changes in phenol content and peroxidase activity during in vitro organogenesis in xanthosoma...
TRANSCRIPT

Changes in phenol content and peroxidase activity during in vitroorganogenesis in Xanthosoma sagittifolium L.
Gabriel Kanmegne1,* and N. Denis Omokolo2
1Faculty of Sciences, Department of Plant Biology, University of Dschang, P.O. Box 67, Dschang, Cameroon;2Department of Biology, University of Yaounde I, Higher Teacher Training College, P.O. Box 47, Yaounde,Cameroon; *Author for correspondence (e-mail: [email protected]; fax: (237) 345 13 81)
Received 5 June 2002; accepted in revised form 20 November 2002
Key words: Cocoyam, Differentiation, Isoperxidases, Phytohormones
Abstract
Direct plant regeneration, multiple shoot formation and callogenesis were induced from cocoyam shoot tips cul-tured in vitro. At different stages of culture, phenol content, peroxidase activity and acidic soluble isoperoxidasepatterns were analysed in plantlets. Results showed that phenol content of plantlets cultured on auxin-free mediadecreased with time, while it increased in those cultured on media supplemented with an auxin. Each form ofmorphogenesis induced with a growth regulator was preceded by an increase in total peroxidase activity. Onhormone-free medium, organogenesis occurred (single shoot development and rhizogenesis), but there was noincrease in total peroxidase activity. The appearance of isoperoxidase A2 was associated with root initiation,while the disappearance of isoperoxidase A5 and the appearance of isoperoxidase A6 preceded multiple shootformation. These results indicate that total peroxidase activity was not a proper marker for organogenesis in co-coyam. Each form of morphogenetic differentiation is associated with an alteration of the acidic isoperoxidasepattern. These enzymes can be used as biochemical markers for rooting and multiple shoot initiation in cocoyam.
Abbreviations: BAP – benzylaminopurine, BM – basal medium, 2,4-D – 2,4-dichlorophenoxyacetic acid, f.w. –fresh weight, IAA – indolylacetic acid, IBA – indolbutyric acid, NAA – naphthalene acetic acid
Introduction
Research on the mechanism of how phytohormonescause differentiation in plants remains open. Indeedplants undergo profound changes in their enzyme ex-pression which in turn regulates different biochemi-cal pathways e.g. gene expression and metabolism ofphenols. Peroxidases (EC 1.11.1.7) are the most stud-ied plant enzymes due to their abundance in plant tis-sues. The many physiological functions of peroxi-dases e.g. lignifying enzyme (Mäder and Füssl 1982),as predominant stress enzyme (Castillo 1992), as in-dolacetic oxidase (Nakajima and Yamazaki 1979) andmany others are well documented. However, the highdiversity of in vitro substrates and the existence ofmany different isoenzymes make it very difficult topostulate a definite physiological role for plant isop-
eroxidases. Phenolics constitute a group of substanceswith a broad spectrum of physiological activities.They are the main substrates of peroxidases. Theyparticipate in redox processes and in plant defensereaction (Svalheim and Robertsen 1990). Indeed, theyintervene in the regulation of developmental pro-cesses such as organogenesis through their interactionwith growth regulators. Their antagonistic effect oncytokinin-like molecules has been demonstrated (Ber-thon et al. 1993; El Hadrami and D’Auzac 1992). Thepreferential oxidation of phenolics by peroxidase pre-vents peroxidase-catalysed oxidation of auxin (Rac-chi et al. 2001).
Organogenesis from tissues cultured in vitro is adirect result of the influence of developmental andenvironmental conditions including components ofthe culture medium. In vitro culture of plant cell and
53Plant Growth Regulation 40: 53–57, 2003.© 2003 Kluwer Academic Publishers. Printed in the Netherlands.

tissue provides a useful model system for the inves-tigation of the morphological, biochemical and mo-lecular events that take place during early develop-ment of plants (De Klerk 1996).
Cocoyam (Xanthosoma sagittifolium, Araceae) isone of the main edible crops in tropical Africa andSouth America. It is cultivated for its young leavesand tubers. Previous studies in our laboratory(Omokolo et al. 1995) have shown that shoot multi-plication, somatic embryogenesis and tuberizationcan be induced in shoot tips of cocoyam (Xanthosomasagittifolium L.) cultured in vitro on Murashige andSkoog medium (Murashige and Skoog 1962) supple-mented with various combinations of IBA, NAA,2,4-D, BAP and kinetin. The biochemical aspects ofinduction of in vitro organogenesis have been inves-tigated in a number of plants including carrot, rice,pea (Chen and Luthe 1987; Choi and Sung 1984;Stirn and Jacobsen 1987). Such information is stilllacking for cocoyam. This work was designed tostudy changes in phenolic content, peroxidase activ-ity and acidic soluble isoperoxidase pattern in co-coyam shoot tips during plant regeneration, multipleshoot formation and callogenesis.
Materials and methods
White cocoyam buds were excised from three-month-old plants grown in the experimental garden of theUniversity of Yaounde I (Cameroon). Shoot tips weretrimmed and sterilized as previously described(Omokolo et al. 1995). Actively growing shoot tipsstock was established by culturing explants for 14 don a basal medium (BM) consisting of minerals andorganic elements of Murashige and Skoog medium(Murashige and Skoog 1962), 0.3% agar and 3% su-crose. Organogenesis was induced by transferring thegrowing shoot tips on BM or on BM supplementedwith either 5 µM 2,4-dichlorophenoxyacetic acid(2,4-D), 5 µM naphthalene acetic acid (NAA) or 5 µMbenzylaminopurine (BAP). Media were adjusted topH 5.7 ± 0.1 with 0.1 M HCl or 0.1 M NaOH andwere autoclaved for 20 min at 120 °C. The cultureswere maintained at 26 ± 2 °C, relative humidity of 75± 4% and irradiance of approximately 90µmol m−2 s−1 provided by cool fluorescent tubes for16-h photoperiod. Forty to fifty explants were usedper medium tested.
Phenol was extracted from plant material withmethanol 70%, and its content was measured spectro-
photometrically at 760 nm using Folin Ciocalteu asreagent (Marigo 1973). The standard curve was es-tablished using chlorogenic acid.
Crude enzyme was extracted with cold Tris-male-ate buffer 0.1 M, pH 6.5 and peroxidase activity wasmeasured spectrophotometrically at 420 nm as previ-ously described (Omokolo et al. 1995), using guaiacolas substrate. Acidic peroxidase isozymes were sepa-rated by non-denaturing polyacrylamide gel electro-phoresis (PAGE) (Davis 1964), and the gel wasstained in a solution containing 0.06% (v/v) H2O2,0.1% (w/v) benzidine and 0.1% (v/v) acetic acid. Thedrawing of the gel was made immediately after stain-ing.
Results and discussion
Results showed that direct plant regeneration was ob-tained on the phytohormone-free medium (Figure 1A)and on the basal medium supplemented with NAA(Figure 1B). Nevertheless plantlets on BM + NAAhad a higher growth rate and produced more roots (12± 2) than those on BM which produced an average of7 ± 1 roots per explant. Roots appeared between d-20and d-25 day of culture on both media. On BM +BAP, multiple shoots (an average of 10 shoots perexplant) appeared after 20 d of culture. On this me-dium, no rhizogenesis occurred (Figure 1C). On BM+ 2,4-D, shoot tips swelled and a vitrified callus wasformed on the surface of leaf primordia (Figure 1D).The appearance of callus was observed on d-15 afterinoculation. There was neither root nor shoot differ-entiation on this medium. These findings confirm thatorganogenesis in X sagittifolium tissues cultured invitro is, like in most plants, under the direct influenceof growth regulators present in the culture medium(Choi and Sung 1984).
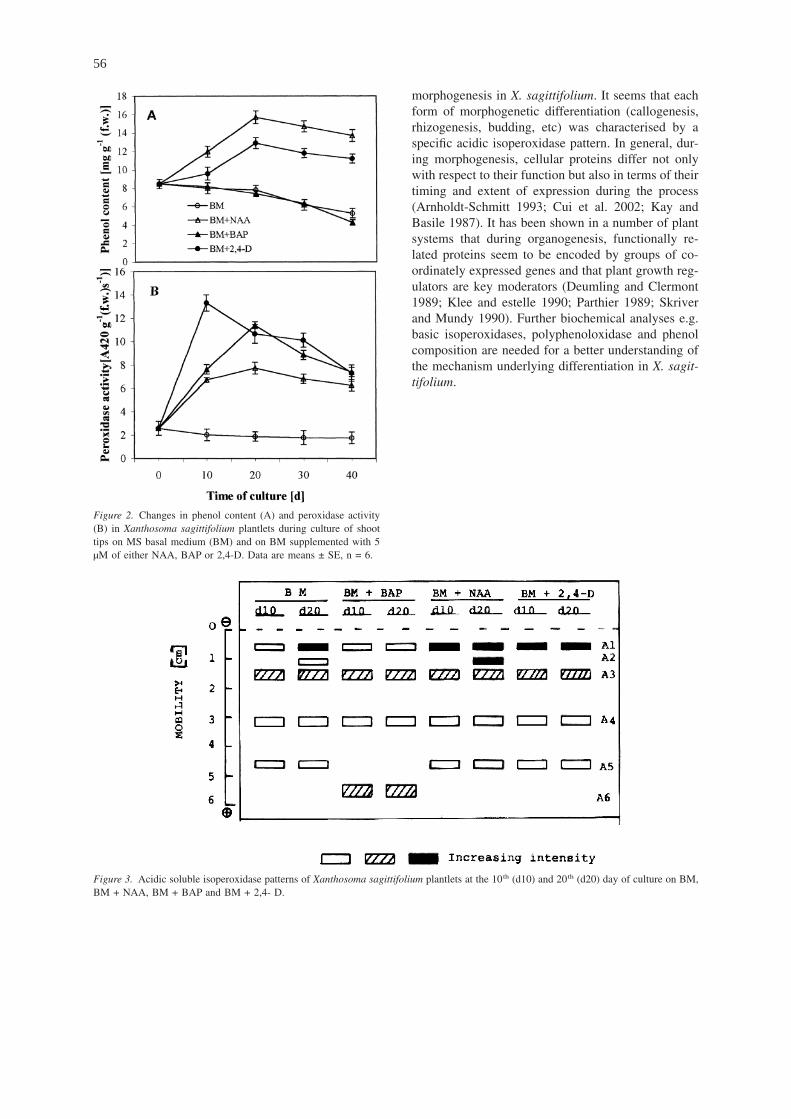
On BM and on BM + BAP phenol content de-creased with ageing, while on BM + NAA and on BM+ 2,4-D (media supplemented with auxins) it in-creased and presented a peak at d-20 after which itdecreased slightly (Figure 2A). Growth and differen-tiation need auxin (Skoog and Miller 1957). On BMand on BM + BAP, no auxin was provided by themedium. The decrease in phenol content in plantletscultured on auxin-free media may be due to the pref-erential use of this compound as substrate by peroxi-dase, leaving endogenous IAA free to promote growthand differentiation (Berthon et al. 1993). Further de-
54

termination of IAA content of the tissues would helpin verifying this hypothesis.
Organogenesis in cocoyam, when induced in thepresence of a growth regulator, was preceded by anincrease in soluble peroxidase activity, followed by adrop after the appearance of organs (Figure 2B). Or-ganogenesis also occurred on BM without a phyto-hormone, but there was no increase in peroxidase ac-tivity. It seems that the increase in enzyme activitywas due more to the presence of the growth regulatorthan to organogenesis. This was confirmed on BM +2,4-D where there was no organ differentiation butperoxidase activity also increased.
Electrophoretic patterns of acidic soluble peroxi-dase isozymes were analysed at d-10 and at d-20 (Fig-ure 3). The band A2 appeared only in conditions ofroot initiation (on BM or on BM + NAA). Moreover,its intensity was higher on BM + NAA where thenumber of roots per explant was high, than on BMwhere the number of roots per explant was low. Thisisoenzyme seems to be associated with root differen-tiation. On BM + BAP (multiple shoot induction me-dium), the band A5 disappeared, while the band A6appeared.
These results indicates that total peroxidase activ-ity was not a proper marker for the orientation of
Figure 1. Thirty-day-old plantlets of Xanthosoma sagittifolium obtained from shoot tips cultured on basal medium (A) or on basal mediumsupplemented with 5 µM of either NAA (B), BAP (C) or 2,4-D (D).
55
morphogenesis in X. sagittifolium. It seems that eachform of morphogenetic differentiation (callogenesis,rhizogenesis, budding, etc) was characterised by aspecific acidic isoperoxidase pattern. In general, dur-ing morphogenesis, cellular proteins differ not onlywith respect to their function but also in terms of theirtiming and extent of expression during the process(Arnholdt-Schmitt 1993; Cui et al. 2002; Kay andBasile 1987). It has been shown in a number of plantsystems that during organogenesis, functionally re-lated proteins seem to be encoded by groups of co-ordinately expressed genes and that plant growth reg-ulators are key moderators (Deumling and Clermont1989; Klee and estelle 1990; Parthier 1989; Skriverand Mundy 1990). Further biochemical analyses e.g.basic isoperoxidases, polyphenoloxidase and phenolcomposition are needed for a better understanding ofthe mechanism underlying differentiation in X. sagit-tifolium.
Figure 2. Changes in phenol content (A) and peroxidase activity(B) in Xanthosoma sagittifolium plantlets during culture of shoottips on MS basal medium (BM) and on BM supplemented with 5µM of either NAA, BAP or 2,4-D. Data are means ± SE, n = 6.
Figure 3. Acidic soluble isoperoxidase patterns of Xanthosoma sagittifolium plantlets at the 10th (d10) and 20th (d20) day of culture on BM,BM + NAA, BM + BAP and BM + 2,4- D.
56

References
Arnholdt-Schmitt B. 1993. Rapid changes in amplification and me-thylation pattern of genomic DNA in cultured carrot root ex-plants (Dacus carota L.). Theor. Appl. Genet. 85: 792–800.
Berthon Y., Battraw M.J. and Gaspar T. 1993. Early test using phe-nolic compounds and peroxidase activity to improve in vitrorooting of Sequoiadendron giganteum. Saussurea 24: 7–14.
Castillo F.J. 1992. Peroxidase and stress. In: Penel C., Gaspar T.h.and Greppin H. (eds), Plant Peroxidases 1980-1990. Topics anddetailed Literature on Molecular, Biochemical and Physiologi-cal Aspects. Univ Geneva, Switzerland, pp. 187–203.
Chen L.J. and Luthe D.S. 1987. Analysis of proteins from embryo-genic and non-embryogenic rice (Oryza sativa L.) calli. PlantSci. 48: 181–188.
Choi J.H. and Sung Z.R. 1984. Two dimensional gel analysis ofcarrot somatic embryogenesis proteins. Plant Mol. Biol. Rep.2: 19–25.
Cui K., Li J., Xing G., Wang L. and Wang Y. 2002. Effect of hy-drogen peroxide on synthesis of proteins during somatic em-bryogenesis in Lycium barbarum. Plant Cell, Tiss. Org. Cult.68: 187–193.
Davis B.J. 1964. Disc electrophoresis. II. Method and applicationto human serum proteins. Ann. New York Acad. Sci. 121: 404–427.
De Klerk G.J. 1996. Markers for adventitious root formation.Agronomie 16: 609–616.
Deumling B. and Clermont L. 1989. Changes in DNA content andchromosomal size during culture and plant regeneration ofScilla siberica: selective chromatin diminution in response toenvironmental conditions. Chromosoma 97: 439–448.
El Hadrami I. and D’Auzac J. 1992. Effects of growth regulatorson polyamine content and peroxidase activity in Hevea brasil-iensis callus. Ann. Bot. 69: 323–325.
Kay L.E. and Basile D.V. 1987. Specific peroxidase isoenzymes arecorrelated with organogenesis. Plant Physiol. 84: 99–105.
Klee H. and estelle M. 1990. Molecular genetic approaches to planthormone biology. Ann. Rev. Plant Physiol. Plant Mol. Biol. 42:529–551.
Mäder M. and Füssl R. 1982. Role of peroxidase in lignification oftobacco cells. II. Regulation by phenolic compounds. PlantPhysiol. 70: 1132–1134.
Marigo G. 1973. Sur une méthode de fractionnement etd’estimation des composés phénoliques chez les végétaux.Analysis 2: 106–110.
Murashige T. and Skoog F. 1962. A revised medium for rapidgrowth and bioassays with tobacco tissue culture. Plant Physiol.15: 473–497.
Nakajima R. and Yamazaki I. 1979. The mechanism of indole-3-acetic acid oxidation by horseradish peroxidase. J. Biol. Chem.254: 872–878.
Omokolo N.D., Tsala N.G., Kanmegne G. and Balangé A.P. 1995.In vitro induction of multiple shoots, plant regeneration and tu-berization from shoot tips of cocoyam. C. R. Acad. Sci. 318:773–778.
Omokolo N.D., Djocgoue P.F., Nana L. and Debost M. 1995. Vari-ation and inheritance of peroxidase activity and phenol andsaccharide content in cacao in relation to susceptibility to blackpod disease. Biol. Plant 37: 429–436.
Parthier B. 1989. Hormone-induced alterations in plant gene ex-pression. Biochem. Physiol. Pflanz 185: 289–314.
Racchi M.L., Bagnoli F., Balla I. and danti S. 2001. Differentialactivity of catalase and superoxide dismutase in seedlings andin vitro micropropagated oak (Quercus robur L.). Plant Cell.Rep. 20: 169–174.
Skoog F. and Miller C.O. 1957. Chemical regulation of growth andorgan formation in plant tissues cultured in vitro. Symp. Soc.Exp. Biol. 11: 118–130.
Skriver K. and Mundy J. 1990. Gene expression in response to ab-cissic acid and osmotic stress. Plant Cell 2: 503–512.
Stirn S. and Jacobsen H.J. 1987. Marker proteins for embryogenicdifferentiation patterns in pea callus. Plant Cell Rep. 6: 50–54.
Svalheim O. and Robertsen B. 1990. Induction of peroxidases incucumber hypocotyls by wounding and fungal infection. Phys-iol. Plant. 78: 261–267.
57
