analysis of synthetic peptides from heptad-repeat domains of herpes simplex virus type 1...
TRANSCRIPT

Analysis of synthetic peptides from heptad-repeatdomains of herpes simplex virus type 1glycoproteins H and B
Stefania Galdiero,1,2,3 Mariateresa Vitiello,2,4 Marina D’Isanto,4
Annarita Falanga,4 Craig Collins,5 Katia Raieta,4 Carlo Pedone,1,2,3
Helena Browne5 and Massimiliano Galdiero2,4
Correspondence
Massimiliano Galdiero
1,2Department of Biological Sciences, Division of Biostructures1 and Centro Interuniversitariodi Ricerca sui Peptidi Bioattivi2, University of Naples Federico II, Via Mezzocannone 16,
80134 Naples, Italy
3Istituto di Biostrutture e Bioimmagini, CNR, Via Mezzocannone 16, 80134 Naples, Italy
4Department of Experimental Medicine, II University of Naples, Via De Crecchio 7, 80138Naples, Italy
5Division of Virology, Department of Pathology, University of Cambridge, Cambridge, UK
Received 22 December 2005
Accepted 20 January 2006
Human herpesviruses enter cells by fusion of their own membrane with a cellular membrane
through the concerted action of multiple viral proteins and cellular receptors. Two conserved
viral glycoproteins, gB and gH, are required for herpes simplex virus type 1 (HSV-1)-mediated
membrane fusion, but little is known of how these proteins cooperate during entry. Both
glycoproteins were shown to contain heptad repeat (HR) sequences predicted to form a-helical
coiled coils, and the inhibitory activity against infection of four sets of synthetic peptides
corresponding to HR1 and HR2 of gB and gH was tested. The interactions between these HR
peptides were also investigated by circular dichroism, native polyacrylamide-gel electrophoresis
and size exclusion high-performance liquid chromatography. gH coiled-coil peptides were more
effective than gB coiled-coils peptides in inhibiting virus infectivity. The peptides did not impair
fusion when added to cells immediately after infection. In contrast, inhibition of infection was
observed, albeit to various extents, when peptides were added to virus before or during inoculation.
The results of biophysical analyses were indicative of the existence of an interaction between
HR1 and HR2 of gH and suggest that the HRs of gB and gH do not interact with each other.
INTRODUCTION
Infection of a target cell by enveloped viruses requires avirally mediated mechanism to promote membrane fusion(Weissenhorn et al., 1999; Earp et al., 2005). This mechan-ism seems surprisingly conserved across several diverse viralfamilies, which makes the fusion process a potentiallyattractive target for antiviral drugs for a number of humanand animal viruses (Dimitrov, 2004).
Enveloped viruses use transmembrane (TM) viral proteinsto mediate fusion with host cell membranes (Hernandezet al., 1996; Eckert &Kim, 2001; Blumenthal et al., 2003; Jahnet al., 2003). Key structural elements shared by most viralfusion glycoproteins are a fusion peptide and a coiled-coilbundle of helices. The fusion peptide is a hydrophobica-helix able to penetrate the membrane of the target cell inorder to destabilize the lipid bilayer organization. By meansof the fusion peptide, inserting in the host cell membrane,and the TM domain, which anchors the protein on the viral
envelope, a bridge is formed between the two membranes.Subsequent rearrangements in the conformation of theseproteins lead to the close juxtaposition of the viral andcellular membranes, culminating in the merging of twobilayers. In addition to the fusion peptide domain, manyfusion proteins also contain one or more heptad-repeat(HR) regions, which are often adjacent to the fusion peptidesequence and to the TM anchor domain (Chambers et al.,1990; Skehel & Wiley, 1998). There are usually two of theseregions: an N-terminal HR region (HR1) adjacent to thefusion peptide and a C-terminal HR region (HR2) close tothe TM anchor. Structural studies of several viral fusionproteins revealed that theHR regions form a six-helix bundlestructure implicated in viral entry (Eckert & Kim, 2001). TheHR1 and HR2 domains associate to form a six-helix bundle,with a central trimeric coiled-coil domain formed by HR1helices; the three HR2 helices pack obliquely in an anti-parallel configuration into the highly hydrophobic grooveson the surface of the central coiled coil. This conformation
0008-1794 G 2006 SGM Printed in Great Britain 1085
Journal of General Virology (2006), 87, 1085–1097 DOI 10.1099/vir.0.81794-0

brings the N-terminal fusion peptide into close proximity tothe TM anchor. Recent evidence suggests that the actual six-helix bundle formation is directly coupled to the merging ofthe membranes (Melikyan et al., 2000; Russell et al., 2001).The similarities in the structures of the six-helix bundlecomplexes elucidated for influenza virus haemagglutinin(HA) (Bullough et al., 1994; Chen et al., 1999), Humanimmunodeficiency virus type 1 (HIV-1) and Simian immuno-deficiency virus (SIV) (Weissenhorn et al., 1997; Caffrey et al.,1998; Chan&Kim, 1998;Malashkevich et al., 1998),MoloneyMurine leukemia virus type 1 gp21 (Fass et al., 1996), Ebolavirus GP2 (Weissenhorn et al., 1998; Malashkevich et al.,1999), human T-cell leukemia virus type I gp21 (Kobe et al.,1999), Visna virus TM (Malashkevich et al., 2001), simianparainfluenza virus 5 (SV5) F1 (Baker et al., 1999) andhuman respiratory syncytial virus (HRSV) F1 (Zhao et al.,2000), all point to a common fusion mechanism for theseviruses. Based on structural similarities, two classes of viralfusion proteins have been distinguished (Jardetzky & Lamb,2004). Proteins containing HR regions and an N-terminalor N-proximal fusion peptide are classified as class I viralfusion proteins. Class II viral fusion proteins (e.g. thealphavirus E1 and the flavivirus E fusion proteins) lack HRregions and have an internal fusion peptide. Their fusionprotein is folded in tight association with a second protein asa heterodimer. Here, fusion activation takes place uponcleavage of the second protein.
Therefore, the role of coiled coils in the entry of severalviruses has been widely investigated, demonstrating thata-helical coiled coils form the basis for critical protein–protein interactions within the fusogenic glycoprotein andplay a pivotal role in membrane fusion. In fact, when solublepeptides comprising the HR sequences are presented tovirus during inoculation into cells, entry of the virus issignificantly impaired (Wild et al., 1994; Rapaport et al.,1995; Lambert et al., 1996; Wild & Buckland, 1997; Joshiet al., 1998; Kilby et al., 1998).
Fusogenic machines of most viruses are composed of oneor two glycoproteins, which, in the majority of cases, canperform both receptor recognition and binding as well asviral penetration. On the other hand, herpesviruses specifymultiple envelope glycoproteins, a subset of which are knownto play a role in membrane fusion (Mettenleiter 2002a, b;Spear & Longnecker, 2003; Spear, 2004). Herpesvirusesinfect their target cells by fusion of the viral envelope withthe cellular plasma membrane at neutral pH. For penetra-tion, a set of envelope glycoproteins that are conservedthroughout the subfamilies of the family Herpesviridae isrequired. This includes glycoproteins B (gB), H (gH) and L(gL). In addition, each subfamily employs different receptor-binding proteins. Herpes simplex virus type 1 (HSV-1)attachment requires gB, gC and gD. The first two bind to cellsurface proteoglycans, whereas gD interacts with cellularsurface receptors from the tumour necrosis factor receptoror immunoglobulin superfamilies (Montgomery et al., 1996;Cocchi et al., 1998; Geraghty et al., 1998;Warner et al., 1998;
Lopez et al., 2000). Interaction of gD with its receptor thentriggers the fusionmachinery, which results in the formationof a continuous membrane consisting of the plasma mem-brane of the infected cell and the viral envelope (Cocchi et al.,2004). Fusion of the HSV-1 envelope with the plasmamembrane is mediated by four glycoproteins, gB, gD, gHand gL (Turner et al., 1998).
The main candidate proteins for performing fusion are gHand gB. Recently, Gianni et al. (2005a) reported that gHpossesses a fusion peptide, although this does not rule outthe possibility that other glycoproteins, such as gB, may alsoplay an active role in the fusion process. Furthermore, ithas also been shown that, besides this region, additionalhydrophobic domains of gH may be required for efficientinduction of fusion (Galdiero et al., 2005).
gH shares features typical of both class I and class II virusfusion proteins. In analogy to type I membrane proteins, itcontains two HR sequences, one located near the putativefusion peptide and one in close proximity to the TM region.However, in common with many class II fusion proteins, gHlacks an N-terminal or even N-proximal fusion peptide inthemembrane-anchored subunit; there is no evidence that itexists as a trimer and the cleavage of gH is not required formembrane fusion.
The role of HSV-1 gB in fusion is less well defined. However,HR regions have been identified in the human cytomegalo-virus (HCMV) gB homologue (Lopper & Compton, 2004).gB may also be a type I membrane glycoprotein and recentcrystallographical evidence suggests that HSV-1 gB mayexist as a trimer (Heldwein et al., 2005).
To understand the molecular events in the early steps ofHSV-1 infection, we sought to determine the functional rolesof different regions of gH and gB a-helical coiled coils inmembrane fusion and virus entry.
A recent report by Gianni et al. (2005b) identified a pre-dicted coil region in HSV-1 gH. In addition, synthetic pep-tides modelled on coiled coils derived from glycoproteins ofother members of the herpesvirus family, namely BHV-1(Okazaki & Kida, 2004) and HCMV (Lopper & Compton,2004), have recently been described.
Using an algorithm to detect potential coiled coils, we identi-fied two HR regions in gH and two in gB and generatedpeptides of different lengths that correspond to the pre-dicted coiled-coil regions. We demonstrated that the gHcoiled-coil peptides were more effective than gB coiled-coilpeptides in inhibiting virus infectivity. The peptides did notimpair fusion when added to cells immediately followinginfection. In contrast, when peptides were added to virusbefore or during inoculation into cells, inhibition of infectionwas observed, albeit to various extents. In the present study,we also applied biophysical methods to determine whetherthe HR regions present in HSV-1 gH and gB were able toestablish intramolecular or intermolecular interactions.
1086 Journal of General Virology 87
S. Galdiero and others

METHODS
Materials. Fluorenylmethoxycarbonyl-protected amino acids werepurchased from INBIOS; NovaSyn TGA resin from Nova Biochem.The reagents (piperidine and pyridine) for the solid phase peptidesynthesis were purchased from Fluka (Sigma-Aldrich); trifluoroaceticacid (TFA) and acetic anhydride were purchased from AppliedBiosystem. H2O, dimethylformamide and CH3CN were supplied byLAB-SCAN.
Proteomics computational methods. The bioinformatic searchfor coils in HSV gH (protein Swiss-Prot accession no. P08356) andgB (protein Swiss-Prot accession no. P10211) was performed withthe program Coils (Lupas et al., 1991), with window widths of 14,21 and 28.
Peptide synthesis. Peptides were synthesized using a standardsolid-phase-9-fluorenylmethoxycarbonyl method, on a PSSM8 multi-specific peptide synthesizer (Shimadzu) as described previously(Galdiero et al., 2005).
Cells and virus. Vero cells were grown in Dulbecco’s modifiedEagle’s medium (DMEM) supplemented with 10% fetal calf serum.HSV-1 carrying a lacZ gene driven by the CMV IE-1 promoter toexpress b-galactosidase was propagated as described previously(Forrester et al., 1992).
Virus entry assays. Peptides were dissolved in DMEM withoutserum and used at a range of concentrations. All experiments wereconducted in parallel with scrambled peptides (peptides with thesame amino acids but in a different order) and no-peptide controls.
To assess the effect of peptides on inhibition of HSV infectivity, fourdifferent ways of treating cell monolayers were performed.
Virus pre-treatment. Approximately 26104 p.f.u. HSV-1 wereincubated in the presence of different concentrations of peptides(10, 100, 250 and 500 mM) for 45 min at 37 uC, then titrated onVero cell monolayers.
Cell pre-treatment. Vero cells were incubated with peptides (10,100, 250 and 500 mM) for 30 min at 4 uC. Peptides were removedand cells were washed with PBS before being infected with serialdilutions of HSV-1 and incubated for 45 min at 37 uC.
Co-treatment. Cells were incubated with increasing concentrationsof the peptides (10, 100, 250 and 500 mM) in the presence of theviral inoculum for 45 min at 37 uC.
Post-treatment. Vero cells monolayers were challenged with HSV-1for 45 min at 37 uC. Different concentrations of peptides (10, 100,250 and 500 mM) were then added to the inoculum, followed by afurther 30 min incubation at 37 uC.
For all treatments, non-penetrated viruses were inactivated by citratebuffer at pH 3?0 after the 45 min incubation with cells at 37 uC.Cells were then incubated for 24 h at 37 uC in DMEM supplementedwith carboxymethylcellulose (CMC). Monolayers were fixed, stainedwith X-Gal (5-bromo-4-chloro-3-indolyl b-D-galactopyranoside) andplaque numbers were scored. Experiments were performed in triplicateand the percentage of inhibition was calculated with respect to no-peptide control experiments.
Circular dichroism (CD) measurements. CD experiments atdifferent concentrations of H-HR1 and H-HR2 (5, 10, 20, 40 and50 mM) were performed in 5 mM phosphate buffer pH 7?4 asdescribed by Galdiero et al. (2005). For mixing experiments, 10 mMof each peptide was prepared.
Toxicity. Peptide cytotoxicity was measured by a lactate dehydro-genase (LDH) assay that was carried out according to manufac-turer’s instructions using a cytotoxicity detection kit (Roche).
SDS-PAGE. Isolated peptides (7?5 mM in phosphate buffer, pH 7?3)were incubated at 100 uC for 5 min (final volume of 5 ml) and analysedby SDS-PAGE in denaturing conditions on 18% polyacrylamide gel.
Native-PAGE (N-PAGE). Isolated peptides and equimolar mixturesof peptides (7?5 mM in phosphate buffer, pH 7?3) were incubatedat 37 uC for 30 min (final volume of 5 ml). After addition of an equalvolume of Tricine sample buffer [0?0625 M Tris/HCl, pH 6?8, 40%(v/v) glycerol, 0?001 g bromophenol blue] the mixture was analysedby PAGE on 12% Tricine gel with a Tricine/glycine running buffer(pH 8?3). Gel electrophoresis was carried out with 25 A constantamperages at room temperature for 2?5 h. The gel was then stainedwith Coomassie blue.
Analytical size exclusion (SE) chromatography. A BioSEP-SEC-S2000 column (30067?8 mm; Phenomenex) was used for SEchromatography (SE-HPLC) to assess oligomer formation. Peptideswere injected into the column connected to an HPLC system (class-LC10 equipped with a diode array detector SPD-M10AV; Shimadzu).HEPES/Na buffer (pH 7?4) was used as a mobile phase with a flowrate of 0?5 ml min21. Peptide mixtures were prepared in the elutionbuffer and injected into the column to assess the formation ofpeptide-complexes. Standards, consisting of ovalbumin (44 kDa),cytochrome C (12 kDa), antp (1?5 kDa) aspy (1?0 kDa) and RPK(0?4 kDa), were used to define the molecular masses of the peptidesand their complex. Antp (Ac-ERQIKIAAQNRR-COOH), aspy (Ac-RKARKAAAR-COOH) and RPK (NH2-RPK-COOH) are peptides ofknown molecular mass.
RESULTS
Bioinformatic analysis
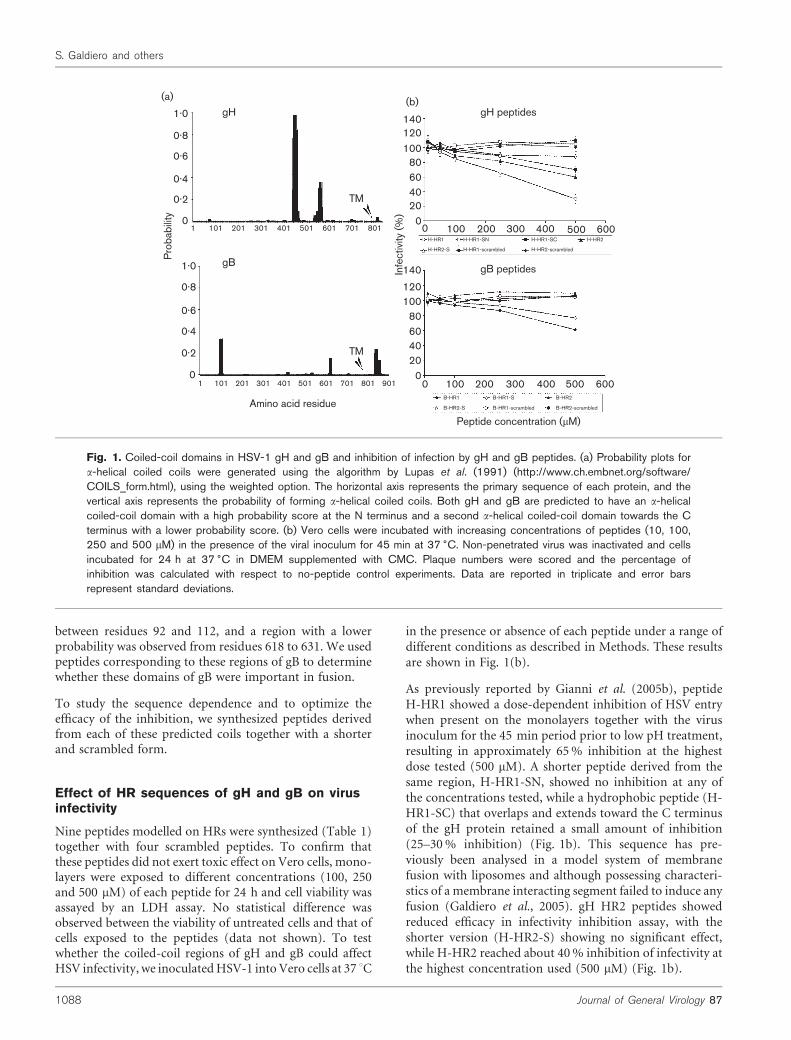
By using the Coils program (Lupas et al., 1991), two HRregions were identified in HSV-1 gH and one in gB, withprobability scores higher than 0?1 (Fig. 1a); moreover athird coil was identified in the cytoplasmatic tail of gB butwas not considered in this study. The two HR motifs in gHwere designated HR-1 and HR-2; HR-1, presents a very highprobability of forming a coiled-coil structure (with a scoreof 0?9) and as recently reported by Gianni et al. (2005b) italso corresponds to residues 444–476 of HSV-1 gH; HR-2was predicted at residues 542–582 with a lower probabilityof forming a coiled coil (0?4 with the weighted option and0?1 with the unweighted option). To avoid false-positivesdue to the presence of highly charged sequences, whichalthough presenting high coiled-coil probabilities do notreally have heptad periodicity and coiled-coil formingpotential, we used the weighted option. It has been recentlyreported (Gianni et al., 2005b) that HR-1 (peptide 444–468),corresponding to the smallest region of gH with the highestprobability to form coils, plays a critical role in the fusionmechanism, so we analysed further the fusion activity ofHR-1 of HSV-1 gH using peptides of different length and, inaddition, verified the activity of HR-2.
To elucidate the potential role played by gB in fusion, weperformed a similar analysis on the gB amino acid sequence.A first coil with a probability higher than 0?3 was found
http://vir.sgmjournals.org 1087
HSV-1 gB and gH heptad repeat regions
between residues 92 and 112, and a region with a lowerprobability was observed from residues 618 to 631. We usedpeptides corresponding to these regions of gB to determinewhether these domains of gB were important in fusion.
To study the sequence dependence and to optimize theefficacy of the inhibition, we synthesized peptides derivedfrom each of these predicted coils together with a shorterand scrambled form.
Effect of HR sequences of gH and gB on virusinfectivity
Nine peptides modelled on HRs were synthesized (Table 1)together with four scrambled peptides. To confirm thatthese peptides did not exert toxic effect on Vero cells, mono-layers were exposed to different concentrations (100, 250and 500 mM) of each peptide for 24 h and cell viability wasassayed by an LDH assay. No statistical difference wasobserved between the viability of untreated cells and that ofcells exposed to the peptides (data not shown). To testwhether the coiled-coil regions of gH and gB could affectHSV infectivity, we inoculatedHSV-1 intoVero cells at 37 uC
in the presence or absence of each peptide under a range ofdifferent conditions as described in Methods. These resultsare shown in Fig. 1(b).
As previously reported by Gianni et al. (2005b), peptideH-HR1 showed a dose-dependent inhibition of HSV entrywhen present on the monolayers together with the virusinoculum for the 45 min period prior to low pH treatment,resulting in approximately 65% inhibition at the highestdose tested (500 mM). A shorter peptide derived from thesame region, H-HR1-SN, showed no inhibition at any ofthe concentrations tested, while a hydrophobic peptide (H-HR1-SC) that overlaps and extends toward the C terminusof the gH protein retained a small amount of inhibition(25–30% inhibition) (Fig. 1b). This sequence has pre-viously been analysed in a model system of membranefusion with liposomes and although possessing characteri-stics of a membrane interacting segment failed to induce anyfusion (Galdiero et al., 2005). gH HR2 peptides showedreduced efficacy in infectivity inhibition assay, with theshorter version (H-HR2-S) showing no significant effect,while H-HR2 reached about 40% inhibition of infectivity atthe highest concentration used (500 mM) (Fig. 1b).
(a)
1.0
0.8
0.6
0.4
0.2
0
gH
TM
(b)
140120100
80604020
1.0
0.8
0.6
0.4
0.2
0
Pro
babi
lity
140
120100
80604020
0
0
gB peptides
0 100 200 300 400 500 600H-HR1
H-HR2-S
H-HR1-SN
H-HR1-scrambled
H-HR1-SC
H-HR2-scrambled
H-HR2
B-HR1
B-HR2-S
B-HR1-S
B-HR1-scrambled
B-HR2
B-HR2-scrambled
0 100 200 300 400 500 600In
fect
ivity
(%)
Amino acid residue
Peptide concentration (mM)
gB
TM
1 101 201 301 401 501 601 701 801
1 101 201 301 401 501 601 701 801 901
gH peptides
Fig. 1. Coiled-coil domains in HSV-1 gH and gB and inhibition of infection by gH and gB peptides. (a) Probability plots fora-helical coiled coils were generated using the algorithm by Lupas et al. (1991) (http://www.ch.embnet.org/software/COILS_form.html), using the weighted option. The horizontal axis represents the primary sequence of each protein, and thevertical axis represents the probability of forming a-helical coiled coils. Both gH and gB are predicted to have an a-helicalcoiled-coil domain with a high probability score at the N terminus and a second a-helical coiled-coil domain towards the Cterminus with a lower probability score. (b) Vero cells were incubated with increasing concentrations of peptides (10, 100,250 and 500 mM) in the presence of the viral inoculum for 45 min at 37 6C. Non-penetrated virus was inactivated and cellsincubated for 24 h at 37 6C in DMEM supplemented with CMC. Plaque numbers were scored and the percentage ofinhibition was calculated with respect to no-peptide control experiments. Data are reported in triplicate and error barsrepresent standard deviations.
1088 Journal of General Virology 87
S. Galdiero and others
The effects of gB peptides were much less pronounced: theshorter and longer versions of B-HR1 showed minor inhibi-tory effects at concentrations of 500 mM and both B-HR2and B-HR2-S peptides failed to have any effect on inhibitingHSV-1 infectivity (Fig. 1b). The inhibitory effects of gH andgB peptides were specific in that scrambled versions of thesepeptides exerted no inhibition (Fig. 1b).
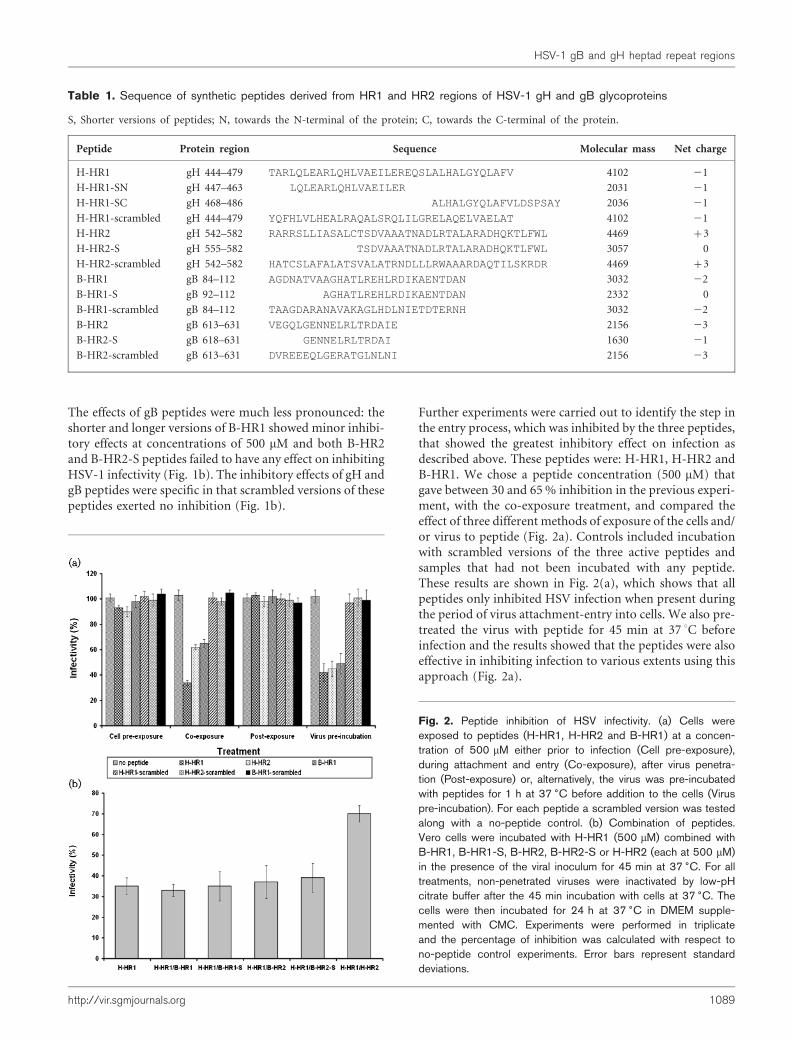
Further experiments were carried out to identify the step inthe entry process, which was inhibited by the three peptides,that showed the greatest inhibitory effect on infection asdescribed above. These peptides were: H-HR1, H-HR2 andB-HR1. We chose a peptide concentration (500 mM) thatgave between 30 and 65% inhibition in the previous experi-ment, with the co-exposure treatment, and compared theeffect of three different methods of exposure of the cells and/or virus to peptide (Fig. 2a). Controls included incubationwith scrambled versions of the three active peptides andsamples that had not been incubated with any peptide.These results are shown in Fig. 2(a), which shows that allpeptides only inhibited HSV infection when present duringthe period of virus attachment-entry into cells. We also pre-treated the virus with peptide for 45 min at 37 uC beforeinfection and the results showed that the peptides were alsoeffective in inhibiting infection to various extents using thisapproach (Fig. 2a).
Table 1. Sequence of synthetic peptides derived from HR1 and HR2 regions of HSV-1 gH and gB glycoproteins
S, Shorter versions of peptides; N, towards the N-terminal of the protein; C, towards the C-terminal of the protein.
Peptide Protein region Sequence Molecular mass Net charge
H-HR1 gH 444–479 TARLQLEARLQHLVAEILEREQSLALHALGYQLAFV 4102 21
H-HR1-SN gH 447–463 LQLEARLQHLVAEILER 2031 21
H-HR1-SC gH 468–486 ALHALGYQLAFVLDSPSAY 2036 21
H-HR1-scrambled gH 444–479 YQFHLVLHEALRAQALSRQLILGRELAQELVAELAT 4102 21
H-HR2 gH 542–582 RARRSLLIASALCTSDVAAATNADLRTALARADHQKTLFWL 4469 +3
H-HR2-S gH 555–582 TSDVAAATNADLRTALARADHQKTLFWL 3057 0
H-HR2-scrambled gH 542–582 HATCSLAFALATSVALATRNDLLLRWAAARDAQTILSKRDR 4469 +3
B-HR1 gB 84–112 AGDNATVAAGHATLREHLRDIKAENTDAN 3032 22
B-HR1-S gB 92–112 AGHATLREHLRDIKAENTDAN 2332 0
B-HR1-scrambled gB 84–112 TAAGDARANAVAKAGLHDLNIETDTERNH 3032 22
B-HR2 gB 613–631 VEGQLGENNELRLTRDAIE 2156 23
B-HR2-S gB 618–631 GENNELRLTRDAI 1630 21
B-HR2-scrambled gB 613–631 DVREEEQLGERATGLNLNI 2156 23
Fig. 2. Peptide inhibition of HSV infectivity. (a) Cells wereexposed to peptides (H-HR1, H-HR2 and B-HR1) at a concen-tration of 500 mM either prior to infection (Cell pre-exposure),during attachment and entry (Co-exposure), after virus penetra-tion (Post-exposure) or, alternatively, the virus was pre-incubatedwith peptides for 1 h at 37 6C before addition to the cells (Viruspre-incubation). For each peptide a scrambled version was testedalong with a no-peptide control. (b) Combination of peptides.Vero cells were incubated with H-HR1 (500 mM) combined withB-HR1, B-HR1-S, B-HR2, B-HR2-S or H-HR2 (each at 500 mM)in the presence of the viral inoculum for 45 min at 37 6C. For alltreatments, non-penetrated viruses were inactivated by low-pHcitrate buffer after the 45 min incubation with cells at 37 6C. Thecells were then incubated for 24 h at 37 6C in DMEM supple-mented with CMC. Experiments were performed in triplicateand the percentage of inhibition was calculated with respect tono-peptide control experiments. Error bars represent standarddeviations.
http://vir.sgmjournals.org 1089
HSV-1 gB and gH heptad repeat regions
We also analysed the possible interaction between differentHSV-1 gB and gH HRs by co-exposure of virus and cellswith combinations of different peptides to determinewhether the inhibitory effect seen with H-HR1 could bereduced in the presence of other HR regions. The rationalebehind this experiment was that, if an inhibitory peptideaggregates with a peptide from another HR domain suchthat they create a structure that could resemble the six-helixbundle of HIV gp41, it would be less efficient at exerting aninhibitory effect on infectivity. The combinations of pep-tides tested and the results obtained are shown in Fig. 2(b).None of the peptides derived from gB affected the inhibitoryeffect exerted by H-HR1 alone. However, when H-HR2 wasincubated together with H-HR1, we observed a significantreduction of the inhibitory activity to approximately 30%from values of approximately 65% inhibition. This observa-tion lends weight to the view that HR1 and HR2 regions ofHSV-1 gH may interact with each other during the fusionprocess.
Structural analysis of HR peptides by CD
Biophysical approaches were employed to characterizefurther the structure of these peptides and to determinewhether any evidence of interaction could be observed.
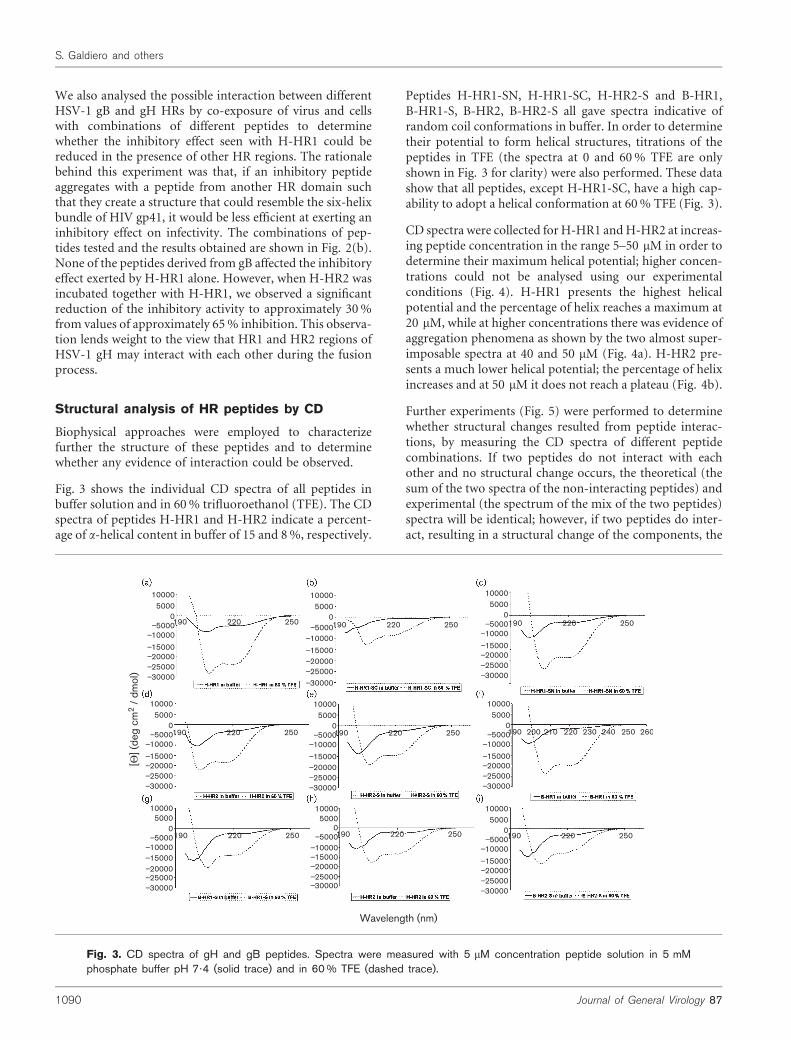
Fig. 3 shows the individual CD spectra of all peptides inbuffer solution and in 60% trifluoroethanol (TFE). The CDspectra of peptides H-HR1 and H-HR2 indicate a percent-age of a-helical content in buffer of 15 and 8%, respectively.
Peptides H-HR1-SN, H-HR1-SC, H-HR2-S and B-HR1,B-HR1-S, B-HR2, B-HR2-S all gave spectra indicative ofrandom coil conformations in buffer. In order to determinetheir potential to form helical structures, titrations of thepeptides in TFE (the spectra at 0 and 60% TFE are onlyshown in Fig. 3 for clarity) were also performed. These datashow that all peptides, except H-HR1-SC, have a high cap-ability to adopt a helical conformation at 60% TFE (Fig. 3).
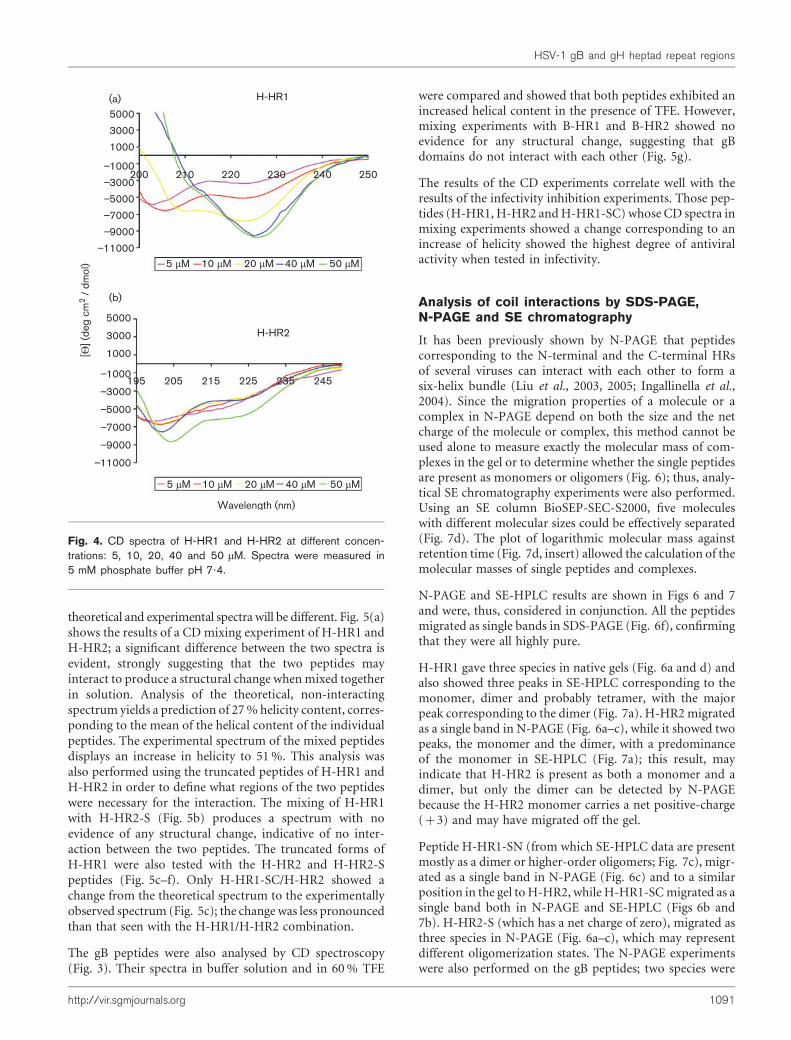
CD spectra were collected for H-HR1 andH-HR2 at increas-ing peptide concentration in the range 5–50 mM in order todetermine their maximum helical potential; higher concen-trations could not be analysed using our experimentalconditions (Fig. 4). H-HR1 presents the highest helicalpotential and the percentage of helix reaches a maximum at20 mM, while at higher concentrations there was evidence ofaggregation phenomena as shown by the two almost super-imposable spectra at 40 and 50 mM (Fig. 4a). H-HR2 pre-sents a much lower helical potential; the percentage of helixincreases and at 50 mM it does not reach a plateau (Fig. 4b).
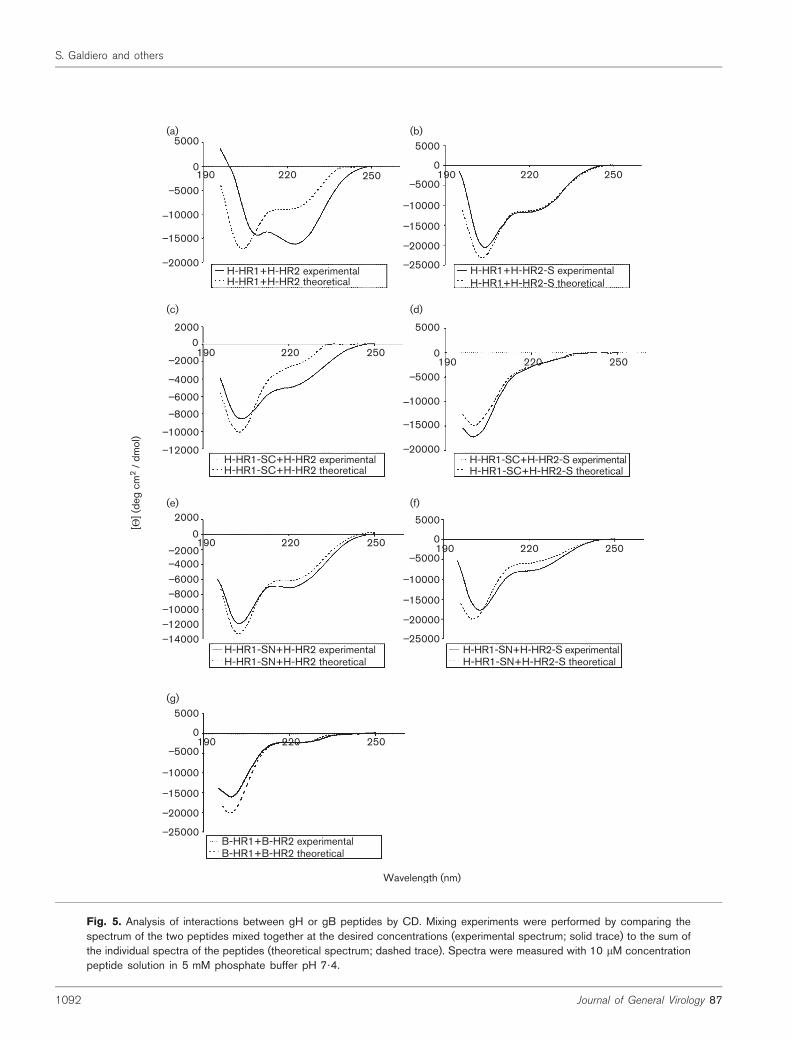
Further experiments (Fig. 5) were performed to determinewhether structural changes resulted from peptide interac-tions, by measuring the CD spectra of different peptidecombinations. If two peptides do not interact with eachother and no structural change occurs, the theoretical (thesum of the two spectra of the non-interacting peptides) andexperimental (the spectrum of the mix of the two peptides)spectra will be identical; however, if two peptides do inter-act, resulting in a structural change of the components, the
10000
0
_10000
5000
_15000_20000
_30000
_25000
_5000
10000
0
_10000
5000
_15000_20000
_30000
_25000
_5000
10000
0
_10000
5000
_15000_20000
_30000
_25000
_5000
10000
0
_10000
5000
_15000_20000
_30000
_25000
_5000
10000
0
_10000
5000
_15000_20000
_30000
_25000
_5000
10000
0
_10000
5000
_15000_20000
_30000
_25000
_5000
10000
0
_10000
5000
_15000_20000
_30000
_25000
_5000
10000
0
_10000
5000
_15000_20000
_30000
_25000
_5000
10000
0
_10000
5000
_15000_20000
_30000
_25000
_5000
190 220 250 190 220 250 190 220 250
190 220 250
190 220 250190 220 250
190 220 250 190 200 210 220 230 240 250 260
Wavelength (nm)
[Θ] (
deg
cm2
/ dm
ol)
190 220 250
Fig. 3. CD spectra of gH and gB peptides. Spectra were measured with 5 mM concentration peptide solution in 5 mMphosphate buffer pH 7?4 (solid trace) and in 60 % TFE (dashed trace).
1090 Journal of General Virology 87
S. Galdiero and others
theoretical and experimental spectra will be different. Fig. 5(a)shows the results of a CD mixing experiment of H-HR1 andH-HR2; a significant difference between the two spectra isevident, strongly suggesting that the two peptides mayinteract to produce a structural change whenmixed togetherin solution. Analysis of the theoretical, non-interactingspectrum yields a prediction of 27% helicity content, corres-ponding to the mean of the helical content of the individualpeptides. The experimental spectrum of the mixed peptidesdisplays an increase in helicity to 51%. This analysis wasalso performed using the truncated peptides of H-HR1 andH-HR2 in order to define what regions of the two peptideswere necessary for the interaction. The mixing of H-HR1with H-HR2-S (Fig. 5b) produces a spectrum with noevidence of any structural change, indicative of no inter-action between the two peptides. The truncated forms ofH-HR1 were also tested with the H-HR2 and H-HR2-Speptides (Fig. 5c–f). Only H-HR1-SC/H-HR2 showed achange from the theoretical spectrum to the experimentallyobserved spectrum (Fig. 5c); the change was less pronouncedthan that seen with the H-HR1/H-HR2 combination.
The gB peptides were also analysed by CD spectroscopy(Fig. 3). Their spectra in buffer solution and in 60% TFE
were compared and showed that both peptides exhibited anincreased helical content in the presence of TFE. However,mixing experiments with B-HR1 and B-HR2 showed noevidence for any structural change, suggesting that gBdomains do not interact with each other (Fig. 5g).
The results of the CD experiments correlate well with theresults of the infectivity inhibition experiments. Those pep-tides (H-HR1, H-HR2 andH-HR1-SC) whose CD spectra inmixing experiments showed a change corresponding to anincrease of helicity showed the highest degree of antiviralactivity when tested in infectivity.
Analysis of coil interactions by SDS-PAGE,N-PAGE and SE chromatography
It has been previously shown by N-PAGE that peptidescorresponding to the N-terminal and the C-terminal HRsof several viruses can interact with each other to form asix-helix bundle (Liu et al., 2003, 2005; Ingallinella et al.,2004). Since the migration properties of a molecule or acomplex in N-PAGE depend on both the size and the netcharge of the molecule or complex, this method cannot beused alone to measure exactly the molecular mass of com-plexes in the gel or to determine whether the single peptidesare present as monomers or oligomers (Fig. 6); thus, analy-tical SE chromatography experiments were also performed.Using an SE column BioSEP-SEC-S2000, five moleculeswith different molecular sizes could be effectively separated(Fig. 7d). The plot of logarithmic molecular mass againstretention time (Fig. 7d, insert) allowed the calculation of themolecular masses of single peptides and complexes.
N-PAGE and SE-HPLC results are shown in Figs 6 and 7and were, thus, considered in conjunction. All the peptidesmigrated as single bands in SDS-PAGE (Fig. 6f), confirmingthat they were all highly pure.
H-HR1 gave three species in native gels (Fig. 6a and d) andalso showed three peaks in SE-HPLC corresponding to themonomer, dimer and probably tetramer, with the majorpeak corresponding to the dimer (Fig. 7a). H-HR2migratedas a single band in N-PAGE (Fig. 6a–c), while it showed twopeaks, the monomer and the dimer, with a predominanceof the monomer in SE-HPLC (Fig. 7a); this result, mayindicate that H-HR2 is present as both a monomer and adimer, but only the dimer can be detected by N-PAGEbecause the H-HR2 monomer carries a net positive-charge(+3) and may have migrated off the gel.
Peptide H-HR1-SN (from which SE-HPLC data are presentmostly as a dimer or higher-order oligomers; Fig. 7c), migr-ated as a single band in N-PAGE (Fig. 6c) and to a similarposition in the gel to H-HR2, while H-HR1-SCmigrated as asingle band both in N-PAGE and SE-HPLC (Figs 6b and7b). H-HR2-S (which has a net charge of zero), migrated asthree species in N-PAGE (Fig. 6a–c), which may representdifferent oligomerization states. The N-PAGE experimentswere also performed on the gB peptides; two species were
H-HR1
5 mM 10 mM 20 mM 40 mM 50 mM
195 205 215 225 235 245
200 210 220 230 240 250
(a)5000
3000
1000_1000_3000_5000_7000_9000
_11000
5000
3000
1000_1000_3000_5000_7000_9000
_11000
H-HR2
Wavelength (nm)
[Θ] (
deg
cm2
/ dm
ol)
(b)
5 mM 10 mM 20 mM 40 mM 50 mM
Fig. 4. CD spectra of H-HR1 and H-HR2 at different concen-trations: 5, 10, 20, 40 and 50 mM. Spectra were measured in5 mM phosphate buffer pH 7?4.
http://vir.sgmjournals.org 1091
HSV-1 gB and gH heptad repeat regions
5000
_5000
_10000
_15000
_20000
0
20000
_2000_4000_6000_8000
_10000_12000
5000
_5000
_10000_15000_20000
0
_25000
190 220 250 190 220 250
190 220 250
H-HR1+H-HR2 experimentalH-HR1+H-HR2 theoretical
H-HR1+H-HR2-S experimentalH-HR1+H-HR2-S theoretical
H-HR1-SC+H-HR2 experimentalH-HR1-SC+H-HR2 theoretical
(a) (b)
(c)
5000
_5000
_10000
_15000
_20000
0190 220 250
(d)
H-HR1-SC+H-HR2-S experimentalH-HR1-SC+H-HR2-S theoretical
2000
0_2000_4000_6000_8000
_10000_12000_14000
190 220 250
H-HR1-SN+H-HR2 experimentalH-HR1-SN+H-HR2 theoretical
(e)
5000
_5000
_10000_15000_20000
0
_25000
190 220 250
H-HR1-SN+H-HR2-S experimentalH-HR1-SN+H-HR2-S theoretical
(f)
5000
_5000
_10000_15000_20000
0
_25000
190 220 250
(g)
B-HR1+B-HR2 experimentalB-HR1+B-HR2 theoretical
Wavelength (nm)
[Θ] (
deg
cm2
/ dm
ol)
Fig. 5. Analysis of interactions between gH or gB peptides by CD. Mixing experiments were performed by comparing thespectrum of the two peptides mixed together at the desired concentrations (experimental spectrum; solid trace) to the sum ofthe individual spectra of the peptides (theoretical spectrum; dashed trace). Spectra were measured with 10 mM concentrationpeptide solution in 5 mM phosphate buffer pH 7?4.
1092 Journal of General Virology 87
S. Galdiero and others
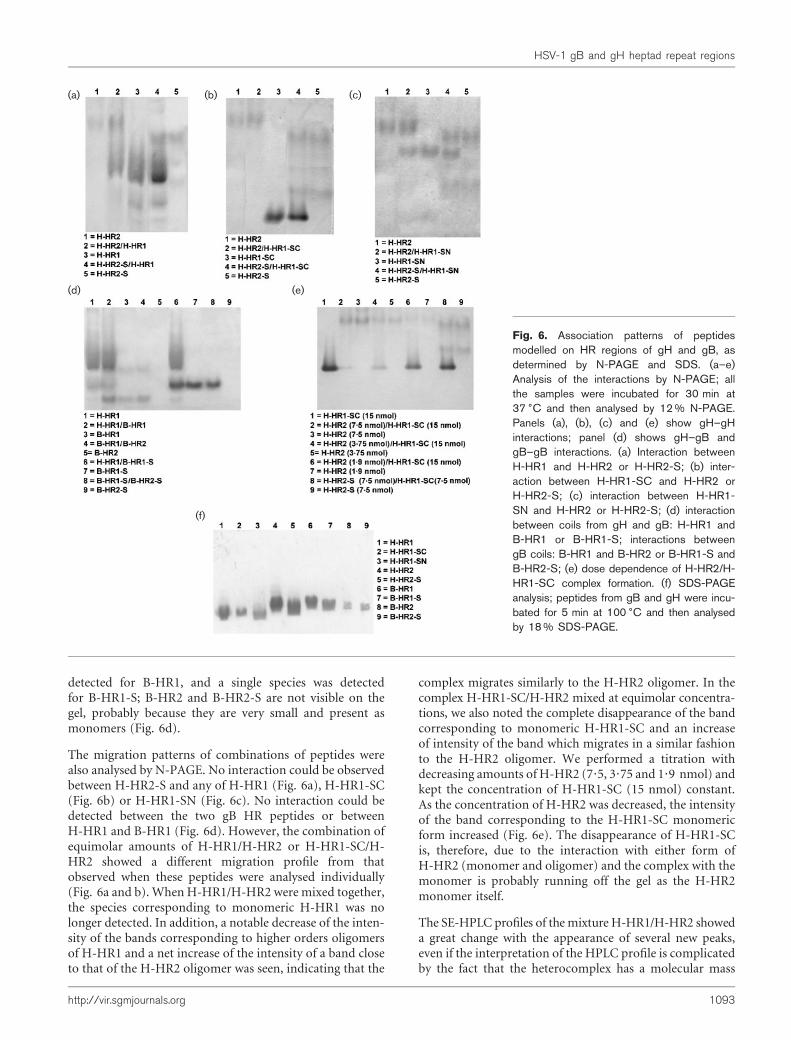
detected for B-HR1, and a single species was detectedfor B-HR1-S; B-HR2 and B-HR2-S are not visible on thegel, probably because they are very small and present asmonomers (Fig. 6d).
The migration patterns of combinations of peptides werealso analysed by N-PAGE. No interaction could be observedbetween H-HR2-S and any of H-HR1 (Fig. 6a), H-HR1-SC(Fig. 6b) or H-HR1-SN (Fig. 6c). No interaction could bedetected between the two gB HR peptides or betweenH-HR1 and B-HR1 (Fig. 6d). However, the combination ofequimolar amounts of H-HR1/H-HR2 or H-HR1-SC/H-HR2 showed a different migration profile from thatobserved when these peptides were analysed individually(Fig. 6a and b). When H-HR1/H-HR2 were mixed together,the species corresponding to monomeric H-HR1 was nolonger detected. In addition, a notable decrease of the inten-sity of the bands corresponding to higher orders oligomersof H-HR1 and a net increase of the intensity of a band closeto that of the H-HR2 oligomer was seen, indicating that the
complex migrates similarly to the H-HR2 oligomer. In thecomplex H-HR1-SC/H-HR2 mixed at equimolar concentra-tions, we also noted the complete disappearance of the bandcorresponding to monomeric H-HR1-SC and an increaseof intensity of the band which migrates in a similar fashionto the H-HR2 oligomer. We performed a titration withdecreasing amounts of H-HR2 (7?5, 3?75 and 1?9 nmol) andkept the concentration of H-HR1-SC (15 nmol) constant.As the concentration of H-HR2 was decreased, the intensityof the band corresponding to the H-HR1-SC monomericform increased (Fig. 6e). The disappearance of H-HR1-SCis, therefore, due to the interaction with either form ofH-HR2 (monomer and oligomer) and the complex with themonomer is probably running off the gel as the H-HR2monomer itself.
The SE-HPLC profiles of themixture H-HR1/H-HR2 showeda great change with the appearance of several new peaks,even if the interpretation of the HPLC profile is complicatedby the fact that the heterocomplex has a molecular mass
(a) (b) (c)
(d) (e)
(f)
Fig. 6. Association patterns of peptidesmodelled on HR regions of gH and gB, asdetermined by N-PAGE and SDS. (a–e)Analysis of the interactions by N-PAGE; allthe samples were incubated for 30 min at37 6C and then analysed by 12 % N-PAGE.Panels (a), (b), (c) and (e) show gH–gHinteractions; panel (d) shows gH–gB andgB–gB interactions. (a) Interaction betweenH-HR1 and H-HR2 or H-HR2-S; (b) inter-action between H-HR1-SC and H-HR2 orH-HR2-S; (c) interaction between H-HR1-SN and H-HR2 or H-HR2-S; (d) interactionbetween coils from gH and gB: H-HR1 andB-HR1 or B-HR1-S; interactions betweengB coils: B-HR1 and B-HR2 or B-HR1-S andB-HR2-S; (e) dose dependence of H-HR2/H-HR1-SC complex formation. (f) SDS-PAGEanalysis; peptides from gB and gH were incu-bated for 5 min at 100 6C and then analysedby 18 % SDS-PAGE.
http://vir.sgmjournals.org 1093
HSV-1 gB and gH heptad repeat regions
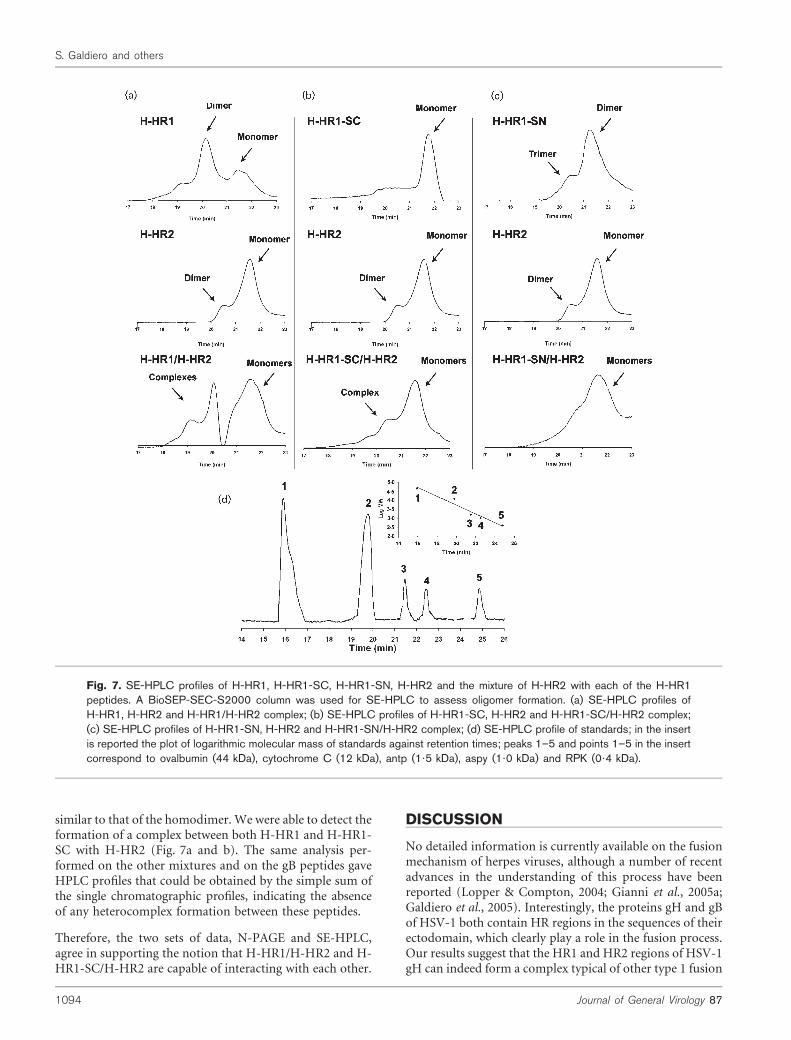
similar to that of the homodimer. We were able to detect theformation of a complex between both H-HR1 and H-HR1-SC with H-HR2 (Fig. 7a and b). The same analysis per-formed on the other mixtures and on the gB peptides gaveHPLC profiles that could be obtained by the simple sum ofthe single chromatographic profiles, indicating the absenceof any heterocomplex formation between these peptides.
Therefore, the two sets of data, N-PAGE and SE-HPLC,agree in supporting the notion that H-HR1/H-HR2 and H-HR1-SC/H-HR2 are capable of interacting with each other.
DISCUSSION
No detailed information is currently available on the fusionmechanism of herpes viruses, although a number of recentadvances in the understanding of this process have beenreported (Lopper & Compton, 2004; Gianni et al., 2005a;Galdiero et al., 2005). Interestingly, the proteins gH and gBof HSV-1 both contain HR regions in the sequences of theirectodomain, which clearly play a role in the fusion process.Our results suggest that the HR1 and HR2 regions of HSV-1gH can indeed form a complex typical of other type 1 fusion
Fig. 7. SE-HPLC profiles of H-HR1, H-HR1-SC, H-HR1-SN, H-HR2 and the mixture of H-HR2 with each of the H-HR1peptides. A BioSEP-SEC-S2000 column was used for SE-HPLC to assess oligomer formation. (a) SE-HPLC profiles ofH-HR1, H-HR2 and H-HR1/H-HR2 complex; (b) SE-HPLC profiles of H-HR1-SC, H-HR2 and H-HR1-SC/H-HR2 complex;(c) SE-HPLC profiles of H-HR1-SN, H-HR2 and H-HR1-SN/H-HR2 complex; (d) SE-HPLC profile of standards; in the insertis reported the plot of logarithmic molecular mass of standards against retention times; peaks 1–5 and points 1–5 in the insertcorrespond to ovalbumin (44 kDa), cytochrome C (12 kDa), antp (1?5 kDa), aspy (1?0 kDa) and RPK (0?4 kDa).
1094 Journal of General Virology 87
S. Galdiero and others

proteins and they function as inhibitors of infection, whilethe HR1 and HR2 regions of gB show lower or absent inhi-bitory activity and no evidence of complex formation, atleast with the techniques used in this study, namely SE-HPLC, CD analysis and N-PAGE.
Predicted coiled-coil regions were identified in both gH andgB, and peptides of different lengths were generated andtested for their effects on virus entry. gH coiled-coil peptidesinhibited HSV-1 infection in a dose-dependent, sequence-specific manner. H-HR1 showed the strongest activity andwe also observed that longer coiled-coil peptides were morepotent inhibitors of virus entry, probably due to increasedability to form more stable secondary structures than theshorter peptides. This observation was confirmed by CDdata that indicated that H-HR1 has a significant helicalcontent in 5 mM buffer solution, which increased uponincreasing peptide concentrations or upon addition of TFE(Figs 3 and 4). Treatment of cells with peptides (H-HR1, H-HR2 and B-HR1) following incubation with virus had noeffect on entry, indicating that the inhibitory peptides actat a step during virus entry and have no effect once the virushas penetrated the cell membrane. Pre-treatment of cellswith peptides, followed by their removal by acid wash, alsodid not impair the ability of HSV-1 to enter cells, suggestingthat the peptides do not irreversibly condition the cellsusceptibility to the virus. Efficient inhibition was observedonly when the gH or gB peptides were incubated with virusat the same time as inoculation into Vero cells, or when thevirus was pre-incubated with peptides for 45 min beforebeing inoculated into the cells. These results are consistentwith previous reports (Lopper & Compton, 2004; Gianniet al., 2005b) and show that the domain with the highestprobability score to form coils, namely the HR1 of gH, is thepeptide which exerts the strongest inhibitory effect.
The interesting observation resulting from these studies wasthat not only was H-HR1 (which showed a high probabilityof forming coiled-coils) active in inhibiting virus entry, butH-HR2 and B-HR1 (which have a low probability score)also were able to inhibit virus entry, albeit with lowerefficacy.
This finding, together with the fact that co-exposure of cellswith a mix of H-HR1 and H-HR2 was able to reduce thelevel of inhibition obtained with H-HR1 alone, prompted usto analyse in further detail the nature of any interactionsbetween coiled-coil sequences of both gH and gB. Whilemost enveloped viruses need a single fusion protein and thetwo interacting HRs are present on the same glycoprotein,we do not know if herpes viruses, which require four differ-ent glycoproteins (Turner et al., 1998) to mediate fusion,established intramolecular contacts between different regionsof the same protein or perhaps intermolecular interactionsbetween gH and gB. The first line of evidence pointingtowards an intramolecular interaction between the twoHRs of gH was the reduction of the inhibitory effect whenwe performed infectivity assays with the mix of peptides. Infact, the only combination of peptides able to reduce the
effect of H-HR1 on infection was that of H-HR1/H-HR2.No reduction was observed with any of the gB peptides. Thisfinding can be explained by the possible sequestration ofH-HR1 peptides. H-HR1 is able to inhibit infection whenfree in the suspension, but unable to inhibit once H-HR2 ispresent.
The results of biophysical analyses were consistent with theexistence of an intramolecular interaction between H-HR1and H-HR2 of gH and would suggest that the HRs of gB andgH do not interact with each other to form intermolecularinteractions. However, it remains a possibility that gB andgH do interact during fusion, but that these interactionsinvolve other domains of these glycoproteins.
It is interesting to note that, in most viral fusion models,synthetic peptides corresponding to the C-terminal HR aregenerally more active in inhibition assays if compared withpeptides corresponding to the N-terminal HR; this is incontrast with our results on gH and gB of HSV-1 and withprevious data on herpesvirus glycoproteins, which onlyanalysed N-terminal HRs due to the higher probability ofthose regions to form coiled-coils. Nevertheless, our resultsindicate that C-terminal HRs in HSV-1 gH can also slightlyinhibit fusion and can interact with the N-terminal part ofthe protein. The only other known case of HR1 domainsbeing more potent than HR2 regions is that of SARS-CoV(Yuan et al., 2004).
In summary, the presence in HSV-1 gH of a putativecanonical fusion peptide (Gianni et al., 2005a), severalmembrane interacting domains capable of fusing artificialmembranes (Galdiero et al., 2005), and both an N-terminaland a C-terminal HR region potentially capable of formingcoiled-coils that clearly interact with each other, demon-strates that gH plays a critical role in membrane fusionmediated by HSV-1. The finding of domains in gH and gBHR1 and HR2 supports the assumption that the HSV fusionmechanism may employ a strategy similar to other class Ienvelope fusion proteins. Further studies are needed tounravel the role of gB, since HR regions are present in thisprotein as well, and the N-terminal HR domain is alsopartially effective in reducing HSV-1 infectivity.
ACKNOWLEDGEMENTS
We gratefully acknowledge support by EU under contract no. QLK2-CT-2002-00810.
REFERENCES
Baker, K. A., Dutch, R. E., Lamb, R. A. & Jardetzky, T. S. (1999).Structural basis for paramyxovirus-mediated membrane fusion.Mol Cell 3, 309–319.
Blumenthal, R., Clague, M. J., Durell, S. R. & Epand, R. M. (2003).Membrane fusion. Chem Rev 103, 53–69.
Bullough, P. A., Hughson, F. M., Skehel, J. J. & Wiley, D. C. (1994).Structure of influenza haemagglutinin at the pH of membranefusion. Nature 371, 37–43.
http://vir.sgmjournals.org 1095
HSV-1 gB and gH heptad repeat regions

Caffrey, M., Cai, M., Kaufman, J., Stahl, S. J., Wingfield, P. T.,Covell, D. G., Gronenenborn, A. M. & Clore, G. M. (1998). Three-dimensional structure of the 44 kDa ectodomain of SIV gp41.
EMBO J 17, 4572–4584.
Chambers, P., Pringle, C. R. & Easton, A. J. (1990). Heptad repeatsequences are located adjacent to hydrophobic regions in several
types of virus fusion glycoproteins. J Gen Virol 71, 3075–3080.
Chan, D. C. & Kim, P. S. (1998). HIV entry and its inhibition. Cell
93, 681–684.
Chen, J., Skehel, J. J. & Wiley, D. C. (1999). N- and C-terminalresidues combine in the fusion-pH influenza hemagglutinin HA2
subunit to form an N cap that terminates the triple-stranded coiledcoil. Proc Natl Acad Sci U S A 96, 8967–8972.
Cocchi, F., Menotti, L., Mirandola, P., Lopez, M. & Campadelli-Fiume, G. (1998). The ectodomain of a novel member of the
immunoglobulin superfamily related to the poliovirus receptor hasthe attributes of a bona fide receptor for herpes simplex virus types 1
and 2 in human cells. J Virol 72, 9992–10002.
Cocchi, F., Fusco, D., Menotti, L., Gianni, T., Eisenberg, R. J., Cohen,G. H. & Campadelli-Fiume, G. (2004). The soluble ectodomain of
herpes simplex virus gD contains a membrane-proximal pro-fusiondomain and suffices to mediate virus entry. Proc Natl Acad Sci U S A
101, 7445–7450.
Dimitrov, D. S. (2004). Virus entry: molecular mechanisms and
biomedical applications. Nat Rev Microbiol 2, 109–122.
Earp, L. J., Delos, S. E., Park, H. E. & White, J. M. (2005). The manymechanisms of viral membrane fusion proteins. Curr Top Microbiol
Immunol 285, 25–66.
Eckert, D. M. & Kim, P. S. (2001). Mechanism of viral membrane
fusion and its inhibition. Annu Rev Biochem 70, 777–810.
Fass, D., Harrison, S. C. & Kim, P. S. (1996). Retrovirus envelopedomain at 1?7 angstrom resolution. Nat Struct Biol 3, 465–469.
Forrester, A., Farrell, H., Wilkinson, G., Kaye, J., Davis-Poynter, N. &Minson, T. (1992). Construction and properties of a mutant of
herpes simplex virus type 1 with glycoprotein H coding sequencesdeleted. J Virol 66, 341–348.
Galdiero, S., Falanga, A., Vitiello, M., Browne, H., Pedone, C. &Galdiero, M. (2005). Fusogenic domains in herpes simplex virustype 1 glycoprotein H. J Biol Chem 280, 28632–28643.
Geraghty, R. J., Krummenacher, C., Cohen, G. H., Eisenberg, R. J. &Spear, P. G. (1998). Entry of alphaherpesviruses mediated by
poliovirus receptor-related protein 1 and poliovirus receptor. Science280, 1618–1620.
Gianni, T., Martelli, P. L., Casadio, R. & Campadelli-Fiume, G.(2005a). The ectodomain of herpes simplex virus glycoprotein Hcontains a membrane a-helix with attributes of an internal fusion
peptide, positionally conserved in the Herpesviridae family. J Virol79, 2931–2940.
Gianni, T., Menotti, L. & Campadelli-Fiume, G. (2005b). A heptadrepeat in herpes simplex virus 1 gH, located downstream of the
a-helix with attributes of a fusion peptide, is critical for virus entryand fusion. J Virol 79, 7042–7049.
Heldwein, K., Lou, H., Whitbeck, J. C., Cohen, G., Eisenberg, R. J. &Harrison, S. (2005). International Herpesvirus Workshop, Turku,Finland.
Hernandez, L. D., Hoffman, L. R., Wolfsberg, T. G. & White, J. M. (1996).Virus-cell and cell-cell fusion. Annu Rev Cell Dev Biol 12, 627–661.
Ingallinella, P., Bianchi, E., Finotto, M., Cantoni, G., Eckert, D. M.,Supekar, V. M., Bruckmann, C., Carfi, A. & Pessi, A. (2004).Structural characterization of the fusion-active complex of severe
acute respiratory syndrome (SARS) coronavirus. Proc Natl Acad SciU S A 101, 8709–8714.
Jahn, R., Lang, T. & Sudhof, T. C. (2003). Membrane fusion. Cell 112,
519–533.
Jardetzky, T. S. & Lamb, R. A. (2004). Virology: a class act. Nature
427, 307–308.
Joshi, S. B., Dutch, R. E. & Lamb, R. A. (1998). A core trimer of the
paramyxovirus fusion protein: parallels to influenza virus hemag-
glutinin and HIV-1 gp41. Virology 248, 20–34.
Kilby, J. M., Hopkins, S., Venetta, T. M. & 12 other authors (1998).Potent suppression of HIV-1 replication in humans by T-20, a pep-
tide inhibitor of gp41-mediated virus entry. Nat Med 4, 1302–1307.
Kobe, B., Center, R. J., Kemp, B. E. & Poumbourios, P. (1999).Crystal structure of human T cell leukemia virus type 1 gp21
ectodomain crystallized as a maltose-binding protein chimera reveals
structural evolution of retroviral transmembrane proteins. Proc Natl
Acad Sci U S A 96, 4319–4324.
Lambert, D. M., Barney, S., Lambert, A. L. & 7 other authors (1996).
Peptides from conserved regions of paramyxovirus fusion (F) pro-
teins are potent inhibitors of viral fusion. Proc Natl Acad Sci U S A
93, 2186–2191.
Liu, S., Zhao, Q. & Jiang, S. (2003). Determination of the HIV-1 gp41
fusogenic core conformation modelled by synthetic peptides: applicable
for identification of HIV-1 fusion inhibitors. Peptides 24, 1303–1313.
Liu, S., Lu, H., Niu, J., Xu, Y., Wu, S. & Jiang, S. (2005). Different fromthe HIV fusion inhibitor C34, the anti-HIV drug Fuzeon (T-20) inhibits
HIV-1 entry by targeting multiple sites in gp41 and gp120. J Biol Chem
280, 11259–11273.
Lopez, M., Cocchi, F., Menotti, L., Avitabile, E., Dubreuil, P. &
Campadelli-Fiume, G. (2000). Nectin2alpha (PRR2alpha or HveB)
and nectin2delta are low-efficiency mediators for entry of herpes
simplex virus mutants carrying the Leu25Pro substitution in glyco-
protein D. J Virol 74, 1267–1274.
Lopper, M. & Compton, T. (2004). Coiled-coil domains in glyco-
proteins B and H are involved in human cytomegalovirus membrane
fusion. J Virol 78, 8333–8341.
Lupas, A., Van Dyke, M. & Stock, J. (1991). Predicting coiled coils
from protein sequences. Science 252, 1162–1164.
Malashkevich, V. N., Chan, D. C., Chutkowski, C. T. & Kim, P. S.(1998). Crystal structure of the simian immunodeficiency virus (SIV)
gp41 core: conserved helical interactions underlie the broad inhibitory
activity of gp41 peptides. Proc Natl Acad Sci U S A 95, 9134–9139.
Malashkevich, V. N., Schneider, B. J., McNally, M. L., Milhollen, M. A.,
Pang, J. X. & Kim, P. S. (1999). Core structure of the envelope
glycoprotein GP2 from Ebola virus at 1.9-A resolution. Proc Natl
Acad Sci U S A 96, 2662–2667.
Malashkevich, V. N., Singh, M. & Kim, P. S. (2001). The trimer-of-
hairpins motif in membrane fusion: visna virus. Proc Natl Acad Sci
U S A 98, 8502–8506.
Melikyan, G. B., Markosyan, R. M., Hemmati, H., Delmedico, M. K.,
Lambert, D. M. & Cohen, F. S. (2000). Evidence that the transition of
HIV-1 gp41 into a six-helix bundle, not the bundle configuration,
induces membrane fusion. J Cell Biol 151, 413–423.
Mettenleiter, T. C. (2002a). Brief overview on cellular virus receptors.
Virus Res 82, 3–8.
Mettenleiter, T. C. (2002b). Herpesvirus assembly and egress. J Virol
76, 1537–1547.
Montgomery, R. I., Warner, M. S., Lum, B. J. & Spear, P. G. (1996).
Herpes simplex virus-1 entry into cells mediated by a novel member
of the TNF/NGF receptor family. Cell 87, 427–436.
Okazaki, K. & Kida, H. (2004). A synthetic peptide from a heptad
repeat region of herpesvirus glycoprotein B inhibits virus replication.
J Gen Virol 85, 2131–2137.
1096 Journal of General Virology 87
S. Galdiero and others

Rapaport, D., Ovadia, M. & Shai, Y. (1995). A synthetic peptidecorresponding to a conserved heptad repeat domain is a potentinhibitor of Sendai virus-cell fusion: an emerging similarity withfunctional domains of other viruses. EMBO J 14, 5524–5531.
Russell, C. J., Jardetzky, T. S. & Lamb, R. A. (2001). Membranefusion machines of paramyxoviruses: capture of intermediates offusion. EMBO J 20, 4024–4034.
Skehel, J. J. & Wiley, D. C. (1998). Coiled coils in both intracellularvesicle and viral membrane fusion. Cell 95, 871–874.
Spear, P. G. (2004). Herpes simplex virus: receptors and ligands forcell entry. Cell Microbiol 6, 401–410.
Spear, P. G. & Longnecker, R. (2003). Herpesvirus entry: an update.J Virol 77, 10179–10185.
Turner, A., Bruun, B., Minson, T. & Browne, H. (1998). GlycoproteinsgB, gD, and gHgL of herpes simplex virus type 1 are necessary andsufficient to mediate membrane fusion in a Cos cell transfectionsystem. J Virol 72, 873–875.
Warner, M. S., Geraghty, R. J., Martinez, W. M., Montgomery, R. I.,Whitbeck, J. C., Xu, R., Eisenberg, R. J., Cohen, G. H. & Spear, P. G.(1998). A cell surface protein with herpesvirus entry activity (HveB)confers susceptibility to infection by mutants of herpes simplex virustype 1, herpes simplex virus type 2, and pseudorabies virus. Virology246, 179–189.
Weissenhorn, W., Dessen, A., Harrison, S. C., Skehel, J. J. & Wiley,D. C. (1997). Atomic structure of the ectodomain from HIV-1 gp41.Nature 387, 426–430.
Weissenhorn, W., Calder, L. J., Wharton, S. A., Skehel, J. J. & Wiley,D. C. (1998). The central structural feature of the membrane fusionprotein subunit from the Ebola virus glycoprotein is a long triple-stranded coiled coil. Proc Natl Acad Sci U S A 95, 6032–6036.
Weissenhorn, W., Dessen, A., Calder, L. J., Harrison, S. C., Skehel,J. J. & Wiley, D. C. (1999). Structural basis for membrane fusion byenveloped viruses. Mol Membr Biol 16, 3–9.
Wild, T. F. & Buckland, R. (1997). Inhibition of measles virus infec-tion and fusion with peptides corresponding to the leucine zipperregion of the fusion protein. J Gen Virol 78, 107–111.
Wild, C. T., Shugars, D. C., Greenwell, T. K., McDanal, C. B. &Matthews, T. J. (1994). Peptides corresponding to a predictive a-helicaldomain of human immunodeficiency virus type 1 gp41 are potentinhibitors of virus infection. Proc Natl Acad Sci U S A 91, 9770–9774.
Yuan, K., Yi, L., Chen, J. & 15 other authors (2004). Suppression ofSARS-CoV entry by peptides corresponding to heptad regions onspike glycoprotein. Biochem Biophys Res Commun 319, 746–752.
Zhao, X., Singh, M., Malashkevich, V. N. & Kim, P. S. (2000).Structural characterization of the human respiratory syncytial virusfusion protein core. Proc Natl Acad Sci U S A 97, 14172–14177.
http://vir.sgmjournals.org 1097
HSV-1 gB and gH heptad repeat regions