an operon from lactobacillus helveticus composed of a proline iminopeptidase gene (pepl) and two...
TRANSCRIPT

Downloaded from www.microbiologyresearch.org by
IP: 54.157.171.172
On: Tue, 16 Aug 2016 02:32:32
Microbiology (1 996), 142, 3459-3468 Printed in Great Britain
An operon f rom Lactobacillus helveticus composed of a proline iminopeptidase gene (pep/) and two genes coding for putative members of the ABC transporter family of proteins
Pekka Varmanen,t Terhi Rantanen and Airi Palva
Author for correspondence: Airi Palva. Tel: +358 16 4188 277. Fax: +358 16 4384 550. e-mail : [email protected]
Ag ricu ltura I Research Centre of Finland, Food Research I nst i tu te, Jokioinen 31600, Finland
A proline iminopeptidase gene (pep/) of an industrial Lactobacillus helveticus strain was cloned and found to be organized in an operon-like structure of three open reading frames (ORFI, ORFZ and ORF3). ORFI was preceded by a typical prokaryotic promoter region, and a putative transcription terminator was found downstream of ORF3, identified as the pep/ gene. Using primer- extension analyses, only one transcription start site, upstream of ORFI, was identifiable in the predicted operon. Although the size of mRNA could not be judged by Northern analysis either with ORFI-, ORFZ- or pep/-specific probes, reverse transcription-PCR analyses further supported the operon structure of the three genes. ORFl, ORFZ and ORF3 had coding capacities for 507,245 and 33.8 kDa proteins, respectively. The ORF3-encoded Pep1 protein showed 65 O/O
identity with the Pep1 proteins from Lactorbacillus delbrueckii subsp. bulgaricus and Lactobacillus delbrueckii subsp. lactis. The ORFI-encoded protein had significant homology with several members of the ABC transporter family but, with two distinct putative ATP-binding sites, it would represent an unusual type among the bacterial ABC transporters. ORFZ encoded a putative integral membrane protein also characteristic of the ABC transporter family. The pep/ gene was overexpressed in Escherichia coli. Purified Pep1 hydrolysed only di- and tripeptides with proline in the first position. Optimum Pep1 activity was observed at pH 7.5 and 40 O C . A gel filtration analysis indicated that Pep1 is a dimer of M, 53000. Pep1 was shown to be a metal-independent serine peptidase having thiol groups at or near the active site. Kinetic studies with proline-pnitroanilide as substrate revealed K,,, and Vmax values of 0 8 mM and 350 mmol min-l mg-l, respectively, and a very high turnover number of 135 000 s-l.
Keywords : Lactobacillus helveticus, proline iminopeptidase, ABC transporter
INTRODUCTION
Lactic acid bacteria (LAB) are limited in their capacity to synthesize amino acids and, therefore, they need ex-
t Present address: Valio Ltd, Research and Development, Helsinki, Finland.
Abbreviations: DIG, digoxigenin-dUTP; LAB, lactic acid bacteria; Pro- pNA, proline-p-nitroanilide; pHMB, p-hydroxymercuribenzoic acid; RT- PCR, reverse transcription-PCR.
The EMBL accession number for the sequence of 3240 bp DNA fragment, carrying the pep/ and ABC transporter genes, reported in this paper is 256283.
ogenous nitrogen sources for optimal growth. When growing in milk, LAB utilize complex proteolytic systems for the degradation of milk proteins. In recent years, the biochemical and genetic characterization of this system has received considerable attention (for reviews, see Kok, 1990; Pritchard & Coolbear, 1993; Tan e t al., 1993; Visser, 1993; Kok & de Vos, 1994). The proteolytic system is composed of a cell-envelope-associated pro- teinase, membrane-bound transport systems, and several cytoplasmic peptidases. The milk proteins contain a high amount of proline (Fox, 1989), resulting in the generation of proline-rich peptides during proteinase action (Kok &
0002-0973 0 1996 SGM 3459

Downloaded from www.microbiologyresearch.org by
IP: 54.157.171.172
On: Tue, 16 Aug 2016 02:32:32
P. V A R M A N E N , T. R A N T A N E N a n d A. P A L V A
de Vos, 1994). Dipeptides and oligopeptides containing an N-terminal proline residue are usually not hydrolysed by general-purpose amino-, di- or tripeptidases. Thus, for degradation of the proline-rich peptides, several prol.ine- specific peptidases with distinct substrate specificities have been evolved in LAB. So far, activities such as prolinase, prolidase, proline iminopeptidase, amino- peptidase P and X-prolyl dipeptidyl aminopeptidase have been detected in various species of LAB (Baankreis & Exterkate, 1991; Meyer-Barton e t al., 1993; Kok 8: de Vos, 1994; Mars & Monnet, 1995; Stucky etal., 1995). It has been previously reported that neither Lactobarillus helveticm nor Lactobacilltls delbrueckii subsp. bdgarictls (hereupon L. bulgaricus) exhibits proline iminopeptidase activity (Kok 8: de Vos, 1994; Baankreis & Exterkate, 1991). However, two recent reports describe the charac- terization of an iminopeptidase gene (pep0 from L. bulgaricus (Atlan e t al., 1994; Gilbert e t al., 1994). On the other hand, a prolinase gene, cloned and characterized from L. helvetictrs (Dudley & Steele, 1994; Varmanen e t al., 1996), possesses an amino acid sequence with 31 YO identity to the proline iminopeptidase from L. btllgaricus (Atlan e t al., 1994) and Lactobacillus delbrueckii subsp. r’actis (Klein e t al., 1994). Although the substrate specificities and other properties of the proline iminopeptidase and prolinase of these two Lactobacillus species show close resemblance (Gilbert e t al., 1994; Dudley & Steele, 1994; Varmanen e t a/., 1996), the level of the amino acid sequence homology does not support that these two enzymes are counterparts.
In this paper, we show that L. he1veticu.r also possesses an iminopeptidase, in addition to the prolinase, by cloning its gene with the aid of a L. bukaricus pep1 probe. Fur- thermore, DNA sequence and mRNA analyses ofpep1 and two ABC transporter genes, all organized in an operon- like structure in L. helveticus 53/7, are described. In addition, overproduction, purification and enzymic characterization of the recombinant L. helveticus Pep1 are reported.
METHODS
Bacterial strains, plasmids and culture conditions. L. helveticus strain 53/7 is an industrial starter from the collection of Valio Ltd (Helsinki, Finland). L. helveticus was routinely grown in MRS or Whey broth at 42OC without shaking. Whey broth comprised 50 g whey permeate (Valio), 20 g casein hydrolysate (Valio) and 10 g yeast extract per 1. For a pH-controlled cultivation, a Biostat M bioreactor was used. The pH was allowed to decrease to 5.6 at which point it was maintained by addition of 1 M NH,. L. bulgaricus B14 (Federal Dairy Research Centre, Kiel, Germany) was grown in MRS broth at 37 “C. Escherichia coli strains DH5aF’ (Woodcock e t al., 1989), JMl05 (Pharmacia) and NM514 (Amersham) were grown in Luria broth. Erythromycin (0.3 mg ml-l) and ampicillin (0-05 mg ml-l) were added when using the vectors pJDC9 (Chen & Morrison, 1987) and pKK223-3 (Pharmacia), respectively, in E. coli.
Construction and screening of a L. helveticus genomic library. Chromosomal DNA was isolated essentially as described by Vidgren e t al. (1992) without a guanidine hydrochloride
treatment, followed by partial digestion with HaeIII. A 3-6 kb fragment pool was selected for a L. helvetictls genomic library established in AgtlO using A-DNA in vitro packaging and cDNA rapid cloning modules (AgtlO; Amersham). Instead of the Amersham cDNA rapid adaptor ligation module, a RiboClone EcoRI Linker System (Promega) was employed. The titration and plating of the phage were performed according to the manufacturers instructions. A cloning efficiency of 1.16 x lo7 p.f.u. (pginsert DNA)-l was reached. The L. helveticus 57/3 genomic library contained 9.5 x lo5 p.f.u., of which 80-90% carried an insert. The diluted library was plated and approximately 7500 plaques were transferred to positively charged nylon membranes (Boehringer Mannheim) and screened by DNA hybridization. For the hybridization probe, a 1-2 kb fragment, carrying the pepI gene from the L. bulgaricus chromosomal DNA (Atlan e t al., 1994), was amplified by PCR using the primer pair 5’-GTCGGTCGACAGACTGGCACG- TCATAGAC-3’ and 5’-GGTTGAATTCGGTTCAGCGAG- CATGTC-3’. Labelling of the probe was with digoxigenin- dUTP (DIG, Boehringer Mannheim). Hybridization was at 60 “C overnight, followed by washes with 0 . 5 ~ SSC, 0.1 % SDS at 50 OC. Isolation of phage DNA was according to the instructions of the cDNA rapid cloning module. Alkaline lysis (Sambrook e t al., 1989) was used for plasmid isolations from E. coli with the DNA purification systems Magic (Promega) and Flexi-Prep (Pharmacia). Other recombinant DNA techniques were performed essentially as described by Sambrook e t al. (1 9 89).
DNA synthesis. The oligonucleotides were synthesized with an Applied Biosystems DNA/RNA synthesizer model 392 and purified by ethanol precipitation or with NAP-10 columns (Pharmacia). For DNA synthesis, performed by PCR ampli- fication, reaction conditions recommended by the manufacturer of DynaZyme DNA polymerase (Finnzymes) were used.
Nucleotide sequencing and sequence analysis. Sequencing was performed on an ALF DNA Sequencer (Pharmacia). The dideoxy sequencing reactions (Sanger e t al., 1977) were per- formed according to the AutoRead Sequencing Kit manual (Pharmacia). Both DNA strands were sequenced using pUC19- specific primers and sequence-specific oligonucleotides for primer walking. DNA sequences were assembled and analysed with the PC/GENE set of programs (release 6.85 ; IntelliGenetics). The PROSITE program of PC/GENE was used to detect specific sites and signatures in protein sequences. Hydropathy analyses were by the method of Kyte & Doolittle (1982) with the SOAP
program of PC/GENE. For homology searches, the databases of EMBL and SWISS-PROT were used both as a CD-ROM version (release 15.0, June 1995) and directly by e-mail with the EMBL BLITZ server.
RNA methods. Total RNA was isolated from L. helveticus cells essentially as described by Palva e t al. (1988) and Vesanto e t al. (1994). RNA agarose gel electrophoresis and Northern blot were performed as described previously (Hames & Higgins, 1985). Hybridization probes were labelled with [a-32P]dCTP (> 3000 Ci mmol-’/lll TBq mmol-l; Amersham) by the protocol of a random-primed DNA labelling kit (Amersham) or with nonradioactive DIG. DIG luminescent kit (Boehringer Mannheim) was used for hybrid detection with the DIG- labelled probe. The primer extensions were performed using an ALF DNA Sequencer essentially as described by Myohanen & Wahlfors (1993) and Vesanto e t al. (1995). The antisense fluorescein-labelled oligonucleotides used in primer extensions were 5’-TTCCGGCATATTTTGGATAG-3’, 5’-AGCGTCG- CAAATATCGTG-3’ and 5’-TTTGTGCTAATTCGTCCAG-
3460

Downloaded from www.microbiologyresearch.org by
IP: 54.157.171.172
On: Tue, 16 Aug 2016 02:32:32
PepI-ABC transporter operon from L. heluetictls
3', being complementary to nucleotides at positions 31 7-337, 1698-1715 and 2319-2337, respectively (Fig. 2). Reverse transcription-PCR (RT-PCR) was performed as follows. Three cDNA preparations were first synthesized with RT (AMV reverse transcriptase, Promega) as described for the primer extension using 20 pg total RNA isolated from L. helvetictls after growth of 4 h. The oligonucleotides used in the first-strand synthesis were P1 , 5'-GCCTCTAATTCTTTCAC- CC-3', P2, 5'-TCGGCAGAGTTGAGCGC-3' and P3, 5'- CCAGGAGATAAATCAACGC-3', being complementary to nucleotides at positions 2431-2449,2011-2027 and 1618-1636, respectively (Fig. 2). After reverse transcription, the three cDNA samples were phenol extracted, precipitated and dis- solved in 16 p1 H,O. Amplifications by PCR were performed with 2 pl cDNA samples. The oligonucletide P4, 5'-CATT- GCGGGACTCTATCC-3', binding to nucleotides 306-323 (Fig. 2), was used as the upstream primer for the cDNA obtained with P1. The oligonucleotide P5, 5'-GCTGATGA- CAAAGCATAATGTG-3', binding to nucleotides 1305-1 326 (Fig. 2) was used as the upstream primer for cDNAs obtained with P2 or P3. As a control for the absence of contaminating DNA, 5 pg total RNA was directly amplified by PCR using oligonucleotide pairs P1 /P4, P2/P5 and P3/P5. Peptidase activity assays. From liquid cultures, peptidase activities in L. helvetiem and E. coli were determined as follows : cells were harvested by centrifugation at 7000g for 15 min, frozen in liquid nitrogen, thawed, washed once with 50mM HEPES (pH 7*0), resuspended in one-third of the original volume of the same buffer, followed by sonication on ice using an Ultrasonic 2000 sonicator (B. Braun) with intervals of 15 s until the cells were disrupted. After removal of the cell debris, peptidase activities were determined according to the method of El Soda & Desmazeaud (1982) using 16.4 mM L-proline-p- nitroanilide (Sigma) as the substrate in 50 mM HEPES (pH 7.0). The enzyme activity measurements with purified PepI were performed at 40 "C using Pro-pNA as the substrate in 50 mM HEPES (pH 7.5). The release ofp-nitroanilide was followed at 410 nm (Perkin Elmer Lambda Bio UV/VIS spectrophoto- meter). A molar extinction coefficient of 8800 M-' cm-' was used to calculate the specific enzyme activity as units (mg protein)-'. Hydrolysis of non-chromogenic peptides (Sigma; Bachem) was determined by two alternative methods : the iminopeptidase activity (proline-liberating) was determined using the modified method of Troll & Lindsley (1955) as described by Baankreis & Exterkate (1991). The hydrolysis of peptides not containing proline as the amino-terminal residue was assayed by measuring the amount of liberated a-amino termini from hydrolysis of peptide substrates using the Cd/ninhydrin method as described by Doi e t al. (1981). The buffer and the incubation temperature used were 50 mM HEPES (pH 7.5) and 40 "C. All activity assays were performed with at least two parallel samples. The protein content was determined by the Bio-Rad protein assay using BSA as the standard. Construction of an overexpression system for the L. helveticus Pep1 in E. coli. The pepI gene was synthesized by PCR using the primer pair 5'-TTAAGAATTCGTTGTT- CATTTTTAAGTAGG-3' and 5'-AAATAAGCTTACAAA- CGCAGTGAAAGAATG-3'. The 1 kb PCR product, carrying the structural gene of pepI with its ribosome-binding site, was ligated as an EcoRI-HindIII-fragment with the pKK223-3 vector and transferred into the E. coli JMl05 host by electro- poration. The expression of pepI was characterized by growing the cells to a density of 50 Klett units (filter 66), followed by 1 mM IPTG induction and estimation of product
accumulation during 5 h by SDS-PAGE and measurement of PepI activity against the Pro-pNA substrate. Purification of Pepl. The crude extract of E. coli JM105, carrying the pepI construct in pKK223-3, was prepared es- sentially as described by Vesanto e t al. (1995). Briefly, the cells, grown in 250 ml L-broth, were harvested 5 h after 1 mM IPTG induction ofpepI expression. After lysing the cells and removal of the cell debris, the cell-free extract was fractionated by 50 and 80 % (NH&SO, precipitations in two steps, respectively. The 80 YO precipitate was collected by centrifugation (4 OC, 6400 g, 30 min), solubilized in 20 mM Tris/HCl, pH 7.5 and dialysed first against deionized water (2 x 30 min) and then overnight against 20 mM Tris/HCl, pH 7.5. The dialysed sample (10.2 ml) was filtered through a 0.45 pm membrane filter (Millex HA ; Millipore) and applied to a DEAE Sepharose FastFlow column (Pharmacia Biotech), equilibrated with 20 mM Tris/HCl, pH 7.5. Proteins were eluted in a linear gradient of 0.15-0.4 M NaCl in starting buffer (flow rate 3 ml min-l, fraction size 5 ml, gradient volume 780 ml), and the fractions were tested for Pro-pNA-hydrolysing activity. One active fraction (5 ml) was concentrated 10-fold by ultrafiltration through a 30 kDa cut-off membrane (Amicon). In the second purification step, the concentrated fraction (500 pl) was applied to a gel filtration column (Superdex 75 HR 10/30, Pharmacia) equilibrated with 25 mM Tris/HCl, pH 7.5, containing 0.15 M NaC1. Proteins were eluted with the same buffer (flow rate 0.4 ml min-l, fraction size 1 ml, gradient volume 28 ml) and assayed for Pro-pNA-hydrolysing activity. Determination of pH and temperature optima. The effect of pH on PepI activity was determined in the range pH 4-1 0 using a buffer system containing either boric acid, MES, HEPES or maleic acid (50 mM each). Reactions were incubated at 37 "C for 5 min and stopped with 0.5 ml 30% (v/v) acetic acid. The absorbance of the samples was measured at 410nm. The optimum temperature was estimated by the same method in the temperature range &70 "C with 50 mM HEPES (pH 7.5). Inhibition studies. Potential inhibitors were added to final concentrations of 0.1 and 1 mM. Reaction mixtures with the purified PepI were preincubated in 50 mM HEPES (pH 7.5) at 4 "C for 30 min. Reactivation was performed by adding the reactivating agent to the reaction mixture followed by in- cubation at 4 "C for an additional 10 min. After addition of the substrate (Pro-pNA), enzyme activity was determined at 40 "C. Molecular mass determination. The molecular mass of the native PepI was determined by gel filtration chromatography (Superdex 75) equilibrated with 25 mM Tris/HCl, pH 7.5, containing 0.15 M NaC1. The column was calibrated with catalase (232 kDa), aldolase (158 kDa), albumin (68 kDa), ovalbumin (43 kDa) and chymotrypsinogen (25 kDa). SDS- PAGE was performed with an 11 YO polyacrylamide gel (Laemmli, 1970). The gel was stained with Coomassie Brilliant Blue R250. LMW-SDS proteins (Pharmacia) were used as marker standards.
RESULTS
Cloning of the proline iminopeptidase region from L. helveticus 5317
T h e homologous counterpart o f the L. bzllgarictls pep1 gene (Atlan e t al., 1994) was identified from the chromo- somal DNA of L. helveticus 53/7 by Southern hybridization using a 1.2 k b pep1-specific probe derived by PCR from L. btllgaricus chromosomal DNA (data n o t shown). T h e probe hybridized to restriction fragments

Downloaded from www.microbiologyresearch.org by
IP: 54.157.171.172
On: Tue, 16 Aug 2016 02:32:32
P. V A R M A N E N , T. R A N T A N E N a n d A. P A L V A
H P E E S P A
OW1 ORE2 pepl
gtlOlcl23
gtlOld4 gtlOldll
- - gtlOldj0
7 p-107 pKTHZlO8
1 kb pIzTIi2109 - pKTH2110
Fig. 7. Partial restriction map of the L. helveticus 53/7 pepl region. The positions and orientations of ORF1, ORF2 and pep/ are indicated by arrows. The inserts of the AgtlO clones gtlOk123, gtlO/clll, gtlOkl4 and gt10k130 and the inserts of the plasmids pKTH2107, pKTH2108, pKTH2109 and pKTH2110 are shown below. Restriction enzymes: El EcoRI; HI Hindlll; P, Pstl; 5, Sall. The open bar with a Hindlll site refers to a DNA segment from AgtlO in pKTH2107.
apart from those detected by a L. helvetictls prolinase (Varmanen e t al., 1996) probe (data not shown). To clone the corresponding pep1 gene, a AgtlO-based genomic library of L. helvetictls 53/7 in E. coli was screened with the above pep1 probe by plaque hybridization. Four hybridization-positive clones, gtlO/cl4, gtlO/clll, gtlO/ c123 and gt10/c130 were chosen for further charac- terization. Phage DNAs of the clones were isolated and digested with various restriction enzymes and further analysed by Southern hybridization. Positive hybridization signals were obtained with 1.2 and 1.6 kb EcoRI fragments from c14 and cl l l , respectively, with a 4.5 kb HikdIII fragment of c123 and with a 0.7 kb EcoRI- Hind111 fragment of c130 (Fig. 1). For sequencing, these fragments were subcloned with p JDC9 and designated pKTH2109, pKTH2108, pKTH2107 and pKTH2110, respectively. A partial restriction enzyme map of the pep1 region in L. helvetictls is shown in Fig. 1.
Nucleotide and amino acid sequence analyses
DNA sequencing of the inserts of pKTH2'108, pKTH2109 and pKTH2110 and partial sequencing of that of pKTH2107 revealed a 3240 bp region of the chromosomal DNA of L. helvetictls carrying three open reading frames (ORF1, ORF2 and ORF3) of 1332, 651 and 882 bp, respectively (Fig. 2). ORFl starts with the GTG codon, ORF2 and ORF3 with ATG. All ORFs are preceded by a putative ribosome-binding site (Fig. 2). ORFl contains 444 codons and has a coding capacity for a 50.7 kDa protein. Conserved -35 and -10 regions (TAGACA-N,,-TACAAT) of a putative promoter were found 52 and 29 nucleotides upstream of the ORFl start codon, respectively. The start codon of ORF2 is located three nucleotides downstream of the stop codon of ORFl. ORF2 contains 217 codons and has a coding capacity for a 24.5 kDa protein. The start codon of ORF3 and the stop codon of ORF2 are separated by eight nucleotides. The ORF3 contains 294 codons and has a coding capacity for a 33.8 kDa protein. ORF3 is followed by an inverted repeat structure one nucleotide downstream of two stop codons.
This hairpin with a AG of -50 kJ mol-' consists of a stem of 9 bp and a loop of 6 bp and is a putative rho- independent transcription terminator. The nucleotide and deduced amino acid sequences of ORF3 were shown to be 64 and 65 % identical with those of the pep1 gene and its product from L. btl&arictl.r, respectively. Thus, ORF3 was confirmed to be the structural pep1 gene of L. helvetiem. The similarity of the L. helvetictls Pep1 protein with PepIs from L. delbrtleckii subsp. lactis (Klein e t al., 1994) and L. bz4Lgarictl.r (Atlan e t al., 1994) and with the L. helvetictls prolinase (Dudley & Steele, 1994; Varmanen e t al., 1996) was shown to be 75, 75 and 39 YO, respectively.
Analysis of the predicted amino acid sequence of the ORF1-encoded 50-7 kDa protein revealed the presence of two ATP/GTP-binding site P-loops also known as Walker A motifs (Walker e t al., 1982; Higgins, 1992) ; one in the N-terminal half of the protein (residues 29-36, Fig. 2) and the other in the C-terminal part (residues 263-270, Fig. 2). In addition, the Walker B motifs (Walker e t al., 1982 ; Higgins, 1992), also involved in ATP-binding, were found in the N- and C-terminal halves (residues 143-147 and 382-386). According to the homology searches performed, the 50.7 kDa protein showed hom- ology to several members of the family of ABC trans- porters. Surprisingly, the highest levels of relatedness of the N- and C-terminal halves of the 50.7 kDa protein were to different proteins. The highest similarities of the N- terminal half was found with Nod1 (50 YO) from RhiTobitlm legtlminosartlm (Evans & Downie, 1986), MalK (44 YO) from E. coli (Gilson e t al., 1982) and LacK (42 %) from Agrobacteritlm radiobacter (Williams e t al., 1992). Instead, the C-terminal half of the 50-7 kDa protein shared the highest similarities with OppF (43 YO) from Lactococctls lactis (Tynkkynen e t al., 1993), GlnQ (40 YO) from Bacilltls stearothermophiltls (Wu & Welker, 1991) and PhnC (37 YO) from E. coli (Makino e t al., 1991). The similarity between the N- and C-terminal parts of the 50.7 kDa protein was 38%. Thus, the ORF1-encoded protein seems to be formed of two different ATP-binding domains fused into a single polypeptide, which represents an unusual ATP- binding protein type among the prokaryotic ABC trans- porters. Hydropathy analysis revealed that the 24.5 kDa protein, encoded by ORF2, contains stretches of hydro- phobic amino acids being able to form five transmembrane helices (data not shown). Thus, the 24.5 kDa protein represents the transmembrane protein of this putative ABC transporter.
Expression of pep/ in L. helveticus
Expression of the pep1 gene in L. helvetiem 53/3 was studied in a pH-controlled culture in Whey broth by measuring peptidase activity against the L-proline-p- nitroanilide substrate and the level of pep1 transcripts as the function of growth (data not shown). The amount of proline-specific enzyme activity increased up to the early stationary phase reaching the highest level 8-10 h after inoculation (Klett,, values 532-530). During the rest of the growth analysed, the activity against the Pro-pNA
3462

Downloaded from www.microbiologyresearch.org by
IP: 54.157.171.172
On: Tue, 16 Aug 2016 02:32:32
PepI-ABC transporter operon from L. heluetictls
1
1 2 1
1
2 4 1 2 0
3 6 1 6 0
4 8 1 1 0 0
6 0 1 14 0
7 2 1 1 8 0
8 4 1 220
9 6 1 260
1 0 8 1 3 0 0
1 2 0 1 340
1 3 2 1 3 0 0
1 4 4 1
C A G C T G C C A T A T G T C C A C C T ~ ~ ~ ~ ~ T A A T A A T A G T A f f A T A A G ~ ~ ~ A T ~ ~ T A ~ ~ T A ~ T A T A T ~ ~ ~ A ~ T A ~ ~ C C ~
T I l T A C G G C A G T A O A C A G G C T C A ~ ~ C A G ~ ~ A ~ ~ T A ~ ~ ~ T C A A T C T A A ~ ~ A ~ A A C C C ~ A ~ C ~ ~ A A T A T G A A T - 3 5 - 1 0 RBS + 0-1
r V N V T F S Y E K E P L I N Q L N M N
A ? T G C A C C C G G ~ A A T C T C f i G A T C G G A C C A A C A G G C P G T I A P G F N L L I G P T G C G K S T L L K I I A G L Y P K Y A G K L T G K I D L
C A T G G T C A A R A A G C G G C C A T G A T G A T G T I T C A A A A T O e r O C C O C H G Q K A A M M F Q N A A E Q F T M T T P R E E I I F A L E N L Q I K A K D Y D
C T C C A C A T ~ C G G n ~ ~ C A G A ~ ~ A C ~ ~ T A C A ~ A T ~ ~ ~ ~ ~ ~ T C T ~ A G C ~ T ~ ~ T I I ; ~ T G G A T L H I K K A V E F T K I A D L L D Q K I N T L S G G Q Q Q H V A L A V L I A M D
G T C G A T G T P r r P C T G C T A G A C G A A C C ~ C T A G T n ; r V D V F L W B L D E P F A S C D P N T R H F L I E K L A S L A E T G R T I I L S D H
G T A ~ A T G A ~ A T G A G ~ G A ~ T G A T C A C ~ A T A ~ A A ~ T A A A A C ~ ~ ~ A A ~ A ~ ~ ~ ~ T ~ ~ ~ ~ T ~ ~ ~ C A ~ V L D D Y E K I C D H L Y Q F E G K T V K E L S A N E K N K L F K Q N K Q F H E
C A A T ( I I T A T P C T T T T G C G T C C A A ~ C A C A C C T G ~ A A T T ~ ~ A C A C A G A T ~ ~ T ~ T ~ ~ ~ ~ ~ ~ ~ T ~ A ~ ~ ~ ~ Q S Y S F A L P T G T P V F E L N K T Q I K Q N R L L L K Q N K L K I Y G K T T
C T A A T T A C P G G T T C A A A T G G T G T C G G C A A A A C A T C A C T A ~ ~ ~ ~ A ~ ~ T G A ~ C ~ T A ~ ~ ~ A C ~ A ~ ~ T A A T C ~ ~ ~ T ~ T A C ~ T L I T - S L F K A M T K M I P Y K G N F T Y L D N E I S K I K Y R
A n A T A m A T C A C A A A T A G C A C ~ ~ C ~ C A A ~ A ~ ~ A A ~ ~ A C A G T T ~ ~ A G A ~ A A ~ ~ T C G ~ T ~ ~ C A ~ ~ K Y L S Q I A Q F F Q K A S D Q F L T V T V K D E I E L S K K D R N N F F T D A
A A A A ~ T C A A T G G C T A G ~ G ~ ~ ~ ~ ~ C A C ~ T ~ T A G T ~ A ~ ~ ~ A T ~ ~ ~ ~ ~ ~ ~ T G A ~ ~ T K I D E W L E K L Q L K Q H L D Q V V Y S L S G G Q Q K K L Q I L L M L M T K H
A A T G T G ? n ; C T A A T I I ; A C G A G C C A T I C I A G C G G A C T A G A N V L L I D E P L S G L D H E S V D L V L Q L M Q E C Q E K L Q Q T F L I I S H
C A G A T A G A C G C G T ~ ~ T T A ~ ~ ~ C A G C T ~ T A T G ~ ~ ~ ~ T T ~ A G ~ ~ ~ A ~ ~ A ~ A T T
WA
RBS + ow2 WB
4 2 0 / 1 Q I D A L A D F C D Y R L V F A D Q Q L K Y V E E M N S S L K F I L A F I I
1 5 6 1 T C C C r C I G A A A T A T C C G A G G C G A G T P T A A C C A C C A A 1 4 S L E I S L K A S L T T N L I V 1 A F A L I Y L L V T R I K I K E L I L L I A V
1 6 8 1 C C A ~ A ~ C G T C G ? T C A C G A T A ~ C G A C G ~ ~ ~ ~ C A C ~ A ~ C T C A T C ~ A ~ A ~ T ~ T ~ A C C ~ ~ ~ T A ~ ~ A T A ~ A C C A ~ ~ T 5 4 P F I A S F T I F A T L F W F S P T P D A Y Y A W N L S T R V Y V Y T L T I A C
1 8 0 1 G T C A C G A G G A A C A C C A C C G C G A C A G A ~ ~ C G T T C G ~ A A ~ ~ C A ~ A C ~ ~ T T ~ ~ A ~ ~ ~ C ~ T ~ ~ T T A ~ C ~ ~ A ~ ~ 9 4 V T R N T T A T D F A R S L E Q N L H L P S K F A Y G V L A A I N I I P R M K T
1 9 2 1 G C A G T G A A G ~ A ~ C G T A C A T ~ C C A T C A T C C G G A 1 3 4 A V K Q I R T S A M M R G M Y L S F W S P V L Y F K A I L V A L N S A D N L A Q
2 0 4 1 G G C A ~ A A T C C C A C G G C T A C G T I I ; ~ A ~ A G ~ C C A ~ T A G ~ T T C ~ ~ ~ ~ A ~ ~ ~ A ~ A ~ ~ T T A A T A ~ ~ 1 7 4 G M E S H G Y V E G Q K R A T I V A I P L T K K D W L I F F T L L I L V N I S L
2 16 1 ' I T C A ? ? m M C T A o ~ ~ C C ~ ~ ~ ~ A T G ~ ~ A ~ A T ~ ~ ~ ~ ~ C A C T ~ ~ ~ C 2 1 4 / 1 F I F K M E I I E G K M P F M G Y E T Y Y R I V G E R S E K P P L V L L H
RBS + O R P ~ w p r )
2 2 8 1 A C G G C G G A C ~ C T C A A G C C A ~ ~ A ~ ~ T G ~ A ~ ~ ~ ~ C ~ A T G G C C G C C G ~ T ~ T T A T G T A ~ A C ~ T T ~ ~ ~ T C ~ ~ ~ T C C ~ A T C 34 G G P G S S H N Y F E V L D E L A Q K D G R R I I M Y D Q L G C G E S S I P D D
2 4 0 1 A T C A T C C T G A A ~ A C A C T ~ ~ ~ T G ~ G A A ~ ~ ~ ~ A C G T C A G C A ~ A G C A T T A ~ ~ A T C ~ ~ ~ A ~ ~ C C A ~ A 74 H P E L Y T K E T W V K E L E A L R E H L A L R K M H L L f Q : W f Y M L A I I
2 5 2 1 ? T f A C A T C T G C G A C T A T C A T C C ~ ~ T A T T ~ ~ ~ A ~ T ~ ~ A C C ~ T ~ T C ~ ~ T C C ~ ~ T ~ ~ C A C ~ ~ A ~ ~ T A ~ C ~ ~ ~ 114 Y M C D Y H P E G I Q S L I L S S T L S S A S L W S K E L H R M I K Y L P I E E
2 6 4 1 A G C A ~ C A G C G A T T C A T A ~ A T A G G G C T G A G ~ A ~ ~ ~ A A ~ A T C ~ ~ A C ~ ~ A A ~ A G C A ~ ~ T C ~ C ~ ~ ~ C A ~ T A T C A ~ A A ~ C ~ ~ A A T 154 Q A A I H R A E L T G N F N D P D Y L K A N E H F M N Q H A I D M T K T W P E C
2 7 6 1 G C G T G A n ; C G C A A A A A G C G T C G G ~ C T G ~ ~ T A T G ~ ~ C A ~ A C ~ A A ~ T A T A ~ C ~ ~ ~ ~ A ~ A ~ T A ~ ~ T ~ ~ G ~ T A A 194 V M R K K R G G T V A Y E T A W G P N E Y T P E G N L H D Y E Y T D K L S K I K
2 0 8 1 A A G ~ C C A A e r C T G A ~ A ~ A G ~ A A C T G A ~ A ~ A ~ T A C G C C A T A T C T A G ~ ~ A C T A T C ~ A T ~ A ~ ~ ~ ~ ~ ~ ~ A ~ ~ ~ C A C A T C T 234 V P T L I T S G T D D L C T P Y V A K T M Q D Q I A S S K W R L F E G C G H M S
Fig. 2. The nucleotide and deduced amino acid sequences of the ABC transporter-pep/ region. ORFl and ORF2 refer to the ATP-binding protein and the transmembrane protein of the ABC transpoi-ter, respectively. The predicted -10 and -35 regions of the promoter are underlined. The 5' end of the common transcript of the ABC transporter genes and pep/, found by primer extension, is marked with a vertical arrow. RBS, predicted ribosome-binding site. The deduced transcription terminator downstream of ORF3 (pep/) is shown with dotted arrows. The Walker motifs, A (WA) and B (WB), characteristic of many nucleotide-binding proteins, are underlined. The conserved residues of the active site region of prolyl oligopeptidases are in bold and marked with asterisks.
3463
Downloaded from www.microbiologyresearch.org by
IP: 54.157.171.172
On: Tue, 16 Aug 2016 02:32:32
P. V A R M A N E N , T. R A N T A N E N a n d A. P i i L V A
mRNA cDNA 1 cDNA 2 cDNA 3
PCR 1 PCR 2
PCR 3
ORFI ORF2 pep1
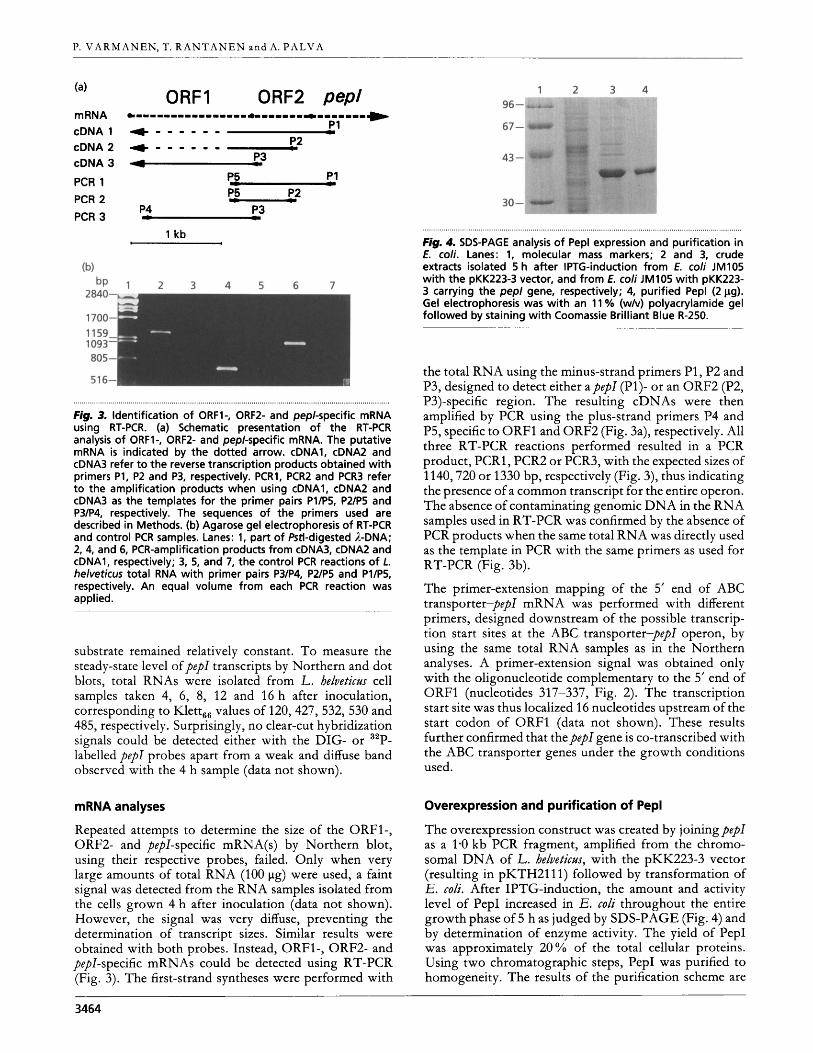
Fig. 3. Identification of ORF1-, ORF2- and pepl-specific mRNA using RT-PCR. (a) Schematic presentation of the RT-PCR analysis of ORFl-, ORF2- and pep/-specific mRNA. The putative mRNA is indicated by the dotted arrow. cDNA1, cDNA2 and cDNA3 refer t o the reverse transcription products obtained with primers P1, P2 and P3, respectively. PCR1, PCR2 and PCR3 refer t o the amplification products when using cDNA1, cDNA2 and cDNA3 as the templates for the primer pairs Pl/P5, P2/P5 and P3/P4, respectively. The sequences of the primers used are described in Methods. (b) Agarose gel electrophoresis of RT-PCR and control PCR samples. Lanes: 1, part of Pstl-digested A-DNA; 2, 4, and 6, PCR-amplification products from cDNA3, cDNA2 and cDNA1, respectively; 3, 5, and 7, the control PCR reactions of L. helveticus total RNA with primer pairs P3/P4, P2/P5 and Pl/P5, respectively. An equal volume from each PCR reaction was applied.
substrate remained relatively constant. To measure the steady-state level ofpep1 transcripts by Northern and dot blots, total RNAs were isolated from L. helveticzrs cell samples taken 4, 6, 8, 12 and 16 h after inoculation, corresponding to Klett66 values of 120,427, 532, 530 and 485, respectively. Surprisingly, no clear-cut hybridization signals could be detected either with the DIG- or 32P- labelled pep1 probes apart from a weak and diffuse band observed with the 4 h sample (data not shown).
mRNA analyses
Repeated attempts to determine the size of the ORF1-, ORF2- and pepl-specific mRNA(s) by Northern blot, using their respective probes, failed. Only when very large amounts of total RNA (100 pg) were used, a faint signal was detected from the RNA samples isolated from the cells grown 4 h after inoculation (data not shown). However, the signal was very diffuse, preventing the determination of transcript sizes. Similar results were obtained with both probes. Instead, ORF1-, ORF2- and pepl-specific mRNAs could be detected using RT-PCR (Fig. 3). The first-strand syntheses were performed with
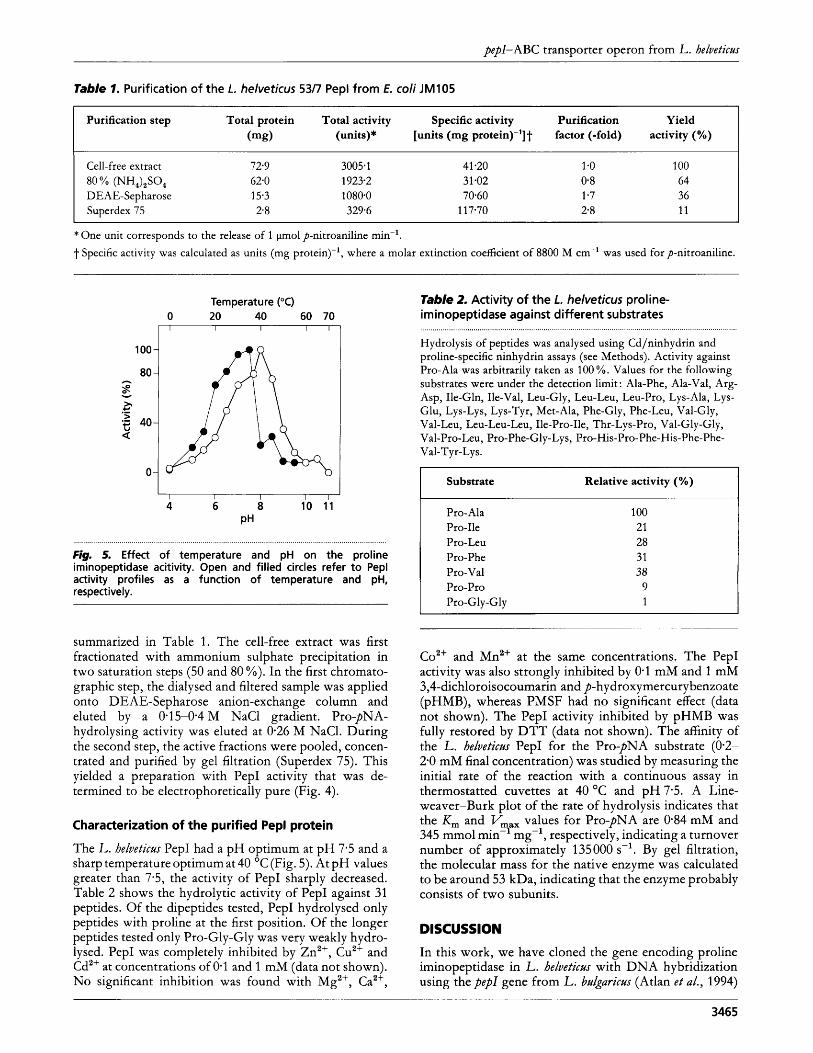
fig. 4. SDS-PAGE analysis o f Pep1 expression and purification in E. coli. Lanes: 1, molecular mass markers; 2 and 3, crude extracts isolated 5 h after IPTG-induction from E. coli JM105 with the pKK223-3 vector, and from E. coli JM105 with pKK223- 3 carrying the pep/ gene, respectively; 4, purified Pep1 (2 pg). Gel electrophoresis was with an 11 % (w/v) polyacrylamide gel followed by staining with Coomassie Brilliant Blue R-250.
the total RNA using the minus-strand primers P1 , P2 and P3, designed to detect either apepl (P1)- or an ORF2 (P2, P3)-specific region. The resulting cDNAs were then amplified by PCR using the plus-strand primers P4 and P5, specific to ORFl and ORF2 (Fig. 3a), respectively. All three RT-PCR reactions performed resulted in a PCR product, PCRl, PCR2 or PCR3, with the expected sizes of 1140,720 or 1330 bp, respectively (Fig. 3), thus indicating the presence of a common transcript for the entire operon. The absence of contaminating genomic DNA in the RNA samples used in RT-PCR was confirmed by the absence of PCR products when the same total RNA was directly used as the template in PCR with the same primers as used for RT-PCR (Fig. 3b).
The primer-extension mapping of the 5’ end of ABC transporter-pep1 mRNA was performed with different primers, designed downstream of the possible transcrip- tion start sites at the ABC transporter-pep1 operon, by using the same total RNA samples as in the Northern analyses. A primer-extension signal was obtained only with the oligonucleotide complementary to the 5’ end of ORFl (nucleotides 31 7-337, Fig. 2). The transcription start site was thus localized 16 nucleotides upstream of the start codon of ORFl (data not shown). These results further confirmed that the pep1 gene is co-transcribed with the ABC transporter genes under the growth conditions used.
Overexpression and purification of Pep1
The overexpression construct was created by joining pep1 as a 1.0 kb PCR fragment, amplified from the chromo- somal DNA of L. helvetictls, with the pKK223-3 vector (resulting in pKTH2111) followed by transformation of E. coli. After IPTG-induction, the amount and activity level of PepI increased in E. d i throughout the entire growth phase of 5 h as judged by SDS-PAGE (Fig. 4) and by determination of enzyme activity. The yield of PepI was approximately 20% of the total cellular proteins. Using two chromatographic steps, PepI was purified to homogeneity. The results of the purification scheme are
3464
Downloaded from www.microbiologyresearch.org by
IP: 54.157.171.172
On: Tue, 16 Aug 2016 02:32:32
pepl-ABC transporter operon from L. helveticm
Table 1. Purification of the L. helveticus 53/7 Pep1 from E. coli JM105
Purification step Total protein Total activity Specific activity Purification Yield (mg) (units)* [units (mg protein)-']t factor (-fold) activity (%)
Cell-free extract
DEAE-Sepharose Superdex 75
80 % (NH,),SO, 72.9 3005.1 62.0 1923.2 15.3 1080.0 2.8 329.6
41.20 31-02 70.60
1 17.70
1.0 100 0.8 64 1-7 36 2.8 11
* One unit corresponds to the release of 1 pmol p-nitroaniline min-'. j- Specific activity was calculated as units (mg protein)-', where a molar extinction coefficient of 8800 M cm-' was used for p-nitroaniline.
Temperature ("C) 40 60 70
100-
80 - n
s
mg 40- a
v
>r + .-
0-
I I I I I
4 6 8 10 11 PH
Fig. 5. Effect o f temperature and pH on the proline iminopeptidase acitivity. Open and filled circles refer t o Pep1 activity profiles as a function o f temperature and pH, respectively.
Table 2. Activity of the L. helveticus proline- iminopeptidase against different substrates ......................................................................................................................................................... . Hydrolysis of peptides was analysed using Cd/ninhydrin and proline-specific ninhydrin assays (see Methods). Activity against Pro-Ala was arbitrarily taken as 100%. Values for the following substrates were under the detection limit : Ala-Phe, Ala-Val, Arg- Asp, Ile-Gln, Ile-Val, Leu-Gly, Leu-Leu, Leu-Pro, Lys- Ala, Lys- Glu, Lys-Lys, Lys-Tyr, Met- Ala, Phe-Gly, Phe-Leu, Val-Gly, Val-Leu, Leu-Leu-Leu, Ile-Pro-Ile, Thr-Lys-Pro, Val-Gly-Gly, Val-Pro-Leu, Pro-Phe-Gly-Lys, Pro-His-Pro-Phe-His-Phe-Phe- Val-Tyr-Lys.
I Substrate Relative activity (%)
Pro- Ala Pro-Ile Pro-Leu Pro-Phe Pro-Val Pro-Pro Pro-G1 y-G1 y
100 21 28 31 38
9 1
summarized in Table 1. The cell-free extract was first fractionated with ammonium sulphate precipitation in two saturation steps (50 and 80 YO). In the first chromato- graphic step, the dialysed and filtered sample was applied onto DEAE-Sepharose anion-exchange column and eluted by a 0.15-0.4 M NaCl gradient. Pro-pNA- hydrolysing activity was eluted at 0.26 M NaC1. During the second step, the active fractions were pooled, concen- trated and purified by gel filtration (Superdex 75). This yielded a preparation with PepI activity that was de- termined to be electrophoretically pure (Fig. 4).
Characterization of the purified Pep1 protein
The L. helveticas PepI had a pH optimum at pH 7.5 and a sharp temperature optimum at 40 OC (Fig. 5). At pH values greater than 7.5, the activity of PepI sharply decreased. Table 2 shows the hydrolytic activity of PepI against 31 peptides. Of the dipeptides tested, PepI hydrolysed only peptides with proline at the first position. Of the longer peptides tested only Pro-Gly-Gly was very weakly hydro- lysed. PepI was completely inhibited by Zn2+, Cu2+ and Cd2+ at concentrations of 0.1 and 1 mM (data not shown). No significant inhibition was found with Mg2+, Ca2+,
Co2+ and Mn2+ at the same concentrations. The PepI activity was also strongly inhibited by 0.1 mM and 1 mM 3,4-dichloroisocoumarin and p-hydroxymercurybenzoate (pHMB), whereas PMSF had no significant effect (data not shown). The PepI activity inhibited by pHMB was fully restored by DTT (data not shown). The affinity of the L. helvetiem PepI for the Pro-pNA substrate (0.2- 2.0 mM final concentration) was studied by measuring the initial rate of the reaction with a continuous assay in thermostatted cuvettes at 40 "C and pH 7.5. A Line- weaver-Burk plot of the rate of hydrolysis indicates that the K, and Vmax values for Pro-pNA are 0.84 mM and 345 mmol min-' mg-l, respectively, indicating a turnover number of approximately 135 000 s-', By gel filtration, the molecular mass for the native enzyme was calculated to be around 53 kDa, indicating that the enzyme probably consists of two subunits.
DISCUSSION
In this work, we have cloned the gene encoding proline iminopeptidase in L. helveticzls with DNA hybridization using the pep1 gene from L. btllgarictls (Atlan e t al., 1994)
3465

Downloaded from www.microbiologyresearch.org by
IP: 54.157.171.172
On: Tue, 16 Aug 2016 02:32:32
P. V A R M A N E N , T. R A N T A N E N and A. P A L V A
as the probe. Although the PepI proteins of L. helvetictls, L. bzllgarictls (Atlan e t al., 1994) and L. delbrzleckii subsp. lactis (Klein e t al., 1994) were revealed to share a relatively high amino acid sequence similarity (75%), the genetic organization of the L. helveticaspep1 locus turned out to be totally different from that of L. btllgarictls and L. delbrtleckii subsp. lactis. The L. helvetictlspepl is preceded by two open reading frames (ORF1 and ORF2), which encode a putative ABC transporter and seem to form an operon with the pep1 gene. A well-conserved promoter is found upstream of ORFl and the only putative transcription terminator locates downstream of pep1 (Fig. 2). The pep1 gene is, however, preceded by a putative weak promoter with a moderately conserved - 35/ - 10 region (TTAA- CT-N,,-TAATAT, position 2128, Fig. 2). Nevertheless, in the 5' end mapping by primer extension, using the total RNA samples from different growth phases, a signal could be detected neither with a primer designed to bind downstream of this putative pep1 promoter (position 2319-2337, Fig. 2) nor with a primer binding downstream of the start site of ORF2 (position 1698-1715, Fig. 2). Instead, the only primer-extension product obtained confirmed the location of the predicted promoter region preceding ORFl . Furthermore, the pep1 gene, cloned under its own putative promoter, was not expressed in E. coli (data not shown). The operon structure was further confirmed by the results of RT-PCR studies, which revealed that at least part of mRNAs derived from the ORFl promoter are transcribed through the pep1 gene (Fig. 3). Unfortunately, the mRNA size determinations by Northern blots were unsuccessful, due to an extremely low concentration of ORF1-, ORF2- and pepl-specific transcript(s) in total RNA. According to the results shown, wc can, howcvcr, conclude that in the growth conditions used the L. belveticzlspepl gene is expressed as a polycistronic transcriptional unit with the two upstream ABC transporter genes. In L. delbrtleckii subsp. lactis and subsp. bzllgariczls, the pepl gene is most probably mono- cistronic, since it is flanked by divergent ORFs encoding proteins not identical with the L. helvetictls ABC trans- porter and the ORFs downstream of pep1 (this study; Atlan e t al., 1994; Klein e t al., 1994).
Analysis of the substrate specificity of the purified PepI revealed that the L. helveticas 53/7 PepI is a proline- specific peptidase with a narrow specificity, similar to that reported for the PepIs from L. btllgarictls (Gilbert et al., 1994) and Lactobacilltls casei (Habibi-Najafi & Lee, 1995). The L. btllgariczls PepI has also been reported to have partial or weak hydrolysing activity against several dipeptides not containing a proline residue (Gilbert e t al. , 1994). A few of these substrates (Ala-Phe, Leu-Gly) were also tested with our PepI, which, however, did not hydrolyse them at a detectable level. It has also been reported that the L. bzllgarictls PepI is equally active against Pro-Ala and Pro-Gly-Gly (Gilbert e t al., 1994 ), whereas a 100-fold difference was observed with the method used in our study (Table 2). It is not known whether these differences are due to real differencies in the enzymic properties of PepIs from L. helvetictls and L. btllgariczls (Gilbert e t al., 1994), or whether they are due to
methodological differences in the measurements of en- zyme activities. The effect of various chemicals against PepI activity was similar to that reported for the L. btllgarictls PepI. The PepI activity is strongly inhibited by 3,4-dichloroisocoumarin and pHMB. The serine protease nature of the L. helvetictls PepI is also confirmed by the presence of the active site region (GQSWGG; at position 103-108, Fig. 2), typical for prolyl oligopeptidases (Rawlings e t al., 1991) and identical with that of the two L. delbrtleckii PepIs. It is noteworthy that PMSF had only a minor effect on PepI activity. The inhibition profiles with divalent cations are similar for the L. helveticzls and L. btllgarictls PepIs (this study; Gilbert e t al., 1994) whereas the L. casei PepI was not strongly inhibited by Cu2+ and Zn2+ (Habibi-Najafi & Lee, 1995). The temperature and pH optima for L. helvetictls PepI (40 "C and pH 7.5) are, instead, identical to those reported for the L. casei Pep1 (Habibi-Najafi & Lee, 1995), whereas the optimum pH for the L. btllgarictls PepI has been shown to be between pH 6.0-7.0, and the enzyme was reported to be unstable at temperatures above 35 "C. The molecular mass determination of the native L. helveticzls PepI indicates that the enzyme probably consists of two subunits. A monomeric and a trimeric form has been reported for PepIs of L. casei and L. btllgariczls (Habibi- Najafi & Lee, 1995; Gilbert e t al., 1994), respectively. To our knowledge, the turnover number of the L. helveticzls PepI with the Pro-pNA substrate (135000 s-'), and that deduced for L. btllgaricus PepI, substantially exceed all those reported for Lactobacilltls peptidases so far. The extremely high turnover number of PepI may also explain the need for a very small amount of pep1 transcripts. The low expression of pep1 is also supported by its codon usage which suggests a low level of translation. Since we were unable to distinguish the PepI activity from that of prolinase (Varmanen e t al., 1996) in L. helveticzls lysates, the ratio and role of these two proline-specific peptidases under the growth conditions used remains to be elucidated.
The ORFl -encoded 50.7 kDa protein shared significant homology with several members of the family of ABC transporters. If ORFl does indeed encode such a trans- porter, then its two putative ATP-binding domains are apparently evolved through an ancient gene duplication or fusion of adjacent genes, thus representing an exception to the more common organization of prokaryotic ABC transporters, where the individual domains are expressed as separate polypeptides (Higgins, 1992). In prokaryotes, only four other ATP-binding proteins from the carbo- hydrate metabolism of E. coli could be found to possess a similar double ATP-binding domain structure to that of the 50.7 kDa protein. These were the transporters of ribose (RbsA) (Buckel e t al., 1986), methylgalactoside (MglA) (Hogg e t al., 1991), L-arabinose (AraG) (Scripture e t al., 1987) and D-xylose (XylG) (Sofia e t al., 1994) of which the RbsA shares the highest similarity with the 50.7 kDa protein (37 YO); these all belong to binding- protein-dependent transport systems. The periplasmic binding protein has been considered to be a unique feature of all bacterial ABC transporters involved in the
3466

Downloaded from www.microbiologyresearch.org by
IP: 54.157.171.172
On: Tue, 16 Aug 2016 02:32:32
PePI-ABC transporter operon from L. helveticus
import of substrates (Fath & Kolter, 1993). However, a gene encoding such a substrate-binding protein was not present in our operon, suggesting that, despite the similarity of the genetic organization of these five ATP- binding proteins, the putative ABC transporter identified in our study may have an export function. This is also supported by the fact that the signature present in all transmembrane proteins of import-specific ABC trans- porter systems (Saurin e t al., 1994) could not be found in the 24.5 kDa putative transmembrane protein encoded by ORF2. Further support for this contention is provided by the observation that the 50.7 kDa protein shares the highest homology with NodI, a transport protein involved in nodulation and probably in the export of a modified /3-1,4-linked N-acetylglucosamine oligo- saccharide (Evans & Downie, 1986). It is unknown whether the common operon structure of the putative ABC transporter genes and pep1 also indicates an interaction of the encoded functions. The structural relationship may be of significance, since the majority of the peptidase genes characterized so far from LAB seem to form a monocistronic transcriptional unit. Interestingly, the L. helveticus prolinase gene is also connected to a putative ABC exporter via the partially overlapping promoter regions (Varmanen e t al., 1996).
ACKNOWLEDGEMENTS
This work was supported by the Ministry of Agriculture and Forestry of Finland. The authors are grateful to Mrs Anneli Virta for the running of the ALF Sequencer and Ms Jaana Jalava for technical assistance.
REFERENCES
Atlan, D., Gilbert, C., Blanc, B. & Portalier, R. (1994). Cloning, sequencing and characterization of the pepIP gene encoding a proline iminopeptidase from Lactobacillus delbrueckii subsp. bulgaricus CNRZ 397. Microbiology 140, 527-535.
Baankreis, R. & Exterkate, F. (1991). Characterization of a peptidase from Lactococcus lactis ssp. cremoris HP that hydrolyses di- and tripeptides containing proline or hydrophobic residues as the aminoterminal amino acid. Syst Appl Microbiol14, 31 7-323.
Buckel, S., Bell, A., Rao, M. & Hermodson, M. (1986). An analysis of the structure of the product of the rbsA gene of Escherichia coli K12. J Biol Chem 261, 7659-7662.
Chen, 1. & Morrison, D. (1 987). Cloning of Streptococcus pneumoniae DNA fragments in Escherichia coli requires vector protected by strong transcriptional terminators. Gene 55, 179-1 87.
Doi, E., Shibata, D. & Matoba, T. (1981). Modified colorimetric ninhydrin methods for peptidase assay. Anal Biochem 118, 173-1 84.
Dudley, E. & Steele, 1. (1994). Nucleotide sequence and distribution of the pepPN gene from Lactobacillus helveticus CNRZ32. FEMS Microbiol Lett 119, 41-46.
El Soda, M. & Desmazeaud, M. (1982). Les peptide hydrolases des lactobacilles du groupe Thermobacterium. I. Mise en Cvidence de ces activitis chez Lactobacillus helveticus, L. acidophilus, L. lactis et L. bulgaricus. Can J Microbiol28, 1181-1 188.
Evans, 1. J. & Downie, J. A. (1986). The nodl gene product of Rhizobium leguminosarum is closely related to ATP-binding bacterial transport proteins ; nucleotide sequence analysis of the nodl and nod] genes. Gene 45, 95-101.
Fath, M. J. & Kolter, R. (1993). ABC transporters: bacterial exporters. Microbiol Rev 57, 995-1017.
FOX, P. (1 989). Proteolysis during cheese manufacture and ripening. J Da iv Sci 72, 1379-1400.
Gilbert, C., Atlan, D., Blanc, 6. & Portalier, R. (1994). Proline iminopeptidase from Lactobacillus delbrueckii subsp. bulgaricus CNRZ 397 : purification and characterization. Microbiolog)r 140, 537-542.
Gilson, E., Nikaido, H. & Hofnung, M. (1982). Sequence of the malK gene in E. coli K12. Nucleic Acids Res 10, 7449-7458.
Habibi-Najafi, M. & Lee, B. (1995). Purification and characterization of proline iminopeptidase from Lactobacillus casei subsp. casei LLG. J Da iv Sci 78,251-259.
Hames, B. & Higgins, S. (1985). Nucleic Acid Hybridisation: a Practical Approach. Oxford & Washington, DC : IRL Press. Higgins, C. (1992). ABC transporters : from microorganisms to man. Annu Rev Cell Biof 8, 67-1 13.
Hogg, R., Voelker C. & Voncarlowitz, 1. (1991). Nucleotide sequence and analysis of the mgl operon of Escherichia coli K12. Mol Gen Genet 229, 453-459.
Klein, J., Schmidt, U. & Plapp, R. (1994). Cloning, heterologous expression, and sequencing of a novel proline iminopeptidase gene, pepI , from Lacto bacillus delbrueckii subs p . lactis D S M 7290. Micro- biology 140, 11 3 3 1 139.
Kok, J. (1990). Genetics of the proteolytic system of lactic acid bacteria. FEMS Microbiol Rev 87, 15-42.
Kok, 1. & de VOS, W. (1994). The proteolytic system of lactic acid bacteria. In Genetics and Biotecbnology of Lactic Acid Bacteria, pp. 169-210. Edited by M. Gasson & W. de Vos. Glasgow: Blackie Academic & Professional. Kyte, 1. & Doolittle, R. F. (1982). A simple method for displaying the hydropathic character of a protein. J Mol BiollS7, 105-132.
Laemmli, U. K. (1970). Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227, 680-685.
Makino, K., Kim, S., Shinagawa, H., Amemura, M. & Nakata, A. (1991). Molecular analysis of the cryptic and functionalphn operons for phosphate use in Escherichia coli K-12. J Bacterioll73,2665-2672.
Mars, 1. & Monnet, V. (1995). An aminopeptidase P from Lactococcus lactis with original specificity. Biochim Biophy Acta 1243, 209-21 5.
Meyer-Barton, E. C., Klein, J. R., Imam, M. & Plapp, R. (1993). Cloning and sequence analysis of the X-propyl-dipeptidyl-amino- peptidase gene (pepx) from Lactobacillus delbrueckii spp. lactis DSM7290. Appl Microbiol Biotechnol40, 82-89.
Myehlnen, 5. & Wahlfors, J. (1993). Automated fluorescent primer extension. Bio Techniques 14, 16-1 7.
Palva, A., Nyberg, K. & Palva, 1. (1988). Quantitation of a-amylase mRNA in Bacilfus subtilis by nucleic acid sandwich hybridization.
Pritchard, G. & Coolbear, T. (1993). The physiology and bio- chemistry of the proteolytic system in lactic acid bacteria. FEMS Microbiol Rev 12, 179-206.
Rawlings, N., Polgar, L. & Barret, A. (1991). A new family of serine type peptidases related to prolyl oligopeptidase. Biochem J 279,
Sambrook, J., Fritsch E. F. & Maniatis, T. (1989). Molecular Cloning: a Laboratov Manual, 2nd edn. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory. Sanger, F., Nicklen, 5. & Coulson, A. R. (1977). DNA sequencing with chain-terminating inhibitors. Proc Natl Acad Sci U S A 74,
Saurin, W., KUster, W. & Dassa, E. (1994). Bacterial binding
D N A 7 , 135-142.
907-9 1 1.
5463-5467.
3467

Downloaded from www.microbiologyresearch.org by
IP: 54.157.171.172
On: Tue, 16 Aug 2016 02:32:32
P. V A R M A N E N , T. R A N T A N E N a n d A. P A L V A
protein-dependent permeases : characterization of distinctive signatures for functionally related integral cytoplasmic membrane proteins. Mol Microbioll2, 993-1004. Scripture, J., Voelker, C., Miller, S., O'Donnell, R., Polgar, L., Rade, J., Horazdovsky, B. & Hogg, R. (1987). High-affinity L-arabinose transport operon nucleotide sequence and analysis of gene products. J Mol Biol197, 37-46. Sofia, H., Burland, V., Daniels, D., Plunkett, G., 111 & Blattner, F. (1994). Analysis of the Escbericbia co l i genome. V. DNA sequence of the region from 76.0 to 81.5 minutes. Nucleic Acids Res 22,
Stucky, K., Klein, J., SchUller, A., Matern, H., Henrich, B. & Plapp, R. (1995). Cloning and DNA sequence analysis ofpepQ, a prolidase gene from Lactobacillus delbrueckii subsp. lactis DSM7290 and partial characterization o f its product. Mol Gen Genet 247, 494-500. Tan, P., Poolman, B. & Konings, W. (1993). Proteolytic enzymes o f Lactococcus lactis. J Dairy Res 60, 269-286. Troll, W. & Lindsley, J. (1955). A photometric method for the determination of proline. J Biol Cbem 215, 655-661. Tynkkynen, S., Buist, G., Kunji, E., Kok, J., Poolman, B., Venema, G. & Haandrikman, A. (1993). Genetic and biochemical charac- terization of the oligopeptide transport system of Lactococcus lactis. J Bacterioll75, 7523-7532. Varmanen, P., Steele, J. & Palva, A. (1996). Characterization of a prolinase gene and its product and an adjacent ABC transporter gene from Lactobacillus helveticus. Microbiology 142, 809-81 6. Vesanto, E., Varmanen, P., Steele, J. L. & Palva, A. (1994). Characterization and expression of the Lactobacillus helveticus pepC gene encoding a general aminopeptidase. Eur J Biocbem 224,
2576-2586.
991-997.
Vesanto, E., Savijoki, K., Rantanen, T., Steele, J. L. & Palva, A. (1 995). An X-prolyl dipeptidyl aminopeptidase (pep% gene from Lactobacillus helveticus. Microbiology 141, 3067-3075. Vidgren, G., Palva, I., Pakkanen, R., Lounatmaa, K. & Palva, A. (1 992). Slayer protein gene of Lactobacillus brevis : cloning by polymerase chain reaction and determination of the nucleotide sequence. J Bacterial 174, 7419-7427. Visser, 5. (1993). Proteolytic enzymes and their relation to cheese ripening and flavor: an overview. J Dairy Sci 76, 329-350. Walker, J., Saraste, M., Runswick, M. & Gay, N. (1982). Distantly related sequences in the a and /I subunits of ATP synthetase, myosin, kinases and other ATP requiring enzymes and a common nucleotide binding fold. EMBO J 1, 945-951. Williams, S. G., Greenwood, J. A. & Jones, C. W. (1992). Molecular analysis o f the lac operon encoding the binding-protein-dependent lactose transport system and /I-galactosidase in Agrobacteritlm radiobacter. Mol Microbioll3, 1755-1768. Woodcock, D., Crowther, P., Doherty, J., Jefferson, 5.. De Cruz, E., Noyer-Weidner, M., Smith, S., Michael, M. & Graham, M. (1989). Quantitative evaluation of Escbericbia coli host strains for tolerance to cytosine methylation in plasmid and phage recombinants. Nucleic Acids Res 17, 3469-3478. Wu, L. & Welker, N. E. (1991). Cloning and characterization of a glutamine transport operon of Bacillus stearotbermopbilus NUB36 : effect of temperature on regulation of transcription. J Bacterioll73, 487774888.
Received 15 May 1996; revised 12 August 1996; accepted 16 August 1996.
3468