altered hippocampal expression of neuropeptides in seizure-prone galr1 knockout mice: hippocampal...
TRANSCRIPT

Epilepsia, 44(8):1022–1033, 2003Blackwell Publishing, Inc.C© 2003 International League Against Epilepsy
Altered Hippocampal Expression of Neuropeptidesin Seizure-prone GALR1 Knockout Mice
∗Sergueı O. Fetissov, †Arie S. Jacoby, ∗Pablo R. Brumovsky, †John Shine,†Tiina P. Iismaa, and ∗Tomas Hokfelt
∗Department of Neuroscience, Karolinska Institutet, Stockholm, Sweden; and †The Garvan Institute of Medical Research,St. Vincent’s Hospital, Sydney, Australia
Summary: Purpose: Mice carrying a deletion of the GALR1galanin receptor have recently showed spontaneous seizure phe-notype with 25% penetrance. To better understand the role ofneuropeptides, which are known to undergo complex plasticitychanges with development of epileptic seizures, we character-ized their expression in the hippocampal formation in GALR1-knockout (-KO) mice with or without seizures and in wild-type(WT) mice.
Methods: Immunohistochemistry and in situ hybridizationwere used to study expression of galanin, neuropeptide Y(NPY), substance P, enkephalin, dynorphin, and cholecystokinin(CCK).
Results: In GALR1-KO mice that had been displayingseizures, a strong upregulation of galanin immunoreactivity (ir)and messenger RNA (mRNA) was found in the polymorph layerof the dentate gyrus; galanin-ir also appeared in a dense fibernetwork in the supragranular layer. A strong upregulation ofenkephalin was found in the granule cells/mossy fibers, whereas
dynorphin mRNA levels were modestly decreased. NPY wasstrongly expressed in the granule cells/mossy fibers, and an in-crease of NPY mRNA levels in the polymorph cells was paral-leled by an increase of NPY-ir in the molecular layer. An up-regulation of substance P-ir was confined to the fibers in thegranule and molecular layers, whereas substance P mRNA wasincreased in the cells of the polymorph layer. Both CCK-ir andmRNA were strongly downregulated in the granule cell/mossyfiber system, but CCK-ir appeared increased in the supragran-ular and molecular layers. No changes in neuropeptide-ir werefound in GALR1-KO mice not displaying seizures.
Conclusions: Complex changes in neuropeptide expression insome principal hippocampal neurons and interneurons appear asa characteristic feature of the spontaneous-seizure phenotype inGALR1-KO mice. However, to what extent causal relations existbetween this “epilepsia peptidergic profile” and developmentof seizures requires further clarification. Key Words: Brain—Epilepsy—Peptidergic plasticity.
Galanin, a 29-aa neuropeptide (1), is widely distributedthroughout the rat (2) and mouse (3) brain. Three recep-tor subtypes for galanin (GALR1, GALR2, and GALR3)have been cloned (4,5). Evidence has been found for in-volvement of galanin and its receptors in multiple neuronaland neuroendocrine functions (6,7). Galanin has been re-ported to have inhibitory properties in several pathwaysin the central nervous system (8–13), including exertingan antiepileptic effect. Thus galanin injected in the hip-pocampal formation has a powerful anticonvulsant effectin rats (14,15), whereas mice ectopically overexpressinggalanin by using different gene promoters display resis-tance for status epilepticus (SE) and kindling epileptogen-esis (16,17). In contrast, mice lacking galanin expressionshow increased propensity to develop SE (16). Recentlymice carrying a deletion of GALR1 have been generated
Accepted May 1, 2003.Address correspondence and reprint requests to Dr. S.O. Fetissov at
Department of Neuroscience B3:4, Karolinska Institutet, Retzius vag. 8,17177, Stockholm, Sweden. E-mail: [email protected]
and phenotypically characterized, and both male and fe-male GALR1 knockout (-KO) mice exhibited spontaneousseizures with a penetrance of 25%, ranging from facialmovements to SE (18).
It has been demonstrated in a number of studies that ex-pression of neuropeptides is dramatically changed, mostlyincreased, in animal models for epilepsy and that this oc-curs in specific subregions, especially in the hippocampalformation (19–22). For example, enkephalin is stronglyupregulated in the granule cell/mossy fiber system (23),and neuropeptide Y (NPY) is apparently aberrantly ex-pressed in the same cells (24,25), whereas cholecystokinin(CCK) is downregulated in the mossy fibers (23). Galaninis upregulated in mossy cells and in the supragranular layer(15,17).
In this work, to better understand the role of peptider-gic plasticity in the development of the seizure-pronephenotype, we used immunohistochemistry and in situhybridization to characterize expression of hippocampalneuropeptides in GALR1-KO mice, to see if any of the
1022
HIPPOCAMPAL NEUROPEPTIDES IN GALR1-KO MICE 1023
neuropeptide changes described in various epileptic an-imal models (see earlier) also occur in the GALR1-KOmouse.
METHODS
Animals and tissue preparationGALR1-KO mice were generated by insertional mu-
tagenesis of the gene encoding GALR1, resulting in ab-sence of normal full-length transcript (18). Spontaneousseizure phenotype occurred with a penetrance of 25%in mice homozygous for mutation on C57BL/6J back-ground, starting at age 11.1 ± 0.2 weeks, with 27% mor-tality (18). Some GALR1-KO mice (75%) were foundnot to develop any seizures when exposed to the standardconditions that consistently evoked seizures in GALR1-KO seizure-prone mice, such as bright overhead lightingor handling. C57BL/6J wild-type (WT) mice were usedas controls. Before killing, 6-month-old mice had beenhoused under controlled environmental conditions withconstant light–dark cycle (light on between 6:00 and 18:00hr), a temperature of 21–22◦C, and a relative humidity of40–50%; food and water were given ad libitum. The daybefore killing, between 16:00 and 17:00, seizure-pronephenotype in GALR1-KO mice was confirmed by brightoverhead lighting, resulting mainly in clonic unilateral orbilateral seizures lasting for ∼1 min. The next day between9:00 and 10:00, GALR1-KO mice (n = 6), which had ex-hibited seizures in their anamnesis but were not in SE andwere not observed to have a seizure in the morning beforekilling (referred to later as GALR1-KO unless specified),GALR1-KO mice that did not display seizures (n = 3), andWT mice (n = 6) were anaesthetized with sodium pento-barbital (0.15 mg/100 g body weight, i.p.) and perfusedvia the ascending aorta with Tyrode’s Ca2+-free solutionat 37◦C, followed by a mixture of 4% paraformaldehydeand 0.4% picric acid in 0.16 M phosphate buffer (pH 6.9),37◦C and then by the same, but ice-cold mixture. The
TABLE 1. Intensity of neuropeptide immunoreactivity in the hippocampal formation of seizure-prone GALR1-KO and WT mice
Galanin NPY SP Enkephalin Dynorphin CCKHippocampalformation KO WT KO WT KO WT KO WT KO WT KO WT
Dentate gyrusGranule cell layer ND ND + ND ++ + ++ +/− +/− +/− +/− +/−Mossy fibers ND ND +++ ND ND ND +++ + ++ ++ +/− ++Polymorph cells ++ +/− + ++ +/− +/− ND ND ND ND ++ +Supragranular layer ++++ +/− + ND + ND ND ND ND ND ++++ +++
HippocampusMolecular layer +/− +/− ++ +/− ++ +/− ND ND ND ND + +/−Stratum lacunosum mol. + ++ +++ + +/− ND + + ND ND ++ +/−Stratum radiatum +/− +/− ++ +/− +/− +/− +/− +/− +/− +/− +++ +++Stratum oriens +/− +/− + +/− + +/− + + +/− +/− +++ +++
Pyramidal cell layersCA1 (cells) ND ND +/− +/− +/− +/− +/− + ND +/− ++ +CA1 (fibers) +/− + +/− +/− ++ +/− +/− + ND +/− ++++ +++CA3 (cells) ND ND +/− ND +/− ND ND ND ND ND + +/−CA3 (fibers) +/− + +/− +/− + ND + +/− + +/− +++ ++
brains were rapidly dissected out, immersed in the samefixative for 90 min, and rinsed with 10% sucrose in 0.1M phosphate buffer (pH 7.4) overnight. Brains were snap-frozen with CO2. Coronal, 14-µm thick brain sectionswere cut on a cryostat (Microm, Heidelberg, Germany),and thaw-mounted on chrome alum-gelatin–coated glassslides.
ImmunohistochemistryThe tyramide signal amplification (TSA) immunohis-
tochemical method (26) and rabbit polyclonal antiseraraised against the following peptides were used: galanin(27), NPY (Walsh J. and H. Wong, unpublished), sub-stance P (28), enkephalin (29), dynorphin (30), and CCK(31). Guinea pig anti–γ -aminobutyric acid (GABA) an-tiserum (1:400; Chemicon, Temecula, CA, U.S.A.) andCCK (1:400) antiserum (31) were used for double stain-ing after processing for the TSA method. Rabbit anti-serum against zinc transporter-3 (ZnT-3), 1:400, was usedas a marker for mossy fibers (32). Incubation with pri-mary antisera (1:4,000) overnight at 4◦C was followedby horseradish peroxidase–conjugated, swine anti-rabbitimmunoglobulin G (IgG; 1:100; Dako A/S, Copenhagen,Denmark) and incubations according to the TSA-PlusFluorescein System protocol (DuPont, New England Nu-clear, Boston, MA, U.S.A.). The specificity of antibodieswas tested by preabsorption tests with an excess (10−6
or 10−5M) of the corresponding peptide, purchased fromPeninsula (Belmont, CA, U.S.A.) or Bachem (Bissendorf,Switzerland). Sections were mounted in a mixture of glyc-erol and 0.1 M phosphate-buffered saline (3:1), pH 7.4,containing 0.1% 1,4-phenylenediamine (Sigma Chemi-cals, St. Louis, MO, U.S.A.) as antifading agent.
After processing, the sections were examined in aRadiance Plus confocal laser scanning system (Bio-Rad,Hemel Hemstead, U.K.), installed on an Eclipse E600fluorescence microscope (Nikon, Tokyo, Japan). Digitalimages resulting from the confocal scanning microscopy
Epilepsia, Vol. 44, No. 8, 2003
1024 S. O. FETISSOV ET AL.
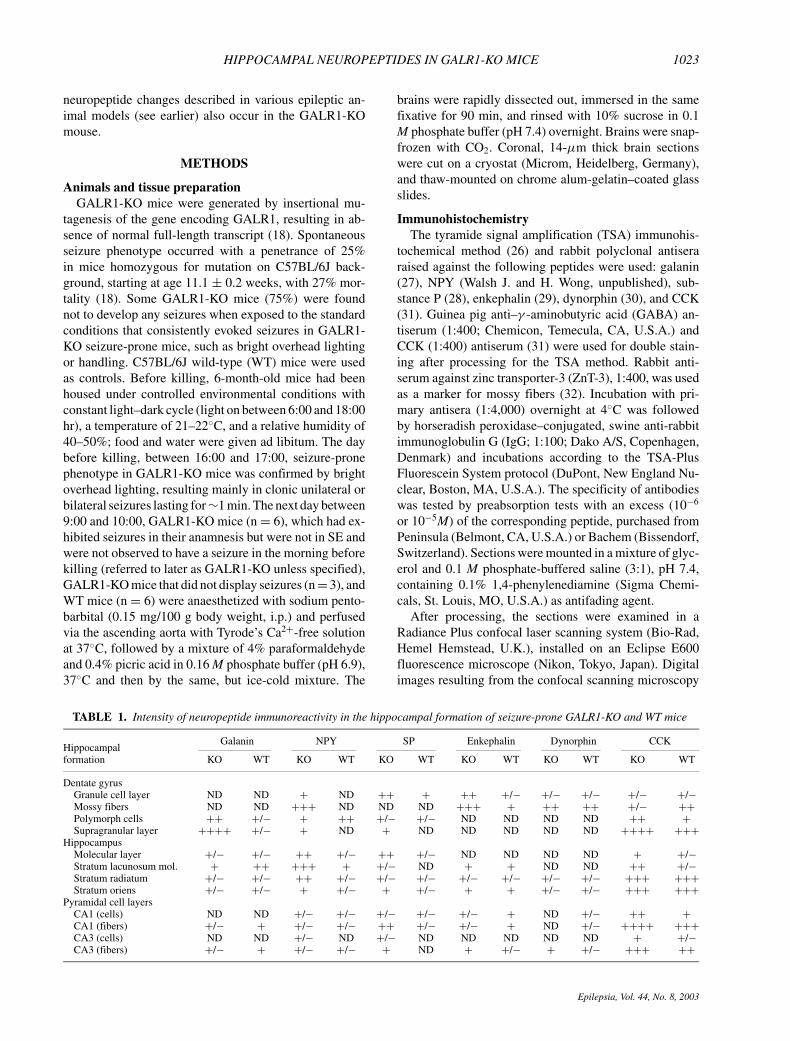
FIG. 1. Galanin (Gal) and chole-cystokinin (CCK) immunoreactivity(-ir) in the dorsal hippocampal for-mation of wild-type (WT) (A) andseizure-prone GALR1-KO (B) miceafter processing for double labeling.Note a strong yellow band reflect-ing overlapping galanin-ir and CCK-ir in the supragranular layer in theKO (B) but not WT (A) mouse. Co-existence (arrowheads) of galanin-irand CCK-ir was found in mossy cellsof the polymorph layer in GALR1-KO mice (C–E). Arrow , a galanin-positive, CCK-negative fiber. Scalebars: 100 µm (A, B); 20 µm (C–E). CA1, CA3, Ammon’s horn, fieldsCA1, CA3; gr, granule cell layer; mf,mossy fibers; mo, molecular layer;po, polymorph layer; sg, supragran-ular layer; slm, stratum lacunosummoleculare; sr, stratum radiatum;so, stratum oriens.
were optimized for image resolution or merged for colorillustrations in PhotoShop 6.0 (Adobe Systems Incorpo-rated, San Jose, CA, U.S.A.). A subjective estimation ofintensity of immunostaining was made in the microscopeby using a rating scale from not detectable (ND), very low(±), low (+), medium (++), high (+++), and very high(++++) levels (Table 1).
In situ hybridizationBetween 9:00 and 10:00, GALR1-KO mice (n = 5) that
had exhibited seizures in their anamnesis according to con-ditions described earlier, and WT mice (n = 5) were killedby decapitation. The brains were dissected, immersedin ice-cold phosphate-buffered saline, immediately there-after frozen, cut at 14-µm thickness with a cryostat (Mi-crom), and thaw-mounted onto “Probe On” slides (FisherScientific, Pittsburgh, PA, U.S.A.).
Antisense oligonucleotides probes complementary tonucleotides 152–199 of galanin mRNA (33), 322–360 ofenkephalin mRNA (34), 1,671–1,714 of NPY mRNA (35),298–341 of CCK mRNA (36), 145–192 of preprotachykinmRNA (37), and 871–918 of dynorphin mRNA (38) weresynthesized by CyberGene AB (Huddinge, Sweden). Theoligonucleotides were labeled at the 3′ end by using ter-minal deoxynucleotidyltransferase (Amersham, Bucking-hamshire, U.K.) with [α-35S]dATP (NEN, Boston, MA,U.S.A.) to a specific activity of 1–4 × 106 cpm/ng oligonu-cleotide. The labeled probes were purified through QIAquick-spin columns (Qiagen/GmbH, Hilden, Germany).Dithiothreitol (DTT) was added to a final concentrationof 10 mM. Sections were hybridized as described pre-
viously (39,40). In brief, air-dried sections were incu-bated in a hybridization buffer [50% formamide, 4× SSC,1 × Denhardt’s solution (1% sarcosyl, 0.02 M phosphatebuffer, 10% dextran sulfate), 500 µg/ml heat-denaturedsalmon sperm DNA, 200 mM DTT, 1 × 107 cpm/ml ofthe labeled probes] in a humidified chamber for 16–18 h at42◦C. After hybridization, the sections were washed in 1×SSC 4 ×15 min at 55◦C and 30 min at room temperature,and then air-dried and dipped into Ilford K5 nuclear emul-sion (Ilford, Mobberly, U.K.) diluted 1:1 with water. Afterexposure at 4◦C for 2–6 days, the slides were developedin D19 (Kodak, Rochester, NY, U.S.A.), fixed in Kodak3000, and mounted in glycerol-phosphate buffer. For thecontrol of specificity of in situ hybridization results, ad-jacent sections were incubated with an excess (×100) ofunlabelled probe, none of which yielded a hybridizationsignal.
Every seventh section from a series through the rostro-caudal extension of the hippocampus, giving in total 10sections for each mRNA, was examined by using a NikonEclipse E600 fluorescence microscope equipped with adark-field condenser. Digital images acquired with NikonDXM1200 digital still camera (using a ×20 objective)were analyzed for intensity of mRNA labeling in ScionImage 4.0 (Frederick, MD, U.S.A.). Each captured imagewas calibrated to 256-pixel grey values (0 = white and256 = black), and every labeled neuronal profile in thepolymorph cell layer was individually selected, and itsmean pixel density was measured. A rectangular selectionover the granule layer upper blade with 200-µm long and100-µm short side was used for the quantification of the
Epilepsia, Vol. 44, No. 8, 2003
HIPPOCAMPAL NEUROPEPTIDES IN GALR1-KO MICE 1025
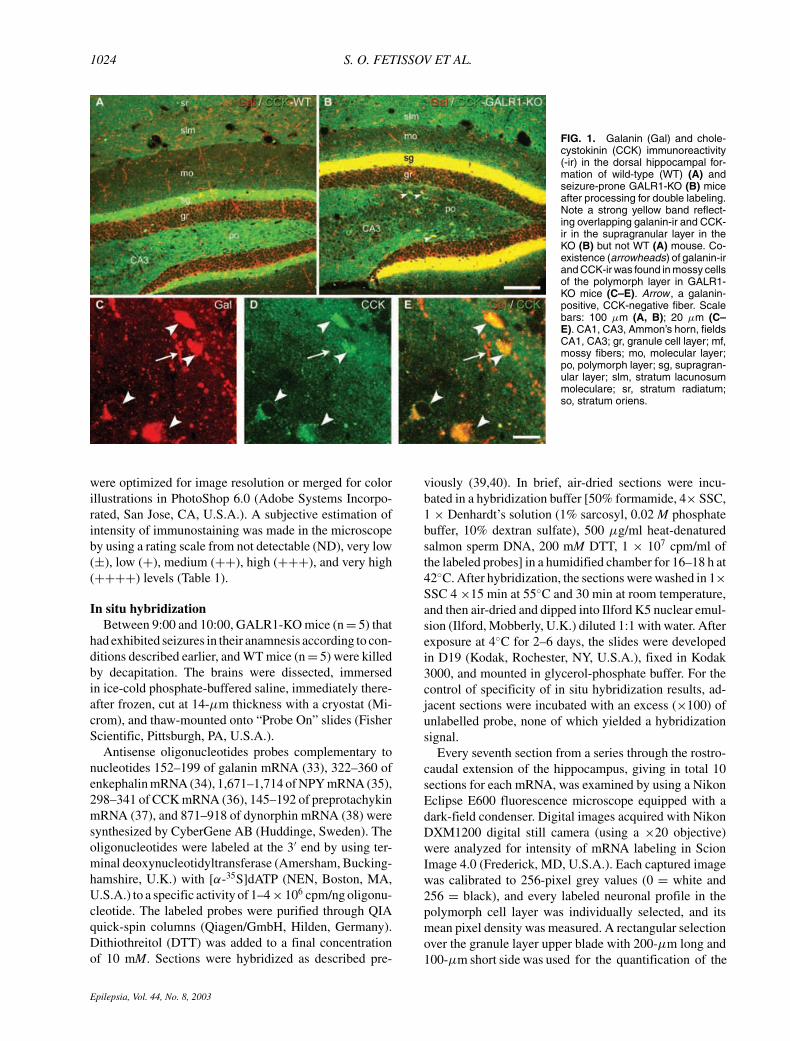
FIG. 2. Galanin-ir in the dorsal hippocampal formation of wildtype (WT) (A, C, E, G), seizure-prone GALR1-KO (B, D, F, H)mice and GALR1-KO mice without seizures (nsz) (I). Note a strongincrease of galanin-ir in the supragranular layer (sg) (B, D) andin cells in the polymorph layer (po) in seizure-prone GALR1-KOs(D). Cells indicated by arrows in C and D correspond to the cellsin G and H, respectively. Scale bar: 100 µm (A, B); 50 µm (C, D);5.5 µm (E–H); 20 µm (I). CA1, CA3, Ammon’s horn, fields CA1,CA3; gr, granule cell layer; mf, mossy fibers; mo, molecular layer;po, polymorph layer; sg, supragranular layer; slm, stratum lacuno-sum moleculare; sr, stratum radiatum; so, stratum oriens.
single in the granule cells. The background levels weremeasured in separate images from outside of the sectionarea. We assumed that silver grain density overlying neu-rons correlates directly with their level of mRNA expres-sion. To estimate silver grain density, the mean pixel den-sity was converted into the relative optic density (ROD)by using formula–log [(256 grey value)/256]. The back-ground ROD level was calculated in the same way andwas ultimately subtracted from the neuronal ROD. Un-paired t test was used to compare means of ROD betweenGALR1-KO and control groups of mice. Data expressedas a percentage of mean ROD relative to the control groupequaled 100% ± SD.
RESULTS
The expression of galanin, enkephalin, NPY, CCK, sub-stance P, and dynorphin peptides and mRNA was studied atdifferent rostrocaudal levels of the hippocampal formationof seizure-prone GALR1-KO mice and compared with thepatterns seen in WT C57BL/6J mice, by using immunohis-tochemistry and in situ hybridization. In terms of generalanatomy and structure of the hippocampal formation, noobvious differences were found between the two groupsof mice. However, it appeared that, in the dorsal dentategyrus, the upper and lower blades of granule cell layerswere more separated in the GALR1-KO than in WT mice,so that in the former, the angle between the two bladeswas significantly larger (27 ± 3.2 vs. 15.5 ± 1.8 degrees;p < 0.001). In the majority of cases, the changes in mRNAand peptide levels were parallel, but the changes in tran-scripts appeared less pronounced than the effects on pep-tide levels. The results of neuropeptide immunostainingin the dorsal hippocampal formation are summarized inTable 1 and shown in Figs. 1–6, and changes in neuropep-tide mRNA levels are demonstrated in Fig. 7. Figure 8shows results from incubation with antiserum to ZnT3, amarker for mossy fibers.
In GALR1-KO mice that did not display seizures,immunohistochemical detection of galanin (Fig. 2I),enkephalin (Fig. 3I), NPY (Fig. 4G), CCK (Fig. 5I), sub-stance P (Fig. 6E), and dynorphin (Fig. 6H) revealed nor-mal distribution of these neuropeptides in the hippocampalformation without apparent differences in the intensity ofstaining when compared with WT mice.
GalaninA moderately dense galanin-positive fiber network was
fairly evenly distributed in the hippocampal formationof WT mice (Figs. 1A and 2A). A marked increase wasseen in intensity of the supragranular layer in GALR1-KOmice, whereas a general decrease was noted of galanin-positive fibers in the hippocampal formation, including thegranule cell layer (cf. Figs. 1B; 2B, D, F with Figs. 1A and2A, C, E). Numerous large neurons with intense galanin
Epilepsia, Vol. 44, No. 8, 2003
1026 S. O. FETISSOV ET AL.
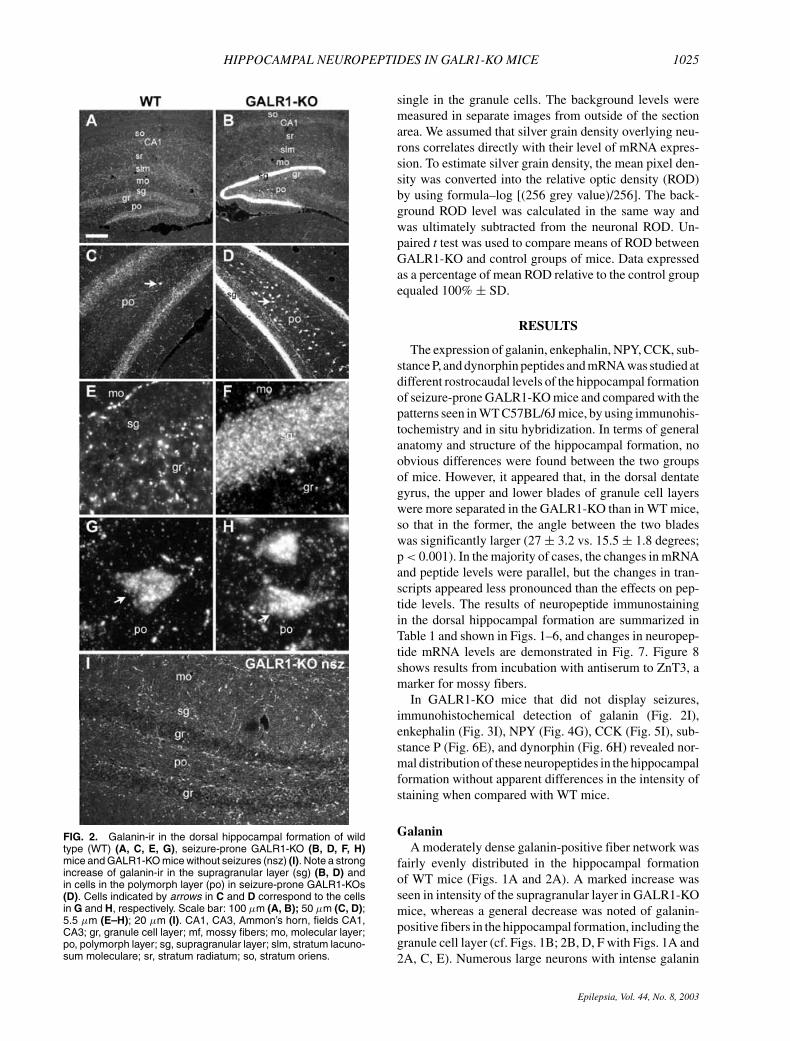
FIG. 3. Enkephalin-ir in the dorsal hippocampal formation ofwild-type (WT) (A, C, E), seizure-prone GALR1-KO (B, D, F) miceand GALR1-KO mice without seizures (nsz) (G). Note a strong in-crease of enkephalin-ir in the granule cells (gr) and mossy fibers(mf) in seizure-prone GALR1-KOs (B, D, F). The enkephalin-positive cells are mainly localized in the external part of the gran-ule cell layer (gr) (E, F). Scale bar: 100 µm (A, B); 50 µm (C, D);5,5 µm (E–F); 20 µm (G). CA3, Ammon’s horn, field CA3; gr,granule cell layer; mf, mossy fibers; po, polymorph layer.
immunoreactivity (-ir) were present, especially in thecaudal parts of the polymorph layer of the GALR1-KOmice (Fig. 2D), whereas fewer such neurons appearedin the WT mice (Fig. 2C). These galanin-ir cells were
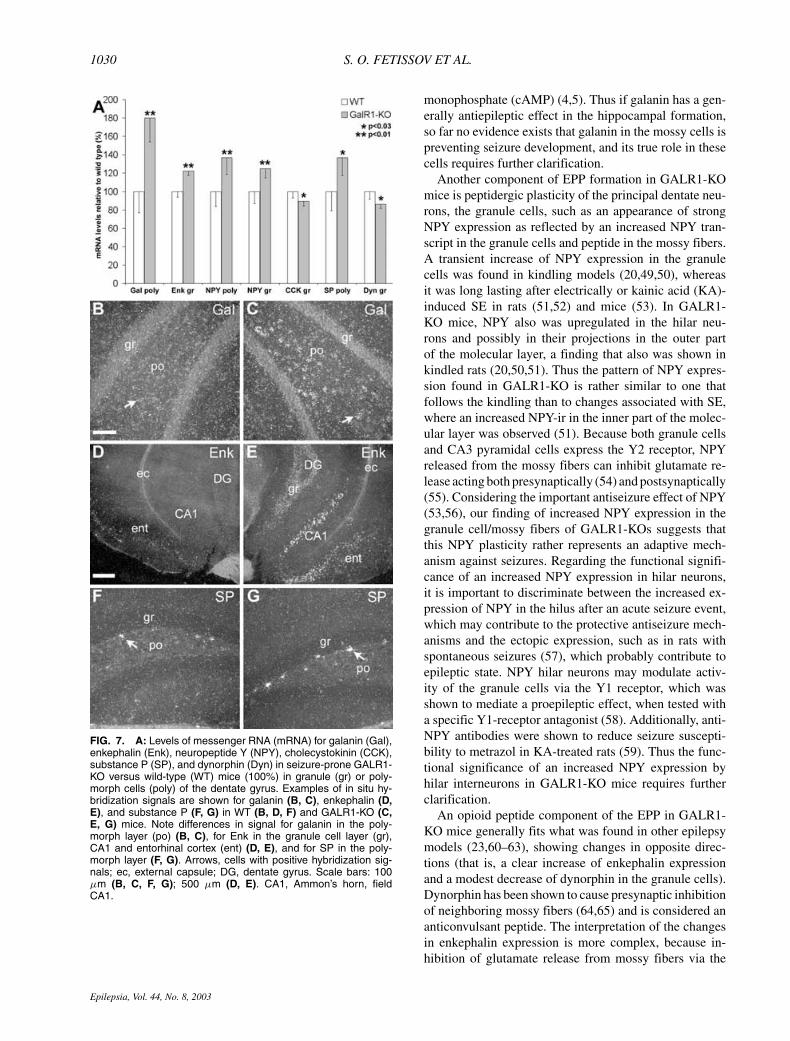
morphologically similar in both groups of mice (Fig. 2Gand H). Moreover, double-labeling experiments showedthat the galanin cells also are CCK positive (Fig. 1C–E),and that the intense bands of galanin- and CCK-ir stronglyoverlap (Fig. 1A and B). In the confocal microscope,coexistence of galanin- and CCK-ir also could be ob-served in nerve endings in the supragranular layer. Ad-ditionally, galanin-positive polymorph neurons did notdisplay GABA-ir. In situ hybridization showed a strongincrease of the galanin mRNA signal in cells of the poly-morph layer in GALR1-KO mice (Fig. 7A; cf. Fig. 7Band C).
EnkephalinA prominent enkephalin expression was present in the
mossy fiber system in both WT and GALR1-KO mice(Fig. 3A and B), but in the KO mice, enkephalin-ir wasmarkedly increased in the hilar region and in stratum lu-cidum, reflecting its high content in mossy fibers (cf. 3Band D with 3A and C). Because of the strong and diffuseenkephalin-ir in the hilar region, it was not clear if anyof the polymorph cells were enkephalin positive. How-ever, we could not detect a distinct enkephalin mRNAsignal in the polymorph layer. In contrast, the granulecells, particularly in the upper blade, displayed an in-creased enkephalin-ir, these cells being predominantly lo-cated in the external part of the granule cell layer (Fig.3F). In accordance with the peptide change, enkephalinmRNA expression was significantly increased in gran-ule cells in GALR1-KO mice, being most prominent inthe ventral part of the dentate gyrus (Fig. 7A; cf. Fig.7D with 7E). Increased transcript levels were also notedin the CA1 region and in the entorhinal cortex (cf. 7Dwith 7E).
Neuropeptide YNPY-positive fibers and interneurons were evenly
distributed in the WT mouse hippocampal formation(Fig. 4A and C). In contrast, in GALR1-KO mice, NPY-ir was increased in several hippocampal/dentate regions,such as the hilus and mossy fibers in stratum lucidum, inthe external part of the molecular layer, and in strata la-cunosum moleculare, radiatum, and oriens (Fig. 4B andD). An increase in the number of NPY-positive fibers wasseen in the granule cell layer (cf. Fig. 4F with 4E). Thesechanges in NPY-ir were corroborated by significantly ele-vated levels of NPY mRNA in both the granule and poly-morph cells (Fig. 7A).
CholecystokininThe WT mice displayed a wide expression of CCK-
ir in the hippocampal formation (Figs. 1A and 5A),whereas GALR1-KO mice showed a mostly similarbut quantitatively different distribution, with an increasein the molecular layer and in the stratum lacunosum
Epilepsia, Vol. 44, No. 8, 2003

HIPPOCAMPAL NEUROPEPTIDES IN GALR1-KO MICE 1027
FIG. 4. Neuropeptide Y immunoreactivity (NPY-ir) in the dor-sal hippocampal formation of wild-type (WT) (A, C, E), seizure-prone GALR1-KO (B, D, F) mice and GALR1-KO mice withoutseizures (nsz) (G). Note that NPY-ir is present in the mossy fibersin seizure-prone GALR1-KO (D) mice. Increases occur but areless pronounced in other layers, including more fibers in the gran-ule cell layer (gr) (E, F). Scale bar: 100 µm (A, B); 50 µm (C, D);5.5 µm (E-F), 20 µm (G). CA3, Ammon’s horn, field CA3; gr, gran-ule cell layer; mf, mossy fibers; mo, molecular layer; po, polymorphlayer.
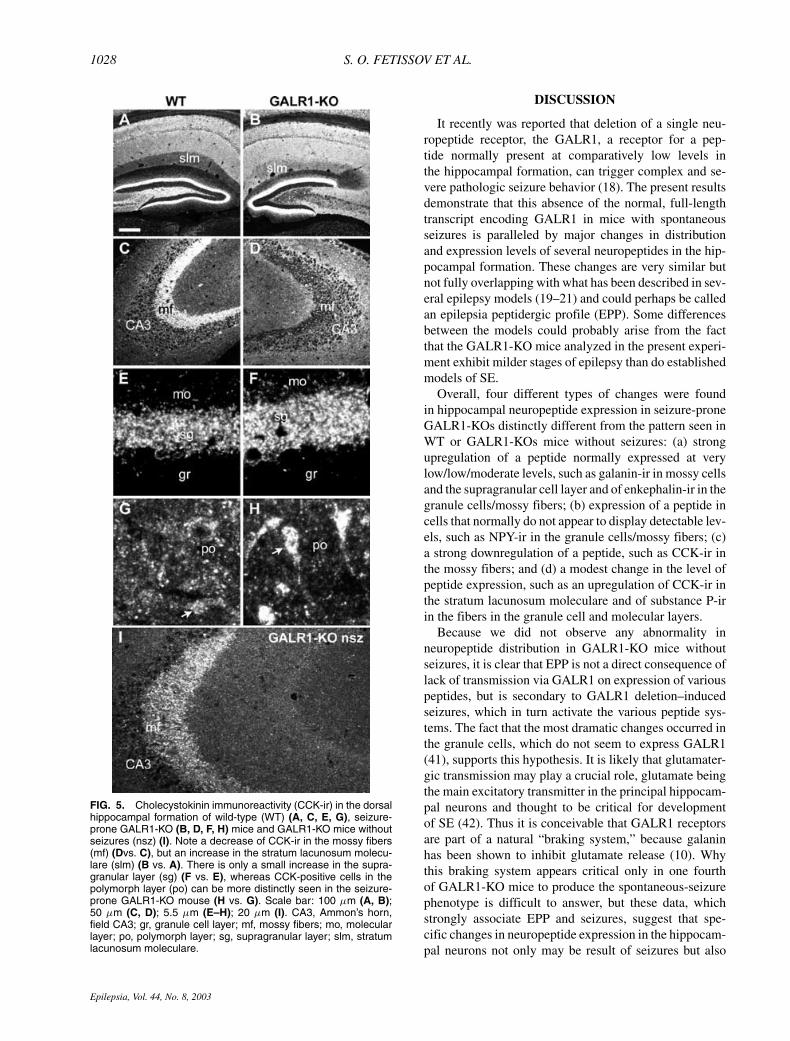
moleculare (Figs. 2B and 5B). The most conspicuouschange, however, was a depletion of CCK-ir in the mossyfiber system, as could be most clearly seen in the stra-tum lucidum (cf. Fig. 5D with 5C). The decrease in the
density of mossy fiber staining in the hilar region ofGALR1-KO mice allowed easy visualization of CCK-positive cells in the polymorph layer (cf. Fig. 5H with5G; see also Fig. 1C–E). These cells were also presentin the WT mice but more weakly fluorescent and, again,masked by dense CCK-positive mossy fiber network andcould be visualized by adjusting the conditions of laserscanning in confocal microscope. Also the supragranu-lar layer in GALR1-KO mice appeared to have a some-what increased CCK-ir (cf. Fig. 5F with 5E); CCK mRNAlevels were not changed significantly in the polymorphcell layer, but it was decreased in the granule cell layer(Fig. 7A).
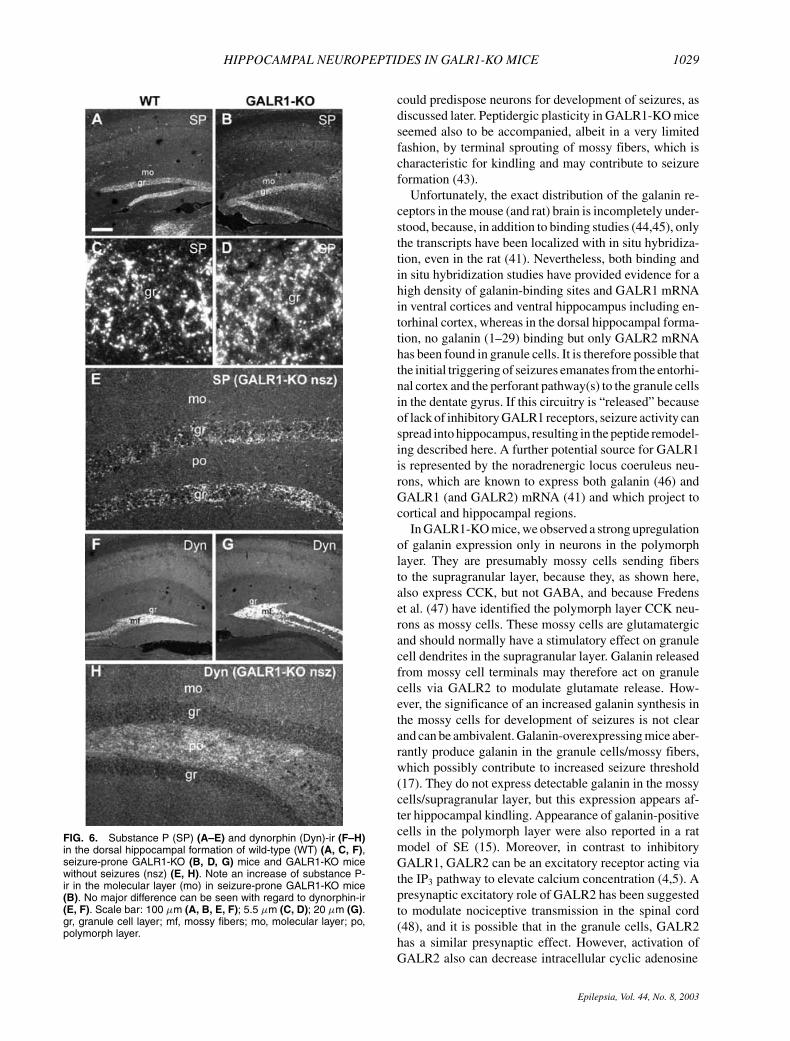
Substance PDot-like substance P-ir was detected in the granule
cell layer of both WT (Fig. 6A and C) and GALR1-KO(Fig. 6B and D) mice, presumably in fibers surround-ing granule cells, because no substance P–positive mossyfibers were found in either type of mouse (Fig. 6Aand B). The intensity of this immunostaining was, how-ever, stronger in GALR1-KO mice (cf. Fig. 6D withC), and in this mouse, a substance P–positive termi-nal network also was found in the molecular layer(Fig. 6B), whereas it was hardly detected in the WTmice (Fig. 6A). In accordance with the peptide change,the substance P (preprotachykinin) mRNA level wasincreased in cells in the polymorph layer, but wasnot detected in the granule cell layer (cf. Fig. 7Gwith F).
DynorphinIn both WT and GALR1-KO mice, the only promi-
nent dynorphin-ir in the hippocampal formation wasdetected in the mossy fiber system (Fig. 6E and F).Even though the intensity of dynorphin immunos-taining appeared unchanged in GALR1-KO mice, insitu hybridization revealed a decrease of the dynor-phin mRNA signal in the granule cells in KO mice(Fig. 7A).
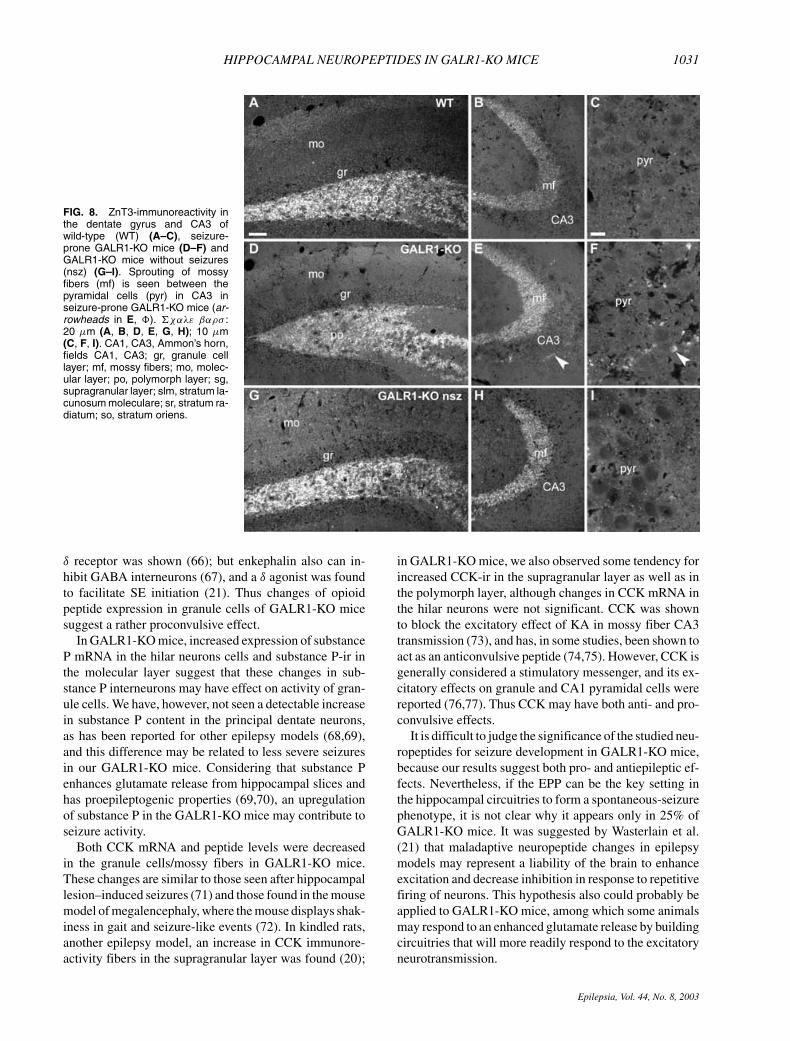
ZnT3Immunohistochemical analysis detected ZnT3 in the
mossy fibers as previously reported (32). No appar-ent difference in ZnT3-ir granules density in the gran-ule cell layer was seen between WT and GALR1-KO mice, suggesting none or a very low level ofcollateral sprouting of mossy fibers (Fig. 8D). How-ever, in the CA3 area, more numerous and continu-ous ZnT3-ir dots were found in seizure-prone GALR1-KO mice (Fig. 8E,F), but not in WT or GALR1-KOmice without seizures, indicating the presence of terminalsprouting of mossy fibers in seizure-prone GALR1-KOmice.
Epilepsia, Vol. 44, No. 8, 2003
1028 S. O. FETISSOV ET AL.
FIG. 5. Cholecystokinin immunoreactivity (CCK-ir) in the dorsalhippocampal formation of wild-type (WT) (A, C, E, G), seizure-prone GALR1-KO (B, D, F, H) mice and GALR1-KO mice withoutseizures (nsz) (I). Note a decrease of CCK-ir in the mossy fibers(mf) (Dvs. C), but an increase in the stratum lacunosum molecu-lare (slm) (B vs. A). There is only a small increase in the supra-granular layer (sg) (F vs. E), whereas CCK-positive cells in thepolymorph layer (po) can be more distinctly seen in the seizure-prone GALR1-KO mouse (H vs. G). Scale bar: 100 µm (A, B);50 µm (C, D); 5.5 µm (E–H); 20 µm (I). CA3, Ammon’s horn,field CA3; gr, granule cell layer; mf, mossy fibers; mo, molecularlayer; po, polymorph layer; sg, supragranular layer; slm, stratumlacunosum moleculare.
DISCUSSION
It recently was reported that deletion of a single neu-ropeptide receptor, the GALR1, a receptor for a pep-tide normally present at comparatively low levels inthe hippocampal formation, can trigger complex and se-vere pathologic seizure behavior (18). The present resultsdemonstrate that this absence of the normal, full-lengthtranscript encoding GALR1 in mice with spontaneousseizures is paralleled by major changes in distributionand expression levels of several neuropeptides in the hip-pocampal formation. These changes are very similar butnot fully overlapping with what has been described in sev-eral epilepsy models (19–21) and could perhaps be calledan epilepsia peptidergic profile (EPP). Some differencesbetween the models could probably arise from the factthat the GALR1-KO mice analyzed in the present experi-ment exhibit milder stages of epilepsy than do establishedmodels of SE.
Overall, four different types of changes were foundin hippocampal neuropeptide expression in seizure-proneGALR1-KOs distinctly different from the pattern seen inWT or GALR1-KOs mice without seizures: (a) strongupregulation of a peptide normally expressed at verylow/low/moderate levels, such as galanin-ir in mossy cellsand the supragranular cell layer and of enkephalin-ir in thegranule cells/mossy fibers; (b) expression of a peptide incells that normally do not appear to display detectable lev-els, such as NPY-ir in the granule cells/mossy fibers; (c)a strong downregulation of a peptide, such as CCK-ir inthe mossy fibers; and (d) a modest change in the level ofpeptide expression, such as an upregulation of CCK-ir inthe stratum lacunosum moleculare and of substance P-irin the fibers in the granule cell and molecular layers.
Because we did not observe any abnormality inneuropeptide distribution in GALR1-KO mice withoutseizures, it is clear that EPP is not a direct consequence oflack of transmission via GALR1 on expression of variouspeptides, but is secondary to GALR1 deletion–inducedseizures, which in turn activate the various peptide sys-tems. The fact that the most dramatic changes occurred inthe granule cells, which do not seem to express GALR1(41), supports this hypothesis. It is likely that glutamater-gic transmission may play a crucial role, glutamate beingthe main excitatory transmitter in the principal hippocam-pal neurons and thought to be critical for developmentof SE (42). Thus it is conceivable that GALR1 receptorsare part of a natural “braking system,” because galaninhas been shown to inhibit glutamate release (10). Whythis braking system appears critical only in one fourthof GALR1-KO mice to produce the spontaneous-seizurephenotype is difficult to answer, but these data, whichstrongly associate EPP and seizures, suggest that spe-cific changes in neuropeptide expression in the hippocam-pal neurons not only may be result of seizures but also
Epilepsia, Vol. 44, No. 8, 2003
HIPPOCAMPAL NEUROPEPTIDES IN GALR1-KO MICE 1029
FIG. 6. Substance P (SP) (A–E) and dynorphin (Dyn)-ir (F–H)in the dorsal hippocampal formation of wild-type (WT) (A, C, F),seizure-prone GALR1-KO (B, D, G) mice and GALR1-KO micewithout seizures (nsz) (E, H). Note an increase of substance P-ir in the molecular layer (mo) in seizure-prone GALR1-KO mice(B). No major difference can be seen with regard to dynorphin-ir(E, F). Scale bar: 100 µm (A, B, E, F); 5.5 µm (C, D); 20 µm (G).gr, granule cell layer; mf, mossy fibers; mo, molecular layer; po,polymorph layer.
could predispose neurons for development of seizures, asdiscussed later. Peptidergic plasticity in GALR1-KO miceseemed also to be accompanied, albeit in a very limitedfashion, by terminal sprouting of mossy fibers, which ischaracteristic for kindling and may contribute to seizureformation (43).
Unfortunately, the exact distribution of the galanin re-ceptors in the mouse (and rat) brain is incompletely under-stood, because, in addition to binding studies (44,45), onlythe transcripts have been localized with in situ hybridiza-tion, even in the rat (41). Nevertheless, both binding andin situ hybridization studies have provided evidence for ahigh density of galanin-binding sites and GALR1 mRNAin ventral cortices and ventral hippocampus including en-torhinal cortex, whereas in the dorsal hippocampal forma-tion, no galanin (1–29) binding but only GALR2 mRNAhas been found in granule cells. It is therefore possible thatthe initial triggering of seizures emanates from the entorhi-nal cortex and the perforant pathway(s) to the granule cellsin the dentate gyrus. If this circuitry is “released” becauseof lack of inhibitory GALR1 receptors, seizure activity canspread into hippocampus, resulting in the peptide remodel-ing described here. A further potential source for GALR1is represented by the noradrenergic locus coeruleus neu-rons, which are known to express both galanin (46) andGALR1 (and GALR2) mRNA (41) and which project tocortical and hippocampal regions.
In GALR1-KO mice, we observed a strong upregulationof galanin expression only in neurons in the polymorphlayer. They are presumably mossy cells sending fibersto the supragranular layer, because they, as shown here,also express CCK, but not GABA, and because Fredenset al. (47) have identified the polymorph layer CCK neu-rons as mossy cells. These mossy cells are glutamatergicand should normally have a stimulatory effect on granulecell dendrites in the supragranular layer. Galanin releasedfrom mossy cell terminals may therefore act on granulecells via GALR2 to modulate glutamate release. How-ever, the significance of an increased galanin synthesis inthe mossy cells for development of seizures is not clearand can be ambivalent. Galanin-overexpressing mice aber-rantly produce galanin in the granule cells/mossy fibers,which possibly contribute to increased seizure threshold(17). They do not express detectable galanin in the mossycells/supragranular layer, but this expression appears af-ter hippocampal kindling. Appearance of galanin-positivecells in the polymorph layer were also reported in a ratmodel of SE (15). Moreover, in contrast to inhibitoryGALR1, GALR2 can be an excitatory receptor acting viathe IP3 pathway to elevate calcium concentration (4,5). Apresynaptic excitatory role of GALR2 has been suggestedto modulate nociceptive transmission in the spinal cord(48), and it is possible that in the granule cells, GALR2has a similar presynaptic effect. However, activation ofGALR2 also can decrease intracellular cyclic adenosine
Epilepsia, Vol. 44, No. 8, 2003
1030 S. O. FETISSOV ET AL.
FIG. 7. A: Levels of messenger RNA (mRNA) for galanin (Gal),enkephalin (Enk), neuropeptide Y (NPY), cholecystokinin (CCK),substance P (SP), and dynorphin (Dyn) in seizure-prone GALR1-KO versus wild-type (WT) mice (100%) in granule (gr) or poly-morph cells (poly) of the dentate gyrus. Examples of in situ hy-bridization signals are shown for galanin (B, C), enkephalin (D,E), and substance P (F, G) in WT (B, D, F) and GALR1-KO (C,E, G) mice. Note differences in signal for galanin in the poly-morph layer (po) (B, C), for Enk in the granule cell layer (gr),CA1 and entorhinal cortex (ent) (D, E), and for SP in the poly-morph layer (F, G). Arrows, cells with positive hybridization sig-nals; ec, external capsule; DG, dentate gyrus. Scale bars: 100µm (B, C, F, G); 500 µm (D, E). CA1, Ammon’s horn, fieldCA1.
monophosphate (cAMP) (4,5). Thus if galanin has a gen-erally antiepileptic effect in the hippocampal formation,so far no evidence exists that galanin in the mossy cells ispreventing seizure development, and its true role in thesecells requires further clarification.
Another component of EPP formation in GALR1-KOmice is peptidergic plasticity of the principal dentate neu-rons, the granule cells, such as an appearance of strongNPY expression as reflected by an increased NPY tran-script in the granule cells and peptide in the mossy fibers.A transient increase of NPY expression in the granulecells was found in kindling models (20,49,50), whereasit was long lasting after electrically or kainic acid (KA)-induced SE in rats (51,52) and mice (53). In GALR1-KO mice, NPY also was upregulated in the hilar neu-rons and possibly in their projections in the outer partof the molecular layer, a finding that also was shown inkindled rats (20,50,51). Thus the pattern of NPY expres-sion found in GALR1-KO is rather similar to one thatfollows the kindling than to changes associated with SE,where an increased NPY-ir in the inner part of the molec-ular layer was observed (51). Because both granule cellsand CA3 pyramidal cells express the Y2 receptor, NPYreleased from the mossy fibers can inhibit glutamate re-lease acting both presynaptically (54) and postsynaptically(55). Considering the important antiseizure effect of NPY(53,56), our finding of increased NPY expression in thegranule cell/mossy fibers of GALR1-KOs suggests thatthis NPY plasticity rather represents an adaptive mech-anism against seizures. Regarding the functional signifi-cance of an increased NPY expression in hilar neurons,it is important to discriminate between the increased ex-pression of NPY in the hilus after an acute seizure event,which may contribute to the protective antiseizure mech-anisms and the ectopic expression, such as in rats withspontaneous seizures (57), which probably contribute toepileptic state. NPY hilar neurons may modulate activ-ity of the granule cells via the Y1 receptor, which wasshown to mediate a proepileptic effect, when tested witha specific Y1-receptor antagonist (58). Additionally, anti-NPY antibodies were shown to reduce seizure suscepti-bility to metrazol in KA-treated rats (59). Thus the func-tional significance of an increased NPY expression byhilar interneurons in GALR1-KO mice requires furtherclarification.
An opioid peptide component of the EPP in GALR1-KO mice generally fits what was found in other epilepsymodels (23,60–63), showing changes in opposite direc-tions (that is, a clear increase of enkephalin expressionand a modest decrease of dynorphin in the granule cells).Dynorphin has been shown to cause presynaptic inhibitionof neighboring mossy fibers (64,65) and is considered ananticonvulsant peptide. The interpretation of the changesin enkephalin expression is more complex, because in-hibition of glutamate release from mossy fibers via the
Epilepsia, Vol. 44, No. 8, 2003
HIPPOCAMPAL NEUROPEPTIDES IN GALR1-KO MICE 1031
FIG. 8. ZnT3-immunoreactivity inthe dentate gyrus and CA3 ofwild-type (WT) (A–C), seizure-prone GALR1-KO mice (D–F) andGALR1-KO mice without seizures(nsz) (G–I). Sprouting of mossyfibers (mf) is seen between thepyramidal cells (pyr) in CA3 inseizure-prone GALR1-KO mice (ar-rowheads in E, �). �χαλε βαρσ :20 µm (A, B, D, E, G, H); 10 µm(C, F, I). CA1, CA3, Ammon’s horn,fields CA1, CA3; gr, granule celllayer; mf, mossy fibers; mo, molec-ular layer; po, polymorph layer; sg,supragranular layer; slm, stratum la-cunosum moleculare; sr, stratum ra-diatum; so, stratum oriens.
δ receptor was shown (66); but enkephalin also can in-hibit GABA interneurons (67), and a δ agonist was foundto facilitate SE initiation (21). Thus changes of opioidpeptide expression in granule cells of GALR1-KO micesuggest a rather proconvulsive effect.
In GALR1-KO mice, increased expression of substanceP mRNA in the hilar neurons cells and substance P-ir inthe molecular layer suggest that these changes in sub-stance P interneurons may have effect on activity of gran-ule cells. We have, however, not seen a detectable increasein substance P content in the principal dentate neurons,as has been reported for other epilepsy models (68,69),and this difference may be related to less severe seizuresin our GALR1-KO mice. Considering that substance Penhances glutamate release from hippocampal slices andhas proepileptogenic properties (69,70), an upregulationof substance P in the GALR1-KO mice may contribute toseizure activity.
Both CCK mRNA and peptide levels were decreasedin the granule cells/mossy fibers in GALR1-KO mice.These changes are similar to those seen after hippocampallesion–induced seizures (71) and those found in the mousemodel of megalencephaly, where the mouse displays shak-iness in gait and seizure-like events (72). In kindled rats,another epilepsy model, an increase in CCK immunore-activity fibers in the supragranular layer was found (20);
in GALR1-KO mice, we also observed some tendency forincreased CCK-ir in the supragranular layer as well as inthe polymorph layer, although changes in CCK mRNA inthe hilar neurons were not significant. CCK was shownto block the excitatory effect of KA in mossy fiber CA3transmission (73), and has, in some studies, been shown toact as an anticonvulsive peptide (74,75). However, CCK isgenerally considered a stimulatory messenger, and its ex-citatory effects on granule and CA1 pyramidal cells werereported (76,77). Thus CCK may have both anti- and pro-convulsive effects.
It is difficult to judge the significance of the studied neu-ropeptides for seizure development in GALR1-KO mice,because our results suggest both pro- and antiepileptic ef-fects. Nevertheless, if the EPP can be the key setting inthe hippocampal circuitries to form a spontaneous-seizurephenotype, it is not clear why it appears only in 25% ofGALR1-KO mice. It was suggested by Wasterlain et al.(21) that maladaptive neuropeptide changes in epilepsymodels may represent a liability of the brain to enhanceexcitation and decrease inhibition in response to repetitivefiring of neurons. This hypothesis also could probably beapplied to GALR1-KO mice, among which some animalsmay respond to an enhanced glutamate release by buildingcircuitries that will more readily respond to the excitatoryneurotransmission.
Epilepsia, Vol. 44, No. 8, 2003

1032 S. O. FETISSOV ET AL.
In conclusion, an intact galaninergic system, includingthe galanin peptide itself and the GALR1, appears impor-tant for maintaining a normal level of neuronal excitability.The spontaneous-seizure phenotype observed in 25% ofGALR1-KO mice was found to be accompanied by com-plex changes in neuropeptide expression in some principalhippocampal neurons and interneurons. These changes,which we call EPP, were dramatic for galanin in mossycells (up), for enkephalin and NPY in granule cells/mossyfibers (up), and for CCK in granule cells/mossy fibers(down) and also occur in most other epilepsy models.However, a more complete understanding of the role ofneuropeptides in seizures and seizure models will requiredetailed knowledge of the localization of, in particular, thepeptide receptors and their transduction mechanisms.
Acknowledgment: This study was supported by the SwedishMRC (04X-2887; 03X-10350), Marianne and Marcus Wallen-berg’s Foundation, Knut and Alice Wallenberg’s Foundation, anUnrestricted Bristol-Myers Squibb Neuroscience Grant, and theEuropean Union (QLK3-CT-2000-00237). S.F. was supportedby the Wenner-Gren Foundations. For the generous supply ofantisera, we thank Drs. I. Christensson-Nylander, Uppsala Uni-versity, Uppsala, Sweden (substance P); P. Frey, Novartis Insti-tute of Biomedical Research, Basel, Switzerland (CCK); HelenWong and the late J.H. Walsh, CURE, Digestive Diseases Re-search Center, Antibody/RIA Core (HIH grant DK41301), LosAngeles, CA, U.S.A. (NPY); L. Terenius, Karolinska Institutet,Stockholm, Sweden (enkephalin, substance P); E. Theodorsson,Linkoping University, Linkoping, Sweden (galanin); E. Weber,CoCensys, Irvine, CA, U.S.A. (dynorphin); and R. Palmiter, Uni-versity of Washington, Seattle, WA, U.S.A. (ZnT3).
REFERENCES
1. Tatemoto K, Rokaeus A❛
, Jornvall H, et al. Galanin: a novelbiologically active peptide from porcine intestine. FEBS Lett1983;164:124–8.
2. Melander T, Hokfelt T, Rokaeus A❛
. Distribution of galaninlike im-munoreactivity in the rat central nervous system. J Comp Neurol1986;248:475–517.
3. Perez SE, Wynick D, Steiner RA, et al. Distribution of galaniner-gic immunoreactivity in the brain of the mouse. J Comp Neurol2001;434:158–85.
4. Iismaa TP, Shine J. Galanin and galanin receptors. Results ProblCell Differ 1999;26:257–91.
5. Branchek TA, Smith KE, Gerald C, et al. Galanin receptor subtypes.Trends Pharmacol Sci 2000;21:109–17.
6. Merchenthaler I, Lopez FJ, Negro-Vilar A. Anatomy and phys-iology of central galanin-containing pathways. Prog Neurobiol1993;40:711–69.
7. Hokfelt T, Bartfai T, Crawley J. (eds.) Galanin: basic researchdiscoveries and therapeutic implications. Ann NY Acad Sci 1998;863:1–469.
8. Fisone G, Wu CF, Consolo S, et al. Galanin inhibits acetylcholinerelease in the ventral hippocampus of the rat: histochemical, autora-diographic, in vivo, and in vitro studies. Proc Natl Acad Sci USA1987;84:7339–43.
9. Dutar P, Lamour Y, Nicoll RA. Galanin blocks the slow cholinergicEPSP in CA1 pyramidal neurons from ventral hippocampus. Eur JPharmacol 1989;164:355–60.
10. Zini S, Roisin MP, Langel U, et al. Galanin reduces release of en-dogenous excitatory amino acids in the rat hippocampus. Eur J Phar-macol 1993;245:1–7.
11. Sakurai E, Maeda T, Kaneko S, et al. Galanin inhibits long-term
potentiation at Schaffer collateral-CA1 synapses in guinea-pig hip-pocampal slices. Neurosci Lett 1996;212:21–4.
12. Fuxe K, Jansson A, Diaz-Cabiale Z, et al. Galanin modulates5-hydroxytryptamine functions: focus on galanin and galaninfragment/5-hydroxytryptamine1A receptor interactions in the brain.Ann N Y Acad Sci 1998;863:274–90.
13. Kehr J, Yoshitake T, Wang FH, et al. Galanin is a potent in vivomodulator of mesencephalic serotonergic neurotransmission. Neu-ropsychopharmacology 2002;27:341–56.
14. Mazarati AM, Halaszi E, Telegdy G. Anticonvulsive effectsof galanin administered into the central nervous system uponthe picrotoxin-kindled seizure syndrome in rats. Brain Res1992;589:164–6.
15. Mazarati AM, Liu H, Soomets U, et al. Galanin modulation ofseizures and seizure modulation of hippocampal galanin in animalmodels of status epilepticus. J Neurosci 1998;18:10070–7.
16. Mazarati AM, Hohmann JG, Bacon A, et al. Modulation ofhippocampal excitability and seizures by galanin. J Neurosci2000;20:6276–81.
17. Kokaia M, Holmberg K, Nanobashvili A, et al. Suppressed kindlingepileptogenesis in mice with ectopic overexpression of galanin. ProcNatl Acad Sci U S A 2001;98:14006–11.
18. Jacoby AS, Hort YJ, Constantinescu G, et al. Critical role forGALR1 galanin receptor in galanin regulation of neuroendocrinefunction and seizure activity. Mol Brain Res 2002;107:195–200.
19. Gall C, Lauterborn J, Isackson P, et al. Seizures, neuropeptide reg-ulation, and mRNA expression in the hippocampus. Prog Brain Res1990;83:371–90.
20. Schwarzer C, Sperk G, Samanin R, et al. Neuropeptides-immunoreactivity and their mRNA expression in kindling: func-tional implications for limbic epileptogenesis. Brain Res Rev1996;22:27–50.
21. Wasterlain CG, Mazarati AM, Naylor D, et al. Short-term plasticityof hippocampal neuropeptides and neuronal circuitry in experimen-tal status epilepticus. Epilepsia 2002;43(suppl 5):20–9.
22. White JD, Gall CM. Differential regulation of neuropeptide andproto-oncogene mRNA content in the hippocampus following re-current seizures. Brain Res 1987;427:21–9.
23. Gall C. Seizures induce dramatic and distinctly different changesin enkephalin, dynorphin, and CCK immunoreactivities in mousehippocampal mossy fibers. J Neurosci 1988;8:1852–62.
24. Sperk G, Marksteiner J, Gruber B, et al. Functional changes in neu-ropeptide Y- and somatostatin-containing neurons induced by limbicseizures in the rat. Neuroscience 1992;50:831–46.
25. Vezzani A, Sperk G, Colmers WF. Neuropeptide Y: emerging evi-dence for a functional role in seizure modulation. Trends Neurosci1999;22:25–30.
26. Adams JC. Biotin amplification of biotin and horseradish per-oxidase signals in histochemical stains. J Histochem Cytochem1992;40:1457–63.
27. Theodorsson E, Rugarn O. Radioimmunoassay for rat galanin: im-munochemical and chromatographic characterization of immunore-activity in tissue extracts. Scand J Clin Lab Invest 2000;60:411–8.
28. Christensson-Nylander I, Herrera-Marschitz M, Staines W, et al.Striato-nigral dynorphin and substance P pathways in the rat. I.Biochemical and immunohistochemical studies. Exp Brain Res1986;64:169–92.
29. Schultzberg M, Lundberg JM, Hokfelt T, et al. Enkephalin-like im-munoreactivity in gland cells and nerve terminals of the adrenalmedulla. Neuroscience 1978;3:1169–86.
30. Weber E, Evans CJ, Barchas JD. Predominance of the amino-terminal octapeptide fragment of dynorphin in rat brain regions.Nature 1982;299:77–9.
31. Frey P. Cholecystokinin octapeptide (CCK 26-33), nonsulfated oc-tapeptide and tetrapeptide (CCK 30-33) in the rat brain: analysis byhigh pressure liquid chromatography (HPLC) and radioimmunoas-say (RIA). Neurochem Int 1983;5:811–5.
32. Wenzel HJ, Cole TB, Born DE, et al. Ultrastructural localizationof zinc transporter-3 (ZnT-3) to synaptic vesicle membranes withinmossy fiber boutons in the hippocampus of mouse and monkey. ProcNatl Acad Sci U S A 1997;94:12676–81.
33. Vrontakis ME, Peden LM, Duckworth ML, et al. Isolation andcharacterization of a complementary DNA (galanin) clone from
Epilepsia, Vol. 44, No. 8, 2003

HIPPOCAMPAL NEUROPEPTIDES IN GALR1-KO MICE 1033
estrogen-induced pituitary tumor messenger RNA. J Biol Chem1987;262:16755–8.
34. Howells RD, Kilpatrick DL, Bhatt R, et al. Molecular cloning andsequence determination of rat preproenkephalin cDNA: sensitiveprobe for studying transcriptional changes in rat tissues. Proc NatlAcad Sci U S A 1984;81:7651–5.
35. Larhammar D, Ericsson A, Persson H. Structure and expressionof the rat neuropeptide Y gene. Proc Natl Acad Sci U S A1987;84:2068–72.
36. Deschenes RJ, Lorenz LJ, Haun RS, et al. Cloning and sequenceanalysis of a cDNA encoding rat preprocholecystokinin. Proc NatlAcad Sci U S A 1984;81:726–30.
37. Krause JE, Chirgwin JM, Carter MS, et al. Three rat prepro-tachykinin mRNAs encode the neuropeptides substance P and neu-rokinin A. Proc Natl Acad Sci U S A 1987;84:881–5.
38. Civelli O, Douglass J, Goldstein A, et al. Sequence and expression ofthe rat prodynorphin gene. Proc Natl Acad Sci U S A 1985;82:4291–5.
39. Schalling M, Seroogy K, Hokfelt T, et al. Neuropeptide tyrosine inthe rat adrenal gland: immunohistochemical and in situ hybridizationstudies. Neuroscience 1988;24:337–49.
40. Dagerlind A❛
, Friberg K, Bean AJ, et al. Sensitive mRNA detectionusing unfixed tissue: combined radioactive and non-radioactive insitu hybridization histochemistry. Histochemistry 1992;98:39–49.
41. O’Donnell D, Ahmad S, Wahlestedt C, et al. Expression of the novelgalanin receptor subtype GALR2 in the adult rat CNS: distinct dis-tribution from GALR1. J Comp Neurol 1999;409:469–81.
42. Olney JW, Collins RC, Sloviter RS. Excitotoxic mechanisms ofepileptic brain damage. Adv Neurol 1986;44:857–77.
43. Represa A, Ben-Ari Y. Kindling is associated with the formationof novel mossy fibre synapses in the CA3 region. Exp Brain Res1992;92:69–78.
44. Skofitsch G, Sills MA, Jacobowitz DM. Autoradiographic distribu-tion of 125I-galanin binding sites in the rat central nervous system.Peptides 1986;7:1029–42.
45. Melander T, Kohler C, Nilsson S, et al. Autoradiographic quantita-tion and anatomical mapping of 125I-galanin binding sites in the ratcentral nervous system. J Chem Neuroanat 1988;1:213–33.
46. Holets VR, Hokfelt T, Rokaeus A❛
, et al. Locus coeruleus neurons inthe rat containing neuropeptide Y, tyrosine hydroxylase or galaninand their efferent projections to the spinal cord, cerebral cortex andhypothalamus. Neuroscience 1988;24:893–906.
47. Fredens K, Stengaard-Pedersen K, Wallace MN. Localization ofcholecystokinin in the dentate commissural-associational system ofthe mouse and rat. Brain Res 1987;401:68–78.
48. Liu HX, Brumovsky P, Schmidt R, et al. Receptor subtype-specificpronociceptive and analgesic actions of galanin in the spinal cord:selective actions via GalR1 and GalR2 receptors. Proc Natl AcadSci U S A 2001;98:9960–4.
49. Tonder N, Kragh J, Finsen BR, et al. Kindling induces transientchanges in neuronal expression of somatostatin, neuropeptide Y,and calbindin in adult rat hippocampus and fascia dentata. Epilepsia1994;35:1299–308.
50. Kopp J, Nanobashvili A, Kokaia Z, et al. Differential regulation ofmRNAs for neuropeptide Y and its receptor subtypes in widespreadareas of the rat limbic system during kindling epileptogenesis. MolBrain Res 1999;72:17–29.
51. Vezzani A, Schwarzer C, Lothman EW, et al. Functional changesin somatostatin and neuropeptide Y containing neurons in the rathippocampus in chronic models of limbic seizures. Epilepsy Res1996;26:267–79.
52. Gruber B, Greber S, Rupp E, et al. Differential NPY mRNA expres-sion in granule cells and interneurons of the rat dentate gyrus afterkainic acid injection. Hippocampus 1994;4:474–82.
53. Baraban SC, Hollopeter G, Erickson JC, et al. Knock-out micereveal a critical antiepileptic role for neuropeptide Y. J Neurosci1997;17:8927–36.
54. Colmers WF, Klapstein GJ, Fournier A, et al. Presynaptic inhibitionby neuropeptide Y in rat hippocampal slice in vitro is mediated bya Y2 receptor. Br J Pharmacol 1991;102:41–4.
55. Greber S, Schwarzer C, Sperk G. Neuropeptide Y inhibitspotassium-stimulated glutamate release through Y2 receptors in rathippocampal slices in vitro. Br J Pharmacol 1994;113:737–40.
56. Woldbye DP, Larsen PJ, Mikkelsen JD, et al. Powerful inhibition ofkainic acid seizures by neuropeptide Y via Y5-like receptors. NatMed 1997;3:761–4.
57. Scharfman HE, Sollas AL, Goodman JH. Spontaneous recur-rent seizures after pilocarpine-induced status epilepticus activatecalbindin-immunoreactive hilar cells of the rat dentate gyrus. Neu-roscience 2002;111:71–81.
58. Gariboldi M, Conti M, Cavaleri D, et al. Anticonvulsant propertiesof BIBP3226, a non-peptide selective antagonist at neuropeptide YY1 receptors. Eur J Neurosci 1998;10:757–9.
59. Vezzani A, Civenni G, Rizzi M, et al. Enhanced neuropeptide Yrelease in the hippocampus is associated with chronic seizure sus-ceptibility in kainic acid treated rats. Brain Res 1994;660:138–43.
60. Douglass J, Grimes L, Shook J, et al. Systemic administration ofkainic acid differentially regulates the levels of prodynorphin andproenkephalin mRNA and peptides in the rat hippocampus. MolBrain Res 1991;9:79–86.
61. Harrison MB, Shumate MD, Lothman EW. Opioid peptide expres-sion in models of chronic temporal lobe epilepsy. Neuroscience1995;65:785–95.
62. Hong JS, Wood PL, Gillin JC, et al. Changes of hippocam-pal met-enkephalin content after recurrent motor seizures. Nature1980;285:231–2.
63. Simpson JN, Zhang WQ, Bing G, et al. Kainic acid-induced sprout-ing of dynorphin- and enkephalin-containing mossy fibers in thedentate gyrus of the rat hippocampus. Brain Res 1997;747:318–23.
64. Wagner JJ, Terman GW, Chavkin C. Endogenous dynorphins in-hibit excitatory neurotransmission and block LTP induction in thehippocampus. Nature 1993;363:451–4.
65. Weisskopf MG, Zalutsky RA, Nicoll RA. The opioid peptidedynorphin mediates heterosynaptic depression of hippocampalmossy fibre synapses and modulates long-term potentiation. Nature1993;362:423–7.
66. Simmons ML, Chavkin C. Endogenous opioid regulation of hip-pocampal function. Int Rev Neurobiol 1996;39:145–96.
67. Madison DV, Nicoll RA. Enkephalin hyperpolarizes interneuronesin the rat hippocampus. J Physiol 1988;398:123–30.
68. Borhegyi Z, Leranth C. Substance P innervation of the rat hippocam-pal formation. J Comp Neurol 1997;384:41–58.
69. Liu H, Mazarati AM, Katsumori H, et al. Substance P is expressed inhippocampal principal neurons during status epilepticus and plays acritical role in the maintenance of status epilepticus. Proc Natl AcadSci U S A 1999;96:5286–91.
70. Liu H, Cao Y, Basbaum AI, et al. Resistance to excitotoxin-inducedseizures and neuronal death in mice lacking the preprotachykinin Agene. Proc Natl Acad Sci U S A 1999;96:12096–101.
71. Gall CM, Pico RM, Lauterborn JC. Focal hippocampal lesions in-duce seizures and long-lasting changes in mossy fiber enkephalinand CCK immunoreactivity. Peptides 1988;9(suppl 1):79–84.
72. Petersson S, Lavebratt C, Schalling M, et al. Expression of chole-cystokinin, enkephalin, galanin and neuropeptide Y is markedlychanged in the brain of the megencephaly mouse. Neuroscience2000;100:297–317.
73. Aitken PG, Jaffe DB, Nadler JV. Cholecystokinin blocks some ef-fects of kainic acid in CA3 region of hippocampal slices. Peptides1991;12:127–9.
74. Kadar T, Pesti A, Penke B, et al. Structure-activity and dose-effectrelationships of the antagonism of picrotoxin-induced seizures bycholecystokinin, fragments and analogues of cholecystokinin inmice. Neuropharmacology 1983;22:1223–9.
75. Zetler G. Anticonvulsant effects of caerulein, cholecystokinin oc-tapeptide (CCK-8) and diazepam against seizures produced inmice by harman, thiosemicarbazide and isoniazid. Neurosci Lett1981;24:175–80.
76. Sinton CM. Cholecystokinin and cholecystokinin antagonists en-hance postsynaptic excitability in the dentate gyrus. Peptides1988;9:1049–53.
77. Dodd J, Kelly JS. The actions of cholecystokinin and related peptideson pyramidal neurones of the mammalian hippocampus. Brain Res1981;205:337–50.
Epilepsia, Vol. 44, No. 8, 2003