association between naturally acquired antibodies to erythrocyte‐binding antigens of plasmodium...
TRANSCRIPT

e50 • CID 2010:51 (15 October) • Richards et al
M A J O R A R T I C L E
Association between Naturally Acquired Antibodiesto Erythrocyte-Binding Antigens of Plasmodiumfalciparum and Protection from Malariaand High-Density Parasitemia
Jack S. Richards,1,2 Danielle I. Stanisic,1,3 Freya J. I. Fowkes,1 Livingstone Tavul,3 Elijah Dabod,3
Jennifer K. Thompson,1 Sanjeev Kumar,4 Chetan E. Chitnis,4 David L. Narum,5 Pascal Michon,3 Peter M. Siba,3
Alan F. Cowman,1 Ivo Mueller,3 and James G. Beeson1
1Walter and Eliza Hall Institute of Medical Research and 2Department of Medical Biology, University of Melbourne, Victoria, Australia;3Papua New Guinea Institute of Medical Research, Madang; 4International Centre for Genetic Engineering and Biotechnology, New Delhi,India; and 5Malaria Vaccine Development Branch, National Institute of Allergy and Infectious Diseases, Bethesda, Maryland
Background. Antibodies targeting blood stage antigens are important in protection against malaria, but theprinciple targets remain unclear. Erythrocyte-binding antigens (EBAs) are important erythrocyte invasion ligandsused by merozoites and may be targets of protective immunity, but there are limited data examining their potentialimportance.
Methods. We examined antibodies among 206 Papua New Guinean children who were treated with antimalarialsat enrolment and observed prospectively for 6 months for reinfection and malaria. Immunoglobulin (Ig) G, IgGsubclasses, and IgM to different regions of EBA175, EBA140, and EBA181 expressed as recombinant proteins wereassessed in comparison with several other merozoite antigens.
Results. High levels of IgG to each of the EBAs were strongly associated with protection from symptomaticmalaria and high density parasitemia, but not with risk of reinfection per se. The predominant IgG subclasseswere either IgG1 or IgG3, depending on the antigen. The predominance of IgG1 versus IgG3 reflected structuralfeatures of specific regions of the proteins. IgG3 was most strongly associated with protection, even for thoseantigens that had an IgG1 predominant response.
Conclusions. The EBAs appear important targets of acquired protective immunity. These findings supporttheir further development as vaccine candidates.
Naturally acquired immunity to symptomatic malaria
develops slowly and probably reflects cumulative ex-
posure over many years. Children bear the greatest bur-
den of disease as they develop immunity to severe ma-
laria and, ultimately, mild symptomatic disease. This
immunity is partly achieved through parasite density
control but does not effectively prevent parasitization
per se. Antibodies against merozoite antigens are likely
Received 13 April 2010; accepted 20 July 2010; electronically published 15September 2010.
Reprints or correspondence: Dr James Beeson, The Walter and Eliza HallInstitute of Medical Research, 1G Royal Parade, Parkville, Victoria, Australia, 3050([email protected]).
Clinical Infectious Diseases 2010; 51(8):e50–e60� 2010 by the Infectious Diseases Society of America. All rights reserved.1058-4838/2010/5108-00E1$15.00DOI: 10.1086/656413
to important in mediating protection, but specific tar-
gets and effector responses are largely unknown [1].
The erythrocyte-binding antigens (EBAs) are mem-
bers of the Duffy binding–like (DBL) superfamily and
have orthologues in Plasmodium vivax and Plasmodium
knowlesi [2]. They are characterized by an N-terminal
cysteine rich domain (Region II), a highly conserved
domain (Region III-V), a C-terminal cysteine rich do-
main (Region VI), and transmembrane and cytoplasmic
domains (Figure 1A). Within P. falciparum, homo-
logues include EBA140 (BAEBL), EBA175, and EBA181
(JESEBL) (Figure 1A). (An additional member, EBL1,
is not examined in this article.) The EBAs are located
within the micronemes, are thought to be released onto
the merozoite surface prior to invasion, and may be
involved in tight junction formation [6]. In P. falci-
parum, Region II comprises 2 DBL domains (F1 and

Antibody-Mediated Immunity against EBAs • CID 2010:51 (15 October) • e51
Figure 1. A, Protein structure of EBA175, EBA140, and EBA181 (adapted from [2]). Region I contains a signal sequence and short exon. Region IIin Plasmodium falciparum has 2 cysteine-rich domains with homology to the Duffy-binding proteins of Plasmodium vivax and Plasmodium knowlesi.These 2 Duffy binding–like (DBL) regions, F1 and F2, are thought to mediate binding to the erythrocyte. Region III-V has unclear function and inEBA175 the region contains either an F or C segment, depending on the allelic variant. Region VI is a C-terminal cysteine-rich domain. Region VIIcomprises a transmembrane domain and 2 cytoplasmic domains. Three additional members of the EBA family are not examined in this article; theseinclude (1) EBA165 (PEBL), a pseudogene [3]; (2) MAEBL, which appears to be sporozoite specific [4], with expression profiles suggesting that it isnot expressed at blood-stage and gene-knock out having no observed change in phenotype in blood stage [4]; and (3) EBL-1, the function of whichhas only recently been described [5]. B, Immunoglobulin (Ig) G subclass responses against EBA antigens and AMA1. Bar graphs show the medianoptical density (OD) and interquartile range. Data from seronegative individuals were excluded from analysis (the seropositivity for each subclassresponse is indicated below each bar graph).
F2), which are involved in binding to sialic acid residues on
the surface of erythrocytes [7]. EBA175 contains mutually ex-
clusive F and C segments in Region III-V, which defines the 2
allelic families of EBA175 and which may bind to the glyco-
phorin peptide backbone [8].
Naturally acquired immunity is directed against numerous
merozoite antigens, but it remains unclear which of these, if
any, mediate protection in humans [1]. The EBAs are par-
ticularly promising because of their known role in erythro-
cyte invasion and established erythrocyte-binding receptors;

e52 • CID 2010:51 (15 October) • Richards et al
Table 1. Correlations between Antibody Responses to EBA Antigens of Plasmodium falciparum
EBA antigen
EBA140 RII EBA140 RIII-V EBA175 F2 EBA175 RIII-V EBA181 RIII-V
IgG IgG1 IgG3 IgM IgG IgG1 IgG3 IgM IgG IgG1 IgG3 IgM IgG IgG1 IgG3 IgM IgG IgG1 IgG3 IgM
EBA140 RII
IgG … … … … … … … … … … … … … … … … … … … …
IgG1 0.95 … … … … … … … … … … … … … … … … … … …
IgG3 0.74 0.75 … … … … … … … … … … … … … … … … … …
IgM 0.68 0.66 0.69 … … … … … … … … … … … … … … … … …
EBA140 RIII-V
IgG 0.66 0.67 0.73 0.57 … … … … … … … … … … … … … … … …
IgG1 0.49 0.52 0.46 0.39 0.65 … … … … … … … … … … … … … … …
IgG3 0.54 0.54 0.68 0.56 0.79 0.56 … … … … … … … … … … … … … …
IgM 0.28 0.28 0.38 0.42 0.39 0.44 0.43 … … … … … … … … … … … … …
EBA175 F2
IgG 0.69 0.69 0.66 0.52 0.66 0.47 0.52 0.35 … … … … … … … … … … … …
IgG1 0.64 0.64 0.59 0.47 0.59 0.47 0.47 0.35 0.93 … … … … … … … … … … …
IgG3 0.46 0.44 0.59 0.48 0.59 0.43 0.62 0.42 0.55 0.48 … … … … … … … … … …
IgM 0.17a 0.19a 0.26 0.38 0.32 0.29 0.34 0.56 0.30 0.26 0.27 … … … … … … … … …
EBA175 RIII-V
IgG 0.70 0.71 0.73 0.62 0.75 0.49 0.64 0.42 0.81 0.77 0.60 0.31 … … … … … … … …
IgG1 0.63 0.63 0.60 0.51 0.65 0.56 0.58 0.43 0.73 0.71 0.54 0.27 0.83 … … … … … … …
IgG3 0.66 0.65 0.75 0.59 0.73 0.48 0.68 0.49 0.74 0.70 0.65 0.35 0.90 0.71 … … … … … …
IgM 0.24 0.27 0.30 0.37 0.36 0.40 0.37 0.62 0.39 0.33 0.32 0.60 0.41 0.39 0.43 … … … … …
EBA181 RIII-V
IgG 0.53 0.56 0.54 0.40 0.68 0.57 0.51 0.33 0.52 0.52 0.44 0.24a 0.59 0.56 0.59 0.37 … … … …
IgG1 0.41 0.43 0.46 0.29 0.50 0.43 0.43 0.21a 0.44 0.45 0.38 0.14a 0.48 0.46 0.48 0.21a 0.73 … … …
IgG3 0.43 0.42 0.54 0.37 0.55 0.44 0.51 0.33 0.47 0.47 0.47 0.15a 0.53 0.45 0.59 0.21a 0.64 0.56 … …
IgM 0.39 0.38 0.45 0.51 0.46 0.39 0.49 0.65 0.45 0.40 0.45 0.54 0.49 0.43 0.55 0.62 0.40 0.32 0.41 …
Schizont
IgG 0.61 0.61 0.66 0.53 0.79 0.57 0.63 0.47 0.70 0.65 0.50 0.36 0.75 0.68 0.72 0.49 0.71 0.59 0.54 0.51
NOTE. Spearman rank correlations were determined using the entire cohort (np206). All P values are !.0001, unless otherwise indicated. Ig, immunoglobulin.a P! .05.
EBA175 and EBA140 bind to glycophorin A and C, respective-
ly (EBA181 binding to erythrocyte protein 4.1 has also been
reported) [7, 9, 10]. The structures of Region II (EBA175,
EBA140, and EBA181) and Region VI (EBA175) have been
determined by crystallography or subsequent modeling [11–
13]. Antibodies induced in experimental animals against these
EBAs appear to have invasion-inhibitory activity [9, 12, 14–
17], and acquired human growth-inhibitory antibodies appear
to target EBA175 and possibly other EBAs [15]. Balancing se-
lection, a possible indicator of immune selection pressure, is
evident for EBA175, but not for EBA140 and EBA181 [12, 18,
19]. Although all parasite strains express the 3 functional EBAs,
the utilization of these ligands and their ability to switch to
alternate invasion pathways varies between isolates [12, 20–23].
This may be a further mechanism of evading protective anti-
bodies but does not seem to influence severity of clinical out-
comes [15, 23]. Presently, only a few prospective studies have
examined EBA175 responses (most looking at RII only) and
protection from malaria, and to our knowledge, none have
examined EBA140 and EBA181 [24–27].
This study provides a unique insight into naturally acquired
antibody responses to the EBAs in a treatment-reinfection study
of children. It has enabled examination of antibodies and the
prospective risk of reinfection, symptomatic malaria, and high-
density parasitemia. We evaluated antibody responses to dif-
ferent regions of EBA175, EBA140, and EBA181, some of which
have not previously been described. Recent studies suggest that
immunoglobulin (Ig) G1 and IgG3 subclass responses may vary
according to the specific antigen being examined and that spe-
cific subclass responses may be required for protective responses
[28–31]. Putative protective effector mechanisms for the EBAs
were explored through examination of IgG subclass responses.
MATERIALS AND METHODS
Details of the study population and ethics approval. Plasma
samples were obtained from a prospective treatment re-infec-
Antibody-Mediated Immunity against EBAs • CID 2010:51 (15 October) • e53
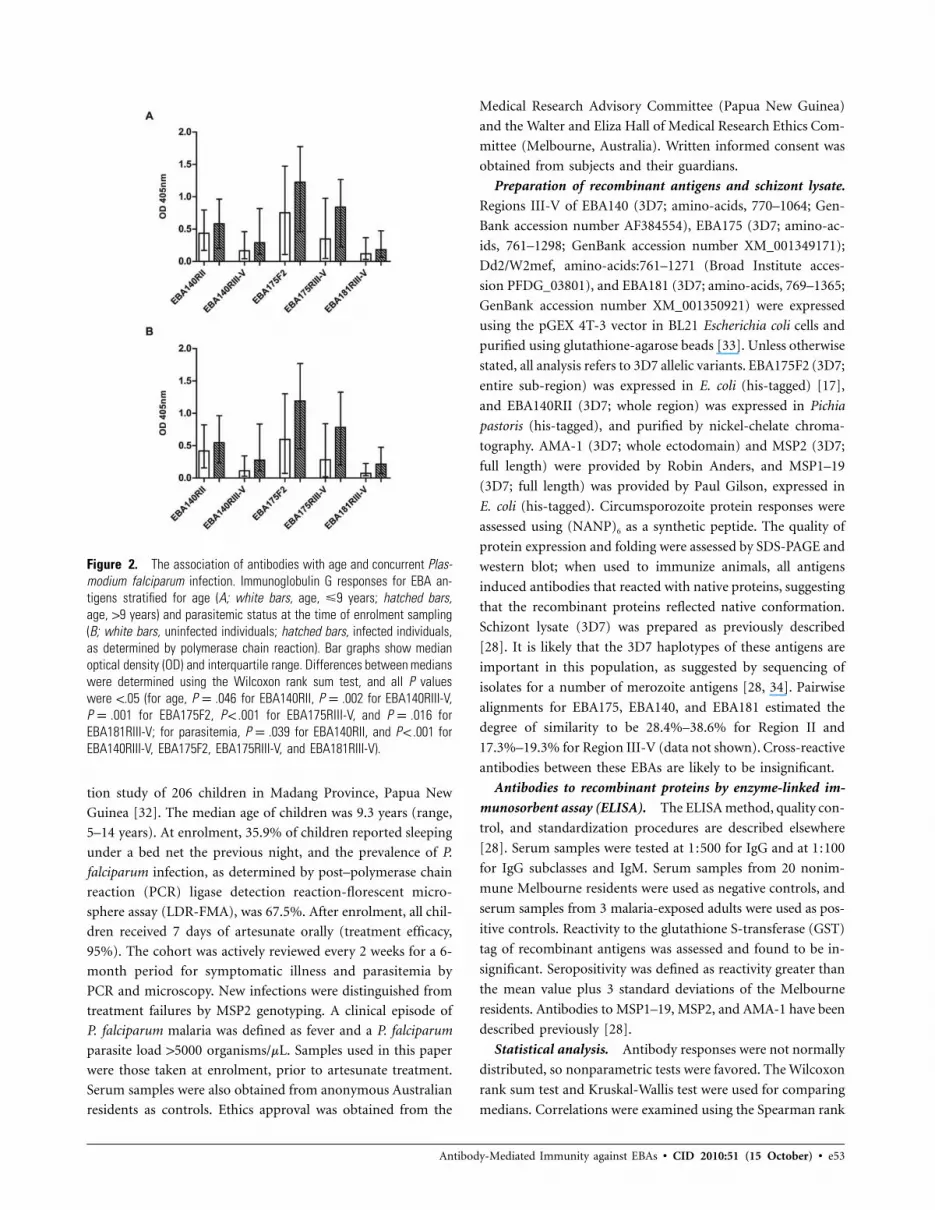
Figure 2. The association of antibodies with age and concurrent Plas-modium falciparum infection. Immunoglobulin G responses for EBA an-tigens stratified for age (A; white bars, age, �9 years; hatched bars,age, 19 years) and parasitemic status at the time of enrolment sampling(B; white bars, uninfected individuals; hatched bars, infected individuals,as determined by polymerase chain reaction). Bar graphs show medianoptical density (OD) and interquartile range. Differences between medianswere determined using the Wilcoxon rank sum test, and all P valueswere !.05 (for age, P p .046 for EBA140RII, P p .002 for EBA140RIII-V,P p .001 for EBA175F2, P ! .001 for EBA175RIII-V, and P p .016 forEBA181RIII-V; for parasitemia, P p .039 for EBA140RII, and P ! .001 forEBA140RIII-V, EBA175F2, EBA175RIII-V, and EBA181RIII-V).
tion study of 206 children in Madang Province, Papua New
Guinea [32]. The median age of children was 9.3 years (range,
5–14 years). At enrolment, 35.9% of children reported sleeping
under a bed net the previous night, and the prevalence of P.
falciparum infection, as determined by post–polymerase chain
reaction (PCR) ligase detection reaction-florescent micro-
sphere assay (LDR-FMA), was 67.5%. After enrolment, all chil-
dren received 7 days of artesunate orally (treatment efficacy,
95%). The cohort was actively reviewed every 2 weeks for a 6-
month period for symptomatic illness and parasitemia by
PCR and microscopy. New infections were distinguished from
treatment failures by MSP2 genotyping. A clinical episode of
P. falciparum malaria was defined as fever and a P. falciparum
parasite load 15000 organisms/mL. Samples used in this paper
were those taken at enrolment, prior to artesunate treatment.
Serum samples were also obtained from anonymous Australian
residents as controls. Ethics approval was obtained from the
Medical Research Advisory Committee (Papua New Guinea)
and the Walter and Eliza Hall of Medical Research Ethics Com-
mittee (Melbourne, Australia). Written informed consent was
obtained from subjects and their guardians.
Preparation of recombinant antigens and schizont lysate.
Regions III-V of EBA140 (3D7; amino-acids, 770–1064; Gen-
Bank accession number AF384554), EBA175 (3D7; amino-ac-
ids, 761–1298; GenBank accession number XM_001349171);
Dd2/W2mef, amino-acids:761–1271 (Broad Institute acces-
sion PFDG_03801), and EBA181 (3D7; amino-acids, 769–1365;
GenBank accession number XM_001350921) were expressed
using the pGEX 4T-3 vector in BL21 Escherichia coli cells and
purified using glutathione-agarose beads [33]. Unless otherwise
stated, all analysis refers to 3D7 allelic variants. EBA175F2 (3D7;
entire sub-region) was expressed in E. coli (his-tagged) [17],
and EBA140RII (3D7; whole region) was expressed in Pichia
pastoris (his-tagged), and purified by nickel-chelate chroma-
tography. AMA-1 (3D7; whole ectodomain) and MSP2 (3D7;
full length) were provided by Robin Anders, and MSP1–19
(3D7; full length) was provided by Paul Gilson, expressed in
E. coli (his-tagged). Circumsporozoite protein responses were
assessed using (NANP)6 as a synthetic peptide. The quality of
protein expression and folding were assessed by SDS-PAGE and
western blot; when used to immunize animals, all antigens
induced antibodies that reacted with native proteins, suggesting
that the recombinant proteins reflected native conformation.
Schizont lysate (3D7) was prepared as previously described
[28]. It is likely that the 3D7 haplotypes of these antigens are
important in this population, as suggested by sequencing of
isolates for a number of merozoite antigens [28, 34]. Pairwise
alignments for EBA175, EBA140, and EBA181 estimated the
degree of similarity to be 28.4%–38.6% for Region II and
17.3%–19.3% for Region III-V (data not shown). Cross-reactive
antibodies between these EBAs are likely to be insignificant.
Antibodies to recombinant proteins by enzyme-linked im-
munosorbent assay (ELISA). The ELISA method, quality con-
trol, and standardization procedures are described elsewhere
[28]. Serum samples were tested at 1:500 for IgG and at 1:100
for IgG subclasses and IgM. Serum samples from 20 nonim-
mune Melbourne residents were used as negative controls, and
serum samples from 3 malaria-exposed adults were used as pos-
itive controls. Reactivity to the glutathione S-transferase (GST)
tag of recombinant antigens was assessed and found to be in-
significant. Seropositivity was defined as reactivity greater than
the mean value plus 3 standard deviations of the Melbourne
residents. Antibodies to MSP1–19, MSP2, and AMA-1 have been
described previously [28].
Statistical analysis. Antibody responses were not normally
distributed, so nonparametric tests were favored. The Wilcoxon
rank sum test and Kruskal-Wallis test were used for comparing
medians. Correlations were examined using the Spearman rank
e54 • CID 2010:51 (15 October) • Richards et al
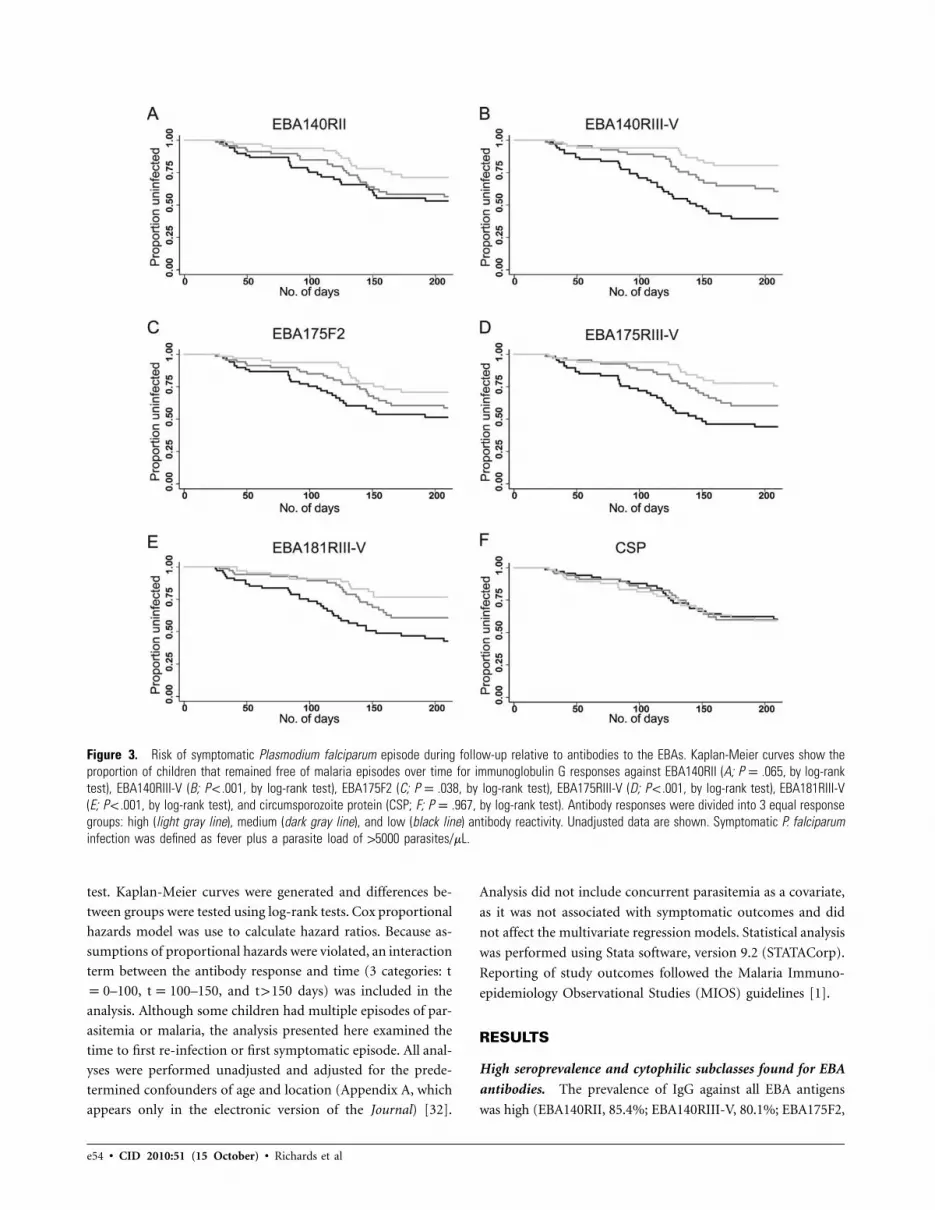
Figure 3. Risk of symptomatic Plasmodium falciparum episode during follow-up relative to antibodies to the EBAs. Kaplan-Meier curves show theproportion of children that remained free of malaria episodes over time for immunoglobulin G responses against EBA140RII (A; P p .065, by log-ranktest), EBA140RIII-V (B; P ! .001, by log-rank test), EBA175F2 (C; P p .038, by log-rank test), EBA175RIII-V (D; P ! .001, by log-rank test), EBA181RIII-V(E; P ! .001, by log-rank test), and circumsporozoite protein (CSP; F; P p .967, by log-rank test). Antibody responses were divided into 3 equal responsegroups: high (light gray line), medium (dark gray line), and low (black line) antibody reactivity. Unadjusted data are shown. Symptomatic P. falciparuminfection was defined as fever plus a parasite load of 15000 parasites/mL.
test. Kaplan-Meier curves were generated and differences be-
tween groups were tested using log-rank tests. Cox proportional
hazards model was use to calculate hazard ratios. Because as-
sumptions of proportional hazards were violated, an interaction
term between the antibody response and time (3 categories: t
p0–100, tp100–150, and t 1 150 days) was included in the
analysis. Although some children had multiple episodes of par-
asitemia or malaria, the analysis presented here examined the
time to first re-infection or first symptomatic episode. All anal-
yses were performed unadjusted and adjusted for the prede-
termined confounders of age and location (Appendix A, which
appears only in the electronic version of the Journal) [32].
Analysis did not include concurrent parasitemia as a covariate,
as it was not associated with symptomatic outcomes and did
not affect the multivariate regression models. Statistical analysis
was performed using Stata software, version 9.2 (STATACorp).
Reporting of study outcomes followed the Malaria Immuno-
epidemiology Observational Studies (MIOS) guidelines [1].
RESULTS
High seroprevalence and cytophilic subclasses found for EBA
antibodies. The prevalence of IgG against all EBA antigens
was high (EBA140RII, 85.4%; EBA140RIII-V, 80.1%; EBA175F2,
Antibody-Mediated Immunity against EBAs • CID 2010:51 (15 October) • e55
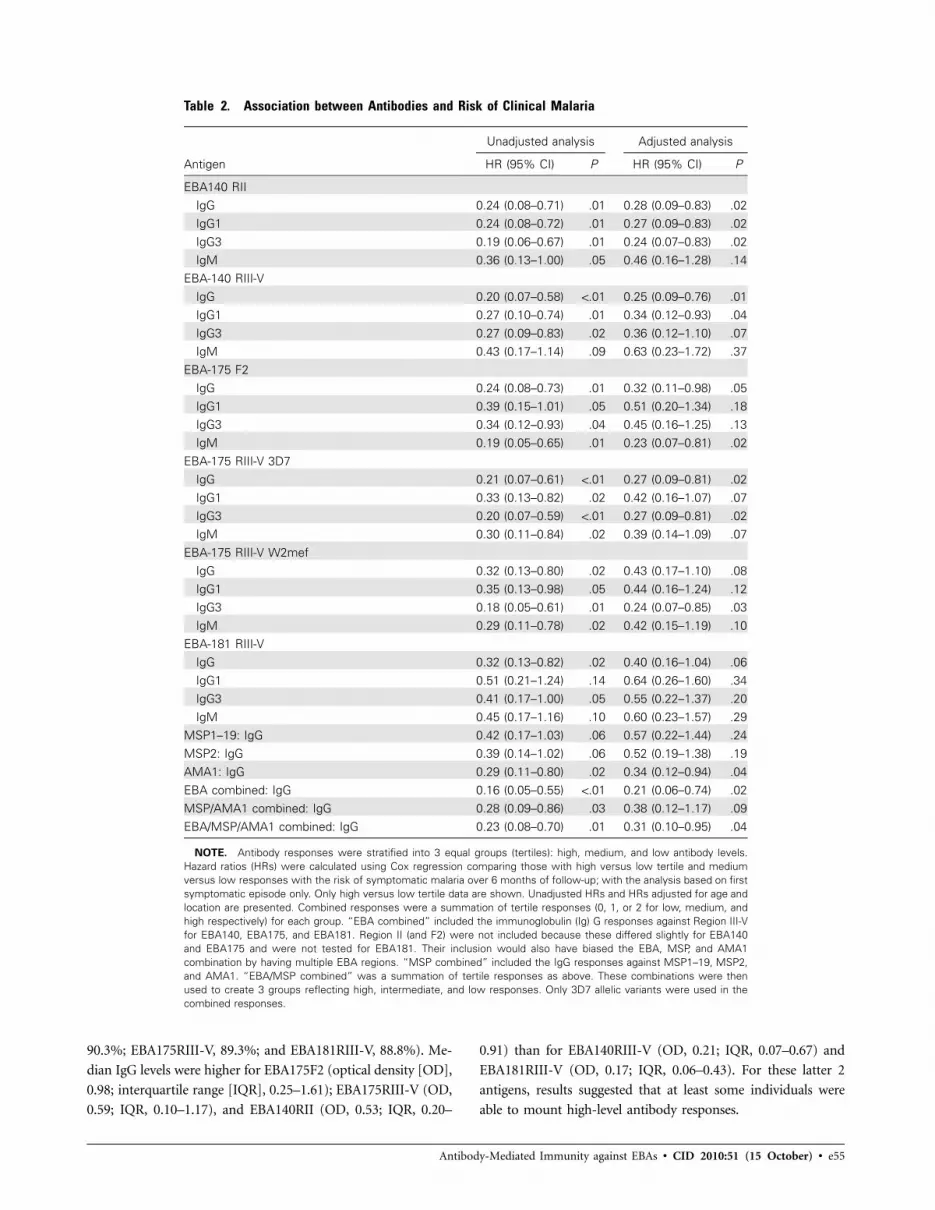
Table 2. Association between Antibodies and Risk of Clinical Malaria
Antigen
Unadjusted analysis Adjusted analysis
HR (95% CI) P HR (95% CI) P
EBA140 RIIIgG 0.24 (0.08–0.71) .01 0.28 (0.09–0.83) .02IgG1 0.24 (0.08–0.72) .01 0.27 (0.09–0.83) .02IgG3 0.19 (0.06–0.67) .01 0.24 (0.07–0.83) .02IgM 0.36 (0.13–1.00) .05 0.46 (0.16–1.28) .14
EBA-140 RIII-VIgG 0.20 (0.07–0.58) !.01 0.25 (0.09–0.76) .01IgG1 0.27 (0.10–0.74) .01 0.34 (0.12–0.93) .04IgG3 0.27 (0.09–0.83) .02 0.36 (0.12–1.10) .07IgM 0.43 (0.17–1.14) .09 0.63 (0.23–1.72) .37
EBA-175 F2IgG 0.24 (0.08–0.73) .01 0.32 (0.11–0.98) .05IgG1 0.39 (0.15–1.01) .05 0.51 (0.20–1.34) .18IgG3 0.34 (0.12–0.93) .04 0.45 (0.16–1.25) .13IgM 0.19 (0.05–0.65) .01 0.23 (0.07–0.81) .02
EBA-175 RIII-V 3D7IgG 0.21 (0.07–0.61) !.01 0.27 (0.09–0.81) .02IgG1 0.33 (0.13–0.82) .02 0.42 (0.16–1.07) .07IgG3 0.20 (0.07–0.59) !.01 0.27 (0.09–0.81) .02IgM 0.30 (0.11–0.84) .02 0.39 (0.14–1.09) .07
EBA-175 RIII-V W2mefIgG 0.32 (0.13–0.80) .02 0.43 (0.17–1.10) .08IgG1 0.35 (0.13–0.98) .05 0.44 (0.16–1.24) .12IgG3 0.18 (0.05–0.61) .01 0.24 (0.07–0.85) .03IgM 0.29 (0.11–0.78) .02 0.42 (0.15–1.19) .10
EBA-181 RIII-VIgG 0.32 (0.13–0.82) .02 0.40 (0.16–1.04) .06IgG1 0.51 (0.21–1.24) .14 0.64 (0.26–1.60) .34IgG3 0.41 (0.17–1.00) .05 0.55 (0.22–1.37) .20IgM 0.45 (0.17–1.16) .10 0.60 (0.23–1.57) .29
MSP1–19: IgG 0.42 (0.17–1.03) .06 0.57 (0.22–1.44) .24MSP2: IgG 0.39 (0.14–1.02) .06 0.52 (0.19–1.38) .19AMA1: IgG 0.29 (0.11–0.80) .02 0.34 (0.12–0.94) .04EBA combined: IgG 0.16 (0.05–0.55) !.01 0.21 (0.06–0.74) .02MSP/AMA1 combined: IgG 0.28 (0.09–0.86) .03 0.38 (0.12–1.17) .09EBA/MSP/AMA1 combined: IgG 0.23 (0.08–0.70) .01 0.31 (0.10–0.95) .04
NOTE. Antibody responses were stratified into 3 equal groups (tertiles): high, medium, and low antibody levels.Hazard ratios (HRs) were calculated using Cox regression comparing those with high versus low tertile and mediumversus low responses with the risk of symptomatic malaria over 6 months of follow-up; with the analysis based on firstsymptomatic episode only. Only high versus low tertile data are shown. Unadjusted HRs and HRs adjusted for age andlocation are presented. Combined responses were a summation of tertile responses (0, 1, or 2 for low, medium, andhigh respectively) for each group. “EBA combined” included the immunoglobulin (Ig) G responses against Region III-Vfor EBA140, EBA175, and EBA181. Region II (and F2) were not included because these differed slightly for EBA140and EBA175 and were not tested for EBA181. Their inclusion would also have biased the EBA, MSP, and AMA1combination by having multiple EBA regions. “MSP combined” included the IgG responses against MSP1–19, MSP2,and AMA1. “EBA/MSP combined” was a summation of tertile responses as above. These combinations were thenused to create 3 groups reflecting high, intermediate, and low responses. Only 3D7 allelic variants were used in thecombined responses.
90.3%; EBA175RIII-V, 89.3%; and EBA181RIII-V, 88.8%). Me-
dian IgG levels were higher for EBA175F2 (optical density [OD],
0.98; interquartile range [IQR], 0.25–1.61); EBA175RIII-V (OD,
0.59; IQR, 0.10–1.17), and EBA140RII (OD, 0.53; IQR, 0.20–
0.91) than for EBA140RIII-V (OD, 0.21; IQR, 0.07–0.67) and
EBA181RIII-V (OD, 0.17; IQR, 0.06–0.43). For these latter 2
antigens, results suggested that at least some individuals were
able to mount high-level antibody responses.

e56 • CID 2010:51 (15 October) • Richards et al
Figure 4. Immunoglobulin (Ig) G, IgG1, and IgG3 responses to EBAs, according to Plasmodium falciparum symptomatic outcomes. Children wereclassified into 3 groups on the basis of their experience of symptomatic malaria episodes in the 6-month period after drug treatment: “protected,”which was defined as no episodes of symptomatic malaria (white bars; n p 73); “single-susceptible,” which was defined as a single episode ofsymptomatic malaria (black bars; n p 27), and “multisusceptible,” which was defined as 11 episode of symptomatic malaria (dashed bars; n p 54).Individuals who were not reinfected or who did not fulfill the category criteria were excluded. Bars indicate median optical density (OD), and errorbars indicate interquartile range. Symptomatic P. falciparum malaria was defined as fever plus a parasite load 15000 parasites/mL. Differences betweenmedians were determined by Kruskal-Wallis test, and all P values were !.05 (except for EBA175F2 IgG1, for which P p .058).
Specific IgG subclasses may determine the type of effector
response. IgG1 and IgG3, known cytophilic subclasses, were of
particular interest. IgG3 was predominant for EBA175RIII-V
and EBA140RIII-V, IgG1 was predominant for EBA140RII and
EBA175F2 (and AMA-1), and a mixed IgG1 and IgG3 response
was predmoniant for EBA181RIII-V (Figure 1B). This suggests
IgG1-predominant responses to cysteine-rich regions (Region
II), but greater IgG3 responses to Regions III-V. There was
negligible IgG2 and IgG4 detected. IgM responses were modest,
with seropositivity rates of 21.4%–60.2% and low antibody
levels (data not shown).
Correlations for IgG and subclass specific responses against
different EBAs. Antibodies to different antigens are thought
to develop concurrently, though probably at different rates.
Therefore, we examined correlations between IgG and IgG
subclasses to each antigen (Table 1). When examining a sin-
gle region, strong correlations were observed between total IgG
levels and the predominant subclass response (eg, EBA140RII
Antibody-Mediated Immunity against EBAs • CID 2010:51 (15 October) • e57
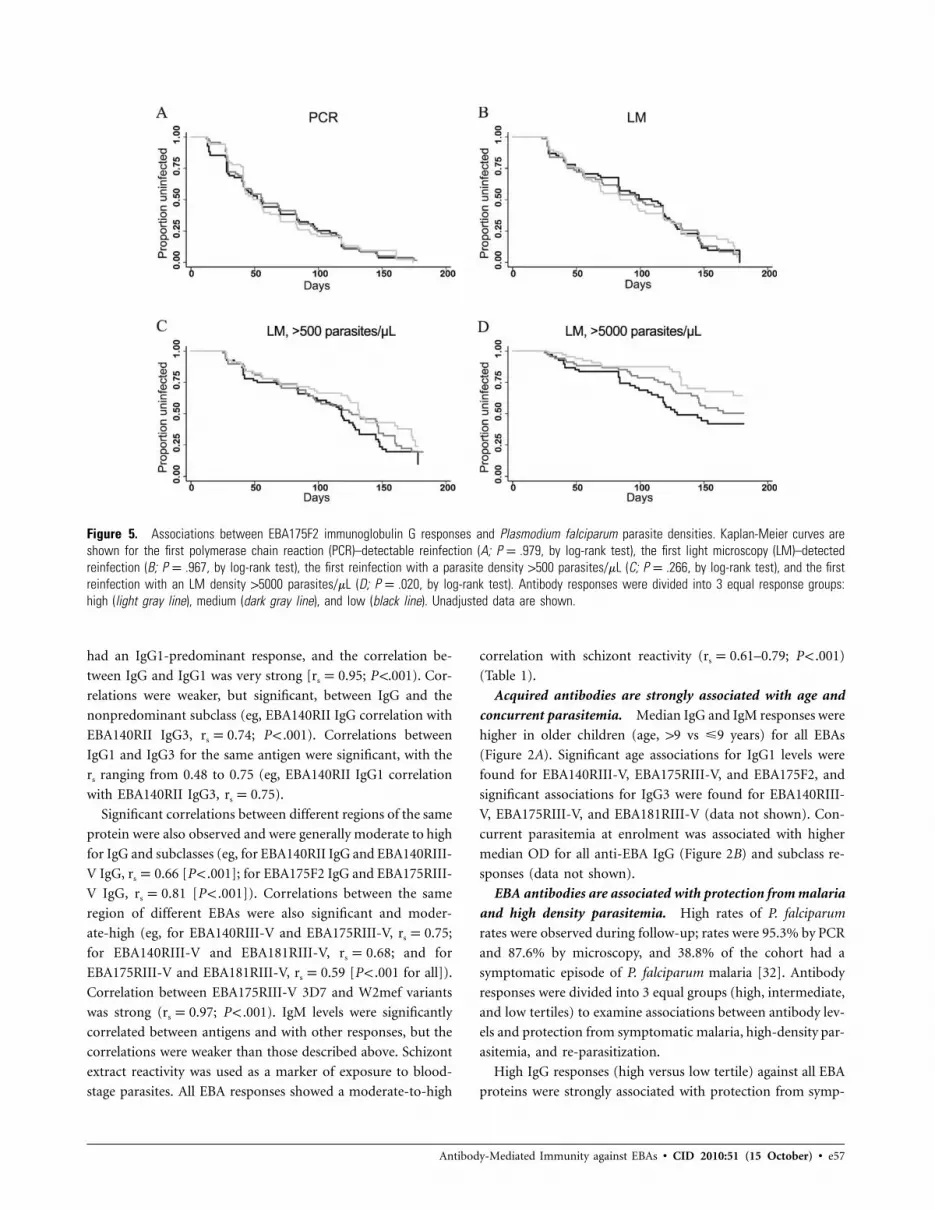
Figure 5. Associations between EBA175F2 immunoglobulin G responses and Plasmodium falciparum parasite densities. Kaplan-Meier curves areshown for the first polymerase chain reaction (PCR)–detectable reinfection (A; P p .979, by log-rank test), the first light microscopy (LM)–detectedreinfection (B; P p .967, by log-rank test), the first reinfection with a parasite density 1500 parasites/mL (C; P p .266, by log-rank test), and the firstreinfection with an LM density 15000 parasites/mL (D; P p .020, by log-rank test). Antibody responses were divided into 3 equal response groups:high (light gray line), medium (dark gray line), and low (black line). Unadjusted data are shown.
had an IgG1-predominant response, and the correlation be-
tween IgG and IgG1 was very strong [rs p0.95; P !.001). Cor-
relations were weaker, but significant, between IgG and the
nonpredominant subclass (eg, EBA140RII IgG correlation with
EBA140RII IgG3, rs p0.74; P ! .001). Correlations between
IgG1 and IgG3 for the same antigen were significant, with the
rs ranging from 0.48 to 0.75 (eg, EBA140RII IgG1 correlation
with EBA140RII IgG3, rs p0.75).
Significant correlations between different regions of the same
protein were also observed and were generally moderate to high
for IgG and subclasses (eg, for EBA140RII IgG and EBA140RIII-
V IgG, rs p0.66 [P ! .001]; for EBA175F2 IgG and EBA175RIII-
V IgG, rs p 0.81 [P ! .001]). Correlations between the same
region of different EBAs were also significant and moder-
ate-high (eg, for EBA140RIII-V and EBA175RIII-V, rs p0.75;
for EBA140RIII-V and EBA181RIII-V, rs p 0.68; and for
EBA175RIII-V and EBA181RIII-V, rs p0.59 [P ! .001 for all]).
Correlation between EBA175RIII-V 3D7 and W2mef variants
was strong (rs p0.97; P ! .001). IgM levels were significantly
correlated between antigens and with other responses, but the
correlations were weaker than those described above. Schizont
extract reactivity was used as a marker of exposure to blood-
stage parasites. All EBA responses showed a moderate-to-high
correlation with schizont reactivity (rs p0.61–0.79; P ! .001)
(Table 1).
Acquired antibodies are strongly associated with age and
concurrent parasitemia. Median IgG and IgM responses were
higher in older children (age, 19 vs �9 years) for all EBAs
(Figure 2A). Significant age associations for IgG1 levels were
found for EBA140RIII-V, EBA175RIII-V, and EBA175F2, and
significant associations for IgG3 were found for EBA140RIII-
V, EBA175RIII-V, and EBA181RIII-V (data not shown). Con-
current parasitemia at enrolment was associated with higher
median OD for all anti-EBA IgG (Figure 2B) and subclass re-
sponses (data not shown).
EBA antibodies are associated with protection from malaria
and high density parasitemia. High rates of P. falciparum
rates were observed during follow-up; rates were 95.3% by PCR
and 87.6% by microscopy, and 38.8% of the cohort had a
symptomatic episode of P. falciparum malaria [32]. Antibody
responses were divided into 3 equal groups (high, intermediate,
and low tertiles) to examine associations between antibody lev-
els and protection from symptomatic malaria, high-density par-
asitemia, and re-parasitization.
High IgG responses (high versus low tertile) against all EBA
proteins were strongly associated with protection from symp-

e58 • CID 2010:51 (15 October) • Richards et al
tomatic malaria (unadjusted hazard ratios (HRs) were 0.24 for
EBA140F2, 0.20 for EBA140RIII-V, 0.24 for EBA175F2, 0.21
for EBA175RIII-V 3D7, and 0.32 for EBA181RIII-V [P ! .05 for
all]) (Figure 3 and Table 2). Adjusting for the primary con-
founders of age and location had little effect on unadjusted
HRs (Table 2). HRs for all EBA constructs (comparing high
versus low tertile responses) were considerably lower than for
the other leading vaccine candidates, MSP1–19 (unadjusted
HR, 0.42 [Pp .06]; adjusted HR, 0.57 [P p .24]) and MSP2
(unadjusted HR, 0.39 [Pp .06]; adjusted HR, 0.52, [Pp .19]),
but were comparable to AMA1 (unadjusted HR, 0.29 [Pp .02];
adjusted HR, 0.34 [Pp .04]). Antibody levels against circum-
sporozoite protein peptide were not associated with protection
against symptomatic malaria (unadjusted HR for high versus
low tertile, 1.58 [95% confidence interval {CI}, 0.65–3.87; P
p .32]; adjusted HR, 1.86 [95% CI, 0.75–4.57; P p .18]). Com-
bined responses for the EBAs (unadjusted HR, .16; P ! .01) and
MSP/AMA1 (unadjusted HR, 0.28; Pp .03) were more strong-
ly associated with protection than were responses to single an-
tigens alone (Table 2). A combined EBA, MSP, and AMA1
response (unadjusted HR, 0.23; P p .01) did not appear to
be associated with stronger protection than the EBA combined
response.
IgG3 responses (high versus low tertile) showed stronger as-
sociations with protection than IgG1 or IgM, even for EBA175F2
and EBA140RII in which the predominant subclass response was
IgG1 (IgG3 unadjusted HR ranging 0.19–0.41 for all EBAs, Table
2). These results were largely unaffected when adjusting for age
and location. Protective associations were also seen for IgG1
responses, although these were more modest than those of IgG3
and generally had a lower level of significance (Table 2). IgM
responses were also associated with protection for all antigen
constructs but these associations were generally lost with ad-
justing for covariates.
The association between antibody levels and protection from
symptomatic malaria appeared to have a dose-response rela-
tionship (Figure 3). In each plot, high responders (upper tertile)
had fewer symptomatic episodes than did medium or low re-
sponders. The unadjusted HR for the medium versus low re-
sponses were strongly associated with protection for RIII-V but
not for RII (data not shown). The importance of antibody levels
in conferring protection was further explored by classifying the
cohort into 3 groups according to symptomatic outcomes: pro-
tected (no symptomatic episodes), single-susceptible (1 symp-
tomatic episode), and multisusceptible (multiple symptomatic
episodes; individuals who were not reinfected were excluded
from this analysis). In all instances, the protected group had
higher median IgG and subclass levels, compared with the sus-
ceptible groups, and there were generally lower antibody levels
among the multisusceptible group than among the single-sus-
ceptible group (Figure 4). The greatest difference was seen for
RIII-V, in which antibodies were 2–8 times higher in the pro-
tected group than the susceptible groups (eg, the median an-
tibody level for EBA175RIII-V was 0.89 for the protected group,
0.43 for the single susceptible group, and 0.11 for the multi-
susceptible group; P ! .001).
IgG responses against all EBA antigens were associated with
protection from high -density infection (parasite load, 15000
parasites/mL) but not with prevention of reinfection (Figure 5).
Findings were similar for IgG3 responses but more moderate
for IgG1 responses (data not shown).
DISCUSSION
These results suggest that EBA antibodies are likely to be im-
portant in protection from P. falciparum malaria. Children with
high levels of antibodies had a greatly reduced risk of symp-
tomatic malaria, and protected children had higher levels of
antibodies than did those who developed clinical malaria. Few
prospective studies have used a treatment reinfection design
that enables the detection of episodes of symptomatic malaria,
high-density parasitemia, and reparasitization, as used here [1].
This study design enabled us to determine that EBA antibodies
were strongly associated with protection from symptomatic ma-
laria and high-density parasitemia but, as expected, did not
prevent reinfection. It is also significant that EBA antibodies
were more strongly associated with protection from symptom-
atic P. falciparum malaria than favored vaccine candidates
MSP1–19 and MSP2 but were similar to AMA-1. The strongest
associations with protective immunity were observed with an-
tibodies to Region III-V rather than Region II (or F2 sub-
domain). Region III-V is free of cysteine residues and has lim-
ited polymorphism. It is also known that a-EBA175 and
a-EBA181 antibodies generated against this region inhibit in
vitro parasite growth, supporting their functional relevance
(A.F.C., unpublished data) [14]. Much of the focus of research
on the EBAs has been on Region II, which mediates erythrocyte
binding, but little is known about the functional role of other
regions of EBA proteins. Our findings support the need for
additional studies to define the function of Region III-V and
its consideration in vaccine development. Combining IgG re-
sponses to several EBAs decreased HRs, suggesting that a broad
EBA response may confer greater protection than to single
EBAs alone.
The prevalence of IgG to the EBAs was high in this cohort
of children from Papua New Guinea, and our results are con-
sistent with previous data describing age-related acquisition of
antibodies to the EBAs [15, 25, 26]. This probably reflects cu-
mulative exposure to parasites over time and was further sug-
gested by finding that levels of EBA antibodies were higher with
increasing reactivity to schizont protein extract. Active para-
sitemia at the time of sampling was found to be associated with
higher antibody levels, suggesting a boosting effect or greater

Antibody-Mediated Immunity against EBAs • CID 2010:51 (15 October) • e59
exposure among these individuals. Almost all prior studies have
detected parasitization with light microscopy only, which un-
derestimates rates of parasitization and thus the important ef-
fect this has on antibody levels [32]. This study utilized sensitive
PCR-based measures of parasitization in addition to light mi-
croscopy; this enabled us to examine whether antibodies pro-
tected from low density reparasitization.
We found that EBA antibodies are predominately of IgG1
or IgG3 subclasses, as reported in other populations, but did
not find evidence of IgG2 or IgG4 responses [24, 25, 35]. An-
tibodies to the EBAs may exert a protective effect by inhibiting
erythrocyte invasion, thereby facilitating control of parasitemia
and decreasing associated morbidity. Antibodies generated in
laboratory animals by vaccination with EBA175 and EBA140
inhibit invasion, supporting this notion [9, 16]. However, the
cytophilic nature of IgG1 and IgG3 suggests that antibodies to
EBAs may also interact with macrophages, granulocytes, and/
or NK cells or activate complement to mediate parasite clear-
ance. Interestingly, protective associations were stronger with
IgG3 than IgG1, even for Region II (or F2), which had an IgG1-
predominant response, implying IgG3 may be functionally
more important. This needs further examination, because any
future vaccine may need to elicit subclass-specific responses to
maximize efficacy.
Acquired immunity to symptomatic malaria is likely to in-
volve responses against a multitude of antigens. Dissecting the
relative contribution of each of these remains a great challenge.
Our findings are consistent with increasing evidence indicating
that the EBAs are promising vaccine candidates [36]. There is
also evidence of balancing selection for EBA175, suggesting the
possibility of immune selection pressure, but the EBAs have
considerably less antigenic diversity than other merozoite pro-
teins. This may minimize the number of variants required for
vaccine development [18]. Aotus monkeys immunized with
EBA175RII (DNA and recombinant antigen) vaccines were af-
forded partial protection from parasite challenge [37, 38], pro-
viding some proof-of-principle for using EBAs as vaccines. A
potential challenge for EBA vaccine development is to over-
come invasion pathway redundancy and the ability of parasites
to use different invasion pathways as a possible immune evasion
strategy [12]. This has been further highlighted by studies show-
ing that parasites remain viable despite genetic disruption of
EBA175, EBA140, and EBA181, either singly or in combination
[12, 33, 39, 40].
This study provides important evidence suggesting that the
EBAs are targets of protective immunity in humans and sup-
ports their evaluation as potential vaccine candidates. Further
research is required to understand mechanisms mediating pro-
tection, the importance and function of specific subclass re-
sponses, and the importance of different regions of the EBAs
in invasion and as targets of protective antibodies.
Acknowledgments
We thank the study participants, staff involved in the study at the PapuaNew Guinea Institute of Medical Research, Madang, and Fiona McCallum,Linda Reiling, and Julie Simpson for their ideas and discussions. Humanerythrocytes and serum were provided by the Australian Red Cross BloodService, Melbourne, Australia.
Financial support. The National Health and Medical Research Councilof Australia (project grant and career development award to J.G.B., post-graduate research fellowship to J.S.R., and training award to F.J.F. [IRIISSgrant 361646]), the Australia-India Strategic Research Fund (Departmentof Innovation, Industry, Science, and Research, Australian Government),Australian Research Council (Future Fellowship to J.G.B.), and the Vic-torian State Government Operational Infrastructure Support grant.
Potential conflicts of interest. All authors: no conflicts.
References
1. Fowkes FJ, Richards JS, Simpson JA, Beeson JG. The relationship be-tween anti-merozoite antibodies and incidence of Plasmodium falci-parum malaria: a systematic review and meta-analysis. PLoS Med 2010;7:e1000218.
2. Adams JH, Blair PL, Kaneko O, Peterson DS. An expanding ebl familyof Plasmodium falciparum. Trends Parasitol 2001; 17:297–299.
3. Triglia T, Thompson JK, Cowman AF. An EBA175 homologue whichis transcribed but not translated in erythrocytic stages of Plasmodiumfalciparum. Mol Biochem Parasitol 2001; 116:55–63.
4. Fu J, Saenz FE, Reed MB, et al. Targeted disruption of maebl in Plas-modium falciparum. Mol Biochem Parasitol 2005; 141:113–117.
5. Mayer DC, Cofie J, Jiang L, et al. Glycophorin B is the erythrocytereceptor of Plasmodium falciparum erythrocyte-binding ligand, EBL-1. Proc Natl Acad Sci USA 2009; 106:5348–5352.
6. Sim BK, Toyoshima T, Haynes JD, Aikawa M. Localization of the 175-kilodalton erythrocyte binding antigen in micronemes of Plasmodiumfalciparum merozoites. Mol Biochem Parasitol 1992; 51:157–159.
7. Sim BK, Chitnis CE, Wasniowska K, Hadley TJ, Miller LH. Receptorand ligand domains for invasion of erythrocytes by Plasmodium fal-ciparum. Science 1994; 264:1941–1944.
8. Kain KC, Orlandi PA, Haynes JD, Sim KL, Lanar DE. Evidence fortwo-stage binding by the 175-kD erythrocyte binding antigen of Plas-modium falciparum. J Exp Med 1993; 178:1497–1505.
9. Maier AG, Duraisingh MT, Reeder JC, et al. Plasmodium falciparumerythrocyte invasion through glycophorin C and selection for Gerbichnegativity in human populations. Nat Med 2003; 9:87–92.
10. Lanzillotti R, Coetzer TL. The 10 kDa domain of human erythrocyteprotein 4.1 binds the Plasmodium falciparum EBA-181 protein. MalarJ 2006; 5:100.
11. Tolia NH, Enemark EJ, Sim BK, Joshua-Tor L. Structural basis for theEBA-175 erythrocyte invasion pathway of the malaria parasite Plas-modium falciparum. Cell 2005; 122:183–193.
12. Maier AG, Baum J, Smith B, Conway DJ, Cowman AF. Polymorphismsin erythrocyte binding antigens 140 and 181 affect function and bindingbut not receptor specificity in Plasmodium falciparum. Infect Immun2009; 77:1689–1699.
13. Withers-Martinez C, Haire LF, Hackett F, et al. Malarial EBA-175 regionVI crystallographic structure reveals a KIX-like binding interface. J MolBiol 2008; 375:773–781.
14. Baum J, Maier AG, Good RT, Simpson KM, Cowman AF. Invasion byP. falciparum merozoites suggests a hierarchy of molecular interactions.PLoS Pathog 2005; 1:e37.
15. Persson KE, McCallum FJ, Reiling L, et al. Variation in use of eryth-rocyte invasion pathways by Plasmodium falciparum mediates evasionof human inhibitory antibodies. J Clin Invest 2008; 118:342–351.
16. Narum DL, Haynes JD, Fuhrmann S, et al. Antibodies against the Plas-modium falciparum receptor binding domain of EBA-175 block inva-sion pathways that do not involve sialic acids. Infect Immun 2000; 68:1964–1966.

e60 • CID 2010:51 (15 October) • Richards et al
17. Pandey KC, Singh S, Pattnaik P, et al. Bacterially expressed and refoldedreceptor binding domain of Plasmodium falciparum EBA-175 elicitsinvasion inhibitory antibodies. Mol Biochem Parasitol 2002; 123:23–33.
18. Baum J, Thomas AW, Conway DJ. Evidence for diversifying selectionon erythrocyte-binding antigens of Plasmodium falciparum and P. vivax.Genetics 2003; 163:1327–1336.
19. Mayer DC, Mu JB, Kaneko O, Duan J, Su XZ, Miller LH. Poly-morphism in the Plasmodium falciparum erythrocyte-binding ligandJESEBL/EBA-181 alters its receptor specificity. Proc Natl Acad SciUSA 2004; 101:2518–2523.
20. Dolan SA, Miller LH, Wellems TE. Evidence for a switching mechanismin the invasion of erythrocytes by Plasmodium falciparum. J Clin Invest1990; 86:618–624.
21. Baum J, Pinder M, Conway DJ. Erythrocyte invasion phenotypes ofPlasmodium falciparum in The Gambia. Infect Immun 2003; 71:1856–1863.
22. Bei AK, Membi CD, Rayner JC, et al. Variant merozoite protein ex-pression is associated with erythrocyte invasion phenotypes in Plasmo-dium falciparum isolates from Tanzania. Mol Biochem Parasitol 2007;153:66–71.
23. Deans AM, Nery S, Conway DJ, Kai O, Marsh K, Rowe JA. Invasionpathways and malaria severity in Kenyan Plasmodium falciparum clin-ical isolates. Infect Immun 2007; 75:3014–3020.
24. Ohas EA, Adams JH, Waitumbi JN, et al. Measurement of antibodylevels against region II of the erythrocyte-binding antigen 175 of Plas-modium falciparum in an area of malaria holoendemicity in westernKenya. Infect Immun 2004; 72:735–741.
25. Okenu DM, Riley EM, Bickle QD, et al. Analysis of human antibodiesto erythrocyte binding antigen 175 of Plasmodium falciparum. InfectImmun 2000; 68:5559–5566.
26. Osier FH, Fegan G, Polley SD, et al. Breadth and magnitude of antibodyresponses to multiple Plasmodium falciparum merozoite antigens areassociated with protection from clinical malaria. Infect Immun 2008;76:2240–2248.
27. John CC, Moormann AM, Pregibon DC, et al. Correlation of highlevels of antibodies to multiple pre-erythrocytic Plasmodium falciparumantigens and protection from infection. Am J Trop Med Hyg 2005; 73:222–228.
28. Stanisic DI, Richards JS, McCallum FJ, et al. Immunoglobulin G sub-class-specific responses against Plasmodium falciparum merozoite an-tigens are associated with control of parasitemia and protection fromsymptomatic illness. Infect Immun 2009; 77:1165–1174.
29. Roussilhon C, Oeuvray C, Muller-Graf C, et al. Long-term clinical
protection from falciparum malaria is strongly associated with IgG3antibodies to merozoite surface protein 3. PLoS Med 2007; 4:e320.
30. Nebie I, Diarra A, Ouedraogo A, et al. Humoral responses to Plas-modium falciparum blood-stage antigens and association with inci-dence of clinical malaria in children living in an area of seasonal ma-laria transmission in Burkina Faso, West Africa. Infect Immun 2008;76:759–766.
31. Courtin D, Oesterholt M, Huismans H, et al. The quantity and qualityof African children’s IgG responses to merozoite surface antigens reflectprotection against Plasmodium falciparum malaria. PLoS One 2009; 4:e7590.
32. Michon P, Cole-Tobian JL, Dabod E, et al. The risk of malarial infec-tions and disease in Papua New Guinean children. Am J Trop MedHyg 2007; 76:997–1008.
33. Thompson JK, Triglia T, Reed MB, Cowman AF. A novel ligand fromPlasmodium falciparum that binds to a sialic acid-containing receptoron the surface of human erythrocytes. Mol Microbiol 2001; 41:47–58.
34. Barry AE, Schultz L, Buckee CO, Reeder JC. Contrasting populationstructures of the genes encoding ten leading vaccine-candidate anti-gens of the human malaria parasite, Plasmodium falciparum. PLoS One2009; 4:e8497.
35. Ford L, Lobo CA, Rodriguez M, et al. Differential antibody responsesto Plasmodium falciparum invasion ligand proteins in individuals livingin malaria-endemic areas in Brazil and Cameroon. Am J Trop MedHyg 2007; 77:977–983.
36. Richards JS, Beeson JG. The future for blood-stage vaccines againstmalaria. Immunol Cell Biol 2009; 87:377–390.
37. Jones TR, Narum DL, Gozalo AS, et al. Protection of Aotus monkeysby Plasmodium falciparum EBA-175 region II DNA prime-proteinboost immunization regimen. J Infect Dis 2001; 183:303–312.
38. Sim BK, Narum DL, Liang H, et al. Induction of biologically activeantibodies in mice, rabbits, and monkeys by Plasmodium falciparumEBA-175 region II DNA vaccine. Mol Med 2001; 7:247–254.
39. Gilberger TW, Thompson JK, Reed MB, Good RT, Cowman AF. Thecytoplasmic domain of the Plasmodium falciparum ligand EBA-175 isessential for invasion but not protein trafficking. J Cell Biol 2003; 162:317–327.
40. Reed MB, Caruana SR, Batchelor AH, Thompson JK, Crabb BS, Cow-man AF. Targeted disruption of an erythrocyte binding antigen inPlasmodium falciparum is associated with a switch toward a sialic acid-independent pathway of invasion. Proc Natl Acad Sci USA 2000; 97:7509–7514.