carotenoids provide the major antioxidant defence in the globally significant n 2 -fixing marine...
TRANSCRIPT

Carotenoids provide the major antioxidant defencein the globally significant N2-fixing marinecyanobacterium Trichodesmiumemi_1913 1897..1908
Dovi Kelman,1* Ami Ben-Amotz,2
Ilana Berman-Frank1
1The Mina and Everard Goodman Faculty of LifeSciences, Bar Ilan University, Ramat Gan 52900, Israel.2The National Institute of Oceanography, IsraelOceanographic and Limnological Research, TelShikmona, POB 8030, Haifa 31080, Israel.
Summary
Photosynthetic oxygen-evolving microorganismscontend with continuous self-production of molecularoxygen and reactive oxygen species. The deleteriouseffects of reactive oxygen species are exacerbatedfor cyanobacterial nitrogen-fixers (diazotrophs) dueto the innate sensitivity of nitrogenase to oxygen.This renders incompatible the processes of oxygen-evolving photosynthesis and N-fixation. We examinedtotal antioxidative potential of various diazotrophicand non-diazotrophic cyanobacteria. We focusedon Trichodesmium spp., a bloom-forming marinediazotroph that contributes significantly to globalnitrogen fixation. Among the species tested, Tri-chodesmium possessed the highest antioxidantactivity. Moreover, while proteins constituted thedominant antioxidative component of all other cyano-bacteria tested, Trichodesmium was unique in thatsmall-molecule natural products provided the major-ity of antioxidant activity, while proteins constitutedonly 13% of total antioxidant activity. Bioassay-guided fractionation followed by high-performanceliquid chromatography profiling of antioxidantpurified fractions identified the highly potent antio-xidant all-trans-b-carotene, and small amounts of9-cis-b-carotene and retinyl palmitate. Search of theTrichodesmium genome identified protein sequenceshomologous to key enzymes in the b-carotene toretinyl palmitate biosynthetic pathway, including33–37% identity to lecithin retinol acyltransferase.The present study demonstrates the importance of
carotenoids in Trichodesmium’s arsenal of defensivecompounds against oxidative damage and protectionof nitrogenase from oxygen and its radicals.
Introduction
A fundamental aspect of aerobic metabolism is theharmful generation of partially reduced species of mole-cular oxygen, generally referred to as reactive oxygenspecies (ROS) or oxyradicals. The predominant ROS,such as hydrogen peroxide (H2O2) and hydroxyl radicals(·OH), are strong oxidants, leading to protein degradation,enzyme inactivation, lipid peroxidation, damage to DNA(Halliwell, 1999) and induction of programmed cell death(Ozben, 2007).
Reactive oxygen species are especially common inaquatic ecosystems, where they accumulate both in theopen ocean and in coastal zones (Blough and Zepp, 1995;Voelker et al., 2000). High irradiances, prevalent in manyregions of the surface oceans, cause the photochemicalproduction of ROS (Andrews et al., 2000; Falkowski andRaven, 2007). Significant ROS production occurs inbiological systems especially by photosynthetic, oxygen-evolving microorganisms (Fridovich, 1998; Falkowski andRaven, 2007). Defences against the harmful effects ofROS are especially important in nitrogen-fixing (diaz-otrophic) cyanobacteria, as the processes of oxygen-evolving photosynthesis and N-fixation are incompatibledue to the innate sensitivity of the oxygen-sensitiveenzyme required for N-fixation, nitrogenase.
To counteract the deleterious effects of potentiallyharmful ROS, aerobic cells have developed complexantioxidant systems based on low-molecular-weightscavengers and specifically adapted enzymes. It is wellestablished that the major enzyme systems superoxidedismutase (SOD), catalase and peroxidase (Halliwell andGutteridge, 1990; Sies, 1991; Halliwell, 1999; Halliwelland Gutteridge, 1999; Asada, 2000) are the primary intra-cellular antioxidant defences in living organisms, includingoxygenic photoautotrophs and photosynthetic diazotro-phs (Mallick and Mohn, 2000; Berman-Frank et al., 2001;Berman-Frank et al., 2004; Wolfe-Simon et al., 2005).
Cells also maintain a variety of non-enzymatic antio-xidants, such as ascorbic acid, reduced glutathione,
Received 15 October, 2008; accepted 22 February, 2009. *Forcorrespondence. E-mail [email protected]; Tel. (+ 972)54 4844842; Fax (+ 972) 3 6357355.
Environmental Microbiology (2009) 11(7), 1897–1908 doi:10.1111/j.1462-2920.2009.01913.x
© 2009 The AuthorsJournal compilation © 2009 Society for Applied Microbiology and Blackwell Publishing Ltd

tocopherols, carotenoids, flavonoids, phenols and aro-matic amines (Halliwell, 1999; Mallick and Mohn, 2000).While the role of antioxidant enzymes is well known, therelative contribution of biologically active small-moleculenatural products to the total antioxidant defences inphotosynthetic organisms and specifically in diazotrophiccyanobacteria such as Trichodesmium is still unclear(Wolfe-Simon et al., 2005).
A variety of assays have been developed to measurethe contribution of specific antioxidants, as well as thetotal antioxidant activity present within living cells. A bio-logical antioxidant has been defined as ‘any substancethat, when present at low concentrations compared withthose of an oxidizable substrate, significantly delays orprevents oxidation of that substrate’ (Halliwell and Gut-teridge, 1995). When ROS are generated in living cells, awide variety of antioxidants come into play (Halliwell andGutteridge, 1990; 1999; Sies, 1991). The relative impor-tance of these as protective agents depends on whichROS is generated, how and where it is generated, and onthe specific target of damage (Halliwell and Gutteridge,1995).
Several radical-scavenging tests were designed tomeasure total antioxidant activity (Prior and Cao, 1999).However, these tests are indirect methods that measurethe ability of antioxidants in the sample to inhibit theoxidative effects of reactive species purposefully gener-ated in the reaction mixture. In contrast, the ferricreducing/antioxidant power (FRAP) assay is a direct testthat measures the total antioxidant levels in a sample(Benzie and Strain, 1996; 1999). The FRAP assay usesantioxidants as reductants in a redox-linked colorimetricmethod. In this method, the reduction of a ferric 2,4,6-tripyridyl-s-triazine (FeIII-TPTZ) complex to the colouredferrous (FeII) form is measured in a linear, reproducibleand robust assay. The assay is considered a direct andvigorous method of testing the ‘total’ antioxidant activitypresent of a given system, as no activity-changing inter-actions between antioxidants occur in this system (Benzieand Strain, 1996).
In the present study, we examined the total antioxidantactivity of diazotrophic and non-diazotrophic cyanobacte-ria with different morphologies and strategies of separat-ing nitrogen fixation from photosynthesis. We specificallyfocused on the marine diazotroph Trichodesmium, whichforms extensive blooms in the oligotrophic subtropical andtropical oceans, and contributes significantly to globalnitrogen fixation (Capone et al., 1997; Davis and McGilli-cuddy, 2006). Protection of nitrogenase from oxygen inTrichodesmium is especially complex as Trichodesmiumhas an intricate strategy of spatial and temporal segrega-tion between nitrogen fixation and photosyntheticallyevolved oxygen during the photoperiod (Chen et al.,1998; 1999; Lin et al., 1998; Berman-Frank et al., 2001;
Kupper et al., 2004). Downregulation of photosystem II(PSII) and an increase in light-dependent oxygen con-sumption via the Mehler reaction occurs during hours ofhigh nitrogen fixation (Berman-Frank et al., 2001; Kupperet al., 2004; Milligan et al., 2007). Yet, ROS are present inTrichodesmium cells during hours of high N2 fixation,including zones where the oxygen-sensitive nitrogenaseis clustered (Berman-Frank et al., 2001), and are impli-cated in the induction of autocatalytic programmed celldeath in natural populations of Trichodesmium (Berman-Frank et al., 2004; 2007). Our results indicated thatTrichodesmium sp. IMS101 possessed the highest anti-oxidant activity among the species tested that was prima-rily due to small-molecule natural products. Consequently,further objectives of the current study were to identify thechemical components of the organic extracts of thiscyanobacterium, and to determine the compounds thatcontribute most significantly to its antioxidant activity anddefences against oxygen and ROS damage.
Results and discussion
The inherent conflict between nitrogen fixation andphotosynthetic oxygen evolution in diazotrophic cyano-bacteria has driven the evolution of many protectivemechanisms against oxygen and ROS. These range fromoxygen avoidance, to temporal separation found in uni-cellulars (e.g. Cyanothece) whereby N-fixation occurs atnight, to spatial separation via specialized cells, hetero-cysts, where N-fixation is isolated from the oxygen-evolving step in photosynthesis (e.g. Anabaena), and tocomplex spatial-temporal systems such as those foundin Trichodesmium (reviewed by Gallon, 1992; 2001;Bergman et al., 1997; Berman-Frank et al., 2001). More-over, a multitude of antioxidative enzymes and other mol-ecules have evolved to counteract ROS in these cells.
Most studies, including those on cyanobacteria, havetested the activity of specific antioxidants, for example,SOD (Eitinger, 2004; Priya et al., 2007), peroxidase(Zhao et al., 2007), phycocyanin (Soni et al., 2008),ascorbic acid (Noctor and Foyer, 1998), glutathione(Noctor and Foyer, 1998), tocopherols (Maeda and Del-laPenna, 2007), carotenoids (Kobayashi et al., 1997; Yanet al., 1999; Schafer et al., 2005) and phlorotannins(Nakai et al., 2006), or looked for novel antioxidant activi-ties to be used in biomedical (Potterat, 1997; Halvorsenet al., 2002; Takamatsu et al., 2003; Li et al., 2007) or infood preservation applications (Araujo et al., 2006), ratherthan examining the total antioxidant activity in the cell. Toprovide a more holistic approach, we tested total antioxi-dant activity, using the FRAP method.
We examined the total antioxidative potential ofdifferent strategies of diazotrophy and compared themwith non-diazotrophic cyanobacteria. Antioxidant activity
1898 D. Kelman, A. Ben-Amotz and I. Berman-Frank
© 2009 The AuthorsJournal compilation © 2009 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 11, 1897–1908

assays were performed on cell extracts of representativediazotrophs: a unicellular diazotroph (Cyanothece), a fila-mentous heterocystous (Anabaena), a non-heterocystousfilamentous (Trichodesmium) and non-diazotrophic fila-mentous (Prochlorothrix) and unicellular (Synechococ-cus) cyanobacteria (Fig. 1). Total antioxidant activity wasdetermined as FRAP value in mM per mg crude extract.Trichodesmium sp. IMS101 showed the highest activityamong the various cyanobacteria tested (Fig. 1A), with amean total FRAP value of 41.8 � 0.88 mM per mg totalcrude extract, containing the organic and protein extracts.This value is four- to eightfold higher than the FRAPvalues measured for all other cyanobacteria tested. Theseother species all displayed very similar antioxidative activ-ity levels (mean total FRAP value of 7.6 � 0.78 mM per mgtotal crude extract, Fig. 1A).
To our knowledge, no other study has been publishedcomparing total antioxidative potential in diazotrophicversus non-diazotrophic cyanobacteria. The application ofFRAP analyses allowed us to test this on crude and spe-cific extracts. To identify which cell component contributedto the total antioxidant activity, antioxidant activity assayswere performed on three different types of extracts: non-polar and polar organic extracts, as well as crude proteinextracts (Fig. 1B). Excluding Trichodesmium, the highestantioxidative activity in all cyanobacteria tested was foundin the protein extracts, which exhibited similar levelsof antioxidant activity with an average FRAP value of19.5 � 2.1 mM per mg crude protein extract (Fig. 1B).Surprisingly, no significant differences (one-way ANOVA,P-value = 0.18) in FRAP activity of the protein extractswere observed between the diazotrophic and non-diazotrophic cyanobacteria, including that measured inTrichodesmium (Fig. 1B).
Trichodesmium is unique in that the highest antioxidantactivity found was in the non-polar and polar organicextracts with mean FRAP values of 66.9 � 0.25 and41.7 � 1.2 mM per mg crude organic extract respectively(Fig. 1B). These values are two- to threefold higher thanthe maximal activities (measured in the protein extracts)from all other cyanobacteria we tested (Fig. 1B). The rela-tive contribution of each extract of Trichodesmium sp.IMS101 to the total antioxidant activity was 54% by thenon-polar organic extract, 33% by the polar organicextract and only 13% by the protein extract (Fig. 1C). Incontrast, the mean relative contribution of each extract ofthe other cyanobacteria tested to the total antioxidantactivity was 86% by the protein extract and only 9% by thenon-polar organic extract and 5% by the polar organicextract (Fig. 1D). The relative contribution of each extractof Trichodesmium is conspicuously comparable to therelative contribution of human plasma antioxidants (alsodetermined by the FRAP assay), where only 10% of theactivity is due to proteins and the remaining activity is due
to small-molecule natural products, mainly uric acid andascorbic acid (Benzie and Strain, 1996).
This remarkable difference in the composition andactivity levels of antioxidants in Trichodesmium, com-pared with those measured for the other cyanobacteria,may result from the challenge of juggling the twoprocesses of N-fixation and photosynthesis during thephotoperiod without specialized cells that can spatiallyseparate the two incompatible processes (Berman-Franket al., 2001). To overcome the deleterious effects ofoxygen during the period of N-fixation, Trichodesmiumdownregulates PSII (Berman-Frank et al., 2001; Kupperet al., 2004), consumes a significant amount of oxygen viaa highly active Mehler cycle (Kana, 1992; 1993; Milliganet al., 2007) and employs antioxidative proteins such asSOD (Cunningham and Capone, 1992; Wolfe-Simonet al., 2005; Priya et al., 2007) and ascorbate peroxidase(I. Berman-Frank, unpublished).
The SOD is considered a major antioxidative enzyme incyanobacteria and phytoplankton. Differences in the SODtypes exist and may also have an effect on the importanceof the antioxidative potential of the proteins in the repre-sentative cyanobacteria. In Anabaena (i.e. Nostoc-type),both FeSOD and MnSOD are found while CyanotheceCCY 0110 has only a MnSOD (C. Dupont, pers. comm.),and Synechococcus possess only the FeSOD types(Wolfe-Simon et al., 2005). In Trichodesmium, MnSODand NiSOD were detected (Wolfe-Simon et al., 2005). Yet,these systems may not be sufficient as the percentageof antioxidant activity from all proteins including theseenzymes in Trichodesmium is ~13% (Fig. 1C), and ROSare still present intracellularly along Trichodesmiumtrichomes even during periods of high N-fixation (Berman-Frank et al., 2001).
In Trichodesmium sp. IMS101, the battle againstoxygen and ROS may be augmented with potent antioxi-dant activity from non-polar small-molecule natural prod-ucts (Fig. 1B). To test the radical-scavenging effect ofthe non-polar crude extract, this extract was run on athin-layer chromatography (TLC) and stained with1,1-diphenyl-2-picrylhydrazyl (DPPH) radical (data notshown). A non-polar component of this extract producedan intense bright yellow zone indicating a strong radical-scavenging activity. High-performance liquid chromatog-raphy (HPLC) profiling of the various components of thisextract (Fig. 2) revealed that the extract contains zeaxan-thin, chlorophyll a, all trans-b-carotene, 9-cis-b-caroteneand some non-pigmented unidentified lipids. In contrast,the polar extract contained chlorophyll a, chlorophyll aderivative and some non-pigmented partially oxidized uni-dentified lipids (Fig. 3). The fact that the polar organicextract of Trichodesmium exhibited moderate antioxidantactivity (Fig. 1B) is not surprising, as chlorophyll a deriva-tives isolated from marine alga (Cahyana et al., 1992), as
Antioxidant defence in the cyanobacterium Trichodesmium 1899
© 2009 The AuthorsJournal compilation © 2009 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 11, 1897–1908
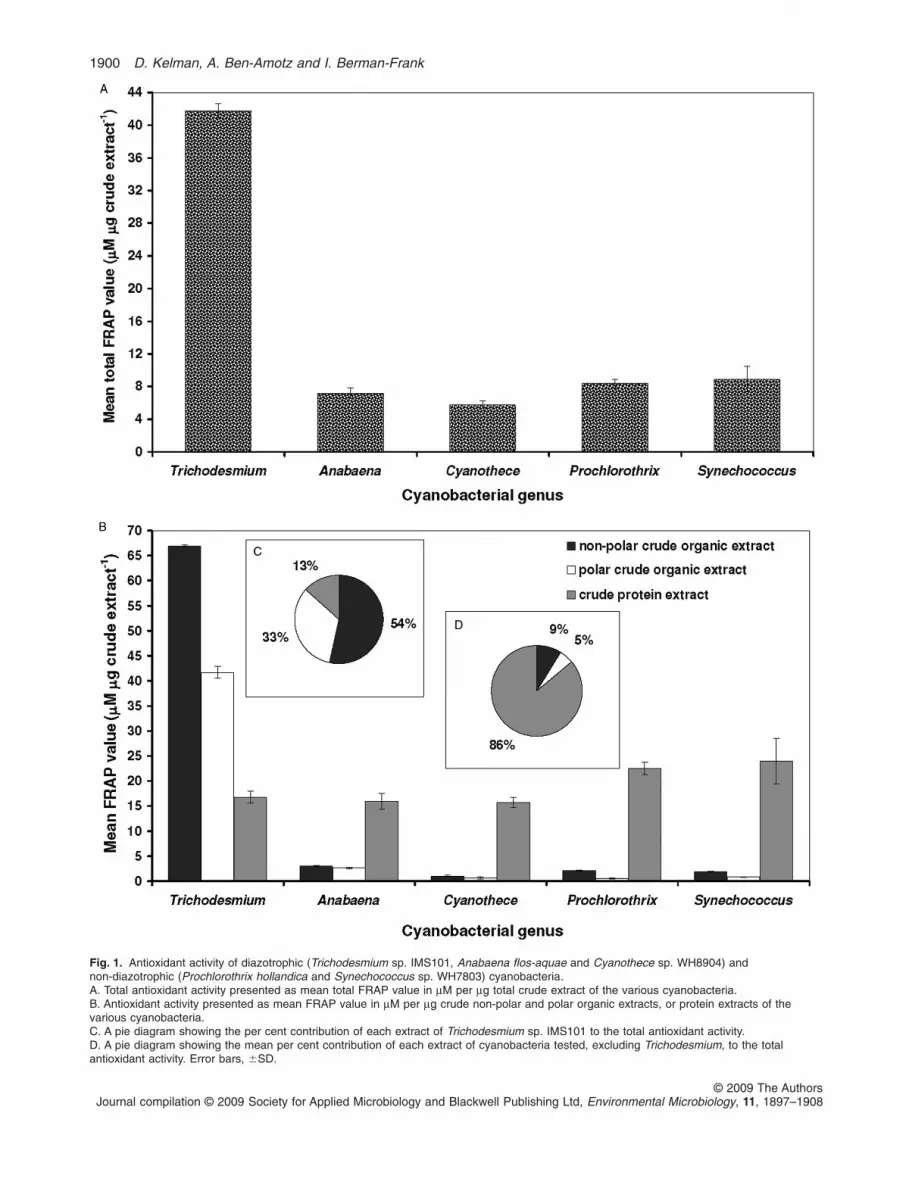
Fig. 1. Antioxidant activity of diazotrophic (Trichodesmium sp. IMS101, Anabaena flos-aquae and Cyanothece sp. WH8904) andnon-diazotrophic (Prochlorothrix hollandica and Synechococcus sp. WH7803) cyanobacteria.A. Total antioxidant activity presented as mean total FRAP value in mM per mg total crude extract of the various cyanobacteria.B. Antioxidant activity presented as mean FRAP value in mM per mg crude non-polar and polar organic extracts, or protein extracts of thevarious cyanobacteria.C. A pie diagram showing the per cent contribution of each extract of Trichodesmium sp. IMS101 to the total antioxidant activity.D. A pie diagram showing the mean per cent contribution of each extract of cyanobacteria tested, excluding Trichodesmium, to the totalantioxidant activity. Error bars, �SD.
1900 D. Kelman, A. Ben-Amotz and I. Berman-Frank
© 2009 The AuthorsJournal compilation © 2009 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 11, 1897–1908

well as from marine bivalves and diatoms (Yamamotoet al., 1992; Watanabe et al., 1993), were shown to exhibitantioxidant activity.
Bioassay-guided fractionation of the active non-polarcrude organic extract resulted in the isolation of a highlyactive purified fraction. Yet, lack of sufficient materialprevented further fractionation of this purified fraction.HPLC profiling of this antioxidant purified fraction(Fig. 4A) revealed that it contained primarily all trans-b-carotene and small amounts of 9-cis-b-carotene, retinyl
palmitate and some non-pigmented unidentified lipids.The identification of all trans-b-carotene (Fig. 4B), 9-cis-b-carotene (Fig. 4C) and retinyl palmitate (data notshown) matched the typical UV-visible absorbancespectra of these compounds present in our internal stan-dard database.
Carotenoids are natural compounds with lipophilicproperties. More than 600 different carotenoids havebeen identified, where b-carotene is the most prominent.Carotenoids possess many important biological functions,
Fig. 2. HPLC profile of Trichodesmium sp. IMS101 non-polar organic extract. Peak identification: (1) unidentified lipids; (2) zeaxanthin(6.244 min, lmax 449.9, 478.0 nm); (3) chlorophyll a (7.258 min; lmax 432.9, 666.1 nm); (4) all trans-b-carotene (18.469 min; lmax 452.4,479.2 nm); and (5) 9-cis-b-carotene (20.730 min; lmax 446.3, 473.1 nm). The numbers indicated in brackets refer to the corresponding retentiontime and absorption maxima for peaks in the HPLC chromatogram.
Fig. 3. HPLC profile of Trichodesmium sp. IMS101 polar organic extract. Peak identification: (1) partially oxidized lipids (2.993 min); (2)chlorophyll a (7.224 min; lmax 432.9, 666.0 nm); and (3) chlorophyll a derivative (8.651 min). The numbers indicated in brackets refer to thecorresponding retention time and absorption maxima (when appropriate) for peaks in the HPLC chromatogram.
Antioxidant defence in the cyanobacterium Trichodesmium 1901
© 2009 The AuthorsJournal compilation © 2009 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 11, 1897–1908

among which are their outstanding antioxidant activities(Burton and Ingold, 1984; Liebler, 1993; Edge et al., 1997;Beutner et al., 2001). Most carotenoids contain anextended system of conjugated double bonds, which isresponsible for their antioxidant activity (Sies and Stahl,1995). In biological systems, carotenoid antioxidant activ-ity is involved in protection against both singlet oxygenand triplet oxygen as radical chain-braking antioxidants,as well as being very effective biological quenchers ofoxygen radicals by efficient energy transfer (Edge et al.,1997; Beutner et al., 2001). Telfer and colleagues (1994)have shown that b-carotene can act as an efficientquencher of singlet oxygen generated within isolated PSIIreaction centres. The ability to quench oxygen radicals isenhanced with increasing number of conjugated doublebonds (Conn et al., 1991). In addition, cis and all-transisomers of carotenoids were shown to have different bio-logical reactivities towards oxygen radicals, where alltrans-b-carotene has a higher reactivity than 9-cis-b-carotene (Conn et al., 1991). As noted by Britton (1995),carotenoids are not just ‘another group of natural pig-ments’. They are substances with special and remarkableproperties in biological systems. In photosynthetic organ-isms, carotenoids can quench triplet chlorophyll, as well
as oxygen radicals (Edge et al., 1997). b-Carotene is con-sidered an unusual type of antioxidant (Burton and Ingold,1984). At low oxygen concentrations (~0.01 atm.), itsantioxidant activity increases substantially. However, athigher oxygen concentrations (~1 atm.), b-carotene losesits antioxidant activity and may even act as a pro-oxidant,particularly at relatively high concentrations (Burton andIngold, 1984). Oxygen gradients within and around indi-vidual Trichodesmium colonies have been measured withmicroelectrodes by Paerl and Bebout (Paerl and Bebout,1992). Depending on the colony examined, they foundinternal oxygen concentrations from 10 to 400 mM(4.1 ¥ 10-7 to 0.0098 atm.). The low oxygen concentra-tions measured in Trichodesmium colonies that resultfrom a combination of the downregulation of PSII(Berman-Frank et al., 2001; Kupper et al., 2004) andhigh Mehler activity (Milligan et al., 2007) suggest thatb-carotene should function predominantly as a highlypotent antioxidant under these conditions.
Additionally, we identified the presence of smallamounts of retinyl palmitate supplementing the two stere-oisomers of b-carotene in the purified fraction of Tri-chodesmium with high antioxidative activity (Fig. 4). Thisidentification was confirmed by mass spectrometry analy-
Fig. 4. A. HPLC profile of the antioxidant purified fraction from Trichodesmium sp. IMS101 non-polar organic extract. Peak identification: (1)unidentified lipids; (2) retinyl palmitate (9.063 min; lmax 326.6 nm); (3) all trans-b-carotene (18.071 min); and (4) 9-cis-b-carotene (20.209 min).The numbers indicated in brackets refer to the corresponding retention time and absorption maxima (when appropriate) for peaks in the HPLCchromatogram.B. The UV-visible absorbance spectrum corresponding to HPLC peak # 3 (18.071 min).C. The UV-visible absorbance spectrum corresponding to HPLC peak # 4 (20.209 min).
1902 D. Kelman, A. Ben-Amotz and I. Berman-Frank
© 2009 The AuthorsJournal compilation © 2009 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 11, 1897–1908

sis (data not shown). The presence of retinyl palmitate inTrichodesmium was surprising, although both retinylpalmitate and its precursor all trans-retinol (vitamin A) arewell-known antioxidants (Sanjeev Kumar et al., 1981;Tesoriere et al., 1997).
Retinyl palmitate originates from b-carotene througha series of biosynthetic steps (Fig. 5). Our BLAST
(Altschul et al., 1990) searches of the Trichodesmiumerythraeum IMS101 genome (http://genome.jgi-psf.org/finished_microbes/trier/trier.home.html) identified severalprotein sequences that are homologous to the keyenzymes in this biosynthetic pathway. A protein sequencein Trichodesmium showed homology to carotenoid oxyge-nase (EC 1.14.99.36), which converts all trans-b-caroteneto all trans-retinal, and was assigned as GenBank acces-sion number YP_723313. Another protein sequence inTrichodesmium had 29–37% identity to retinal reductase(EC 1.1.1.105), which converts all trans-retinal to all trans-retinol. Moreover, we detected a protein sequence inTrichodesmium with 33–37% identity to lecithin retinolacyltransferase (LRAT, EC 2.3.1.76), the enzyme respon-sible for the conversion of all trans-retinol into retinylesters, primarily into retinyl palmitate (Ruiz et al., 1999;Zolfaghari et al., 2002; Batten et al., 2004).
With the exception of Vibrio cholerae and the diaz-otrophic heterocystous cyanobacterium Anabaena,LRAT and its orthologues have been found only ineukaryotes, including plants and in animal viruses(Anantharaman and Aravind, 2003). Phylogenetic analy-sis of the LRAT homologue found in V. cholerae sug-gested that it had been acquired through lateral gene
transfer from a vertebrate source (Anantharaman andAravind, 2003). LRAT belongs to a unique and ancientgene family (Anantharaman and Aravind, 2003; Jahnget al., 2003), and to the N1pC/P60 protein superfamilyof cell-wall peptidases found in various bacterial lin-eages (Anantharaman and Aravind, 2003). Amino acidsequence alignment of the major known LRATsequences in the protein databases to the sequenceidentified in the BLAST search against T. erythraeumIMS101 genome (Fig. 6) revealed similarities in con-served positions, including structural homology with cys-teines. LRAT is the founder member of a unique class ofenzymes, which include class II tumour suppressors,proteins essential for development and putative pro-teases (Jahng et al., 2003). All of these proteins possesscysteine residues. LRAT, which is considered a thiolacyltransferase, contains a cysteine at position 161(marked in Fig. 6 as the first asterisk) and is alsopresent in the Trichodesmium LRAT protein sequence.This cysteine was found to be the essential nucleophilicresidue critical for catalysis (Mondal et al., 2000). Areverse search of Trichodesmium LRAT proteinsequence confirmed the homology to mammalian LRATproteins and to the hypothetical protein Tery_4214(GenBank accession number YP_723688) in theT. erythraeum IMS101 genome. This hypotheticalprotein was found to belong to the putative conservedN1pC/P60 domain. Members of this family are charac-terized by containing a well-conserved NCEHF motif(Fig. 6). In vertebrates, LRAT is important in storage andmobilization of retinol as esters in peripheral tissues and
Fig. 5. The biosynthetic pathway of retinyl palmitate from all trans-b-carotene.
Antioxidant defence in the cyanobacterium Trichodesmium 1903
© 2009 The AuthorsJournal compilation © 2009 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 11, 1897–1908

for generating an intermediate in the synthesis of therhodopsin chromophore in the visual tissues (Ananthara-man and Aravind, 2003). The presence of LRAT ortho-logues in plants suggests that similar retinol processingoccurs also in plants (Anantharaman and Aravind,2003). The evolutionary history as well as the role andfunction of LRAT and the retinyl palmitate biosyntheticpathway in Trichodesmium, or in other cyanobacteria,are presently unknown. In this comparative genomicsstudy, we identified and discussed the possible presenceof LRAT in the Trichodesmium genome, as thesequence homology contained the major domain of thisenzyme, including its catalytic site. As we did not identifythe other domains of this enzyme in the Trichodesmiumgenome, this finding should be considered with cautionand further research is warranted.
Trichodesmium is an ancient cyanobacterium thatflourishes in present-day subtropical and tropical oceanswith characteristic surface blooms extending over thou-sands of kilometres. The exposure to high irradiances,at these latitudes, combined with the concurrent pro-duction of photosynthetically evolved oxygen and (non-
heterocystous) nitrogen fixation, require an exceptionalstrategy to tackle and reduce oxidative stress. We showthat carotenoids, and not antioxidative proteins, providethe bulk of the activity in Trichodesmium’s arsenal ofdefensive compounds against oxidative damage. Byreducing the large investment in energy and resources,typical for protein synthesis and mobilization, Trichodes-mium can maintain an extremely high total antioxidantactivity allowing it to thrive and persist in today’s oxygen-ated surface oceans.
Experimental procedures
Cultures and growth conditions
Trichodesmium sp. IMS101 cultures were grown in YBCIImedium (Chen et al., 1996) at 25°C with a 12:12 light/darkcycle under 80 mmol of quanta m-2 s-1, and constant aerationwith ambient air. Anabaena flos-aquae (strain UTEX 2557)cultures were grown in Bold 3N medium (Starr and Zeikus,1993). Cyanothece sp. WH8904 cultures were grown in Arti-ficial Seawater medium (Goldman and McCarthy, 1978).Prochlorothrix hollandica cultures were grown in BG-11
Fig. 6. Amino acid sequence alignment of partial LRAT protein sequences from Rattus norvegicus (LRAT_RAT, Norway rat, Q9JI61), Xenopustropicalis (LRAT_FROG, Western clawed frog, Q6P884), Danio rerio (LRAT_DANIO, Zebrafish, AAH95753), Bos taurus (LRAT_BOVIN,Domestic cow, Q9BGL2), Homo sapiens (LRAT_HUMAN, Human, O95237), Mus musculus (LRAT_MOUSE, House mouse, Q9JI60) andTrichodesmium erythraeum IMS101 [LRAT_TRICHO, from a BLAST search to T. erythraeum IMS101 complete genome, CP000393(http://genome.jgi-psf.org/finished_microbes/trier/trier.home.html)]. Homologous sequences (black-shaded characters denote identity and greydenotes similarity) highlight similarities in highly conserved positions, including the structural homology with cysteines (indicated withasterisks).
1904 D. Kelman, A. Ben-Amotz and I. Berman-Frank
© 2009 The AuthorsJournal compilation © 2009 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 11, 1897–1908

medium (Rippka et al., 1979). Synechococcus sp. WH7803cultures were grown in F/2 medium (Guillard, 1975).
Extractions
Cells for organic extractions were sampled at midday andfiltered on 47 mm GF/F glass microfibre filters (Whatman,Middlesex, UK) and frozen at -20°C. The filters were thenextracted for 24 h in ethyl acetate at room temperature. Thenon-polar (ethyl-acetate) extracts were then filtered andevaporated to dryness under N2 flow. The cells that remainedon the GF/F filters were further extracted with methanol(MeOH) for 24 h at room temperature. The polar (MeOH)extracts were then filtered and evaporated to dryness with aSpeedVac. At all stages, the extracts were protected fromlight with aluminium foil. The crude extracts were weighedand kept at -20°C for further analysis.
Cells for protein extractions were also sampled at the sametime of day and filtered on 13 mm 0.22 mm Durapore mem-brane filters (Millipore, Billerica, MA, USA), immediatelyfrozen in liquid N2, and then kept at -80°C. For the proteinextraction, the filters were placed in 200 ml of 10 mM Tris–HCl(pH 7.5), vortexed for 1 min and then the filter was removedfrom solution. The samples were sonicated for 20–30 s andcentrifuged for 10 min at 10 000 r.p.m. The supernatant wasthen removed and placed in a new 1.5 ml Eppendorf micro-centrifuge tube. The protease inhibitor phenylmethylsulfonylfluoride (PMSF) was added at a final concentration of 5 mMto the supernatant to prevent the destruction of proteins in thesample, and then vortexed well. The protein extract sampleswere then stored at -80°C for further analysis. Protein con-centration was determined using the BCA (bicinchoninic acid)Protein Assay Kit (Pierce, Rockford, IL, USA) according to themanufacturer’s instructions.
Antioxidant activity assay
The FRAP assay was modified from Benzie and Strain’sprotocol (Benzie and Strain, 1996; 1999). The working FRAPreagent was produced by mixing 300 mM acetate buffer(pH 3.6), 10 mM 2,4,6-tripyridyl-s-triazine (TPTZ) solutionand 20 mM FeCl3·6H2O in a 10:1:1 ratio prior to use andheated to 37°C. The 300 mM acetate buffer was prepared bymixing 3.1 g of sodium acetate trihydrate (C2H3NaO2·3H2O)with 16 ml glacial acetic acid and brought to 1 l with ddH2O.The TPTZ solution was prepared by mixing 10 mM TPTZ with40 mM HCl.
One hundred fifty microlitres of working FRAP reagent wasadded to each well of a 96-well microtiter plate. A blankreading was then taken at 595 nm using a Multiskan RC(Labsystems) plate reader. Twenty microlitres of sample wasthen added to each well in triplicate. For the organic extractsamples, each extract was dissolved in DMSO at a concen-tration of 10 mg ml-1. For the protein extracts, each samplewas taken directly from the Tris–HCl protein extract. Controltests of DMSO or Tris–HCl (with PMSF) were also added.Triplicate samples were added to the FRAP reagent, incu-bated for 8 min at room temperature and read at 595 nm.Triplicate standards of known FeII concentrations were runsimultaneously using concentrations between 50 and
1000 mM of FeSO4·7H2O. A standard curve was plotted andthe FRAP values (in mM) for each sample were determined.As results may vary between plates, a new standard curvewas prepared for each plate.
Bioassay-guided fractionation, isolation andidentification of compounds
To isolate and characterize the antioxidant component/s ofthe cyanobacterium, bioassay-guided fractionation was per-formed on a 10 l full-density culture of Trichodesmiumsp. IMS101. The culture was grown, harvested and extractedas outlined above. The non-polar (ethyl-acetate) crude extractwas subjected to the FRAP antioxidant activity assay asoutlined above. This bioassay was used as a guide throughoutthe fractionation process. The crude extract was fractionatedby solvent partitioning with n-heptane and dichloromethane assolvent systems against aq. MeOH (10–20% H2O). The result-ing fractions were evaporated to dryness and assayed forantioxidant activity. Subsequent fractionation of the activeheptane phase employed liquid chromatography on a Diolbonded silica gel Sep-Pak column (Baker, Phillipsburg, NJ,USA), eluted with solvents of increasing polarity (from 100%n-heptane to 100% ethyl-acetate). The resulting fractions wereevaporated to dryness and assayed for antioxidant activity.
The most active fraction (determined using the FRAPassay) was further fractionated using HPLC. The HPLC wasbased on a Waters HPLC system (Millipore, Marlborough,MA, USA), connected to a Waters 996 photodiode arraydetector, and managed by Waters EMPOWER Chromato-graphy Software. The column used was a 25 cm ¥ 4.6 mm(internal diameter) C18 reverse-phase Vydac 201 TP54column (The Separation Group, Hysperia, CA, USA). Thecolumn was maintained at 30 � 0.2°C in HPLC Column 7955Heater/Chiller (Jones Chromatography, Glamorgan, UK).Elution was performed with an isocratic solvent, HPLCgrade MeOH : acetonitrile (9:1, v/v), at a constant flow of1.0 ml min-1. This HPLC system was used to determine thevarious components of the polar and non-polar crude organicextracts of the cyanobacterium Trichodesmium, as well as thepurified active fraction. This system is well documented forthe simultaneous separation and identification of the differentcarotenoids and their isomers, as well as chlorophylls, retin-ols, their derivatives and tocopherols (Ben-Amotz, 1995;Ben-Amotz and Fishler, 1998).
TLC and DPPH radical-scavenging effect
The radical-scavenging effect of crude extracts and purifiedcompounds were detected on silica gel 60 TLC plates (Merck,Darmstadt, Germany), using a spray reagent composed of a0.2% (w/v) solution of DPPH radical in MeOH (Son et al.,2002; Takamatsu et al., 2003). Plates were kept in the darkunder a stream of N2 for 30 min after spraying and observed.Active compounds are observed as bright yellow spotsagainst the purple background.
Acknowledgements
We thank L. Sherman, A. Post and O. Prasil for generous giftsof culture inocula for Cyanothece sp. WH8904, Synechococ-
Antioxidant defence in the cyanobacterium Trichodesmium 1905
© 2009 The AuthorsJournal compilation © 2009 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 11, 1897–1908

cus sp. WH7803 and P. hollandica. We wish to thank I. Mehrfor assistance with the HPLC work, R. Persky for the MassSpectrometry analysis, O. Beja and B. Shomer for advice inthe bioinformatics and M. Hyams and S. Aharoni for technicalassistance. The helpful comments made by the three anony-mous reviewers are greatly appreciated. D.K. was supportedby the Soref-Kolman Foundation Postdoctoral Fellowship.
References
Altschul, S.F., Gish, W., Miller, W., Myers, E.W., and Lipman,D.J. (1990) Basic local alignment search tool. J Mol Biol215: 403–410.
Anantharaman, V., and Aravind, L. (2003) Evolutionaryhistory, structural features and biochemical diversity ofthe NlpC/P60 superfamily of enzymes. Genome Biol 4:R11.
Andrews, S.S., Caron, S., and Zafiriou, O.C. (2000) Photo-chemical oxygen consumption in marine waters: a majorsink for colored dissolved organic matter? Limnol Ocean-ogr 45: 267–277.
Araujo, K.G.L., Domingues, J.R., Srur, A., and da Silva,A.J.R. (2006) Production of antioxidants by AnaboenaPCC 7119 and evaluation of their protecting activity againstoxidation of soybean oil. Food Biotechnol 20: 65–77.
Asada, K. (2000) The water-water cycle as alternative photonand electron sinks. Philos Trans R Soc Lond B Biol Sci 355:1419–1430.
Batten, M.L., Imanishi, Y., Maeda, T., Tu, D.C., Moise, A.R.,Bronson, D., et al. (2004) Lecithin-retinol acyltransferase isessential for accumulation of all-trans-retinyl esters in theeye and in the liver. J Biol Chem 279: 10422–10432.
Ben-Amotz, A. (1995) Simultaneous profiling and identifi-cation of carotenoids, retinols, and tocopherols byhigh-performance liquid-chromatography equipped with3-dimensional photodiode-array detection. J Liq Chro-matogr 18: 2813–2825.
Ben-Amotz, A., and Fishler, R. (1998) Analysis of carotenoidswith emphasis on 9-cis beta-carotene in vegetables andfruits commonly consumed in Israel. Food Chem 62: 515–520.
Benzie, I.F.F., and Strain, J.J. (1996) The ferric reducingability of plasma (FRAP) as a measure of ‘antioxidantpower’: the FRAP assay. Anal Biochem 239: 70–76.
Benzie, I.F.F., and Strain, J.J. (1999) Ferric reducing antioxi-dant power assay: Direct measure of total antioxidantactivity of biological fluids and modified version for simul-taneous measurement of total antioxidant power andascorbic acid concentration. Meth Enzymol 299: 15–27.
Bergman, B., Gallon, J.R., Rai, A.N., and Stal, L.J. (1997) N2
fixation by non-heterocystous cyanobacteria. FEMS Micro-biol Rev 19: 139–185.
Berman-Frank, I., Lundgren, P., Chen, Y.B., Kupper, H.,Kolber, Z., Bergman, B., and Falkowski, P. (2001) Segre-gation of nitrogen fixation and oxygenic photosynthesis inthe marine cyanobacterium Trichodesmium. Science 294:1534–1537.
Berman-Frank, I., Bidle, K.D., Haramaty, L., and Falkowski,P.G. (2004) The demise of the marine cyanobacterium,Trichodesmium spp., via an autocatalysed cell deathpathway. Limnol Oceanogr 49: 997–1005.
Berman-Frank, I., Rosenberg, G., Levitan, O., Haramaty, L.,and Mari, X. (2007) Coupling between autocatalytic celldeath and transparent exopolymeric particle production inthe marine cyanobacterium Trichodesmium. Environ Micro-biol 9: 1415–1422.
Beutner, S., Bloedorn, B., Frixel, S., Blanco, I.H., Hoffmann,T., Martin, H.D., et al. (2001) Quantitative assessment ofantioxidant properties of natural colorants and phytochemi-cals: carotenoids, flavonoids, phenols and indigoids. Therole of beta-carotene in antioxidant functions. J Sci FoodAgric 81: 559–568.
Blough, N.V., and Zepp, R.G. (1995) Reactive oxygenspecies in natural waters. In Active Oxygen in Chemistry.Foote, C.S., Valentine, J.S., Greenberg, A, and Liebman,J.F (eds). New York, NY, USA: Blackie Academic and Pro-fessional, pp. 280–333.
Britton, G. (1995) Structure and properties of carotenoids inrelation to function. FASEB J 9: 1551–1558.
Burton, G.W., and Ingold, K.U. (1984) Beta-carotene- anunusual type of lipid antioxidant. Science 224: 569–573.
Cahyana, A.H., Shuto, Y., and Kinoshita, Y. (1992)Pyropheophytin a as an antioxidative substance from themarine alga, Arame (Eisenia bicyclis). Biosci BiotechnolBiochem 56: 1533–1535.
Capone, D.G., Zehr, J.P., Paerl, H.W., Bergman, B.,and Carpenter, E.J. (1997) Trichodesmium, a globallysignificant marine cyanobacterium. Science 276: 1221–1229.
Chen, Y.B., Zehr, J.P., and Mellon, M. (1996) Growth andnitrogen fixation of the diazotrophic filamentous nonhetero-cystous cyanobacterium Trichodesmium sp. IMS 101 indefined media: evidence for a circadian rhythm. J Phycol32: 916–923.
Chen, Y.B., Dominic, B., Mellon, M.T., and Zehr, J.P. (1998)Circadian rhythm of nitrogenase gene expression in thediazotrophic filamentous nonheterocystous Cyanobacte-rium Trichodesmium sp. strain IMS101. J Bacteriol 180:3598–3605.
Chen, Y.B., Dominic, B., Zani, S., Mellon, M.T., and Zehr,J.P. (1999) Expression of photosynthesis genes in relationto nitrogen fixation in the diazotrophic filamentous nonhet-erocystous cyanobacterium Trichodesmium sp. IMS 101.Plant Mol Biol 41: 89–104.
Conn, P.F., Schalch, W., and Truscott, T.G. (1991) Thesinglet oxygen and carotenoid interaction. J PhotochemPhotobiol B-Biol 11: 41–47.
Cunningham, K.A., and Capone, D.G. (1992) Supoeroxidedismutase as a protective enzyme against oxygen toxicity:an overview and initial studies in Trichodesmium. InMarine Pelagic Cyanobacteria: Trichodesmium and OtherDiazotrophs. Carpenter, E.J., Capone, D.G., and Reuter,J.G (eds). Dordecht, the Netherlands: Kluwer AcademicPublishers, pp. 331–341.
Davis, C.S., and McGillicuddy, D.J. (2006) Transatlanticabundance of the N2-fixing colonial cyanobacteriumTrichodesmium. Science 312: 1517–1520.
Edge, R., McGarvey, D.J., and Truscott, T.G. (1997) Thecarotenoids as anti-oxidants – a review. J Photochem Pho-tobiol B-Biol 41: 189–200.
Eitinger, T. (2004) In vivo production of active nickel super-oxide dismutase from Prochlorococcus marinus MIT9313
1906 D. Kelman, A. Ben-Amotz and I. Berman-Frank
© 2009 The AuthorsJournal compilation © 2009 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 11, 1897–1908

is dependent on its cognate peptidase. J Bacteriol 186:7821–7825.
Falkowski, P.G., and Raven, J.A. (2007) Aquatic Photosyn-thesis, 2nd edn. Princeton, NJ, USA: Princeton UniversityPress.
Fridovich, I. (1998) Oxygen toxicity: a radical explanation.J Exp Biol 201: 1203–1209.
Gallon, J.R. (1992) Reconciling the Incompatible – N2 Fixa-tion and O2. New Phytol 122: 71–609.
Gallon, J.R. (2001) N2 fixation in phototrophs: adaptation to aspecialized way of life. Plant Soil 230: 39–48.
Goldman, J.C., and McCarthy, J.J. (1978) Steady-stategrowth and ammonium uptake of a fast-growing marinediatom. Limnol Oceanogr 23: 695–703.
Guillard, R.R.L. (1975) Culture of phytoplankton for feedingmarine invertebrates. In Culture of Marine InvertebrateAnimals. Smith, W.L., and Chanley, M.H. (eds). New York,NY, USA: Plenum Press, pp. 26–60.
Halliwell, B. (1999) Antioxidant defence mechanisms: fromthe beginning to the end (of the beginning). Free Rad Res31: 261–272.
Halliwell, B., and Gutteridge, J.M.C. (1990) The antioxidantsof human extracellular fluids. Arch Biochem Biophys 280:1–8.
Halliwell, B., and Gutteridge, J.M.C. (1995) The definition andmeasurement of antioxidants in biological systems. FreeRad Biol Med 18: 125–126.
Halliwell, B., and Gutteridge, J.M.C. (1999) Free Radicals inBiology and Medicine. Oxford, UK: Oxford UniversityPress.
Halvorsen, B.L., Holte, K., Myhrstad, M.C.W., Barikmo, I.,Hvattum, E., Remberg, S.F., et al. (2002) A systematicscreening of total antioxidants in dietary plants. J Nutr 132:461–471.
Jahng, W.J., Xue, L.L., and Rando, R.R. (2003) Lecithinretinol acyltransferase is a founder member of a novelfamily of enzymes. Biochemistry 42: 12805–12812.
Kana, T.M. (1992) Oxygen cycling in cyanobacteria withspecific reference to oxygen protection in Trichodesmiumspp. In Marine Pelagic Cyanobacteria: Trichodesmiumand Other Diazotrophs. Carpenter, E.J., Capone, D.G.,and Reuter, J.G. (eds). Dordecht, the Netherlands: KluwerAcademic Publishers, pp. 29–41.
Kana, T.M. (1993) Rapid oxygen cycling in Trichodesmiumthiebautii. Limnol Oceanogr 38: 18–24.
Kobayashi, M., Kakizono, T., Nishio, N., Nagai, S., Kurimura,Y., and Tsuji, Y. (1997) Antioxidant role of astaxanthin inthe green alga Haematococcus pluvialis. Appl MicrobiolBiotechnol 48: 351–356.
Kupper, H., Ferimazova, N., Setlik, I., and Berman-Frank, I.(2004) Traffic lights in Trichodesmium: regulation of photo-synthesis for nitrogen fixation studied by chlorophyllfluorescence kinetic microscopy. Plant Physiol 135: 2120–2133.
Li, H.B., Cheng, K.W., Wong, C.C., Fan, K.W., Chen, F., andJiang, Y. (2007) Evaluation of antioxidant capacity andtotal phenolic content of different fractions of selectedmicroalgae. Food Chem 102: 771–776.
Liebler, D.C. (1993) Antioxidant reactions of carotenoids. AnnNew York Acad Sci 691: 20–31.
Lin, S.J., Henze, S., Lundgren, P., Bergman, B., and Carpen-
ter, E.J. (1998) Whole-cell immunolocalization of nitroge-nase in marine diazotrophic cyanobacteria, Trichodesmiumspp. Appl Environ Microbiol 64: 3052–3058.
Maeda, H., and DellaPenna, D. (2007) Tocopherol functionsin photosynthetic organisms. Curr Opin Plant Biol 10: 260–265.
Mallick, N., and Mohn, F.H. (2000) Reactive oxygen species:response of algal cells. J Plant Physiol 157: 183–193.
Milligan, A.J., Berman-Frank, I., Gerchman, Y., Dismukes,G.C., and Falkowski, P.G. (2007) Light-dependent oxygenconsumption in nitrogen-fixing cyanobacteria plays a keyrole in nitrogenase protection. J Phycol 43: 845–852.
Mondal, M.S., Ruiz, A., Bok, D., and Rando, R.R. (2000)Lecithin retinol acyltransferase contains cysteine residuesessential for catalysis. Biochemistry 39: 5215–5220.
Nakai, M., Kageyama, N., Nakahara, K., and Miki, W. (2006)Phlorotannins as radical scavengers from the extract ofSargassum ringgoldianum. Mar Biotechnol 8: 409–414.
Noctor, G., and Foyer, C.H. (1998) Ascorbate and glu-tathione: keeping active oxygen under control. Annu RevPlant Physiol Plant Mol Biol 49: 249–279.
Ozben, T. (2007) Oxidative stress and apoptosis: impact oncancer therapy. J Pharm Sci 96: 2181–2196.
Paerl, H.W., and Bebout, B.M. (1992) Oxygen dynamicsin Trichodesmium spp. aggregates. In Marine PelagicCyanobacteria: Trichodesmium and Other Diazotrophs.Carpenter, E.J., Capone, D.G., and Reuter, J.G. (eds).Dordecht, the Netherlands: Kluwer Academic Publishers,pp. 43–59.
Potterat, O. (1997) Antioxidants and free radical scavengersof natural origin. Curr Org Chem 1: 415–440.
Prior, R.L., and Cao, G.H. (1999) In vivo total antioxidantcapacity: comparison of different analytical methods. FreeRad Biol Med 27: 1173–1181.
Priya, B., Premanandh, J., Dhanalakshmi, R.T., Seethalak-shmi, T., Uma, L., Prabaharan, D., and Subramanian, G.(2007) Comparative analysis of cyanobacterial superoxidedismutases to discriminate canonical forms. BMC Genom-ics 8: 435.
Rippka, R., Deruelles, J., Waterbury, J.B., Herdman, M., andStanier, R.Y. (1979) Generic assignments, strain historiesand properties of pure cultures of cyanobacteria. J GenlMicrobiol 111: 1–61.
Ruiz, A., Winston, A., Lim, Y.H., Gilbert, B.A., Rando, R.R.,and Bok, D. (1999) Molecular and biochemical character-ization of lecithin retinol acyltransferase. J Biol Chem 274:3834–3841.
Sanjeev Kumar, P., Vasudevan, D.M., and Jayanthi Bai, N.(1981) Antioxidant role of retinyl-palmitate on carbontetra-chloride toxicity. Acta Vitaminol Enzymol 3: 214–218.
Schafer, L., Vioque, A., and Sandmann, G. (2005) Functionalin situ evaluation of photo synthesis-protecting carotenoidsin mutants of the cyanobacterium SynechocystisPCC6803. J Photochem Photobiol B-Biol 78: 195–201.
Sies, H. (1991) Oxidative Stress: Oxidants and Antioxidants.London, UK: Academic Press.
Sies, H., and Stahl, W. (1995) Vitamin-E and vitamin-C, beta-carotene, and other carotenoids as antioxidants. Am J ClinNutr 62: S1315–S1321.
Son, B.W., Kim, J.C., Choi, H.D., and Kang, J.S. (2002) Aradical scavenging farnesylhydroquinone from a marine-
Antioxidant defence in the cyanobacterium Trichodesmium 1907
© 2009 The AuthorsJournal compilation © 2009 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 11, 1897–1908

derived fungus Penicillium sp. Arch Pharmacal Res 25:77–79.
Soni, B., Trivedi, U., and Madamwar, D. (2008) A novelmethod of single step hydrophobic interaction chromatog-raphy for the purification of phycocyanin from Phormidiumfragile and its characterization for antioxidant property.Bioresour Technol 99: 188–194.
Starr, R.C., and Zeikus, J.A. (1993) UTEX – the culturecollection of algae at the University of Texas at Austin 1993list of cultures. J Phycol 29: 1–106.
Takamatsu, S., Hodges, T.W., Rajbhandari, I., Gerwick,W.H., Hamann, M.T., and Nagle, D.G. (2003) Marinenatural products as novel antioxidant prototypes. J NatProd 66: 605–608.
Telfer, A., Dhami, S., Bishop, S.M., Phillips, D., and Barber, J.(1994) Beta-carotene quenches singlet oxygen formed byisolated photosystem-II reaction centers. Biochemistry 33:14469–14474.
Tesoriere, L., Darpa, D., Re, R., and Livrea, M.A. (1997)Antioxidant reactions of all-trans retinol in phospholipidbilayers: effect of oxygen partial pressure, radical fluxes,and retinol concentration. Arch Biochem Biophys 343:13–18.
Voelker, B.M., Sedlak, D.L., and Zafiriou, O.C. (2000) Chem-istry of superoxide radical in seawater: reactions withorganic Cu complexes. Environ Sci Technol 34: 1036–1042.
Watanabe, N., Yamamoto, K., Ihshikawa, H., Yagi, A.,Sakata, K., Brinen, L.S., and Clardy, J. (1993) Newchlorophyll-a-related compounds isolated as antioxidantsfrom marine bivalves. J Nat Prod 56: 305–317.
Wolfe-Simon, F., Grzebyk, D., Schofield, O., and Falkowski,P.G. (2005) The role and evolution of superoxide dismu-tases in algae. J Phycol 41: 453–465.
Yamamoto, K., Sakata, K., Watanabe, N., Yagi, A., Brinen,L.S., and Clardy, J. (1992) Chlorophyllonic acid a methyl-ester, a new chlorophyll-a related compound isolated as anantioxidant from short-necked clam, Ruditapes philippi-narum. Tetrahedron Lett 33: 2587–2588.
Yan, X.J., Chuda, Y., Suzuki, M., and Nagata, T. (1999)Fucoxanthin as the major antioxidant in Hijikia fusiformis, acommon edible seaweed. Biosci Biotechnol Biochem 63:605–607.
Zhao, W.X., Ye, Z., and Zhao, J.D. (2007) RbrA, a cyanobac-terial rubrerythrin, functions as a FNR-dependent peroxi-dase in heterocysts in protection of nitrogenase fromdamage by hydrogen peroxide in Anabaena sp PCC 7120.Mol Microbiol 66: 1219–1230.
Zolfaghari, R., Wang, Y.P., Chen, Q.Y., Sancher, A., andRoss, A.C. (2002) Cloning and molecular expressionanalysis of large and small lecithin: retinol acyltransferasemRNAs in the liver and other tissues of adult rats. BiochemJ 368: 621–631.
1908 D. Kelman, A. Ben-Amotz and I. Berman-Frank
© 2009 The AuthorsJournal compilation © 2009 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 11, 1897–1908