characterization of a novel and selective cannabinoid cb1 receptor inverse agonist, imidazole 24b,...
TRANSCRIPT

Characterization of a novel and selective cannabinoid CB1 receptor inverse
agonist, Imidazole 24b, in rodents
Lauren P. Shearman a, D. Sloan Stribling a, Ramon E. Camacho a, Kimberly M. Rosko a,Junying Wang a, Sharon Tong a, Yue Feng b, Donald J. Marsh b, Hong Yu b, Xiaoming Guan b,
Stephanie K. Spann a, Douglas J. MacNeil b, Tung M. Fong b, Joseph M. Metzger a,Mark T. Goulet c, William K. Hagmann c, Christopher W. Plummer c, Paul E. Finke c,
Sander G. Mills c, Shrenik K. Shah c, Quang Truong c, L.H.T. Van der Ploeg b,D. Euan MacIntyre a, Alison M. Strack a,⁎
a Department of Pharmacology, Merck Research Laboratories, Rahway, NJ 07065, United Statesb Department of Metabolic Disorders, Merck Research Laboratories, Rahway, NJ 07065, United Statesc Department of Medicinal Chemistry, Merck Research Laboratories, Rahway, NJ 07065, United States
Received 3 January 2007; received in revised form 11 September 2007; accepted 16 October 2007
Available online 25 October 2007
Abstract
We document in vitro and in vivo effects of a novel, selective cannabinoid CB1 receptor inverse agonist, Imidazole 24b (5-(4-chlorophenyl)-N-
cyclohexyl-4-(2,4-dichlorophenyl)-1-methyl-imidazole-2-carboxamide). The in vitro binding affinity of Imidazole 24b for recombinant human
and rat CB1 receptor is 4 and 10 nM, respectively. Imidazole 24b binds to human cannabinoid CB2 receptor with an affinity of 297 nM; in vitro, it
is a receptor inverse agonist at both cannabinoid CB1 and CB2 receptors as it causes a further increase of forskolin-induced cAMP increase. Oral
administration of Imidazole 24b blocked CP-55940-induced hypothermia, demonstrating cannabinoid CB1 receptor antagonist efficacy in vivo.
Using ex vivo autoradiography, Imidazole 24b resulted in dose-dependent increases in brain cannabinoid CB1 receptor occupancy (RO) at 2h post-
dosing in rats, indicating that ∼50% receptor occupancy is sufficient for attenuation of receptor agonist-induced hypothermia. Imidazole 24b
administered to C57Bl/6 mice and to dietary-induced obese (DIO) Sprague–Dawley rats attenuated overnight food intake with a minimal effective
dose of 10 mg/kg, p.o. Administration had no effect in cannabinoid CB1 receptor-deficient mice. DIO rats were dosed orally with vehicle,
Imidazole 24b (1, 3 or 10 mg/kg), or dexfenfluramine (3 mg/kg) for 2 weeks. At 3 mg/kg, Imidazole 24b reduced cumulative food intake, leading
to a non-significant decrease in weight gain. Imidazole 24b at 10 mg/kg and dexfenfluramine treatment inhibited food intake and attenuated
weight gain. These findings suggest that selective cannabinoid CB1 receptor inverse agonists such as Imidazole 24b have potential for the
treatment of obesity.
© 2007 Elsevier B.V. All rights reserved.
Keywords: Cannabinoid; Food intake; Obesity; Body weight; Diet-induced obesity; Adiposity
1. Introduction
Cannabinoid receptors and associated endocannabinoids are
found in numerous physiological systems and most recently
have received attention for their potential in the pharmacolog-
ical treatment of obesity. The endocannabinoid system is
comprised of several endogenous ligands (anandamide, 2-
arachidonoyl glycerol (2-AG), 2-arachidonyl glyceryl ether and
virodhamine (Devane et al., 1992; Mechoulam et al., 1995)).
Two cannabinoid receptor subtypes, cannabinoid CB1 receptor
(Matsuda et al., 1990) and cannabinoid CB2 receptor (Munro
et al., 1993) have been cloned and characterized. Both receptors
are primarily coupled to Gi/o proteins and act to inhibit adenylyl
cyclase (Howlett, 1995) and other signaling pathways (Boua-
boula et al., 1997, 1995). Cannabinoid CB1 receptors are
Available online at www.sciencedirect.com
European Journal of Pharmacology 579 (2008) 215–224www.elsevier.com/locate/ejphar
⁎ Corresponding author. RY80Y-145, Department of Pharmacology, Merck
Research Laboratories, PO Box 2000, Rahway, NJ 07065, United States. Tel.:
+1 732 594 8367; fax: +1 732 594 3841.
E-mail address: [email protected] (A.M. Strack).
0014-2999/$ - see front matter © 2007 Elsevier B.V. All rights reserved.
doi:10.1016/j.ejphar.2007.10.033

present in the central nervous system (CNS), with high levels of
expression in brain areas implicated in motivation, mood, and
appetite regulation (Devane et al., 1992; Matsuda et al., 1990;
Munro et al., 1993 Westlake et al., 1994). In contrast,
cannabinoid CB2 receptors are predominantly localized in
peripheral tissues and immune cells (Galiegue et al., 1995;
Munro et al., 1993) and in microglial cells (Walter et al., 2003).
There are a growing number of observations, particularly in
brain and isolated heart and blood vessel preparations that
suggest that other cannabinoid receptor subtypes may exist
although the genes that encode them have not been identified
(Breivogel et al., 2001; Jarai et al., 1999).
An increasing body of evidence underscores the role of
cannabinoid CB1 receptor in appetite and in reward aspects of
food consumption. Endocannabinoids and Δ9-tetrahydrocan-
nabinol (Δ9-THC) stimulate food intake in animals and man
(Abel, 1975; Cota et al., 2003a; Foltin et al., 1988).
Hypothalamic endocannabinoid levels are elevated in leptin-
deficient or leptin receptor-deficient mice (Di Marzo et al.,
2001). Anandamide and 2-AG administered into specific brain
nuclei stimulate food intake in rodents (Jamshidi and Taylor,
2001; Williams and Kirkham, 1999). Furthermore, extracellular
concentrations of endocannabinoids fluctuate during fasting and
upon feeding in brain regions that are regulated by the anorectic
peptide leptin (Di Marzo et al., 2001; Kirkham et al., 2002).
Besides these effects on food intake, cannabinoid receptor
agonists also induce hypothermia, ataxia, catalepsy, and
analgesia in rodents (Martin et al., 1991).
Conversely, cannabinoid CB1 receptor antagonists/inverse
agonists (e.g. SR141716A) are anorexigenic and cause weight
loss in rodents (Colombo et al., 1998; Hildebrandt et al., 2003;
Ravinet Trillou et al., 2003) and in humans (Pi-Sunyer et al.,
2006). Blockade of cannabinoid CB1 receptor by SR141716A
decreases food consumption in rats (Arnone et al., 1997;
Rinaldi-Carmona et al., 1994) and marmosets (Simiand et al.,
1998) and attenuates alcohol drinking in rodents (Arnone et al.,
1997; Wang et al., 2003). SR141716A inhibits the orexigenic
effects of endocannabinoids and Δ9-THC in rats (Kirkham
et al., 2002; Williams and Kirkham, 1999, 2002). SR141716A
inhibits feeding in non-deprived rats (Colombo et al., 1998),
genetically obese animals (Bensaid et al., 2003; Di Marzo et al.,
2001; Vickers et al., 2003) and animals raised on high fat diets
(Hildebrandt et al., 2003; Ravinet Trillou et al., 2003).
Deletion of the murine gene coding for cannabinoid CB1
receptor (Cnr1) leads to leanness, resistance to diet-induced
obesity and enhanced leptin sensitivity (Cota et al., 2003b;
Ravinet Trillou et al., 2004). When maintained on standard
chow, cannabinoid CB1 receptor-deficient (Cnr1−/−) mice
exhibit reduced spontaneous food intake and body weight
gain. Cnr1−/− mice also have reduced food intake following a
fast (Di Marzo et al., 2001). The anorexigenic effects of
SR141716A are absent in Cnr1−/− mice (Di Marzo et al., 2001;
Ravinet Trillou et al., 2004). Importantly, hypothalamic
cannabinoid CB1 receptor mRNA is co-expressed in neurons
containing neuropeptides which modulate food intake, partic-
ularly corticotropin-releasing hormone, cocaine–amphetamine
regulated transcript, melanin concentrating hormone, and
prepro-orexin (Cota et al., 2003b), further implicating canna-
binoid CB1 receptor in the pathways known to control appetite.
Clearly, potential pharmacological manipulations of this
system could lead to the development of novel compounds for
the treatment of diseases in which cannabinoid CB1 receptors
are implicated. Here we describe and characterize the in vitro
characteristics, CNS receptor occupancy and in vivo effects on
indices of behaviour and of energy balance in rodents of a
novel, potent and orally active cannabinoid CB1 receptor
inverse agonist, Imidazole 24b (Plummer et al., 2005).
2. Materials and methods
2.1. Animals
All testing protocols described below were reviewed and
approved by the Merck Research Laboratories Institutional
Animals Care and Use Committee in Rahway, NJ. Rats and
mice were maintained in a 12 h/12 h light–dark cycle with free
access to food and water in group housing conditions in a
temperature controlled environment (22 °C).
Sprague–Dawley CD and DIO (Dietary-Induced Obese
Sprague–Dawley CD) rats were obtained from Charles River
Labs (Wilmington, DE). DIO rats received a moderately high
fat, high sucrose diet of 4.41 total kcal/g (D12266B, Research
Diets, Brunswick, NJ; 32% dietary fat, 16% protein, 51%
carbohydrate) starting at 4 weeks of age while chow-fed rats
were maintained on a standard rodent chow of 3.82 total kcal/g
(#7001, Teklad, Madison, WI, 4% dietary fat, 24% protein, 72%
carbohydrate). DIO rats were used at 12–16 weeks of age.
Cannabinoid CB1 receptor-deficient (Cnr1−/−) and wild-type
(Cnr1+/+) male mice (Taconic, USA) weighing 25 + 2 g were
housed at constant temperature (22 °C) and humidity (30–
70%), with food (Teklad Diet #7012, 5% dietary fat; 3.75 kcal/
g) and water available ad libitum. Cnr1−/− mice were licensed
from A. Zimmer (Zimmer et al., 1999). These mice were
backcrossed onto the C57BL/6J genetic background for ten
generations by A. Zimmer prior to N10 homozygous Cnr1−/−
mice being rederived at Taconic Farms onto the C57BL/6N
genetic background. The resulting heterozygous mice were
intracrossed to yield mice of all three possible genotypes
(Cnr1+/+; Cnr1+/−, heterozygous; Cnr1−/−). The resulting
wild-type and CB1 receptor-deficient mice were then used to
set up time-matched homozygous and wild-type intracrosses.
These intracrosses were used to generate age-matched Cnr1+/+
and Cnr1−/− mice. Mice were maintained at all times on an
irradiated rodent chow (Harlan Teklad #7012) except during the
time of study.
2.2. Automated food intake system
Rats were caged individually in an automated food intake
system in Nalgene cages with metabolism feeders attached to
them. The food cups were external to the feeder and were placed
on individual balances. Each balance was connected to a central
computer which collected readings every 5 min (weight of food
in grams to 0.1 g); for visual clarity, data from every 30′ was
216 L.P. Shearman et al. / European Journal of Pharmacology 579 (2008) 215–224

graphed. These data were analyzed to measure grams of food
consumed. Rats were transferred into the caging with
specialized feeders for at least 3 days before experimentation
to allow for acclimation.
2.3. Compounds
Tween 80 was purchased from Fisher Scientific (Pittsburgh,
PA). CP55940 (5-(1,1-dimethylheptyl)-2-[5-hydroxy-2-(3-
hydroxypropyl)cyclohexyl]phenol) was purchased from Tocris
Cookson (Ellisville, MO). S-(+)-fenfluramine (dexfenflura-
mine) was purchased from Research Biochemicals Inc. (Natick,
MA). The cannabinoid CB1 receptor inverse agonist, Imidazole
24b (5-(4-chlorophenyl)-N-cyclohexyl-4-(2,4-dichlorophenyl)-
1-methyl-imidazole-2-carboxamide; Fig. 1) was synthesized
and prepared in the Medicinal Chemistry Department at Merck
Research Laboratories, Rahway, NJ (Plummer et al., 2005).
[125I]-AM-2233 (R-2-[125I]iodophenyl-(1-(1-methylpiperidin-
2-ylmethyl)-1H-indol-3-yl)methanone) was synthesized and
labeled in-house.
2.4. Experiments
2.4.1. Radioligand binding
The long form of the human cannabinoid CB1 receptor, the
human cannabinoid CB2 receptor, and the rat cannabinoid CB1
receptor were cloned based on GenBank sequences, and were
stably expressed in clonal CHO cell lines. The binding affinity
of Imidazole 24b was measured by incubating various
concentrations of Imidazole 24b with 0.5 nM [3H] CP55940,
1.5 μg of recombinant cannabinoid CB1 receptor-CHO
membranes (or 0.1 μg of CBR2 CHO membranes) in 50 mM
Tris–HCl pH 7.4, 5 mM MgCl2, 2.5 mM EDTA, 0.5 mg/mL
fatty acid free Bovine Serum Albumin (BSA), 1× proteinase
inhibitor mix (P8340, Sigma), and 1% DMSO. After 1h
incubation at 37 °C, the reaction was stopped by filtration and
bound radioligand was separated from free radioligand by
washing the filter plate. Total specifically bound radiolabel was
10% or less of the total added radiolabel. Inhibitory IC50 values
were calculated through non-linear curve fitting.
2.4.2. Adenylyl cyclase assays
The intrinsic activity of activating or inhibiting Gi by the
cannabinoid CB1 receptor-ligand complex was measured by
incubating recombinant cannabinoid CB1 receptor-CHO cells
with various concentrations of test ligand in the presence of
10 μM forskolin, 200 μM phosphodiesterase inhibitor, 3-
isobutyl-1-methylxanthine (IBMX) in the assay buffer (Earle's
balanced salt solution supplemented with 5 mMMgCl2, 10 mM
HEPES pH7.3, 1 mg/mL BSA) at room temperature for 30 min.
Cells were lysed by boiling and intracellular cAMP level was
determined using an cAMP SPA kit (Amersham). The maximal
CP55940-mediated inhibition of forskolin-stimulated cAMP
increase is defined as 100% receptor agonist efficacy, and the
intrinsic activity of all other compounds is relative to the efficacy
of CP55940. Negative efficacy denotes inverse agonism.
Functional antagonism of receptor agonist response by
Imidazole 24b was determined by the Gi-cAMP assay as above,
except that both the CB receptor agonist 2-AG and Imidazole
24b were present in the incubation. A series of dose response
curves of 2-AG was performed in the absence or presence of
various concentrations of Imidazole 24b. The EC50 values were
determined and a Schild's plot was constructed, from which the
Kb value was determined.
To test for potential off-target activities, Imidazole 24b was
tested in 169 assays of receptor binding and enzyme activity by
MDS Pharma Services (King of Prussia, PA).
2.4.3. Receptor occupancy of Imidazole 24b in rats
Sprague–Dawley rats (male, 350 to 400g; n=3/group) were
dosed with Imidazole 24b (3, 10 and 30 mg/kg, p.o.) or vehicle
(10% Tween 80 in water). Two hours later, rats were euthanized
by CO2 asphyxiation. Plasma samples were taken by cardiac
puncture, and brains were quickly dissected and frozen in
isopentane at −40 °C and stored at −80 °C until use. The degree
of brain receptor occupancy (reciprocal of receptor binding) was
determined by quantitative receptor autoradiography in vitro,
using [125I]-AM-2233 as the radioligand. Thin (20 μm) coronal
brain sections encompassing the substantial nigra (20 μm) were
cut on a cryostat at −17 °C, and thaw-mounted onto microslides
and dried completely at room temperature. Receptor binding
was initiated by incubating the brain sections in the binding
buffer (Tris–HCl: 50 mM, pH. 7.3, MgCl2: 2 mM, CaCl2:
1 mM, KCl: 5 mM, BSA: 0.5%) containing 0.5 nM [125I]-AM-
2233 for 30 min at room temperature. Non-specific binding was
defined by similar incubation conditions in the presence of
10 μM Imidazole 24b. Brain sections were washed 3×4 min in
ice-cold washing buffer (Tris–HCl: 50 mM, pH. 7.3, MgCl2:
2 mM, CaCl2: 1 mM, KCl: 5 mM, Triton X-100: 0.05% (v/v)),
and then rinsed briefly in ice-cold distilled water before drying
at room temperature. Brain sections were exposed to X-ray film
overnight, and films were developed following standard
procedures.
Autoradiographic images were captured and analyzed with
an MCID/M2 image analysis system. Relative optical density
was determined for vehicle- or compound-treated groups (n=3
for vehicle or each dose of compound treatment). Specific
binding was obtained by subtracting the non-specific binding
Fig. 1. Structure of cannabinoid CB1 receptor inverse agonist Imidazole 24b
(5-(4-chlorophenyl)-N-cyclohexyl-4-(2,4-dichlorophenyl)-1-methyl-imidaz-
ole-2-carboxamide).
217L.P. Shearman et al. / European Journal of Pharmacology 579 (2008) 215–224

from the corresponding total binding. The maximal binding was
defined by specific binding in the vehicle treated rat brains. Oral
dosing of Imidazole 24b resulted in a dose-dependent reduction
of [125I]-AM-2233 binding to cannabinoid CB1 receptor, and %
receptor occupancy is derived by the reciprocal of % maximal
binding.
2.4.4. Reversal of cannabinoid receptor agonist-evoked
hypothermia
Sprague–Dawley rats (male, average body weight 162g)
were dosed with vehicle (10% Tween 80 in H2O) or Imidazole
24b (3, 10, or 30 mg/kg, p.o.). After 120 min, vehicle treated
animals were dosed with vehicle (0.1% Tween 80/H2O, i.p.) or
the cannabinoid receptor agonist CP-55940 (1 mg/kg, i.p.). All
Imidazole 24b treated animals were challenged with CP-55940.
Rectal temperatures were measured with a thermocouple probe
(BAT 10 Microprobe thermometer and RET-2 temperature
probe; Physitemp Instruments Inc., Clifton, NJ) prior to the p.o.
dose (t=− 120 min), prior to the i.p. dose (t=0 min) and at 45,
60 and 75 min post-CP55940 challenge.
2.4.5. Imidazole 24b effects on mouse food intake
Two experiments to test the suppression of food intake in mice
were performed. First, ad libitum fed male C57BL/6N wild-type
mice (n=9–10 per group, 11 week old, mean body weight 33 g)
were dosed with vehicle or Imidazole 24b (5 or 10 mg/kg, p.o.). In
the second experiment, to test specificity of Imidazole 24b for the
CB1 receptor, ad libitum fed,11-week oldmale wild-type (Cnr1+/+
or cannabinoid CB1 receptor-deficient (Cnr1−/−)mice were dosed
with vehicle or Imidazole 24b (10 or 16 mg/kg, p.o.).
In both experiments, Imidazole 24b was dissolved or
dispersed as a fine homogeneous suspension in 0.225%
methylcellulose/10% Tween 80 in water for subsequent oral
dosing. All mice were weighed and vehicle or Imidazole 24b
was administered by oral gavage to male mice ∼30 min prior to
the onset of the dark phase of the light cycle. A pre-weighed
aliquot of a highly palatable moderate high fat diet (Research
Diets D12266Bi; 25% kcal from sucrose, 32% kcal from fat,
4.41 kcal/g) was provided in the food hopper of the wire cage
top∼5 min prior to the onset of the dark phase of the light–dark
cycle and weighed 2 and 18 h after the onset of the dark cycle.
Additionally, all mice were weighed 18 h after the onset of the
dark phase of the light cycle. All dosing studies were of
crossover design.
2.4.6. Imidazole 24b effects on acute food intake in DIO rat
At 1h prior to dark cycle onset, male DIO rats (n=6–8 per
group, 536 g average body weight) were orally gavaged with
vehicle (10% Tween 80 in H2O) or the cannabinoid CB1
receptor inverse agonist Imidazole 24b (1, 3 or 10 mg/kg). Rats
were fed milled diet during acclimation to the caging and during
the experiment. Overnight food intake and body weight changes
were measured.
2.4.7. Imidazole 24b pharmacokinetics in DIO rat
A pharmacokinetic study with Imidazole 24b was performed
with male DIO rats (n=4). Rats were dosed at 3 mg/kg, p.o. in
10% tween/H2O. Blood samples were collected at 0.25, 0.5, 1,
2, 4, 6 and 8 h post-dosing. Plasma compound concentrations
were measured by Liquid Chromatography/Mass Spectrometry.
2.4.8. Imidazole 24b 14-day study in DIO rat
Baseline body composition was determined by DEXAscan
(Fan Beam X-Ray Bone Densitometer, QDR4500; Hologics,
Inc., Waltham, MA) 7 days prior to day 1 of dosing. Male DIO
rats (∼550 g avg. body weight; 22% body fat) were dosed orally
with vehicle (10% Tween80 in water), Imidazole 24b (1, 3 or
10 mg/kg), or S-(+)-fenfluramine (dexfenfluramine; 3 mg/kg).
Rats were dosed 1h before the onset of the dark cycle. Food
intake and body weight were recorded daily. Upon termination
on day 14, body composition was again determined by
DEXAscan analysis and brains and blood were collected.
White adipose tissues (WAT; retroperitoneal, mesenteric,
epididymal) were removed and weighed.
2.4.9. Data analysis
Data are presented as mean + S.E.M. For the in vivo
pharmacological studies, statistical analysis was performed
using Student's t-test or 1-way ANOVA followed by Dunnett's
t-test for post hoc analysis. Two-way ANOVA was used to
analyze the effects of Imidazole 24b in Cnr1+/+ and Cnr1−/−
mice and for the hypothermia study; a Bonferroni post hoc
analysis was used in conjunction with the 2-way ANOVA.
Significance level was set at Pb0.05.
3. Results
3.1. Radioligand binding
The binding affinity of Imidazole 24b for the recombinant
human or rat cannabinoid CB1 receptor or human cannabinoid
CB2 receptor was determined by measuring its ability to inhibit
the binding of [3H] CP55940. Imidazole 24b binds to human
and rat cannabinoid CB1 receptor with 4 and 10 nM affinity,
respectively. Imidazole 24b is selective for cannabinoid CB1
receptor, and it binds to the human cannabinoid CB2 receptor
with a binding affinity of 297 nM (Table 1).
As shown in Fig. 2 and Table 1, Imidazole 24b is an inverse
agonist at both cannabinoid CB1 receptor and cannabinoid CB2
receptor. It causes a further increase of forskolin-induced cAMP
increase. In contrast, a typical receptor agonist such as CP55940
Table 1
Binding IC50 value and receptor inverse agonist EC50 value of Imidazole 24b
IC50, nM EC50, nM (%act)
Human cannabinoid CB1 receptor 4 (n=16) 17 (−110%) (n=4)
Human cannabinoid CB2 receptor 297 (n=6) 165 (−74%) (n=2)
Rat cannabinoid CB1 receptor 10 (n=4) 4 (−110%) (n=2)
Binding assays were run in CHO membranes containing stably expressed
recombinant cannabinoid receptor. 0.5 nM [3H] CP55940 was used for
competition. To test functional inverse agonism, Imidazole 24b was tested in
adenylyl cyclase assays in cannabinoid receptor-containing CHO cells.
CP44940-mediated inhibition of forskolin-stimulated cAMP was used to define
100% receptor agonist efficacy. Data represent mean values with the number of
independent measurements indicated in parentheses.
218 L.P. Shearman et al. / European Journal of Pharmacology 579 (2008) 215–224

inhibits the forskolin-induced cAMP increase. These data
demonstrate that Imidazole 24b possesses intrinsic receptor
inverse agonist activity in the absence of receptor agonist.
In the presence of receptor agonist, the competitive receptor
antagonist activity of Imidazole 24b was demonstrated by
determining the dose response curve of the endogenous receptor
agonist 2-AG in the presence of various concentrations of
Imidazole 24b. Imidazole 24b shifted the receptor agonist dose
response to the right, and the Kb value from Schild's plot (4 nM)
was consistent with the binding affinity of Imidazole 24b (Fig. 3).
These data demonstrate that Imidazole 24b is a competitive
receptor antagonist when receptor agonist is present.
Imidazole 24b was tested in 169 assays of receptor binding
and enzyme activity by MDS Pharma Services (King of Prussia,
PA). In addition to its binding activity at cannabinoid CB1
receptor and cannabinoid CB2 receptor, Imidazole 24b affected
only 4 targets with IC50b10 μM (rat A3 receptor=0.5 μM;
rabbit VMAT2=0.3 μM; human HERG=4 μM; human
pp60SRC=8.7 μM). Thus, Imidazole 24b is a highly selective
cannabinoid CB1 receptor inverse agonist.
3.2. Receptor occupancy study in rats
Oral administration of Imidazole 24b resulted in a dose-
dependent increase in brain cannabinoid CB1 receptor occu-
pancy (23±1, 49±1, 66±18% receptor occupancy for 3, 10 and
30 mg/kg Imidazole 24b, respectively) at 2h post-dosing.
3.3. Imidazole 24b temperature study
The in vivo pharmacological antagonism of Imidazole 24b for
brain cannabinoid receptors was studied in a hypothermia model
(Fig. 4). The cannabinoid receptor agonist CP-55940 evoked
significant reductions in temperature relative to vehicle at 45, 60
and 75 min with a maximal decrease of 4.2 °C at 75 min post-
treatment (Dose: F(1.47)=263, Pb0.001; time: F(3, 47)=81.4,
Pb0.001; interaction: F(3, 47)=74.8, Pb0.001)). Imidazole
24b inhibited CP-55940-induced hypothermia in a dose-related
manner (Dose: F(3,95)=7.5, Pb0.001; time: F(3, 95)=273,
Pb0.001; interaction: F(9, 95)=5.6, Pb0.001)). Imidazole 24b
was not tested alone in this study. Historically, other CB1 receptor
inverse agonists that we have tested have not altered temperature
in the rat.
3.4. C57BL/6 and Cnr1−/− mouse food intake studies
In C57BL/6 mice, Imidazole 24b (10 mg/kg, p.o.) signi-
ficantly reduced spontaneous food intake at 2 and 18 h (2 h:
Fig. 3. Competitive receptor antagonist effect of Imidazole 24b. (A) Effect of
Imidazole 24b on the dose response curves of the receptor agonist 2-AG.
Specified concentrations of Imidazole 24b were included in each dose response
curve of 2-AG. (B) Schild's plot derived from (A).
Fig. 4. Imidazole 24b treatment inhibits cannabinoid CB1 receptor agonist-
induced hypothermia in rats. Imidazole 24b (0, 3, 10, or 30 mg/kg, p.o. in 10%
tween/H2O) was administered 2 h prior to the cannabinoid CB1 receptor agonist,
CP-55940 (1 mg/kg, IP in 1% tween/H2O). ⁎=Pb0.05 vs. the vehicle/CP-
55940 group. #=Pb0.05 for CP-55940 vs. the all vehicle/vehicle group at their
respective time points.
Fig. 2. Inverse receptor agonist activity of Imidazole 24b at the human
cannabinoid CB1 receptor expressed in CHO cells. To test functional inverse
agonism, Imidazole 24b was tested in adenylyl cyclase assays in cannabinoid
receptor-containing CHO cells. CP44940-mediated inhibition of forskolin-
stimulated cAMP was used to define 100% agonist efficacy.
219L.P. Shearman et al. / European Journal of Pharmacology 579 (2008) 215–224

F(29)=7.2, P=0.003; 18 h: F(29)=16.1, Pb0.001; Table 2).
Dose-dependent reductions in overnight body weight gain were
observed following treatment with 5 and 10 mg/kg of Imidazole
24b (F (29)=21.3, Pb0.001; Table 2). Imidazole 24b treatment
dose dependently reduced 2-hour (phenotype: F (1,56)=1.7,
P=0.2; dose: F(2,56)=7.8, P=0.001; interaction: F(2,56)=
8.0, Pb0.001) and overnight food intake (phenotype: F (1,56)=
1.1, P=0.3; dose: F(2,56)=3.8, P=0.03; interaction: F(2,56)=
7.2, Pb0.01) in Cnr1 +/+ mice but was without effect in Cnr1−/−
mice (Fig. 5A and B). Imidazole 24b treatment only elicited
dose-dependent reductions in body weight gain in Cnr1 +/+ mice
(Fig. 5C; phenotype: F (1,56)=0.3, P=0.6; dose: F(2,56)=7.4,
P=0.001; interaction: F(2,56)=4.7, P=0.014)).
3.5. Imidazole 24b acute DIO rat food intake study
Imidazole 24b, at 10 mg/kg, significantly reduced cumulative
overnight food intake by 28% (Fig. 6A). Reductions in food
intake were seen as early as 10 h post-dosing with 10 mg/kg
Imidazole 24b. At 18 h, food intake was significantly reduced
F(3,28)=4.8, Pb0.01). Neither Imidazole 24b at 1 nor 3 mg/kg
altered overnight food intake significantly, although a trend
towards food intake reduction, ∼10%, was observed at 3 mg/
kg. Vehicle treated rats gained an average of 5g of body weight
overnight. Dose-related reductions in overnight weight gain
were observed F(3,28) = 4.8, P=0.014); Imidazole 24b
Table 2
Food intake and body weight effects of Imidazole 24b in mice
Treatment Food intake (g) Overnight change in
body weight (g)
2 h Overnight
Vehicle 0.96±0.08 4.41±0.09 1.5±0.11
5 mg/kg 0.79±0.09 4.22±0.15 0.97±0.15
10 mg/kg 0.37±0.14 a 2.58±0.52 a−0.57±0.47 a
Imidazole 24b decreases food intake and body weight in C57Bl/6 mice
significantly (Pb0.01) by 1-way ANOVA. See text for detailed statistics.a Represents difference from vehicle of Pb0.05 by Dunnett's post hoc
analysis.
Fig. 5. Cannabinoid CB1 receptor-mediated food intake suppression by
Imidazole 24b decreases food intake in Cnr1+/+ but not Cnr1−/− mice at
(A) 2 h and (B) 24 h. C. Similarly, body weight loss evoked by Imid24 b is
observed in Cnr1+/+ but not Cnr1−/− mice. Cnr1+/+, n=10; Cnr1−/−, n=9.⁎Pb0.05 vs. corresponding vehicle; #Pb0.05 from Cnr1+/+ vehicle group.
Fig. 6. (A) Acute Imidazole 24b treatment 1 h prior to dark onset (Time 0)
reduces cumulative overnight food intake in DIO rats in a dose-related manner.
Data points in the 10 mg/kg group are statistically lower than vehicle treatment
from 10 h post-treatment onward. (B) Acute Imidazole 24b treatment reduces
overnight body weight gain in DIO rats. ⁎Pb0.05 vs. vehicle by Dunnett's post
hoc analysis; n=6 rats/group.
220 L.P. Shearman et al. / European Journal of Pharmacology 579 (2008) 215–224
treatment at 3 mg/kg led to no net change in body weight, while
10 mg/kg evoked a ∼2 g weight loss (Fig. 6B).
3.6. Pharmacokinetics of Imidazole 24b in DIO rats
Oral administration of Imidazole 24b led to peak plasma
levels of 502±190 nM and an 8 h area under the curve of 1.57±
0.34 μM⁎h.
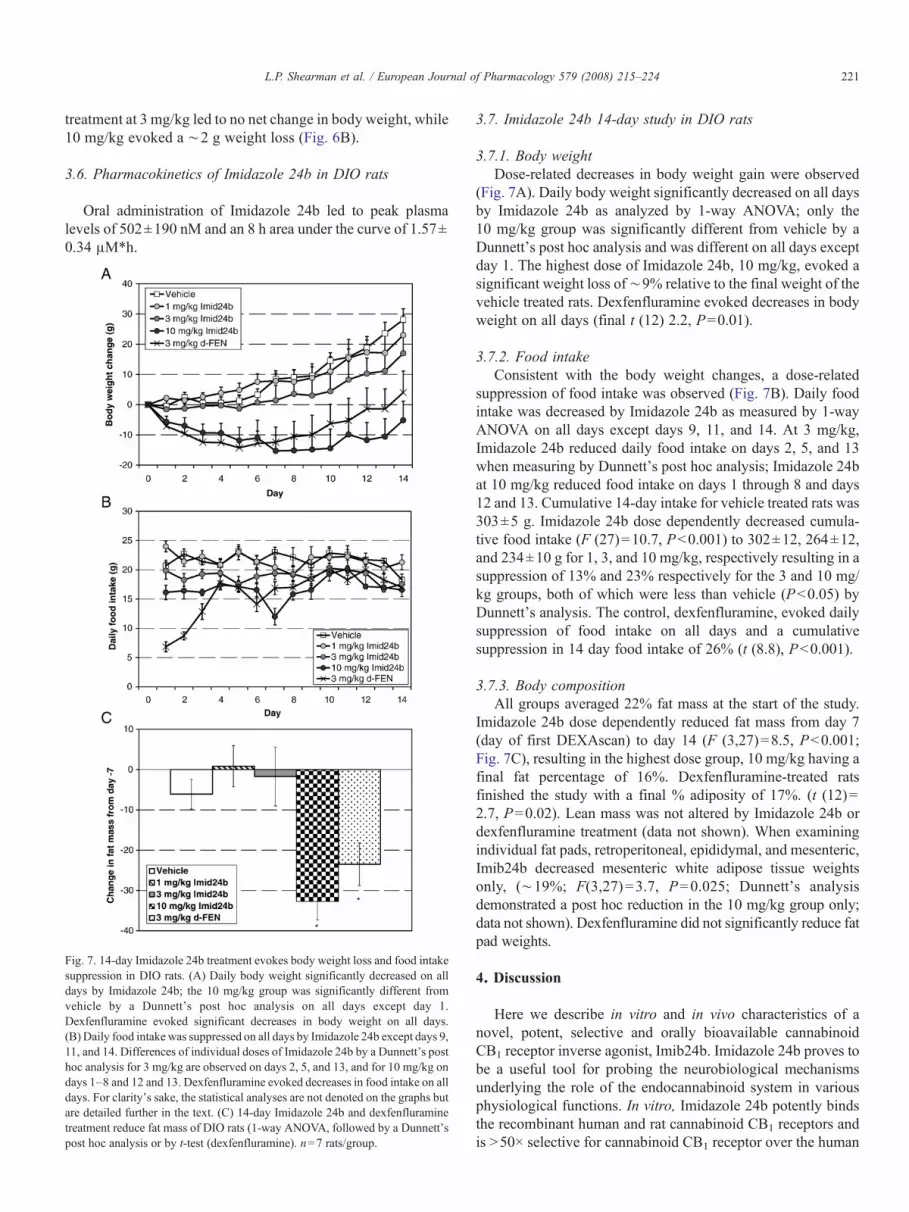
3.7. Imidazole 24b 14-day study in DIO rats
3.7.1. Body weight
Dose-related decreases in body weight gain were observed
(Fig. 7A). Daily body weight significantly decreased on all days
by Imidazole 24b as analyzed by 1-way ANOVA; only the
10 mg/kg group was significantly different from vehicle by a
Dunnett's post hoc analysis and was different on all days except
day 1. The highest dose of Imidazole 24b, 10 mg/kg, evoked a
significant weight loss of∼9% relative to the final weight of the
vehicle treated rats. Dexfenfluramine evoked decreases in body
weight on all days (final t (12) 2.2, P=0.01).
3.7.2. Food intake
Consistent with the body weight changes, a dose-related
suppression of food intake was observed (Fig. 7B). Daily food
intake was decreased by Imidazole 24b as measured by 1-way
ANOVA on all days except days 9, 11, and 14. At 3 mg/kg,
Imidazole 24b reduced daily food intake on days 2, 5, and 13
when measuring by Dunnett's post hoc analysis; Imidazole 24b
at 10 mg/kg reduced food intake on days 1 through 8 and days
12 and 13. Cumulative 14-day intake for vehicle treated rats was
303±5 g. Imidazole 24b dose dependently decreased cumula-
tive food intake (F (27)=10.7, Pb0.001) to 302±12, 264±12,
and 234±10 g for 1, 3, and 10 mg/kg, respectively resulting in a
suppression of 13% and 23% respectively for the 3 and 10 mg/
kg groups, both of which were less than vehicle (Pb0.05) by
Dunnett's analysis. The control, dexfenfluramine, evoked daily
suppression of food intake on all days and a cumulative
suppression in 14 day food intake of 26% (t (8.8), Pb0.001).
3.7.3. Body composition
All groups averaged 22% fat mass at the start of the study.
Imidazole 24b dose dependently reduced fat mass from day 7
(day of first DEXAscan) to day 14 (F (3,27)=8.5, Pb0.001;
Fig. 7C), resulting in the highest dose group, 10 mg/kg having a
final fat percentage of 16%. Dexfenfluramine-treated rats
finished the study with a final % adiposity of 17%. (t (12)=
2.7, P=0.02). Lean mass was not altered by Imidazole 24b or
dexfenfluramine treatment (data not shown). When examining
individual fat pads, retroperitoneal, epididymal, and mesenteric,
Imib24b decreased mesenteric white adipose tissue weights
only, (∼19%; F(3,27)=3.7, P=0.025; Dunnett's analysis
demonstrated a post hoc reduction in the 10 mg/kg group only;
data not shown). Dexfenfluramine did not significantly reduce fat
pad weights.
4. Discussion
Here we describe in vitro and in vivo characteristics of a
novel, potent, selective and orally bioavailable cannabinoid
CB1 receptor inverse agonist, Imib24b. Imidazole 24b proves to
be a useful tool for probing the neurobiological mechanisms
underlying the role of the endocannabinoid system in various
physiological functions. In vitro, Imidazole 24b potently binds
the recombinant human and rat cannabinoid CB1 receptors and
is N50× selective for cannabinoid CB1 receptor over the human
Fig. 7. 14-day Imidazole 24b treatment evokes body weight loss and food intake
suppression in DIO rats. (A) Daily body weight significantly decreased on all
days by Imidazole 24b; the 10 mg/kg group was significantly different from
vehicle by a Dunnett's post hoc analysis on all days except day 1.
Dexfenfluramine evoked significant decreases in body weight on all days.
(B) Daily food intake was suppressed on all days by Imidazole 24b except days 9,
11, and 14. Differences of individual doses of Imidazole 24b by a Dunnett's post
hoc analysis for 3 mg/kg are observed on days 2, 5, and 13, and for 10 mg/kg on
days 1–8 and 12 and 13. Dexfenfluramine evoked decreases in food intake on all
days. For clarity's sake, the statistical analyses are not denoted on the graphs but
are detailed further in the text. (C) 14-day Imidazole 24b and dexfenfluramine
treatment reduce fat mass of DIO rats (1-way ANOVA, followed by a Dunnett's
post hoc analysis or by t-test (dexfenfluramine). n=7 rats/group.
221L.P. Shearman et al. / European Journal of Pharmacology 579 (2008) 215–224

cannabinoid CB2 receptor. It is a receptor inverse agonist at both
cannabinoid CB1 receptor and cannabinoid CB2 receptor and in
the presence of a cannabinoid receptor agonist, Imidazole 24b is a
competitive receptor antagonist. In vivo, Imidazole 24b occupies
CNS cannabinoid CB1 receptor in a manner correlative with
Imidazole 24b functional inhibition of cannabinoid receptor
agonist modulated hypothermia and suppresses food intake and
modulates energy balance in rodents.
Endocannabinoids and synthetic cannabinoid receptor ago-
nists produce a panoply of cannabinoid-induced physiological
effects, one of which is hypothermia (Martin et al., 1991). We
demonstrated that oral (and intravenous, data not shown)
administration of Imidazole 24b attenuated cannabinoid
receptor agonist (CP-55940)-induced hypothermia, indicating
that Imidazole 24b is an effective cannabinoid CB1 receptor
antagonist in vivo. As cannabinoid receptor agonist-induced
hypothermia is believed to be centrally mediated (Fitton and
Pertwee, 1982; Rawls et al., 2002), these results also indicate
sufficient cannabinoid CB1 receptor occupancy and brain
penetration by Imidazole 24b. Indeed, using a novel ex vivo
autoradiographic method, oral administration of Imidazole 24b
resulted in a dose-dependent increase in brain cannabinoid CB1
receptor occupancy (ranging from 23 to 66% receptor
occupancy for 3, 10 and 30 mg/kg Imidazole 24b) at 2 h
post-dosing in rats, with ∼50% receptor occupancy needed for
significant attenuation of cannabinoid receptor agonist-induced
hypothermia in the rat. Others using [131I]AM281 as an in vivo
tracer/radioligand have suggested that the locomotor effects of
cannabinoids in mouse occur at ∼35–40% receptor occupancy
(Cosenza et al., 2000). In both these instances, administration of
receptor inverse agonists led to pharmacological effects at low
to moderate occupancy of the receptor. In contrast, Need (Need
et al., 2006) found ligand-specific relationships between
receptor occupancy were required for catecholaminergic
changes induced by two cannabinoid CB1 receptor inverse
agonists. Dopamine and norepinephrine efflux in the prefrontal
cortex was observed only above 65% receptor occupancy with
either compound, suggesting a very high occupancy require-
ment. However, when suppression of palatable food intake was
examined, food intake suppression by SLV319 was observed at
11–30% receptor occupancy, whereas when SR141716A was
studied, food intake suppression was only observed at receptor
occupancy of 67% or greater. Hence, generalization about
occupancy required may depend on factors such as ligand of
use, brain regions and the specific endpoint being measured.
The endocannabinoid system regulates appetitive behavior.
Cannabinoid CB1 receptors are present in brain regions
implicated in feeding and reward pathways. Our present work
demonstrates that Imidazole 24b reduces appetite in a dose-
dependent manner in both mice and DIO rats. Imidazole 24b
(10 mg/kg, p.o.) reduced food intake significantly in lean mice
at both 2 and 18 h after administration. Dose-dependent
reductions in overnight body weight were observed following
treatment with 5 and 10 mg/kg of Imidazole 24b in chow-fed
mice. Importantly, the lack of efficacy of acute Imidazole 24b
(up to 16 mg/kg, p.o.) administration in Cnr1−/− mice demon-
strates that the anorexigenic effects of Imidazole 24b are
receptor-mediated. Imidazole 24b was ineffective at 1 and 3 mg/
kg on food intake or body weight in DIO rats, but at 10 mg/kg
did inhibit food intake by ∼28%. Significant reductions in food
intake were observed as early as 5 h after administration of
10 mg/kg which led to a significant overnight body weight loss.
Furthermore, we demonstrated that when chronically
administered, Imidazole 24b evokes long-lasting dose-related
reductions in food intake, body weight gain and adiposity in
DIO rats. Chronic administration of Imidazole 24b at 3 mg/kg
reduced food intake, resulting in sustained effects on body
weight gain in DIO rats. Chronic Imidazole 24b treatment at
10 mg/kg evoked a significant decrease in body weight and
adiposity (e.g., decreased fat mass and white adipose tissue
weights). The observation that cannabinoid CB1 receptor
inverse agonists have sustained effects on body weight despite
partial tachyphylaxis of food intake inhibition have led to the
suggestion that the anti-obesity effects of cannabinoid CB1
receptor inverse agonists involve mechanisms in addition to
anorexigenesis, such as increased energy expenditure. In
support of this hypothesis, SR141716 has been shown to
increase oxygen consumption in ob/ob mice (Liu et al., 2005).
Thus, treatment of DIO rats for 14 days with the cannabinoid
CB1 receptor inverse agonist Imidazole 24b reduced body
weight gain and adipose tissue weight (particularly mesenteric
fat) without effects on lean mass and thus, likely results from
both food intake and energy expenditure changes. These
findings are consistent with the effects of chronic pharmaco-
logical blockade of cannabinoid CB1 receptors by SR141716 or
its analog, AM251 in rats (Colombo et al., 1998; Vickers et al.,
2003) and in mice (Hildebrandt et al., 2003; Ravinet Trillou
et al., 2003).
Cannabinoid CB1 receptors reportedly modulate energy
balance by direct peripheral effects in addition to the known
CNS regulation. Cannabinoid CB1 receptor mRNA is observed
in adipocytes isolated from epididymal white adipose tissue in
Cnr1+/+, but not Cnr1−/− mice. The cannabinoid CB1 receptor
agonist, WIN-55,212 at high doses stimulates lipoprotein lipase
activity in mouse adipocytes, activity which is blocked by
SR141716 (Cota et al., 2003b). Treatment of obese Zucker rats
with SR141716 has been shown to increase adiponectin
(Acrp30) expression in adipocytes (Bensaid et al., 2003). In
turn, adiponectin secreted from adipocytes, has been shown to
induce free fatty acid oxidation and decrease body weight in
mice (Fruebis et al., 2001; Masaki et al., 2003). We did not
measure serum adiponectin levels in the chronic Imidazole 24b
study and do not know if levels had been modulated by
Imidazole 24b treatment. Thus, we cannot eliminate the
possibility that a component of the effects on energy balance
may derive in part from contributions by peripheral cannabinoid
CB1 receptor mechanisms.
Together the in vitro and in vivo characteristics of Imidazole
24b demonstrate its usefulness for elucidating the roles of
cannabinoid CB1 receptor. Our demonstration of cannabinoid
CB1 receptor occupancy in the CNS and its correlation with a
physiological measure (i.e., attenuation of hypothermia)
establish that Imidazole 24b is a valuable tool to further
illuminate the CNS roles of the cannabinoid CB1 receptor and of
222 L.P. Shearman et al. / European Journal of Pharmacology 579 (2008) 215–224

endocannabinoids. Indeed, another cannabinoid CB1 receptor
inverse agonist has been recently demonstrated to reduce body
weight ∼5% in humans after a year of treatment (Pi-Sunyer et
al., 2006). Thus, these results further confirm that cannabinoid
CB1 receptor inverse agonists may represent an additional
approach to the pharmacotherapy of obesity and the better
understanding of CNS regulation of energy homeostasis.
Acknowledgments
The authors would like to thank Andreas Zimmer for
supplying the Cnr1−/− mice.
References
Abel, E., 1975. Cannabis: effects on hunger and thirst. Behav. Biol. 15,
255–281.
Arnone, M., Maruani, J., Chaperon, F., Thiebot, M., Poncelet, M., Soubrie, P.,
Le Fur, G., 1997. Selective inhibition of sucrose and ethanol intake by SR
141716, an antagonist of central cannabinoid (CB1) receptors. Psychophar-
macology (Berl.) 132, 104–106.
Bensaid, M., Gary-Bobo, M., Esclangon, A., Maffrand, J., Le Fur, G., Oury-
Donat, F., Soubrie, P., 2003. The cannabinoid CB1 receptor antagonist
SR141716 increases Acrp30 mRNA expression in adipose tissue of obese
fa/fa rats and in cultured adipocyte cells. Mol. Pharmacol. 63, 908–914.
Bouaboula, M., Poinot-Chazel, C., Bourrie, B., Canat, X., Calandra, B., Rinaldi-
Carmona, M., Le Fur, G., Casellas, P., 1995. Activation of mitogen-activated
protein kinases by stimulation of the central cannabinoid receptor CB1.
Biochem. J. 312, 637–641.
Bouaboula, M., Perrachon, S., Milligan, L., Canat, X., Rinaldi-Carmona, M.,
Portier, M., Barth, F., Calandra, B., Pecceu, F., Lupker, J., Maffrand, J.-P., Le
Fur, G., Casellas, P., 1997. A selective inverse agonist for central cannabinoid
receptor inhibits mitogen-activated protein kinase activation stimulated by
insulin or insulin-like growth factor. J. Biol. Chem. 272, 22330–22339.
Breivogel, C., Griffin, G., Di Marzo, V., Martin, B., 2001. Evidence for a new G
protein-coupled cannabinoid receptor in mouse brain. Mol. Pharmacol. 60,
155–163.
Colombo, G., Agabio, R., Diaz, G., Lobina, C., Reali, R., Gessa, G., 1998.
Appetite suppression and weight loss after the cannabinoid antagonist SR
141716. Life Sci. 63, PL113–PL117.
Cosenza, M., Gifford, A., Gatley, S., Pyatt, B., Liu, Q., Makriyannis, A.,
Volkow, N., 2000. Locomotor activity and occupancy of brain cannabinoid
CB1 receptors by the antagonist/inverse agonist AM281. Synapse 38,
477–482.
Cota, D., Marsicano, G., Lutz, B., Vicennati, V., Stalla, G., Pasquali, R., Pagotto,
U., 2003a. Endogenous cannabinoid system as a modulator of food intake.
Int. J. Obes. Relat. Metab. Disord. 27, 289–301.
Cota, D., Marsicano, G., Tschop, M., Grubler, Y., Flachskamm, C., Schubert,
M., Auer, D., Yassouridis, A., Thone-Reineke, C., Ortmann, S., Tomassoni,
F., Cervino, C., Nisoli, E., Linthorst, A.C.E., Pasquali, R., Lutz, B., Stalla,
G.K., Pagotto, U., 2003b. The endogenous cannabinoid system affects
energy balance via central orexigenic drive and peripheral lipogenesis.
J. Clin. Invest. 112, 423–431.
Devane, W., Hanus, L., Breuer, A., Pertwee, R., Stevenson, L., Griffin, G.,
Gibson, D., Mandelbaum, A., Etinger, A., Mechoulam, R., 1992. Isolation
and structure of a brain constituent that binds to the cannabinoid receptor.
Science 258, 1882–1884.
Di Marzo, V., Goparaju, S., Wang, L., Liu, J., Batkai, S., Jarai, Z., Fezza, F.,
Miura, G., Palmiter, R., Sugiura, T., Kunos, G., 2001. Leptin-regulated
endocannabinoids are involved in maintaining food intake. Nature 410,
822–825.
Fitton, A., Pertwee, R., 1982. Changes in body temperature and oxygen
consumption rate of conscious mice produced by intrahypothalamic and
intracerebroventricular injections of delta 9-tetrahydrocannabinol. Br. J.
Pharmacol. 75, 409–414.
Foltin, R., Fischman, M., Byrne, M., 1988. Effects of smoked marijuana on food
intake and body weight of humans living in a residential laboratory. Appetite
11, 1–14.
Fruebis, J., Tsao, T., Javorschi, S., Ebbets-Reed, D., Erickson, M., Yen, F.,
Bihain, B., Lodish, H., 2001. Proteolytic cleavage product of 30-kDa
adipocyte complement-related protein increases fatty acid oxidation in
muscle and causes weight loss in mice. Proc. Natl. Acad. Sci. U. S. A. 98,
2005–2010.
Galiegue, S., Mary, S., Marchand, J., Dussossoy, D., Carriere, D., Carayon, P.,
Bouaboula, M., Shire, D., Le Fur, G., Casellas, P., 1995. Expression of
central and peripheral cannabinoid receptors in human immune tissues and
leukocyte subpopulations. Eur. J. Biochem. 232, 54–61.
Hildebrandt, A., Kelly-Sullivan, D., Black, S., 2003. Antiobesity effects of
chronic cannabinoid CB1 receptor antagonist treatment in diet-induced
obese mice. Eur. J. Pharmacol. 462, 125–132.
Howlett, A.C., 1995. Pharmacology of cannabinoid receptors. Annu. Rev.
Pharmacol. Toxicol. 35, 607–634.
Jamshidi, N., Taylor, D., 2001. Anandamide administration into the ventrome-
dial hypothalamus stimulates appetite in rats. Br. J. Pharmacol. 134,
1151–1154.
Jarai, Z., Wagner, J., Varga, K., Lake, K., Compton, D., Martin, B., Zimmer, A.,
Bonner, T., Buckley, N., Mezey, E., Razdan, R., Zimmer, A., Kunos, G.,
1999. Cannabinoid-induced mesenteric vasodilation through an endothelial
site distinct from CB1 or CB2 receptors. Proc. Natl. Acad. Sci. U. S. A. 96,
14136–14141.
Kirkham, T., Williams, C., Fezza, F., Di Marzo, V., 2002. Endocannabinoid
levels in rat limbic forebrain and hypothalamus in relation to fasting, feeding
and satiation: stimulation of eating by 2-arachidonoyl glycerol. Br. J.
Pharmacol. 136, 550–557.
Liu, Y., Connoley, I., Wilson, C., Stock, M., 2005. Effects of the cannabinoid
CB1 receptor antagonist SR141716 on oxygen consumption and soleus
muscle glucose uptake in Lep(ob)/Lep(ob) mice. Int. J. Obes. (Lond). 29,
183–187.
Martin, B., Compton, D., Thomas, B., Prescott, W., Little, P., Razdan, R.,
Johnson, M., Melvin, L., Mechoulam, R., Ward, S., 1991. Behavioral,
biochemical, and molecular modeling evaluations of cannabinoid analogs.
Pharmacol. Biochem. Behav. 40, 471–478.
Masaki, T., Chiba, S., Yasuda, T., Tsubone, T., Kakuma, T., Shimomura, I.,
Funahashi, T., Matsuzawa, Y., Yoshimatsu, H., 2003. Peripheral, but not
central, administration of adiponectin reduces visceral adiposity and
upregulates the expression of uncoupling protein in agouti yellow (Ay/a)
obese mice. Diabetes 52, 2266–2273.
Matsuda, L., Lolait, S., Brownstein, M., Young, A., Bonner, T., 1990. Structure
of a cannabinoid receptor and functional expression of the cloned cDNA.
Nature 346, 561–564.
Mechoulam, R., Ben-Shabat, S., Hanus, L., Ligumsky, M., Kaminski, N.,
Schatz, A., Gopher, A., Almog, S., Martin, B., Compton, D., Pertweee, R.,
Griffine, G., Bayewitchf, M., Bargf, J., Vogel, Z., 1995. Identification of an
endogenous 2-monoglyceride, present in canine gut, that binds to
cannabinoid receptors. Br. J. Pharmacol. 50, 83–90.
Munro, S., Thomas, K., Abu-Shaar, M., 1993. Molecular characterization of a
peripheral receptor for cannabinoids. Nature 365, 61–65.
Need, A., Davis, R., Alexander-Chacko, J., Eastwood, B., Chernet, E., Phebus,
L., Sindelar, D., Nomikos, G., 2006. The relationship of in vivo central CB1
receptor occupancy to changes in cortical monoamine release and feeding
elicited by CB1 receptor antagonists in rats. Psychopharmacology (Berl.)
184, 26–35.
Pi-Sunyer, F.X., et al., for the RIO-North America Study Group, 2006. Effect of
rimonabant, a cannabinoid-1 receptor blocker, on weight and cardiometa-
bolic risk factors in overweight or obese patients: RIO-North America: a
randomized controlled trial. JAMA 295, 761–775.
Plummer, C., Finke, P., Mills, S., Wang, J., Tong, X., Doss, G., Fong, T., Lao, J.,
Schaeffer, M., Chen, J., Shen, C., Stribling, D., Shearman, L., Strack, A.,
Van der Ploeg, L., 2005. Synthesis and activity of 4,5-diarylimidazoles as
human CB1 receptor inverse agonists. Bioorg. Med. Chem. Lett. 15,
1441–1446.
Ravinet Trillou, C., Arnone, M., Delgorge, C., Gonalons, N., Keane, P.,
Maffrand, J., Soubrie, P., 2003. Anti-obesity effect of SR141716, a CB1
223L.P. Shearman et al. / European Journal of Pharmacology 579 (2008) 215–224

receptor antagonist, in diet-induced obese mice. Am. J. Physiol., Regul.
Integr. Comp. Physiol. 284, R345–R353.
Ravinet Trillou, C., Delgorge, C., Menet, C., Arnone, M., Soubrie, P., 2004.
CB1 cannabinoid receptor knockout in mice leads to leanness, resistance to
diet-induced obesity and enhanced leptin sensitivity. Int. J. Obes. Relat.
Metab. Disord. 28, 640–648.
Rawls, S.M., Cabassa, J., Geller, E.B., Adler, M.W., 2002. CB1 receptors in the
preoptic anterior hypothalamus regulate WIN 55212-2 [(4,5-dihydro-2-
methyl-4(4-morpholinylmethyl)-1-(1-naphthalenyl-carbonyl)-6H-pyrrolo
[3,2,1ij]quinolin-6-one]-induced hypothermia. J. Pharmacol. Exp. Ther.
301, 963–968.
Rinaldi-Carmona, M., Barth, F., Heaulme, M., Shire, D., Calandra, B., Congy,
C., Martinez, S., Maruani, J., Neliat, G., Caput, D., Ferrara, P., Soubrie, P.,
Breliere, J., Le Fur, G., 1994. SR141716A, a potent and selective antagonist
of the brain cannabinoid receptor. FEBS Lett. 350, 240–244.
Simiand, J., Keane, M., Keane, P., Soubrie, P., 1998. SR 141716, a CB1
cannabinoid receptor antagonist, selectively reduces sweet food intake in
marmoset. Behav. Pharmacol. 9, 179–181.
Vickers, S., Webster, L., Wyatt, A., Dourish, C., Kennett, G., 2003. Preferential
effects of the cannabinoid CB1 receptor antagonist, SR 141716, on food
intake and body weight gain of obese (fa/fa) compared to lean Zucker rats.
Psychopharmacology (Berl.) 167, 103–111.
Walter, L., Franklin, A., Witting, A., Wade, C., Xie, Y., Kunos, G., Mackie, K.,
Stella, N., 2003. Nonpsychotropic cannabinoid receptors regulate microglial
cell migration. J. Neurosci. 23, 1398–1405.
Wang, L., Liu, J., Harvey-White, J., Zimmer, A., Kunos, G., 2003.
Endocannabinoid signaling via cannabinoid receptor 1 is involved in
ethanol preference and its age-dependent decline in mice. Proc. Natl. Acad.
Sci. U. S. A. 100, 1393–1398.
Westlake, T., Howlett, A., Bonner, T., Matsuda, L., Herkenham, M., 1994.
Cannabinoid receptor binding and messenger RNA expression in human
brain: an in vitro receptor autoradiography and in situ hybridization
histochemistry study of normal aged and Alzheimer's brains. Neuroscience
63, 637–652.
Williams, C., Kirkham, T., 1999. Anandamide induces overeating: mediation by
central cannabinoid (CB1) receptors. Psychopharmacology (Berl.) 143,
315–317.
Williams, C., Kirkham, T., 2002. Reversal of delta 9-THC hyperphagia by
SR141716 and naloxone but not dexfenfluramine. Pharmacol. Biochem.
Behav. 71, 333–340.
Zimmer, A., Zimmer, A.M., Hohmann, A.G., Herkenham, M., Bonner, T.I.,
1999. Increased mortality, hypoactivity, and hypoalgesia in cannabinoid
CB1 receptor knockout mice. Proc. Natl. Acad. Sci. U. S. A. 96, 5780–5785.
224 L.P. Shearman et al. / European Journal of Pharmacology 579 (2008) 215–224