nicotinic acetylcholine receptor distribution in relation to spinal neurotransmission pathways
TRANSCRIPT

Nicotinic Acetylcholine ReceptorDistribution in Relation to Spinal
Neurotransmission Pathways
IMRAN KHAN,1* HITOSHI OSAKA,1 SHANAKA STANISLAUS,1 ROSA M. CALVO,1
TOM DEERINCK,3 TONY L. YAKSH,1,2AND PALMER TAYLOR1
1Department of Pharmacology, University of California, San Diego,La Jolla, California 92093-0636
2Department of Anesthesiology, University of California, San Diego,La Jolla, California 92093-0636
3National Center for Microscopy and Imaging Research, University of California,San Diego, La Jolla, California 92093-0636
ABSTRACTNeuronal nicotinic receptors (nAChR) are pentameric assemblies of subunits of a gene
family where specified combinations of � and � subunits form functional receptors. To extendour understanding of the role of spinal nAChR in the processing of sensory stimuli andregulation of autonomic and motor responses, we initiated investigations to localize nAChRsubunit expression within discrete spinal regions and cell types. High-affinity epibatidinebinding was present in the superficial dorsal and ventral horns, the mediolateral and centralcanal regions. RT-PCR identified transcripts for �3, �4, �5, �2, and �4 in both spinal cordparenchyma and dorsal root ganglia (DRG). Our affinity-purified antibodies against �3, �4,�5, �2, and �4 subunits identified specific protein bands of appropriate molecular mass(preadsorbed with the respective antigens) in specific tissues and cells that express nicotinicreceptors, including the spinal cord and DRG neurons. Having established the absence ofcrossreactivity with related subunits, specific fluorescence labeling of nerve terminals andcell bodies was achieved and correlated with the distribution of defined marker proteins andnicotinic receptor binding sites determined autoradiographically. Our findings indicate that�3, �4, �5, �2, and �4 subunits are all expressed on primary afferents (IB4-positive termi-nals) in the spinal cord. The predominant presynaptic (synaptophysin colocalization) labelingis in the superficial layer of the dorsal horn. These receptor subunits, except for �4, are alsopresent in postsynaptic autonomic (anti-bNOS-positive) and somatic motor neurons (anti-VAChT-positive). The �3, �5, and �2 subunits showed additional staining in glial (anti-GFAP-positive) cells. These studies reveal a dense and distinguishable distribution of nAChRsubunits in the spinal cord and point toward future therapeutic targeting for specific spinalactions. J. Comp. Neurol. 467:44–59, 2003. © 2003 Wiley-Liss, Inc.
Indexing terms: spinal nicotinic receptor; neuronal nicotinic receptor subunits; receptor
autoradiography; immunohistochemistry; primary afferent terminals; presynaptic
receptors; cholinergic neurons
Nicotinic receptors within the central nervous system(CNS) arise from assembly of multiple homologous subunitsencoded by at least 12 separate genes to form pentamericreceptors (Lukas et al., 1999). While some of the subunits,�7, �8, and �9, assemble as homomeric pentamers in neu-rons, most � subunits, �2, �3, �4, and �6, typically assembleas two copies with three copies of an assortment of �2, �4,�3, and �5 subunits to form pentamers. Typically, the het-eromeric receptors are thought to have two binding sites thatare found at the interface ofthe � and � subunits (Lukas etal., 1999; Lindstrom, 2000; Bertrand, 2001).
Grant sponsor: U.S. Public Health Service; Grant number: HL-35018; Grantsponsor: the California Tobacco-Related Disease Research Program (P.T.).
*Correspondence to: Imran Khan, Department of Pharmacology, Univer-sity of California San Diego, 9500 Gilman Drive, La Jolla, CA 92093-0636.E-mail: [email protected]
Received 9 July 2002; Revised 8 May 2003; Accepted 14 July 2003DOI 10.1002/cne.10913Published online the week of October 13, 2003 in Wiley InterScience
(www.interscience.wiley.com).
THE JOURNAL OF COMPARATIVE NEUROLOGY 467:44–59 (2003)
© 2003 WILEY-LISS, INC.

In the rat, in situ hybridization studies have mapped thelocation of nAChR transcripts to various regions of the spinalcord and higher centers of the CNS (Wada et al., 1989, 1990;Boyd et al., 1991; Genzen et al., 2001; Azam et al., 2002). Inour previous studies, we observed that intrathecal nicotinicagonists elicit pressor, heart rate, nociceptive, and antinoci-ceptive responses, the nature of which is agonist selectiveand blocked by specific nicotinic antagonists (Khan et al.,1994b, 1998, 2001). Thus, the role of multiple nicotinic re-ceptors can, in part, be identified on the basis of functionalparameters. Ligand binding studies with radiolabeled nico-tinic agonists also reveal the existence of multiple subtypesin the spinal cord (Khan et al., 1994c, 1997). The capacity ofnicotinic agonists to release other transmitters as detectedby microdialysis suggests that the receptors may regulatetransmitter release from presynaptic sites (Khan et al.,1996a), as appears to be the case elsewhere in the CNS(MacDermott et al., 1999; Kiyosawa et al., 2001).
To identify specific nicotinic receptor subtypes in the spi-nal cord and link them to function, we employed antibodiesdirected to the abundant subunits in the spinal cord to lo-calize through immunochemistry the expression of nicotinicreceptor proteins in various spinal neurons and nerve termi-nals. The spinal cord offers a unique setting for such studies,since agonists can be administered at discrete segmentallevels and functional responses ascertained. In turn, modi-fication of receptor expression and function through selectivelesioning enables one to correlate altered responses of func-tioning nerve tracts with receptor distribution.
In this study, we employed a series of subunit selectiveantibodies to localize �3, �4, �5, �2, and �4 nicotinicreceptor subunits in spinal neurons and terminals. Inturn, the localization of receptor subunit expression iscorrelated with the distribution of binding sites in thespinal cord as determined by autoradiography. These find-ings provide an important step in correlating spinal nico-tinic receptor subtype and localization with function ofspinal nerve tracts and pathways in the conscious animal.
MATERIALS AND METHODS
Experimental animals
Male Sprague-Dawley rats (300–350 g) and male NewZealand White rabbits (2–4 kg) were purchased from Har-lan (Indianapolis, IN) and Irish Farm (Norco, CA), respec-tively. All animals were housed in the animal facility andwere maintained on 12-hour light/dark cycles. All studieswere carried out according to protocols reviewed and ap-proved by the Institutional Animal Care Committee.
[3H]-Epibatidine binding autoradiography in
spinal cord sections
Procedures for autoradiographic localization of nicotinicreceptors in spinal sections were described previously
(Khan et al., 1996b). Briefly, 16 �m-thick lumbar andthoracic spinal cord transverse sections were mounted onFisher (Pittsburgh, PA) lysine-precoated glass slides. Theslides were dipped in assay buffer for 10 minutes followedby incubation in assay buffer (pH 7.4) containing either0.05 nM or 0.8 nM of [3H]-epibatidine for 60 minutes at4°C. After incubation, the slides were washed by foursuccessive dippings of 1 minute in ice-cold assay bufferfollowed by two dips in ice-cold deionized distilled water.Nonspecific binding was determined by coincubation oftissue sections with [3H]-epibatidine and 60 �M ofl-nicotine. Dried slides were placed against Amersham(Piscataway, NJ) [3H]-Hyperfilm for 8 –10 weeks, thenthe films were hand-processed with D-19 developer andfixer.
Autoradiograms on films were scanned with an Epson(Expression 1600) scanner, processed with Adobe Photo-Shop, and binding intensities analyzed using NIH-Image.The varying intensities of [3H]-epibatidine binding in spi-nal cord tissue were artificially color coded with a color-table prepared using Adobe PhotoShop.
Fig. 1. Autoradiograms illustrating [3H]-epibatidine binding in ratspinal cord sections. A: Binding of 0.05 nM epibatidine to lumbarsections. B,E: Binding of 0.8 nM [3H]-epibatidine to lumbar andthoracic spinal cord sections, respectively. C,D,F: Artificial colorcoded intensities of the actual binding in A,B,E, respectively. Theyellow circles demarcate the IML region in the thoracic spinal cord.The rectangular bars in the bottom of the composite represent thecolor table gradient for the corresponding white to black gradient inthe autoradiograms. Scale bar � 500 �m.
Abbreviations
bNOS brain nitric oxide synthaseCy cyaninDRG dorsal root ganglionFITC fluorescein isothiocyanateGFAP glial fibrillary acidic proteinIML intermediolateral cell columnnAChR nicotinic acetylcholine receptorNeuN B neuronal nuclei BRT-PCR reverse transcriptase-polymerase chain reactionVAChT vesicular acetylcholine transporter protein
45NICOTINIC RECEPTOR SUBUNITS IN SPINAL TISSUE

Fig. 2. Detection of nAChR subunits �3, �4, �5, �2, and �4 inspinal cord regions and dorsal root ganglia by reverse-transcriptasepolymerase chain reaction (RT-PCR). Total RNA samples (3–5 �g)isolated from dorsal and ventral lumbar spinal cord and dorsal rootganglion were used as template for first strand synthesis. cDNAs werethen amplified using the appropriate primers for specific � and �
subunits. In the top panel, the 1 Kb bands represent 1,000, 850, 650,500, and 400 basepair lengths, whereas, in the bottom panel theladder bands show 400, 300, and 200 basepair lengths. DRG, dorsalroot ganglion tissue; SC-D, dorsal spinal cord; SC-V, ventral spinalcord.
Fig. 3. Western blots demonstrating specificity of affinity-purifiedpolyclonal antibodies against (A) �4, (B) �3, (C) �5, (D) �2, and (E) �4receptor subunits. Equal concentrations of different GST-nAChR sub-unit fusion proteins were electrophoresed and immobilized on theblot. The numbers indicate prestained molecular weight markers(Gibco, Gaithersburg, MD); they run faster than the unstained mo-lecular weight standards (Novex). Accordingly, the GST-fusion pro-
teins of predicted molecular mass of �4 (37 kD), �3 (34 kD), �5 (34kD), �2 (34 kD), and �4 (35 kD) run slightly slower than theprestained markers when compared on a mass basis. Following elec-trophoresis, Coomassie staining of the GST-proteins detects migra-tion rates that correspond with the Western blot detected proteins.The ratios indicate dilution of the original stock solution (�200 ng/lane) of the GST-fusion proteins.
46 I. KHAN ET AL.

Detection of RNA species
RNA templates for RT-PCR assays were isolated fromvarious tissue or cell culture preparations with TRI-Reagent (Sigma, St. Louis, MO). RT-PCR reactions wereperformed according to the protocol provided in RT-PCRkit from Epicenter Technologies (Madison, WI). The RT-PCR product obtained for each nicotinic receptor subunitwas verified by restriction enzyme digestion and by nucle-otide sequencing. The primer sequences for the variousalpha and beta subunits used in this study are describedin http://phrtayl0.ucsd.edu/ikhan.
GST-fusion protein constructs, synthesis,and purification
DNA fragments encoding segments of the cytoplasmicdomain (between transmembrane spans 3 and 4) of a
specific nicotinic receptor subunit were PCR-amplifiedfrom the cDNA encoding the subunits. The cDNA sourcesemployed have been described in previous publications(Boulter et al., 1987; Bertrand, et al., 1990; Galzi et al.,1991). PCR-amplified purified DNA fragments with theappropriate 5� overhang were ligated into pGEX-2T vectorat a Bam HI restriction site. Sequences of the plasmids inthe transformants from E. coli were verified by restrictionenzyme digestions and then DNA sequencing.
High expression of GST-nicotinic receptor subunit fu-sion proteins was achieved by transformation into E. coliBL21 and induction by IPTG for 3 hours. Bacterially syn-thesized GST-fusion proteins were purified using a B-PERGST Fusion Protein Purification Kit from Pierce (Rock-ford, IL). SDS-PAGE and Coomassie blue staining identi-fied the overexpressed GST fusion proteins. The authen-ticity of the specific GST-fusion protein was verified byN-terminal amino acid sequencing of the thrombin di-gested peptide fragments (data not shown).
Design of synthetic peptide antigens andgeneration of antibodies
Peptide fragments, 22–26 amino acids in length, of se-quence within the cytoplasmic domain between trans-membrane segments 3 and 4 of the neuronal nAChR sub-units (�3, �4, �5, �2, and �4), were synthesized byGenmed (San Francisco, CA). Sequences of the syntheticpeptides from the segments of nAChR subunits containedin the GST-fusion proteins referred to above are describedin http://phrtayl0.ucsd.edu/ikhan.
Rabbits (2–4 kg) were inoculated intramuscularly with200 �g of the KLH-conjugated peptide followed by 4–6
Fig. 4. Western blots illustrating the molecular masses of variousnAChR subunits in solubilized membrane preparations from varioustissue and cell line sources. Solubilized membranes were run onSDS-polyacrylamide gels and then immobilized on nitrocellulosemembranes. The blots were probed with antibodies against the fol-lowing subunits: �3 (A1–A3), �4 (B1–B3), �5 (C), �2 (D), and �4(E1,E2). In A–E, lanes left to the MWM lane represent blots probedwith the particular antibodies for that panel that were preadsorbedwith a specific antigen for that antibody. The specific bands for theparticular subunit proteins are indicated by the respective arrows.Note loss of the specific bands in the respective preadsorbed blots. Ineach panel the numbers indicate the prestained molecular weightmarker (MWM) bands. The residual bands in the preadsorbed blotresults from the interaction of HRP-conjugated secondary antibodies
to membrane proteins, as evidenced by the presence of these bandswhen blotting was conducted in the absence of antinicotinic receptorsubunit antibody addition (data not shown). A2,A3: More porous gelthan A1 to show band separation; A2 prolonged exposure; A3 shortexposure. As evident in A2 and A3, the nonspecific bands (around 80kD and 52 kD) are present in both the preadsorbed and control blots,whereas the specific band is present in control blots only. Moreover, inthe autoradiogram of shorter exposure, the specific bands are moreevident than the nonspecific bands. The antibody against �4 used inB1 is from a different rabbit as compared to the anti-�4 antibody usedin B2, B3. DRG-PCC, primary cell cultures of dorsal root ganglia;MTRNR, primary cell cultures of motor neurons; PC12, pheochromo-cytoma cell line; DRG-T, dorsal root ganglia from rats; SC, rat spinalcord.
TABLE 1. Predicted Molecular Mass on the Basis of Amino Acid Sequencesof Neuronal Nicotinic Receptor Subunit Proteins and the DeterminedMolecular Masses of the Protein Bands in Western Blots that Stained
Specifically for Antibodies against the Subunit Proteins
SubunitsPredicted
molecular massDetermined
molecular mass
alpha 3 51 51alpha 4 62 55alpha 5 47 56beta 2 50 52beta 4 49 50
The predicted molecular mass does not take into account glycosylation of the subunitproteins, which may result from potential N-linked glycosylation sites in theN-terminal region of the subunit proteins.
47NICOTINIC RECEPTOR SUBUNITS IN SPINAL TISSUE

Fig. 5. Photomicrographs depicting immunofluorescence (red) fol-lowing incubation of lumbar spinal sections with anti-�3 subunit(A–H) and with anti-�4 subunit antibodies (I–P). A,B,I,J: Fluores-cence labeling in the dorsal horn region. C,D,K,J: Labeling in ventralhorn region. For each spinal region, the left panels (A,C,I,K) and rightpanels (B,D,J,L) depict immunofluorescence labeling following incu-bation of the spinal tissue sections with untreated antibodies andantibodies preadsorbed with the specific antigens, respectively. Greenand blue fluorescence in A–D and I–L represent IB4 lectin bindingand immunofluorescence labeling with anti-neuronal nuclei B (NeuNB), respectively, in the same sections. Colocalization of the �3 nAChRsubunit epitope and B4 lectin yields yellow fluorescence, as shown in
A. A lesser degree of colocalization is observed between B4 lectin andanti-�4, as shown in I. Similarly, colocalization of nAChR subunit-likeprotein and NeuN B yields purple fluorescence, which is illustrated inA,C,I,K. E–H and M–P are identical to A–D and I–L, respectively,except they depict the immunofluorescence to antinicotinic receptorsubunits only. The dorsal and ventral spinal regions imaged arerepresented by the blue and orange boxes, respectively, on the lumbarspinal cord section regional map shown in Figure 9. The photomicro-graphs shown here and in Figures 6–10 are representative of pho-tomicrographs from at least three separate experiments. Scale bars �50 �m.

booster injections of 100 �g every 3–4 weeks. Blood wascollected from the rabbit ear vein every 3–4 weeks after thethird booster. Terminal bleeding was done usually 3–4weeks after the final booster. After storage at 4°C overnight,the clotted blood was centrifuged at 10,000 rpm. The serum
was decanted and stored at –20°C until the antibody wasaffinity-purified. The same protocol was followed when therabbits were injected with GST-fusion proteins.
Approximately 2 mg of the synthetic peptide antigenwas coupled to beads using an UltraLink Immobilization
Fig. 6. Photomicrographs depicting immunofluorescence labeling(red) following incubation of dorsal lumbar spinal sections withanti-�5 (A–D), anti-�2 (E–H), and with anti-�4 (I–L) antibodies.A,E,I, and B,F,J depict immunofluorescence following incubation ofthe spinal tissue sections with untreated antibodies and antibodiespreadsorbed with the specific antigens, respectively. A,B,E,F,I,J alsoillustrate green and blue fluorescence that represent IB4 lectin bind-ing and immunofluorescence labeling with anti-neuronal nuclei B(NeuN B), respectively, in the same sections. Colocalization of nAChR
subunit-like protein and IB4 lectin yields yellow fluorescence, asshown in A,E,I. Note, in the preadsorbed section, mostly IB4 lectinand anti-NeuN B stains can be observed (B,F,J). C,D,G,H,K,L areidentical to panels A,B,E,F,I,J, respectively, except that they illus-trate only anti-subunit antibody staining. The map on the bottomright illustrates a cross section of the lumbar spinal cord. The coloredbox on the diagram of spinal cross-section shows the approximatelocation of the areas imaged. Scale bar � 50 �m.
49NICOTINIC RECEPTOR SUBUNITS IN SPINAL TISSUE
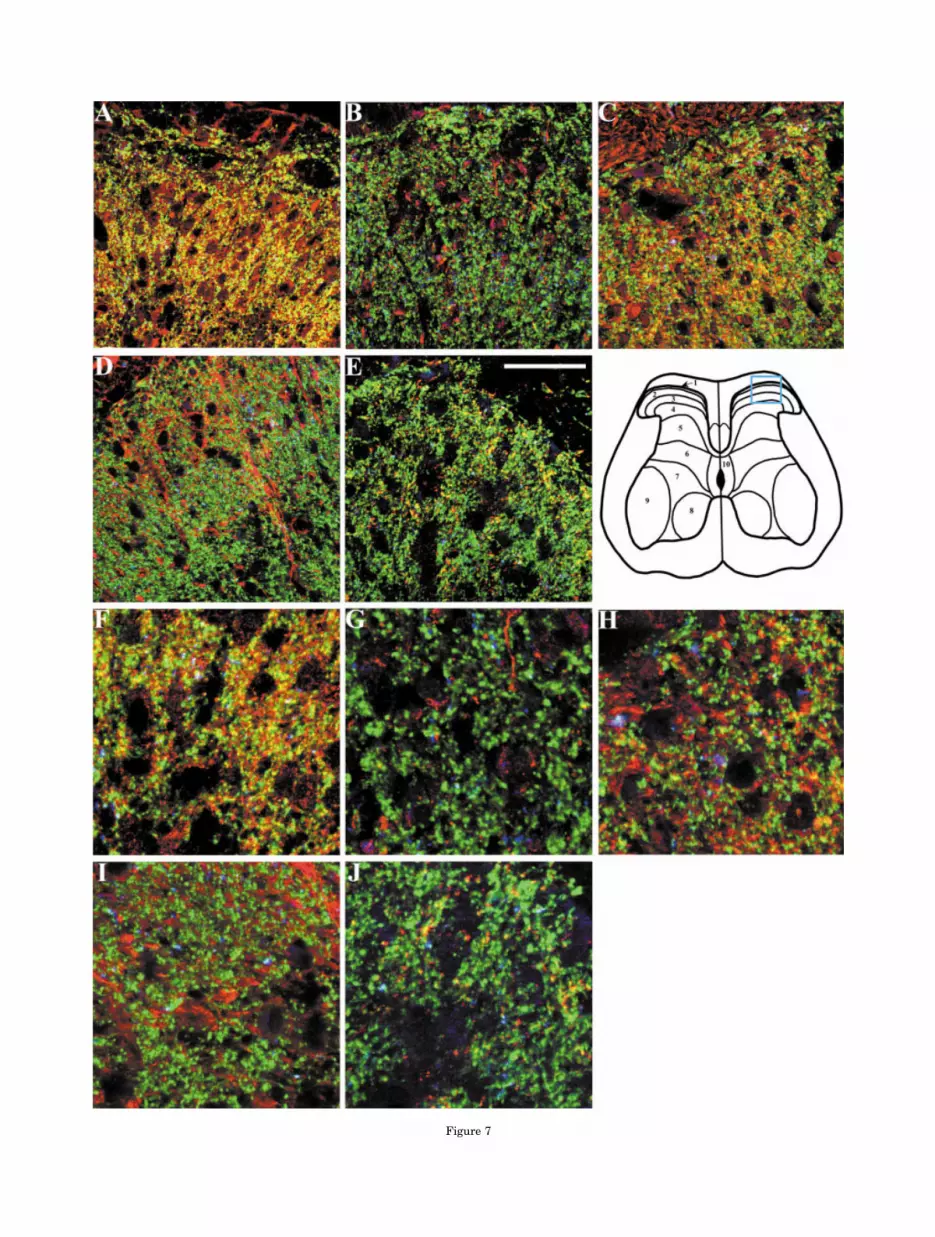
Figure 7

kit (Pierce). The immobilized peptide was packed in acolumn and washed with PBS, pH 7.4. The collected se-rum was diluted 1:10 with 0.02 M Tris.HCl, pH 7.5, andrecycled through the column for 10–12 hours with aid of apump. The column was successively washed with 0.1%Brij in 0.01M Tris.HCl, 0.1% Brij, and 500 mM NaCl in0.01 M Tris.HCl, and 500 mM NaCl in 0.01 M Tris.HClbuffers, pH 7.5. The antibody was eluted in 10 fractions bysequential additions of 2 M glycine (pH 3), a buffer washand 2 M triethylamine (pH 11). Eluants were collected inone-tenth bed volume of 1 M Tris.HCl (pH 8.0) to effectneutralization. For long-term storage, 0.04% of sodiumazide was added to the affinity-purified antibodies.
Western blotting
Tissues or cells were homogenized as previously de-scribed (Khan et al., 1997) with protease inhibitor cocktail(Sigma). After centrifugation at 15,000g, 15 min at 4°C,the pellet was solubilized with RIPA buffer (50 mM TrisBase, 150 mM NaCl, 1 mM EDTA, 0.01 M sodium-deoxycholate, 1% NP-40, 7 mM sodium dodecyl sulfate, pH8.0) with gentle rotation for 30 minutes at 4°C. Aftercentrifugation at 15,000g for 15 minutes, the supernatantwas collected. The solubilized proteins were diluted 1:1 inLaemmli sample buffer, electrophoresed on prefabricatedpolyacrylamide gels (Novex, San Diego, CA), and trans-ferred to nitrocellulose using a blot apparatus.
Blots were incubated with TBS containing 5% powderedmilk (Carnation), washed in Tris-buffered saline (TBS),pH 7.4, containing 0.05% Tween-20, and then incubatedwith primary antibodies (1:1,000 in 0.05% Tween-20 inTBS) for 1 hour. The blots were then rigorously washedand incubated with an appropriate horseradishperoxidase-conjugated antibody (1:10,000) for 1 hour. Pri-mary antibody binding was visualized using an ECLstaining kit (Pierce).
Immunohistochemical assays
For confocal microscopy, spinal cord tissue fixed in 4%paraformaldehyde was sectioned (100 �m thick) with avibratome. Tissue sections were incubated with 0.05 Mglycine in 0.1 M PBS, pH 7.5, for 30 min, washed in PBS,and incubated in blocking buffer (1% BSA, 1% cold waterfish gelatin, 1% normal donkey serum and 0.1% Triton in0.1 M PBS, pH 7.5) for 1 hour. The sections were thenincubated overnight with primary antibodies (�1 �g/ml)in working buffer (1:1 dilution of blocking buffer in PBS,pH 7.5). Other antibodies used in the assay were: anti-synaptophysin (1:75 of stock, Chemicon, Temecula, CA)anti-GFAP (1:100 of stock, Sigma) anti-VAChT (1:200 of
stock, Promega, Madison, WI) anti-NeuN B (1:200 ofstock, Chemicon) anti-bNOS (1:1,000 of stock, Sigma). Theprimary afferent nerve terminal marker isolectin B4 thatwas conjugated to fluorescein isothiocyanate (FITC) wasused at recommended concentrations (Sigma).
After incubation with primary antibodies, the sectionswere washed in working buffer and then incubated withappropriate secondary antibodies (1:75 in buffer) conju-gated to Cy™5, FITC or Lissamine-Rhodamine fluores-cence probes (Jackson ImmunoResearch, West Grove,PA). Sections were then washed, mounted on glass slides,covered with antifading reagent, and coverslips. Immuno-fluorescence labeling of the sections was observed using aBio-Rad (Hercules, CA) MRC-1024 confocal system at-tached to a Zeiss Axiovert 35 M microscope. Images col-lected were processed using Adobe PhotoShop.
To compare fluorescence intensities between two im-ages, images were collected with the same laser intensityand gain setting. Postcapture manipulation of the imagesfor gray levels and contrast were conducted with identicalsettings.
RESULTS
Localization of nicotinic binding sites inspinal cord regions by autoradiography
In situ localizations of nicotinic agonist binding sites inspinal cord regions were determined at two concentrationsof [3H]-epibatidine. Epibatidine binds to two classes ofsites in spinal cord membranes; the ratio of high-affinitysites (KD � 0.08 nM) to lower affinity sites (KD � 0.4 nM)is �2:1 (Khan et al., 1997). Thus, below 0.08 nM of [3H]-epibatidine, greater than 85% binding of the radioligandto the spinal cord tissue section will represent binding tothe high-affinity sites. However, above 0.4 nM of epibati-dine, appreciable binding will occur at both sites. Figure1A,C shows the binding of �0.05 nM epibatidine to lum-bar spinal cord sections. The higher-affinity [3H]-epibatidine binding sites are primarily located in the su-perficial dorsal horn region, the mediolateral region, andthe central canal region. High-density binding is observedin the superficial layer, where the primary afferent termi-nals are located in the lumbar spinal cord (Redmond et al.,1997).
At higher concentrations, 0.8 nM, [3H]-epibatidineshowed stronger labeling of the same areas, but also la-beled the ventral spinal cord region (Fig. 1B,D). This in-dicates that neuronal nicotinic receptors may also bepresent on motor neurons. At the higher concentrations ofepibatidine, similar regions in thoracic spinal cord sec-tions show epibatidine binding (Fig. 1E,F). Interestingly,autoradiograms from the thoracic cord also showed bind-ing to IML (intermediolateral cell column) regions (indi-cated by the yellow circle) where the autonomic sympa-thetic preganglionic fibers emerge (Dun et al., 2002).
Nicotinic receptor subunit transcripts inspinal cord tissue
RT-PCR was used to identify nicotinic receptor subunittranscripts in the spinal cord parenchyma. Since our dataindicate nicotinic receptor sites may be present on pri-mary afferent terminals, we also looked for the presence ofnAChR subunit transcripts in dorsal root ganglia (DRG).�3, �4, �5, �2, and �4 transcripts were found to be presentin both spinal cord parenchyma as well as in DRG (Fig. 2).
Fig. 7. Photomicrographs depicting immunofluorescence labelingfollowing incubation of dorsal lumbar spinal sections with a specificanti-nicotinic receptor subunit (red), anti-synaptophysin (green), andanti-VAChT (blue) antibodies. Immunofluorescence labeling withanti-�3 (A), anti-�4 (B), anti-�5 (C), anti-�2 (D), and anti-�4 (E).F–J: A �3 magnification of a selected region from panels A–E, re-spectively. The approximate location of the areas imaged (in A–E) isshown by the colored box in diagram of the spinal cord (second row).Colocalization of nAChR subunit-like protein, synaptophysin, andVAChT yields white fluorescence as shown in panels A,F, and to alesser extent in C,H, whereas colocalization of nAChR subunit-likeprotein and synaptophysin yields yellow fluorescence as shown inpanels A,C,E,F,H,J and to a lesser degree in panels B,D,G,I. Scalebar � 50 �m.
51NICOTINIC RECEPTOR SUBUNITS IN SPINAL TISSUE
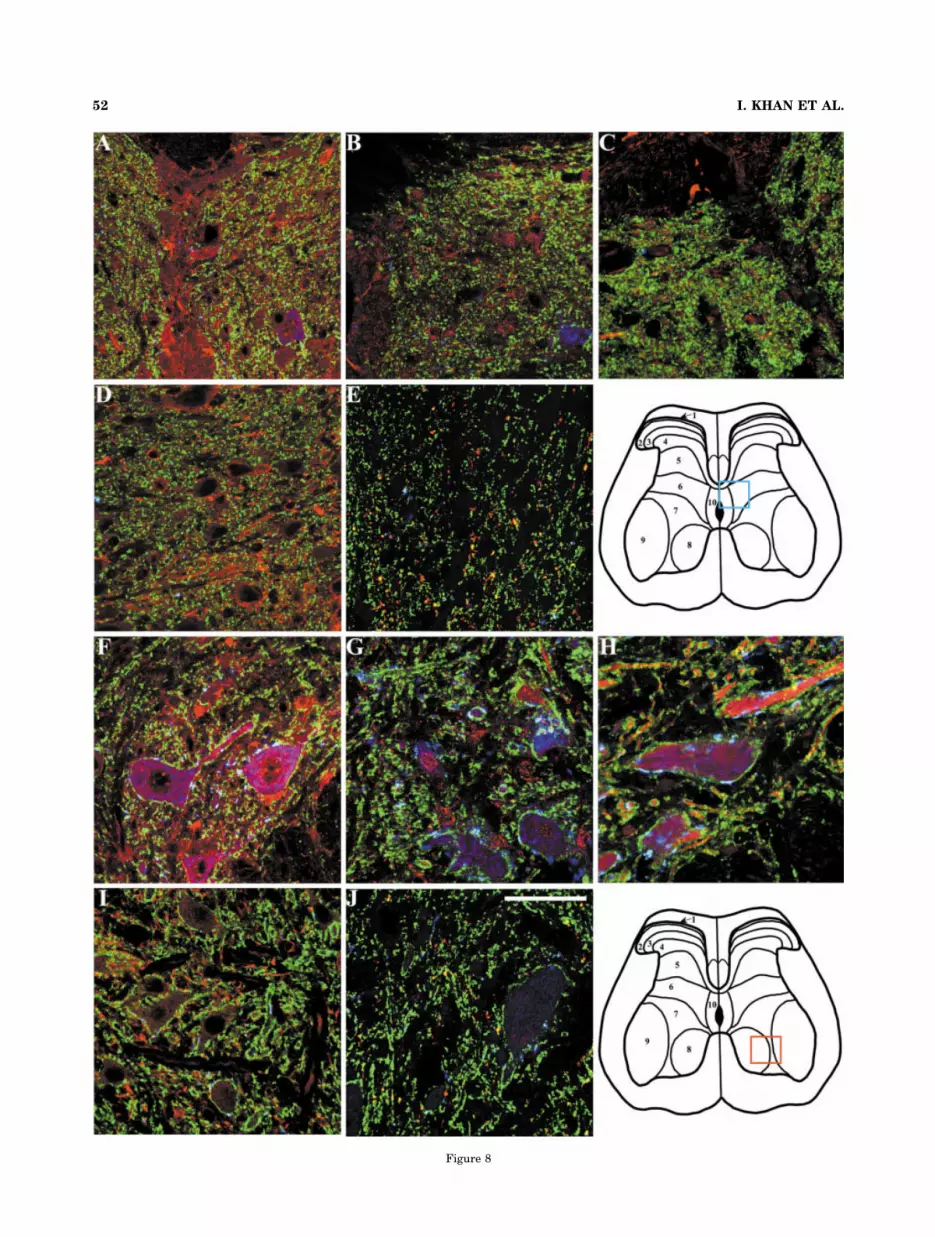
Figure 8
52 I. KHAN ET AL.

The authenticity of the RT-PCR products was confirmedby both restriction enzyme digestions and by nucleotidesequencing of the RT-PCR products (data not shown).Hence, the potential for expression of all of the subunitgenes exists in cells of the DRG and spinal cord nuclei.
Specificities of antibodies against nicotinicreceptor subunits
Expression of nicotinic receptor subunit proteins in spi-nal neurons and terminals was assessed by immunohisto-chemical assays.
Antibodies to GST-fusion proteins and to isolated
receptor peptides. Antibodies were made to the GST fu-sion proteins as well as to isolated peptides of 24–26 aminoacids from the same cytoplasmic region of the receptor sub-unit between transmembrane spans 3 and 4. For the fusionproteins, encoding sequences ranging between molecularmasses of 8.3–11.4 kDa were fused to the GST protein yield-ing fusion proteins between 34 and 37 kDa. The purifiedantibodies used in our studies selectively bound to the spe-cific antigen at a dilution of 1:1,000 or greater (stock anti-body concentration �0.5 �g/�l). Figure 3 shows an absenceof crossreactivity when the antipeptide antibodies were usedin Western blots against the various GST fusion proteinsmade from homologous subunits. Apart from some degrada-tion of the purified fusion proteins, the antibody interactionsseemed to be subunit-specific. In fact, the migrations in theblot correspond closely with the migration of the GST fusionprotein when detected by Coomassie blue staining. In ouranalysis, we compared the antibodies raised against the twoderived antigens, the GST fusion protein and the isolatedpeptide. We then employed the antibodies with the greatertiter and selectivity.
Western blots of the antibodies and preadsorption
with the GST fusion proteins. As shown in Figure 4,affinity-purified antibodies against �3, �4, �5, �2, and �4subunits also identify specific protein bands in particularcells and tissues including the spinal cord and DRG neurons.The specific bands disappear following incubation of theantibodies with an excess of the respective antigens (Fig. 4,arrow marks). The molecular masses of the specific bandsrepresenting specific nAChR subunits match well with thepredicted size from the amino acid sequences of the subunits(Table 1). Figure 4A1–4A3 shows the specific molecular formdetected by anti-�3 in various tissues and cell lines. A spe-cific band for an apparent molecular weight of 51 kD can bedetected in spinal cord tissue as well as in DRG and motorneurons. The same band size is also observed in extractsfrom PC12 cells (which constitutively express �3, �5, �2, �4,
but not �4 nAChR subunits). The band for �3 was not ob-served in HEK cell extract (data not shown). The fact thatbands of similar size are present in spinal cord tissue, DRGtissue, primary cell cultures of DRG, and motor neurons aswell as PC12 cells, but not in HEK cells, indicate that thespecific band likely represents the �3 subunit. Similarly, theantibody against �4 peptide sequence identified a specificband of apparent molecular weight of 55 kD in DRG neuronsand spinal cord tissue (Fig. 4B1–4B3). A band of same sizewas not observed in HEK or PC12 cell extracts. The antibodyagainst �4 used in panel 4B1 is from a different rabbit ascompared to the anti-�4 antibody used in panels 4B2 and4B3. As illustrated, the affinity-purified anti-�4 antibodiesfrom two different hosts identify the same molecular massproteins in rat spinal cord and DRG. For �2, �4, �5, and �3,two specific bands are apparent. This could result from dif-ferential N-glycosylation and partial proteolysis of the recep-tor subunits.
The specific bands for �2, �4, and �5 could also beobserved in PC12, but not in HEK cell extracts (data notshown). In the pre-adsorbed blots, the remaining majorbands result from the binding of secondary antibodies tothe immobilized proteins as ascertained by blotting in theabsence of primary antibody (data not shown). The ap-pearance of these nonspecific bands depends primarily onthe exposure time of the film to visualize the specific bands(also see Fig. 4 legend).
Immunohistochemical labeling in spinal cord sec-
tions with anti-subunit antibodies. Figures 5 and 6show immunolabeling for antibodies against nAChR sub-units in spinal cord sections and reveal specificity of fluores-cence labeling. Figure 5 shows the fluorescence labeling byuntreated and preadsorbed antibodies against �3 (Fig.5A–H) and �4 (5I–P) nAChR subunits. As is evident from thephotomicrograph, positive labeling by anti-�3 and anti-�4subunit antibodies in the spinal cord can be observed in thesuperficial layer of the dorsal horn (anti-�3: Fig. 5A,B,E,F;anti-�4: 5I,J,M,N), and in the ventral horn (anti-�3: Fig.5C,D,G,H; anti-�4: 5K,L,O,P). Although anti-�3 shows es-sentially no nonspecific immunolabeling, trace nonspecificfluorescence labeling by anti-�4 can be observed in bothdorsal and ventral horns of the spinal cord.
Specific fluorescence labeling in dorsal horn spinal cordsections similar to anti-�3 was also observed for antibodiesagainst �5, �2, and �4 subunit proteins (Fig. 6). Immunola-beling specificities of these three antibodies were also ob-served in the spinal ventral horn region (data not shown).However, anti-�2 shows some nonspecific fluorescence label-ing of the blood vessels in the ventral horn (data not shown).Overall, the specificities of the polyclonal antibodies againstthe nAChR subunits, as determined by Western blot andimmunohistochemical assays, indicate that they can be uti-lized to identify the localized expression of various nAChRsubunits in spinal cord regions.
The antibodies against each subunit fusion protein orpeptide antigen were raised in multiple rabbits and puri-fied antibody from at least two of these inoculated rabbitsidentified the same band in Western blots for a particularsubunit protein. These antibodies also exhibited similarimmunohistochemical staining pattern and specificity asdetermined by preadsorption. Only antibodies showingsuch matching patterns were used in these studies.
Fig. 8. Photomicrographs depicting immunofluorescence labelingfollowing incubation of central and ventral lumbar spinal sectionswith a specific anti-nicotinic receptor subunit (red), anti-synaptophysin (green), and anti-VAChT (blue) antibodies. Immuno-fluorescence labeling with anti-�3 (A,F), anti-�4 (B,G), anti-�5 (C,H),anti-�2 (D,I), and anti-�4 (E,J). A–E, and F–J represent photomicro-graphs from the central canal and ventral lumbar regions, respec-tively. The colored boxes on the diagram of spinal cross-section showthe approximate location of the areas imaged. Colocalization ofnAChR subunit-like protein, synaptophysin and VAChT yields a lim-ited amount of white fluorescence, as evident in F,H, whereas colo-calization of nAChR subunit-like protein and synaptophysin yieldsyellow fluorescence, as evident in A,C,E,F,H,J and to a lesser degreein B,D,G,I. Scale bar � 50 �m.
53NICOTINIC RECEPTOR SUBUNITS IN SPINAL TISSUE

Figure 9
54 I. KHAN ET AL.

Colocalization of nicotinic receptorsubunits with cellular markers and their
localization in various spinal regions
To determine the types of spinal neurons and terminalslabeled by the nAChR antibodies, we double- or triple-labeled the sections with nAChR subunit antibodies andspecific marker antibodies.
IB4 binding sites. Isolectin B4 is a specific marker forprimary afferent terminals (Guo et al., 1999). As shown inFigures 5 and 6, all of the five nAChR subunit proteinsexhibit colocalization with this primary afferent terminalmarker in the superficial dorsal horn region of the spinalcord (as indicated by yellow fluorescence). In multiplesections of spinal cord from individual rats, we have con-sistently observed that anti-�3, anti-�5, and anti-�4 anti-bodies colocalize with isolectin B4. However, anti-�4 andanti-�2 appear to exhibit less extensive colocalization withIB4 as compared to the other three antibodies.
Neuronal markers. All of the antibodies against thenAChR subunits exhibit some binding to potential postsyn-aptic sites on spinal neurons, as indicated by colocalization ofsubunit antibodies with anti-NeuN B (Rosenberg et al.,2001) both in the dorsal and ventral spinal horn regions(Figs. 5, 9). The subunit antibodies, particularly anti-�3,anti-�4, anti-�5, and anti-�2, also exhibit immunofluores-cence on motor neurons in the ventral spinal horn. However,the �4 subunit protein has very limited, if any, expression inthese dorsal and ventral neurons. For all of the antibodiesagainst various nicotinic receptor subunits, expression of thereceptor subunit proteins appears greater in the dorsal thanthe ventral spinal cord. This is consistent with the density ofnicotinic agonist binding sites seen autoradiographically inthese regions (Fig. 1).
Presynaptic markers. As observed in Figure 7, all fiveantibodies against specific nAChR subunits exhibit colocal-ization with synaptophysin indicating presynaptic (Ohta etal., 1993) expression of the five nicotinic receptor subunitsexamined. The presynaptic location of the nicotinic receptorsubunits also parallels the colocalization of the subunits withIB4 lectin. Accordingly, anti-�3, anti-�5, and anti-�4 showstrong colocalization with synaptophysin over a diffuse re-gion within the dorsal lumbar spinal cord. This is evidentboth in the low magnification images as well as in highermagnification of selected regions (Fig. 7). In contrast, bothanti-�4 and anti-�2 exhibit very weak colocalization withsynaptophysin in the superficial layer of the dorsal lumbarspinal cord. In the central canal and ventral spinal hornregions, colocalization of nAChR subunits with synaptophy-sin is limited, suggesting a distinct distribution (Fig. 8).
Cholinergic neuron and terminal markers. Anti-vesicular acetylcholine transporter protein (anti-VAChT),a cholinergic neuron marker (Hellstrom et al., 1999), ex-hibits colocalization with anti-�3, anti-�4, anti-�5, andanti-�2 in the dorsal spinal cord. As illustrated in Figure7, in the dorsal horn region anti-synaptophysin, anti-VAChT, and anti-nicotinic receptor subunits all colocalize,indicating that these nicotinic receptor subunit proteinsare expressed presynaptically in cholinergic neurons.However, anti-�4 and anti-�4 show only minor colocaliza-tion with both anti-synaptophysin and anti-VAChT. In thecentral canal region, although sporadic anti-VAChT fluo-rescent labeling was observed, no colocalization with anyof antinicotinic receptor subunit antibodies was evident(Fig. 8). As reported earlier by other investigators (Hell-strom et al., 1999), in the ventral horn region, in additionto labeling the motor neurons, anti-VAChT also appears tofluorescently label some presynaptic varicosities, whichare in apposition to the motor neurons. Interestingly,anti-�3 and anti-�5 are distinctively colocalized with anti-synaptophysin and anti-VAChT in these varicosities (Fig.8). Although anti-�4 and anti-�2 antibodies exhibitedfaint fluorescence labeling of the varicosities, anti-�4showed a virtual absence of labeling of the varicosities.
Glial markers. The glial marker anti-GFAP (Stuesseet al., 2001) showed strong colocalization with anti-�2 inthe dorsal horn, central canal, and ventral horn regions ofthe spinal cord. Although anti-�4 and anti-�4 exhibitedvirtually no colocalization with anti-GFAP in any of thespinal regions, significant colocalization in the dorsal hornregion was observed between GFAP and either anti-�3and anti-�5 (Figs. 9, 10).
Autonomic motor neurons. Anti-bNOS, a marker forautonomic motor neurons located in the IML region ofthoracic spinal cord (Reuss and Reuss, 2001), exhibitedstrong colocalization with anti-�3, anti-�5, and anti-�2.Colocalization of anti-�4 and anti-�4 with anti-bNOS wasfar weaker than with the other three nicotinic receptorsubunit antibodies (Fig. 10).
DISCUSSION
Nicotinic receptor binding sites inrat spinal cord
An extensive pharmacological analysis of the cardiovas-cular and behavioral responses evoked by nicotinic ago-nists and antagonist selectivity in blocking the agonistresponses indicate the existence of multiple nicotinic re-ceptor subtypes in spinal cord (Khan et al., 1994b, 1996a,1998, 2001). Membrane binding measurements corrobo-rate this multiplicity by demonstrating multiple classes ofnicotinic binding sites in the spinal cord (Khan et al.,1994c, 1997, 2001). In situ binding of [3H]-epibatidineidentifies nicotinic receptor sites throughout the spinalcord, but with far greater density in the superficial layer ofthe dorsal horn. Hence, localization of [3H]-epibatidinebinding sites overlaps the area where the primary afferentterminals project into the spinal cord. In the dorsal horn,about two-thirds of the nicotinic receptors are of the high-affinity class (Khan et al., 1997), with a prevalence in thesuperficial layer. Our findings also demonstrate specificnicotinic agonist binding sites in the IML region of tho-racic spinal cord. Assignment of a functional role for thesesites comes from observations that spinal nicotinic ago-
Fig. 9. Photomicrographs depicting immunofluorescence labelingfollowing incubation of dorsal and ventral lumbar spinal sections witha specific anti-nicotinic receptor subunit (red), anti-GFAP (green), andanti-NeuN B (blue) antibodies. Immunofluorescence labeling withanti-�3 (A,F), anti-�4 (B,G), anti-�5 (C,H), anti-�2 (D,I), and anti-�4(E,J). A–E and F–J represent photomicrographs from the dorsal andventral lumbar regions, respectively. The colored boxes on the dia-gram of spinal cross-section show the approximate location of theareas imaged. Colocalization of nAChR subunit-like protein andGFAP yields yellow fluorescence that is strongly evident in D. Yellowfluorescence is weaker, but evident in A,C,F,I, and virtually absent inE,J. The colored boxes on the diagram of spinal cross-section show theapproximate location of the areas imaged. Scale bar � 50 �m.
55NICOTINIC RECEPTOR SUBUNITS IN SPINAL TISSUE

Figure 10

nists elicit nociception and analgesia by an action on nic-otinic binding sites in the superficial layer of the lumbardorsal horn (Lawand et al., 1999; Khan et al., 2001).Similar studies suggest that the cardiovascular responsesto spinal nicotinic agonists result primarily from stimula-tion of the nicotinic binding sites on IML neurons in tho-racic spinal cord (Khan et al., 1994a).
An important question relates to the nature of the nico-tinic receptor(s) that mediates the observed behavioral ef-fects in the several spinal regions. In the gene family ofneuronal nicotinic receptors, at least eight � and three �subunits have been identified (Lukas et al., 1999; Lindstrom,2000; Bertrand, 2001). In situ hybridization studies havelocalized �3, �4, �5, �7, and �2 subunits in spinal cord tissue(Wada et al., 1989, 1990; Boyd et al., 1991). In addition to thetranscripts for the same five subunits, �4 subunit mRNA hasalso been identified in DRG neurons (Genzen et al., 2001).Recently, �2, �6, and �9 subunit transcripts have beenmapped in various rat brain regions (Lena et al., 1999;Keiger and Alker, 2000; Klink et al., 2001). These subunitshave yet to be identified in the spinal cord tissue. However,in neurons from trigeminal and dorsal root ganglia tran-scripts for �2–�7 and �2–�4 have been described (Keigerand Alker, 2000; Genzen et al., 2001). Yet the role of �2, �6,and �3 in forming functional nicotinic receptors in intactanimals remains inconclusive (Lena et al., 1999; Keiger andAlker, 2000; Sheffield et al., 2000; Klink et al., 2001). �7, �8,and �9 form homomeric nicotinic receptors (Lindstrom,2000) that are sensitive to �-bungarotoxin (Alkondon et al.,1999).
In our previous studies, antagonism of the physiologicalresponses to spinal nicotinic agonists or displacement of theradiolabeled nicotinic agonist binding with �-bungarotoxinhas been minimal (Khan et al., 1994b,c). �7 was not consid-ered initially since we noted no blockade of pressor or noci-ceptive response to spinal nicotinic agonists after intrathecal�-bungarotoxin (Khan et al., 1994b). Also, �7 receptors havebeen known to desensitize rapidly and might be expected tohave less influence on prolonged exposure to agonists (Mikeet al., 2000). Thus, we focused initially on examining theexpression of �3, �4, �5, �2, and �4 subunits in spinal neu-rons and terminals. �3, �5, and �4 subunits are encoded bya gene cluster localized to rat chromosome 8. They appear tobe candidate genes for the spontaneously hypertensive ratsphenotype (Khan et al., 2002). As reported in studies withdorsal root and peripheral ganglia (Keiger and Alker,2000; Genzen et al., 2001), we identified transcripts for
the five nicotinic receptor subunits in DRG neurons. Inspinal cord tissue, we also identified the mRNA encod-ing the five nicotinic receptor subunits. �4 transcripts inspinal cord tissues had not been previously reported.RT-PCR assays used in the present study may be moresensitive for low-level detection of �4 transcripts inspinal tissue.
Although several studies have identified multiple nico-tinic receptor subunit transcripts in spinal cord tissues orsensory ganglia (Wada et al., 1989, 1990; Boyd et al., 1991;Lena et al., 1999; Keiger and Alker, 2000; Klink et al.,2001), data actually localizing expression of the individualreceptor subunits in spinal neurons and terminals are farmore limited. Accordingly, we raised and affinity-purifiedantibodies against the �3, �4, �5, �2, and �4 subunits.Antibodies against a particular subunit appear specific forthat subunit based on the absence of crossreactivity toother GST-fusion proteins (Fig. 3).
Western blot analysis of solubilized proteins from spinalcord tissue and DRG neurons identified specific bands for theantibodies against the individual nicotinic receptor subunits.The preadsorbed bands identified molecular masses (Fig. 4,Table 1) that match the predicted size for the subunits andelectrophoretic migration reported for the identified sub-units (Elliot et al., 1996; Boulter et al., 1990; Gotti et al.,1997; Martin-Ruiz et al., 1999; Guan et al., 2000, 2001; Yehet al., 2001a,b). The molecular weights for the specific bandsidentified by antibodies against �3, �5, �2, and �4 subunitsalso closely match the molecular weight determined from theamino acid sequences as well as those reported in variousspecies, including rats (Martin-Ruiz et al., 1999; Guan et al.,2000, 2001; Gotti et al., 1997; Yeh et al 2001a,b). However,there are also observations that report molecular weights of�4 subunits that differ from those described in this study(Marubio et al., 1999; Viseshakul et al., 1998; Arroya-Jimenez et al., 1999). Arroya-Jimenez et al. (1999), utilizinga commercially available antibody against the �4 subunit,identified a 75 kD protein band in Western analysis from ratbrain extract. Other investigators, using the same antibody,identified a protein band of similar size, but also identifiedthe major protein bands to be smaller than 75 kD in ratbrain tissue (Martin-Ruiz et al., 1999). Owing to autolyticdegradation processes during harvesting, storage, and prep-aration of protein samples containing the �4 nAChR sub-unit, more than one molecular form of this subunit proteinmay be identified in Western or similar analytical proce-dures (Madhock et al., 1992; Martin-Ruiz et al., 1999).
Overall, the data presented here emphasize that theantibodies are specific for the specific nicotinic receptorsubunit proteins. This assertion is based on several obser-vations: 1) the respective antibodies do not show crossre-activity with other GST fusion proteins; 2) the antibodiesidentify specific proteins in Western assays that are sim-ilar in size to that of predicted size based on amino acidsequences; 3) staining appears differentially distributedin particular tissues and cells; 4) two antibodies raised indifferent rabbits against a particular subunit typicallyshow remarkably similar staining patterns; and 5) thestaining pattern of our antibodies against �3, �4, and �2 iscomparable to that using commercially available antibod-ies (data not shown can be downloaded from http://phrtayl0.ucsd.edu/ikhan) (Marubio et al., 1999; Arroya-Jimenez et al., 1999; Guan et al., 2000). Thus, while it isimpossible to exclude absolutely that other proteins maybe recognized, several convergent observations provide
Fig. 10. Top panels (A–E): Photomicrographs depicting immunoflu-orescence labeling following incubation of central lumbar spinal sectionswith a specific anti-nicotinic receptor subunit (red), anti-GFAP (green),and anti-NeuN B (blue) antibodies. Immunofluorescence labeling withanti-�3 (A), anti-�4 (B), anti-�5 (C), anti-�2 (D), and anti-�4 (E). Colo-calization of nAChR subunit-like protein and GFAP yields yellow fluo-rescence that is evident primarily in D. The colored box on the diagramof spinal cross-section show the approximate location of the areas im-aged. Bottom panels (F–J): Photomicrographs depicting immunofluores-cence labeling following incubation of the intermediolateral (IML) cellcolumn region of the thoracic spinal cord with a specific anti-nicotinicreceptor subunit (red) and anti-bNOS (green) antibodies. Immunofluo-rescence labeling with anti-�3 (F), anti-�4 (G), anti-�5 (H), anti-�2 (I),and anti-�4 (J). Colocalization of nAChR subunit-like protein and bNOSyields yellow fluorescence that is evident in F,H,I. The approximatelocation of the areas imaged is shown by the colored box in the autora-diogram illustrating [3H]-epibatidine binding to the thoracic spinal cordsection (bottom right panel). Scale bars � 50 �m.
57NICOTINIC RECEPTOR SUBUNITS IN SPINAL TISSUE

substantial support for the specificity of the antibodies forthe respective nicotinic receptor subunits.
Nicotinic receptor subunit expression invarious spinal regions
Similar to the Western blots, immunolabeling with thesepolyclonal antibodies appears to be antigen-specific, as evi-denced by loss of fluorescence labeling following preadsorp-tion of the antibodies with specific antigens. Only the anti-�2antibody shows some nonspecific fluorescence localizedaround vascular walls that cannot be preadsorbed.
Immunolabeling for the five anti-subunit antibodies ismost intense in the dorsal horn region, followed by the cen-tral and ventral horn regions. In the dorsal horn, the immu-nolabeling is particularly strong in the superficial layer com-prising laminae II and III. However, anti-�4 also appears tohave strong fluorescence labeling in the central canal region.In the dorsal horn region, based on overlap with anti-synaptophysin (a presynaptic marker) fluorescence, all of theantibodies exhibited presynaptic fluorescence labeling. Lo-calization at presynaptic sites for �3, �5, and �4 appearsgreater than for �4 and �2 in the superficial layer. Except foranti-�4 subunit antibody, the presynaptic localization wasmost apparent in the dorsal horn.
In the superficial layer of the dorsal horn, we also observedsignificant colocalization between anti-synaptophysin, apresynaptic marker (Ohta et al., 1993) and anti-VAChT, amarker for cholinergic neurons, an observation made pre-viously in spinal cord tissue (Hellstrom et al., 1999). Ourdata demonstrate that all of the five antibodies also fluo-rescently label cholinergic terminals in the dorsal hornregion. These cholinergic presynaptic terminals may rep-resent primary afferent terminals (Sann et al., 1995) aswell as presynaptic terminals of glycinergic inhibitory in-terneurons (Ribeiro-Da-Silva and Cuello, 1990). This isconsistent with the findings that nociceptive and anti-nociceptive responses to intrathecal nicotinic agonistsmay be due to the spinal release of excitatory (Khan et al.,1996a, 1997) and inhibitory (Khan et al., 2001; Kiyosawaet al., 2001) neurotransmitters, respectively. Further-more, expression of nicotinic receptor subunits on theseterminals also suggests that a large fraction of the nico-tinic receptors are positioned presynaptically to modulatethe release of acetylcholine from these terminals.
In addition to their superficial layer distribution, �3, �4,�5, and �2 subunits show presynaptic localization in theventral horn area, particularly in the synaptic varicositiesthat fluorescently label for cholinergic markers. Theseterminals are identified as cholinergic type C-terminals(Hellstrom et al., 1999). The functional significance ofthese terminals is at present uncertain.
Besides the presynaptic terminals, the antibodies, exceptfor anti-�4, also localize with potential postsynaptic sites inthe dorsal and ventral horns, intermediolateral cell column(IML), and the central canal spinal regions. Hence, nicotinicreceptors are present on motor neurons in the ventral hornand autonomic motor neurons in the IML region. The latterlocalization would suggest that they play a role in pregan-glionic sympathetic activity, an observation consistent withintrathecal nicotinic agonist administration directly affect-ing blood pressure and heart rate (Khan et al., 1994a).
Although most of the fluorescence labeling for the fiveantibodies was localized on the presynaptic terminals andthe postsynaptic neurons, anti-�3, anti-�5, and �2 anti-bodies show fluorescence labeling on some glial cells. Thisobservation is intriguing, since glial cells can regulate the
extracellular concentrations of several transmitters andhormones, including excitatory amino acids and prosta-glandins (Schousboe, 2003; Svensson and Yaksh, 2002),which are known to play a major facilitatory role in spinalnociceptive transmission (Yaksh et al., 1999).
Overall, the immunolabeling patterns of the antibodiesshow a close correlation with the expression of the nicotinicreceptors in the spinal cord, demonstrated by receptor auto-radiography (Fig. 1). Nicotinic receptors are expressed prin-cipally on the primary afferent terminals as well as otherpresynaptic terminals in the superficial dorsal horn region.Moreover, nicotinic receptor expression appears in interneu-rons in the dorsal horn region and the central canal region.These observations are consistent with our previous findingssuggesting that these postsynaptic neurons are a secondarysite for nicotinic agonists eliciting nociceptive and antinoci-ceptive responses (Khan et al., 1998, 2001). Recently, a dif-ferential distribution of various subtypes of nicotinic recep-tors was found by Perry et al. (2002) using 125I-epibatidineand competition with other nicotinic agonists showing par-tial receptor selectivity. Our present observations combiningligand autoradiography and subunit selective antibodies lo-calize further spinal receptor distribution in regional as wellas synaptic sites.
In conclusion, the data presented here provide a de-tailed map to localize expression of nicotinic receptor sub-unit proteins in spinal terminals and neurons. Determi-nation of receptor localization is critical to analyzing theinfluence of selective lesioning on receptor expression.Further studies involving immunoprecipitation and char-acterization of the subunit composition of the immunopre-cipitates will be required to determine which of the sub-units are involved in assembling the composite nicotinicreceptors in the discrete spinal locations.
LITERATURE CITED
Alkondon M, Pereira EF, Eisenberg HM, Albuquerque EX. 1999. Cholineand selective antagonists identify two subtypes of nicotinic acetylcho-line receptors that modulate GABA release from CA1 interneurons inrat hippocampal slices. J Neurosci 19:2693–2705.
Arroyo-Jimenez MM, Bourgeois J-P, Marubio LM, Le Sourd AM, OttersenOP, Rinvik E, Fairen A, Changeux J-P. 1999. Ultrastructural localiza-tion of the �4-subunit of the neuronal acetylcholine nicotinic receptorin the rat substantia nigra. J Neurosci 19:6475–6487.
Azam L, Winzer-Serhan UH, Chen Y, Leslie FM. 2002. Expression ofneuronal nicotinic acetylcholine receptor subunit mRNAs within mid-brain dopamine neurons. J Comp Neurol 444:260–274.
Bertrand LS. 2001. Neuronal nicotinic receptors: from structure to func-tion. Nicotine Tob Res 3:203–223.
Bertrand D, Ballivet M, Rungger D. 1990. Activation and blocking ofneuronal nicotinic acetylcholine receptor reconstituted in Xenopus oo-cytes. Proc Natl Acad Sci U S A 87:1993–1997.
Boulter J, Connolly J, Denneris E, Goldman D, Heinemann S, Patrick J.1987. Functional expression of two neuronal nicotinic acetylcholinereceptors from cDNA clones identifies a gene family. Proc Natl Acad SciU S A 84:7763–7767.
Boulter J, O’Shea-Greenfield A, Duvoisin RM, Connolly JG, Wada E, JensenA, Gardner PD, Ballivet M, Deneris ES, McKinnon D. 1990. �3, �5, and�4: Three members of the rat neuronal nicotinic acetylcholine receptor-related gene family form a gene cluster. J Biol Chem 265:4472–4482.
Boyd RT, Jacob MH, McEachern AE, Caron S, Berg DK. 1991. Nicotinicacetylcholine receptor mRNA in dorsal root ganglion neurons. J Neu-robiol 22:1–14.
Dun SL, Ng YK, Brailoiu GC, Ling EA, Dun NJ. 2002. Cocaine- andamphetamine-regulated transcript peptide-immunoreactivity in ad-renergic C1 neurons projecting to the intermediolateral cell column ofthe rat. J Chem Neuroanat 23:123–132.
Elliot KJ, Ellis SB, Berckhan KJ, Urrutia A, Chavez-Noriega LE, JohnsonED, Velicelebli G, Harpold, MM. 1996. Comparative structure of hu-
58 I. KHAN ET AL.

man neuronal �2-�7 and �2-�4 nicotinic acetylcholine receptor sub-units and functional expression of the �2, �3, �4, �7, �2 and �4subunits. J Mol Neurosci 7:217–228.
Galzi J-L, Revah F, Bessis A, Changeux J-P. 1991. Functional architectureof the nicotinic acetylcholine receptor: from electric organ to brain.Annu Rev Pharmacol 31:37–72.
Genzen JR, Van Cleve W, McGehee DS. 2001. Dorsal root ganglion neuronsexpress multiple nicotinic acetylcholine receptor subtypes. J Neuro-physiol 86:773–782.
Gotti C, Moretti M, Maggi R, Longhi R, Hanke W, Klinke N, Clementi F.1997. �7 and �8 nicotinic receptor subtypes immunopurified from chickretina have different immunological, pharmacological and functionalproperties. Eur J Neurosci 9:1201–1211.
Guan Z-Z, Zhang X, Ravid R, Nordberg A. 2000. Decreased protein levels ofnicotinic receptor subunits in the hippocampus and temporal cortex ofpatients with Alzheimer’s disease. J Neurochem 74:237–243.
Guan Z-Z, Zhang X, Mousavi M, Tian J-Y, Unger C, Nordberg A. 2001.Reduced expression of neuronal nicotinic acetylcholine receptors dur-ing the early stages of damage by oxidative stress in PC12 cells.J Neurosci Res 66:551–558.
Guo A, Vulchanova L, Li WX, Elde R. 1999. Immunocytochemical localiza-tion of the vanilloid receptor 1 (VR1): relationship to neuropeptides, theP2X3 purinoceptor and IB4 binding sites. Eur J Neurosci 11:946–958.
Hellstrom J, Arvidsson U, Elde R, Cullheim S, Meister B. 1999. Differentialexpression of nerve terminal protein isoforms in VAChT-containing vari-cosities of the spinal cord ventral horn. J Comp Neurol 411:578–590.
Keiger CJH, Alker JC. 2000. Individual variation in the expression profilesof nicotinic receptors in the olfactory bulb and trigeminal ganglion andidentification of �2, �6, �9, and �3 transcripts. Biochem Pharmacol59:233–240.
Khan IM, Taylor P, Yaksh TL. 1994a. Stimulatory pathways and sites ofactions of intrathecally administered nicotinic agonists. J PharmacolExp Ther 271:1550–1557.
Khan IM, Taylor P, Yaksh TL. 1994b. Cardiovascular and behavioralresponses to nicotinic agonists administered intrathecally. J Pharma-col Exp Ther 270:150–158.
Khan IM, Yaksh TL, Taylor P. 1994c. Ligand specificity of nicotinic ace-tylcholine receptors in rat spinal cord: studies with nicotine and cy-tisine. J Pharmacol Exp Ther 270:159–166.
Khan IM, Marsala M, Printz MP, Taylor P, Yaksh TL. 1996a. Intrathecalnicotinic agonist-elicited release of excitatory amino acids as measured byin vivo spinal microdialysis in rats. J Pharmacol Exp Ther 278:97–106.
Khan IM, Youngblood KL, Yaksh TL, Printz MP, Taylor P. 1996b. Spinalnicotinic receptor expression in spontaneously hypertensive rats. Hy-pertension 28:1093–1098.
Khan IM, Yaksh TL, Taylor P. 1997. Epibatidine binding sites and activityin the spinal cord. Brain Res 753:269–282.
Khan IM, Buerkle H, Taylor P, Yaksh TL. 1998. Nociceptive and antino-ciceptive responses to intrathecally administered nicotinic agonists.Neuropharmacology 37:1515–1525.
Khan IM, Stanislaus S, Zhang L, Taylor P, Yaksh TL. 2001. A-85380 andepibatidine each interact with disparate spinal nicotinic receptor sub-types to achieve analgesia and nociception. J Pharmacol Exp Ther297:230–239.
Khan IM, Singletary E, Alemayehu A, Stanislaus S, Printz MP, Yaksh TL, TaylorP. 2002. Nicotinic receptor gene cluster on rat chromosome 8 in nociceptiveand blood pressure hyperresponsiveness. Physiol Genom 11:65–72.
Kiyosawa A, Katsurabayashi S, Akaike N, Pang ZP, Akaike N. 2001.Nicotine facilitates glycine release in the rat spinal dorsal horn.J Physiol 536:101–110.
Klink R, de Kerchove d’Exaerde A, Zoli M, Changeux J-P. 2001. Molecularand physiological diversity of nicotinic acetylcholine receptors in themidbrain dopaminergic nuclei. J Neurosci 21:1452–1463.
Lawand NB, Lu Y, Westlund KN. 1999. Nicotinic cholinergic receptors:potential targets for inflammatory pain relief. Pain 80:291–299.
Lena C, d’Exaerde AK, Cordero-Erausquin M, Nicolas N, Arroyo-JimenezMM, Changeux JP. 1999. Diversity and distribution of nicotinic ace-tylcholine receptors in the locus ceruleus neurons. Proc Natl Acad SciU S A 96:12126–12131.
Lindstrom J. 2000. The structure of neuronal nicotinic receptors. In: Clem-enti F, Fornasari D, Gotti C. editors. Neuronal nicotinic receptors. NewYork: Springer. p 101–162.
Lukas RJ, Changeux J-P, Le Novere N, Albuquerque EX, Balfour DJK,Berg DK, Bertrand D, Chiappinelli VA, Clarke PBS, Collins AC, DaniJA, Grady SR, Kellar KJ, Lindstrom JM, Marks MJ, Quik M, TaylorPW, Wonnacott S. 1999. International Union of Pharmacology. XX.
Current status of the nomenclature for nicotinic acetylcholine receptorsand their subunits. Pharmacol Rev 51:397–401.
MacDermott AB, Role LW, Siegelbaum SA. 1999. Presynaptic ionotropicreceptors and the control of transmitter release. Annu Rev Neurosci22:443–485.
Madhock TC, Bjercke RJ, Langone JJ, Sharp BM. 1992. Brain nicotinicreceptors isolated by a monospecific antibody against a synthetic �3subunit receptor peptide compared to a monoclonal anti-idiotypic (tonicotine) antibody. Biochem Biophys Res Commun 182:303–1308.
Martin-Ruiz CM, Court JA, Molnar E, Lee M, Gotti C, Mamalaki A, TsouloufisT, Tzartos S, Ballard C, Perry RH, Perry EK. 1999. �4 but not �3 and �7nicotinic acetylcholine receptor subunits are lost from the temporal cortexin Alzheimer’s Disease. J Neurochem 73:1635–1640.
Marubio LM, Arroyo-Jimenez MM, Cordero-Erausquin M, Lena C, NovereNL, d’Exaerde AK, Huchet M, Damaj MI, Changeux J-P. 1999. Re-duced antinociception in mice lacking neuronal nicotinic receptor sub-units. Nature 398:805–810.
Mike A, Castro NG, Albuquerque EX. 2000. Choline and acetylcholine havesimilar kinetic properties of activation and desensitization on the alpha7nicotinic receptors in rat hippocampal neurons. Brain Res 882:155–168.
Ohta M, Kitamoto T, Iwaki T, Ohgami T, Fukui M, Tateishi J. 1993.Immunohistochemical distribution of amyloid precursor protein duringnormal rat development. Dev Brain Res 75:151–161.
Perry DC, Xiao Y, Nguyen HN, Musachio JL, Davila-Garcia MI, Kellar KJ. 2002.Measuring nicotinic receptors with characteristics of �4�2, �3�2 and �3�4subtypes in rat tissues by autoradiography. J Neurochem 82:68–481.
Redmond L, Xie H, Ziskind-Conhaim L, Hockfield S. 1997. Cues intrinsic tothe spinal cord determine the pattern and timing of primary afferentgrowth. Dev Biol 182:205–218.
Reuss MH, Reuss S. 2001. Nitric oxide synthase neurons in the rodentspinal cord: distribution, relation to Substance P fibers, and effects ofdorsal rhizotomy. J Chem Neuroanat 21:181–196.
Ribeiro-Da-Silva A, Cuello AC. 1990. Choline acetyltransferase-immunoreactive profiles are presynaptic to primary sensory fibers inthe rat superficial dorsal horn. J Comp Neurol 295:370–384.
Rosenberg GA, Cunningham LA, Wallace J, Alexander S, Estrada EY,Grossetete M, Razhagi A, Mille RK, Gearing A. 2001. Immunohisto-chemistry of matrix metalloproteinases in reperfusion injury to ratbrain: activation of MMP-9 linked to stromelysin-1 and microglia in cellcultures. Brain Res 893:104–112.
Sann H, McCarthy PW, Mader M, Schemann M. 1995. Cholineacetyltransferase-like immunoreactivity in small diameter neurons ofthe rat dorsal root ganglion. Neurosci Lett 198:17–20.
Schousboe A. 2003. Role of astrocytes in the maintenance and modulationof glutamatergic and GABAergic neurotransmission. Neurochem Res28:347–352.
Sheffield EB, Quick MW, Lester RAJ. 2000. Nicotinic acetylcholine recep-tor subunit mRNA expression and channel function in medial habenulaneurons. Neuropharmacology 39:2591–2603.
Stuesse SL, Crisp T, McBurney DL, Schechter JB, Lovell JA, Cruce WLR.2001. Neuropathic pain in aged rats: behavioral responses and astro-cytic activation. Exp Brain Res 137:219–227.
Svensson CI, Yaksh TL. 2002. The spinal phospholipase-cyclooxygenase-prostanoid cascade in nociceptive processing. Annu Rev PharmacolToxicol 42:553–583.
Viseshakul N, Figl A, Lytle C, Cohen BN. 1998. The �4 subunit of rat �4�2nicotinic receptors is phosphorylated in vivo. Mol Brain Res 59:100–104.
Wada E, Wada K, Boulter J, Deneris E, Heinemann S, Patrick J, SwansonLW. 1989. Distribution of alpha2, alpha3, alpha4 and beta2 neuronalnAChR subunit mRNAs in the central nervous system: a hybridizationhistochemical study in the rat. J Comp Neurol 284:314–335.
Wada E, McKinnon D, Heinemann S, Patrick J, Swanson LW. 1990. Thedistribution of mRNA encoded by a new member of the neuronalnicotinic acetylcholine receptor gene family (�5) in the rat centralnervous system. Brain Res 526:45–53.
Yaksh TL, Hua XY, Kalcheva I, Nozaki-Taguchi N, Marsala M. 1999. Thespinal biology in humans and animals of pain states generated bypersistent small afferent input. Proc Natl Acad Sci USA 96:7680–7686.
Yeh JJ, Yasuda P, Davila-Garcia MI, Xiao Y, Ebert S, Gupta T, Keller K,Wolfe BB. 2001a. Neuronal nicotinic acetylcholine �3 subunit protein inrat brain and sympathetic ganglion measured using a subunit-specificantibody: regional and ontogenic expression. J Neurochem 77:336–346.
Yeh JJ, Ferreira M, Ebert S, Yasuda RP, Kellar KJ, Wolfe BB. 2001b.Axotomy and nerve growth factor regulate levels of neuronal nicotinicacetylcholine receptor �3 subunit in the rat superior cervical ganglion.J Neurochem 79:258–265.
59NICOTINIC RECEPTOR SUBUNITS IN SPINAL TISSUE