baff/baff-r involved in antibodies production of rats with collagen-induced arthritis via...
TRANSCRIPT

Ba
PJI
a
ARR3AA
KPSBCP
1
toetitgsuTig
(
0d
Journal of Ethnopharmacology 141 (2012) 290– 300
Contents lists available at SciVerse ScienceDirect
Journal of Ethnopharmacology
jo ur nal homep age : www.elsev ier .com/ locate / je thpharm
AFF/BAFF-R involved in antibodies production of rats with collagen-inducedrthritis via PI3K-Akt-mTOR signaling and the regulation of paeoniflorin
ei-Pei Li1, Dan-Dan Liu1, Yun-Jie Liu, Shan-Shan Song, Qing-Tong Wang, Yan Chang, Yu-Jing Wu,ing-Yu Chen, Wen-Di Zhao, Ling-Ling Zhang ∗, Wei Wei ∗
nstitute of Clinical Pharmacology, Anhui Medical University, Key Laboratory of Anti-inflammatory and Immunopharmacology of Education Ministry of China, 230032 Hefei, China
r t i c l e i n f o
rticle history:eceived 9 July 2011eceived in revised form0 November 2011ccepted 2 February 2012vailable online 27 February 2012
eywords:I3K/Akt/mTORignalingAFF/BAFF-Rollagen induced arthritisaeoniflorin
a b s t r a c t
Ethnopharmacological relevance: Paeoniflorin (Pae) is extracted from the root of paeonia lactiflora whichhave attracted attention for anti-rheumatic and immune modulating properties.Aim of the study: To investigate the role of PI3K/Akt/mTOR signaling mediated by BAFF/BAFF-R in antibod-ies production and the regulation of Pae on the signaling pathway in rats with collagen-induced arthritis(CIA).Materials and methods: CIA rats were randomly separated into different groups and treated with Pae (25,100 mg/kg) from day 18 to day 38 after immunization. The effects of Pae on B lymphocytes of CIA ratswere evaluated by the levels of BAFF, anti-CII antibody, IgA, IgG and IgM, and the expressions of BAFF-R,PI3K, p-Akt and mTOR.Results: In CIA rats, the levels of anti-CII antibody, IgA, IgG and IgM in serum enhanced, BAFF, BAFF-R, PI3K,p-Akt and mTOR were highly expressed. Pae (100 mg/kg) obviously decreased arthritis score, relievedankle and paw swelling, improved spleen histopathology in CIA rats, decreased the levels of IgA, IgM,
IgG and anti-CII antibody, and significantly decreased the expressions of BAFF, BAFF-R, PI3K, p-Akt andmTOR.Conclusion: PI3K/Akt/mTOR signaling mediated by BAFF/BAFF-R participates in antibodies production byB lymphocytes of CIA rats. Pae had therapeutic effects on rats with CIA. These effects might be relative toregulating PI3K/Akt/mTOR signal mediated by BAFF/BAFF-R, and down regulate the antibodies production further.. Introduction
Rheumatoid arthritis (RA) is a systemic autoimmune diseasehat mainly characterized by chronic inflammation targeting syn-vial membrane, following with cartilage and bone erosion (Seylert al., 2005). Although the precise pathogenesis of RA is unclear,he presence of germinal centers, plasma cells, neutrophils, andnflammation in the synovium and at extra articular sites suggesthat both B cell and T cell activation are involved in disease patho-enesis (Yanaba et al., 2008). B cells are not only a precursor cellecreting antibody, but also can they can produce cytokines to reg-late the function of antigen-presenting cells (APCs) and result in
cell activation (Cohen, 2010). It suggests B cells are also involvedn the development of subprime lymphocytes and production oferminal center. Thus, it is reasonable to presume B cells affecting
∗ Corresponding authors. Tel.: +86 551 516 1208; fax: +86 551 516 1208.E-mail addresses: [email protected] (L.-L. Zhang), [email protected]
W. Wei).1 These authors contributed equally to this work.
378-8741/$ – see front matter © 2012 Elsevier Ireland Ltd. All rights reserved.oi:10.1016/j.jep.2012.02.034
© 2012 Elsevier Ireland Ltd. All rights reserved.
the pathogenesis of RA and it is necessary to investigate the rolesof B cells in RA.
B cell-activating factor belonging to the TNF family (BAFF) isa B cell survival and maturation factor. Excessive BAFF produc-tion can destroy B cell self tolerance status, which maybe resultin systemic autoimmune disease. BAFF increase humoral immuneresponse, which induces B cell proliferation, differentiation andimmunoglobulin production. BAFF, as a cytokine, combines its spe-cific receptors, and then causes a series of intracellular response. Inclinical research, the high level of BAFF in serum has been demon-strated in RA patients. This suggests that BAFF play a vital role inthe development of autoimmune disorders.
BAFF has three receptors, including BAFF-R, transmembraneactivator and calcium modulator and cyclophilin ligand interac-tor (TACI), and B cell maturation Ag (BCMA) (Nq et al., 2004; Mayneet al., 2008). Although all these three receptors bind to BAFF, BAFF-Rin vivo appears as a most crucial receptor for the survival of mature B
cells and homeostasis in the peripheral lymphoid organs. BAFF-R isexpressed initially in developing B cells, which exit from the bonemarrow then enter into the mature follicular (FO) and marginalzone (MZ) pools of spleen. BAFF regulates these preimmune B cells
pharm
pwS
its(itpdpmt(aPrm(aCctgti
t(bag(eaTT2s(Tuwar
m
cao Biological Medicine, Shanghai; rabbit anti-BAFF-R antibody(FL-175, sc-28989), mouse anti-PI3-kinase p110� antibody (A-8, sc-55589), rabbit anti-p-Akt1/2/3 antibody (Ser 473, sc-135651) andmouse anti-mTORC1 (10E10, sc-81537) were obtained from Santa
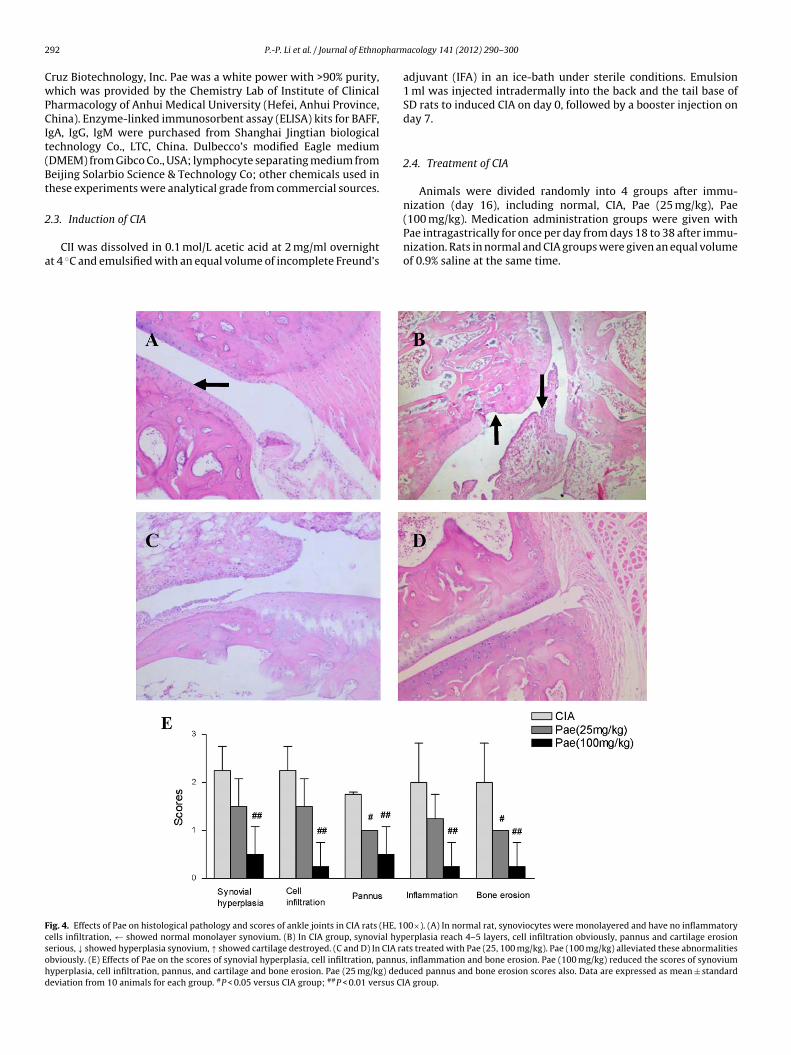
Fig. 2. Effects of Pae on arthritis score of CIA rats. Arthritis score were measured fromday 18 every four days. Arthritis score of CIA rats increased, with a peak score fromday 26 to day 34. Pae (100 mg/kg) reduced the arthritis scores at day 22 (P < 0.05).From day 26 to day 38, the two medication administration groups all decreased
P.-P. Li et al. / Journal of Ethno
ools via survival signals delivered through BAFF-R.FO and MZ cellsill die in the absence of either BAFF-R or BAFF (Zhang et al., 2005;
cholz et al., 2008).BAFF/BAFF-R not only activate anti-apoptotic signal pathway by
nducing the expression of NF-�B and Bcl-2, resulting in the forma-ion of anti-apoptotic protein, but also induce expression of B cellurface CD21 and CD23, and promoting the survival of mature B cellDebnath et al., 2007). Collectively, BAFF/BAFF-R signaling pathways essential for B cell maturation. However, it is unclear that howhe signaling pathway downstream is mediated by BAFF/BAFF-Rarticipating in B cells proliferation, survival and antibody pro-uction in RA. In recent years, some research indicate that thehosphatidylinositol 3-kinase (PI3K) catalytic subunit p110� pro-ote the survival and growth of B cell mediated by BAFF, and
he PI3K/Akt pathway is likely to be important for BAFF responsesHenley et al., 2008). Mammalian target of rapamycin (mTOR) is
serine-threonine kinase and one of downstream molecules ofI3K/Akt. mTOR acts as a sensor kinase that coordinates cellularesponse to growth factors, energy availability and nutrients inammalian cells (Shor et al., 2009). Early and late B cell receptor
BCR) signals are blocked by mTOR inhibitor. The mTOR inhibitorlso suppresses the activation of splenic B cells in mice throughD40, resulting in reduced proliferation and differentiation of Bells (Thomson et al., 2009). However, document retrievals showedhat PI3K/Akt/mTOR pathways were almost discussed in the patho-enesis and treatment method of malignant cancer. It is unknownhat whether BAFF/BAFF-R regulate B cell proliferation and survivaln RA through PI3K/Akt/mTOR pathways.
There are many drugs available in clinic practice for thereatment of RA, including non-steroidal anti-inflammatory drugsNSAIDs), disease modifying anti-rheumatic drugs (DMARDs) andiologic agents. But most of these drugs have disadvantage aboutdverse effects. In the past decade, arthritis patients have seen arowing interest in the use of herbal therapies. Paeoniflorin (Pae)Fig. 1) is one of the components of total glucosides of paeony (TGP)xtracted from the root of paeonia lactiflora. Anti-inflammatory,nalgesia, antihepatic injury and immunoregulatory activities ofGP have been extensively proved in our laboratory for many years.GP was approved for marketing in China in 1998 (Zhou and Li,003; Zheng and Wei, 2005). Pae belongs to a monoterpene gluco-ide and it is extracted and purified from TGP. In adjuvant arthritisAA) rats, Pae has been demonstrated inducing Th1 cells shifted toh2, Th3 cells mediated activities to take effect the immunoreg-latory function (Wu et al., 2007). G protein-coupled signalingas related to the pathogenesis of synovitis in collagen-induced
rthritis (CIA) rats. Pae has anti-inflammatory effects on CIA rats byegulating G protein-coupled signaling (Zhang et al., 2008).
CIA is an established experimental model of polyarthritis withany histopathological features similar to RA. The model is used
Fig. 1. Chemical structure of Pae (formula C23H28O11, molecular weight 480.45).
acology 141 (2012) 290– 300 291
frequently in research RA pathology and new drugs for RA. In thecurrent studies, we investigated whether PI3K/Akt/mTOR path-ways were mediated by BAFF/BAFF-R to participate in antibodiesproduction of B cell. In addition, we evaluated the regulation of Paeon antibodies production and PI3K/Akt/mTOR signaling pathwaysimultaneously on the establishment of CIA model.
2. Materials and methods
2.1. Animals
Sprague–Dawley (SD) rats (male, 150–180 g. Grade II, CertificateNo. 006) were purchased from the Animal Department of AnhuiMedical University (Hefei, Anhui Province, China). All animals werehoused in a room with a controlled ambient temperature (22 ± 2 ◦C)and humidity (50 ± 10%). These animals were acclimated to thehousing conditions for 7 days before experiments, and the EthicsReview Committee approved all experiments for Animal Experi-mentation of the Institute of Clinical Pharmacology, Anhui MedicalUniversity.
2.2. Drugs and reagents
Chicken type II collagen (CII) was provided by Institute of Ben-
arthritis scores significantly. Data are expressed as mean ± standard deviation from10 animals for each group. #P < 0.05 versus CIA group; ##P < 0.01 versus CIA group.
Fig. 3. Effects of Pae on paw swelling of CIA rats. From day 18, paw swelling ofCIA rats were more serious than that of normal rats (P < 0.01). From day 26 to day30, Pae (100 mg/kg) decreased the paw swelling obviously. From day 34 to day 38,the two medication administration groups all decreased the paw swelling. Data areexpressed as mean ± standard deviation from 10 animals for each group. **P < 0.01versus normal group; #P < 0.05 versus CIA group; ##P < 0.01 versus CIA group.
2 pharm
CwPCIt(Bt
2
a
Fcsohd
92 P.-P. Li et al. / Journal of Ethno
ruz Biotechnology, Inc. Pae was a white power with >90% purity,hich was provided by the Chemistry Lab of Institute of Clinical
harmacology of Anhui Medical University (Hefei, Anhui Province,hina). Enzyme-linked immunosorbent assay (ELISA) kits for BAFF,
gA, IgG, IgM were purchased from Shanghai Jingtian biologicalechnology Co., LTC, China. Dulbecco’s modified Eagle mediumDMEM) from Gibco Co., USA; lymphocyte separating medium fromeijing Solarbio Science & Technology Co; other chemicals used inhese experiments were analytical grade from commercial sources.
.3. Induction of CIA
CII was dissolved in 0.1 mol/L acetic acid at 2 mg/ml overnightt 4 ◦C and emulsified with an equal volume of incomplete Freund’s
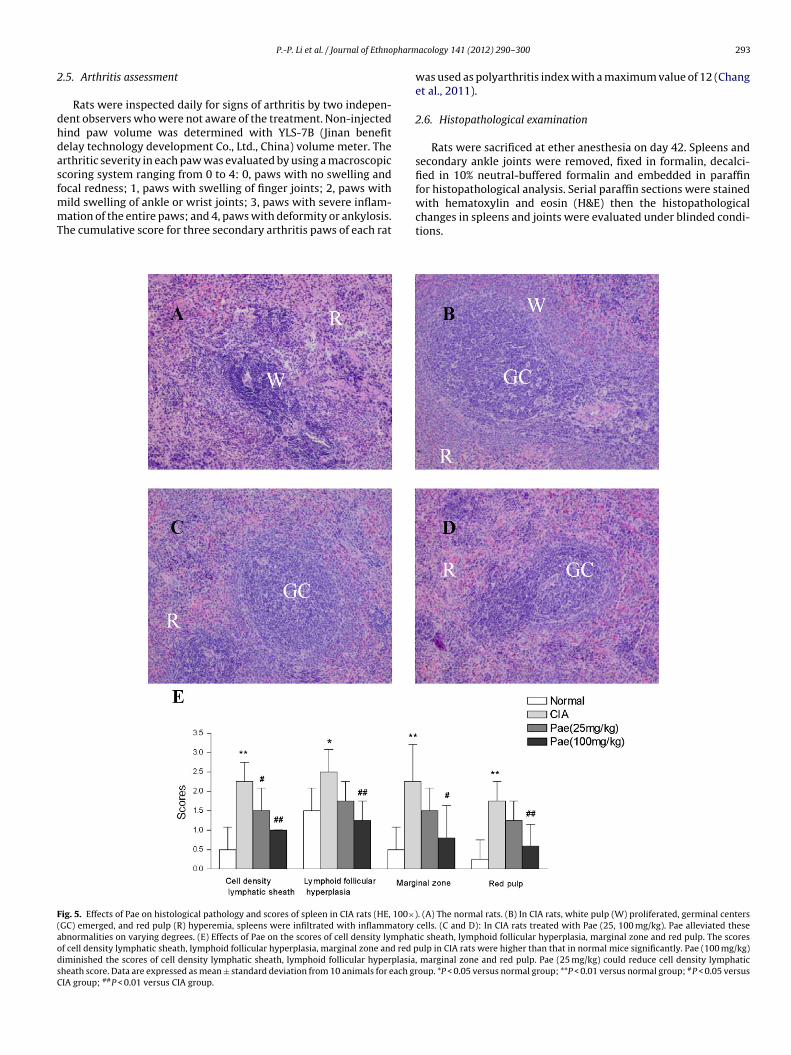
ig. 4. Effects of Pae on histological pathology and scores of ankle joints in CIA rats (HE, 1ells infiltration, ← showed normal monolayer synovium. (B) In CIA group, synovial hyperious, ↓ showed hyperplasia synovium, ↑ showed cartilage destroyed. (C and D) In CIA rabviously. (E) Effects of Pae on the scores of synovial hyperplasia, cell infiltration, pannusyperplasia, cell infiltration, pannus, and cartilage and bone erosion. Pae (25 mg/kg) dedeviation from 10 animals for each group. #P < 0.05 versus CIA group; ##P < 0.01 versus CI
acology 141 (2012) 290– 300
adjuvant (IFA) in an ice-bath under sterile conditions. Emulsion1 ml was injected intradermally into the back and the tail base ofSD rats to induced CIA on day 0, followed by a booster injection onday 7.
2.4. Treatment of CIA
Animals were divided randomly into 4 groups after immu-nization (day 16), including normal, CIA, Pae (25 mg/kg), Pae
(100 mg/kg). Medication administration groups were given withPae intragastrically for once per day from days 18 to 38 after immu-nization. Rats in normal and CIA groups were given an equal volumeof 0.9% saline at the same time.00×). (A) In normal rat, synoviocytes were monolayered and have no inflammatoryerplasia reach 4–5 layers, cell infiltration obviously, pannus and cartilage erosionts treated with Pae (25, 100 mg/kg). Pae (100 mg/kg) alleviated these abnormalities, inflammation and bone erosion. Pae (100 mg/kg) reduced the scores of synoviumuced pannus and bone erosion scores also. Data are expressed as mean ± standardA group.
pharm
2
dhdasfmmT
F(aodsC
P.-P. Li et al. / Journal of Ethno
.5. Arthritis assessment
Rats were inspected daily for signs of arthritis by two indepen-ent observers who were not aware of the treatment. Non-injectedind paw volume was determined with YLS-7B (Jinan benefitelay technology development Co., Ltd., China) volume meter. Therthritic severity in each paw was evaluated by using a macroscopiccoring system ranging from 0 to 4: 0, paws with no swelling and
ocal redness; 1, paws with swelling of finger joints; 2, paws withild swelling of ankle or wrist joints; 3, paws with severe inflam-ation of the entire paws; and 4, paws with deformity or ankylosis.
he cumulative score for three secondary arthritis paws of each rat
ig. 5. Effects of Pae on histological pathology and scores of spleen in CIA rats (HE, 100×)GC) emerged, and red pulp (R) hyperemia, spleens were infiltrated with inflammatory
bnormalities on varying degrees. (E) Effects of Pae on the scores of cell density lymphatf cell density lymphatic sheath, lymphoid follicular hyperplasia, marginal zone and red piminished the scores of cell density lymphatic sheath, lymphoid follicular hyperplasiaheath score. Data are expressed as mean ± standard deviation from 10 animals for each grIA group; ##P < 0.01 versus CIA group.
acology 141 (2012) 290– 300 293
was used as polyarthritis index with a maximum value of 12 (Changet al., 2011).
2.6. Histopathological examination
Rats were sacrificed at ether anesthesia on day 42. Spleens andsecondary ankle joints were removed, fixed in formalin, decalci-fied in 10% neutral-buffered formalin and embedded in paraffin
for histopathological analysis. Serial paraffin sections were stainedwith hematoxylin and eosin (H&E) then the histopathologicalchanges in spleens and joints were evaluated under blinded condi-tions.. (A) The normal rats. (B) In CIA rats, white pulp (W) proliferated, germinal centerscells. (C and D): In CIA rats treated with Pae (25, 100 mg/kg). Pae alleviated theseic sheath, lymphoid follicular hyperplasia, marginal zone and red pulp. The scoresulp in CIA rats were higher than that in normal mice significantly. Pae (100 mg/kg)
, marginal zone and red pulp. Pae (25 mg/kg) could reduce cell density lymphaticoup. *P < 0.05 versus normal group; **P < 0.01 versus normal group; #P < 0.05 versus

2 pharmacology 141 (2012) 290– 300
pp
(
(
(
(
lr(
(
(
(
(
2
owC2Ipit4
Fig. 6. Effects of Pae on the concentration of BAFF in serum. Compared with normalrats, the level of BAFF was increase in CIA rats (P < 0.01). The administration of Pae(100 mg/kg) decreased BAFF levels obviously and Pae (25 mg/kg) had no effect on
94 P.-P. Li et al. / Journal of Ethno
The severity of arthritis in joint was graded from synovial hyper-lasia, cell infiltration, pannus, and bone erosion, as describedreviously (Liu et al., 2011).
1) Synovium hyperplasia: grade 0, hyperplasia was absent; grade1, hyperplasia was mild with two to four layers of reactivesynoviocytes; grade 2, hyperplasia was moderate with fourplus layers of reactive synoviocytes, increased mitotic activity,and mild or absent synovial cell invasion of adjacent bone andconnective tissue; grade 3, hyperplasia was severe and charac-terized by invasion and effacement of joint space and adjacentcartilage, bone, and connective tissue.
2) Cellular infiltration: grade 0, no changes; grade 1, few focal infil-trates; grade 2, extensive focal infiltrates; grade 3, extensiveinfiltrates invading the capsule with aggregate formation.
3) Pannus formation: grade 0, no changes; grade 1, pannus for-mation at up to two sites; grade 2, pannus formation at up tofour sites, with infiltration or flat overgrowth of joint surface;grade 3, pannus formation at more than four sites or extensivepannus formation at two sites.
4) Bone erosion: grade 0, no changes; grade 1, superficial, local-ized cartilage degradation in more than one region; grade 2,localized deep cartilage degradation; grade 3, extensive deepcartilage degradation at several locations.
Histopathological evaluation in spleen was based on cell densityymphatic sheath, lymphoid follicular hyperplasia, marginal zone,ed pulp and the appearance of prominent germinal center (GC)Liu et al., 2011).
1) Cell density lymphatic sheath: grade 0, no changes; grade 1, celldensity was mild increased; grade 2, cell density was moderateaccretion, and congest cells were observed; grade 3, cell densitywas severe accretion, overlying cells were observed.
2) Lymphoid follicular hyperplasia: grade 0, hyperplasia wasabsent; grade 1, hyperplasia was mild; grade 2, hyperplasia wasmoderate with few GCs; grade 3, hyperplasia was severe andcharacterized more GCs.
3) Marginal zone hyperplasia: grade 0, hyperplasia was absentand accompany with few erythrocytes; grade 1, hyperplasiawas mild, marginal zone broaden mildly; grade 2, hyperplasiawas moderate with broad marginal zone; grade 3, hyperplasiawas severe, and the boundary between white pulp and red pulpvague.
4) Red pulp congestion: grade 0, no congestion; grade 1, conges-tion was mild; grade 2, congestion was moderate, inherentcells were reduced in red pulp; grade 3, red pulp were filledwith blood, and inherent cells were reduced in red pulpremarkably.
.7. Measurement of anti-CII antibody in serum
Rats were sacrificed at ether anesthesia on day 42. The serumf rats was collected and diluted into 1:50. And then the serumas seeded into 96-well culture plates, which were bagged withII overnight. These 96-well culture plates were incubated for
h at room temperature and then were seeded goat-anti-mousegG-HRP 100 �l (1:400), incubating at room temperature 1 h. O-
henylenediamine was seeded into 96-well culture plate for 1 hn room temperature. At last, H2SO4 (2 mol/L) 50 �l suspendhe reaction. The concentration was calculated by absorbance at90 nm.
BAFF level. Data are expressed as mean ± standard deviation from 10 animals foreach group. **P < 0.01 versus normal group; ##P < 0.01 versus CIA group.
2.8. Measurement of BAFF, IgA, IgG and IgM
The level of BAFF, IgA, IgG and IgM in serum was measuredusing ELISA kits, respectively. Each sample was assayed in threeduplicates.
2.9. Immunohistochemistry
Immunohistochemical (IHC) analysis was performed by theavidin–biotin peroxidase complex method. In brief, deparaffinized5-�m-thick tissue section was placed in a microwave oven in10 mM citrate buffer (pH 6.0) for 10 min for antigen retrieval. Aftertissue sections were blocked with 0.3% hydrogen peroxidase inphosphate-buffered saline for 10 min at room temperature, eachsection was incubated overnight at 4 ◦C with primary antibodiesfor BAFF-R (1:50 dilution), PI3-kinase p110� (1:50 dilution), or p-Akt1/2/3 (Ser473) (1:25 dilution).
2.10. Western blotting analysis
Rats were sacrificed at ether anesthesia on day 42. Spleenswere removed under sterile conditions and spleenocytes were col-lected. Then cells were suspended in DMEM medium. The cellsuspension was purified with lymphocyte separating medium.Then the cells were washed with phosphate-buffered saline buffertwice and lysed in RIPA lysis buffer (50 mM TRIS (tris (hydrox-ymethyl) aminomethane)–HCl, pH 7.4, 150 mM NaCl, 10 mMphenylmethylsulfonyl fluoride (PMSF), 1 mM ethylene diaminetetraacetic acid (EDTA), 0.1% sodium dodecyl sulfate (SDS), 1% Tri-ton x-100, 1% sodium deoxycholate for 20–30 min on ice. Proteinconcentration was determined with Lowry Protein Assay. The pro-teins were resolved by sodium dodecyl-sulfate-polyacrylamide gelelectrophoresis (SDS-PAGE) and then transferred to polyvinyli-dene fluoride (PVDF) membranes (Millipore, Bedford, MA), andincubated with blocking buffer (Tris-buffered saline/Tween 20(TBST)/5% nonfat drymilk) overnight at 4 ◦C. Immunoblot wasdone with the indicated primary antibody followed by the appro-priate horseradish peroxidase (HRP)-conjugated goat anti-mouse(1:30,000/1:40,000) IgG and immunodetection with enhancedchemiluminescence (ECL, Pierce, Rockford, IL) using hydrogenperoxide and luminol as substrate with Kodak X-AR film. Autora-diographs were scanned using a GS-700 Imaging Densitometer
(Bio-Rad, Hercules, CA). All the experiments reported in this studywere performed three times and the results were reproducible(Zhao et al., 2011).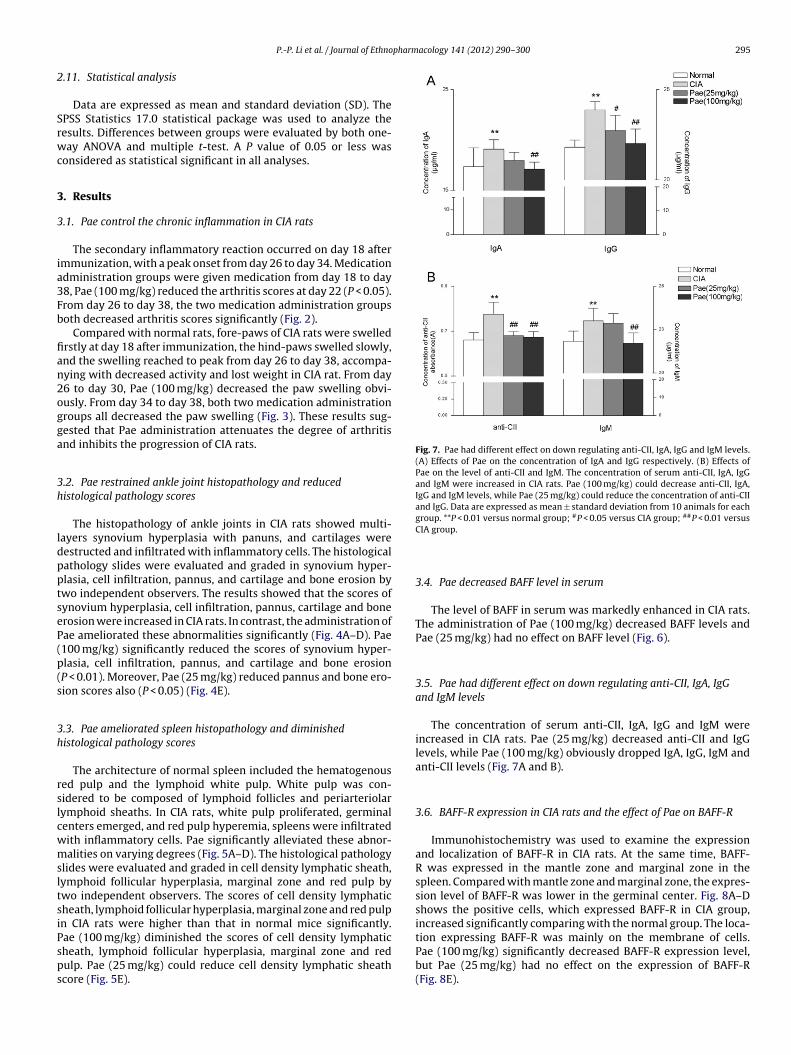
pharmacology 141 (2012) 290– 300 295
2
Srwc
3
3
ia3Fb
fian2ogga
3h
ldpptseP(p(s
3h
rslcwmsltsiPsps
Fig. 7. Pae had different effect on down regulating anti-CII, IgA, IgG and IgM levels.(A) Effects of Pae on the concentration of IgA and IgG respectively. (B) Effects ofPae on the level of anti-CII and IgM. The concentration of serum anti-CII, IgA, IgGand IgM were increased in CIA rats. Pae (100 mg/kg) could decrease anti-CII, IgA,IgG and IgM levels, while Pae (25 mg/kg) could reduce the concentration of anti-CIIand IgG. Data are expressed as mean ± standard deviation from 10 animals for eachgroup. **P < 0.01 versus normal group; #P < 0.05 versus CIA group; ##P < 0.01 versus
P.-P. Li et al. / Journal of Ethno
.11. Statistical analysis
Data are expressed as mean and standard deviation (SD). ThePSS Statistics 17.0 statistical package was used to analyze theesults. Differences between groups were evaluated by both one-ay ANOVA and multiple t-test. A P value of 0.05 or less was
onsidered as statistical significant in all analyses.
. Results
.1. Pae control the chronic inflammation in CIA rats
The secondary inflammatory reaction occurred on day 18 aftermmunization, with a peak onset from day 26 to day 34. Medicationdministration groups were given medication from day 18 to day8, Pae (100 mg/kg) reduced the arthritis scores at day 22 (P < 0.05).rom day 26 to day 38, the two medication administration groupsoth decreased arthritis scores significantly (Fig. 2).
Compared with normal rats, fore-paws of CIA rats were swelledrstly at day 18 after immunization, the hind-paws swelled slowly,nd the swelling reached to peak from day 26 to day 38, accompa-ying with decreased activity and lost weight in CIA rat. From day6 to day 30, Pae (100 mg/kg) decreased the paw swelling obvi-usly. From day 34 to day 38, both two medication administrationroups all decreased the paw swelling (Fig. 3). These results sug-ested that Pae administration attenuates the degree of arthritisnd inhibits the progression of CIA rats.
.2. Pae restrained ankle joint histopathology and reducedistological pathology scores
The histopathology of ankle joints in CIA rats showed multi-ayers synovium hyperplasia with panuns, and cartilages wereestructed and infiltrated with inflammatory cells. The histologicalathology slides were evaluated and graded in synovium hyper-lasia, cell infiltration, pannus, and cartilage and bone erosion bywo independent observers. The results showed that the scores ofynovium hyperplasia, cell infiltration, pannus, cartilage and bonerosion were increased in CIA rats. In contrast, the administration ofae ameliorated these abnormalities significantly (Fig. 4A–D). Pae100 mg/kg) significantly reduced the scores of synovium hyper-lasia, cell infiltration, pannus, and cartilage and bone erosionP < 0.01). Moreover, Pae (25 mg/kg) reduced pannus and bone ero-ion scores also (P < 0.05) (Fig. 4E).
.3. Pae ameliorated spleen histopathology and diminishedistological pathology scores
The architecture of normal spleen included the hematogenoused pulp and the lymphoid white pulp. White pulp was con-idered to be composed of lymphoid follicles and periarteriolarymphoid sheaths. In CIA rats, white pulp proliferated, germinalenters emerged, and red pulp hyperemia, spleens were infiltratedith inflammatory cells. Pae significantly alleviated these abnor-alities on varying degrees (Fig. 5A–D). The histological pathology
lides were evaluated and graded in cell density lymphatic sheath,ymphoid follicular hyperplasia, marginal zone and red pulp bywo independent observers. The scores of cell density lymphaticheath, lymphoid follicular hyperplasia, marginal zone and red pulpn CIA rats were higher than that in normal mice significantly.
ae (100 mg/kg) diminished the scores of cell density lymphaticheath, lymphoid follicular hyperplasia, marginal zone and redulp. Pae (25 mg/kg) could reduce cell density lymphatic sheathcore (Fig. 5E).CIA group.
3.4. Pae decreased BAFF level in serum
The level of BAFF in serum was markedly enhanced in CIA rats.The administration of Pae (100 mg/kg) decreased BAFF levels andPae (25 mg/kg) had no effect on BAFF level (Fig. 6).
3.5. Pae had different effect on down regulating anti-CII, IgA, IgGand IgM levels
The concentration of serum anti-CII, IgA, IgG and IgM wereincreased in CIA rats. Pae (25 mg/kg) decreased anti-CII and IgGlevels, while Pae (100 mg/kg) obviously dropped IgA, IgG, IgM andanti-CII levels (Fig. 7A and B).
3.6. BAFF-R expression in CIA rats and the effect of Pae on BAFF-R
Immunohistochemistry was used to examine the expressionand localization of BAFF-R in CIA rats. At the same time, BAFF-R was expressed in the mantle zone and marginal zone in thespleen. Compared with mantle zone and marginal zone, the expres-sion level of BAFF-R was lower in the germinal center. Fig. 8A–Dshows the positive cells, which expressed BAFF-R in CIA group,increased significantly comparing with the normal group. The loca-tion expressing BAFF-R was mainly on the membrane of cells.Pae (100 mg/kg) significantly decreased BAFF-R expression level,
but Pae (25 mg/kg) had no effect on the expression of BAFF-R(Fig. 8E).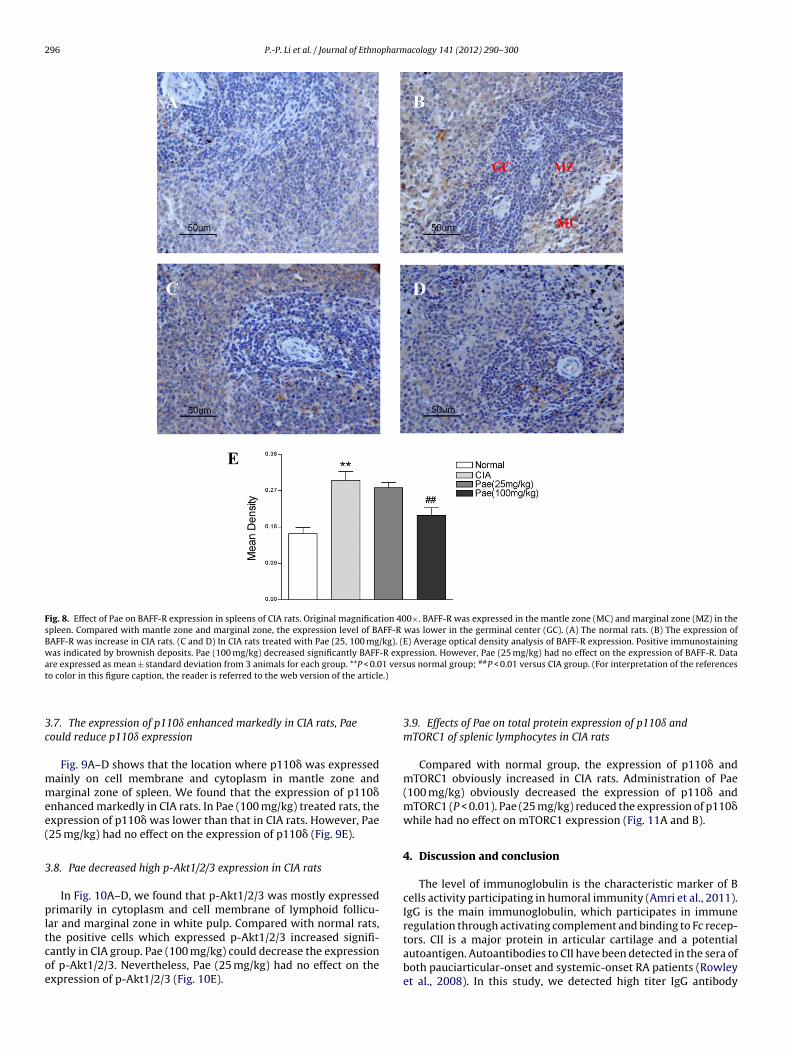
296 P.-P. Li et al. / Journal of Ethnopharmacology 141 (2012) 290– 300
Fig. 8. Effect of Pae on BAFF-R expression in spleens of CIA rats. Original magnification 400×. BAFF-R was expressed in the mantle zone (MC) and marginal zone (MZ) in thespleen. Compared with mantle zone and marginal zone, the expression level of BAFF-R was lower in the germinal center (GC). (A) The normal rats. (B) The expression ofBAFF-R was increase in CIA rats. (C and D) In CIA rats treated with Pae (25, 100 mg/kg). (E) Average optical density analysis of BAFF-R expression. Positive immunostainingwas indicated by brownish deposits. Pae (100 mg/kg) decreased significantly BAFF-R expression. However, Pae (25 mg/kg) had no effect on the expression of BAFF-R. Dataa 1 vers ##
t .)
3c
mmee(
3
pltcoe
re expressed as mean ± standard deviation from 3 animals for each group. **P < 0.0o color in this figure caption, the reader is referred to the web version of the article
.7. The expression of p110ı enhanced markedly in CIA rats, Paeould reduce p110ı expression
Fig. 9A–D shows that the location where p110� was expressedainly on cell membrane and cytoplasm in mantle zone andarginal zone of spleen. We found that the expression of p110�
nhanced markedly in CIA rats. In Pae (100 mg/kg) treated rats, thexpression of p110� was lower than that in CIA rats. However, Pae25 mg/kg) had no effect on the expression of p110� (Fig. 9E).
.8. Pae decreased high p-Akt1/2/3 expression in CIA rats
In Fig. 10A–D, we found that p-Akt1/2/3 was mostly expressedrimarily in cytoplasm and cell membrane of lymphoid follicu-
ar and marginal zone in white pulp. Compared with normal rats,
he positive cells which expressed p-Akt1/2/3 increased signifi-antly in CIA group. Pae (100 mg/kg) could decrease the expressionf p-Akt1/2/3. Nevertheless, Pae (25 mg/kg) had no effect on thexpression of p-Akt1/2/3 (Fig. 10E).us normal group; P < 0.01 versus CIA group. (For interpretation of the references
3.9. Effects of Pae on total protein expression of p110ı andmTORC1 of splenic lymphocytes in CIA rats
Compared with normal group, the expression of p110� andmTORC1 obviously increased in CIA rats. Administration of Pae(100 mg/kg) obviously decreased the expression of p110� andmTORC1 (P < 0.01). Pae (25 mg/kg) reduced the expression of p110�while had no effect on mTORC1 expression (Fig. 11A and B).
4. Discussion and conclusion
The level of immunoglobulin is the characteristic marker of Bcells activity participating in humoral immunity (Amri et al., 2011).IgG is the main immunoglobulin, which participates in immuneregulation through activating complement and binding to Fc recep-
tors. CII is a major protein in articular cartilage and a potentialautoantigen. Autoantibodies to CII have been detected in the sera ofboth pauciarticular-onset and systemic-onset RA patients (Rowleyet al., 2008). In this study, we detected high titer IgG antibody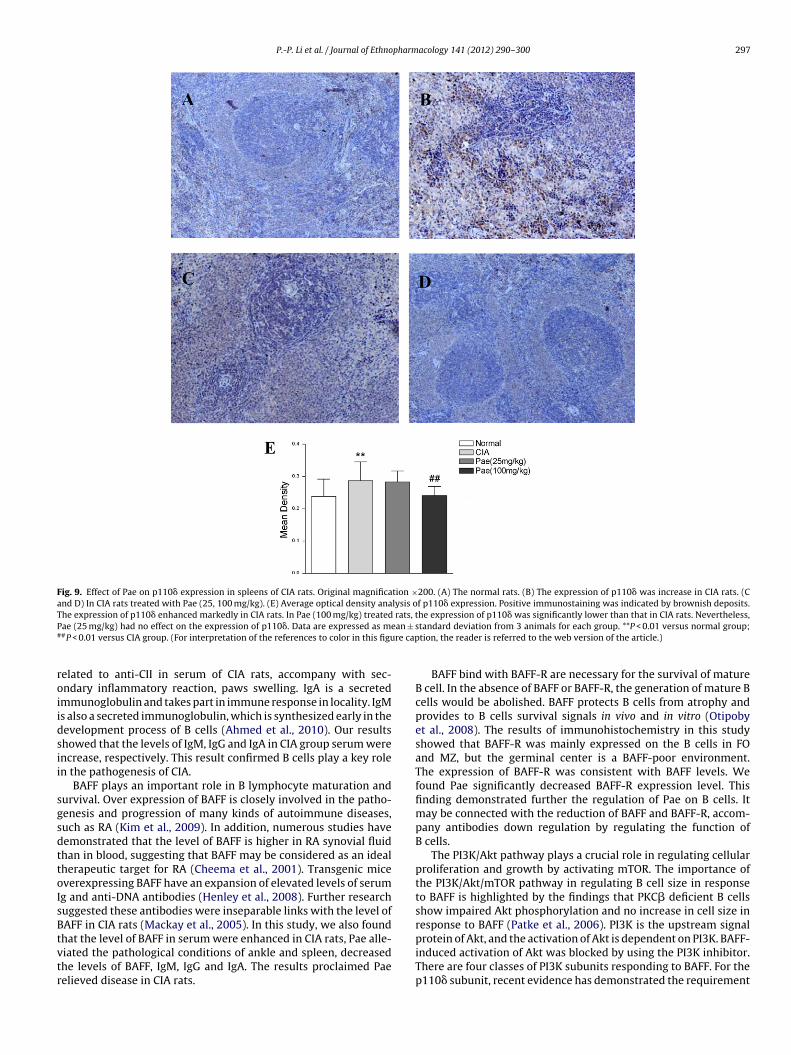
P.-P. Li et al. / Journal of Ethnopharmacology 141 (2012) 290– 300 297
Fig. 9. Effect of Pae on p110� expression in spleens of CIA rats. Original magnification ×200. (A) The normal rats. (B) The expression of p110� was increase in CIA rats. (Cand D) In CIA rats treated with Pae (25, 100 mg/kg). (E) Average optical density analysis of p110� expression. Positive immunostaining was indicated by brownish deposits.T rats, tP ean ±
# re cap
roiidsii
sgsdttoIsBtvtr
he expression of p110� enhanced markedly in CIA rats. In Pae (100 mg/kg) treatedae (25 mg/kg) had no effect on the expression of p110�. Data are expressed as m#P < 0.01 versus CIA group. (For interpretation of the references to color in this figu
elated to anti-CII in serum of CIA rats, accompany with sec-ndary inflammatory reaction, paws swelling. IgA is a secretedmmunoglobulin and takes part in immune response in locality. IgMs also a secreted immunoglobulin, which is synthesized early in theevelopment process of B cells (Ahmed et al., 2010). Our resultshowed that the levels of IgM, IgG and IgA in CIA group serum werencrease, respectively. This result confirmed B cells play a key rolen the pathogenesis of CIA.
BAFF plays an important role in B lymphocyte maturation andurvival. Over expression of BAFF is closely involved in the patho-enesis and progression of many kinds of autoimmune diseases,uch as RA (Kim et al., 2009). In addition, numerous studies haveemonstrated that the level of BAFF is higher in RA synovial fluidhan in blood, suggesting that BAFF may be considered as an idealherapeutic target for RA (Cheema et al., 2001). Transgenic miceverexpressing BAFF have an expansion of elevated levels of serumg and anti-DNA antibodies (Henley et al., 2008). Further researchuggested these antibodies were inseparable links with the level ofAFF in CIA rats (Mackay et al., 2005). In this study, we also found
hat the level of BAFF in serum were enhanced in CIA rats, Pae alle-iated the pathological conditions of ankle and spleen, decreasedhe levels of BAFF, IgM, IgG and IgA. The results proclaimed Paeelieved disease in CIA rats.he expression of p110� was significantly lower than that in CIA rats. Nevertheless,standard deviation from 3 animals for each group. **P < 0.01 versus normal group;tion, the reader is referred to the web version of the article.)
BAFF bind with BAFF-R are necessary for the survival of matureB cell. In the absence of BAFF or BAFF-R, the generation of mature Bcells would be abolished. BAFF protects B cells from atrophy andprovides to B cells survival signals in vivo and in vitro (Otipobyet al., 2008). The results of immunohistochemistry in this studyshowed that BAFF-R was mainly expressed on the B cells in FOand MZ, but the germinal center is a BAFF-poor environment.The expression of BAFF-R was consistent with BAFF levels. Wefound Pae significantly decreased BAFF-R expression level. Thisfinding demonstrated further the regulation of Pae on B cells. Itmay be connected with the reduction of BAFF and BAFF-R, accom-pany antibodies down regulation by regulating the function ofB cells.
The PI3K/Akt pathway plays a crucial role in regulating cellularproliferation and growth by activating mTOR. The importance ofthe PI3K/Akt/mTOR pathway in regulating B cell size in responseto BAFF is highlighted by the findings that PKC� deficient B cellsshow impaired Akt phosphorylation and no increase in cell size inresponse to BAFF (Patke et al., 2006). PI3K is the upstream signal
protein of Akt, and the activation of Akt is dependent on PI3K. BAFF-induced activation of Akt was blocked by using the PI3K inhibitor.There are four classes of PI3K subunits responding to BAFF. For thep110� subunit, recent evidence has demonstrated the requirement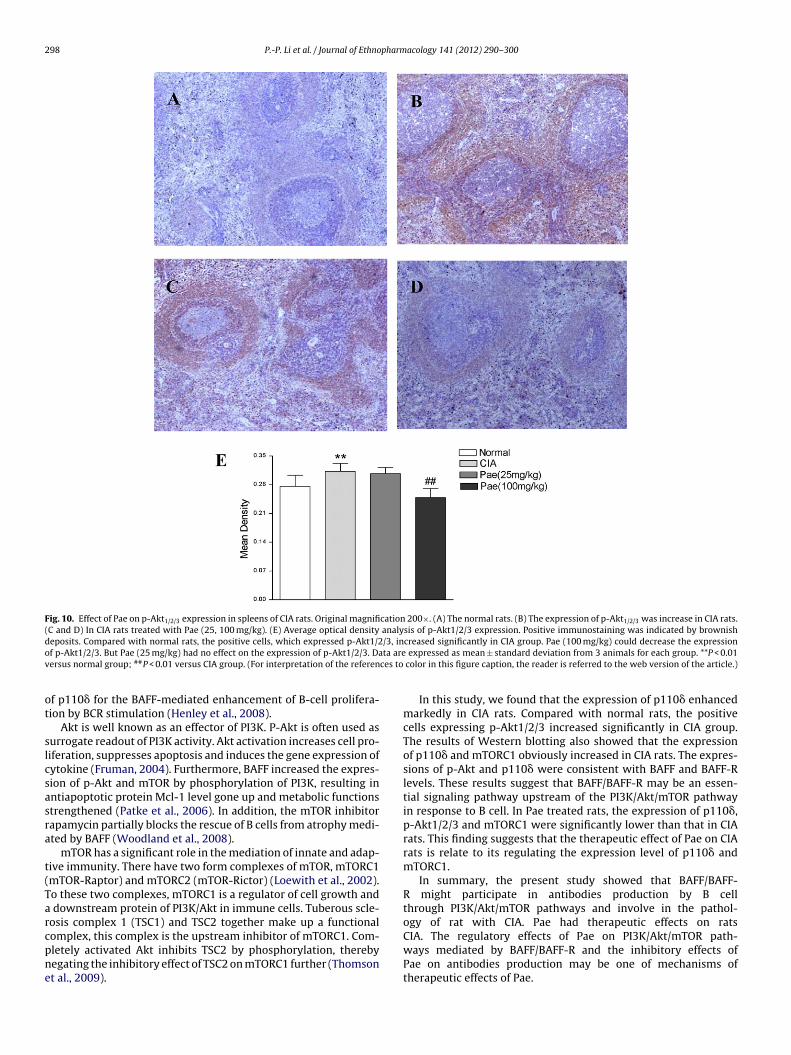
298 P.-P. Li et al. / Journal of Ethnopharmacology 141 (2012) 290– 300
Fig. 10. Effect of Pae on p-Akt1/2/3 expression in spleens of CIA rats. Original magnification 200×. (A) The normal rats. (B) The expression of p-Akt1/2/3 was increase in CIA rats.(C and D) In CIA rats treated with Pae (25, 100 mg/kg). (E) Average optical density analysis of p-Akt1/2/3 expression. Positive immunostaining was indicated by brownishd /3, ino ta arev ces to
ot
slcsasra
t(Tarcpne
eposits. Compared with normal rats, the positive cells, which expressed p-Akt1/2f p-Akt1/2/3. But Pae (25 mg/kg) had no effect on the expression of p-Akt1/2/3. Daersus normal group; ##P < 0.01 versus CIA group. (For interpretation of the referen
f p110� for the BAFF-mediated enhancement of B-cell prolifera-ion by BCR stimulation (Henley et al., 2008).
Akt is well known as an effector of PI3K. P-Akt is often used asurrogate readout of PI3K activity. Akt activation increases cell pro-iferation, suppresses apoptosis and induces the gene expression ofytokine (Fruman, 2004). Furthermore, BAFF increased the expres-ion of p-Akt and mTOR by phosphorylation of PI3K, resulting inntiapoptotic protein Mcl-1 level gone up and metabolic functionstrengthened (Patke et al., 2006). In addition, the mTOR inhibitorapamycin partially blocks the rescue of B cells from atrophy medi-ted by BAFF (Woodland et al., 2008).
mTOR has a significant role in the mediation of innate and adap-ive immunity. There have two form complexes of mTOR, mTORC1mTOR-Raptor) and mTORC2 (mTOR-Rictor) (Loewith et al., 2002).o these two complexes, mTORC1 is a regulator of cell growth and
downstream protein of PI3K/Akt in immune cells. Tuberous scle-osis complex 1 (TSC1) and TSC2 together make up a functional
omplex, this complex is the upstream inhibitor of mTORC1. Com-letely activated Akt inhibits TSC2 by phosphorylation, therebyegating the inhibitory effect of TSC2 on mTORC1 further (Thomsont al., 2009).creased significantly in CIA group. Pae (100 mg/kg) could decrease the expression expressed as mean ± standard deviation from 3 animals for each group. **P < 0.01
color in this figure caption, the reader is referred to the web version of the article.)
In this study, we found that the expression of p110� enhancedmarkedly in CIA rats. Compared with normal rats, the positivecells expressing p-Akt1/2/3 increased significantly in CIA group.The results of Western blotting also showed that the expressionof p110� and mTORC1 obviously increased in CIA rats. The expres-sions of p-Akt and p110� were consistent with BAFF and BAFF-Rlevels. These results suggest that BAFF/BAFF-R may be an essen-tial signaling pathway upstream of the PI3K/Akt/mTOR pathwayin response to B cell. In Pae treated rats, the expression of p110�,p-Akt1/2/3 and mTORC1 were significantly lower than that in CIArats. This finding suggests that the therapeutic effect of Pae on CIArats is relate to its regulating the expression level of p110� andmTORC1.
In summary, the present study showed that BAFF/BAFF-R might participate in antibodies production by B cellthrough PI3K/Akt/mTOR pathways and involve in the pathol-ogy of rat with CIA. Pae had therapeutic effects on rats
CIA. The regulatory effects of Pae on PI3K/Akt/mTOR path-ways mediated by BAFF/BAFF-R and the inhibitory effects ofPae on antibodies production may be one of mechanisms oftherapeutic effects of Pae.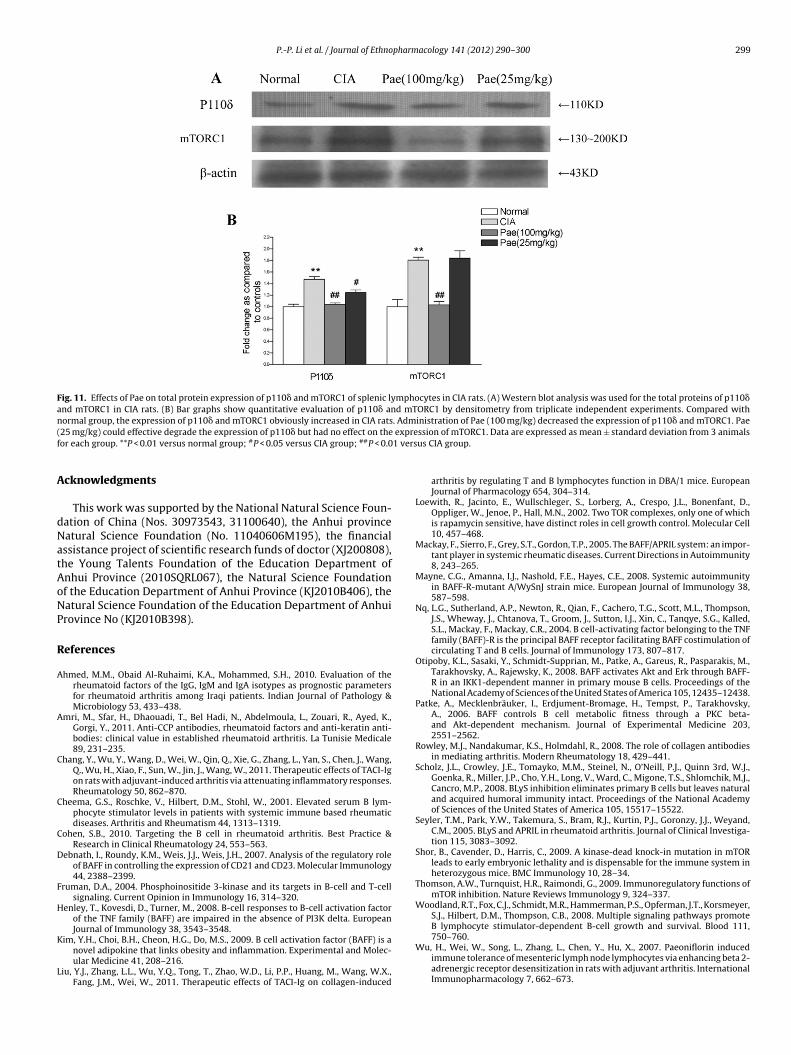
P.-P. Li et al. / Journal of Ethnopharmacology 141 (2012) 290– 300 299
Fig. 11. Effects of Pae on total protein expression of p110� and mTORC1 of splenic lymphocytes in CIA rats. (A) Western blot analysis was used for the total proteins of p110�and mTORC1 in CIA rats. (B) Bar graphs show quantitative evaluation of p110� and mTORC1 by densitometry from triplicate independent experiments. Compared withn . Adm( expref 1 vers
A
dNatAoNP
R
A
A
C
C
C
D
F
H
K
L
ormal group, the expression of p110� and mTORC1 obviously increased in CIA rats25 mg/kg) could effective degrade the expression of p110� but had no effect on theor each group. **P < 0.01 versus normal group; #P < 0.05 versus CIA group; ##P < 0.0
cknowledgments
This work was supported by the National Natural Science Foun-ation of China (Nos. 30973543, 31100640), the Anhui provinceatural Science Foundation (No. 11040606M195), the financialssistance project of scientific research funds of doctor (XJ200808),he Young Talents Foundation of the Education Department ofnhui Province (2010SQRL067), the Natural Science Foundationf the Education Department of Anhui Province (KJ2010B406), theatural Science Foundation of the Education Department of Anhuirovince No (KJ2010B398).
eferences
hmed, M.M., Obaid Al-Ruhaimi, K.A., Mohammed, S.H., 2010. Evaluation of therheumatoid factors of the IgG, IgM and IgA isotypes as prognostic parametersfor rheumatoid arthritis among Iraqi patients. Indian Journal of Pathology &Microbiology 53, 433–438.
mri, M., Sfar, H., Dhaouadi, T., Bel Hadi, N., Abdelmoula, L., Zouari, R., Ayed, K.,Gorgi, Y., 2011. Anti-CCP antibodies, rheumatoid factors and anti-keratin anti-bodies: clinical value in established rheumatoid arthritis. La Tunisie Medicale89, 231–235.
hang, Y., Wu, Y., Wang, D., Wei, W., Qin, Q., Xie, G., Zhang, L., Yan, S., Chen, J., Wang,Q., Wu, H., Xiao, F., Sun, W., Jin, J., Wang, W., 2011. Therapeutic effects of TACI-Igon rats with adjuvant-induced arthritis via attenuating inflammatory responses.Rheumatology 50, 862–870.
heema, G.S., Roschke, V., Hilbert, D.M., Stohl, W., 2001. Elevated serum B lym-phocyte stimulator levels in patients with systemic immune based rheumaticdiseases. Arthritis and Rheumatism 44, 1313–1319.
ohen, S.B., 2010. Targeting the B cell in rheumatoid arthritis. Best Practice &Research in Clinical Rheumatology 24, 553–563.
ebnath, I., Roundy, K.M., Weis, J.J., Weis, J.H., 2007. Analysis of the regulatory roleof BAFF in controlling the expression of CD21 and CD23. Molecular Immunology44, 2388–2399.
ruman, D.A., 2004. Phosphoinositide 3-kinase and its targets in B-cell and T-cellsignaling. Current Opinion in Immunology 16, 314–320.
enley, T., Kovesdi, D., Turner, M., 2008. B-cell responses to B-cell activation factorof the TNF family (BAFF) are impaired in the absence of PI3K delta. EuropeanJournal of Immunology 38, 3543–3548.
im, Y.H., Choi, B.H., Cheon, H.G., Do, M.S., 2009. B cell activation factor (BAFF) is anovel adipokine that links obesity and inflammation. Experimental and Molec-ular Medicine 41, 208–216.
iu, Y.J., Zhang, L.L., Wu, Y.Q., Tong, T., Zhao, W.D., Li, P.P., Huang, M., Wang, W.X.,Fang, J.M., Wei, W., 2011. Therapeutic effects of TACI-Ig on collagen-induced
inistration of Pae (100 mg/kg) decreased the expression of p110� and mTORC1. Paession of mTORC1. Data are expressed as mean ± standard deviation from 3 animalsus CIA group.
arthritis by regulating T and B lymphocytes function in DBA/1 mice. EuropeanJournal of Pharmacology 654, 304–314.
Loewith, R., Jacinto, E., Wullschleger, S., Lorberg, A., Crespo, J.L., Bonenfant, D.,Oppliger, W., Jenoe, P., Hall, M.N., 2002. Two TOR complexes, only one of whichis rapamycin sensitive, have distinct roles in cell growth control. Molecular Cell10, 457–468.
Mackay, F., Sierro, F., Grey, S.T., Gordon, T.P., 2005. The BAFF/APRIL system: an impor-tant player in systemic rheumatic diseases. Current Directions in Autoimmunity8, 243–265.
Mayne, C.G., Amanna, I.J., Nashold, F.E., Hayes, C.E., 2008. Systemic autoimmunityin BAFF-R-mutant A/WySnJ strain mice. European Journal of Immunology 38,587–598.
Nq, L.G., Sutherland, A.P., Newton, R., Qian, F., Cachero, T.G., Scott, M.L., Thompson,J.S., Wheway, J., Chtanova, T., Groom, J., Sutton, I.J., Xin, C., Tanqye, S.G., Kalled,S.L., Mackay, F., Mackay, C.R., 2004. B cell-activating factor belonging to the TNFfamily (BAFF)-R is the principal BAFF receptor facilitating BAFF costimulation ofcirculating T and B cells. Journal of Immunology 173, 807–817.
Otipoby, K.L., Sasaki, Y., Schmidt-Supprian, M., Patke, A., Gareus, R., Pasparakis, M.,Tarakhovsky, A., Rajewsky, K., 2008. BAFF activates Akt and Erk through BAFF-R in an IKK1-dependent manner in primary mouse B cells. Proceedings of theNational Academy of Sciences of the United States of America 105, 12435–12438.
Patke, A., Mecklenbräuker, I., Erdjument-Bromage, H., Tempst, P., Tarakhovsky,A., 2006. BAFF controls B cell metabolic fitness through a PKC beta-and Akt-dependent mechanism. Journal of Experimental Medicine 203,2551–2562.
Rowley, M.J., Nandakumar, K.S., Holmdahl, R., 2008. The role of collagen antibodiesin mediating arthritis. Modern Rheumatology 18, 429–441.
Scholz, J.L., Crowley, J.E., Tomayko, M.M., Steinel, N., O’Neill, P.J., Quinn 3rd, W.J.,Goenka, R., Miller, J.P., Cho, Y.H., Long, V., Ward, C., Migone, T.S., Shlomchik, M.J.,Cancro, M.P., 2008. BLyS inhibition eliminates primary B cells but leaves naturaland acquired humoral immunity intact. Proceedings of the National Academyof Sciences of the United States of America 105, 15517–15522.
Seyler, T.M., Park, Y.W., Takemura, S., Bram, R.J., Kurtin, P.J., Goronzy, J.J., Weyand,C.M., 2005. BLyS and APRIL in rheumatoid arthritis. Journal of Clinical Investiga-tion 115, 3083–3092.
Shor, B., Cavender, D., Harris, C., 2009. A kinase-dead knock-in mutation in mTORleads to early embryonic lethality and is dispensable for the immune system inheterozygous mice. BMC Immunology 10, 28–34.
Thomson, A.W., Turnquist, H.R., Raimondi, G., 2009. Immunoregulatory functions ofmTOR inhibition. Nature Reviews Immunology 9, 324–337.
Woodland, R.T., Fox, C.J., Schmidt, M.R., Hammerman, P.S., Opferman, J.T., Korsmeyer,S.J., Hilbert, D.M., Thompson, C.B., 2008. Multiple signaling pathways promoteB lymphocyte stimulator-dependent B-cell growth and survival. Blood 111,
750–760.Wu, H., Wei, W., Song, L., Zhang, L., Chen, Y., Hu, X., 2007. Paeoniflorin inducedimmune tolerance of mesenteric lymph node lymphocytes via enhancing beta 2-adrenergic receptor desensitization in rats with adjuvant arthritis. InternationalImmunopharmacology 7, 662–673.

3 pharm
Y
Z
Z
00 P.-P. Li et al. / Journal of Ethno
anaba, K., Bouaziz, J.D., Matsushita, T., Magro, C.M., St Clair, E.W., Tedder, T.F., 2008.B-lymphocyte contributions to human autoimmune disease. Immunology 223,284–299.
hang, L.L., Wei, W., Wang, N.P., Wang, Q.T., Chen, J.Y., Chen, Y., Wu, H., Hu, X.Y.,2008. Paeoniflorin suppresses inflammatory mediator production and regulates
G protein-coupled signaling in fibroblast-like synoviocytes of collagen inducedarthritic rats. Inflammation Research 57, 388–395.hang, X., Park, C.S., Yoon, S.O., Li, L., Hsu, Y.M., Ambrose, C., Choi, Y.S., 2005. BAFFsupports human B cell differentiation in the lymphoid follicles through distinctreceptors. International Immunology 17, 779–788.
acology 141 (2012) 290– 300
Zhao, W., Tong, T., Wang, L., Li, P.P., Chang, Y., Zhang, L.L., Wei, W., 2011. Chicken typeII collagen induced immune tolerance of mesenteric lymph node lymphocytesby enhancing beta2-adrenergic receptor desensitization in rats with collagen-induced arthritis. International Immunopharmacology 11, 12–18.
Zheng, Y.Q., Wei, W., 2005. Total glucosides of paeony suppresses adjuvant arthritis
in rats and intervenes cytokine-signaling between different types of synovio-cytes. International Immunopharmacology 5, 1560–1573.Zhou, Q., Li, Z.G., 2003. Pharmaceutical effects of total glucosides of peony and itsapplication in autoimmune disease. Chinese Journal of New Drugs and ClinicalRemedies 22, 687–691.