androgen spares androgen-insensitive motoneurons from apoptosis in the spinal nucleus of the...
TRANSCRIPT

Hormones and Behavior 30, 424–433 (1996)Article No. 0047
Androgen Spares Androgen-InsensitiveMotoneurons from Apoptosis in the SpinalNucleus of the Bulbocavernosus in Rats
Louise M. Freeman,*,1 Neil V. Watson,*,2 andS. Marc Breedlove*,†*Department of Psychology and †Graduate Group in Neuroscience, 3210 Tolman Hall,University of California, Berkeley, California 94720-1650
ing a site of androgen action other than the motoneuronsThe spinal nucleus of the bulbocavernosus (SNB) is athemselves. q 1996 Academic Presssexually dimorphic motor nucleus in the rat lumbar spi-
nal cord. The sex difference arises through the andro-genic sparing of the motoneurons and their target mus-cles from ontogenetic cell death. Indirect evidence sug-gests that androgen acts on the target muscles rather Neuronal death is a widespread phenomenon in thethan directly on SNB motoneurons to spare them from developing central nervous system (CNS) and one ofdeath. The testicular feminization mutation (Tfm), a de-
the major factors shaping the brain. In rats, most popu-fect in the androgen receptor (AR), blocks androgeniclations of motoneurons undergo ontogenetic cell deathsparing of SNB motoneurons and their targets. The pat-prenatally (Harris and McCaig, 1984; Oppenheim,tern of AR immunocytochemistry was previously found1986); however, for motoneurons of the spinal nucleusto be different in adult Tfm and wild-type rats: immuno-of the bulbocavernosus (SNB), this apoptosis is delayed.staining was nuclear in most SNB cells of wild-type rats,The SNB is a lumbar motor nucleus that is influencedbut very few SNB cells display nuclear AR immunostain-
ing in affected Tfm rats. Because the Tfm mutation is by androgen during both development and adulthood.carried on the X chromosome, random X inactivation In male rats the SNB innervates two sexually dimorphicduring development makes female carriers of Tfm (// striated muscles, the bulbocavernosus (BC) and levatorTfm) genetic mosaics for androgen sensitivity. Tfm carri- ani (LA), as well as the sexually monomorphic externalers, their wild-type sisters, and affected Tfm males were anal sphincter (Breedlove and Arnold, 1980; Schroder,treated with perinatal testosterone and immunocyto- 1980). A perinatal surge of testosterone in males rescueschemistry was used to detect androgen receptor in the
the BC and LA muscles from degeneration (Cihak, Gut-SNB when the rats reached adulthood. Mosaic femalesmann, and Hanzlikova, 1970) and also rescues somecould be distinguished from their wild-type sisters bySNB motoneurons from apoptosis (Nordeen, Nordeen,external morphology. In such perinatally androgenizedSengelaub, and Arnold, 1985). Therefore, adult malesmosaics, adult SNB cells were equally divided betweenhave more and larger SNB motoneurons than do fe-wild-type and Tfm genotype, as indicated by AR immu-males (Breedlove and Arnold, 1980, 1981). Treating fe-nocytochemistry. In contrast, the pattern of AR immuno-males pups with androgen during this same criticalcytochemistry in target muscles of mosaics appeared
similar to that of wild-type females. These results indi- period can permanently masculinize their SNB cellcate that early androgen spared both androgen-sensitive number (Breedlove and Arnold, 1983). Male rats withand -insensitive motoneurons from cell death, confirm- the sex-linked recessive testicular feminization muta-
tion (Tfm), a point mutation that renders 85 to 90%of androgen receptors (ARs) nonfunctional (Yarbrough,1 Present address:Department of Biology, 229 Gilmer Hall, Univer-Quarmby, Simental, Joseph, Sart, Lubahn, Olsen,sity of Virginia, Charlottesville, VA 22903.French, and Wilson, 1990), have a feminine SNB2 Present address: Department of Psychology, Simon Fraser Univer-
sity, Burnaby, British Columbia V5A 1S6, Canada. (Breedlove and Arnold, 1981). Thus it seems clear that
424
0018-506X/96 $18.00Copyright q 1996 by Academic Press
All rights of reproduction in any form reserved.
AID H&B 1351 / 6807$$$321 01-02-97 15:54:05 habas AP: H & B

425Androgen Spares Tfm Motoneurons
androgenic activation of the AR is crucial for survival This analysis indicates testosterone is just as likely tospare androgen-insensitive SNB motoneurons as wild-of SNB motoneurons in male rats.
In which tissues is activation of the AR crucial for type SNB cells from apoptosis. The crucial site for an-drogen action to spare SNB cells must therefore be inSNB motoneuron survival? There is circumstantial evi-
dence that androgen maintains the muscles and thereby tissue(s) other than the motoneurons themselves.indirectly spares SNB motoneurons (Forger, Hodges,Roberts, and Breedlove, 1992, review). For example, an- MATERIALS AND METHODSdrogen can spare the developing BC from involutionin females that have had the lumbosacral spinal cord Breeding and Identification of Androgenized Tfm(including SNB motoneurons) removed (Fishman and CarriersBreedlove, 1988). Furthermore, AR expression has been
Eight Tfm carrier females from our breeding colonyfound in the target muscles, but not the motoneurons,(identified by having previously given birth to affectedduring the critical period for androgen action (Fishman,Tfm males) were paired with males. Vaginal smearsChism, Firestone, and Breedlove, 1990; Jordan,were taken daily; the day of positive sperm smear wasBreedlove, and Arnold, 1991), and local treatment ofrecorded as the first embryonic day (E1). Dams werethe muscle with antiandrogen blocks masculinizationinjected with testosterone propionate (TP, 2 mg in ses-of the SNB more effectively than the same dose of anti-ame oil subcutaneously) on E16–20, the critical periodandrogen given systemically (Fishman and Breedlove,for androgenic elimination of nipple formation (Gold-1992). Removing the target muscles results in death ofman, Shapiro, and Neuman, 1976). On E23, the damsSNB motoneurons despite androgen treatment (Kurz,were anesthetized with ether, the pups were deliveredCover, and Sengelaub, 1992), but it is possible that theby cesarean section, and the dams were sacrificed bymotoneurons themselves normally respond directly topentobarbital overdose. Pups were dried and warmed,androgen, but cannot do so when deprived of a target.injected with TP (1 mg subcutaneously) and cross-fos-In this study, we attempted to eliminate the motoneu-tered to another lactating dam. All pups received a sec-ron as the site of action for the androgenic sparing ofond 1-mg TP injection on Postnatal Day 3 (P3). In wild-SNB cells from cell death in rats by producing genetictype rats, the prenatal androgen regimen had been re-mosaics for the AR, i.e., rats having some cells that areported to block the development of external nipples inTfm and others that are wild type. We did this by givingrats (Goldman et al., 1976), a result confirmed by pilotperinatal androgen to Tfm carrier pups (XXTfm). Becausestudies of our own (Freeman, unpublished observa-random X inactivation occurs in the embryonic femaletions). The postnatal androgen treatments were added(Lyon, 1992), roughly half of the motoneurons of het-to maximally masculinize the SNB system in females.erozygous Tfm carrier pups should be androgen insen-
Pups were weaned on P30 and classified by pheno-sitive. If androgen acts at the target muscles to sparetype at that time (Table 1). Males had palpable testes,SNB cells from apoptosis during development, both an-either descended into the scrotal sac (wild-type males)drogen-sensitive and -insensitive motoneurons shouldor in the abdomen (affected Tfm males). Wild-typesurvive. If the motoneurons are the site of action, wemales also displayed no nipples, but affected Tfm maleswould expect to see predominantly wild-type motoneu-developed nipples despite their endogenous androgenrons in adulthood (Fig. 1). We previously demonstratedproduction and the perinatal androgen treatment.that immunocytochemistry (ICC) with the polyclonalAmong the animals without testes (females), thoseantibody PG21 (a gift of G. Prins) can detect ARs in thewithout visible nipples were classified as wild-type fe-lumbar spinal cord, and that motoneurons in Tfm malesmales. Females in which TP had failed to prevent nipplediffer from those in wild-type males by their failureformation were judged to be partially androgen insensi-to show nuclear immunostaining (Freeman, Padgett,tive and, therefore, considered Tfm mosaics. This classi-Prins, and Breedlove, 1995). In this study we used AR-fication of females as wild type or mosaic was con-ICC to distinguish wild-type and Tfm motoneuronsfirmed by other measures to be discussed later. We referwithin the SNB of mosaic female rats.to these females as wild type and mosaic hereafter.We now describe (1) a method of perinatally mascu-
linizing Tfm carrier rats and distinguishing them fromAR-ICC in Androgenized Tfm Mosaicstheir wild-type sisters, and (2) the pattern of AR-ICC
staining in such perinatally androgenized Tfm carriers, Seventeen mosaics were selected at random from thelitters and assigned to one of two adult hormone treat-their wild-type sisters, and affected Tfm male controls.
AID H&B 1351 / 6807$$$321 01-02-97 15:54:05 habas AP: H & B
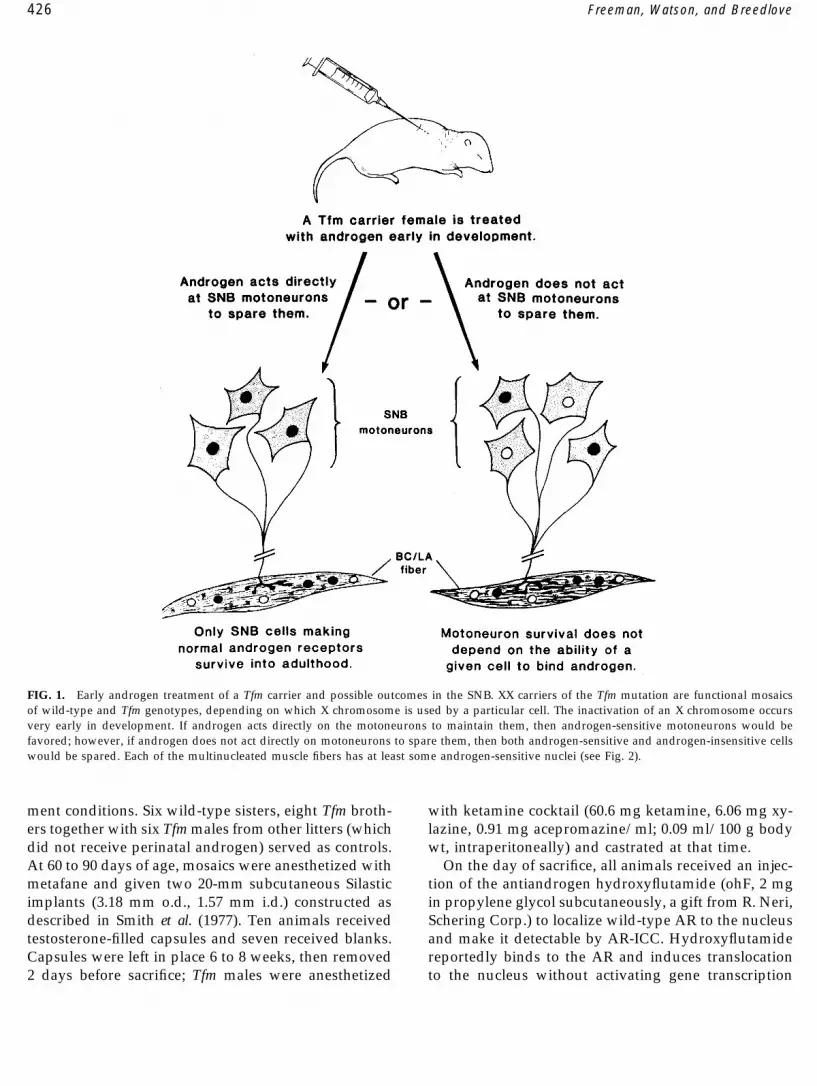
426 Freeman, Watson, and Breedlove
FIG. 1. Early androgen treatment of a Tfm carrier and possible outcomes in the SNB. XX carriers of the Tfm mutation are functional mosaicsof wild-type and Tfm genotypes, depending on which X chromosome is used by a particular cell. The inactivation of an X chromosome occursvery early in development. If androgen acts directly on the motoneurons to maintain them, then androgen-sensitive motoneurons would befavored; however, if androgen does not act directly on motoneurons to spare them, then both androgen-sensitive and androgen-insensitive cellswould be spared. Each of the multinucleated muscle fibers has at least some androgen-sensitive nuclei (see Fig. 2).
ment conditions. Six wild-type sisters, eight Tfm broth- with ketamine cocktail (60.6 mg ketamine, 6.06 mg xy-lazine, 0.91 mg acepromazine/ml; 0.09 ml/100 g bodyers together with six Tfm males from other litters (which
did not receive perinatal androgen) served as controls. wt, intraperitoneally) and castrated at that time.On the day of sacrifice, all animals received an injec-At 60 to 90 days of age, mosaics were anesthetized with
metafane and given two 20-mm subcutaneous Silastic tion of the antiandrogen hydroxyflutamide (ohF, 2 mgin propylene glycol subcutaneously, a gift from R. Neri,implants (3.18 mm o.d., 1.57 mm i.d.) constructed as
described in Smith et al. (1977). Ten animals received Schering Corp.) to localize wild-type AR to the nucleusand make it detectable by AR-ICC. Hydroxyflutamidetestosterone-filled capsules and seven received blanks.
Capsules were left in place 6 to 8 weeks, then removed reportedly binds to the AR and induces translocationto the nucleus without activating gene transcription2 days before sacrifice; Tfm males were anesthetized
AID H&B 1351 / 6807$$$321 01-02-97 15:54:05 habas AP: H & B

427Androgen Spares Tfm Motoneurons
TABLE 1
Phenotypes and Presumed Genotypes of Offspring from Eight Perinatally Androgenized Litters of Tfm Carrier Damsa
Gonads Ovaries Ovaries Testes TestesNipples Absent Present Absent PresentPresumed genotype XX XXTfm XY XTfmYClassification Wild-type females Mosaic females Wild-type males Affected Tfm malesNumber of pups 30 24 26 26
a x2 test shows that the numbers of each phenotype obtained are consistent with the expected 1:1:1:1 ratio (P ú 0.05).
(Wong, Zhou, Sar, and Wilson, 1994); in SNB motoneu- for AR-ICC and one of which served as a no-primarycontrol.rons, it is apparently the absence of ligand binding that
reduces nuclear PG21 immunostaining in Tfm males One immunoreacted slide from each animal was cov-erslipped without counterstain and examined under(Freeman et al., 1995). Four mosaic and four Tfm animals
received ohF 8 hr before sacrifice; the remaining ani- light microscopy. Putative SNB motoneuron nucleiwere identified as darkly stained circles located justmals were injected 2 hr before sacrifice. All animals
received a lethal dose of sodium pentobarbital and were ventrolateral to the central canal. A camera lucidadrawing of each section was made, mapping the loca-perfused intracardially with phosphate-buffered saline
(PBS) followed by 4% paraformaldehyde. Gonadal sex tion of apparent motoneuron nuclei. Darkly labeled nu-clei were scored as 2 and lightly labeled as 1. Coverslipswas confirmed by dissection, and the LA muscles from
wild-type females and mosaics were removed and were then soaked off and the tissue counterstained withneutral red Nissl stain and recoverslipped. SNB moto-stored in 4% paraformaldehyde. Later, they were
trimmed, blotted dry, and weighed. neurons were identified by their location, large size,and dark Nissl stain. The identities of the putative mo-Spinal cords were removed, postfixed 1 to 3 hr, then
soaked overnight in 20% buffered sucrose. The follow- toneuron nuclei seen in the uncounterstained conditionwere confirmed. Additional motoneurons whose nucleiing day the lumbosacral segments were frozen-sec-
tioned in the frontal plane at 50 mm and the free-floating had no visible immunolabeling were also mapped andscored as 0’s. Total number of SNB cells was thensections were rinsed in a phosphate-buffered gelatin–
Triton solution (PGT, 0.1% gelatin, 0.3% Triton in PBS). counted and the percentage of labeled cells calculated.When calculating total number of SNB cells, all appar-Consecutive sections were divided between three wells,
so every third section could be processed without the ent motoneurons were counted (0’s, 1’s, and 2’s); how-ever, when calculating the percentage of labeled moto-primary antiserum.
Sections were incubated first for 1 hr at room temper- neurons, the lightly labeled cells (1’s) were eliminatedfrom the analysis because their identity as wild-type orature in 10% normal goat serum (NGS), then for 48 hr
at 47C in the primary antiserum, PG21 (0.186 mg/ml Tfm motoneurons was less certain.LA muscles were also removed from five wild-type,solution in PGT with 4% NGS). PG21 is a polyclonal
antiserum raised against the first 21 amino acids of the three testosterone-treated mosaic, and four blank-treated mosaic animals. The muscles were frozen-sec-rat androgen receptor (Prins, Birch, and Greene, 1991).
Tissue was rinsed in PGT, treated with avidin and bio- tioned at a thickness of 50 mm, reacted for AR-ICC asabove, and coverslipped without counterstain, and thetin blocking solutions (obtained from Vector Labora-
tories, Burlingame, CA), rinsed again, then reacted with AR-positive nuclei within a region of approximately140,000 mm2 were counted with the aid of an eyepieceavidin–biotin–peroxidase complex kit (Vectastain Elite
goat anti-rabbit IgG kit, Burlingame, CA). Following a grid. Coverslips were removed, all nuclei stained withhematoxylin, and the slides coverslipped again. The1-hr incubation in biotinylated secondary antibody
(goat anti-rabbit), tissue was rinsed in PGT, then incu- same region of the muscle was located by use of stagemicrometers on the microscope and the total numberbated for 1 hr in avidin–biotin complex. Staining was
visualized with H2O2 and diaminobenzidine and inten- of nuclei within that region counted. The proportion ofAR-positive nuclei was calculated as the number of AR-sified with 1% NiCl2 . Tissue from each well was
mounted on a separate slide, so each animal’s cord was positive nuclei divided by the number of total nucleiseen with hematoxylin stain.divided into three parts, two of which were stained
AID H&B 1351 / 6807$$$321 01-02-97 15:54:05 habas AP: H & B

428 Freeman, Watson, and Breedlove
All counts were made without knowledge of pheno-type or adult hormone treatment condition. Differencesin total SNB cell number were evaluated by t test. Dif-ferences in the percentage of immunolabeled SNB cellsand in the percentage of AR-positive muscle nucleiwere evaluated by Mann–Whitney U test.
RESULTS
Breeding and Identification of Androgenized TfmCarriers
The eight perinatally androgenized litters yielded 106offspring, and in seven of eight litters, all four of thepredicted phenotypes were present. The different phe-notypes were predicted to be present in equal numbers,and x2 tests showed that they did not differ signifi-cantly from the expected value (Table 1, P ú 0.05).
FIG. 2. Androgen receptor immunocytochemistry in the levator aniMuscle Survival and AR-ICC muscle of an androgenized female mosaic rat. Note the large number
of immunopositive nuclei found across muscle fibers, which run fromGross dissection showed LA muscles had survived top to bottom in this view. Later hematoxylin staining (not shown)
of all nuclei indicates that about one-third of nuclei in levator aniin all wild-type females and in 16 of 17 mosaic females.muscle are AR-immunoreactive in either mosaic or wild-type andro-Among animals who were not treated with androgengenized females.in adulthood, LA muscles from wild-type females were
significantly heavier (197 { 11 mg) than those frommosaics that received blank capsules in adulthood (87
SNB Motoneurons{ 17 mg, P õ 0.01, two-tailed t test), concordant withtheir partial androgen insensitivity. In mosaics, adult In agreement with previous findings in wild-type rats
(Breedlove and Arnold, 1981), adult androgen treat-androgen treatment increased muscle size (214 { 16mg) compared with the blank-treated mosaics (P õ ment did not significantly affect the total SNB cell num-
ber in the mosaics (P ú 0.10); neither did it alter the0.01, two-tailed t test).A large number of AR-immunoreactive nuclei were percentage of SNB motoneurons showing PG21 immu-
nolabeling (P ú 0.10). Whether the ohF injection wasfound in the LA muscles of all animals. The pattern ofstaining in the LA of mosaic animals indicated that all given 8 or 2 hr before sacrifice also had no significant
effect on cell number (P ú 0.10) or percentage showingmuscle fibers had at least some AR-positive nuclei (Fig.2). The percentage of AR-positive nuclei in the LA mus- immunolabeling (Pú 0.05). Therefore, data from all the
mosaic animals were combined.cles of T-treated mosaics (29.9 { 5.37%) was not signifi-cantly different from that of blank-treated mosaics (39.1 Among the affected Tfm males, littermates of the
mosaics (who consequently received the same pre-{ 8.78%). Therefore, the data from mosaics were com-bined (35.1{ 5.44%) and compared with the percentage and postnatal androgen treatment) showed no sig-
nificant differences in either SNB cell number (P úof AR-positive nuclei in muscles from wild-type fe-males (37.8 { 5.2%). There were no significant differ- 0.20) or percentage immunolabeled (P ú 0.20) when
compared with the other Tfm animals that had re-ences between any of the groups for this measure (Pú 0.40 for all comparisons by Mann–Whitney U test), ceived no perinatal androgen. Eight-hour versus 2-hr
ohF treatment also did not influence cell number (Pindicating that the proportion of androgen-sensitive nu-clei was equivalent across groups. ú 0.50) or percentage labeled (P ú 0.05), so all Tfm
AID H&B 1351 / 6807$$$321 01-02-97 15:54:05 habas AP: H & B
429Androgen Spares Tfm Motoneurons
control animals were treated as one group for theremainder of the analysis.
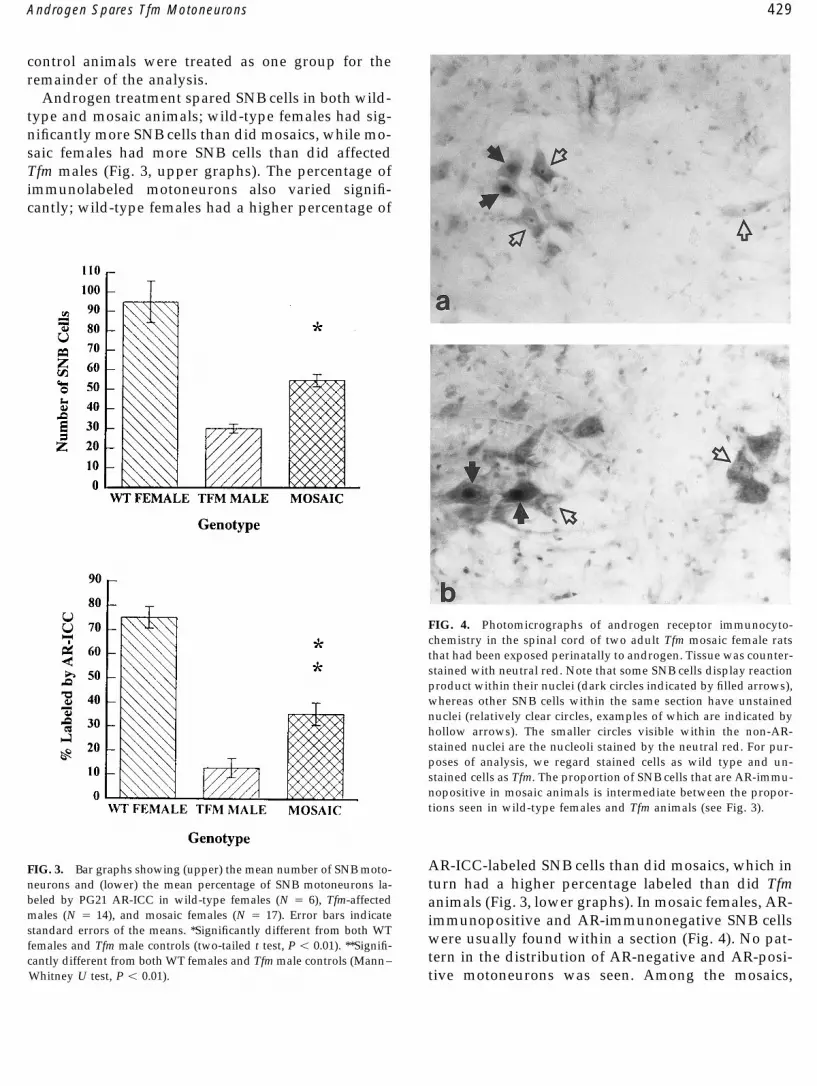
Androgen treatment spared SNB cells in both wild-type and mosaic animals; wild-type females had sig-nificantly more SNB cells than did mosaics, while mo-saic females had more SNB cells than did affectedTfm males (Fig. 3, upper graphs). The percentage ofimmunolabeled motoneurons also varied signifi-cantly; wild-type females had a higher percentage of
FIG. 4. Photomicrographs of androgen receptor immunocyto-chemistry in the spinal cord of two adult Tfm mosaic female ratsthat had been exposed perinatally to androgen. Tissue was counter-stained with neutral red. Note that some SNB cells display reactionproduct within their nuclei (dark circles indicated by filled arrows),whereas other SNB cells within the same section have unstainednuclei (relatively clear circles, examples of which are indicated byhollow arrows). The smaller circles visible within the non-AR-stained nuclei are the nucleoli stained by the neutral red. For pur-poses of analysis, we regard stained cells as wild type and un-stained cells as Tfm. The proportion of SNB cells that are AR-immu-nopositive in mosaic animals is intermediate between the propor-tions seen in wild-type females and Tfm animals (see Fig. 3).
AR-ICC-labeled SNB cells than did mosaics, which inFIG. 3. Bar graphs showing (upper) the mean number of SNB moto-neurons and (lower) the mean percentage of SNB motoneurons la- turn had a higher percentage labeled than did Tfmbeled by PG21 AR-ICC in wild-type females (N Å 6), Tfm-affected animals (Fig. 3, lower graphs). In mosaic females, AR-males (N Å 14), and mosaic females (N Å 17). Error bars indicate immunopositive and AR-immunonegative SNB cellsstandard errors of the means. *Significantly different from both WT
were usually found within a section (Fig. 4). No pat-females and Tfm male controls (two-tailed t test, P õ 0.01). **Signifi-tern in the distribution of AR-negative and AR-posi-cantly different from both WT females and Tfm male controls (Mann–
Whitney U test, P õ 0.01). tive motoneurons was seen. Among the mosaics,
AID H&B 1351 / 6807$$$321 01-02-97 15:54:05 habas AP: H & B
430 Freeman, Watson, and Breedlove
drogens; therefore, the differential responses to thetreatment must be the result of differential sensitivityto androgen. Since random X inactivation predicts thatTfm carriers will be partially insensitive to the receptor-mediated effects of androgen, it is logical to concludethat the animals in which nipples were not eliminatedare Tfm carriers, whereas the remaining females inwhich nipples were eliminated are homozygous wild-type females. Second, the ratio of phenotypes con-formed to the predicted patterns. Mendelian geneticspredicts that half the female offspring of female carriersof a sex-linked recessive trait will themselves be carri-ers, and that the four expected genotypes produced bythe cross of a Tfm carrier with a normal male (wild-type males, Tfm males, wild-type females, and carrierfemales) will be present in equal numbers. We did ob-serve four different phenotypes among the offspring ofsuch a cross:
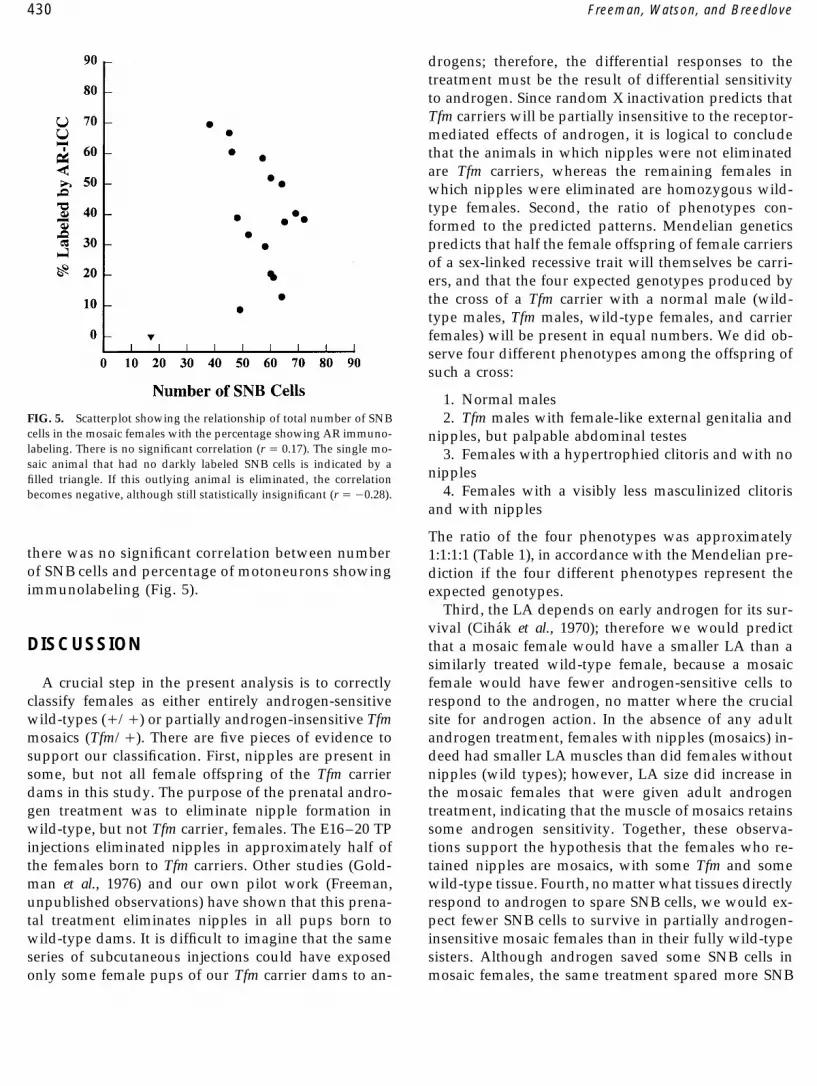
1. Normal malesFIG. 5. Scatterplot showing the relationship of total number of SNB 2. Tfm males with female-like external genitalia andcells in the mosaic females with the percentage showing AR immuno- nipples, but palpable abdominal testeslabeling. There is no significant correlation (r Å 0.17). The single mo- 3. Females with a hypertrophied clitoris and with nosaic animal that had no darkly labeled SNB cells is indicated by a
nipplesfilled triangle. If this outlying animal is eliminated, the correlation4. Females with a visibly less masculinized clitorisbecomes negative, although still statistically insignificant (r Å 00.28).
and with nipples
The ratio of the four phenotypes was approximatelythere was no significant correlation between number 1:1:1:1 (Table 1), in accordance with the Mendelian pre-of SNB cells and percentage of motoneurons showing diction if the four different phenotypes represent theimmunolabeling (Fig. 5). expected genotypes.
Third, the LA depends on early androgen for its sur-vival (Cihak et al., 1970); therefore we would predict
DISCUSSION that a mosaic female would have a smaller LA than asimilarly treated wild-type female, because a mosaicfemale would have fewer androgen-sensitive cells toA crucial step in the present analysis is to correctly
classify females as either entirely androgen-sensitive respond to the androgen, no matter where the crucialsite for androgen action. In the absence of any adultwild-types (///) or partially androgen-insensitive Tfm
mosaics (Tfm//). There are five pieces of evidence to androgen treatment, females with nipples (mosaics) in-deed had smaller LA muscles than did females withoutsupport our classification. First, nipples are present in
some, but not all female offspring of the Tfm carrier nipples (wild types); however, LA size did increase inthe mosaic females that were given adult androgendams in this study. The purpose of the prenatal andro-
gen treatment was to eliminate nipple formation in treatment, indicating that the muscle of mosaics retainssome androgen sensitivity. Together, these observa-wild-type, but not Tfm carrier, females. The E16–20 TP
injections eliminated nipples in approximately half of tions support the hypothesis that the females who re-tained nipples are mosaics, with some Tfm and somethe females born to Tfm carriers. Other studies (Gold-
man et al., 1976) and our own pilot work (Freeman, wild-type tissue. Fourth, no matter what tissues directlyrespond to androgen to spare SNB cells, we would ex-unpublished observations) have shown that this prena-
tal treatment eliminates nipples in all pups born to pect fewer SNB cells to survive in partially androgen-insensitive mosaic females than in their fully wild-typewild-type dams. It is difficult to imagine that the same
series of subcutaneous injections could have exposed sisters. Although androgen saved some SNB cells inmosaic females, the same treatment spared more SNBonly some female pups of our Tfm carrier dams to an-
AID H&B 1351 / 6807$$$321 01-02-97 15:54:05 habas AP: H & B

431Androgen Spares Tfm Motoneurons
cells in the wild-type females, again confirming our explained, given the following assumptions: (1) in fe-males and Tfm males, a small pool of motoneurons (ap-classification of these females. Perinatal androgen treat-
ment of mice, in which coat color markers can be used proximately 30, according to the counts in this study)normally survives even in the absence of androgen toto classify Tfm carrier and wild-type females, resulted
in a pattern similar to that based on our classification of innervate the external anal sphincter; (2) in wild-typefemales, where all motoneurons are androgen sensitive,rats: fewer SNB cells were spared by perinatal androgen
treatment of carriers than their wild-type sisters (Olsen, perinatal androgen treatments save 60 to 70 more moto-neurons, which survive to innervate the BC/LA mus-Wagner, and Clemens, 1988). Finally, females with nip-
ples had a lower percentage of SNB motoneurons la- cles. If androgen acted directly on the motoneurons tosave them, we would expect the same treatment to savebeled by AR-ICC than did their wild-type sisters, as
would be predicted if some motoneurons were express- only about half that number (30–40) in the mosaics,since, on average, only half the motoneurons would being the Tfm gene in mosaics. We are therefore extremely
confident that nipple retention in gonadal females who expected to express the functional AR. Perinatal andro-gen treatment would result in a SNB cell number higherreceived perinatal testosterone correctly identifies ge-
netically mosaic Tfm carriers. than that of Tfm animals but lower than that of wild-type androgenized females. Furthermore, if we add theA second crucial issue is whether we accurately iden-
tified wild-type and Tfm SNB cells within the mosaic (untested) assumption that nondimorphic external analsphincter motoneurons are much less likely to showfemales. In this case we know that our classification is
imperfect. The AR-ICC results in the wild-type female, PG21 immunostaining than are BC/LA motoneurons,this scenario could also explain the lower percentageand Tfm male control groups indicate that our accuracy
in distinguishing androgen-sensitive from -insensitive of motoneurons showing immunostaining in mosaics.External anal sphincter motoneurons would make up amotoneurons is not absolute; wild-type motoneurons
indicate a ‘‘false-negative’’ rate of approximately 25%; greater percentage of total SNB motoneurons in mosaicfemales than in wild-type; therefore we would expect ai.e., only 75% of SNB cells are strongly AR-ICC positive
under present conditions, even in entirely wild-type lower percentage of SNB cells to show immunolabeling.We, however, believe this explanation is unlikely.females. Similarly, Tfm motoneurons indicate a ‘‘false-
positive’’ rate of around 13% (Fig. 3, middle bar in Among the mosaics there was a wide range in bothSNB cell number (17–72) and percentage showing AR-lower graph). Despite these limitations, AR-ICC labels
a significantly smaller proportion of SNB motoneurons ICC (0–70%). If wild-type motoneurons are selectivelyspared from cell death during development, we wouldin mosaics than in wild-type females, indicating that at
least some of the motoneurons spared by early andro- expect a positive correlation between SNB cell numberand percentage of SNB cells showing nuclear AR im-gen express the Tfm rather than the wild-type AR. Mo-
saics have significantly more AR-positive SNB cells munostaining. No such correlation is seen (Fig. 5, r Å0.17). Furthermore, one mosaic animal had no apparentthan the Tfm males and significantly fewer AR-positive
cells than wild-type females, indicating a difference ro- LA muscle in adulthood; this animal (indicated as afilled triangle in Fig. 5) also had the fewest SNB cellsbust enough to be detected despite the presence of false-
positive and false-negative classifications. Therefore, (17) and no darkly immunostained motoneurons. Thelack of masculinization in this mosaic may be due tothe site of androgen’s action in sparing SNB cells from
cell death must be other than the motoneurons them- an especially high proportion of Tfm cells. If this animalis eliminated from the analysis, the correlation becomesselves. This study agrees with the finding of a similar
sparing of androgen-insensitive motoneurons in two negative, although still statistically insignificant (r Å00.28). This strongly argues against any type of selec-sex-reversed mice (Breedlove, 1986). This interpretation
also conforms to other studies that provide indirect evi- tive sparing of wild-type SNB cells; if anything, themosaics with the most SNB cells tend to have propor-dence that the site of action in rats is the muscle (Fish-
man and Breedlove, 1985, 1988, 1992; Fishman et al., tionately fewer motoneurons expressing AR immuno-staining.1990).
One alternative interpretation is that only wild-type It is also possible that fewer cells were counted inthe mosaics because cells were less likely to be recog-SNB motoneurons can be spared from cell death by
early androgen, and that the reason fewer cells survive nized as motoneurons. For instance, Tfm motoneuronscould have smaller somata or lighter Nissl staining andin mosaics is that fewer androgen-sensitive motoneu-
rons were initially available. Our results could then be therefore be less likely to be counted as a motoneuron
AID H&B 1351 / 6807$$$321 01-02-97 15:54:05 habas AP: H & B

432 Freeman, Watson, and Breedlove
under light microscopy. Having a darkly immuno- ics, it should be possible to compare the two popula-tions in a within-subject design. The two populationsstained nucleus, on the other hand, may make a cell
more likely to be counted as a motoneuron, resulting would presumably receive similar signals from the tar-get muscles or supraspinal afferents, so any differencesin more discernible motoneurons in wild-type than in
mosaic animals. The lack of a correlation between num- between the two populations after androgen treatmentcan be attributed to AR-mediated response of the indi-ber of SNB cells and percentage showing AR-ICC stain-
ing among mosaics, however, also argues against wild- vidual neuron. Both soma size (Breedlove and Arnold,1981) and dendritic extent (Kurz, Sengelaub, and Ar-type motoneurons being more detectable than Tfm mo-
toneurons. It is therefore likely that androgen is less nold, 1986) are regulated by androgen in adulthood;comparing Tfm with wild-type motoneurons could de-effective in sparing SNB cells from death in mosaics
than in wild types because the target muscles of mosa- termine if androgen acts on the motoneuron to affecteither morphological measure. In addition, double-la-ics are less androgen sensitive than those of wild types.
The smaller size of the muscles in mosaics is consistent beling techniques could be used to identify proteinscoexpressed with, and possibly regulated by, the AR.with that hypothesis. Despite its decreased efficacy in
mosaics, androgen can spare both wild-type and Tfm The identification of such factors would be an im-portant step in understanding how androgens influencemotoneurons from ontogenetic cell death, eliminating
the motoneuron itself as a site of action for that process the development and plasticity of the nervous system.of sexual differentiation.
What of the BC/LA target muscles? Our predictionis that the BC/LA target muscles of mosaics would also ACKNOWLEDGMENTbe mosaic for functional AR. Because muscle fibers aremultinucleated, each muscle fiber would still be ex- This work was supported by NIH Grant NS28421.pected to possess some functional ARs. In fact, wefound that the LA muscles of mosaic animals possessabout the same proportion of AR-immunoreactive nu- REFERENCESclei as seen in wild-type animals—about one-third ofthe total nuclei in the muscle. Furthermore, that andro-
Breedlove, S. M. (1986). Cellular analyses of hormone influence ongen spares the BC/LA muscles in mosaic females, butmotoneuronal development and function. J. Neurobiol. 17, 157–176.
not in Tfm males, confirms that the muscles of mosaics Breedlove, S. M., and Arnold, A. P. (1980). Hormone accumulation inare at least partially sensitive to androgen. On the other a sexually dimorphic motor nucleus in the rat spinal cord. Science
210, 564–566.hand, the same perinatal androgen treatment results inBreedlove, S. M., and Arnold, A. P. (1981). Sexually dimorphic motora smaller adult BC/LA muscle in mosaics than in wild-
nucleus in the rat lumbar spinal cord: Response to adult hormonaltype females, presumably because the mosaic femalesmanipulation, absence in androgen-insensitive adult rats. Brain Res.
begin with only partially sensitive precursors to form 225, 297–307.the muscles. Breedlove, S. M., and Arnold, A. P. (1983). Hormonal control of a
developing neuromuscular system. II. Sensitive periods for the an-Whether the proportion of AR-positive nucleidrogen-induced masculinization of the rat spinal nucleus of thechanges during the development of mosaics, i.e.,bulbocavernosus. J. Neurosci. 3, 424–432.whether androgen promotes the proliferation of AR-
Cihak, R., Gutmann, E., and Hanzlıkova, V. (1970). Involution andpositive myoblasts more than AR-negative myoblasts, hormone-induced persistence of the M. sphincter (levator) ani inwas not addressed in this study, but androgen has been female rats. J. Anat. 106, 93–100.
Fishman, R. B., and Breedlove, S. M. (1985). The androgenic inductionreported to promote proliferation in vitro of myoblastsof spinal sexual dimorphism is independent of supraspinal affer-derived from newborn rat thigh muscle (Powers andents. Dev. Brain Res. 23, 255–258.Florini, 1975). Determining whether androgen has such
Fishman, R. B., and Breedlove, S. M. (1988). Neonatal androgen main-an effect on the rat BC/LA in vivo would be difficult tains sexually dimorphic muscles in the absence of innervation.because only electron microscopy permits distinguish- Muscle Nerve 11, 553–560.
Fishman, R. B., and Breedlove, S. M. (1992). Local perineal implantsing muscle fiber nuclei from other muscle cells.of anti-androgen block masculinization of the SNB. Dev. Brain Res.Perinatally androgenized Tfm carriers may prove to70, 283–286.be a valuable tool for investigating the sites of androgen
Fishman, R. B., Chism, L., Firestone, G. L., and Breedlove, S. M. (1990).action in both development and adulthood. Since both Evidence for androgen receptors in sexually dimorphic perinealandrogen-sensitive and -insensitive motoneurons are muscles of neonatal male rats: Absence of androgen accumulation
by the perineal motoneurons. J. Neurobiol. 21, 694–704.present in the adult spinal cord of androgenized mosa-
AID H&B 1351 / 6807$$$321 01-02-97 15:54:05 habas AP: H & B

433Androgen Spares Tfm Motoneurons
Forger, N. G., Hodges, L. L., Roberts, S. L., and Breedlove, S. M. Olsen, K. L., Wagner, C. K., and Clemens, L. G. (1988). Relation-(1992). Regulation of motoneuron death in the spinal nucleus of ship between male sexual behaviors and dorsal medial spinalthe bulbocavernosus, J. Neurobiol. 23, 1192–1203. nucleus in androgen-insensitive Tfm mice. Soc. Neurosci. Abstr.
Freeman, L. M., Padgett, B. A., Prins, G. S., and Breedlove, S. M. 14, 283.(1995). Distribution of androgen receptor immunoreactivity in the Oppenheim, R. W. (1986). The absence of significant postnatal moto-spinal cord of wild-type, androgen-insensitive and gonadectomized neuronal death in the brachial and lumbar spinal cord of the rat.male rats. J. Neurobiol. 27, 51–59. J. Comp. Neurol. 246, 281–286.
Goldman, A. S., Shapiro, B. H., and Neuman, F. (1976). Role of testos- Powers, M. L., and Florini, F. R. (1975). A direct effect of testosteroneterone and its metabolites in the differentiation of the mammary on muscle cells in tissue culture. Endocrinology 97, 1043–1047.gland in rats. Endocrinology 99, 1490–1495. Prins, G. S., Birch, L., and Greene, G. L. (1991). Androgen receptor
Harris, A. J., and McCaig, C. D. (1984). Motoneuron death and motor localization in different cell types of the adult rat prostate. Endocri-unit size during embryonic development of the rat. J. Neurosci. 4, nology 129, 3187–3199.13–24. Schroder, H. D. (1980). Organization of the motoneurons innervating
Jordan, C. L., Breedlove, S. M., and Arnold, A. P. (1991). Ontogeny of the pelvic muscles of the male rat. J. Comp. Neurol. 192, 567–587.steroid accumulation in spinal lumbar motoneurons of the rat. J.
Smith, E. R., Damassa, D. A., and Davidson, J. M. (1977). HormoneComp. Neurol. 313, 441–448.
administration: peripheral and intracranial implants. In R. D. Mey-Kurz, E. M., Cover, A. R., and Sengelaub, D. R. (1992). Testosteroneers (Ed.), Methods in Psychobiology, Vol. III. Academic Press, Newfails to save androgen-sensitive rat motoneurons following earlyYork.target removal. Dev. Brain Res. 70, 181–189.
Wong, C.-I., Zhou, Z.-X., Sar, M., and Wilson, E. M. (1994). SteroidKurz, E. M., Sengelaub, D. R., and Arnold, A. P. (1986). Androgensrequirement for androgen receptor dimerization and DNA binding.regulate the dendritic length of mammalian motoneurons in adult-J. Biol. Chem. 268, 19004–190012.hood. Science 232, 395–398.
Yarbrough, W. G., Quarmby, V. E., Simental, J. A., Joseph, D. R., Sart,Lyon, M. F. (1992). Some milestones in the history of X-chromosomeM., Lubahn, D. B., Olsen, K. L., French, F. S., and Wilson, E. M.inactivation. Annu. Rev. Genetics 26, 17–28.(1990). A single base mutation in the androgen receptor gene causesNordeen, E. J., Nordeen, K. W., Sengelaub, D. R., and Arnold, A. P.androgen insensitivity in the testicular feminized rat. J. Biol. Chem.(1985). Androgens prevent normally occurring cell death in a sexu-
ally dimorphic spinal nucleus. Science 229, 671–673. 265, 8893–8900.
AID H&B 1351 / 6807$$$321 01-02-97 15:54:05 habas AP: H & B