biphasic effects of copper on neurotransmission in rat hippocampal neurons
TRANSCRIPT

Copper is an essential transition metal that participates inthe regulation of brain physiology, being a key structuralcomponent of various proteins and a co-factor for enzymesthat are critical for brain function, including enzymesinvolved in antioxidant defense and cellular respiration(Mathie et al. 2006). More recently, some reports havedescribed the effect of copper at the synaptic level, where itmodulates complex parameters such as LTP (Goldschmithet al. 2005; Leiva et al. 2009) and receptor pharmacology
Received April 13, 2011; revised manuscript received July 31, 2011;accepted August 2, 2011.Address correspondence and reprint requests to Dr Carlos Opazo,
Departamento de Fisiologıa, Facultad de Ciencias Biologicas, Univers-idad de Concepcion, P.O. Box 160-C, Concepcion Chile.E-mail: [email protected] used: AD, Alzheimer’s Disease; AMPA, a-amino-
3-hydroxy-5-methyl-4-isoxazolepropionic acid; APV, DL-2-amino-5-phosphonovaleric acid; CNQX, 6-cyano-7-nitroquinoxaline-2,3-dione;GABA, c-aminobutyric acid.
*Laboratorio de Neurobiometales Departamento de Fisiologıa, Facultad de Ciencias Biologicas,
Universidad de Concepcion, Chile
�Laboratorio de Neurofisiologıa, Departamento de Fisiologıa, Facultad de Ciencias Biologicas,
Universidad de Concepcion, Chile
�Facultad de Ciencias de la Salud, Universidad San Sebastian, Concepcion, Chile
§Centro de Investigaciones Biomedicas Facultad de Ciencias Biologicas, Universidad Andres Bello,
Santiago, Chile
Abstract
The importance of copper in the CNS is well documented, but
the mechanisms related to its brain functions are poorly
understood. Copper is released at the synaptic cleft, where it
may modulate neurotransmission. To understand the func-
tional impact of copper on the neuronal network, we have
analyzed the synaptic activity of primary rat hippocampal
neurons by using different approaches including whole cell
patch clamp, recording of calcium transients, immunofluo-
rescence and western blot. Here, we show that copper
produces biphasic changes in neurotransmission. When
copper is acutely applied to the plate it blocks neurotrans-
mission. Interestingly, when it is applied for 3 h to hippo-
campal neurons it mainly increases the frequency and
amplitude of a-amino-3-hydroxy-5-methyl-4-isoxazoleprop-
ionic acid (AMPA)ergic currents (control: 0.21 ± 0.05 Hz/22.9
± 1.3 pA; copper: 0.68 ± 0.16 Hz/30.5 ± 2.5 pA), intracellular
calcium transients (control: 0.05 ± 0.013 Hz; copper:
0.11 ± 0.02 Hz) and evoked AMPA currents (control: EC50
8.3 ± 0.5 lM; copper: EC50 2.9 ± 0.2 lM). Moreover, our
results suggest that copper increases GluA1 subunit levels of
the AMPA receptor through the anchorage of AMPA receptors
to the plasma membrane as a result of PSD-95 accumulation.
We also found that copper-treated neurons displayed an
undistinguishable neurotransmission to control neurons after
24 h of treatment, indicating that changes in neurotransmis-
sion induced by copper at 3 h of incubation are homeostati-
cally regulated after long-term exposure to the metal.
Together, our data reveal an unexpected biphasic effect of
copper on neurotransmission, which may be relevant to
understand the effects of this ion in brain diseases that display
copper dyshomeostasis such as that observed in Alzheimer’s
disease (AD).
Keywords: AMPA, calcium, copper, GABA, neurotransmis-
sion.
J. Neurochem. (2011) 10.1111/j.1471-4159.2011.07417.x
JOURNAL OF NEUROCHEMISTRY | 2011 doi: 10.1111/j.1471-4159.2011.07417.x
� 2011 The AuthorsJournal of Neurochemistry � 2011 International Society for Neurochemistry, J. Neurochem. (2011) 10.1111/j.1471-4159.2011.07417.x 1

(Weiser and Wienrich 1996; Vlachova et al. 1996; Sharonovaet al. 1998). The average copper concentration in the CSF hasbeen estimated to be approximately 70 lM, whereas thenormal extracellular copper concentration in the brain is0.2–1.7 lM (Stuerenburg 2000; Tarohda et al. 2004).Nevertheless, these levels may be exceeded in the synapticcleft under either normal or pathological conditions becausecopper can be released to the extracellular space fromsynaptosomes (Kardos et al. 1989). In fact, copper is releasedin a calcium dependent manner under high potassium (Kardoset al. 1989) and chemical stimulation driven by NMDAactivation (Schlief et al. 2005). Moreover, micromolar con-centration of copper (400 lM) is present in senile plaques inAD brains (Lovell et al. 1998), which could be a source ofcopper for the neurons surrounding these pathological struc-tures. In addition, there are several reports that support a clearlink between brain copper dyshomeostasis and AD (Bush2003).
Under both scenarios, physiological or pathological, post-synaptic receptors may be targets for copper. Indeed, it hasbeen observed that acute applications of copper rapidlyblocks GABA and NMDA neurotransmission on rat olfac-tory bulb neurons (Trombley and Shepherd 1996), and alsoinhibits AMPAergic neurotransmission on rat cortical neu-rons (Weiser and Wienrich 1996). Indeed, Cu2+ inhibitsAMPA/kainate receptors expressed by rat cortical neurons inculture, with an IC50 of 4.3 ± 0.6 lM (with 100 lM kainate,holding potential )60 mV) (Weiser and Wienrich 1996);thus, suggesting that copper has an acute modulatory role onthe AMPA receptor in neurotransmission. To date, the effectsthat copper may exerts on neurotransmission for prolongedperiods of incubation remain unknown and therefore here wehave studied the neurotransmission of primary cultures ofhippocampal neurons exposed to low micromolar concen-trations of copper at different times (0, 3 and 24 h).Remarkably our results suggest that copper after 3 h ofincubation, induces an increase in synaptic activity throughAMPA receptor at the post-synaptic density via a mechanismthat involves the accumulation of PSD95 protein. Thus, ourfindings represent a novel mechanism for the action ofcopper, which may have implication for neurophysiologyand neuropathology of CNS.
Materials and methods
Primary cultures of rat hippocampal neuronsHippocampal neurons were obtained from 18-day pregnantSprague-Dawley rats and maintained for 10–14 days in vitro(DIV) as previously described (Aguayo and Pancetti 1994). Animalswere obtained from the animal house of Catholic University ofChile (Santiago, Chile). All animals were handled in strictaccordance with NIH recommendations and approved by theappropriate committee at the University of Concepcion (Concep-cion, Chile).
ElectrophysiologyExperiments were performed in the ‘whole-cell’ configuration.Recording pipettes were pulled from borosilicate glass (WPI,Sarasota, FL, USA) in a horizontal puller (Sutter Instruments,Novato, CA, USA). Membrane currents were measured using anAxopatch-200B amplifier (Molecular Devices, LLC, Sunnyvale,CA, USA) and an inverted microscope (Eclipse TE200-U; NikonInstruments Inc., Tokyo, Japan). Data were collected, stored andanalyzed using a data acquisition system card (Molecular Devices,LLC) and the pClamp9 software (Molecular Devices, LLC). Forsynaptic activity records, data were analyzed using the Minianalysissoftware, obtaining the frequency, amplitude and decay time of therecords. All experiments were performed at room temperature (20–25�C) using a membrane potential of )60 mV. Data are given asmeans ± SEM and are obtained from more than five experiments.
Solutions and drugsThe intracellular medium contained (in mM): 120 KCl, 2 MgCl2,2 ATP-Na2, 10 BAPTA, 0.5 GTP and 10 HEPES (pH 7.4). Theextracellular medium contained (in mM): 150 NaCl, 5.4 KCl, 2CaCl2, 1 MgCl2, 10 glucose and 10 HEPES (pH 7.4). Recordswere performed in the presence of 100 nM TTX to inhibit actionpotentials as described (Sepulveda et al. 2009). AMPA currentswere isolated with bicuculline (5 lM), DL-2-amino-5-phosphono-valeric acid (APV) (5 lM) and strychnine (1 lM). For GABAcurrents we used 6-cyano-7-nitroquinoxaline-2,3-dione (CNQX)(4 lM), APV (5 lM) and strychnine (1 lM).
ImmunofluorescenceHippocampal neurons plated in 35 mm dish were washed inphosphate-buffered saline (PBS) (pH 7.4) and fixed with coldmethanol ()20�C) for 10 min. Then, the dish was washed again inPBS and neurons were permeabilized and blocked for 30 min withPBST (PBS + triton 0.1%) and bovine serum albumin 10%. Then,cells were incubated with the following primary antibodies for 16 h:anti-MAP2, 1 : 600 (Santa Cruz Biotechnology, CA, USA); anti-GluA1, 1 : 200 (Santa Cruz Biotechnology); anti-GABAA a-1subunit, 1 : 200 (Neuromab, CA, USA); anti-PSD95, 1 : 200(NeuroMab); and anti-GluA2, 1 : 200 (Chemicon, Temecula, CA,USA). Secondary antibodies conjugated with FITC, Cy3 and Cy5were used for fluorescent staining (Jackson ImmunoResearchLaboratories, PA, USA). All of them were used at 1 : 200 for 2 h.Finally, samples were mounted in fluorescent mounting medium(Dako, CA, USA) and images were obtained under a Nikon Eclipseconfocal microscope (Nikon Instruments Inc.) The immunoreactiv-ity of the different synaptic proteins was quantified at the primaryprocesses with the aid of the ImageJ software (NIH). Fluorescentsignal was quantified as relative units using a region of interest of10 lm of length. To study the immunoreactivity of GluA1 subunitsin non-permeabilized neurons, the neuronal primary cultures werefixed with paraformaldehyde (4%) for 10 min, as previouslyreported (Noel et al. 1999). We confirm the non-permeabilizedstate of the neurons by negative immunostaining with MAP2antibody.
Western blotsEqual amounts of proteins were separated by 10–12% sodiumdodecyl sulfate–polyacrylamide gel electrophoresis as previously
Journal of Neurochemistry � 2011 International Society for Neurochemistry, J. Neurochem. (2011) 10.1111/j.1471-4159.2011.07417.x� 2011 The Authors
2 | C. Peters et al.

described (Avila et al. 2010). Protein bands were transferred ontonitrocellulose membranes, blocked with 5% milk and incubated withthe following primary antibodies: anti-a-tubulin, 1 : 5000 (Sigma,St Louis, MO, USA); anti-GluA1, 1 : 500 (extracellular N-terminalepitope; Santa Cruz Biotechnology); anti-GABAA a-1 subunit,1 : 500 (Neuromab); anti-PSD95 1 : 1000 (Neuromab); b-actin,1 : 2000 (Santa Cruz Biotechnology); and anti-Ctr1, 1 : 500 (kindlyprovided by Professor Dennis Thiele and Dr. Yasuhiro Nose).Immunoreactive bands were detected with secondary antibodiesconjugated with HRP (Santa Cruz Biotechnology) and werevisualized with ECL plus western blotting detection system(PerkinElmer, MA, USA).
Analysis of intracellular calcium transientsNeurons were loaded with Fluo-4 AM (1 lM in pluronic acid/dimethylsulfoxide; Molecular Probes, Eugene, OR, USA) for30 min at 37�C and then washed twice with external solution asdescribed above and incubated for 30 min at 37�C. Neurons weremounted in a perfusion chamber that was placed on the stage of aninverted fluorescent microscope (Eclipse TE), equipped with axenon lamp and a 40· objective (22–24�C). Cells were subse-quently illuminated for 200 ms using a computer-controlledLambda 10-2 filter wheel (Sutter Instruments) and regions ofinterest were simultaneously selected on neuronal somata contain-ing Fluo-4 fluorescence (Ex:Em;480 : 510 nm) in a field havingusually more than 10 cells. Images were collected at 2–5 s intervalsduring a continuous 5-min period of recording with a 12-bit cooled
SensiCam camera (PCO, Kelheim, Germany). Finally, calciumtransients, as defined by their TTX sensitivity (Gu et al. 1994),were acquired and analyzed off line with Axon InstrumentsWorkbench 2.2 software.
Results
Copper increases synaptic activity in primary cultures of rathippocampal neuronsThe effect of copper on synaptic activity has not beendescribed in detail to date. So far, it is known that copperinhibits AMPAergic activity in cortical neurons when it isacutely applied to neurons by direct perfusion (Weiser andWienrich 1996). In fact, AMPA evoked currents inhippocampal neurons in the presence of copper (acuteapplication) were decreased compared with the controlneurons treated with AMPA alone (Fig. 1a). On theseprimary neurons copper inhibited AMPA currents with anIC50 22.9 ± 3.8 lM (Fig. 1b). Moreover, the synapticactivity decreases in the presence of copper (Fig. 1c–e).These results are in agreement with previous reports thatdescribed the inhibitory effect of copper on neurotransmis-sion when it is applied acutely to the cortical neurons(Weiser and Wienrich 1996). To evaluate if these changeswere maintained after hours of exposure, rat primary
Fig. 1 Acute application of copper inhibits AMPA evoked currents and
decreases synaptic activity in rat hippocampal neurons. (a) Repre-
sentative traces of AMPA evoked currents in the absence or presence
of copper (up to 500 lM). (b) The graph summarizes the inhibitory
effect of copper on AMPA evoked currents (IC50 = 22 ± 3.8 lM). (c)
Typical miniature synaptic currents of rat hippocampal neurons treated
acutely (perfusion) with copper 10 lM. Graphs summarize data ob-
tained in these experiments with CuCl2 (10 lM): (d) frequency (Hz);
(e) amplitude (pA). The bars are mean ± SEM from at least six
neurons (*p < 0.05, **p < 0.01).
� 2011 The AuthorsJournal of Neurochemistry � 2011 International Society for Neurochemistry, J. Neurochem. (2011) 10.1111/j.1471-4159.2011.07417.x
Copper enhances CNS neurotransmission | 3
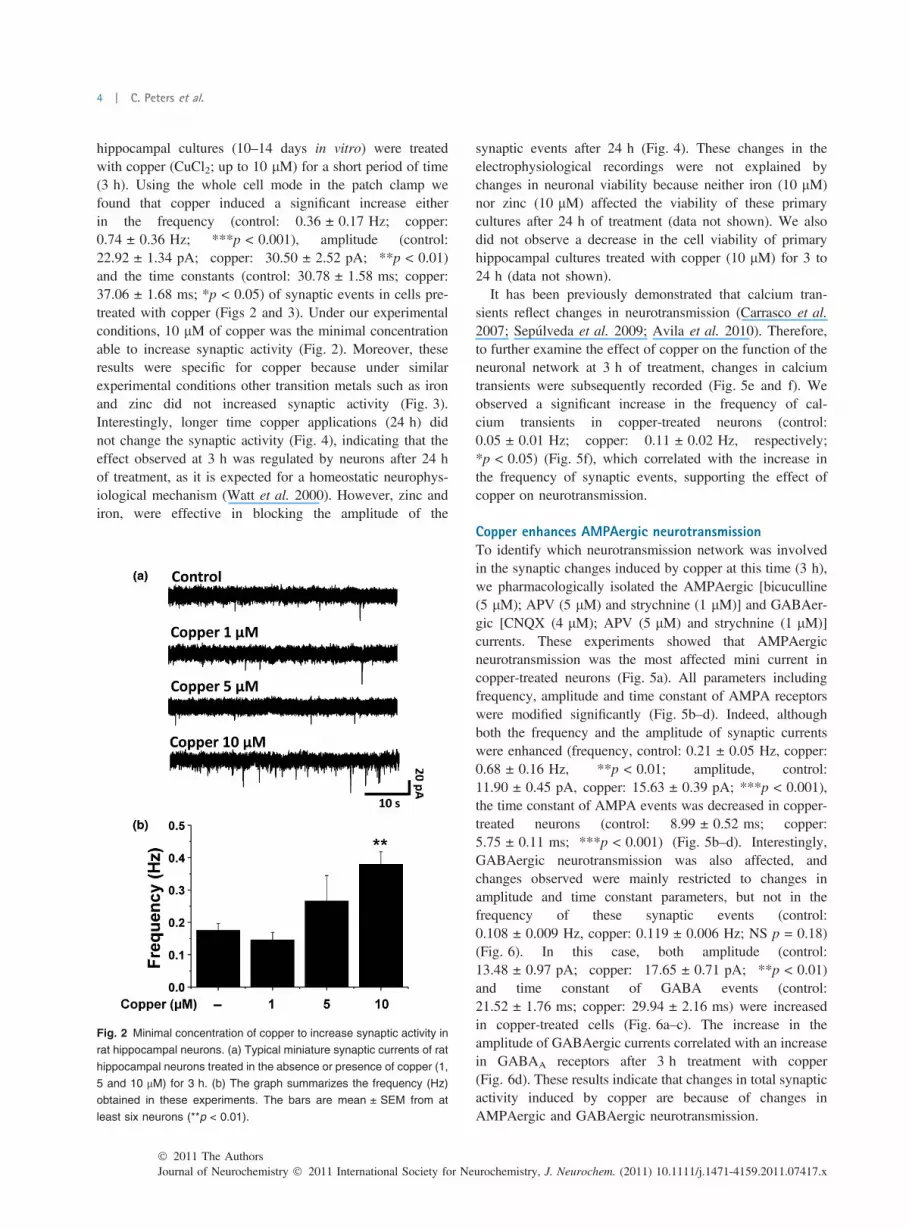
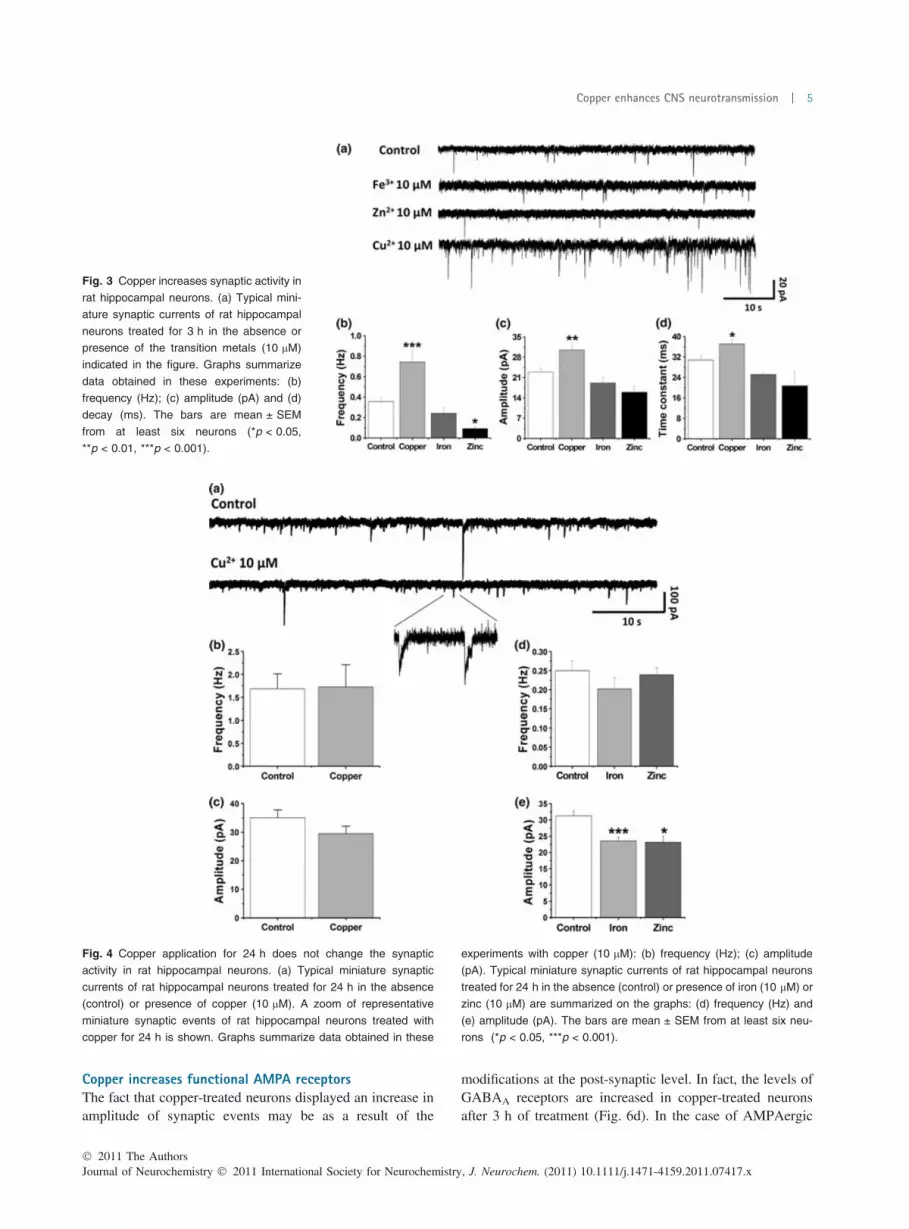
hippocampal cultures (10–14 days in vitro) were treatedwith copper (CuCl2; up to 10 lM) for a short period of time(3 h). Using the whole cell mode in the patch clamp wefound that copper induced a significant increase eitherin the frequency (control: 0.36 ± 0.17 Hz; copper:0.74 ± 0.36 Hz; ***p < 0.001), amplitude (control:22.92 ± 1.34 pA; copper: 30.50 ± 2.52 pA; **p < 0.01)and the time constants (control: 30.78 ± 1.58 ms; copper:37.06 ± 1.68 ms; *p < 0.05) of synaptic events in cells pre-treated with copper (Figs 2 and 3). Under our experimentalconditions, 10 lM of copper was the minimal concentrationable to increase synaptic activity (Fig. 2). Moreover, theseresults were specific for copper because under similarexperimental conditions other transition metals such as ironand zinc did not increased synaptic activity (Fig. 3).Interestingly, longer time copper applications (24 h) didnot change the synaptic activity (Fig. 4), indicating that theeffect observed at 3 h was regulated by neurons after 24 hof treatment, as it is expected for a homeostatic neurophys-iological mechanism (Watt et al. 2000). However, zinc andiron, were effective in blocking the amplitude of the
synaptic events after 24 h (Fig. 4). These changes in theelectrophysiological recordings were not explained bychanges in neuronal viability because neither iron (10 lM)nor zinc (10 lM) affected the viability of these primarycultures after 24 h of treatment (data not shown). We alsodid not observe a decrease in the cell viability of primaryhippocampal cultures treated with copper (10 lM) for 3 to24 h (data not shown).
It has been previously demonstrated that calcium tran-sients reflect changes in neurotransmission (Carrasco et al.2007; Sepulveda et al. 2009; Avila et al. 2010). Therefore,to further examine the effect of copper on the function of theneuronal network at 3 h of treatment, changes in calciumtransients were subsequently recorded (Fig. 5e and f). Weobserved a significant increase in the frequency of cal-cium transients in copper-treated neurons (control:0.05 ± 0.01 Hz; copper: 0.11 ± 0.02 Hz, respectively;*p < 0.05) (Fig. 5f), which correlated with the increase inthe frequency of synaptic events, supporting the effect ofcopper on neurotransmission.
Copper enhances AMPAergic neurotransmissionTo identify which neurotransmission network was involvedin the synaptic changes induced by copper at this time (3 h),we pharmacologically isolated the AMPAergic [bicuculline(5 lM); APV (5 lM) and strychnine (1 lM)] and GABAer-gic [CNQX (4 lM); APV (5 lM) and strychnine (1 lM)]currents. These experiments showed that AMPAergicneurotransmission was the most affected mini current incopper-treated neurons (Fig. 5a). All parameters includingfrequency, amplitude and time constant of AMPA receptorswere modified significantly (Fig. 5b–d). Indeed, althoughboth the frequency and the amplitude of synaptic currentswere enhanced (frequency, control: 0.21 ± 0.05 Hz, copper:0.68 ± 0.16 Hz, **p < 0.01; amplitude, control:11.90 ± 0.45 pA, copper: 15.63 ± 0.39 pA; ***p < 0.001),the time constant of AMPA events was decreased in copper-treated neurons (control: 8.99 ± 0.52 ms; copper:5.75 ± 0.11 ms; ***p < 0.001) (Fig. 5b–d). Interestingly,GABAergic neurotransmission was also affected, andchanges observed were mainly restricted to changes inamplitude and time constant parameters, but not in thefrequency of these synaptic events (control:0.108 ± 0.009 Hz, copper: 0.119 ± 0.006 Hz; NS p = 0.18)(Fig. 6). In this case, both amplitude (control:13.48 ± 0.97 pA; copper: 17.65 ± 0.71 pA; **p < 0.01)and time constant of GABA events (control:21.52 ± 1.76 ms; copper: 29.94 ± 2.16 ms) were increasedin copper-treated cells (Fig. 6a–c). The increase in theamplitude of GABAergic currents correlated with an increasein GABAA receptors after 3 h treatment with copper(Fig. 6d). These results indicate that changes in total synapticactivity induced by copper are because of changes inAMPAergic and GABAergic neurotransmission.
Fig. 2 Minimal concentration of copper to increase synaptic activity in
rat hippocampal neurons. (a) Typical miniature synaptic currents of rat
hippocampal neurons treated in the absence or presence of copper (1,
5 and 10 lM) for 3 h. (b) The graph summarizes the frequency (Hz)
obtained in these experiments. The bars are mean ± SEM from at
least six neurons (**p < 0.01).
Journal of Neurochemistry � 2011 International Society for Neurochemistry, J. Neurochem. (2011) 10.1111/j.1471-4159.2011.07417.x� 2011 The Authors
4 | C. Peters et al.
Copper increases functional AMPA receptorsThe fact that copper-treated neurons displayed an increase inamplitude of synaptic events may be as a result of the
modifications at the post-synaptic level. In fact, the levels ofGABAA receptors are increased in copper-treated neuronsafter 3 h of treatment (Fig. 6d). In the case of AMPAergic
Fig. 3 Copper increases synaptic activity in
rat hippocampal neurons. (a) Typical mini-
ature synaptic currents of rat hippocampal
neurons treated for 3 h in the absence or
presence of the transition metals (10 lM)
indicated in the figure. Graphs summarize
data obtained in these experiments: (b)
frequency (Hz); (c) amplitude (pA) and (d)
decay (ms). The bars are mean ± SEM
from at least six neurons (*p < 0.05,
**p < 0.01, ***p < 0.001).
Fig. 4 Copper application for 24 h does not change the synaptic
activity in rat hippocampal neurons. (a) Typical miniature synaptic
currents of rat hippocampal neurons treated for 24 h in the absence
(control) or presence of copper (10 lM). A zoom of representative
miniature synaptic events of rat hippocampal neurons treated with
copper for 24 h is shown. Graphs summarize data obtained in these
experiments with copper (10 lM): (b) frequency (Hz); (c) amplitude
(pA). Typical miniature synaptic currents of rat hippocampal neurons
treated for 24 h in the absence (control) or presence of iron (10 lM) or
zinc (10 lM) are summarized on the graphs: (d) frequency (Hz) and
(e) amplitude (pA). The bars are mean ± SEM from at least six neu-
rons (*p < 0.05, ***p < 0.001).
� 2011 The AuthorsJournal of Neurochemistry � 2011 International Society for Neurochemistry, J. Neurochem. (2011) 10.1111/j.1471-4159.2011.07417.x
Copper enhances CNS neurotransmission | 5

currents, amplitude was also increased after 3 h treatmentwith copper (Fig. 5), suggesting that a major number ofreceptors should be at the plasma membrane. We analyzedthe localization of AMPA receptors using an antibody thatdetects the GluA1 subunit of these excitatory receptors. Wefound that GluA1 staining was significantly increased(***p < 0.001) at MAP2-positive dendritic zones of cop-
per-treated neurons (Fig. 7a and b), which seemed to belocalized at the plasma membrane. Moreover, we found thatanother subunit of the AMPA receptor, GluA2, was alsoincreased after 3 h of treatment with copper (Fig. 6d). Toconfirm that AMPA receptors were functionally active andeffectively localized at the post-synaptic membrane, weperformed studies of evoked currents using the AMPA ligand
Fig. 5 Synaptic activity induced by copper
is mediated by an increase in AMPA cur-
rents and calcium transients. (a) Repre-
sentative traces of AMPA minis isolated by
pharmacology maneuvers [APV (5 lM);
strychnine (1 lM); bicuculline (5 lM)] in
control neurons and copper-treated
neurons for 3 h. Graphs summarize data
obtained in these experiments: (b) fre-
quency (Hz); (c) amplitude (pA) and (d)
decay (ms). The bars are mean ± SEM
from at least six neurons (**p < 0.01,
***p < 0.001). (e) Calcium transient records
of control and copper-treated (10 lM)
neurons for 3 h. (f) Frequency of calcium
transient showing an increase after copper
treatment (CuCl2, 10 lM) for 3 h (*p < 0.05).
Fig. 6 Copper does not affect the fre-
quency of GABA currents, but increases the
amplitude and decay time. GABA mini cur-
rents were isolated by pharmacology
maneuvers [AP-V (5 lM); strychnine
(1 lM); CNQX (4 lM)] in control neurons
and copper-treated (10 lM) neurons by 3 h.
Graphs summarize data obtained in these
experiments for: (a) frequency (Hz); (b)
amplitude (pA) and (c) decay (ms). The
bars are mean ± SEM from at least six
neurons (**p < 0.01). (d) Confocal micro-
photographs of untreated (control) or cop-
per-treated (10 lM) hippocampal neurons
for 3 h, showing immunoreactivity for GluA2
(red) and GABAA a-1 (green) subunits.
Right panels show the merge (yellow) of
GluA2 (red) and GABAA a-1 (green)
subunits immunoreactivity. Scale bar
represents 25 lm.
Journal of Neurochemistry � 2011 International Society for Neurochemistry, J. Neurochem. (2011) 10.1111/j.1471-4159.2011.07417.x� 2011 The Authors
6 | C. Peters et al.

(Fig. 7c). These studies revealed that copper-treated neuronswere more sensitive to AMPA compared with controlneurons, showing a significant shift to the left in the dose–response curve. In fact, the EC50 for AMPA observed incontrol neurons (EC50 8.3 ± 0.5 lM) was decreased to anEC50 of 2.9 ± 0.2 lM in copper-treated (10 lM; 3 h)neurons (Fig. 7d). Furthermore, the desensitization of AMPAreceptors was slower in copper-treated neurons as shown bythe values for peak/plateau of the currents evoked by AMPA(control: 1.84 ± 0.58 pA; copper-treated: 1.28 ± 0.09 pA;*p < 0.05), which favor a longer opening time for AMPAreceptors. However, when copper was co-applied withAMPA (‘acute application of copper’), AMPA evokedcurrents were effectively inhibited by the presence of copper(IC50 22.9 ± 3.8 lM) (Fig. 1a and b), in agreement withprevious results recorded in cortical neurons (Weiser andWienrich 1996). All together, our data indicate that neuronsbehave differently to copper under acute versus prolongedincubation time, through mechanisms that may or may not beconnected. For example, 3 h of copper treatment may arisefrom a permanent block of AMPA receptors throughhomeostatic or anti-homeostatic mechanisms (Carrasco et al.2007), which then may increase these receptors at thesynaptic membrane. Thus, to explore this idea using adifferent pharmacological approach, we incubated hippo-campal cultures for 3 h with 4 lM of the AMPA blockerCNQX (See Figure S1), at a concentration that efficientlyinhibits AMPA receptors (Carrasco et al. 2007). After 3 h of
incubation, we did not observe any change in total synapticactivity (See Figure S1), indicating that blockade of AMPAreceptors at 3 h did not induce a compensatory response atthe synapses, suggesting that the effect of copper onneurotransmission after 3 h of incubation is probably notrelated to the acute inhibitory effect of copper on AMPAcurrents (Fig. 1) (Weiser and Wienrich 1996).
To evaluate if the changes observed for AMPA andGABAA receptors by immunofluorescence were because ofchanges in the expression of these receptors, we performedwestern blot analysis using anti-GluA1 and anti-GABAA
a-1 subunit antibodies. Interestingly, this study showed nodifferences in the levels of both proteins between controland copper-treated (3 h) neurons (Fig. 8a–c), indicatingthat the mechanism behind the effect of copper onneurotransmission should be explained by changes in thelocalization of receptors. In fact, immunoflurescence studiesperformed in non-permeabilized neurons show the presenceof large clusters of GluA1 subunits at the plasmamembrane, which were undetectable in control neurons(Fig. 8d and e). The result supports the idea that copperincreases AMPAergic neurotransmission by the clusteringof AMPA receptors at the plasma membrane. Interestingly,and in agreement with the literature (Nose et al. 2010), weobserved that the total levels of the main copper transporterlocated at the plasma membrane, Ctr1, were decreased(Fig. 8f and g) after 3 h of treatment with copper,indicating that this brain primary culture possesses the
Fig. 7 Membrane anchorage of functional AMPA receptors is induced
by copper in hippocampal neurons. (a) Confocal microphotographs of
untreated (control) or copper-treated (10 lM) hippocampal neurons for
3 h, showing immunoreactivity for GluA1 subunit (red) and MAP2
(green). Scale bar represents 20 lm. (b) The graph shows a
quantitative analysis of GluA1 subunit immunoreactivity shown in pa-
nel A. The bars are mean ± SEM from at least six neurons
(***p < 0.001). (c) Representative traces of AMPA evoked currents on
untreated neurons and copper-treated (10 lM) neurons for 3 h. (d)
AMPA dose–response curves for untreated (EC50 = 8.26 ± 0.5 lM)
and copper-treated (10 lM) neurons for 3 h (EC50 = 2.89 ± 0.2 lM)
(p < 0.001).
� 2011 The AuthorsJournal of Neurochemistry � 2011 International Society for Neurochemistry, J. Neurochem. (2011) 10.1111/j.1471-4159.2011.07417.x
Copper enhances CNS neurotransmission | 7

Fig. 8 Copper increases the clustering of AMPA receptors at the
plasma membrane. (a) Western blot of control and copper-treated
(10 lM) neurons for 3 h, showing the levels of GluA1 and GABAA a-1
subunits. The graph shows a quantitative analysis of the immunore-
activity for (b) GluA1 and (c) GABAA a-1 subunits. (d) Confocal
microphotographs of untreated (control) or copper-treated (10 lM)
hippocampal neurons for 3 h, showing immunoreactivity for GluA1
subunits in non-permeabilized neurons. The arrow heads indicate
small clusters, and the arrows indicate large clusters. Scale bar rep-
resents 15 lm. (e) The graph shows a quantitative analysis of the size
of GluA1 clusters (lm2; *p < 0.05). (f) Western blot of control (left line)
and copper-treated (10 lM; right line) neurons for 3 h, showing the
immunoreactivity for the copper transporter Ctr1. b-actin was used as
internal control. (g) The graph shows a quantitative analysis of the
immunoreactivity for Ctr1 (*p < 0.05).
Fig. 9 PSD95 levels are regulated by cop-
per. (a) Confocal microphotographs of un-
treated (control) or copper-treated (10 lM)
neurons for 3 h, showing immunoreactivity
for PSD95. Scale bar of 30 lm. (b) The
graph shows a quantitative analysis of
PSD95 immunoreactivity shown in panel A.
The bars are mean ± SEM from at least six
neurons (***p < 0.001). (c) Western blot
showing the increase of PSD95 levels from
neurons treated with copper (up to 10 lM).
a-tubulin was used as internal control. (d)
Western blot showing the decrease of
PSD95 levels from neurons treated with the
copper chelator, TTM (25 lM).
Journal of Neurochemistry � 2011 International Society for Neurochemistry, J. Neurochem. (2011) 10.1111/j.1471-4159.2011.07417.x� 2011 The Authors
8 | C. Peters et al.

machinery to regulate the copper uptake at the plasmamembrane. These results also indicate that total levels ofAMPA and GABAA receptors are not affected by copperin the same fashion than Ctr1.
To analyze whether changes in AMPA receptor wereaccompanied with an increase in scaffolding proteins neces-sary for its accumulation at the synaptic membrane, wefurther evaluated the levels and localization of PSD95, acritical protein for AMPA receptor anchorage at the synapses(Colledge et al. 2003). Remarkably, these studies indicatedthat protein levels of PSD95 are indeed increased in copper-treated neurons (Fig. 9a–c), thus offering a potentialexplanation for the clustering of AMPA receptors in thepost-synaptic membranes. Interestingly, when hippocampalneurons were treated with a copper chelator, TTM (25 lM),the levels of PSD95 were decreased (Fig. 9d), indicating theimportance of copper on the regulation of PSD95 and thefunction of AMPA receptors.
Discussion
The importance of copper in the CNS is well documented,but the mechanisms behind its brain functions are unknown(Linder and Hazegh-Azam 1996). Brain copper deficiency isobserved in Menke’s disease, which affects brain physiology,as patients display gray matter degeneration, Purkinje cellabnormalities and hippocampal neuronal loss (Okeda et al.1991). AD is another pathology characterized by neurode-generation that produces a broad spectrum of symptoms thathave been linked to copper brain depletion as cupro-proproteins such as ceruloplasmin are decreased (Connoret al. 1993) or less active as observed by superoxidedismutase (Omar et al. 1999; Maynard et al. 2005). Cur-rently, the effects of copper in CNS diseases are thus poorlyunderstood.
It has been suggested that copper is released at the synapticcleft (Hartter and Barnea 1988; Hopt et al. 2003), where itmay modulate neurotransmission (Weiser and Wienrich1996; Schlief et al. 2005, 2006). In this regard, this is thefirst time that copper is shown to enhance synaptic activityafter 3 h of incubation (Figs 2 and 3). This effect is specificbecause it is not produced by 3 h applications of othertransition metals, such as zinc or iron (Fig. 3). Bothfrequency and amplitude of total synaptic events wereincreased in copper-treated neurons, indicating that pre- andpost-synaptic components are involved (Fig. 3b and c). Forinstance, a pre-synaptic mechanism is supported by theobservation that the frequency of calcium transients was alsoaugmented in copper-treated neurons (Fig. 5e and f). On theother hand, a post-synaptic mechanism is suggested by theincrease in amplitude displayed in copper-treated neurons(Fig. 3c), which also correlated with an increase of GluA1synaptic levels (Fig. 7). Interestingly, we have found that thisincrease was related to an increase in PSD95 (Fig. 9),
indicating that AMPA receptor anchorage at post-synapticmembranes may be occurring under these experimentalconditions. We also found that TTM, a copper II chelator,mirrored the effect of copper on PSD95, indicating thatsomehow PSD95 expression is under the regulation ofcopper (Fig. 9), through a direct or indirect mechanism. Forexample, copper could directly interact with PSD95 increas-ing its stability or decreasing its degradation by theproteasome (Colledge et al. 2003). In fact, PSD95 has inits structure several metal-coordinating amino acids, such ashistidine and tyrosine that may be targeted by copper.Conserved histidine residues (H130 and H225) are located inthe PSD95 PDZ domains, which are important for theinteraction of PDS95 with stargazin (Schnell et al. 2002).Therefore, these histidines could be targeted by copper,increasing AMPA clustering driven by PSD95-stargazininteraction. However, other alternative mechanisms mayexplain the effect of copper on neurotransmission. Forexample, copper could increase phosphorylation of serine-295 of PSD95 (Kim et al. 2007), which is important forAMPAergic neurotransmission (Kim et al. 2007). Moreover,copper could regulate PSD-95 palmitoylation, which occurson cysteine residues (Craven et al. 1999) that could also be apossible target for copper. All these possible mechanismsdeserve further examination.
All together, these results suggest that neurons exposed toa copper-enriched environment display a more activesynaptic transmission, which might be important in neuro-degenerative diseases characterized by an imbalance incopper homeostasis (Bush 2003).
AMPA-evoked currents were significantly increased incopper-treated neurons, showing a ligand shift to the leftreflected by a significant change in EC50 (control:8.3 ± 0.5 lM; copper-treated: 2.9 ± 0.2 lM, p < 0.001),indicating that these neurons have more AMPA receptorsat the plasma membranes as it is suggested by theimmunofluorescence analysis and AMPA-evoked currents(Fig. 7). Moreover, desensitization was also decreased inthese neurons keeping AMPA channels open for longer times(Fig. 7c). However, we cannot discard that other auxiliaryproteins, such as TARP, may be involved in this effect(Farrant and Cull-Candy 2010).
Finally, copper also increased the amplitude of theGABAergic transmission but without affecting frequency ofthese synaptic events (Fig. 6), indicating only a post-synapticmechanism probably involving changes in receptor pharma-cology, as time constant of GABA events was increased incopper-treated neurons by a mechanism that was not exploredin the present work, which may involve homeostatic or anti-homeostatic mechanisms (Carrasco et al. 2007).
The molecular mechanism of copper (3 h) in neurotrans-mission seemed to be unrelated to a compensatory actionresulting from the blockade of AMPA receptor, which wasable to inhibit AMPA evoked currents in hippocampal
� 2011 The AuthorsJournal of Neurochemistry � 2011 International Society for Neurochemistry, J. Neurochem. (2011) 10.1111/j.1471-4159.2011.07417.x
Copper enhances CNS neurotransmission | 9

neurons when copper was acutely applied to neurons (Fig. 1aand b), as it has been previously described in cultured ratcortical neurons (Weiser and Wienrich 1996). In this regard,we examined the total synaptic events in neurons afterblockade of AMPA receptors for 3 h with CNQX, which is aspecific and potent antagonist of AMPA currents. Thispharmacological maneuver did not affect any parameter ofthe total mini currents, indicating that at this time frame acompensatory mechanism is not responsible for receptorblockade (See Figure S1). Therefore, we speculate that themechanism behind the effect of copper on this neuronalnetwork involves intracellular changes independent ofAMPA receptor blockade. Interestingly, after 24 h exposureof copper, the synaptic activity returned to the control levels,indicating that changes in neurotransmission observed at 3 hare under homeostatic regulation.
In summary, in this work, we have shown that copper, in atime and concentration dependent manner, may inducedifferent effects on neurotransmission, indicating that afine-tuning of this essential metal is needed by neuronal cellsto maintain adequate synaptic function. In fact, an increase inthe brain concentration of copper, as well as a decrease in thelevels of this metal, can lead to serious illness (Bush 2003).Therefore, future studies are needed to better understand themechanisms behind copper effects on living neurons, whichmay lead to prevent or to treat copper-related neurodegen-erative diseases and neurological conditions, which remainuntreatable to date.
Acknowledgements
We would like to thank the members of the laboratory, especiallyClaudia Lopez and Laurie Aguayo for their technical assistance.This work was supported by the Chilean Government (Anillo-PBCTACT-04 project to LGA, GVD and CO), FONDECYT 1100502(LGA), FONDECYT 1100942 (GDV) and DIUC grant Nº205.033.101-1.0 (CO). CP and FJS are recipients of CONICYTfellowships. We thank Professor Dennis Thiele and Dr YasuhiroNose for providing us anti-Ctr1 antibody. We are grateful to the‘‘Centro de Microscopıa Avanzada (CMA) Bıo Bıo for technicalassistance in confocal microscopy. The authors declare no conflictof interest.
Supporting information
Additional supporting information may be found in the onlineversion of this article:
Figure S1. The AMPA receptor antagonist CNQX does notincrease the synaptic activity after 3 h of incubation.
As a service to our authors and readers, this journal providessupporting information supplied by the authors. Such materials arepeer-reviewed and may be re-organized for online delivery, but arenot copy-edited or typeset. Technical support issues arising fromsupporting information (other than missing files) should beaddressed to the authors.
References
Aguayo L. G. and Pancetti F. C. (1994) Ethanol modulationof the c-aminobutyric acid A- and glycine-activated Cl) current incultured mouse neurons. J. Pharmacol. Exp. Ther. 270, 61–69.
Avila M. E., Sepulveda F. J., Burgos C. F., Moraga-Cid G., Parodi J.,Moon R. T., Aguayo L. G., Opazo C. and De Ferrari G. V. (2010)Canonical Wnt3a modulates intracellular calcium and enhancesexcitatory neurotransmission in hippocampal neurons. J. Biol.Chem. 285, 18939–18947.
Bush A. I. (2003) The metallobiology of Alzheimer’s disease. TrendsNeurosci. 26, 207–214.
Carrasco M. A., Castro P. A., Sepulveda F. J., Cuevas M., Tapia J. C.,Izaurieta P., van Zundert B. and Aguayo L. G. (2007) Anti-homeostatic synaptic plasticity of glycine receptor function afterchronic strychnine in developing cultured mouse spinal neurons.J. Neurochem. 100, 1143–1154.
Colledge M., Snyder E. M., Crozier R. A., Soderling J. A., Jin Y.,Langeberg L. K., Lu H., Bear M. F. and Scott J. D. (2003)Ubiquitination regulates PSD-95 degradation and AMPA receptorsurface expression. Neuron 40, 595–607.
Connor J. R., Tucker P., Johnson M. and Snyder B. (1993) Cerulo-plasmin levels in the human superior temporal gyrus in aging andAlzheimer’s disease. Neurosci. Lett. 159, 88–90.
Craven S. E., El-Husseini A. E. and Bredt D. S. (1999) Synaptic tar-geting of the postsynaptic density protein PSD-95 mediated bylipid and protein motifs. Neuron. 22, 497–509.
Farrant M. and Cull-Candy S. G. (2010) Neuroscience. AMPA recep-tors-another twist? Science 327, 1463–1465.
Goldschmith A., Infante C., Leiva J., Motles E. and Palestini M. (2005)Interference of chronically ingested copper in long-term potentia-tion (LTP) of rat hippocampus. Brain Res. 1056, 176–182.
Gu X., Olson E. C. and Spitzer N. C. (1994) Spontaneous neuronalcalcium spikes and waves during early differentiation. J. Neurosci.14, 6325–6335.
Hartter D. E. and Barnea A. (1988) Evidence for release of copper in thebrain: depolarization-induced release of newly taken-up 67copper.Synapse 2, 412–415.
Hopt A., Korte S. and Fink H. (2003) Methods for studying synapto-somal copper release. J. Neurosci. Methods 128, 159–172.
Kardos J., Kovacs I., Hajos F., Kalman M. and Simonyi M. (1989)Nerve endings from rat brain tissue release copper upon depolar-ization. A possible role in regulating neuronal excitability. Neu-rosci. Lett. 103, 139–144.
Kim M. J., Futai K., Jo J., Hayashi Y., Cho K. and Sheng M. (2007)Synaptic accumulationof PSD-95 and synaptic function regulated byphosphorylation of serine-295 of PSD-95. Neuron. 56, 488–502.
Leiva J., Palestini M., Infante C., Goldschmidt A. and Motles E. (2009)Copper suppresses hippocampus LTP in the rat, but does not alterlearningormemory in themorriswatermaze.BrainRes.1256, 69–75.
Linder M. C. and Hazegh-Azam M. (1996) Copper biochemistry andmolecular biology. Am. J. Clin. Nutr. 63, 797S–811S.
Lovell M. A., Robertson J. D., Teesdale W. J., Campbell J. L. andMarkesbery W. R. (1998) Copper, iron and zinc in Alzheimer’sdisease senile plaques. J. Neurol. Sci. 158, 47–52.
Mathie A., Sutton G. L., Clarke C. E. and Veale E. L. (2006) Zinc andcopper: pharmacological probes and endogenous modulators ofneuronal excitability. Pharmacol. Ther. 111, 567–583.
Maynard C. J., Bush A. I., Masters C. L., Cappai R. and Li Q. (2005)Metals and amyloid-b in Alzheimer’s disease. Int. J. Exp. Pathol.86, 147–159.
Noel J., Ralph G. S., Pickard L., Williams J., Molnar E., Uney J. B.,Collingridge G. L. and Henley J. M. (1999) Surface expression of
Journal of Neurochemistry � 2011 International Society for Neurochemistry, J. Neurochem. (2011) 10.1111/j.1471-4159.2011.07417.x� 2011 The Authors
10 | C. Peters et al.

AMPA receptors in hippocampal neurons is regulated by an NSF-dependent mechanism. Neuron. 23, 365–376.
Nose Y., Wood L. K., Kim B. E., Prohaska J. R., Fry R. S., Spears J. W.and Thiele D. J. (2010) Ctr1 is an apical copper transporter inmammalian intestinal epithelial cells in vivo that is controlled at thelevel of protein stability. J. Biol. Chem. 285, 32385–32392.
Okeda R., Gei S., Chen I., Okaniwa M., Shinomiya M. and MatsubaraO. (1991) Menkes’ kinky hair disease: morphological and immu-nohistochemical comparison of two autopsied patients. Acta Neu-ropathol. 81, 450–457.
Omar R. A., Chyan Y. J., Andorn A. C., Poeggeler B., Robakis N. K.and Pappolla M. A. (1999) Increased expression but reducedactivity of antioxidant enzymes in Alzheimer’s disease. J. Alzhei-mers Dis. 1, 139–145.
Schlief M. L., Craig A. M. and Gitlin J. D. (2005) NMDA receptoractivation mediates copper homeostasis in hippocampal neurons.J. Neurosci. 25, 239–246.
Schlief M. L., West T., Craig A. M., Holtzman D. M. and Gitlin J. D.(2006) Role of the Menkes copper-transporting ATPase in NMDAreceptor-mediated neuronal toxicity. Proc. Natl Acad. Sci. USA103, 14919–14924.
Schnell E., Sizemore M., Karimzadegan S., Chen L., Bredt D. S. andNicoll R. A. (2002) Direct interactions between PSD-95 and star-gazin control synaptic AMPA receptor number. Proc. Natl Acad.Sci. USA 99, 13902–13907.
Sepulveda F. J., Opazo C. and Aguayo L. G. (2009) Alzheimerb-amyloid blocks epileptiformactivity in hippocampal neurons.Mol. Cell. Neurosci. 41, 420–428.
Sharonova I. N., Vorobjev V. S. and Haas H. L. (1998) High-affinitycopper block of GABA(A) receptor-mediated currents in acutelyisolated cerebellar Purkinje cells of the rat. Eur. J. Neurosci. 10,522–528.
Stuerenburg H. J. (2000) CSF copper concentrations, blood-brain barrierfunction, and coeruloplasmin synthesis during the treatment ofWilson’s disease. J. Neural. Transm. 107, 321–329.
Tarohda T., Yamamoto M. and Amamo R. (2004) Regional distributionof manganese, iron, copper, and zinc in the rat brain duringdevelopment. Anal. Bioanal. Chem. 380, 240–246.
Trombley P. Q. and Shepherd G. M. (1996) Differential modulation byzinc and copper of amino acid receptors from rat olfactory bulbneurons. J. Neurophysiol. 76, 2536–2546.
Vlachova V., Zemkova H. and Vyklicky L. (1996) Copper modulationof NMDA responses in mouse and rat cultured hippocampal neu-rons. Eur. J. Neurosci. 8, 2257–2264.
Watt A. J., van Rossum M. C., MacLeod K. M., Nelson S. B. andTurrigiano G. G. (2000) Activity coregulates quantal AMPA andNMDA currents at neocortical synapses. Neuron. 26, 659–670.
Weiser T. and Wienrich M. (1996) The effects of copper ions on glu-tamate receptors in cultured rat cortical neurons. Brain Res. 742,211–218.
� 2011 The AuthorsJournal of Neurochemistry � 2011 International Society for Neurochemistry, J. Neurochem. (2011) 10.1111/j.1471-4159.2011.07417.x
Copper enhances CNS neurotransmission | 11