auto- and heterosynthetic processes in developing ovarian follicles of bacillus rossius (rossi)...
TRANSCRIPT

Int. J. InsectMorphoL & Embryol., Vol. 15, No. 1/2, pp. 49 to64, 1986. 0020-7322/8653.00 + .00 Printed in Great Britain. (~ 1986 Pergamon Press Ltd.
A U T O - A N D H E T E R O S Y N T H E T I C P R O C E S S E S IN D E V E L O P I N G
O V A R I A N F O L L I C L E S O F B A C I L L U S R O S S I U S (ROSSI)
( P H A S M A T O D E A • B A C I L L I D A E )
M. MAZZINI Istituto di Zoologia, Universita di Siena, Siena, Italy.
M . M A S E T T I
Istituto di Zoologia e Anatomia Comparata , Universita di Pisa, Pisa, Italy.
and
F. GIORGI Istituto di lstologia e Embriologia, Universita_ di Pisa, Pisa, Italy.
(Accepted 14 June 1985)
Abs t rac t - -The relative contribution to y01k deposition by heterosynthetic and autosynthetic processes has been examined in developing ovarian follicles of Bacillus rossius (Phasmatodea : Bacillidae). To accomplish this goal, early and late vitellogenic follicles have been examined ultrastructurally. In addition, follicle proteins, labelled both after in vivo and in vitro exposure to radioactive amino acids, have been characterized by gel electrophoresis on polyacrylamide gels. Finally, the intracellular distribution of radioactivity, due to incorporation of (~H)-leucine under both in vivo and in vitro conditions, has been examined in developing ovarian follicles by autoradiography and scintillation counting.
Based on the evidence obtained in the present study, it is concluded that most, if not all, yolk deposited in developing ovarian follicles of B. rossius is obtained by exogenous processes, that is, taken up from the hemolymph by endocytosis. The endogenous contribution is limited to the formation of yolk-like vesicular bodies, whose major function has been interpreted in relation to the intracellular digestion and recycling of aged cell org~tnelles.
Index descriptors (In addition to those in title): Ovarian development, oocyte growth, gel electrophoresis, autoradiography, immunoprecipitat ion assay, stick insect, transmission elect:ton microscopy.
I N T R O D U C T I O N
IN MOST oviparous species, including higher insects (Postlethwait and Giorgi, 1985) and non-mammalian vertebrates (Wallace, 1978), eggs are known to acquire their load of yolk reserve from an extraovarian source. The process, whereby oocytes in these species take up yolk precursors from the external milieu, was defined as heterosynthetic in the early literature (Schechtman, 1955). It is now well documented that heterosynthetic processes in oocytes occur by endocytic sequestration of several proteins from the hemolymph (Telfer et al., 1982), mainly vitellogenins (Wyatt and Pan, 1978), paravitellin and lipophorin (Telfer et aL, 1981) and that the fat body is the tissue capable of releasing them into the hemolymph (Pan et al., 1969; Wyatt, 1980; Wyatt et al., 1984).
*Research ~;ponsored by grants of M.P.I . , 60°70 and 4007o funds. This is the fourth paper in the series "Vitellogenesis in stick insects".
49

50 M. MAZZINI, M. MASETTI and F. GIORGI
The alternative process to form yolk in developing oocytes is by autosynthesis, that is, by the oocyte's capability to synthesize its own yolk storage. From a phyletic standpoint, autosynthetic yolk is believed to be characteristic of oocytes in lower invertebrates where pinocytosis is poorly developed (Huebner and Anderson, 1976; Beams and Kessel, 1962, 1963, 1973). Such processes, however, are not completely disposed of in oocytes of higher oviparous species. Earlier reports (Lanzavecchia, 1968; Kress and Spornitz, 1972) and more recent ones (Ward, 1978; Kessel and Beams, 1980) have, in fact, provided ultrastructural evidence suggesting an oocytes's capability to elaborate its yolk storage even in species where sequestration of hemolymph proteins by endocytosis seems to play a predominant role (Kessel and Ganion, 1980a,b; Blades-Eckelbarger and Youngbluth, 1984).
The present study was undertaken to clarify how auto- and heterosynthetic processes in oocytes of Bacillus rossius contribute to the overall yolk content of fully mature eggs. To provide an answer to this question, we have largely relied on an ultrastructural analysis of developing ovarian follicles as well as on their capability to incorporate radioactive amino acids under in vivo and in vitro conditions.
M A T E R I A L S A N D M E T H O D S
Adult vitellogenic females of Bacillus rossius were maintained under standard laboratory conditions and reared on a diet of fresh bramble leaves. Ovarian follicles from mature females were dissected in a physiological saline solution or, whenever appropriate, directly in the fixative.
Ultrastruclural analysis Ovarian follicles at different stages of development were fixed for 1 hr in 5% glutaraldehyde - 4%
formaldehyde in 0.1 M cacodylate buffer at pH 7.2, maintained at a temperature of 4°C according to Karnovsky's method (1965). Following a prolonged rinse in the buffer, the ovarian follicles were post-fixed for 1 hr in 1% osmium tetroxide in 0.1 M cacodylate buffer at pH 7.2 and subsequently dehydrated in a graded series of alcohol. Eventually, they were embedded in a Epon - Araldite mixture and polymerized for 3 days at 60°C. Silver to pale gold sections were cut on a Reichert ultramicrotome, stained with uranyl acetate and lead citrate, and examined in a Philips EM 300 transmission electron microscope.
Light microscope autoradiography One micrometer-thick sections were cut from Epon Araldite blocks and placed on gelatin-coated slides. The
slides were then coated with a layer of Kodak NTB2 photographic emulsion by the dipping technique and eventually stored in light-proof boxes at 4°C for periods of 2 - 4 weeks. After this exposure time, the slides were developed for 3 min in DI9 developer (Kodak) and rapidly fixed in kodafix. Developed slides were stained with Methylene blue Toluidine blue in borax and photographed in a Zeiss photomicroscope Ill.
Incorporation o f radioactivity For incorporation studies, a few adult females were injected with (3H)-leucine, ("C)-leucine or
(~S)-methionine (Radiochemical Centre, Amersham). After varying times of exposure to the radioisotopes, ovarian follicles were dissected and processed as described below. Incorporation of TCA-precipitable radioactivity was determined by placing aliquots of tissue extracts on paper disks (3MM Whatman), followed by processing in 3-chloro-acetic acid (TCA), alcohol, and ether according to Mans and Novelli (1961). The paper disks treated this way were eventually placed in scintillation vials with 8 ml of toluene containing 0.32% diphenyloxazole (PPO) and counted in a Beckman LS-100C counter.
In vitro culturing A few ovarian follicles were excised from the abdominal body wall and cultured overnight (16 hr) in Grace's
medium (Gibco). One hundred ~l of ("C)-leucine or (~H)-leucine were added to 1 ml of sterile-filtered culture medium. At the end of the culture period, ovarian follicles were washed in a saline solution and soon processed for immunoprecipitation assay or autoradiography.
lmmunoprecipitation assay Antibodies to yolk proteins were raised in rabbits as previously described (Giorgi et al., 1982). Extracts from
ovarian follicles radiolabelled either in vivo or in vitro were made to react with equal volumes of anti-yolk antisera. After centrifugation in a high-speed bench centrifuge, the supernatant and the immune precipitate were counted separately.

Auto- and Heterosynthetic Processes in Developing Ovarian Follicles 51
Electrophoresis and fluorography Sodium dodecyl sulphate (SDS) gel electrophoresis was carried out in a 5-15% polyacrylamide gradient
according to Laemmli (1970) by a vertical slab gel apparatus. Ovarian follicles were dissolved in SDS sample buffer (2% sodium dodecyl sulphate in 0.05 M Tris - HCI buffer at pH 6.8, containing 5% 2-mercaptoethanol), boiled for 3 min and then stored in small aliquots at - 20°C until used. After electrophoresis, gels were stained for 2 hr in 1% Coomassie brilliant Blue G250 in 50°70 methanol- 10°70 acetic acid and destained in a continuous flow of 7% acetic acid. For fluorography, gels were immersed for 30 min in Amplifiy (Radiochemical Centre, Amersham), dried under vacuum at 60°C and mounted with Kodak SO-282 X-ray films and stored for 1 -3 weeks at - 70"C.
R E S U L T S Each panoistic ovariole in B. rossius consists of a series of developing ovarian follicles,
whose sizes increase along the ovariole in an anteroposterior direction. While the last follicle in the ovariole is vitellogenic, the penultimate one is blocked either in previtellogenesis or early vitellogenesis.
In order to evaluate the extent of the relative contribution given by auto- and heterosynthetic processes to oocyte growth in B. rossius, we have examined ultrastructurally ovarian follicles at varying developmental stages. In particular, we have focused our attention on 2 aspects of the oocyte ultrastructure: the oocyte plasma membrane for evidence of endocytic incorporation of yolk precursors from the external milieu (heterosynthesis) and the internal ooplasm for evidence of any contribution provided by the endoplasmic reticulum and/or the Golgi apparatus (autosynthesis). Figure 1 shows the follicle cell/oocyte interface from a follicle occupying the penultimate position in 1:he ovariole. The structural appearance of the cortical ooplasm in this oocyte is typical of an early vitellogenic stage, characterized by a few incipient yolk spheres and several coated pits and vesicles. Both of these are 3-layered organelles with a mean diameter of about 60 nm. They comprise a fuzzy layer onto the luminal side of the vesicle, a unit membrane continuous with the oocyte plasma membrane and an outer spiked layer characterized by regularly spaced clathrin subunits (Fig. 2). Besides these organelles, the cortical ooplasm of the penultimate follicle contains a bewildering variety of small vesicles and several mitochondria.
Figure 3 depicts the central ooplasm of the penultimate follicle in the ovariole. A distinct feature of the ooplasm at this developmental stage is the presence of vesicular bodies of varying electron densities, enclosing such organelles as myelin figures and mitochondria. Besides these organelles, the central ooplasm also contains numerous vesicles, tubular elements and only scant rough endoplasmic reticulum. Some of the vesicular bodies in the central ooplasm may represent initial phases in the degradation of such organelles as mitochondria (Fig. 3). Subsequent development of these bodies is likely to entail further condensation of the mitochondrial matrix (Figs. 4; 5). Later still, the appearance of myelin figures reduces the electron density in these bodies (Figs. 6; 7). Based on these observations, we conclude that the penultimate follicle in the ovariole is structurally characterized by a low rate of endocytic uptake and by the occurrence of an extensive autolysis presumably related to the turn-over of aged cell organelles. In no instances could we witness granules assembled in regions of the cytoplasm close to the endoplasmic reticulum or Golgi apparatus.
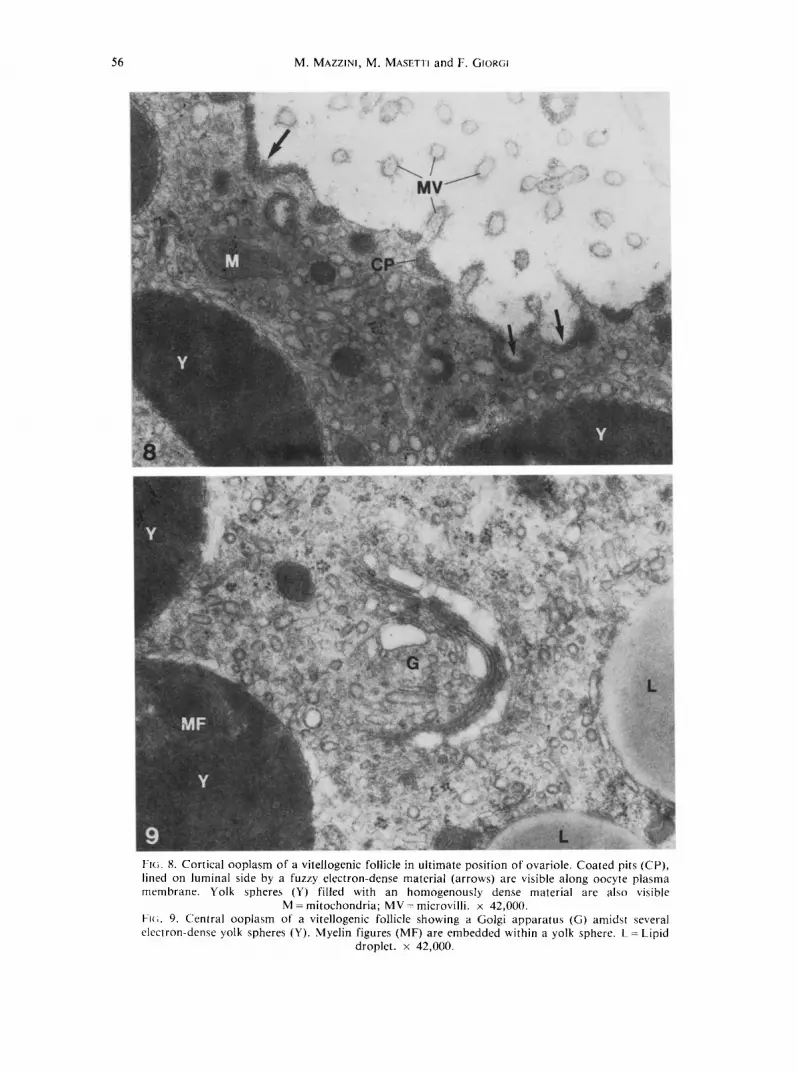
Having established that early vitellogenic follicles are practically incapable of performing endogenous activities, we subsequently examined how terminal oocytes in the ovariole undergo rapid vitellogenic growth. Figures 8 and 9 depict the cortical and central ooplasm, respectively, of the last ovarian follicle in the ovariole. The presence of large coated pits and vesicles with a mean diameter of 180 nm witnesses an extensive endocytic

52 M. MAZZINI, M. MASETTI and F. GIORGI
Fl(~. 1. Low power view of foll icle ce l l /oocy te ( F C / O C ) in ter face f rom an ear ly vi te l logenic follicle in penu l t ima te pos i t ion , in ovariole . L = l ipid drople t ; M - mi tochondr i a ; Y = yolk. x 14,000.
FJ(;. 2. High power view of a coated pit (CP) fo rmed a long oocyte p l a sma m e m b r a n e of an early vi tel logenic follicle. FC = follicle cell; MV - microvi l l i ; OC = oocyte, x 38,000.

Auto- and Heterosynthetic Processes in Developing Ovarian Follicles 53
activity at these developmental stages. Forming yolk spheres have become so large as to have reduced the cortical ooplasm to a narrow fringe underneath the oolemma (Fig. 8). Quite likely, once divested of their lattice coat, coated vesicles deliver their load of membrane-bounded proteins to these large spheres without undergoing intermediate fusion with small vesicles. Other apparently empty vesicles are present both in the cortical ooplasm as well as in the central one. They are likely to be part of a system of smooth endoplasmic reticulum functionally related either to membrane recycling or synthesis of non-protein materials.
The central ooplasm of these vitellogenic oocytes is filled with numerous lipid droplets and large yolk spheres, some of which contain myelin figures within a homogeneously electron-dense matrix (Fig. 9). The Golgi apparatus often lies close to the yolk spheres as if it were transferring vesicles to or f rom them (Fig. 9).
As in the case of the penultimate follicle, the observations carried out in this section suggest that the oocyte is actually incapable to carry out any endogenous activity, and at the same time stress the importance of endocytosis as the only mechanism for yolk formation.
Another approach to define the relative extent of autosynthetic and heterosynthetic processes in oocytes is to determine their protein content as a function of development and to corrtpare it with that of the hemolymph. A common protein content between the ovarian follicle and the hemolymph would in fact be highly suggestive of a heterosynthetic origin of yolk.
Figure 10 shows several ovarian follicles analyzed at varying developmental stages by eletrophoresis in a 5 - 1 5 % polyacrylamide gel gradient under denaturing conditions. There appear to be at least 5 major protein fractions that accumulate with development and are common both to ovarian follicles and hemolymph. In order to establish whether these protein fractions are primarily contributed by the oocyte or by the associated follicle cells, a few ovarian follicles were dissected and each tissue was analyzed separately by gel electrophoresis. Figure 11 shows that all major protein fractions contributed by the hemolymph are also present in the oocyte, whereas fractions in other regions of the gel are primarily due to follicle cells. The fact that some of the fractions present in the oocyte are also shared by a follicle cell preparat ion is most likely due to contamination arising during manual dissection. Owing, in fact, to microvilli interlocking at the follicle cell/oocyte interface, most of the cortical ooplasm remains stuck to the follicle cell epithelium.
Even though gel electrophoresis gave us a good indication in favour of the heterosynthetic nature of most of the protein fractions present in vitellogenic oocytes, it does not necessarily prove it. This can only be attained by exposing follicles to radioactive amino acids. Since under in vivo conditions, both auto- and heterosynthetic proteins would be expected to be labelled, it becomes imperative to compare it with that carried out in vitro. Both of these experiments are reported in Figs. 12 and 13 where labelling of ovarian follicles for 4 and 8 hr is compared with that of hemolymph proteins over the same time exposure (4 hr). It can be clearly seen that those proteins that are common to both hemolymph and oocytes are indeed labelled only after in vivo exposure, and that those that are labelled following in vitro culturing are exclusive of the follicle cells. The extent of labelling attained by individual polypeptides for any given time of exposure to the radioisotope is likely to depend upon their relative methionine content and /o r their differentia] rate of uptake into the oocyte.

54 M, MAZZINI, M. MA~;E111 and F. GIOR~I
Ft~;. 3. Central ooplasm of an early vitellogenic follicle. Note variety of vesicular bodies (VB) containing electron-dense material at varying degrees of condensation. Myelin figures (MF) are also
visible in some of them. M mitochondria. × 40,000.

Auto- and Heterosynthetic Processes in Developing Ovarian Follicles 55
Fl(;s. 4--7. Central ooplasm of an early vitellogenic follicle showing an alleged sequence of a progressive transformation of electron-dense vesicular bodies (VB) into myelin figures (MF). M =
mitochondria, x 40,000.
56 M. MAZZINI, M. MASETTI and F. GIORGI
Fl(;. 8. Cortical ooplasm of a vitellogenic follicle in ultimate position of ovariole. Coated pits (CP), lined on luminal side by a fuzzy electron-dense material (arrows) are visible along oocyte plasma membrane. Yolk spheres (Y) filled with an homogenously dense material are also visible
M - mitochondria; MV = microvilli, x 42,000. Fie;. 9. Central ooplasm of a vitellogenic follicle showing a Golgi apparatus (G) amidst several electron-dense yolk spheres (Y). Myelin figures (MF) are embedded within a yolk sphere. L = Lipid
droplet. × 42,000.

Auto- and Heterosynthetic Processes in Developing Ovarian Follicles 57
! OF (rn m) He
I I I
10 FIG. 10. Gel electrophoresis of hemolymph (He) and several ovarian follicles (OF) of succeeding developmental stages analyzed under denaturing conditions in a 5- 15% polyacrylamide gradient. FractiorL bands arrowed are common to both ovarian follicles and hemolymph. Follicles are staged
according to length of major axis.
As a furl;her criterion to define heterosynthetic vs autosynthetic processes in developing oocytes of Bacillus, we have relied on the compartmentat ion attained by label after exposure to a radioactive amino acid. I f indeed most, if not all, yolk proteins in Bacillus are heterosynthetic by nature, that is due to endocytic incorporation f rom the hemolymph, then, yolk spheres should be labelled only in vivo, but not in vitro, except perhaps fc,r proteins contributed by the follicle cells and transferred to the oocyte via endocytic uptake. Figure 14 shows that oocytes exposed in vivo to (3H)-leucine for 12 hr are almost exclusively labelled over the yolk spheres irrespective of the developmental stage achieved. When similarly staged ovarian follicles are labelled under in vitro conditions, autoradiographic grains appear preferentially over the follicle cells and not over the yolk spheres (Fig. 15).
Finally, to provide a quantitative account of the relative contribution by yolk proteins, ovarian follicles were labelled with (~4C)-leucine both in vivo and in vitro culturing and subsequendy immunoprecipitated with an anti-yolk anti-serum. Table 1 shows that under these experimental conditions, follicles are progressively more labelled as they proceed through development. This is likely to depend upon the relative protein content of each ovarian follicle that is also known to increase with development (unpublished data). On the other hand, a real estimate of the contribution provided by heterosynthetic processes under these conditions is expressed by the ratio of the immunoprecipitate counts over those in the supernatant. Figure 16 shows that such a ratio becomes significantly higher with development only in follicles radiolabelled in vivo. This figure demonstrates that yolk proteins accumulate into the oocyte only in follicles cultured in vivo suggesting that yolk can originate only f rom the hemolymph and hence be exogenous.

58 M. MAZZINI, M. MASETTI and F. GIORGI
F C O C O F i . . . . . i ~ I I
m 11
Flo. 11. Gel electrophoresis of a dissected ovarian follicle (OF) analyzed as in previous figure and showing protein content of both follicle cells (FC) and oocyte (OC). Arrows point fraction bands
common to hemolymph.
O F (8 hr) H e I I t I
12
O F (4 hr ) I
w
FJci. 12. Extent of (~%)-methionine incorporated into ovarian protein fractions as detected by fluorography. Follicles at varying developmental stages were exposed to (~%)-methionine in vivo for 4 and 8 hr. Note how protein fractions labelled under these conditions have an electrophoretic
counterpart in hemolymph (arrows).

Auto- and Heterosynthetic Processes in Developing Ovarian Follicles 59
OF (8hr) He OF(4hr) J I I I I I
~ I .S | . S rnm
13
r
FIG. 13. Extent of (35S)-methionine incorporated into ovarian protein fractions, as detected by fluorography. Follicles at varying developmental stages were exposed to (l~S)_methionine in vitro for 4 and 8 hr. No major protein fraction labelled under these experimental conditions has a counterpart in
hemolymph (arrows).
D I S C U S S I O N
There are several animal species that synthesize endogenously all their yolk reserves (Gremigni, 1983; Mazzini et al., 1984). From a structural point of view, developing oocytes in these species are characterized by a substantial development of the endoplasmic reticulum and by an equally extensive Golgi apparatus (Kessel, 1966; Ganion and Kessel, 1972). Since this is typical of lower invertebrates, including spiralians (Huebner and Anderson, 1976) and apterygote insects (Bilinski, 1977, 1979; Bitsch and Bitsch, 1982), it is interpreted as a phylogenetically primitive condition (Boyer, 1972). More evolved oviparous species have developed mechanisms whereby the oocyte is practically dispensed from synthesizing its own yolk (Wallace, 1978; Giorgi, 1980; Giorgi and Mazzini, 1984). Under these conditions a tissue other than the ovary, namely the fat body in insects (Engelmann, 1979) and the liver in non-mammalian vertebrates (Clemens, 1974), can synthesize and secrete yolk precursors into the blood stream. In addition, the oocyte has developed the capacity to endocytose them by receptor-mediated endocytosis (Roth and Woods, 1982).
Our ultrastructural analysis of ovarian follicles in Bacillus has shown that very little differentiation occurs in the endoplasmic reticulum during vitellogenesis and, at the same time, stresses the role of endocytosis as the major, if not the only, source of yolk precursors in the oocyte. Yet, in spite of this, we have repeatedly observed vesicular bodies that appeared filled with electron-dense material morphologically similar to yolk. Based on similar observations, a number of earlier investigators have suggested that oocytes that are already capable of undergoing endocytosis may also elaborate autosynthetically part of their yolk (Droller and Roth, 1966; Beams and Kessel, 1973; Kessel and Beams, 1980). However, this conclusion is valid only if the morphological evidence is corroborated by a quantitative analysis on yolk formation. In addition, the

60 M. MAZZINI, M. MASETTI and F. GIORGI
FIo. 14. Light microscope autoradiograph of an early vitellogenic follicle exposed in vivo for 12 hr to ('H)-leucine. Note how autoradiographic grains are preferentially located over yolk spheres.
FC - follicle cells; OC - oocyte; Y - yolk spheres. × 600. Fro. 15. Light microscope autoradiograph of an early vitellogenic follicle exposed in vitro to (~H)-leucine up to 12 hr. Note how autoradiographic grains are preferentially located over follicle cell
epithelium (FC). OC oocyte; Y = yolk spheres. × 600.

Auto- and Heterosynthetic Processes in Developing Ovarian Follicles 61
chemical nature of the material accumulated in vesicular bodies during early vitellogenesis needs also to be chemically ascertained. As far as we are aware, no such information is as yet available, except for Xenopus in which the extent of yolk contributed by endogenous processes, if any, appears to be negligible (Wallace et al., 1972).
In the present study, the relative contribution by auto- and heterosynthetic processes to yolk deposition in B. rossius has been evaluated by examining how follicles become labelled after exposure to radioactive amino acids under both in vivo and in vitro conditions. Our data show that vitellogenic follicles exposed in vivo to (~S)-methionine are labelled in those protein fractions that have an electrophoretic counterpart in the female hemolymph. By contrast, vitellogenic follicles exposed to (35S)-methionine after in vitro culturing are labelled only in those proteins that are unique to the follicle itself. Since our dissecting analysis of developing ovarian follicles proved the follicle cells to be the major source of these proteins, the pattern of labelling obtained in vitro is in all likelihood attributable to the synthetic processes of the follicle cells and not to that of the oocyte. The autoradiographic analysis of vitellogenic follicles has, in addition, shown that yolk spheres become labelled only when oocytes are exposed to (3H)-leucine under in vivo conditions. The possibility that follicle cells in B. rossius could synthesize proteins that are subsequently contributing to the overall yolk content cannot be completely ruled out on the basis of the present evidence. However, analysis by scintillation counting of in vitro cultured fo]licles has shown that endogenously contributed proteins may at most amount to 2-3°7o of the total protein content. In view of these findings, our ultrastructural observations could be safely interpreted as indicating a predominant, if not exclusive, role of beterosynthetic yolk formation over the autosynthetic one.
The occurrence of electron-dense vesicular bodies in early vitellogenic follicles is probably due to a focal degradation of aged cell organelles rather than to endogenous synthesis. One may in fact assume that, since oocytes are long-lasting cells relative to the life span of their organelles, there must be a device to get rid of, and possibly recycle, aged organelles. It is with these considerations in mind that we interpret the images depicted in Figs. 4 - 7 where mitochondria are assumed to condense into an electron-dense material and subsequenly undergo intracellular digestion.
TABLE I. IMMUNO ASSAY OF FOLLICLE HOMOGENATE PRECIPITATED WITH ANTI YOlK SERUM AFTER in vil20 OR in vitro EXPOSURE TO (J4C)-LEUCINE
Folli~.les* In vivo~f In vitro{ (mm) T S IMP/y T S IMP/y
0.76 117 26 23 203 112 188 1.16 407 55 1305 371 202 215 1.26 596 75 2247 574 300 520 1.50 980 85 3730 584 285 485 1.33 2047 423 5432 2674 1254 892 2.10 610 181 3235 3632 2047 2406 2.50 506 156 1510 3372 1862 2395
* Follicle length (mm). ~ Follicles exposed in vivo to (~'C)- leucine for 15 hr. { Follicles exposed in vitro to ('4C)- leucine for 15 hr. (T) TCA-precipitable counts per follicle of a given size. (S) TCA-precipitable counts of supernatant after immunoprecipitation with anti-yolk antiserum
per follicle of a given size. (IMP/y) TCA insoluble counts of immunoprecipitate with anti-yolk antiserum per follicle of a
given size,

62 M. MAZZINI, M. MASETTI and F. GIORGI
One question raised by this interpretation is whether these previtellogenic yolk-like vesicular bodies contribute to the yolk spheres of later stages. This would be consistent with the observations by us (see Fig. 9) and other (Kress and Spornitz, 1972) that yolk-like vesicular bodies seem to fuse with endocytotically derived vesicles to yield fully formed yolk spheres. These observations could be accounted for if exogenous and endogenous compartments were assumed to fuse. Under such circumstances, proteins gathered into the oocytes by endocytosis would be conveyed into the same cell compartment where endogenously derived material is being recycled during pre- or early vitellogenesis (Busson-Mabillot, 1984).
IMPy/S
50.
40-
30"
20.
10,
e - - e in vivo o - - o in vitro
eO~o~O--o~o__ o--O
1.0 1.5 2.0 2.5 m m
ovarian follicles
FiG. 16. Diagram showing ratio of radiolabelled immunoprecipitate over supernatant (IMPy/S) as plotted against size (ram) of ovarian follicles.
R E F E R E N C E S
BEAMS, H. W. and R. KESSEL. 1962. lntracisternal granules of the endoplasmic reticulum in the crayfish oocyte. J. Cell BioL 13: 158-62 .
BEAMS, H. W. and R. KESSEL. 1963. Electron microscopic studies on developing crayfish oocytes with special reference to the origin of yolk J. Cell Biol. 18:621 44.
BEAMS, H. W. and R. KESSEL. 1973. Oocyte structure and early vitellogenesis in the trout, Salmo gairdneri. Amer. J. Anat. 136:105 22.
BnANSKL S. 1977. Oogenesis in Acerentomon gallicum Jonescu (Protura). Previtellogenic and vitellogenic stages. Cell Tissue Res. 179:401 - 12.
BnJrqSK1, S. 1979. Oogenesis in Campodea sp. (Diplura). The ultrastructure of the egg chamber during vitellogenesis. Cell Tissue Res. 202: 133-43 .
B;'rscn, J. and C. BITSCH. 1982. Les etapes de la vitellogen~se chez Thermobia domestica (Packard) (Thysannra : Lepismatidae). Int. J. Insect MorphoL EmbryoL 11: 197 - 212.
BI_ADES-ECKELBARGER, P. 1. and M. J. YOUNGBLUTH. 1984. The ultrastructure of oogenesis and yolk formation in Labidocera aestiva (Copepoda : Calanoida). J. Morphol. 1"/9:33 - 46.
BORER, B. C. 1972. Ultrastructural studies of differentiation in the oocyte of the polyclad turbellarian, Prostheceraeus floridanus. J. Morphol. 1 3 6 : 2 7 3 - 9 6 .
BUSSON-MABn l.OT, S. 1984. Endosomes transfer yolk proteins to lysosomes in the vitellogenic oocyte of the trout? Biol. Cell 51:53 - 66.
C] t~MENS, M. J. 1974. The regulation of egg yolk protein synthesis by steroid hormones. Prog. Biophys. MoL Biol. 28:71 - 107.
DROLCER, M. J. and T. F. ROTH. 1966. An electron microscope study of yolk formation during oogenesis in Lebistes reticulatus guppyi. J. Cell Biol. 28:209 32.

Auto- and Heterosynthetic Processes in Developing Ovarian Follicles 63
ENGELMANN, IF'. 1979. Insect vitellogenin: identification, biosynthesis and role in vitellogenesis. Adv. Insect PhysioL 14: 4 9 - 109.
GANION, L. R. and R. G. KESSEL. 1972. Intracellular synthesis, transport and packaging of proteinaceous yolk in oocytes of Orconectes immunis. J. Cell Biol. 52:420 37.
GIORGI, F. 1980. Coated vesicles in the oocyte, pp. 135 - 77. In C. D. OCKLEFORD and A. WHYTE (eds.) Coated Vesicles, Cambridge University Press, Cambridge.
GIORGI, F. and M. MAZZ1NI. 1984. Vitellogenesis in the stick insect Bacillus rossius (Rossi) (Insecta Phasmatodea Bacillidae). 2. Ultrastructural observations on developing oocytes. Monit. Zool. ltal. (N.S.) 18: 259- 73.
GIORGI, F., G. BALDIN1, A. L. SIMONINI and M. MENGHERI. 1982. Vitellogenesis in the stick insect Carausius morosus. II: Purification and biochemical characterization of two vitellins from eggs. Insect Biochem. 12: 5 5 3 - 6 2 .
GREMIGNI, V. 1983. Platyhelminthes--Turbel lar ia , pp. 6 7 - 107. In K. G. ADIYODI and R. G. ADIYODI (eds.) Reproductive Biology of Invertebrates, vol. 1, John Wiley, New York.
HUEBNER, E. and E. ANDERSON. 1967. Comparat ive spiralian oogenesis. Structural aspects: an overview. Amer. Zool. 1 6 : 3 1 5 - 4 3 .
KARNOVSKY, M. H. 1965. A fo rmaldehyde-g lu ta ra ldehyde fixative of high osmolality for use in electron microscopy. J. Cell Biol. 27: 1 3 7 A - 8 A .
KESSEL, R. G. 1966. Electron microscope studies on the origin and maturat ion of yolk in oocytes of the tunicate, Cion~ intestinalis. Z. Zellforsch. Mikrosk. Anat. 7 1 : 5 2 5 - 4 4 .
KESSEL, R. G. and H. W. BEAMS. 1980. Cytodifferentiation and vitellogenesis during oogenesis in Arachnida" cytological studies on developing oocytes of Harvestman. J. MorphoL 163: 1 7 5 - 90.
KESSEL, R. G. and L. R. GANION. 1980a. Electron microscopic and autoradiographic studies on vitellogenesis in Necti~rus maculosus. J. Morphol. 164: 2 1 5 - 33.
KESSEL, R. G. and,L. R. GANION. 1980b. Cytodifferentiation in the Rana pipiens oocyte. V1. The origin and morphogenesis of primary yolk precursor complexes. J. Submicrosc. Cytol. 12: 6 4 7 - 54.
KRESS, A. and U. M. SPORNITZ. 1972. Ultrastructural studies of oogenesis in some European amphibians. 1. Rant' esculenta and Rana temporaria. Z. Zellforsch. Mikrosk. Anat. 128:438 - 56.
LAEMMLI, U. K. 1970. Cleavage of structural proteins during the assembly of the head bacteriophage T 4. Nature (Lond.) 227: 6 8 0 - 5.
LANZAVECCHIA, G. 1968. Problems of the yolk formation in Amphibia. Acc. Naz. Lincei. Problemi attuali di scienza e cultura. Quaderno 104:101 9.
MANS, R. L. and D. G. NOVELLI. 1961. Measurement of incorporation of radioactive amino acids into proteins by a filter paper disk method. Arch. Bioc~m. t~iophys. 94: 4 8 - 63.
MAZZINI, M., G. CALLAINI and C. MENCARELL1. 1984. A comparative analysis of the evolution of the egg envelopes and the origin of the yolk. Boll. ZooL 51:35 - 101.
PAN, M. L., W. J. BELL and W. H. TELFER. 1969. Vitellogenic blood protein synthesis by insect fat body. Science (Wash., D. C.) 1 6 5 : 3 9 3 - 4 .
POSTLETHWAI':, J. H. and F. GIORG1. 1985. Vitellogenesis in insects, pp. 8 5 - 1 2 6 . In L. BROWDER (ed.) Development Biology: A Comprehensive Synthesis. Plenum Press, New York.
ROTH, T. F. and J. W. WOODS. 1982. Fundamental questions in receptor-mediated endocytosis, pp. 163 - 81. In V. T MARCHESl and R. C. GALLO (eds.) Differentiation and Function of Hemopoietic Cell Surfaces. A. R. Liss, New York.
SCHECHTMAN, A. M. 1955. Ontogeny of the blood and related antigens and their significance for the theory of differentiation, pp. 3 - 3 1 . In E. G. BUTLER (ed.) Biological Specificity and Growth. Princeton University Press, Princeton, N.J.
TELFER, W. H., E. RUBENS'rEIN and M. L. PAN. 1981. How the ovary makes yolk in Hyalophora, pp. 637 - 54. In F. SEHNAL, A. ZABZA, J. J. MENN and B. CVMBOROWSKI (eds.) Regulation oflnsect Development and t3ehavior. Technical University, Wroclaw.
TELFER, W. H., E. HUEBNER and D. S. SMITH. 1982. The cell biology of vitellogenic follicles in Hyalophora and Rhodnius, pp. 118-46 . In R. C. KING and H. AKAI (eds.) Insect Ultrastructure, vol. 1, Plenum Press, New York.
WALLACE, R. A. 1978. Oocyte growth in non-mammal ian vertebrates, pp. 4 6 9 - 5 0 2 . In R. E. JONES (ed.) The Vertebrate Ovary. Plenum Press, New York.
WALLACE, R. A., J. M. NICKOL, T. HO and D. W. JArrED. 1972. Studies on amphibian yolk. X. The relative roles of autosynthetic and heterosynthetic processes during yolk protein assembly by isolated amphibian oocytes. Dev. BioL 29: 2 5 5 - 72.
WARD, R. T. ! 978. The origin of protein and fatty yolk in Rana pipiens. IV. Secondary vesicular yolk formation in frog oocytes. Tissue Cell 10:525 - 34.
WYATT, G. R 1980. The fat body as a protein factory, pp. 201 - 25. In M. LOCKE and D. S. SMITH (eds.) Insect Bioh~gy in the Future. Academic Press, New York.

64 M. MAZZINI, M. MASETTI and F. GIORGI
WYATT, G. R. and M. L. PAN. 1978. Insect plasma proteins. Annu. Rev. Biochem. 47: 7 7 9 - 817. WYATT, G. R., M. LOCKE and J. Y. BRADFIELD. 1984, The vitellogenin genes of Locusta migratoria and other
insects, pp. 7 3 - 80. In W. ENGELS (ed.) Advances in Invertebrates Reproduction. vol. 3. Elsevier Science Publishers, Amsterdam.