anion- and proton-dependent gating of clc-4 anion/proton transporter under uncoupling conditions
TRANSCRIPT

Biophysical Journal Volume 100 March 2011 1233–1241 1233
Anion- and Proton-Dependent Gating of ClC-4 Anion/Proton Transporterunder Uncoupling Conditions
Gokce Orhan,† Christoph Fahlke,†‡ and Alexi K. Alekov†*†Institut fur Neurophysiologie, Medizinische Hochschule, Hannover, Germany; and ‡Zentrum fur Systemische NeurowissenschaftenHannover, Hannover, Germany
ABSTRACT ClC-4 is a secondary active transporter that exchanges Cl� ions and Hþ with a 2:1 stoichiometry. In externalSCN�, ClC-4 becomes uncoupled and transports anions with high unitary transport rate. Upon voltage steps, the number ofactive transporters varies in a time-dependent manner, resembling voltage-dependent gating of ion channels. We here inves-tigated modification of the voltage dependence of uncoupled ClC-4 by protons and anions to quantify association of substrateswith the transporter. External acidification shifts voltage dependence of ClC-4 transport to more positive potentials and leads toreduced transport currents. Internal pH changes had less pronounced effects. Uncoupled ClC-4 transport is facilitated byelevated external [SCN�] but impaired by internal Cl� and I�. Block by internal anions indicates the existence of an internalanion-binding site with high affinity that is not present in ClC channels. The voltage dependence of ClC-4 coupled transportis modulated by external protons and internal Cl� in a manner similar to what is observed under uncoupling conditions. Ourdata illustrate functional differences but also similarities between ClC channels and transporters.
INTRODUCTION
ClC channels and transporters are present in almost allprokaryotic and eukaryotic cells and fulfill a variety ofphysiological functions. Four human isoforms, ClC-1,ClC-2, ClC-Ka, and ClC-Kb, represent anion channels. Theremaining ones (ClC-3–ClC-7) function under physiologicalconditions as anion-proton exchangers (1). Channel-medi-ated transmembrane anion flux regulates cellular excitability(2–4) andmediates epithelial salt transport (5–9). In contrast,ClC transporters contribute to pH regulation in variousintracellular compartments (10–13).
ClC transporters can also mediate uncoupled aniontransport in polyatomic anions or decreased externalpH (14–16). Uncoupled ClC anion transport has beendescribed thermodynamically as slippage mode, facilitateddiffusion with very high transport rate, or as channelconductance. However, the underlying mechanisms ofuncoupling still remain elusive. For ClC-4, we describeda sequence of increased uncoupling potency of externalanions in the order I� < Br� < NO3
� <SCN�. Thissequence correlates with the apparent external bindingaffinity of the transport substrates to the transporter (14).Similar data was reported from calorimetric studies onthe bacterial ClC transporter homolog ClC-ec1 showingweaker binding for uncoupling anions when compared toCl� (17).
Voltage-dependent gating in ClC channels has beenintensively studied. Its physiological importance is illus-
Submitted June 9, 2010, and accepted for publication January 24, 2011.
*Correspondence: [email protected]
G. Orhan’s present address is Abt. Neurologie mit Schwerpunkt Epilepto-
logie, Hertie Institut fur Klinische Hirnforschung, Universitatsklinikum
Tubingen, Hoppe-Seyler-Str. 3, D-72076 Tubingen, Germany.
Editor: Tzyh-Chang Hwang.
� 2011 by the Biophysical Society
0006-3495/11/03/1233/9 $2.00
trated by several disease-causing mutations (4,18–20).ClC channel gating displays various unique features.There are two structurally distinct gating processes thatreflect the double-barreled architecture of these channels,protopore gating and common gating (21–24). In addition,a substantial number of channels remain open even atextreme voltages (25–27) deviating from standard Boltz-mann behavior. Lastly, ClC channel gating is intimatelycoupled to the concentration of permeant anions and pH(25,26,28–31). Gating of ClC channels developed prob-ably from conformational changes in a Cl�/Hþ antiportcycle (31,32). In particular, they seem to function asbroken transporters in which one of the conformationalstates is characterized with high conductance for anions(31,33).
ClC transporters, such as ClC-3, ClC-4, and ClC-5,exhibit pronounced time- and voltage-dependent alterationof current amplitudes (16,34–37). Noise analysis on ClC-4under uncoupling conditions (14) and on coupled ClC-5transport (16) revealed that time-dependent changes in thecurrent amplitudes upon voltage steps are due to alterationsin the number of active transporters, resembling the gatingprocesses observed in ion channels.
We here study the effects of anions and protons onvoltage-dependent gating of ClC-4 under uncouplingconditions to quantify interactions of the transport sub-strates with the transporter protein. This approach hasbeen taken for various neurotransmitter transporters (38–40)as well as for ion channels evolved from transporters (32).Moreover, it allows direct comparison with mammalianClC channels for which the modification of channel gatingby anions and protons has been extensively studied.
doi: 10.1016/j.bpj.2011.01.045

1234 Orhan et al.
METHODS
Cell culture
HEK293 cell lines, stably expressing hClC-4 (36), were cultured in stan-
dard modified Eagle’s medium supplemented with 10% fetal bovine serum
and 900 mg/mL Geneticin (G418; Invitrogen, Karlsruhe, Germany). Two
different oligoclonal cell lines with indistinguishable functional character-
istics were used.
Mammalian cells often express endogenous anion channels. However,
we recently demonstrated that endogenous HEK293 currents do not inter-
fere with the characterization of ClC-4 gating in stably transfected cells,
as their amplitudes are much smaller than ClC-4-mediated currents (14).
Electrophysiology
Standard whole-cell patch-clamp (41) was performed using an Axopatch
200B (Axon Instruments, Union City, CA) and HEKA EPC-10 (HEKA,
Lambrecht, Germany) amplifiers. Borosilicate pipettes were pulled with
resistances of 1–2 MU. Capacitive artifacts and series resistance errors
were reduced by capacitance cancellation and series resistance compensa-
tion. The calculated voltage error due to series resistance was <5 mV.
Currents were digitized with 100 kHz sampling rate after 5 kHz analog
filtering. The composition of the standard solutions was 1), extracellular
145 mM NaSCN, 4 mM KSCN, 2 mM CaCl2, 1 mM Mg(Gluconate)2,
and 5 mM HEPES, pH 7.4; and 2), intracellular 95 mM NaI, 2.5 mM
MgCl2, 2.5 mM Na2ATP, 5 mM EGTA, and 10 mM HEPES, pH 7.4. For
variation of pH, HEPES was replaced on equimolar basis with MES
(2-(n-morpholino)ethanesulphonic acid) or AMPSO (n-(1,1-Dimethyl-2-
hydroxyethyl)-3-amino-2-hydroxy-propanesulfonic acid). For experiments
with different anion concentrations, intracellular I� or extracellular
SCN� were substituted on an equimolar basis with Cl� or Gluconate�. Inall experiments, we used external and internal agar salt bridges made
from plastic tubing filled with 3 M KCl in 0.8% agar to connect the
Ag/AgCl electrodes to the bath/pipette solution. Junction potentials were
calculated using the JPCalc module in Clampex (Axon Instruments) (42)
and corrected.
Data analysis
Data were analyzed with Clampfit 8 (Axon Instruments), FitMaster
(HEKA), Origin 6 (OriginLab, Northampton, MA), and EXCEL (Micro-
soft, Redmond, WA). Time constants were obtained from exponential fits
of the ClC-4 current time course. Instantaneous current amplitudes were
resolved by extrapolating monoexponential fits of the initial activation
time course to the voltage step and subsequently normalized to one. Voltage
dependence of activation was described as a Boltzmann function
Popen ¼ PSS þ 1� PSS
1þ eðV�V0:5Þ=k;
with PSS being the minimum open probability and V0.5 the midpoint of acti-
vation. The slope factor (k) is equivalent to RT=zF, with z denoting the
apparent charge moved during the voltage-dependent transition, and R, T,
and F representing the universal gas constant, absolute temperature, and
the Faraday constant, respectively. The voltage dependence of the rate
constants was assumed to follow the Eyring law:
a ¼ a0ecV :
Assuming a simple C 4 O gating model with forward and backward
rate constants a and b, the voltage dependence of the time constants (t)
was fitted with the inverted sum of two exponential functions with
opposite argument sign ðt ¼ 1=ðaþ bÞÞ: All summary data are shown as
mean 5 SE.
Biophysical Journal 100(5) 1233–1241
RESULTS
Voltage-dependent gating of uncoupled hClC-4
In native cells, ClC-4 is located mainly in intracellularcompartments. However, heterologous expression inmammalian cells results also in surface insertion and permitswhole-cell ClC-4 current recordings (14,36). External SCN�
results in a significant number of ClC-4 proteins operating inan uncoupled slippage mode with unitary transport ratesmuch larger compared to those remaining in the coupledtransporter mode. Therefore, anion currents are dominatedby uncoupled transport which allows characterization ofthis particular functional mode of ClC-4 without significantcontamination due to coupled ion transport. Internal iodideslows down ClC-4 kinetics and therefore greatly improvesthe temporal resolution of the measurements (14,36).
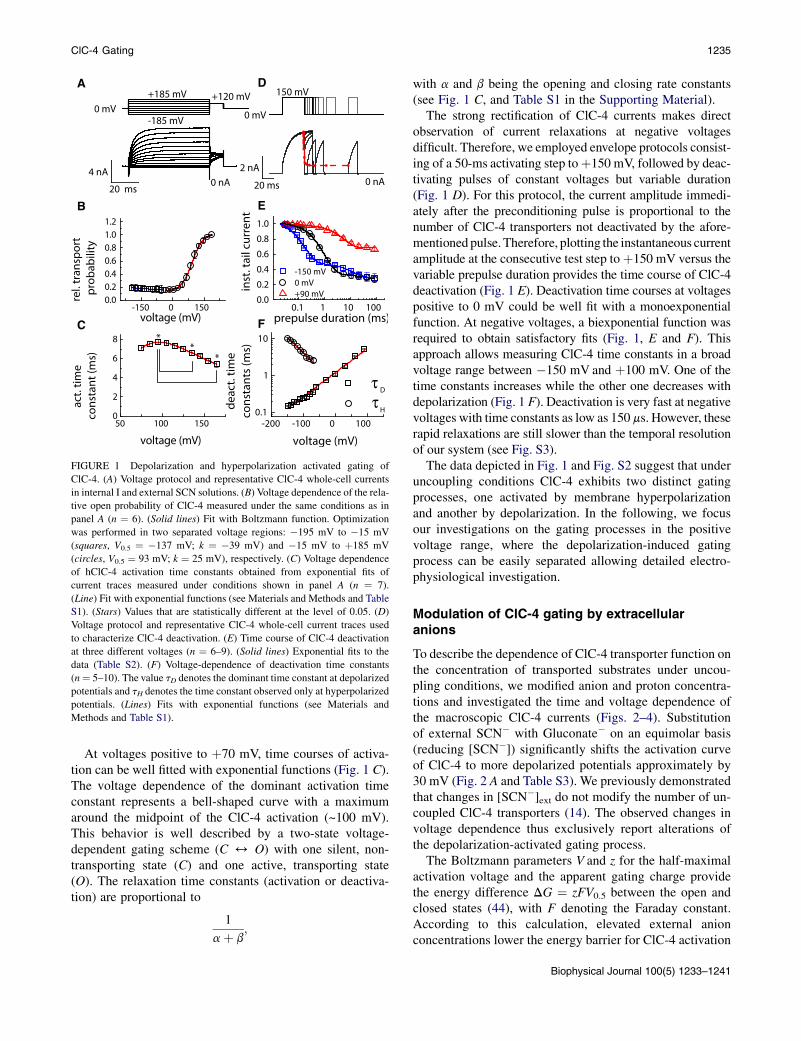
Fig. 1 shows the time- and voltage dependence of anioncurrents from mammalian cells stably expressing humanClC-4 measured in external SCN� and internal I�. ClC-4currents are small at negative voltages but exhibit an instan-taneous rise followedby a slower activation at potentials posi-tive to þ40 mV (Fig. 1 A). We recently demonstrated thatunder these conditions current activation is due to changesin the number of active uncoupled transporters (14). To deter-mine the number of active transporters at a given voltage weemployed a procedure that was originally developed to quan-tify steady-state open probabilities of ion channels. We usedtail current protocols consisting of conditioning prepulses toreach steady state at the voltage of interest, followed bya constant test pulse to þ120 mV (Fig. 1 A). Tail currentanalysis might be complicated by the existence of capacitivecurrents that were recently reported for ClC-5 (43).We testedwhether such currents exist also for ClC-4 but could notobserve any under our experimental conditions (see Fig. S1in the Supporting Material).
Because the unitary transport rate at the test pulse and thenumber of transporters per cell are not affected by the pre-pulse potential, plotting the instantaneous tail current ampli-tude at the test step versus the preceding potential providesthe relative number of active transporters (Fig. 1 B). Hence-forth, we will use the term ‘‘transport probability’’ for theprobability of a transporter to reside in an active transport-ing state and the term ‘‘activation’’ for the transition froman inactive into an active transporting state.
The voltage-dependent transport probability of the depo-larization-activated gating process of ClC-4 can be fittedwith a Boltzmann curve with nonzero open probability.We also observed a slight but distinct increase in the tailcurrent amplitudes at negative voltages which demonstratesthe existence of a second independent process, activatedwith hyperpolarization. This process is uncovered and betterobserved when depolarizing prepulses are applied to recruitadditional transporters from the silent nontransporting state(due to the depolarization-activated gating process; seeFig. S2).
B
C
A D
E
F
ττ
FIGURE 1 Depolarization and hyperpolarization activated gating of
ClC-4. (A) Voltage protocol and representative ClC-4 whole-cell currents
in internal I and external SCN solutions. (B) Voltage dependence of the rela-
tive open probability of ClC-4 measured under the same conditions as in
panel A (n ¼ 6). (Solid lines) Fit with Boltzmann function. Optimization
was performed in two separated voltage regions: �195 mV to �15 mV
(squares, V0.5 ¼ �137 mV; k ¼ �39 mV) and �15 mV to þ185 mV
(circles, V0.5 ¼ 93 mV; k ¼ 25 mV), respectively. (C) Voltage dependence
of hClC-4 activation time constants obtained from exponential fits of
current traces measured under conditions shown in panel A (n ¼ 7).
(Line) Fit with exponential functions (see Materials and Methods and Table
S1). (Stars) Values that are statistically different at the level of 0.05. (D)
Voltage protocol and representative ClC-4 whole-cell current traces used
to characterize ClC-4 deactivation. (E) Time course of ClC-4 deactivation
at three different voltages (n ¼ 6–9). (Solid lines) Exponential fits to the
data (Table S2). (F) Voltage-dependence of deactivation time constants
(n¼ 5–10). The value tD denotes the dominant time constant at depolarized
potentials and tH denotes the time constant observed only at hyperpolarized
potentials. (Lines) Fits with exponential functions (see Materials and
Methods and Table S1).
ClC-4 Gating 1235
At voltages positive to þ70 mV, time courses of activa-tion can be well fitted with exponential functions (Fig. 1 C).The voltage dependence of the dominant activation timeconstant represents a bell-shaped curve with a maximumaround the midpoint of the ClC-4 activation (~100 mV).This behavior is well described by a two-state voltage-dependent gating scheme (C 4 O) with one silent, non-transporting state (C) and one active, transporting state(O). The relaxation time constants (activation or deactiva-tion) are proportional to
1
aþ b;
with a and b being the opening and closing rate constants(see Fig. 1 C, and Table S1 in the Supporting Material).
The strong rectification of ClC-4 currents makes directobservation of current relaxations at negative voltagesdifficult. Therefore, we employed envelope protocols consist-ing of a 50-ms activating step toþ150 mV, followed by deac-tivating pulses of constant voltages but variable duration(Fig. 1 D). For this protocol, the current amplitude immedi-ately after the preconditioning pulse is proportional to thenumber of ClC-4 transporters not deactivated by the afore-mentioned pulse.Therefore, plotting the instantaneous currentamplitude at the consecutive test step toþ150 mV versus thevariable prepulse duration provides the time course of ClC-4deactivation (Fig. 1 E). Deactivation time courses at voltagespositive to 0 mV could be well fit with a monoexponentialfunction. At negative voltages, a biexponential function wasrequired to obtain satisfactory fits (Fig. 1, E and F). Thisapproach allows measuring ClC-4 time constants in a broadvoltage range between �150 mV and þ100 mV. One of thetime constants increases while the other one decreases withdepolarization (Fig. 1 F). Deactivation is very fast at negativevoltages with time constants as low as 150 ms. However, theserapid relaxations are still slower than the temporal resolutionof our system (see Fig. S3).
The data depicted in Fig. 1 and Fig. S2 suggest that underuncoupling conditions ClC-4 exhibits two distinct gatingprocesses, one activated by membrane hyperpolarizationand another by depolarization. In the following, we focusour investigations on the gating processes in the positivevoltage range, where the depolarization-induced gatingprocess can be easily separated allowing detailed electro-physiological investigation.
Modulation of ClC-4 gating by extracellularanions
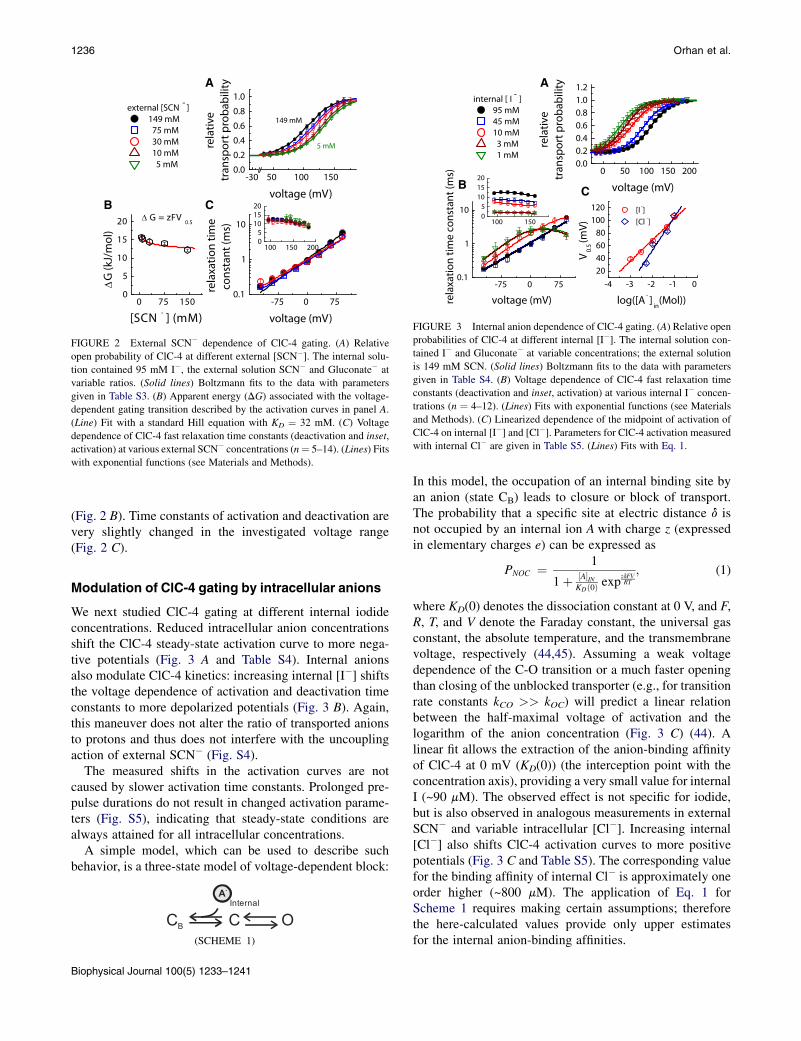
To describe the dependence of ClC-4 transporter function onthe concentration of transported substrates under uncou-pling conditions, we modified anion and proton concentra-tions and investigated the time and voltage dependence ofthe macroscopic ClC-4 currents (Figs. 2–4). Substitutionof external SCN� with Gluconate� on an equimolar basis(reducing [SCN�]) significantly shifts the activation curveof ClC-4 to more depolarized potentials approximately by30 mV (Fig. 2 A and Table S3). We previously demonstratedthat changes in [SCN�]ext do not modify the number of un-coupled ClC-4 transporters (14). The observed changes involtage dependence thus exclusively report alterations ofthe depolarization-activated gating process.
The Boltzmann parameters V and z for the half-maximalactivation voltage and the apparent gating charge providethe energy difference DG ¼ zFV0.5 between the open andclosed states (44), with F denoting the Faraday constant.According to this calculation, elevated external anionconcentrations lower the energy barrier for ClC-4 activation
Biophysical Journal 100(5) 1233–1241
A
B C
FIGURE 3 Internal anion dependence of ClC-4 gating. (A) Relative open
probabilities of ClC-4 at different internal [I�]. The internal solution con-
tained I� and Gluconate� at variable concentrations; the external solution
is 149 mM SCN. (Solid lines) Boltzmann fits to the data with parameters
given in Table S4. (B) Voltage dependence of ClC-4 fast relaxation time
constants (deactivation and inset, activation) at various internal I� concen-
trations (n ¼ 4–12). (Lines) Fits with exponential functions (see Materials
and Methods). (C) Linearized dependence of the midpoint of activation of
ClC-4 on internal [I�] and [Cl�]. Parameters for ClC-4 activation measured
with internal Cl� are given in Table S5. (Lines) Fits with Eq. 1.
A
B C
FIGURE 2 External SCN� dependence of ClC-4 gating. (A) Relative
open probability of ClC-4 at different external [SCN�]. The internal solu-tion contained 95 mM I�, the external solution SCN� and Gluconate� at
variable ratios. (Solid lines) Boltzmann fits to the data with parameters
given in Table S3. (B) Apparent energy (DG) associated with the voltage-
dependent gating transition described by the activation curves in panel A.
(Line) Fit with a standard Hill equation with KD ¼ 32 mM. (C) Voltage
dependence of ClC-4 fast relaxation time constants (deactivation and inset,
activation) at various external SCN� concentrations (n¼ 5–14). (Lines) Fits
with exponential functions (see Materials and Methods).
1236 Orhan et al.
(Fig. 2 B). Time constants of activation and deactivation arevery slightly changed in the investigated voltage range(Fig. 2 C).
Modulation of ClC-4 gating by intracellular anions
We next studied ClC-4 gating at different internal iodideconcentrations. Reduced intracellular anion concentrationsshift the ClC-4 steady-state activation curve to more nega-tive potentials (Fig. 3 A and Table S4). Internal anionsalso modulate ClC-4 kinetics: increasing internal [I�] shiftsthe voltage dependence of activation and deactivation timeconstants to more depolarized potentials (Fig. 3 B). Again,this maneuver does not alter the ratio of transported anionsto protons and thus does not interfere with the uncouplingaction of external SCN� (Fig. S4).
The measured shifts in the activation curves are notcaused by slower activation time constants. Prolonged pre-pulse durations do not result in changed activation parame-ters (Fig. S5), indicating that steady-state conditions arealways attained for all intracellular concentrations.
A simple model, which can be used to describe suchbehavior, is a three-state model of voltage-dependent block:
C O
A-
Internal
CB
(SCHEME 1)
Biophysical Journal 100(5) 1233–1241
In this model, the occupation of an internal binding site byan anion (state CB) leads to closure or block of transport.The probability that a specific site at electric distance d isnot occupied by an internal ion A with charge z (expressedin elementary charges e) can be expressed as
PNOC ¼ 1
1þ ½A�INKDð0Þ exp
zdFVRT
; (1)
where KD(0) denotes the dissociation constant at 0 V, and F,
R, T, and V denote the Faraday constant, the universal gasconstant, the absolute temperature, and the transmembranevoltage, respectively (44,45). Assuming a weak voltagedependence of the C-O transition or a much faster openingthan closing of the unblocked transporter (e.g., for transitionrate constants kCO >> kOC) will predict a linear relationbetween the half-maximal voltage of activation and thelogarithm of the anion concentration (Fig. 3 C) (44). Alinear fit allows the extraction of the anion-binding affinityof ClC-4 at 0 mV (KD(0)) (the interception point with theconcentration axis), providing a very small value for internalI (~90 mM). The observed effect is not specific for iodide,but is also observed in analogous measurements in externalSCN� and variable intracellular [Cl�]. Increasing internal[Cl�] also shifts ClC-4 activation curves to more positivepotentials (Fig. 3 C and Table S5). The corresponding valuefor the binding affinity of internal Cl� is approximately oneorder higher (~800 mM). The application of Eq. 1 forScheme 1 requires making certain assumptions; thereforethe here-calculated values provide only upper estimatesfor the internal anion-binding affinities.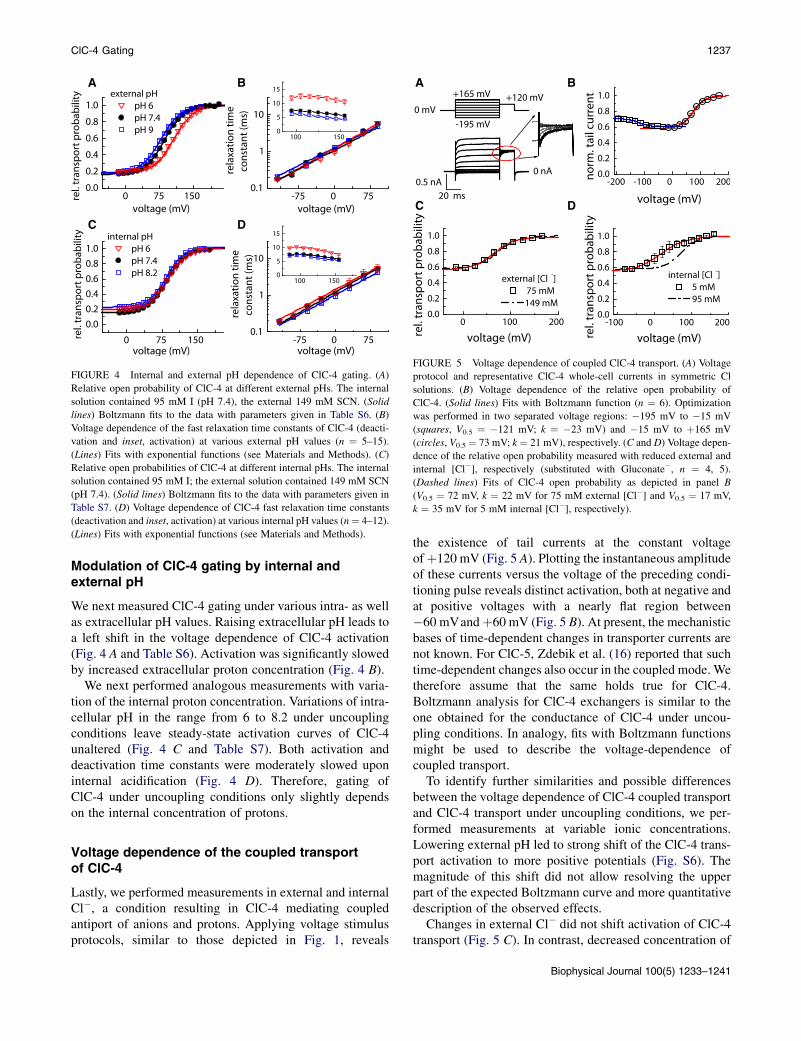
A B
C D
FIGURE 5 Voltage dependence of coupled ClC-4 transport. (A) Voltage
protocol and representative ClC-4 whole-cell currents in symmetric Cl
solutions. (B) Voltage dependence of the relative open probability of
ClC-4. (Solid lines) Fits with Boltzmann function (n ¼ 6). Optimization
was performed in two separated voltage regions: �195 mV to �15 mV
(squares, V0.5 ¼ �121 mV; k ¼ �23 mV) and �15 mV to þ165 mV
(circles, V0.5 ¼ 73 mV; k¼ 21 mV), respectively. (C and D) Voltage depen-
dence of the relative open probability measured with reduced external and
internal [Cl�], respectively (substituted with Gluconate�, n ¼ 4, 5).
(Dashed lines) Fits of ClC-4 open probability as depicted in panel B
(V0.5 ¼ 72 mV, k ¼ 22 mV for 75 mM external [Cl�] and V0.5 ¼ 17 mV,
k ¼ 35 mV for 5 mM internal [Cl�], respectively).
A B
C D
FIGURE 4 Internal and external pH dependence of ClC-4 gating. (A)
Relative open probability of ClC-4 at different external pHs. The internal
solution contained 95 mM I (pH 7.4), the external 149 mM SCN. (Solid
lines) Boltzmann fits to the data with parameters given in Table S6. (B)
Voltage dependence of the fast relaxation time constants of ClC-4 (deacti-
vation and inset, activation) at various external pH values (n ¼ 5–15).
(Lines) Fits with exponential functions (see Materials and Methods). (C)
Relative open probabilities of ClC-4 at different internal pHs. The internal
solution contained 95 mM I; the external solution contained 149 mM SCN
(pH 7.4). (Solid lines) Boltzmann fits to the data with parameters given in
Table S7. (D) Voltage dependence of ClC-4 fast relaxation time constants
(deactivation and inset, activation) at various internal pH values (n¼ 4–12).
(Lines) Fits with exponential functions (see Materials and Methods).
ClC-4 Gating 1237
Modulation of ClC-4 gating by internal andexternal pH
We next measured ClC-4 gating under various intra- as wellas extracellular pH values. Raising extracellular pH leads toa left shift in the voltage dependence of ClC-4 activation(Fig. 4 A and Table S6). Activation was significantly slowedby increased extracellular proton concentration (Fig. 4 B).
We next performed analogous measurements with varia-tion of the internal proton concentration. Variations of intra-cellular pH in the range from 6 to 8.2 under uncouplingconditions leave steady-state activation curves of ClC-4unaltered (Fig. 4 C and Table S7). Both activation anddeactivation time constants were moderately slowed uponinternal acidification (Fig. 4 D). Therefore, gating ofClC-4 under uncoupling conditions only slightly dependson the internal concentration of protons.
Voltage dependence of the coupled transportof ClC-4
Lastly, we performed measurements in external and internalCl�, a condition resulting in ClC-4 mediating coupledantiport of anions and protons. Applying voltage stimulusprotocols, similar to those depicted in Fig. 1, reveals
the existence of tail currents at the constant voltageofþ120 mV (Fig. 5 A). Plotting the instantaneous amplitudeof these currents versus the voltage of the preceding condi-tioning pulse reveals distinct activation, both at negative andat positive voltages with a nearly flat region between�60 mVandþ60 mV (Fig. 5 B). At present, the mechanisticbases of time-dependent changes in transporter currents arenot known. For ClC-5, Zdebik et al. (16) reported that suchtime-dependent changes also occur in the coupled mode. Wetherefore assume that the same holds true for ClC-4.Boltzmann analysis for ClC-4 exchangers is similar to theone obtained for the conductance of ClC-4 under uncou-pling conditions. In analogy, fits with Boltzmann functionsmight be used to describe the voltage-dependence ofcoupled transport.
To identify further similarities and possible differencesbetween the voltage dependence of ClC-4 coupled transportand ClC-4 transport under uncoupling conditions, we per-formed measurements at variable ionic concentrations.Lowering external pH led to strong shift of the ClC-4 trans-port activation to more positive potentials (Fig. S6). Themagnitude of this shift did not allow resolving the upperpart of the expected Boltzmann curve and more quantitativedescription of the observed effects.
Changes in external Cl� did not shift activation of ClC-4transport (Fig. 5 C). In contrast, decreased concentration of
Biophysical Journal 100(5) 1233–1241

1238 Orhan et al.
internal anions led to a pronounced left-shift of the activa-tion of ClC-4 transport (Fig. 5 D). This altered voltagedependence results in higher ClC-4 transport currents atdepolarized potentials near zero mV (Fig. S7).
DISCUSSION
ClC-4 exhibits pronounced time- and voltage-dependentchanges in current amplitude. Under uncoupling conditions,these changes are based on altered numbers of active trans-porters -resembling gating of ion channels (14). We herestudied under uncoupling conditions the gating propertiesof human ClC-4 at different internal and external anion andproton concentrations. Our investigations provide insightsinto interactions of the transported substrates with mamma-lian ClC-type anion transporters.
Voltage-dependent gating in ClC transporters
Time- and voltage-dependent changes have been studied forClC-4 and ClC-5 by three independent groups (14,16,43,46).Based on noise analysis and substrate concentrationdependence, we interpreted the time- and voltage depen-dence of uncoupled ClC-4 transport as change in the numberof open and closed channels (14). Zdebik et al. (16) proposeda similar gating mechanism for coupled transport activityof ClC-5. They postulated burstlike transporter activityoriginating from a voltage-dependent gating process thatcauses ClC-5 to reside in either an active transporting or aninactive, nontransporting state. These authors suggestedthat within each burst, transport is very fast, resulting indiscrete microscopic current events, resembling openingand closing of ion channels with averaged single channelconductance.
Both models imply that transporter gating can be mathe-matically described using the classical framework devel-oped for ion channels without imposing scientific bias.These two models are technically identical, indicating thatour analysis can be used for channel as well as for mecha-nistically undefined slippage modes. The analysis of thisstudy does not require exact knowledge about the under-lying transport mechanisms and does not aim to distinguishbetween a channel or transporter interpretation of the un-coupled mode in ClC-4.
Separation of coupled and uncoupled ClC-4currents
To study ClC-4 under uncoupling conditions we used SCN�
as sole permeant external anion. This maneuver results inmore-than-tenfold excess of uncoupled ClC-4 anioncurrents. With internal iodide, this proportion is slightlyhigher (Fig. S4). Therefore, the different unitary transportrates allow determining properties of the uncoupled ClC-4
Biophysical Journal 100(5) 1233–1241
transport in isolation, even though a number of ClC-4proteins might still function as coupled transporters (14).
With Cl� as sole permeant anion, ClC-4 functions exclu-sively as coupled transporter (14). Changed ionic conditionsmight theoretically lead to different degree of uncoupling ofClC-4 in addition to the described gating alterations.However, we have already demonstrated that variation ofexternal [SCN�] does not change the number of transportersdetected by noise analysis (14). Moreover, similar measure-ments with internal I� revealed no changes in the couplingstoichiometry (Fig. S4). Taken together, these results indi-cate that (under our experimental conditions) changes inthe concentration of the permeant anions do not modifythe uncoupling action of external SCN�—therefore permit-ting accurate determination of gating alteration of coupledtransport and of transport under uncoupling conditions.
Gating of uncoupled ClC exchangers
The anion-proton exchanger ClC-4 can exhibit definedvoltage dependence (14). We report data showing thatsome properties of ClC-4 gating under uncoupling condi-tions resemble gating of ClC channels. First, the apparentgating charge associated with depolarization-induced gatingof ClC-4 (~�1.3e, see Fig. 1) is very similar to valuesobtained for the fast gating process of ClC channels(25,26,29). Second, activation of uncoupled ClC-4 deviatesfrom the canonical Boltzmann behavior in exhibiting non-zero open probability at extreme voltages. Lastly, ClC-4gating is modulated by external anions (Fig. 2) in a mannerthat resembles ClC channel behavior.
The depolarization activated gating process of ClC-4 ischaracterized by a nonzero open probability at negative volt-ages (Fig. 1 and Fig. S2). Such behavior can be describedonly by a gating scheme containing at least two separatepathways of activation with opposite voltage dependence(25,26). Simplistically, this behavior can be depicted usingthe following scheme:
C1/D
O)HC2 (SCHEME 2)
with pathways (D) and (H) activated by depolarization andhyperpolarization. Applying voltage pulses at depolarizedpotentials would increase the population of proteins acti-vating along the (D)-pathway and decrease the populationalong the (H)-pathway. However, ClC-4 behaves differently:preconditioning pulses at depolarized potentials markedlyenhance the transport probability at negative voltages(Fig. S2). Therefore, we conclude that ClC-4 gating iscontrolled by at least two distinct processes with differentvoltage dependences.
Gating of ClC channels is currently thought to occur bya degraded transporter mechanism. In a previous work(14), we proposed a schematic transporter mechanismfor uncoupling of ClC-4 by polyatomic anions. In this

ClC-4 Gating 1239
mechanism, anions different from Cl� tend to lock the ClCtransporters in a longer lasting permeable conformation de-pending on the applied voltage. The complexity of ClC-4gating under uncoupling conditions, however, suggeststhat at least one additional voltage-dependent gating processexists. Similarly to ClC channels, the voltage dependence ofthese processes might originate from proton- or anionbinding.
Dependence of ClC-4 gating on extracellularand intracellular anion concentration
Under uncoupling conditions, voltage-dependent gating ofClC-4 is modified both by intracellular and extracellularanions. External anions promote activation, resembling theanion dependence of ClC channel gating. This indicatesthat binding of external SCN� is voltage-dependent.
In contrast to the behavior of ClC-4 under uncouplingconditions, we did not detect gating alternations of coupledtransport caused by lower external [Cl�] (Fig. 5). This issimilar to the described effects of external anions onthe gating of ClC-5 (46). Our explanation for this observa-tion is the much higher binding affinity of external Cl�,when compared to SCN� (14,17). Assuming sigmoidalMichaelis-Menten binding, large gating alternations areexpected in the region around the KM value. In contrast,at high substrate concentrations a saturating behavior isto be expected. Therefore, to detect significant gatingchanges, one has to lower the external [Cl�] to values aslow as 20 mM. Unfortunately, the transport currents atsuch concentrations are strongly reduced, which preventsmeasurements of steady-state activation curves.
The effects of intracellular anions on ClC-4 gatingare qualitatively different from those on ClC channels.Increased intracellular anion concentration activates ClC-0by causing a shift in the voltage dependence of the fast gatingprocess to more negative potentials and by increasedminimum open probability (25). In ClC-4, increased concen-trations of I� and Cl� oppose opening (Figs. 3 and 5). Thiseffect suggests the existence of an additional high-affinityion binding site at the intracellular side of the protein, andthat this site crucially determines the time- and voltagedependence of ClC-4 currents. Experimental evidence forsuch a binding site has been provided previously bydescribing diverging effects of intra- and extracellular I(and SCN) on ClC-4 currents (36).
The existence of binding sites with different affinities hasbeen proposed as an explanation for the asymmetric rectifi-cation of ClC transport (17). In accordance with this hypoth-esis, the equilibrium binding affinities of ClC-4 for externalSCN� and internal Cl� and I� differ by approximately threeorders of magnitude, which would explain the strong recti-fication of ClC-4 currents in the negative range. Remark-ably, the binding affinities of Cl� to the internal andcentral site in the bacterial ClC protein ClC-ec1, obtained
by Picollo et al. (17), are identical with the estimated valueprovided by our analysis here (~800 mM). This points tohigh structural conservation between ClC-ec1 and themammalian ClC transporters and suggests that anionbinding to one of the two sites (Sint and Scent) in ClC-4 isan important part involved in voltage-dependent gating inmammalian ClC exchangers. A recent crystallographicstudy of a eukaryotic ClC exchanger suggests a pore trans-porter mechanism with very low slippage rate for anions atthe internal side of the protein (33). Based on these twostudies, we propose that the strong block by intracellularanions observed in ClC-4 originates from binding to ananion site, accessible for internal anions (Sint or Scent).This will imply a very intimate relation between transportand voltage-dependent gating in ClC transporters. A similarintimate relation has been established for the channel branchof the ClC protein family (31,32,47).
External and internal pH dependence of ClC-4gating
The effects of internal and external pH on ClC-4 differfrom previously reported pH modulation of ClC channelgating and are not predicted by any existing models.Lowering the external as well as the internal pH opensClC-0 and ClC-1 (26,30,31,47–49). In contrast, macro-scopic ClC-4 and ClC-5 currents are reduced by decreasingthe external pH (35,50–52). Physiologically, the blockingaction of external protons might play a role as a feedbackmechanism preventing endosomal overacidification. Wehere demonstrated that this pH-dependent current reductionis due to changes in gating — extracellular acidificationshifts steady-state activation curves of ClC-4 to more depo-larized potentials reducing the amount of active transporters(Fig. 4). We observed similar behavior also for the coupledmode of ClC-4. Block by external protons has beenreported for ClC-K and ClC-2 channels (8,53,54). Inboth cases, the effects are not conferred by protonation ofthe so-called gating glutamate (Gluex). ClC-K channelslack this residue. In ClC-2, recent work has demonstratedprotonation of H532 to be responsible for regulation byexternal pH (54). Therefore, additional protonation sitesmight be responsible for the effects of external protonsobserved in ClC-4.
We did not detect shifts in the voltage dependence of acti-vation for 100-fold different internal proton concentrations.This suggests that the depolarization-activated gatingprocess is insignificantly dependent on the intracellularpH (Fig. 4). This is in clear contrast to many ClC channelsfor which proton transport is coupled to channel gating(28,31,49,55). This differencemight originate in the differenttransport mechanisms of ClC channels and transporters. Thecoupled antiport of anions and protons requires a strictcoupling between the transport of internal and externalsubstrates, a feature not present in ClC channels. The
Biophysical Journal 100(5) 1233–1241

1240 Orhan et al.
uncoupled anion transport therefore might fail to respond tochanges in intracellular pH.
Smith and Lippiat (43) detected voltage-dependentconformational changes in ClC-5 in the absence of permeantanions, Therefore, a certain proportion of the gating chargeinvolved in the voltage-dependent gating of mammalianClC transporters might originate from protein conforma-tional changes. However, such a gating charge might alsobe caused from the binding and unbinding of protons.Because internal protons do not shift the voltage depen-dence of ClC-4 transport (Fig. 4), the origin might be,instead, proton binding to the external gating glutamate.This hypothesis is supported by the voltage-dependentproton block, described for ClC-5 (52).
Structural basis of ClC-4 gating
For ClC channels, the existence of two structurally distinctgating processes is already well established (21). Whereasthe fast gating process opens and closes individual proto-pores, there is a slow gating process mediating cooperativeopening and closing of both protopores. A conserved externalglutamate residue (Gluex) located at the entry to the selec-tivity filter is an important determinant of fast protoporegating (33,56,57). ForClC-4, there is some evidence support-ing the idea that the depolarization-induced gating processcorresponds to the fast gating in ClC channels. For example,it is eliminated by neutralizing the conserved glutamate sidechain, the gating glutamate involved in fast gating (16,35,56).However, because mutations at this position affect slowgating as well (24,48,58), this finding is not sufficient forthe unambiguous assignment of the observed ClC-4 gatingprocesses to the fast or slow gating of ClC channels.
Physiological relevance of voltage-dependentgating in ClC transporters
Recent data suggested that endosomal accumulation of Cl�
by intracellular ClC exchangers such as ClC-5 and ClC-7 isphysiologically important (59,60). There is no indicationthat ClC-4 should be considered different. Because of itsintracellular localization, ClC-4 is exposed to low [Cl�] atits extraendosomal side. Lowering of cytoplasmic [Cl�]will reduce the driving force for Cl�-Hþ exchange andimpair anion accumulation. Gating of ClC exchangers mightcompensate for this effect. At a concentration of 10 mM, thetransport probability strongly increases at ~0 mV (Figs. 3and 5). Such changes will affect macroscopic transport ratesand thus modify endosomal pH and [Cl�]. Enhanced activa-tion at low intracellular [Cl�] might therefore keep aniontransport constant despite the lowered electromotive drivingforce.
Some of the findings reported here are obtained exclu-sively under strong uncoupling conditions. Thereforefurther studies will be necessary to define the physiologicalimportance of these processes.
Biophysical Journal 100(5) 1233–1241
For many years, voltage-dependent gating of intracellularClC exchangers has been assumed unimportant. We herereported what we believe to be novel regulatory roles ofanions and protons that might ensure endosomal acidifica-tion and anion accumulation over varying cellular condi-tions and point to the physiological significance of thesefunctional features.
SUPPORTING MATERIAL
Seven supplementary figures (Figs. S1, S2, S3, S4, S5, S6, and S7) and
seven tables (Tables S1, S2, S3, S4, S5, S6 and S7) are available at
http://www.biophysj.org/biophysj/supplemental/S0006-3495(11)00129-9.
The authors thank Drs. Martin Fischer and Gabriel Stoelting for helpful
discussions.
This work was supported by a HiLF grant of the Medical School Hannover.
REFERENCES
1. Jentsch, T. J. 2008. CLC chloride channels and transporters: from genesto protein structure, pathology and physiology. Crit. Rev. Biochem.Mol. Biol. 43:3–36.
2. Koch, M. C., K. Steinmeyer, ., T. J. Jentsch. 1992. The skeletalmuscle chloride channel in dominant and recessive human myotonia.Science. 257:797–800.
3. George, Jr., A. L., M. A. Crackower, ., G. C. Ebers. 1993. Molecularbasis of Thomsen’s disease (autosomal dominant myotonia congenita).Nat. Genet. 3:305–310.
4. Saint-Martin, C., G. Gauvain, ., C. Depienne. 2009. Two novelCLCN2 mutations accelerating chloride channel deactivation are asso-ciated with idiopathic generalized epilepsy. Hum. Mutat. 30:397–405.
5. Simon, D. B., R. S. Bindra, ., R. P. Lifton. 1997. Mutations in thechloride channel gene, CLCNKB, cause Bartter’s syndrome type III.Nat. Genet. 17:171–178.
6. Catalan,M.,M. I. Niemeyer,., F. V. Sepulveda. 2004.Basolateral ClC-2 chloride channels in surface colon epithelium: regulation by a directeffect of intracellular chloride. Gastroenterology. 126:1104–1114.
7. Janssen, A. G., U. Scholl, ., C. Fahlke. 2009. Disease-causingdysfunctions of barttin in Bartter syndrome type IV. J. Am. Soc.Nephrol. 20:145–153.
8. Estevez, R., T. Boettger,., T. J. Jentsch. 2001. Barttin is a Cl� channelb-subunit crucial for renal Cl� reabsorption and inner ear Kþ secretion.Nature. 414:558–561.
9. Riazuddin, S., S. Anwar, ., C. Fahlke. 2009. Molecular basis ofDFNB73: mutations of BSND can cause nonsyndromic deafness orBartter syndrome. Am. J. Hum. Genet. 85:273–280.
10. Lloyd, S. E., S. H. S. Pearce, ., R. V. Thakker. 1996. A commonmolecular basis for three inherited kidney stone diseases. Nature.379:445–449.
11. Kornak, U., D. Kasper, ., T. J. Jentsch. 2001. Loss of the ClC-7chloride channel leads to osteopetrosis in mice and man. Cell.104:205–215.
12. Poet, M., U. Kornak,., T. J. Jentsch. 2006. Lysosomal storage diseaseupon disruption of the neuronal chloride transport protein ClC-6. Proc.Natl. Acad. Sci. USA. 103:13854–13859.
13. Graves, A. R., P. K. Curran, ., J. A. Mindell. 2008. The Cl�/Hþ anti-porter ClC-7 is the primary chloride permeation pathway in lysosomes.Nature. 453:788–792.
14. Alekov, A. K., and Ch. Fahlke. 2009. Channel-like slippage modes inthe human anion/proton exchanger ClC-4. J. Gen. Physiol. 133:485–496.

ClC-4 Gating 1241
15. Matsuda, J. J., M. S. Filali, ., F. S. Lamb. 2010. The ClC-3 Cl�/Hþ
antiporter becomes uncoupled at low extracellular pH. J. Biol. Chem.285:2569–2579.
16. Zdebik, A. A., G. Zifarelli,., M. Pusch. 2008. Determinants of anion-proton coupling in mammalian endosomal CLC proteins. J. Biol.Chem. 283:4219–4227.
17. Picollo, A., M. Malvezzi, ., A. Accardi. 2009. Basis of substratebinding and conservation of selectivity in the CLC family of channelsand transporters. Nat. Struct. Mol. Biol. 16:1294–1301.
18. Fahlke, Ch., R. Rudel, ., A. L. George, Jr. 1995. An aspartic acidresidue important for voltage-dependent gating of human muscle chlo-ride channels. Neuron. 15:463–472.
19. Pusch, M., K. Steinmeyer, ., T. J. Jentsch. 1995. Mutations in domi-nant human myotonia congenita drastically alter the voltage depen-dence of the CIC-1 chloride channel. Neuron. 15:1455–1463.
20. Wu, F. F., A. Ryan,., C. Fahlke. 2002. Novel CLCN1 mutations withunique clinical and electrophysiological consequences. Brain.125:2392–2407.
21. Miller, C. 1982. Open-state substructure of single chloride channelsfrom Torpedo electroplax. Philos. Trans. R. Soc. Lond. B Biol. Sci.299:401–411.
22. Accardi, A., and M. Pusch. 2000. Fast and slow gating relaxations inthe muscle chloride channel CLC-1. J. Gen. Physiol. 116:433–444.
23. Zuniga, L., M. I. Niemeyer, ., F. V. Sepulveda. 2004. The voltage-dependent ClC-2 chloride channel has a dual gating mechanism.J. Physiol. 555:671–682.
24. de Santiago, J. A., K. Nehrke, and J. Arreola. 2005. Quantitative anal-ysis of the voltage-dependent gating of mouse parotid ClC-2 chloridechannel. J. Gen. Physiol. 126:591–603.
25. Chen, T. Y., and C. Miller. 1996. Nonequilibrium gating and voltagedependence of the ClC-0 Cl� channel. J. Gen. Physiol. 108:237–250.
26. Fahlke, Ch., A. Rosenbohm, ., R. Rudel. 1996. Mechanism ofvoltage-dependent gating in skeletal muscle chloride channels.Biophys. J. 71:695–706.
27. Scholl, U., S. Hebeisen, ., C. Fahlke. 2006. Barttin modulates traf-ficking and function of ClC-K channels. Proc. Natl. Acad. Sci. USA.103:11411–11416.
28. Hanke, W., and C. Miller. 1983. Single chloride channels from Torpedoelectroplax. Activation by protons. J. Gen. Physiol. 82:25–45.
29. Pusch, M., U. Ludewig, ., T. J. Jentsch. 1995. Gating of the voltage-dependent chloride channel CIC-0 by the permeant anion. Nature.373:527–531.
30. Rychkov, G. Y., M. Pusch, ., A. H. Bretag. 1996. Concentration andpH dependence of skeletal muscle chloride channel ClC-1. J. Physiol.497:423–435.
31. Lısal, J., and M. Maduke. 2008. The ClC-0 chloride channel isa ‘broken’ Cl�/Hþ antiporter. Nat. Struct. Mol. Biol. 15:805–810.
32. Chen, T. Y., and T. C. Hwang. 2008. CLC-0 and CFTR: chloride chan-nels evolved from transporters. Physiol. Rev. 88:351–387.
33. Feng, L., E. B. Campbell, ., R. MacKinnon. 2010. Structure ofa eukaryotic CLC transporter defines an intermediate state in the trans-port cycle. Science. 330:635–641.
34. Steinmeyer, K., B. Schwappach, ., T. J. Jentsch. 1995. Cloning andfunctional expression of rat CLC-5, a chloride channel related tokidney disease. J. Biol. Chem. 270:31172–31177.
35. Friedrich, T., T. Breiderhoff, and T. J. Jentsch. 1999. Mutational anal-ysis demonstrates that ClC-4 and ClC-5 directly mediate plasmamembrane currents. J. Biol. Chem. 274:896–902.
36. Hebeisen, S., H. Heidtmann,., C. Fahlke. 2003. Anion permeation inhuman ClC-4 channels. Biophys. J. 84:2306–2318.
37. Zifarelli, G., and M. Pusch. 2009. Conversion of the 2 Cl�/1 Hþ anti-porter ClC-5 in a NO3
�/Hþ antiporter by a single point mutation.EMBO J. 28:175–182.
38. Otis, T. S., andM. P. Kavanaugh. 2000. Isolation of current componentsand partial reaction cycles in the glial glutamate transporter EAAT2.J. Neurosci. 20:2749–2757.
39. Watzke, N., E. Bamberg, and C. Grewer. 2001. Early intermediates inthe transport cycle of the neuronal excitatory amino acid carrierEAAC1. J. Gen. Physiol. 117:547–562.
40. Kovermann, P., J. P. Machtens, ., C. Fahlke. 2010. A conservedaspartate determines pore properties of anion channels associatedwith excitatory amino acid transporter 4 (EAAT4). J. Biol. Chem.285:23676–23686.
41. Hamill, O. P., A. Marty, ., F. J. Sigworth. 1981. Improved patch-clamp techniques for high-resolution current recording from cellsand cell-free membrane patches. Pflugers Arch. 391:85–100.
42. Barry, P. H. 1994. JPCalc, a software package for calculating liquidjunction potential corrections in patch-clamp, intracellular, epithelialand bilayer measurements and for correcting junction potentialmeasurements. J. Neurosci. Methods. 51:107–116.
43. Smith, A. J., and J. D. Lippiat. 2010. Voltage-dependent charge move-ment associated with activation of the CLC-5 2Cl�/1Hþ exchanger.FASEB J. 24:3696–3705.
44. Cui, J., D. H. Cox, and R. W. Aldrich. 1997. Intrinsic voltage depen-dence and Ca2þ regulation of mslo large conductance Ca-activatedKþ channels. J. Gen. Physiol. 109:647–673.
45. Woodhull, A. M. 1973. Ionic blockage of sodium channels in nerve.J. Gen. Physiol. 61:687–708.
46. Smith, A. J., and J. D. Lippiat. 2010. Direct endosomal acidificationby the outwardly rectifying CLC-5 Cl�/Hþ exchanger. J. Physiol.588:2033–2045.
47. Miller, C. 2006. ClC chloride channels viewed through a transporterlens. Nature. 440:484–489.
48. Traverso, S., G. Zifarelli,., M. Pusch. 2006. Proton sensing of CLC-0mutant E166D. J. Gen. Physiol. 127:51–65.
49. Zifarelli, G., A. R. Murgia, ., M. Pusch. 2008. Intracellular protonregulation of ClC-0. J. Gen. Physiol. 132:185–198.
50. Mo, L., H. L. Hellmich, ., N. K. Wills. 1999. Comparison ofamphibian and human ClC-5: similarity of functional properties andinhibition by external pH. J. Membr. Biol. 168:253–264.
51. Vanoye, C. G., and A. L. George, Jr. 2002. Functional characterizationof recombinant human ClC-4 chloride channels in cultured mammaliancells. J. Physiol. 539:373–383.
52. Picollo, A., M. Malvezzi, and A. Accardi. 2010. Proton block of theCLC-5 Cl�/Hþ exchanger. J. Gen. Physiol. 135:653–659.
53. Waldegger, S., N. Jeck, ., H. W. Seyberth. 2002. Barttin increasessurface expression and changes current properties of ClC-K channels.Pflugers Arch. 444:411–418.
54. Niemeyer, M. I., L. P. Cid, ., F. V. Sepulveda. 2009. Voltage-depen-dent and -independent titration of specific residues accounts forcomplex gating of a ClC chloride channel by extracellular protons.J. Physiol. 587:1387–1400.
55. Chen, M. F., and T. Y. Chen. 2001. Different fast-gate regulation byexternal Cl� and Hþ of the muscle-type ClC chloride channels.J. Gen. Physiol. 118:23–32.
56. Dutzler, R., E. B. Campbell, and R. MacKinnon. 2003. Gating theselectivity filter in ClC chloride channels. Science. 300:108–112.
57. Fischer, M., A. G. Janssen, and C. Fahlke. 2010. Barttin activatesClC-K channel function by modulating gating. J. Am. Soc. Nephrol.21:1281–1289.
58. Fahlke, Ch., H. T. Yu, ., A. L. George, Jr. 1997. Pore-formingsegments in voltage-gated chloride channels. Nature. 390:529–532.
59. Novarino, G., S. Weinert, ., T. J. Jentsch. 2010. Endosomal chloride-proton exchange rather than chloride conductance is crucial for renalendocytosis. Science. 328:1398–1401.
60. Weinert, S., S. Jabs, ., T. J. Jentsch. 2010. Lysosomal pathology andosteopetrosis upon loss of Hþ-driven lysosomal Cl� accumulation.Science. 328:1401–1403.
Biophysical Journal 100(5) 1233–1241