-4, Mch-5, andMch-6 (for reviews, see Alnemri et al., 1996; Fraserand Evan, 1996; Vaux and Strasser, 1996). The overallcommon feature of this family of proteases is the con-servation of the active site QACXG (where X is R,Q, or G) pentapeptide to induce apoptosis and theabsolute requirement for proteolytic cleavage of proen-zymes and substrates at a conserved aspartate residue
Received January 8, 1997; revised manuscript received February12, 1997; accepted February 13, 1997.
Address correspondence and reprint requests to Dr. R. Posmanturat Department of Neuroscience Therapeutics. Parke-Davis Pharma-ceutical Research, Warner-Lambert Company. 2800 Plymouth Road,Ann Arbor, Ml 48105. U.S.A.
Abbreviations used: Ac-DEVD-MCA, acetyl-DEVD-7-amido-4-methylcoumarin; Ac-YVAD-MCA. acctyl-YVAD-7-amido-4-meth-ylcoumarin; ICE, interleukin- I ß-converting enzyme: LDH, lactatedehydrogenase; PAGE, polyacrylamide gel electrophoresis; PARP.poly( ADP-ribose) polymerase; PBS, phosphate-buffered saline;SDS, sodium dodecyl sulfate: Z-D-DCB, carbobenzoxy-Asp-CH2OC (0) -2.6-dichlorobenzene.
2328

CPP32-LIKE ACTIVITY IN SH-SY5Y NEUROBLASTOMA 2329
(Cerretti et al., 1992; Thornberry et al., 1992). Paststudies have demonstrated that overexpression of ICEin fibroblasts can lead to apoptosis (Miura et al.,1993). In addition, inhibitors of ICE-like homologuessuch as the viral protein Crm-A and peptide inhibitorscan block Fas-mediated T cell apoptosis (Enari et al.,1995, 1996; Tewari and Dixit, 1995; Tewari et al.,1995). However, other studies using ICE knockoutmice have shown that ICE itself participates but is notabsolutely required for mammalian apoptosis (Kuidaet aI., 1995; Li et al., 1995). These studies suggestthe involvement of other caspases or another class ofproteases in mammalian apoptosis (Martin and Green,1995).
Among the caspase homologues, CPP32 (caspase-3) has been deemed an attractive candidate as a puta-tive mediator of mammalian apoptosis. Following theinduction of apoptosis CPP32 can become maximallyactivated when the proenzyrne is cleaved from a 32-kDa isoform into active 17- and 13-kDa fragments(Darmon et al., 1994; Nicholson et al., 1995; Orth etal., 1996). Activated CPP32 can proteolytically cleaveseveral important intracellular, membrane, and nuclearproteins that when degraded can facilitate cell deathduring apoptosis (Fernandes-Alnemri et al., 1995;Nicholson et al., 1995; Tewari et al., 1995). Preferredsubstrates of CPP32 include poly(ADP-ribose) poly-merase (PARP), an enzyme involved in DNA repair(Lazebnik et al., 1994; Nicholson et al., 1995); a-spectrin, an integral membrane skeletal protein (Martinand Green, 1995; Martin et al., 1995; Nath et al., 1996;Vanags et al., 1996); the 70-kDa protein componentof the UI -ribonucleoprotein; and the catalytic subunitof the DNA-dependent protein kinase (Casciola-Rosenet al., 1996). In addition, Goldberg et al. (1996) havedemonstrated that the HD gene product, huntingtin, iscleaved by CPP32. Furthermore, caspases have beenidentified as important mediators in other models ofneuronal apoptosis (Nath et al., 1996). Most recently,CPP32 knockout mice were found to have reducedneuronal death during brain development (Kuida et al.,1996). Consequently, an understanding of caspases inneuronal apoptotic cell death can provide new thera-peutic insights in disease processes where these prote-ases have been implicated.
In this study, we explored the role of two caspaseproteases, ICE (caspase-l) and CPP32 (caspase-3)lollowing staurospori ne-induced apoptosis in SH-SY5Y cells. We detected multiple lines of evidencefor the role of CPP32-like activity in staurosporine-induced apoptosis. Specifically, neurons challengedwith staurosporine accumulate the CPP32-like-derived55-kDa PARP cleavage product and the I 7-kDa CPP32breakdown product and demonstrate increased hydro-lysis of the fluorometric CPP32 substrate acetyl-DEVD-7-amido-4-methylcoumarin (Ac-DEVD-MCA). Moreover, a pan caspase inhibitor [Z-D-DCB;carbobenzoxy-Asp-CH2OC(O) -2,6-dichlorobenzene I(Dolle et al., 1994) was able to block all indicators of
CPP32-like activation, immunohistochemical alter-ations, and cell death following staurosporine chaI-lenge in SH-SY5Y cells.
MATERIALS AND METHODS
ChemicalsHoechst 33258 DNA stain (bisbenzimide) was obtained
from Sigma Chemical Co. (St. Louis, MO, U.S.A.). Stauro-sporine was obtained from Calbiochem (La Jolla, CA,U.S.A.). The pan caspase inhibitor Z-D-DCB was synthe-sized at Warner-Lambert Co. (Dolle et al., 1994). The fluo-rometric peptide substrates acetyl-YVAD-7-amido-4-meth-ylcoumarin (Ac-YVAD-MCA) and Ac-DEVD-MCA werepurchased from Bachem Bioscience (King of Prussia, PA,U.S.A.). Lactate dehydrogenase (LDH) Cytox kit 96 wasobtained from Promega (Madison, WI, U.S.A.). The DC(detergent-compatible) protein assay was purchased fromBio-Rad (Hercules, CA, U.S.A.). Phosphate-buffered saline(PBS) was ordered from Celox (Hopkins, MN, U.S.A.).Fluorescent secondary anti-mouse EgG antibodiesconjugatedwith fluorescein isothiocyanate was ordered from VectorLaboratories (Burlingame, CA, U.S.A.).
Staurosporine challenge of SH-SY5Yneuroblastoma cells
Human neuroblastorna SH-SY5Y cells (provided by SteveFisher as a generous gift; University of Michigan) werecultured on 12-well plates until confluency (~-.2X l0~‘perwell). Cells were periodically fed with Dulbecco‘s modifiedEagle‘s medium supplemented with 10% fetal bovine serum,100 lU/mI penicillin, 100 pg/mI streptomycin, and 2.5 pg/ml amphotericin B (Fungizone). All cells were washed threetimes with serum-free Dulbecco‘s modified Eagle‘s mediumbefore experimental protocols. Some treatment groups werepreincubated with Z-D-DCB (50 pM) for I h before stauro-sporine challenge (0.5 pM).
LDH quantificationApoptotic cell death was assessed in SH-SY5Y cells by
measuring the release of the cytosolic enzyme LDH into themedium (25 p1 of each sample) due to secondary necrosis.Methods were identical to those described in recent studies(Nath et al., 1996).
Hoechst staining of apoptotic nucleiCells were rinsed twice with phosphate buffer solution
and incubated with I pg/mi DNA dye Hoechst 33258 stainin PBS for 5 min at room temperature. All photomicrographswere taken using a Lietz microscopic imaging system at amagnification of 320><.
Protein extraction, sodium dodecyl sulfate(SDS) —polyacrylamide gel electrophoresis(PAGE), and western blot methodology
Following treatment with staurosporine, cells werewashed twice with Tris-buffered saline containing EDTA[20 mM Tris-HC1 (pH 7.4), 155 mM NaCI, and 1 mMEDTA1. SDS protein extraction of cell lysates was per-formed in the presence of protease inhibitors as previouslydescribed (Nath et al., 1996). The extraction procedure wascompleted by precipitating the proteins with trichioroaceticacid, and resolubilization of the pellet was achieved with 3M Tris hase. Protein content determination of samples wasperformed using a modified Folio phenol reagent assay and
J. Neuro,hein., Vol. 68, No. 6, 1997

2330 R. POSMANTUR ET AL.
bovine serum albumin standards. Protein samples containing5—15 pg of protein were eleetrophoresed on 4—20% acryl-amide gradient gels using a Tris glycine running buffer.Following separation of proteins by SDS-PAGE, proteinswere transferred onto a PVDF membrane using a Tris glycinebuffer (48 mM Tris base, 39 mM glycine, and 10% metha-nol, pH 9.2) and semidry electrotransferring unit (Bio-Rad)at 20 mV for 2 h. All lanes contained identical amounts ofprotein. To ensure consistency of loading, gels were rou-tinely stained with Coomassie Brilliant Blue dye. Followingtransfer of the proteins to PVDF membrane paper, nonspe-cific sites were blocked with an incubation of 5% nonfat drymilk in 20 mM Tris HCI, 0.05% Tween 20, and 0.02% NaN~at 4°C.Incubation of the primary antibody was performedwith either anti-PARP (Santa Cruz Biotechnologies. SantaCruz, CA, U.S.A.) or anti-CPP32 (clone 19: TransductionLaboratories, Lexington, KY, U.S.A.) monoclonal antibod-ies. Visualization of proteins was performed with bio-tinylated secondary antibodies and streptavidin-alkalinephosphatase (Amersham, Heightsville, IL, U.S.A.) using5-bromo-4-chloro-3-indolyl phosphate/nitro blue tetrazo-hum (Sigma) as the active chromogen.
Triton protein extraction for assaying ICE andCPP32-like activity
On completion of the experimental protocol, nontrypsin-ized SH-SY5Y cells were scraped from three wells of a 12-well plate, collected in 15-ml centrifuge tubes, and washedtwice with 5 ml of Tris-buffered saline containing EDTAbuffer by centrifugation for 5 min at 4°C.Cell pellets wereresuspended and lysed in a buffer containing 20 mM Tris-HCI (pH 7.4), 150 mM NaC1, 1 mM dithiothreitol, 5 mMEDTA, 5 mM EGTA, and 1% Triton X-100 for 60 min at4°C. Cell lysates were then recovered after centrifugationand stored in 50% glycerol at —70°C. All cell lysates wereassayed for protein concentration with a modified Folin phe-nol reagent assay. ICE and CPP32-Iike activities were as-sayed for proteolytic activity by using the fluorometric pep-tide substrates Ac-YVAD-MCA and Ae-DEVD-MCA, re-spectively. Cell lysates were added to a buffer (200 pI)containing 100 pMpeptide substrate, 100 mM HEPES, 10%glycerol, I Ifl M EDTA, 10 mM dithiothreitol, and the generalcaspase inhibitor Z-D-DCB at 10pM. Fluorescence (opticaldensity at an excitation wavelength of 380 ± 15 nni and anemission wavelength of 460 ±15 nm) was measured every15 min up to 60 min with a Millipore Cytoflor 2300 fluores-cence plate reader.
Immunofluorescence studiestmmunofluorescenee studies used SH-SY5Y cells cultured
on German glass coverslips (Fisher, Springfield, NJ, U.S.A.)at medium density. At the desired time point cells from allexperimental treatment groups were fixed in 4% paraformal-dehyde for 2 h at room temperature. Following fixation cellswere washed twice with PBS (136 mM NaCI, 81 mM KCI,1.6 mM Na,HPO4, and 14 mM KH2PO4, pH 7.4). Cultureswere first incubated in 5% nonfat dry milk at 4°Cfor 2 h.Primary antibodies (anti-CPP32 and anti-PARP) in blockersolution {Tween-PBS; 10 mM sodium phosphate (pH 7.5),0.9% NaCI, 0.1% Tween 20, and 5% nonfat dry milki wereincubated for 3 h at 25°C.The cultures were then washedthree times in blocking solution at 10-min intervals. Second-ary antibodies (anti-mouse 1gO) linked to a specific fluoro-phore, fluorescein isothiocyanate, were applied for 2 h. Thecoverslips were then washed three times in PBS, mounted
FIG. 1. Time course of CPP32 activation and cell death followingstaurosporine challenge in SH-SY5Y neuroblastoma cells.CPP32 (A) and PARP (B) immunoreactivity in SH-SY5Y cell ly-sates at O (lane 1), 1 (lane 2), 3 (lane 3), 6 (lane 4), and 24 h(lane 5) following staurosporine challenge was examined bySDS-PAGE and western blotting techniques. Proteins were visu-alized on PVDF transfers using anti-CPP32 and anti-PARP anti-bodies. Staurosporine-treated SH-SY5Y cells contained a 17-kDa CPP32 breakdown product (A) and an 85-kDa PARP break-down product (B) suggestive of CPP32-like proteolysis. C: LDHrelease sampled from control (O) and staurosporine-treated (•)SH-SY5Y lysates at 0, 1, 3, 6, 12, and 24 h following stauro-sporine challenge. Data are mean ±SEM (bars) values (n = 3).
on slides containing Elvanol (anti-fade agent; Du Pont, Wil-mington, DE, U.S.A.), and allowed to dry. Slides were storedin the dark before immunofluorescence microscopic exami-nation. Control cultures without primary antibodies did notstain.
RESULTS
Staurosporine challenge in SH-SY5Yneuroblastoma cells produces proteolytic cleavageproducts suggestive of CPP32-like-mediatedproteolysis
We used SDS-PAGE and western blotting method-ology on protein extracts from SH-SY5Y cells at 0, 1,3, 6, 16, and 24 h following staurosporine challengeto establish the temporal profile for CPP32-like activa-tion. Western blots were probed with antibodiesagainst the caspase CPP32 and the nuclear enzymePARP to detect the presence of lower-molecular-massbreakdown products suggestive of caspase proteolysis.Reduction of the parent 32-kDa CPP32 subunit oc-curred as early as 3 h (Fig. lA). However, the active
J. Neurochern., Vol. 68, No. 6, 1997
CPP32-LIKE ACTIVITY IN SI-I-SY5Y NEUROBLASTOMA 2331
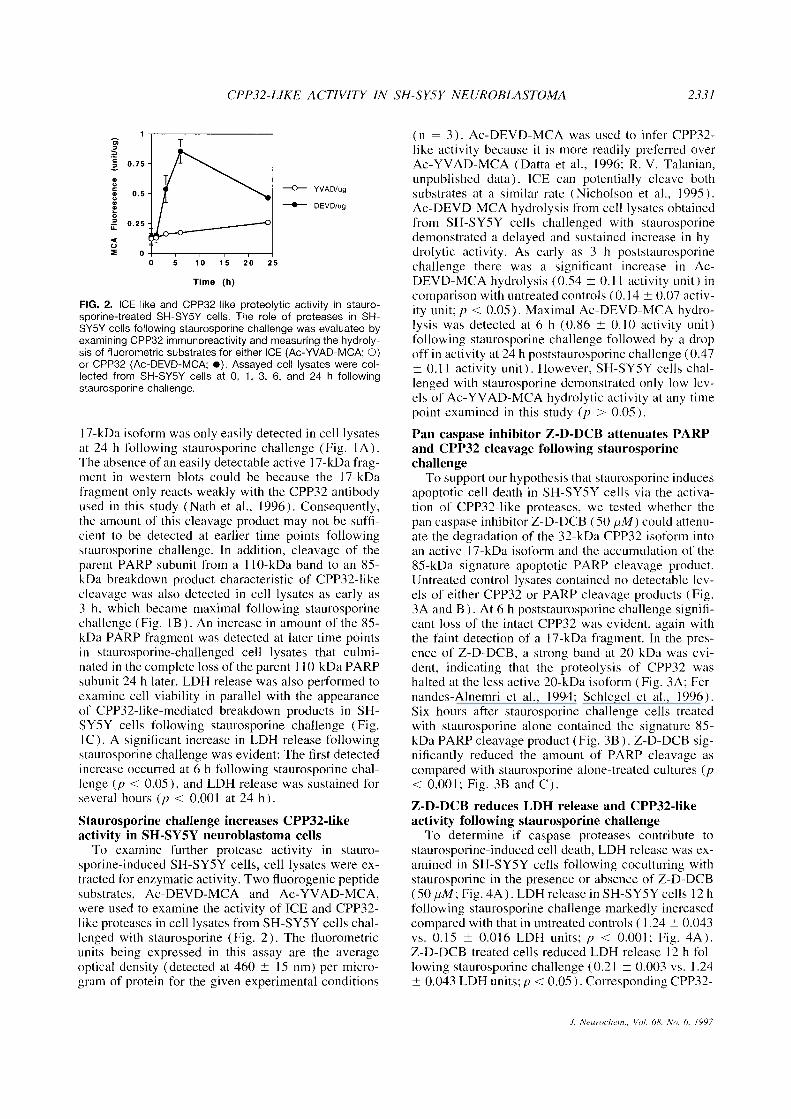
FIG. 2. ICE-like and CPP32-like proteolytic activity in stauro-sporine-treated SH-SY5Y cells. The role of proteases in SH-SY5Y cells following staurosporine challenge was evaluated byexamining CPP32 immunoreactivity and measuring the hydroly-sis of fluorometric substrates for either ICE (Ac-YVAD-MCA; O)or CPP32 (Ac-DEVD-MCA; •). Assayed cell lysates were col-lected from SH-SY5Y cells at O, 1, 3, 6, and 24 h followingstaurosporine challenge.
I 7-kDa isoform was only easily detected in cell lysatesat 24 h following staurosporine challenge (Fig. IA).The absence of an easily detectable active 17-kDa frag-ment in western blots could be because the 17-kDafragment only reacts weakly with the CPP32 antibodyused in this study (Nath et al., 1996). Consequently,the amount of this cleavage product may not be suffi-cient to be detected at earlier time points followingstaurosporine challenge. In addition, cleavage of theparent PARP subunit from a 1 l0-kDa band to an 85-kDa breakdown product characteristic of CPP32-likecleavage was also detected in cell lysates as early as3 h, which became maximal following staurosporinechallenge (Fig. lB). An increase in amount of the 85-kDa PARP fragment was detected at later time pointsin staurosporine-challenged cell lysates that culmi-nated in the complete loss of the parent I l0-kDa PARPsubunit 24 h later. LDH release was also performed toexamine cell viability in parallel with the appearanceof CPP32-like-mediated breakdown products in SH-SYSY cells following staurosporine challenge (Fig.IC). A significant increase in LDH release followingstaurosporine challenge was evident: The first detectedincrease occurred at 6 h following staurosporine chal-lenge (p < 0.05), and LDH release was sustained forseveral hours (p < 0.001 at 24 h).
Staurosporine challenge increases CPP32-likeactivity in SH-SY5Y neuroblastoma cells
To examine further protease activity in stauro-sporine-induced SH-SY5Y cells, cell lysates were ex-tracted for enzymatic activity. Two fluorogenic peptidesubstrates, Ac-DEVD-MCA and Ac-YVAD-MCA,were used to examine the activity of ICE and CPP32-like proteases in cell lysates from SH-SY5Y cells chal-lenged with staurosporine (Fig. 2). The fluorometricunits being expressed in this assay are the averageoptical density (detected at 460 ± 15 nm) per micro-gram of protein for the given experimental conditions
(n = 3). Ac-DEVD-MCA was used to infer CPP32-like activity because it is more readily preferred overAc-YVAD-MCA (Datta et al., 1996; R. V. Talanian,unpublished data). ICE can potentially cleave bothsubstrates at a similar rate (Nicholson et aI., 1995).Ac-DEVD-MCA hydrolysis from cell lysates obtainedfrom SH-SY5Y cells challenged with staurosporinedemonstrated a delayed and sustained increase in hy-drolytic activity. As early as 3 h poststaurosporinechallenge there was a significant increase in Ac-DEVD-MCA hydrolysis (0.54 ±0.11 activity unit) incomparison with untreated controls (0.14 ±0.07 activ-ity unit; p < 0.05). Maximal Ac-DEVD-MCA hydro-lysis was detected at 6 h (0.86 ±0.10 activity unit)following staurosporine challenge followed by a dropoff in activity at 24h poststaurosporine challenge (0.47±0.11 activity unit). However, SH-SY5Y cells chal-lenged with staurosporine demonstrated only low lev-els of Ac-YVAD-MCA hydrolytic activity at any timepoint examined in this study (p > 0.05).
Pan caspase inhibitor Z-D-DCB attenuates PARPand CPP32 cleavage following staurosporinechallenge
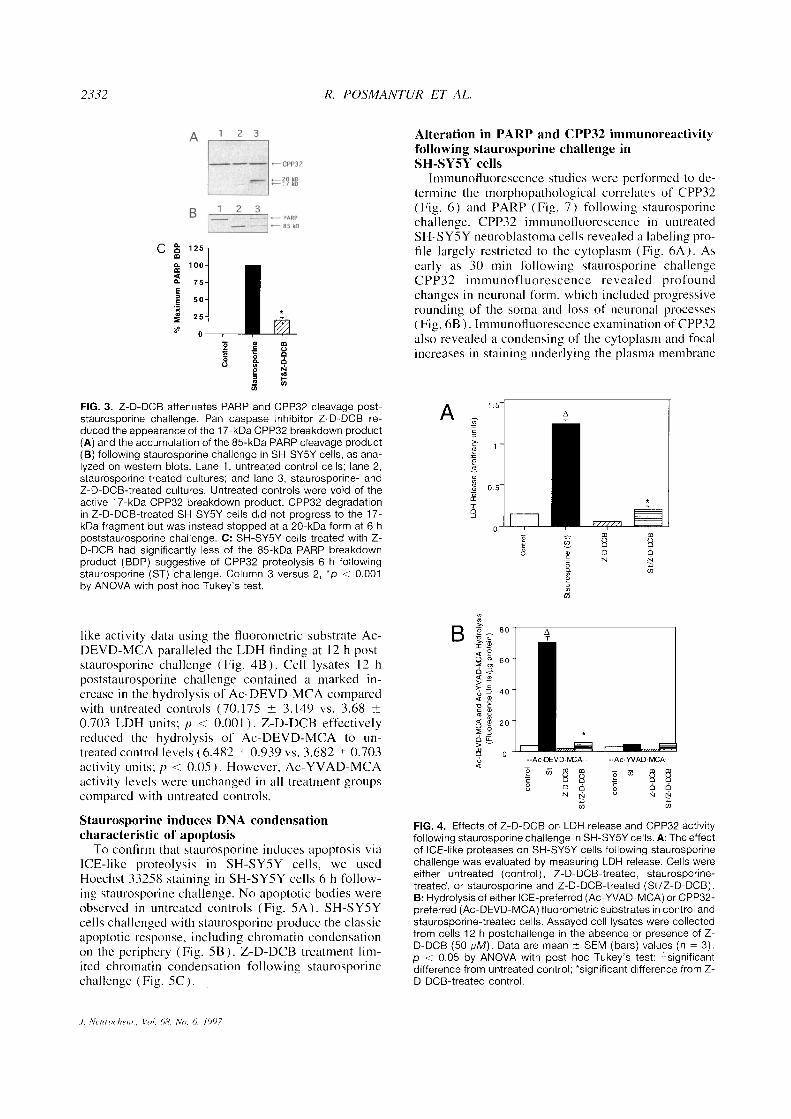
To support our hypothesis that staurosporine inducesapoptotic cell death in SH-SY5Y cells via the activa-tion of CPP32-like proteases, we tested whether thepan caspase inhibitor Z-D-DCB (50 pM) could attenu-ate the degradation of the 32-kDa CPP32 isoform intoan active I 7-kDa isoform and the accumulation of the85-kDa signature apoptotic PARP cleavage product.Untreated control lysates contained no detectable lev-els of either CPP32 or PARP cleavage products (Fig.3A and B). At 6 h poststaurosporine challenge signifi-cant loss of the intact CPP32 was evident, again withthe faint detection of a l7-kDa fragment. In the pres-ence of Z-D-DCB, a strong band at 20 kDa was evi-dent, indicating that the proteolysis of CPP32 washalted at the less active 20-kDa isoform (Fig. 3A; Fer-nandes-Alnemri et al., 1994; Schlegel et al., 1996).Six hours after staurosporine challenge cells treatedwith staurosporine alone contained the signature 85-kDa PARP cleavage product (Fig. 3B). Z-D-DCB sig-nificantly reduced the amount of PARP cleavage ascompared with staurosporine alone-treated cultures (p< 0.001; Fig. 3B and C).
Z-D-DCB reduces LDH release and CPP32-likeactivity following staurosporine challenge
To determine if caspase proteases contribute tostaurosporine-induced cell death, LDH release was ex-amined in SH-SY5Y cells following coculturing withstaurosporine in the presence or absence of Z-D-DCB(50 pM; Fig. 4A). LDH release in SH-SY5Y cells 12hfollowing staurosporine challenge markedly increasedcompared with that in untreated controls (1.24 ±0.043vs. 0.15 ±0.016 LDH units; p < 0.001; Fig. 4A).Z-D-DCB-treated cells reduced LDH release 12 h fol-lowing staurosporine challenge (0.21 ±0.003 vs. 1.24±0.043 LDH units; p < 0.05). Corresponding CPP32-
J. Neurochern., Vol. 68, No. 6, 1997
2332 R. POSIVIANTUR ET AL.
FIG. 3. Z-D-DCB attenuates PARR and CPP32 cleavage post-staurosporine challenge. Pan caspase inhibitor Z-D-DCB re-duced the appearance ofthe 1 7-kDa CPP32 breakdown product(A) and the accumulation of the 85-kDa PARP cleavage product(B) following staurosporine challenge in SH-SY5Y cells, as ana-lyzed on western blots. Lane 1, untreated control cells; lane 2,staurosporine-treated cultures; and lane 3, staurosporine- andZ-D-DCB-treated cultures. Untreated controls were void of theactive 1 7-kDa CPP32 breakdown product. CPP32 degradationin Z-D-DCB-treated SH-SY5Y cells did not progress to the 17-kDa fragment but was instead stopped at a 20-kDa form at 6 hpoststaurosporine challenge. C: SH-SY5Y cells treated with Z-D-DCB had significantly less of the 85-kDa PARR breakdownproduct (BDP) suggestive of CPP32 proteolysis 6 h followingstaurosporine (ST) challenge. Column 3 versus 2, °p< 0.001by ANOVA with post hoc Tukey‘s test.
like activity data using the fluorometric substrate Ac-DEVD-MCA paralleled the LDH finding at 12 h post-staurosporine challenge (Fig. 4B). Cell lysates 12 hpoststaurosporine challenge contained a marked in-crease in the hydrolysis of Ac-DEVD-MCA comparedwith untreated controls (70.175 ± 3.149 vs. 3.68 ±0.703 LDH units; p < 0.001). Z-D-DCB effectivelyreduced the hydrolysis of Ac-DEVD-MCA to un-treated control levels (6.482 ±0.939 vs. 3.682 ±0.703activity units; p < 0.05). However, Ac-YVAD-MCAactivity levels were unchanged in all treatment groupscompared with untreated control s.
Alteration in PARP and CPP32 immunoreactivityfollowing staurosporine challenge inSH-SY5Y cells
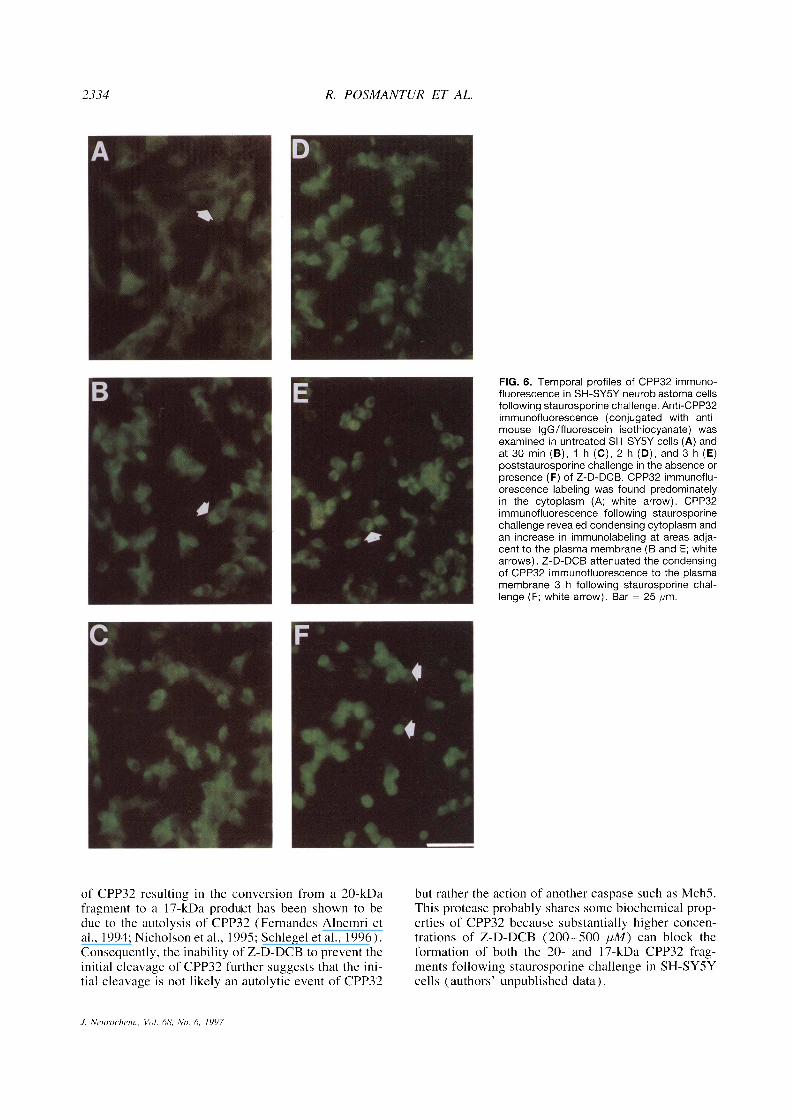
Immunofluorescence studies were performed to de-termine the morphopathological correlates of CPP32(Fig. 6) and PARP (Fig. 7) following staurosporinechallenge. CPP32 immunofluorescence in untreatedSH-SY5Y neuroblastoma cells revealed a labeling pro-file largely restricted to the cytoplasm (Fig. 6A). Asearly as 30 min following staurosporine challengeCPP32 immunofluorescence revealed profoundchanges in neuronal form, which included progressiverounding of the soma and loss of neuronal processes(Fig. 6B). Immunofluorescence examination of CPP32also revealed a condensing of the cytoplasm and focalincreases in staining underlying the plasma membrane
Staurosporine induces DNA condensationcharacteristic of apoptosis
To confirm that staurosporine induces apoptosis viaICE-like proteolysis in SH-SYSY cells, we usedHoechst 33258 staining in SH-SY5Y cells 6 h follow-ing staurosporine challenge. No apoptotic bodies wereobserved in untreated controls (Fig. SA). SH-SY5Ycells challenged with staurosporine produce the classicapoptotic response, including chromatin condensationon the periphery (Fig. SB). Z-D-DCB treatment lim-ited chromatin condensation following staurosporinechallenge (Fig. SC).
FIG. 4. Effects of Z-D-DCB on LDH release and CPP32 activityfollowing staurosporine challenge in SH-SY5Y cells. A: The effectof ICE-like proteases on SH-SY5Y cells following staurosporinechallenge was evaluated by measuring LDH release. Cells wereeither untreated (control), Z-D-DCB-treated, staurosporine-treated, or staurosporine and Z-D-DCB-treated (St/Z-D-DCB).B: Hydrolysisof either ICE-preferred (Ac-YVAD-MCA) or CPP32-preferred (Ac-DEVD-MCA) fluorometric substrates in control andstaurosporine-treated cells. Assayed cell lysates were collectedfrom cells 12 h postchallenge in the absence or presence of Z-D-DCB (50 pM). Data are mean ±SEM (bars) values (n = 3).p < 0.05 by ANOVA with post hoc Tukey‘s test: -significantdifference from untreated control; *significant difference from Z-D-DCB-treated control.
J. Neurorhern., Vol. 68, No. 6. 1997

CPP32-LIKE ACTIVITY IN SH-SY5Y iVEUROBLASTOMA 2333
FIG. 5. Z-D-DCB prevents hallmark features of apoptotic cell death detected by Hoechst staining following staurosporine challenge.Hoechst-stained SH-SY5Y cells were examined in untreated controls (A) and 6 h following staurosporine challenge in the absence(B) or presence (C) of Z-D-DCB (50 pM). Untreated controls were void of any evidence of apoptotic nuclei (A). Photomicrographsof cells challenged with staurosporine revealed large numbers of condensed and apoptotic nuclei (B). SH-SY5Y cells challenged withstaurosporine in the presence of Z-D-DCB contained fewer apoptotic nuclei (C). Bar = 25 pm.
as early as I h (Fig. 6C). A continued progression ofcondensing cytoplasm and focal increases in stainingunderlying plasma membrane was also observed at 2h (Fig. 6D) and became maximal 3 h (Fig. 6E) post-staurosporine challenge. Z-D-DCB treatment was ableto attenuate partially changes associated with stauro-sporine-induced apoptosis (Fig. 6F).
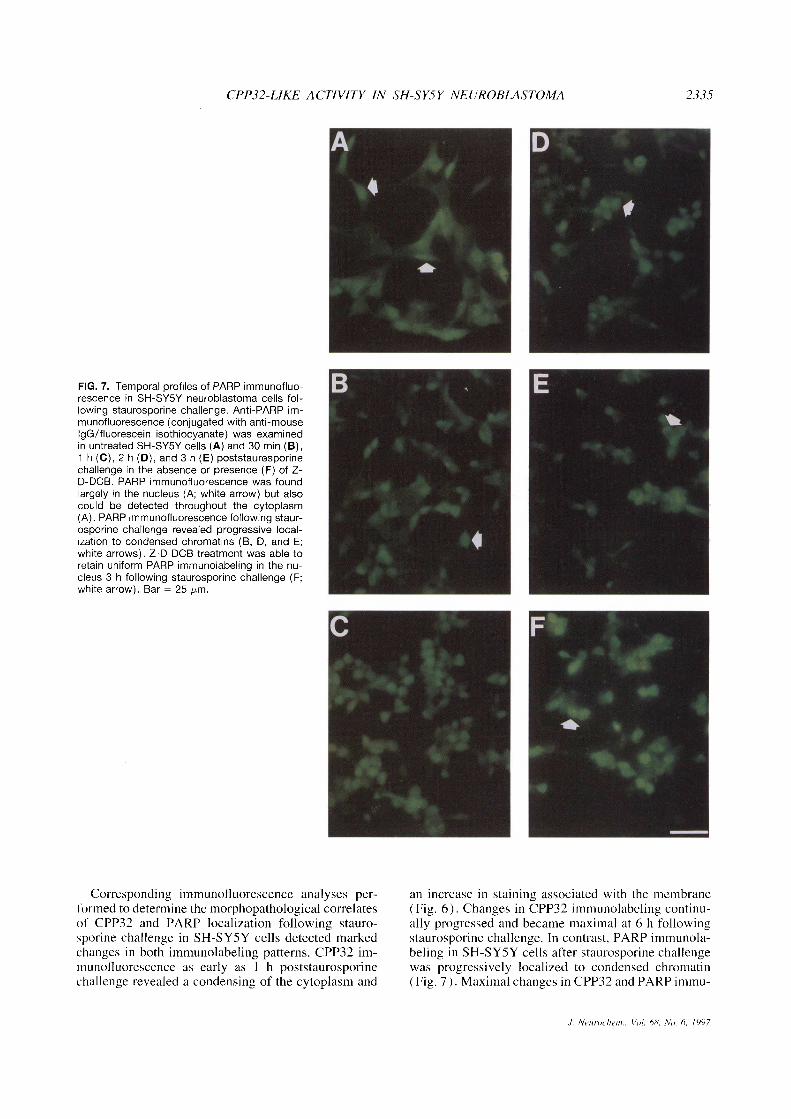
The distribution of PARP immunolabeling in un-treated SH-SYSY controls was predominately local-ized in the nucleus but also detected diffusely through-out the cytoplasm (Fig. 7A). Alterations in PARP im-munofluorescence were detected as early as 30 min(Fig. 7B) poststaurosporine challenge in SH-SYSYcells, including the loss of neuronal processes and cel-lular retraction. PARP immunofluorescence in SH-SY5Y cells at 1 (Fig. 7C), 2 (Fig. 7D), and 3 h (Fig.7E) poststaurosporine challenge revealed progressivelocalization to condensed chromatins. It is importantthat Z-D-DCB was able to preserve uniform PARPimmunolabeling in the nucleus 3 h poststaurosporinechallenge in SH-SY5Y cells but was not able to pre-serve completely neuronal form as observed in un-treated controls (Fig. 7F).
DISCUSSION
Staurosporine, a general protein kinase inhibitor, hasbeen reported to induce apoptosis in many cell types(Jacobson et al., 1996; Weil et al., 1996) includingneurons (Koh et al., 1995; Wiesner and Dawson, 1996).In our current study we demonstrated that staurosporine-challenged SH-SYSY neuroblastoma cells undergo apo-ptotic cell death via caspase protease activation. Severalmethods were established for detecting CPP32-like ac-tivity, including PARP cleavage (Lazebnik et al., 1994;Nicholson et al., 1995), the appearance of active CPP32
breakdown products (Cerretti et al., 1992; Darmon etal., 1994; Fernandes-Alnemri et al., 1994; Nicholson etal., 1995; Schlegel et al., 1996), and the cleavage ofthe fluorometric CPP32 substrate Ac-DEVD-MCA(Nicholson et al., 1995). In this study, using the above-mentioned techniques, we demonstrated CPP32-like ac-tivation in neuronal apoptosis (Figs. I and 2). However,ICE activity was not detected in staurosporine-treatedSH-SYSY cells at any time point examined in this study(Fig. 2). Furthermore, the time course of CPP32-likeactivity as inferred by both the detection of specificbreakdown product patterns and fluorometric activityprecedes cell death [as measured by LDH release (Fig.
I vs. Fig. 2)1. These findings are of importancebecausethey identify a candidate protease in neuronal apoptosisthat degrades several important proteins necessary forcell survival and provide an attractive target for thera-peutic intervention.
The pan caspase inhibitor Z-D-DCB attenuated allindicators of CPP32-like activity, including Ac-DEVD-MCA hydrolytic activity (Fig. 4B), the accu-mulation of the 85-kDa PARP cleavage product, andthe formation of the active 17-kDa CPP32 form (Fig.3), as well as a reduction in cell death as measuredby LDH release up to 12 h following staurosporinechallenge (Fig. 4A). Moreover, the accumulation of aless active 20-kDa CPP32 form (Fernandes-Alnemriet al., 1994; Schlegel et al., 1996) in Z-D-DCB-treatedcells poststaurosporine challenge is consistent with theliterature on the order of proteolytic processing ofCPP32 in apoptotic paradigms. The initial cleavage ofCPP32 from a 32-kDa protein to a lower-molecular-mass 20-kDa fragment has been shown to be producedby Mch4 (Fernandes-Alnemri et al., 1996), MchS(Srinivasula et al., 1996), or potentially other relatedproteases (Darmon et al., 1994). The second cleavage
J. Neuro/hen,., V//I. 68. No. 6. 1997
2334 R. POSMANTUR ET AL.
FIG. 6. Temporal profiles of CPP32 immuno-fluorescence in SH-SY5Y neuroblastoma cellsfollowing staurosporine challenge. Anti-CPP32immunofluorescence (conjugated with anti-mouse lgG/fluorescein isothiocyanate) wasexamined in untreated SH-SY5Y cells (A) andat 30 min (B), 1 h (C), 2 h (D), and 3 h (E)poststaurosporine challenge in the absence orpresence (F) of Z-D-DCB. CPP32 immunoflu-orescence labeling was found predominatelyin the cytoplasm (A; white arrow). CPP32immunofluorescence following staurosporinechallenge revealed condensing cytoplasm andan increase in immunolabeling at areas adja-cent to the plasma membrane (B and E; whitearrows). Z-D-DCB attenuated the condensingof CPP32 immunofluorescence to the plasmamembrane 3 h following staurosporine chal-lenge (F; white arrow). Bar = 25 pm.
of CPP32 resulting in the conversion from a 20-kDafragment to a 17-kDa produ~cthas been shown to bedue to the autolysis of CPP32 (Fernandes-Alnemri etal., 1994; Nicholson et al., 1995; Schlegel et al., 1996).Consequently, the inability of Z-D-DCB to prevent theinitial cleavage of CPP32 further suggests that the ini-tial cleavage is not likely an autolytic event of CPP32
but rather the action of another caspase such as MchS.This protease probably shares some biochemical prop-erties of CPP32 because substantially higher concen-trations of Z-D-DCB (200—500 pM) can block theformation of both the 20- and 17-kDa CPP32 frag-ments following staurosporine challenge in SH-SY5Ycells (authors‘ unpublished data).
J. Neuroc/jem., Vol. 68, No. 6. 1997
CPP32-LIKE ACTIViTY IN SH-SY5Y NEUROBLASTOMA 2335
FIG. 7. Temporal profiles of PARP immunofluo-rescence in SH-SY5Y neuroblastoma cells fol-lowing staurosporine challenge. Anti-PARR im-munofluorescence (conjugated with anti-mouselgG/fluorescein isothiocyanate) was examinedin untreated SH-SY5Y cells (A) and 30 min (B),1 h (C), 2 h (D), and 3 h (E) poststauresporinechallenge in the absence or presence (F) of Z-D-DCB. PARR immunofluorescence was foundlargely in the nucleus (A; white arrow) but alsocould be detected throughout the cytoplasm(A). PARR immunofluorescence following staur-osporine challenge revealed progressive local-ization to condensed chromatins (B, D, and E;white arrows). Z-D-DCB treatment was able toretain uniform PARR immunolabeling in the nu-cleus 3 h following staurosporine challenge (F;white arrow). Bar = 25 pm.
Corresponding immunofluorescence analyses per-formed to determine the morphopathological correlatesof CPP32 and PARP localization following stauro-sporine challenge in SH-SY5Y cells detected markedchanges in both immunolabeling patterns. CPP32 im-munofluorescence as early as I h poststaurosporinechallenge revealed a condensing of the cytoplasm and
an increase in staining associated with the membrane(Fig. 6). Changes in CPP32 immunolabeling continu-ally progressed and became maximal at 6 h followingstaurosporine challenge. In contrast, PARP immunola-beling in SH-SYSY cells after staurosporine challengewas progressively localized to condensed chromatin(Fig. 7). Maximal changes in CPP32 and PARP immu-
J. Neuroehen,., V//I. 68, No. 6, 1997

2336 R. POSMANTUR ET AL.
nolabeling patterns paraileled but preceded the appear-ance of apoptotic features, i.e., membrane blebbing andchromatin condensation, detected by Hoechst staining(Fig. 4). It is important that the pan caspase inhibitorZ-D-DCB was able to reduce CPP32 immunolabelingassociated with the plasma membrane and retain PARPimmunolabeling within defined nucleus. These datacollectively suggest that derangements of CPP32-likesubstrates are delayed and evolutionary in nature andthat the induction of CPP32-like proteases is correlatedwith cell death and hallmark apoptotic features.
In summary, we demonstrated that staurosporine in-duces apoptosis in SH-SYSY neuroblastoma cells viaCPP32-like proteolysis. We produced multiple lines ofevidence using the detection of specific breakdownproducts and enzymatic activity assays to implicateCPP32-like activity rather than ICE-like activity fol-lowing staurosporine challenge in SH-SY5Y cells.Western blot analyses of CPP32 revealed breakdownproducts consistent with past findings suggesting a dis-tinct molecular ordering in the activation of CPP32 incells undergoing apoptosis. Moreover, we were ableto block effectively all indicators of CPP32-like pro-teolytic activity using the pan caspase inhibitor Z-D-DCB. In conclusion, we argue that the biochemicaldata outlined here with the alterations in CPP32 andPARP immunolabeling patterns detected followingstaurosporine challenge suggest that CPP32-like acti-vation is an important and necessary component ofneuronal apoptosis following staurosporine challenge.
REFERENCES
Alnemri E. S., Livingston D. J., Nicholson D. W., Salvesen G.,Thornberry N. A., Wong W. W., and Yuan J. (1996) HumanICE/CED-3 protease nomenclature. Cell 87, 171.
Brown D. G., Sun X. M., and Cohen G. M. (1993) Dexamethasone-induced apoptosis involves cleavage of DNA to large fragmentsprior to internucleosomal fragmentation. J. Biol. Chern. 268,3037— 3039.
Casciola-Rosen L., Nicholson D. W., Chong T., Rowan K. R.,Thornberry N. A., Miller D. K., and Rosen A. (1996) Apopain/CPP32 cleaves proteins that are essential for cellular repair: afundamental principle of apoptotic death. J. Exp. Med. 183,1957— 1964.
Cerretti P. G., Kozlosky C. J., Mosley B., Netson N., Ness K. V.,Greenstreet T. A.. March C. J., Kronheim S. R., Druck T., Can-nizzaro L. A., Huebner K., and Black R. A. (1992) Molecutarcloning of the interleukin-lß converting enzyme. Science 256,97— 101.
Darmon A. J., Ehrman N., Caputo A., Fujinaga J., and BleackleyR. C. (1994) The cytotoxic T cell proteinase granzyme B doesnot activate interleukin-l-beta-converting enzyme. J. Biol.Chem. 269, 32043—32050.
Datta R., Banach D., Kojima H., Talanian R. V., Alnemri E. S.,Wong W. W., and Kufe D. W. (1996) Activation of the CPP32protease in apoptosis induced by I -beta-u-arabinofuranosylcy-tosine. BloodS8, 1936—1943.
Dolle R. E., Hoyer D., Prasad C. V. C., Schmidt S. J., HelaszekC. T., Miller R. E., and Ator M. A. (1994) P
1 aspartate-basedpeptide a-((2,6-dichlorobenzoyt)oxy)methyl ketones as potenttime-dependent inhibitors of interleukin- 1 ß-converting enzyme.J. Med. Chem. 37, 563—564.
Enari M., Hug H., and Nagata S. (1995) Involvement of an tCE-like protease in Fas-mediated apoptosis. Nature 375, 78—81.
Enari M., Hug H., Hayakawa M., Ito F., Nishimura Y., and NagataS. (1996) Different apoptotic pathways mediated by Fas andthe tumor-necrosis-factor receptor. Cytosolic phospholipase A2is not involved in Fan-mediated apoptosis. Enir. J. Biochem.236, 533—538.
Fernandes-Alnemri T., Litwack G., and Alnemri E. (1994) CPP32.a novel human apoptotic protein with homology to Caenorhah-ditis eleganz cell death protein Ced-3 and mammalian l-~con-verting enzyme. J. Biol. CItein. 269, 30761— 30764.
Fernandes-Alnemri T., Takahashi A.. Armstrong R., Krebs J., FritzL., Tomaselli K. J., Wang L., Yu Z., Croce C. M., Salveson G.,Earnshaw W. C., Litwack G., and Alnernri E. 5. (1995) Meh3.a novel human apoptotic cysteine protease highly related toCPP32. Cancer Ren. 55, 6045—6052.
Fernandes-Alnemri T., Armstrong R. C., Krebs J., Srinivasula S. M.,Wang L., Bullrich F., Fritz L. C., Trapani J. A., Tomaselli K. J.,Litwack G., and Alnemri E. S. (1996) In vitro activation ofCPP32 and Mch3 by Mch4, a novel human apoptotic cysteineprotease containing two FADD-like domains. Proc. Natl. Acad.Sci. USA 93, 7464—7469.
Fraser A. and Evan G. (1996) A license to kill. Cell 85, 781 —784.Gavrieli Y., Sherman Y., and Ben-Sasson 5. (1992) Identification
of programmed cell death. Early synthesis and DNA degrada-tion. J. Cell Biol. 119, 493—501.
Goldberg Y. P., Nicholson D. W., Rasper D. M.. Kalehman M. A.,Koide H. B., Graham R. K., Bromm M., Kazemi-Esfarjani P.,Thornberry N. A., Vaillancourt J. P.. and Hayden M. R. (1996)Cleavage of huntingtin by apopain, in a proapoptotic cysteineprotease, is modulated by the polyglutamine tract. Nat. Genet.13, 442—449.
Gschwind M., Martin J. R., Moreau J. L.. and HuberG. (1996) BetaAPP cognitive function vs. beta-amyloid induced cell death.Ann. NYAcad. Sri. 777, 293—296.
Hill I.E., MacManus J. P.. Rasquinha I., and Tuor U. 1. (1995)DNA fragmentation indicative of apoptosis following unilateralcerebral hypoxia—isehemia in the neonatal rat. Brain Ren. 676,398—403.
Homma S., Yaginuma H., and Oppenheim R. W. (1994) Pro-grammed cell death during the earliest stages of spinal corddevelopment in the chick embryo: a possible means of earlyphenotypic selection. J. Comp. Neurol. 345, 377—395.
Jacobson M. D., Weil M., and Rat‘f M. C. (1996) Role of Ced-3/ICE-family in staurosporine induced programmed cell death. J.Cell Biol. 133, 1041—1051.
Kerr J. F. R. and Harmon B. V. (1991) Definition and incidence ofapoptosis: an historical perspective, in Apoptosi.n.‘ The Molecu-lar Basis of Cell Death (Tomeiand L. D. and Cope F. O., eds),p. 5. Cold Spring Harbor Laboratory Press, Cold Spring Harbor,New York.
Koh J. Y., Wie M. B., Sensi S. L., Canzoniero L. M., Demaro L.,Cernansky C., and Choi D. W. (1995) Staurosporine-inducedneuronal apoptosis. Exp. Neurol. 135, 153—159.
Kuida K., Lippke J. A., Ku G., Harding M. W., Livingston D. J., SuM. S., and Flavell R. A. (1995) Altered cytokine export andapoptosis in mice deficient in interleukin-l beta converting en-zyme. Science 267, 2000—2003.
Kuida K., Zheng T., Na S., Kuan C., Yang D., Karasuyama H.,Rakic P., and Flavell A. (1996) Decreased apoptosis in thebrain and premature lethality in CPP32-deficient mice. Nature384, 368—370.
Lazebnik Y. A., Kaufman S. H., Desnoyers S., Poirier G. G., andEarnshaw W. C. (t994) Cleavage of poly (ADP-ribose) poly-merase by a proteinase with properties like ICE. Nature 371,346—347.
Li P., Allen H., Banerjee S., Franklin S., Herzog L., Johnston C.,McDowell J., Paskind M., Rodman L., Salfeld J., Towne E.,Tracey D., Wardwell S., Wei F-Y., Wong W., Kamen R., andSeshadri T. (1995) Mice deficient in IL-l beta-converting en-zyme are defective in production of mature IL-l beta and resis-tant to endotoxin shock. Cell 80, 401—411.
Linnik M.D., Zobrist R. H., and Hatfield M.D. (1993) Evidence
J. Neuroche/fl., Vol. 68, No. 6, 1997

CPP32-LIKE ACTIVITY IN SH-SY5Y NEUROBLASTOMA 2337
supporting a role for programmed cell death in focal cerebralischemia in rats. Stroke 24, 2002—2008.
MacManus J. P., Buchan A. M., Hill I. E., Rasquinha I., and PrestonE. (1993) Global ischemia can cause DNAfragmentation indic-ative of apoptosis in rat brain. Neuro.nci. Lett. 164, 89—92.
Martin S. J. and Green D. R. (1995) Protease activation duringapoptosis: death by a thousand cuts? Cell 82, 349—352.
Martin S. J., O‘Brien G. A., Nishioka W. K., McGahon A. J., Mah-boubia T., Saido T. C., and Green D. (1995) Proteolysis offodrin during apoptosis. J. Bio!. Chem. 270, 6425—6428.
Marx J. (1996) Mutant enzyme provides new insights into the causeof ALS. Science 272, 446—447.
Miura M., Zhu H., Rotello R., Hartwieg E. A., and Yuan j. (1993)Induction of apoptosis in fibroblasts by IL-I beta-convertingenzyme, a mammalian homolog of the C. elegans cell deathgene Ced-3. Cell 75, 65 3—660.
Nath R., Raser K. J., Stafford D., Hajimohammadreza I., Posner A.,Allen H., Talanian R., Yuen P. Y., Gilbertsen R. B., and WangK. W. (1996) Non-erythroid a-spectrin breakdown by calpainand interleukin lB-converting enzyme like proteases in apo-ptotic cells: contributory roles of both protease families in neu-ronal apoptosis. Biochem. J. 318, 683—690.
Nicholson D. W., Ambereen A. N., Thornberry N. A., VaillancourtJ. P., Ding, C. K., Gallant M., Gareau Y., Griffin P. R., LabelleM., Lazebnik Y. A., Munday N. A., Raju S. M., Smulson M. E.,Yamin T-T., Yu V. L., and Miller D. K. (1995) Identificationand inhibition of the ICE/CED-3 protease necessary for mam-malian apoptosis. Nature 376, 37—43.
Nixon R. A., Saito K. I., Grynspan F., Griffin W. R., Katayama S.,Honda T., Mohan P. S., Shea T. B., and Beermann M. (1994)Calpain activated neutral protease (calpain) system in agingand Alzheimer‘s disease. Ann. NYAcad. Sei. 747, 77—91.
Orth K., O‘Rourke K., Salvesen G. S., and Dixit V. M. (1996) Mo-lecular ordering of apoptotic mammalian CED-3/ICE-Iike pro-teases. J. Bio!. Chem. 271, 20977—20980.
Portera-Cailliau C., Hedreen J. C., Price D. L., and Koliatsos V. E.(1995) Evidence for apoptotic cell death in Huntington diseaseand excitotoxic animal models. J. Neuro,vci. 15, 3775—3787.
Satou T., Cummings B. J., and Cotman C. W. (1995) Immunoreac-tivity for BCI-2 protein with neurons in the Alzheimers diseasebrain increases with disease severity. Brain Ren. 697, 35—43.
Schlegel J., Peters t., Orrenius S., Miller D. M., Thornberry N. A.,Yainin Y. Y., and Nicholson D. W. (1996) CPP32/apopain isa key interleukin lß converting enzyme-like protease involvedin Fan-mediated apoptosis. J. Bio!. Chern. 271, 1841—1844.
Srinivasula S. M., Ahmad M., Fernandes-Alnemri T., Litwack G.,and Alnemri E. (1996) Molecular ordering of the Fas-apoptoticpathway: the Fas/APO-l protease Mch5 is a CrrnA-inhibitableprotease that activates multiple CED-3/ICE-Iike cysteine prote-uses. Proc. Nat!. A cad. Sei. USA 93, 14486—14491.
Tewari M. and Dixit V. M. (1995) Fas- and tumor necrosis factor-
induced apoptosis is inhibited by the poxvirus crmA gene prod-uct. J. Biol. C/scm. 270, 3255—3260.
Tewari M., Quan L. T., O‘Rourke K., Desnoyers S., Zeng Z., BedlerD. R., Poirier S. S., and Thompson C. B. (1995) Apoptosis inthe pathogenesis and treatment of disease. Science 267, 1456.
l‘hornberry N. A., Herbert G. B., Calaycay J. R., Chapman K. T.,Howard A. D., Kostura M. J., Miller D. K., Molineaux S. M.,Weidner J. R., Aunins J., et aI. (1992) A novel heterodimericcysteine protease is required for interleukin-IB processing inmonocytes. Nature 356, 768—774.
Troost D., Aten J., Morsink F., and de-long J. M. (1995) Apoptosisin ALS is not restricted to motor neurons: Bel-2 expressionincreased in post-central cortex, adjacent to the affected motorcortex. J. Neurol. Sei. 129, 79—80.
Vanags D. M., Porn-Ares M. I., Coppola S., Burgess D. H., andOnenius S. (1996) Protease involvement in fodrin cleavageand phosphatidylserine exposure in apoptosis. J. Bio!. C/scm.271, 31075—31081.
Vaux D. L. and Strasser A. (1996) The molecular biology ofapoptosis. Proc. Nati. Acad. Sei. USA 93, 2239—2244.
Vito P., Lacana E., and D‘adamio L. (1996) Interfering withapoptosis: Ca
2~binding protein ALG-2 and Alzheimers diseasegene ALG-3. Science 271, 521 —525.
Walkinshaw G. and Waters C. M. (1995) Induction of apoptosis incatecholaminergic PCI2 cells by L-DOPA, Implications for thetreatment of Parkinson‘s disease. J. Clin. Invest. 95, 2458—2464.
Weil M., Jacobson M. D., Coles H. S., Davies T. J., Gardner R. L.,and Raff M. C. (1996) Constitutive expression of the machineryfor programmed cell death. J. Cell Bio!. 133, 1053—1059.
Wiesner D. A. and Dawson G. (1996) Staurosporine induces pro-grammed cell death in embryonic neurons and activation of theceramide pathway. J. Neurochens. 66, 1418—1425.
Yaginuma H., Tomita M., Takashita N., McKay S. E., Cardwell C.,Yin Q. W., and Oppenheim R. W. (1996) A novel type ofprogrammed neuronal death in the cervical spinal cord of thechick embryo. J. Neuro,nci. 16, 3685-3703.
Yamatsuji T., Matsui T., Okamoto T., Kornatsuzaki K., et al. (1996)G-protein-mediated neuronal DNA fragmentation induced byfamilial Alzheimer‘s disease-associated mutants of APP. Sci-ence 272, 1349—1352.
Zanienhof S. and Guthrie D. (1995) Programmed cell death en-hances uniformity in rat cerebral hemispheres. Dcv. Neuro,sci.17, 264—266.
Zeitlin S., Liu J. P., Chapman D. L., Papaioannou V. E., and Efstrati-adis A. (1995) Increased apoptosis and early embryonic lethal-ity in mice nullizygous for the Huntington‘s disease gene homo-logue. Nat. Genel. 11, 155—163.
Ziv I., Melamed E., Nardi N., Luria D.. Achiron A., Offen D., andBarzilai A. (1994) Dopamine induces apoptosis-like cell deathin cultured chick sympathetic neurons—a possible novel patho-genetic mechanism in Parkinson‘s disease. Neuro,sci. Let!. 170,136— 140.
J. Neuroche,n., Vol. 68, No. 6, /997