apoptosis in mouse primordial germ cells: a study by transmission and scanning electron microscope
TRANSCRIPT

Anat Embryol (1994) 189:435M40
Anatomy and Embryology
�9 Springer-Verlag 1994
Apoptosis in mouse primordial germ cells: a study by transmission and scanning electron microscope Maurizio Pesce, Massimo De Feliei
Department of Public Health and Cell Biology, Section of Histology and Embryology, University of Rome "Tor Vergata', Via O. Raimondo 8, 1-00173 Rome, Italy
Accepted: 6 January 1994
Abstract. A detailed study of the death in vitro of mouse primordial germ cell (PGCs) by means of transmission and scanning electron microscopy is reported. The re- sults show that after 4-5 h of culture 1 5 ~ 0 % PGCs as- sume the typical morphological features of apoptotic cells, including chromatin condensation in dense masses under the nuclear membrane, compaction of the cyto- plasm, crowding of organelles and surface protuberances. Cells then break up into discrete fragments (apoptotic bodies) which eventually degenerate by "secondary ne- crosis". It is possible that apoptosis plays a biologically useful role in avoiding uncontrolled PGC proliferation and in eliminating misplaced germ cells whose survivial might be harmful to the animal.
Key words: Apoptosis - Primordial germ cells - Mouse embryo - Gametogenesis
Introduction
In normal embryonic development cell death occurs at specific times and sites, hence the name of programmed cell death, which may be considered a type of apoptotic (controlled) cell death. Apoptosis is characterized by specific morphological changes, including chromatin condensation, nuclear and cytoplasmic fragmentation and usually by D N A degradation into fragments with sizes of one or more nucleosomes (DNA ladder). This form of cell death is triggered by changes in the level of specific physiological stimuli (for reviews, see Arends and Wyllie 1991; Kerr and Harmon 1991).
We recently reported that apoptosis may occur in pri- mordial germ cells (PGCs) in the mouse embryo, and that PGCs obtained from mouse embryos at 11.5-12.5 days
Correspondence to: M. De Felici
post coitum (dpc) cultured in the absence of suitable feed- er layers and growth factors rapidly show apoptotic fea- tures in culture, such as nucleosomic D N A fragmenta- tion and high expression of tissue transglutaminase (Pesce et al. 1993). The present study provides a time course and a detailed morphological characterization of PGC apoptosis in culture by means of transmission and scanning electron microscopy.
Materials and methods
PGCs were collected, purified and identified from 12.5 dpc CD-1 mice fetal gonads according to the method of De Felici and McLaren (1983). Briefly, gonads were cleared from mesonephros and incubated in phosphate-buffered saline (PBS) containing 0.01% EDTA, at room temperature for 10 15 rain. Cells were released by pricking the tissues with fine needles to yield a cell suspension containing about 70-80% PGCs, as determined by their identifica- tion with alkaline phosphatase staining.
For transmission electron microscope (TEM) observations, PGCs were cultured for 4.6 h in suspension in 0.5 ml modified Eagle's MEM supplemented with 5% horse serum and 2.5% fetal calf serum (De Felici and Dolci 1991), in 1.5-ml Eppendorf tubes. After culture, the cells were centrifuged, washed in PBS and fixed for 1 h in 2% glutaraldehyde in 0.1 M sodium cacodylate buffer (pH 7.3). They were then washed three times with 0.1 M sodium cacodylate, post-fixed with a 1% osmium tetroxide aqueous solu- tion and pre-stained en bloc with a solution of 2% uranyl acetate in 30% ethanol. PGCs were dehydrated in ethanol, cleared in toluene and embedded in Epon 812. Sections were cut at 0.1 gm with a LKB Ultrotome V and attached onto 150-mesh copper grids pre-treated with Formvar. Grids were stained with lead hydroxide and ob- served under a $7000 Hitachi TEM.
For the scanning electron microscope (SEM), PGCs were cul- tured for 4-18 h on glass coverslips (diameter 11 ram) coated wth 50 gl of Matrigel (a reconstituted basal membrane from Collabora- tive Research to which 12.5-dpc PGCs are able to attach, our un- published observations). Cells were fixed for 1 h with 2% glu- taratdehyde in 0.1 M sodium cacodylate (pH 7.3) containing 2% sucrose. After several washes with 0.1 M sodium cacodylate con- taining 2% sucrose, PGCs were post fixed with 0.1% osmium te- traoxide in 0.2 M sodium cacodylate for 30 min, dehydrate in an ethanol series and dried by the critical point method. After gold sputteirng the samples were examined under a Hitachi $570 SEM.
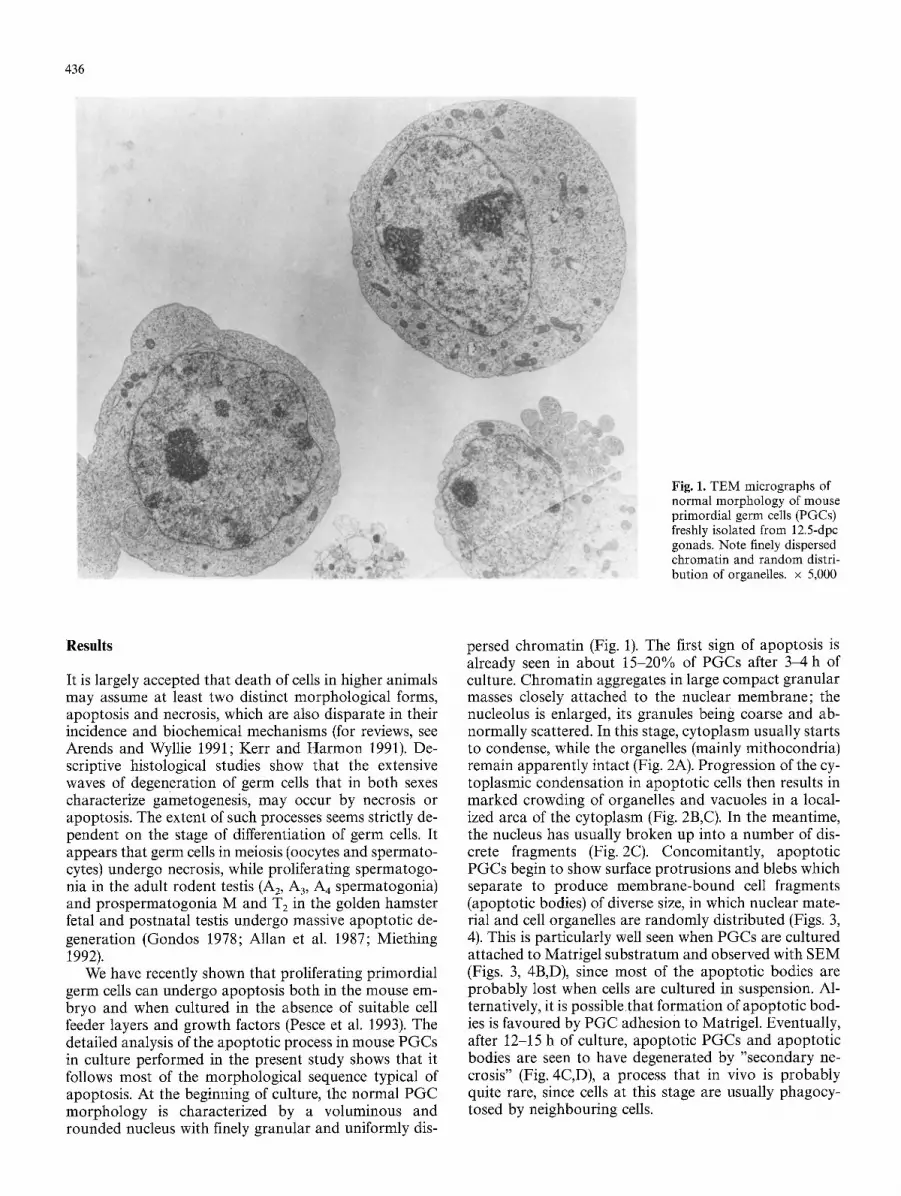
436
Fig. 1. TEM micrographs of normal morphology of mouse primordial germ cells (PGCs) freshly isolated from 12.5-dpc gonads. Note finely dispersed chromatin and random distri- bution of organelles, x 5,000
Results
It is largely accepted that death of cells in higher animals may assume at least two distinct morphological forms, apoptosis and necrosis, which are also disparate in their incidence and biochemical mechanisms (for reviews, see Arends and Wyllie 1991; Kerr and Harmon 1991). De- scriptive histological studies show that the extensive waves of degeneration of germ cells that in both sexes characterize gametogenesis, may occur by necrosis or apoptosis. The extent of such processes seems strictly de- pendent on the stage of differentiation of germ cells. It appears that germ cells in meiosis (oocytes and spermato- cytes) undergo necrosis, while proliferating spermatogo- nia in the adult rodent testis (A2, A 3, A4 spermatogonia) and prospermatogonia M and T 2 in the golden hamster fetal and postnatal testis undergo massive apoptotic de- generation (Gondos 1978; Allan et al. 1987; Miething 1992).
We have recently shown that proliferating primordial germ cells can undergo apoptosis both in the mouse em- bryo and when cultured in the absence of suitable cell feeder layers and growth factors (Pesce et al. 1993). The detailed analysis of the apoptotic process in mouse PGCs in culture performed in the present study shows that it follows most of the morphological sequence typical of apoptosis. At the beginning of culture, the normal PGC morphology is characterized by a voluminous and rounded nucleus with finely granular and uniformly dis-
persed chromatin (Fig. 1). The first sign of apoptosis is already seen in about 15-20% of PGCs after 3-4 h of culture. Chromatin aggregates in large compact granular masses closely attached to the nuclear membrane; the nucleolus is enlarged, its granules being coarse and ab- normally scattered. In this stage, cytoplasm usually starts to condense, while the organelles (mainly mithocondria) remain apparently intact (Fig. 2A). Progression of the cy- toplasmic condensation in apoptotic cells then results in marked crowding of organelles and vacuoles in a local- ized area of the cytoplasm (Fig. 2B,C). In the meantime, the nucleus has usually broken up into a number of dis- crete fragments (Fig. 2C). Concomitantly, apoptotic PGCs begin to show surface protrusions and blebs which separate to produce membrane-bound cell fragments (apoptotic bodies) of diverse size, in which nuclear mate- rial and cell organelles are randomly distributed (Figs. 3, 4). This is particularly well seen when PGCs are cultured attached to Matrigel substratum and observed with SEM (Figs. 3, 4B,D), since most of the apoptotic bodies are probably lost when cells are cultured in suspension. Al- ternatively, it is possible that formation of apoptotic bod- ies is favoured by PGC adhesion to Matrigel. Eventually, after 12-15 h of culture, apoptotic PGCs and apoptotic bodies are seen to have degenerated by "secondary ne- crosis" (Fig. 4C,D), a process that in vivo is probably quite rare, since cells at this stage are usually phagocy- tosed by neighbouring cells.
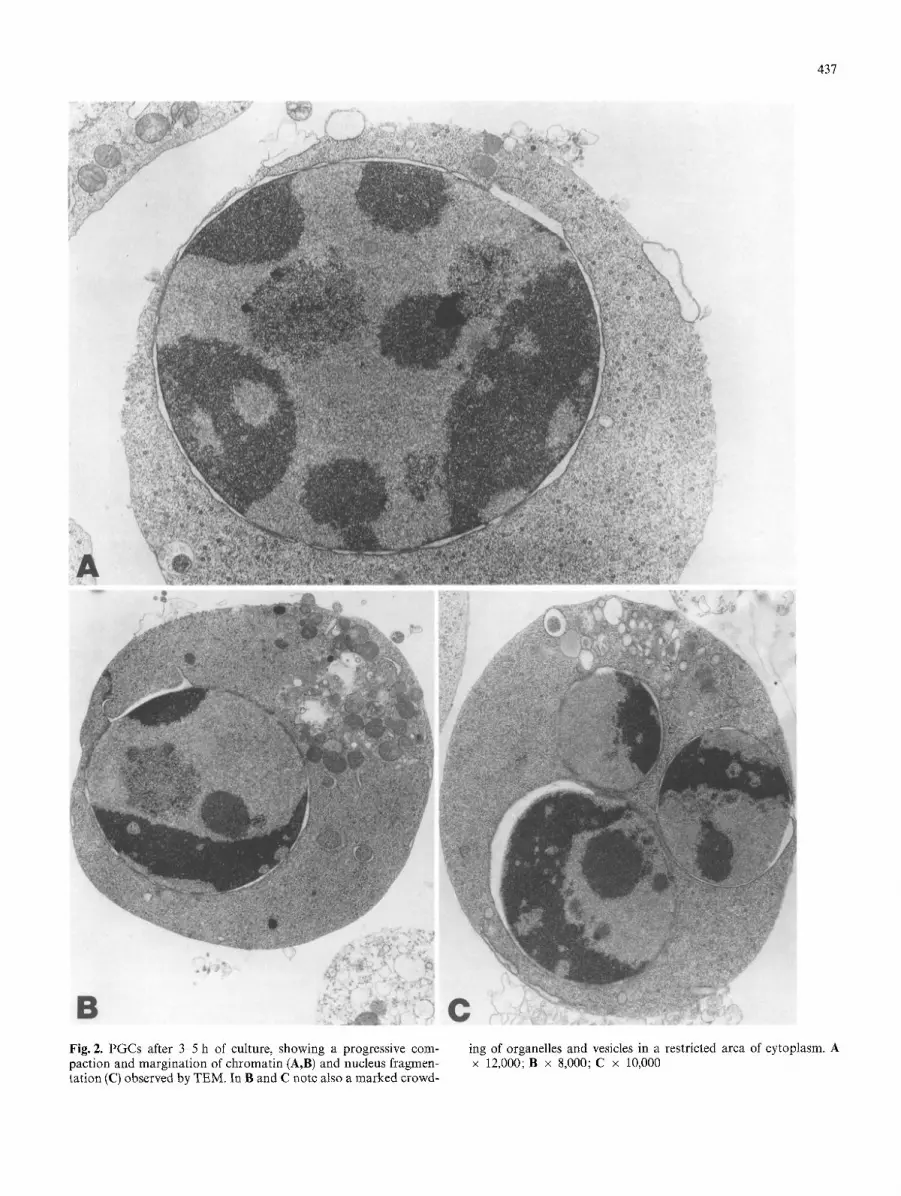
437
Fig. 2. PGCs after 3 5 h of cuIture, showing a progressive com- paction and margination of chromatin (A,B) and nucleus fragmen- tation (C) observed by TEM. In B and C note also a marked crowd-
ing of organelles and vesicles in a restricted area of cytoplasm. A x 12,000; B x 8,000; C x 10,000
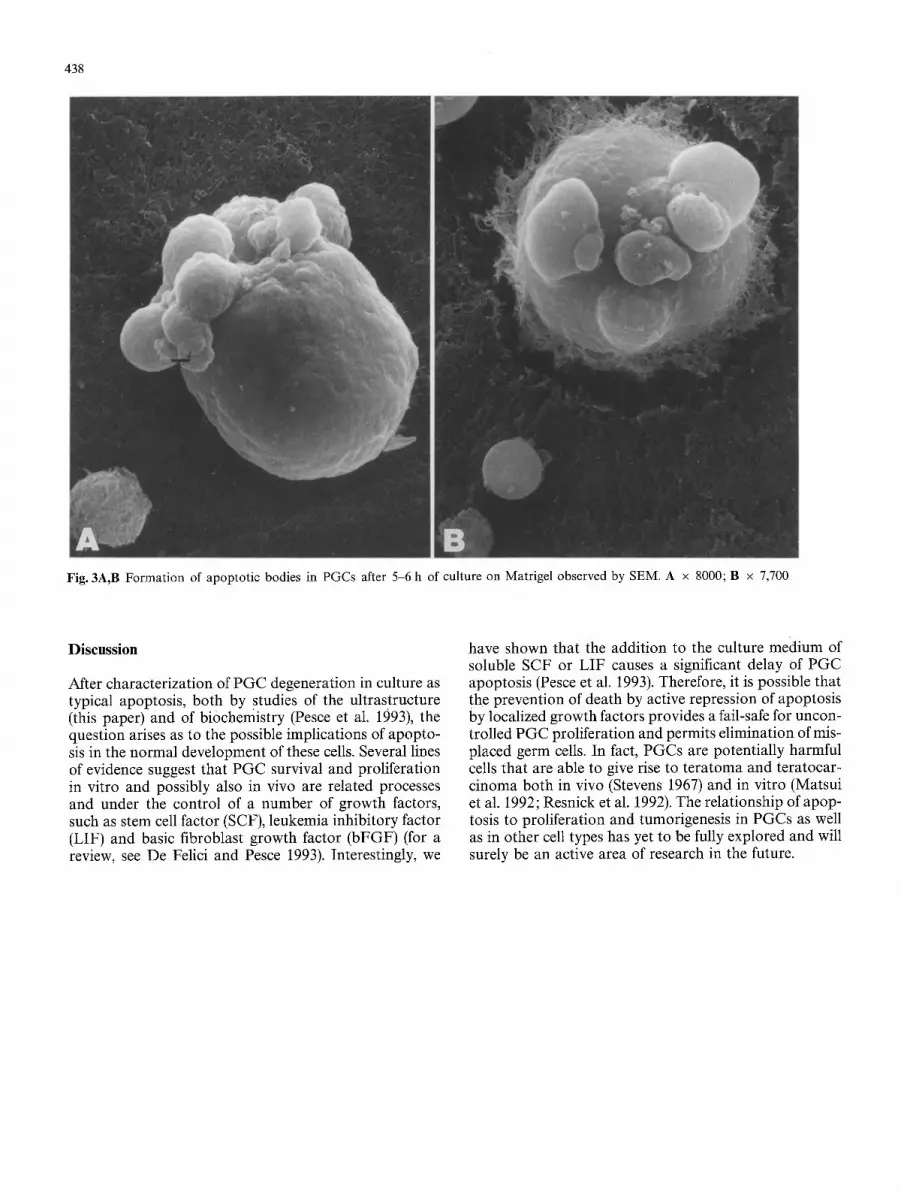
438
Fig. 3A,B Formation of apoptotic bodies in PGCs after 5-6 h of culture on Matrigel observed by SEM. A x 8000; B x 7,700
Discussion
After characterization of PGC degeneration in culture as typical apoptosis, both by studies of the ultrastructure (this paper) and of biochemistry (Pesce et al. 1993), the question arises as to the possible implications of apopto- sis in the normal development of these cells. Several lines of evidence suggest that PGC survival and proliferation in vitro and possibly also in vivo are related processes and under the control of a number of growth factors, such as stem cell factor (SCF), leukemia inhibitory factor (LIF) and basic fibroblast growth factor (bFGF) (for a review, see De Felici and Pesce 1993). Interestingly, we
have shown that the addition to the culture medium of soluble SCF or LIF causes a significant delay Of PGC apoptosis (Pesce et al. 1993). Therefore, it is possible that the prevention of death by active repression of apoptosis by localized growth factors provides a fail-safe for uncon- trolled PGC proliferation and permits elimination of mis- placed germ cells. In fact, PGCs are potentially harmful cells that are able to give rise to teratoma and teratocar- cinoma both in vivo (Stevens 1967) and in vitro (Matsui et al. 1992; Resnick et al. 1992). The relationship of apop- tosis to proliferation and tumorigenesis in PGCs as well as in other cell types has yet to be fully explored and will surely be an active area of research in the future.
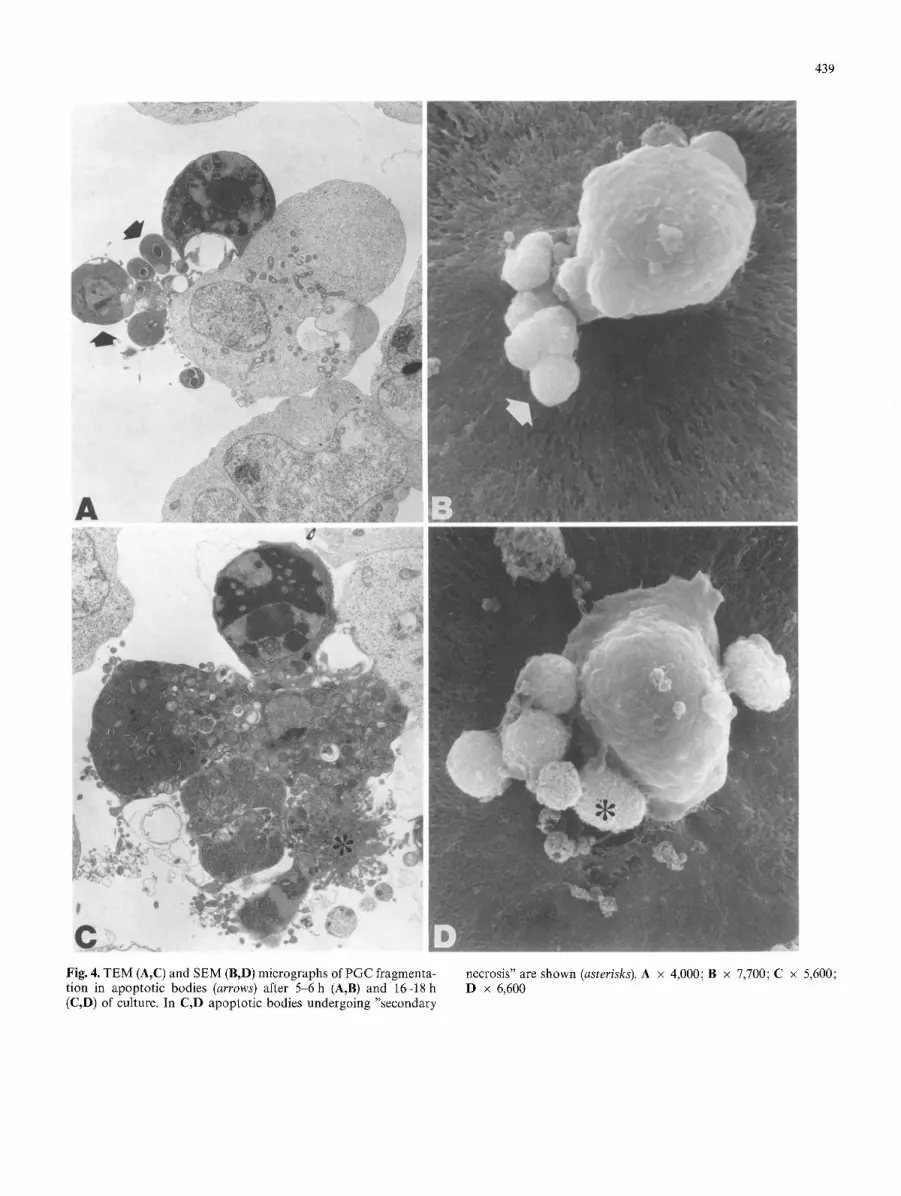
439
Fig. 4. TEM (A,C) and SEM (B,D) micrographs of PGC fragmenta- tion in apoptotic bodies (arrows) after 5 6 h (A,B) and 16-18 h (C,D) of culture. In C,D apoptotic bodies undergoing "secondary
necrosis" are shown (asterisks). A x 4,000; B • 7,700; C • 5,600; D x 6,600

440
Acknowledgements. We wish to thank Prof. Mario Stefanini for the use of transmission and scanning electron microscopes. We also thank Mr. Quinto Giustiniani for skilful technical assistance and Mr. Graziano Bonelli for excellent photographic services. Financial support from M.U.R.S.T. (40% and 60%) and C.N.R. (Progetto Finalizzato FAT.MA, grant no. 93.00738) is gratefully acknowl- edged.
References
Allan D J, Harmon BV, Kerr JFR (1987) Cell death in spermatogen- esis. In: Potten CS (ed) Perspectives on mammalian cell death. Oxford University Press, Oxford, pp 229-258
Arends MJA, Wytlie CC (1991) Apoptosis mechanism and role in pathology. Int Rev Exp Pathol 32:223-254
De Felici M, Dolci S (1991) Leukemia inhibitory factor sustains the survival of mouse primordial germ cells cultured on TM 4 feeder layers. Dev Biol 147:281-284
De Felici M, McLaren A (1983) In vitro culture of mouse primor- dial germ cells. Exp Cell Res 144:417-427
De Felici M, Pesce M (1993) Growth factors in mouse primordial germ cell migration and proliferation. Prog Growth Factor Res (in press)
Gondos B (1978) Oogonia and oocytes in mammals. In: Jones RE (ed) The vertebrate ovary. Comparative biology and evolution. Plenum Press, New York, pp 83-120
Kerr JFR, Harmon BV (1991) Definition and incidence of apopto- sis: an historical perspective. In: Tomei LD, Coppe FO (eds) Apoptosis: the molecular basis of cell death. Cold Spring Har- bor Laboratory Press, Cold Spring Harbor, New York, pp 5-29
Matsui Y, Zsebo K, Hogan B (1992) Derivation of pluripotential embryonic stem cells from murine primordial germ cells in cul- ture. Cell 70:841-847
Miething A (1992) Germ-cell death during prespermatogenesis in the testis of the golden hamster. Cell Tisuse Res 267:583-590
Pesce M, Farrace MG, Dolci S, Piacentini M, De Felici M (1993) Stem cell factor and leukemia inhibitory factor promote primor- dial germ cell survival by suppressing programmed cell death (apoptosis). Development 118 : 1089-1094
Resnick JL, Bixler LS, Cheng L, Donovan PJ (1992) Long-term proliferation of mouse primordial germ cells in culture. Nature 359:550-551
Stevens LC (1967) Origin of testicular teratomas from primordial germ cells in mice. J Natl Cancer Inst 37:859-861
Approaches for identifying germ cell mutagens: Report of the 2013 IWGT workshop on germ cell assays☆
Zebrafish Staufen1 and Staufen2 are required for the survival and migration of primordial germ cells