an alkali-inducible flotillin-like protein frombacillus haloduransc-125
TRANSCRIPT

An Alkali-Inducible Flotillin-like Protein fromBacillus halodurans C-125
Hui-Min Zhang,1,2
Zhijun Li,1Mikiko Tsudome,
1Susumu Ito,
1Hideto Takami,
1and Koki
Horikoshi1
Received November 10, 2004
Flotillins are markers of lipid microdomains, and have emerged as a key concept in cellularbiology. However, it remains unclear whether flotillin proteins exist in prokaryotic cells.The amino acid sequence of the BH3500 protein from Bacillus halodurans was 30% identical
to that of flotillin-1. Motif analysis revealed that several specific residues (SPFH and flotillindomains, an AEA-repeat structure) and five potential phosphorylation sites are conservedin the BH3500 protein. In addition, the BH3500 protein was found to possess two trans-
membrane-spanning domains at the N-terminus, which is consistent with the common prop-erties of flotillin-1. The BH3500 protein was detected in the Triton-insoluble, buoyantmembrane fraction of B. halodurans by mass spectrometry and Western blotting. Interest-ingly, BH3500 was expressed strongly in alkaline conditions at both transcriptional and
translational levels, which implies that it is one of the alkali-inducible proteins.
KEY WORDS: Alkali-inducible protein; Bacillus halodurans; flotillin.
1. INTRODUCTION
Flotillin-1 and )2 are major integral membraneproteins that have been shown to be enriched ondetergent-resistant plasma membrane (DRM)microdomains (Bickel et al., 1997; Dermine et al.,2001). In many eukaryotic cell types, flotillinsappear to be markers of lipid rafts (sphingolipid-and cholesterol-rich DRM) (Bickel et al., 1997;Schulte et al., 1997). Although the molecular func-tion of flotillins is still unknown, they seem to beinvolved in organizing the structure of lipid raftsand other DRM domains (Volonte et al., 1999).Also, flotillins presumably participate in molecularsignaling across the plasma membrane, such as insu-lin signaling (Baumann et al., 2000) and T-cellreceptor signaling (Simons and Toomre, 2000).Flotillin-1 accumulates in brain cells with the
development of Alzheimer’s pathology (Kokuboet al., 2000), and flotillin-2 plays a critical role inthe structuring and maintenance of the epidermis(Schroeder et al., 1994).
Flotillins have been identified in goldfish(Schulte et al., 1997), rats (Lang et al., 1998), mice(Bickel et al., 1997; Cho et al., 1995), humans(Schroeder et al., 1994), and Drosophila (Galbiatiet al., 1998). However, a prokaryotic flotillin proteinhas not been discovered to date. In our previouswork, two epidermal surface antigen (flotillin-2)-likegenes were found in the genomic sequences of Bacillushalodurans (Takami et al., 2000) and Oceanobacillusiheyensis (Takami et al., 2002). In addition, threeflotillin-like genes were found in B. subtilis,B. cereus, and B. anthracis (Ivanova et al., 2003;Kunst et al., 1997). As a first step toward
1 Extremobiosphere Research Center, Japan Agency for Marine-
Earth Science and Technology (JAMSTEC), 2–15 Natsushima,
Yokosuka, Kanagawa 237-0061, Japan.2 To whom correspondence should be addressed. E-mail: huimin
@jamstec.go.jp
Abbreviations: SPFH, stomatin/prohibitin/flotillin/HflK/C; DRM,
detergent-resistant plasma membrane; PCR, polymerase chain
reaction; TBS, Tris-buffered saline; SDS-PAGE, sodium dodecyl
sulfate-polyacrylamide gel electrophoresis; PVDF, polyvinylidine
fluoride; PHB, prohibitin homology; IPTG, isopropyl-b-D-thioga-
lactopyranoside; FLO-2, flotillin-2; FLO-L, flotillin-like.
1251572-3887/05/0200-0125/0 � 2005 Springer Science+Business Media, Inc.
The Protein Journal, Vol. 24, No. 2, February 2005 (� 2005)DOI: 10.1007/s10930-004-1519-3

understanding the role of these flotillin-like genes inprokaryotic cells, we focused on the BH3500 gene,which was annotated as a flotillin homologue. Here,we report the sequence analysis, extraction, andpurification of the BH3500 protein, along with itsexpression pattern.
2. MATERIALS AND METHODS
2.1. Strains and Growth Conditions
Bacillus halodurans C-125 (JCM 9153) cangrow well at pH 10.5 and at pH 7 in the presenceof excess sodium chloride (1–10%). It was grownwith shaking in Horikoshi II basal medium (Takamiand Horikoshi, 1999) at 37�C. Alkalinity and/orsodium ions are necessary for good growth andmotility of the strain (Kitada and Horikoshi, 1977).For production of BH3500 protein by the strain,cells were grown in three different media: HorikoshiIIA (adjusted to pH 10 with � 2% Na2CO3), Hori-koshi IIN (containing 2% NaCl, pH 7), and Hori-koshi IIAK (adjusted to pH 10 with KOH insteadof Na2CO3). For Northern blot analysis, the bacter-ium was cultivated in Horikoshi II medium contain-ing 0.5% NaCl adjusted with KOH to various pHlevels, and the corresponding number of moles ofKCl (8–48 moles) was added to the pH 7 culture toadjust the concentration of Kþ ions. The cells wereharvested at the logarithmic phase of growth(absorbance at 600 nm � 1.0).
Escherichia coli BL21 (DE3) (Invitrogen, Carlsbad,CA) was grown at 37�C in Luria-Bertani broth con-taining 100 mg/l ampicillin to an absorbance at600 nm of � 0.5, and then the expression of theBH3500 gene was induced with 0.5 mM isopropyl-b-D-thiogalactopyranoside (IPTG) for 3–5 h. Thecells were harvested by centrifugation, and lysed byB-PER Bacterial Protein Extraction Reagent (PierceBiotechnology, Rockford, IL).
2.2. Cloning, Overexpression, and Purification of the
BH3500 Protein
The BH3500 gene fragment was amplified by PCRusing a specific primer set (50-CCATATGACGGTTGTGACGTTTCTTGC-30 and 50-TCCGTTATTTTTCTTCACTGTCACTG-30) possessing theNdeI (GGAATT) or BamHI (CGCGGA) restrictionsite on the 50 side. After digestion and purification,
the whole BH3500 gene was ligated with the NdeIand BamHI sites of the pET-15b vector carrying anN-terminal His-Tag sequence (Novagen, Madison,WI). E. coli BL21 (DE3) was transformed with theligation mixture. The coding sequence for theBH3500 gene was verified by direct sequencing.
Crude recombinant BH3500 protein was sus-pended in 100 mM phosphate buffer (pH 8.0) plus8 M urea. The His-tagged proteins were purified byaffinity chromatography using a Ni-NTA agarosecolumn (U 0.7 cm, 1.5 cm, QIAGEN, Valencia,CA), and identification was confirmed by massspectrometry.
2.3. Extraction of DRM from B. halodurans C-125
DRM fractions containing only integral pro-teins were separated as described previously (Salzerand Prohaska, 2001). The bacterial cells were dis-rupted by a French press with ice-cold 1% TritonX-100 in lysis buffer (10 mM Tris-HCl, pH 7.4;RNase A; DNase I; PMSF; MgCl2), incubated for20 min on ice, and centrifuged for 10 min at 4�C(15,000 g). The pellet was resuspended in ice-cold1% Triton X-100 in TBS (10 mM Tris-HCl,150 mM NaCl, pH 7.5) and mixed with an equalvolume of 80% sucrose in 0.2 M Na2CO3. A totalof 500 ll of this suspension was transferred to ultra-centrifuge tubes, overlaid with 1.5 ml 30% sucrosein TBS and then with 1 ml 10% sucrose in TBS.The mixture was centrifuged in an SW55Ti rotor(Beckman, Fullerton, CA) for 17 h at 4�C(230,000 g). Fractions (500 ll) were collected fromthe top; the DRM fractions were diluted with anequal volume of TBS and centrifuged at 100,000 gfor 1 h. Aliquots of these fractions were subjectedto 5–20% gradient SDS-PAGE electrophoresis.
2.4. Mass Spectrometry and Western Blot Analysis
The proteins were manually excised from thegel and digested with trypsin (12.5 ng ll; Promega,Madison, WI). Mass spectrometric analysis was per-formed as described previously (Ohta et al., 2004).For Western blot analysis, the purified recombinantBH3500 protein was used to immunize rabbits withFreund’s adjuvant. The anti-BH3500 antibody waspurified using a PD-10 column (Amersham PharmaciaBiotech, Piscataway, NJ). DRM was prepared bysucrose density gradient centrifugation. It was
126 Zhang, Li, Tsudome, Ito, Takami, and Horikoshi

separated by 5–20% gradient SDS-PAGE and elec-tro-transferred to PVDF membranes (Amersham).The membranes were subsequently incubated withthe rabbit anti-BH3500 antibody (1:5000 dilution)and assayed for anti-rabbit IgG secondary antibodyconjugated to horseradish peroxidase (Promega).
2.5. Northern Blot Analysis
After cultivation under various pH conditions,B. halodurans cells were harvested, and total RNAswere isolated by the method of Igo and Losick (Igoand Losick, 1986). BH3500 cDNA probes werelabeled with primers (50-TGGTGTTCCGAT-CATTGCTG-30; 50-ATTGCTCTGCTTTTGCTCTCG-30) with digoxigenin-dUTP using a DIG DNALabeling and Detection kit (Roche Diagnostics,Indianapolis, IN). Hybridization signals weredetected as per the manufacturer’s instructions.
3. RESULTS AND DISCUSSION
3.1. Comparative Analysis of BH3500 Sequence
The amino acid sequence of the BH3500 proteinwas obtained from ExtremoBase (http://www.jamstec.go.jp/jamstec-j/bio/exbase.html), and sequence align-ment was performed with ClustalX software. Figure 1shows the alignment of the BH3500 protein sequencewith known protein sequences of human flotillins. TheBH3500 protein showed 30% identity to human flotil-lin-1 (accession no. NM_005803) and 29% identity tohuman flotillin-2 (NM_004475).
Flotillins usually contain several specific resi-dues and potential phosphorylation sites. These spe-cific residues and sites are generally known to becharacteristic of the proteins categorized into theflotillin family (Galbiati et al., 1998; Morrow et al.,2002; Schroeder et al., 1994; Tavernarakis et al.,1999). To examine whether these residues and sitesalso exist in BH3500, motif scans were donethrough access via the Hits homepage (http://hits.isb-sib.ch) (Fig. 1). Firstly, the N-terminus ofthe BH3500 protein contained an SPFH (stomatin/prohibitin/flotillin/HflK/C)-domain in the regionfrom the 90th to the 231st amino acid (aa), and thedomain showed 36 and 34% identity with humanflotillin-1 and flotillin-2, respectively. The SPFHdomain is also called the prohibitin homology
(PHB) domain and was recently identified in humanflotillins (Morrow et al., 2002; Tavernarakis et al.,1999). Secondly, the flotillin domain, which is astructural motif of the flotillin protein (Schroederet al., 1994), was observed in the region from the236th to the 412th aa of the BH3500 protein. Thedomain shows 32 and 27% identity with humanflotillin-1 and flotillin-2, respectively. Thirdly, in theC-terminal side of the BH3500 protein, an AEA-repeat structure was observed, as in the cases ofother flotillin proteins. Such a repeat is of structuralinterest and is predicted to adopt an a-helical con-formation in human flotillins (Schroeder et al.,1994). Finally, BH3500 protein contains two poten-tial casein kinase II phosphorylation sites, twopotential tyrosine kinase phosphorylation sites, anda potential protein kinase C phosphorylation site.These sites are widely conserved in flotillins fromflies to mammals (Galbiati et al., 1998).
The SPFH/PHB domain is shared by severaleukaryotic and prokaryotic membrane proteins,including stomatin, podocin, and prohibitin, whichhave apparently diverse functions (Tavernarakiset al., 1999). Flotillin is a transmembrane proteinwith a hydrophobic region in the SPFH/PHBdomain (Gkantiragas et al., 2001). Flotillin differsfrom podocin, stomatin, and prohibitin in that ithas a well-defined hydrophobic putative intramem-brane domain. Hydropathy analysis of the BH3500protein was performed using the Tmpred program(http://www.ch.embnet.org/software/TMPRED_form.html). It was suggested that two hydrophobicdomains (the 1st to the 28th aa and the 102nd tothe 128th aa) might function as potential transmem-brane-spanning domains. The hydrophobic regionfrom the 102nd to the 128th aa of the BH3500 pro-tein corresponds to the PHB domain. This is alsoconsistent with the common properties of flotillin-1(Bickel et al., 1997). These results strongly suggestthat the BH3500 protein might be a newly discov-ered ancestral member of the flotillin family.
To further clarify the evolutionary relationship,phylogenetic analysis was performed. Although theneighbor-joining tree defines four major groups,BH3500 did not belong to known eukaryoticflotillin-1 and flotillin-2 groups, suggesting a noveltype of flotillin (Fig. 2). BH3500 formed a newgroup with O. iheyensis. B. halodurans and O. iheyensisare similar in alkaliphilic phenotype, the relation-ship between the two flotillin-like proteins andalkaliphily remains to be further clarified.
127Alkali-Inducible Flotillin-like Protein

3.2. Purification and Characterization of BH3500
Protein
Approximately 40 lg of recombinant BH3500 pro-tein (58 kDa) was obtained from 1 ml of culture(absorbance at 600 nm � 0.7) (Fig. 3a). The proteinwas insoluble in Triton X-100 at 4�C, showing acommon characteristic of flotillin. On the other
hand, the crude extract of B. halodurans was incu-bated with Trion X-100 on ice, followed by centri-fugation in order to separate the detergent-insolublematerials from the soluble membrane proteins andother matter. In order to distinguish between theintegral and peripheral membrane proteins, thepellet was mixed with buffer containing 0.1 MNa2CO3 because peripheral membrane proteins are
Fig. 1. Multiple amino acid sequence alignment of BH3500. The sequence of the BH3500 protein is compared with human flotillin-1
(h.FLO-1) and flotillin-2 (h.FLO-2). The SPFH-domain is shaded in gray and the flotillin-domain is boxed and shaded in gray. The poten-
tial sites for phosphorylation by casein kinase II (@), protein kinase C (U), the tyrosine kinases (#), and the C-terminal AEA-repeat struc-
ture (4) are indicated. *, identical amino acids in all three sequences; :, conserved substitutions; ., semi-conserved substitutions; -, gap in
the sequence.
128 Zhang, Li, Tsudome, Ito, Takami, and Horikoshi

dissociated from membranes by alkaline extraction,while integral membrane proteins remain firmlyattached. Step gradient ultracentrifugation of thispellet yielded an opalescent band floating in thelow-density region of the gradient (10–30% sucroseinterface). The eukaryotic flotillins have beencharacteristically observed in these low-densityDRM complexes (Salzer and Prohaska, 2001). Thefractions were collected and analyzed by mass spec-trometry and Western blotting. The prominent58 kDa band in the low-density fraction (fraction 3)was identified as the BH3500 protein by mass spec-trometry (Fig. 3b and c) and with the help of ananti-BH3500 rabbit polyclonal antibody (Fig. 3d).The BH3500 protein is a Triton-insoluble, buoyant,integral membrane protein, further confirming thatit would be categorized as a new member of theflotillin family.
3.3. Expression of BH3500 Gene
Western blot analysis showed that the rate ofgeneration of the BH3500 protein at pH 10 wasabout three times greater than at pH 7.0 (Fig. 3Dand E). This increase was also observed at pH 10
without sodium ions, suggesting that the increase inthe amount of BH3500 protein is alkaline- but notsodium-dependent.
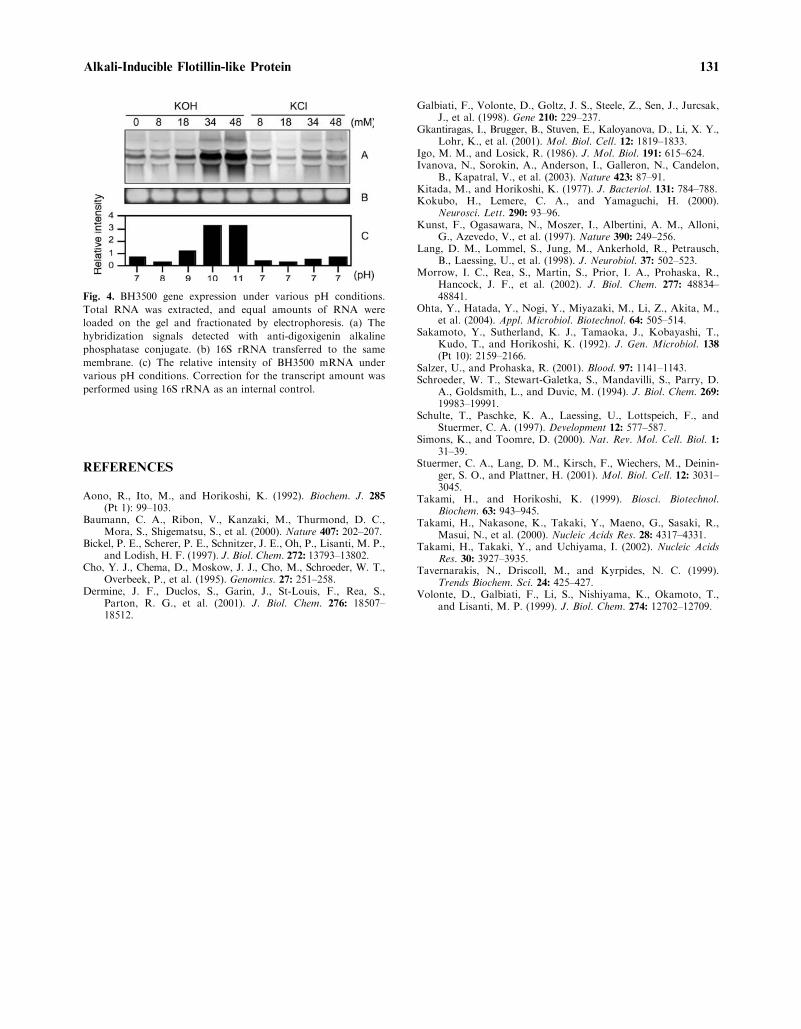
Northern blot analysis was performed toexamine the expression pattern of the BH3500gene at the transcriptional level under various pHconditions (Fig. 4). To eliminate the influence ofsodium ions, cells were cultured with low levels ofsodium (0.5% NaCl). A single transcript of about1.5 kb was detected during growth at pH 7–11.The mRNA of the BH3500 gene was significantlyup-regulated under alkaline conditions (pH10–11). The level of expression at pH 11 was upto approximately 4.5 times that at pH 7. Thisincreased expression was not observed when Kþ
ion used as a control at pH 7. These findingssupport the view that the BH3500 protein shouldbe alkali-inducible.
Bacillus halodurans is an alkaliphilic bacteriumthat can grow in a wide range of pH conditions (pH7–11) (Takami and Horikoshi, 1999). The flagellin(hag) of the organism was reported to be alkali-induc-ible (Aono et al., 1992). Moreover, Sakamoto et al.,have demonstrated that B. halodurans cells swim fasterunder alkaline conditions than under neutral conditions(Sakamoto et al., 1992). We have no idea at presentwhether the BH3500 protein is directly related to thealkaliphilic phenotype. In mammalian cells, flotillins play
0.1
Drosophila FLO-2
goldfish FLO-2
rat FLO-2
mouse FLO-2human FLO-2
B.subtilis FLO-LB.anthracis FLO-L
B.cereus FLO-L
B.halodurans BH3500
O. iheyensis FLO-L
Drosophila FLO-1
goldfish FLO-1
mouse FLO-1rat FLO-1
human FLO-1
Fig. 2. Phylogenetic analysis of BH3500 protein. An alignment of full-length coding sequences was used for construction of the phyloge-
netic tree. The accession numbers for the flotillin-1 (FLO-1) sequences of Drosophila, goldfish, mouse, rat, and human are AF044734,
U33556, NM_008027, U60976, and NM_005803, respectively; the accession numbers for the flotillin-2 (FLO-2) sequences in Drosophila,
goldfish, mouse, rat, and human are AF044916, L36867, NM_008028, AF023303, and NM_004475, respectively; the accession numbers
for the flotillin-like (FLO-L) sequences in B. subtilis, B. cereus, B. anthracis, and O. iheyensis are Z99119, AE017266, AAT29654, and
OB0205, respectively. Bar, 0.1 Knuc unit.
129Alkali-Inducible Flotillin-like Protein

a central role in cellular organization (Bickel et al., 1997;Schroeder et al., 1994), membrane trafficking events(Morrow et al., 2002), and specific signaling cascades(Lang et al., 1998; Stuermer et al., 2001). It is unclear whatbiological roles BH3500 protein plays in prokaryotes. Toclarify these questions, gene knockout experiments to
disrupt the BH3500 gene in B. halodurans by homologousrecombination have being carried out. Further proteomeand transcriptome analyses using gene knockout mutantsare necessary. In any event, the flotillin-like proteins thatare conserved even in prokaryotic cells seem to be veryactive across the plasmamembrane in fundamental ways.
Fig. 3. Characterization of BH3500 protein. (a) The expression of the His-tagged BH3500 protein in E. coli. (b) Separation of DRM frac-
tions by sucrose-gradient centrifugation. Six 500-ll fractions (lanes 1–6) were collected from the top. Aliquots of these samples were ana-
lyzed by polyacrylamide gel electrophoresis. (c) Identification by mass spectrometry of the 58 kDa protein (*) present in the low-density
fraction (lane 3). The bold and underlined letters indicate those peptides identified by mass spectrometry and linked to the BH3500
sequence. (d) Detection of the BH3500 protein by Western blotting. Lane 1, Horikoshi IIAK; lane 2, Horikoshi IIA; lane 3, Horikoshi
IIN. (E) Relative intensity of Western blotting.
130 Zhang, Li, Tsudome, Ito, Takami, and Horikoshi
REFERENCES
Aono, R., Ito, M., and Horikoshi, K. (1992). Biochem. J. 285
(Pt 1): 99–103.Baumann, C. A., Ribon, V., Kanzaki, M., Thurmond, D. C.,
Mora, S., Shigematsu, S., et al. (2000). Nature 407: 202–207.Bickel, P. E., Scherer, P. E., Schnitzer, J. E., Oh, P., Lisanti, M. P.,
and Lodish, H. F. (1997). J. Biol. Chem. 272: 13793–13802.Cho, Y. J., Chema, D., Moskow, J. J., Cho, M., Schroeder, W. T.,
Overbeek, P., et al. (1995). Genomics. 27: 251–258.Dermine, J. F., Duclos, S., Garin, J., St-Louis, F., Rea, S.,
Parton, R. G., et al. (2001). J. Biol. Chem. 276: 18507–18512.
Galbiati, F., Volonte, D., Goltz, J. S., Steele, Z., Sen, J., Jurcsak,J., et al. (1998). Gene 210: 229–237.
Gkantiragas, I., Brugger, B., Stuven, E., Kaloyanova, D., Li, X. Y.,Lohr, K., et al. (2001). Mol. Biol. Cell. 12: 1819–1833.
Igo, M. M., and Losick, R. (1986). J. Mol. Biol. 191: 615–624.Ivanova, N., Sorokin, A., Anderson, I., Galleron, N., Candelon,
B., Kapatral, V., et al. (2003). Nature 423: 87–91.Kitada, M., and Horikoshi, K. (1977). J. Bacteriol. 131: 784–788.Kokubo, H., Lemere, C. A., and Yamaguchi, H. (2000).
Neurosci. Lett. 290: 93–96.Kunst, F., Ogasawara, N., Moszer, I., Albertini, A. M., Alloni,
G., Azevedo, V., et al. (1997). Nature 390: 249–256.Lang, D. M., Lommel, S., Jung, M., Ankerhold, R., Petrausch,
B., Laessing, U., et al. (1998). J. Neurobiol. 37: 502–523.Morrow, I. C., Rea, S., Martin, S., Prior, I. A., Prohaska, R.,
Hancock, J. F., et al. (2002). J. Biol. Chem. 277: 48834–48841.
Ohta, Y., Hatada, Y., Nogi, Y., Miyazaki, M., Li, Z., Akita, M.,et al. (2004). Appl. Microbiol. Biotechnol. 64: 505–514.
Sakamoto, Y., Sutherland, K. J., Tamaoka, J., Kobayashi, T.,Kudo, T., and Horikoshi, K. (1992). J. Gen. Microbiol. 138(Pt 10): 2159–2166.
Salzer, U., and Prohaska, R. (2001). Blood. 97: 1141–1143.Schroeder, W. T., Stewart-Galetka, S., Mandavilli, S., Parry, D.
A., Goldsmith, L., and Duvic, M. (1994). J. Biol. Chem. 269:19983–19991.
Schulte, T., Paschke, K. A., Laessing, U., Lottspeich, F., andStuermer, C. A. (1997). Development 12: 577–587.
Simons, K., and Toomre, D. (2000). Nat. Rev. Mol. Cell. Biol. 1:31–39.
Stuermer, C. A., Lang, D. M., Kirsch, F., Wiechers, M., Deinin-ger, S. O., and Plattner, H. (2001). Mol. Biol. Cell. 12: 3031–3045.
Takami, H., and Horikoshi, K. (1999). Biosci. Biotechnol.Biochem. 63: 943–945.
Takami, H., Nakasone, K., Takaki, Y., Maeno, G., Sasaki, R.,Masui, N., et al. (2000). Nucleic Acids Res. 28: 4317–4331.
Takami, H., Takaki, Y., and Uchiyama, I. (2002). Nucleic AcidsRes. 30: 3927–3935.
Tavernarakis, N., Driscoll, M., and Kyrpides, N. C. (1999).Trends Biochem. Sci. 24: 425–427.
Volonte, D., Galbiati, F., Li, S., Nishiyama, K., Okamoto, T.,and Lisanti, M. P. (1999). J. Biol. Chem. 274: 12702–12709.
Fig. 4. BH3500 gene expression under various pH conditions.
Total RNA was extracted, and equal amounts of RNA were
loaded on the gel and fractionated by electrophoresis. (a) The
hybridization signals detected with anti-digoxigenin alkaline
phosphatase conjugate. (b) 16S rRNA transferred to the same
membrane. (c) The relative intensity of BH3500 mRNA under
various pH conditions. Correction for the transcript amount was
performed using 16S rRNA as an internal control.
131Alkali-Inducible Flotillin-like Protein