altered odor-induced expression of c-fos and arg 3.1 immediate early genes in the olfactory system...
TRANSCRIPT

Altered Odor-Induced Expression of c-fos and arg3.1 Immediate Early Genes in the Olfactory Systemafter Familiarization with an Odor
M. Montag-Sallaz,1 N. Buonviso2
1 Research Group Neurogenetics, Leibniz Institute for Neurobiology, D-39118 Magdeburg, Germany
2 Neurosciences et systemes sensoriels, CNRS-Universite Claude Bernard Lyon I, F-69622Villeurbanne Cedex, France
Received 16 October 2001; accepted 15 February 2002
ABSTRACT: In adult rats, repeated exposure toan odorant, in absence of any experimentally deliveredreinforcement, leads to a drastic decrease in mitral/tufted (M/T) cell responsiveness, not only for the famil-iar odor but also for other novel odors. In the presentstudy, using two different and complementary in situhybridization methods, we analyzed the effect of famil-iarization with an odorant on c-fos and arg 3.1 mRNAexpression levels, and we examined the odor specificityof this effect. Odor exposure induces a specific increasein c-fos and arg 3.1 expression in some particular olfac-tory bulb quadrants. Previous familiarization with thetest odor results in a decreased expression of both IEGsin these quadrants, leading to the alteration of the odor-specific pattern of c-fos and arg 3.1 expression. In con-
trast, this odor-specific pattern is not affected whendifferent odors are used for familiarization and test.Similarly, an odor-specific familiarization effect leadingto a reduced c-fos and arg 3.1 expression was also de-tected in the cingulate cortex and in the anterior piri-form cortex. These results support our hypothesis thatthe decrease in M/T cell responsiveness following a pre-ceding familiarization with an odorant may be related toa particular form of synaptic plasticity involvingchanges at the genomic level, and reveals further insightin olfactory information processing and the cellularmechanisms underlying familiarization in the olfactorysystem. © 2002 Wiley Periodicals, Inc. J Neurobiol 52: 61–72, 2002
Keywords: olfactory bulb; familiarization; neural plas-ticity; novelty; piriform cortex
INTRODUCTION
Recently, we reported that a simple exposure of adultrats to an odorant, in the absence of any experimen-tally delivered reinforcement, leads to a drastic de-crease in mitral/tufted (M/T) cell responsiveness(Buonviso et al., 1998; Buonviso and Chaput, 2000).Surprisingly, excitatory responses were decreased notonly to the familiar odor but also to other novel odors.These effects were (1) rapidly induced, because they
can appear after the first exposure to isoamyl acetate,(2) long-lasting as they were still observed 10 daysafter the end of exposure. To explain these findings,several hypotheses were considered (Buonviso et al.,1998; Buonviso and Chaput, 2000), but the underly-ing mechanisms remained unknown. The characteris-tics of the decrease in M/T cell responsiveness fol-lowing familiarization with an odorant indicate thatthis effect may be related to a particular form ofsynaptic plasticity. The implication of immediateearly genes (IEGs) in synaptic plasticity and memoryformation has been widely described (for review, seeHerdegen and Leah, 1998; Tischmeyer and Grimm,1999). Expression of c-fos and c-jun, two proto-on-cogenes among the best characterized modulator
Correspondence to: M. Montag-Sallaz ([email protected])© 2002 Wiley Periodicals, Inc.Published online in Wiley InterScience (www.interscience.wiley.com).DOI 10.1002/neu.10069
61

genes (Kaczmarek and Nikolajew, 1990; Silva andGiese, 1994), has been well studied in the olfactorysystem. It was reported that a 20–30 min exposure toan odor induced differential expression of these twotranscription factors in the glomerular and/or granulelayers of the olfactory bulb (OB) (Sallaz and Jourdan,1993; Guthrie and Gall, 1995; Baba et al., 1997).Complementary studies showed that the c-fos expres-sion pattern in the OB after presentation of an odorantis odor-specific, and depends not only on the afferentolfactory input, but also on the cholinergic and nor-adrenergic centrifugal systems (Sallaz and Jourdan,1996), both functionally involved in mechanisms ofOB plasticity associated with olfactory learning (Ger-vais et al., 1988; Ravel et al., 1992). In the context ofolfactory familiarization, the expression of anotherIEG is particularly interesting to study. The so-calledactivity-regulated gene 3.1 (arg 3.1; Link et al., 1995)or activity-regulated cytoskeleton-associated protein(arc; Lyford et al., 1995) has been recently identifiedin brain using differential cloning techniques, on thebasis of its rapid induction after seizure stimulation.In adult rats, arg 3.1 expression can be triggered indifferent manners: metrazole-induced seizure, high-frequency stimulation of the perforant path inducinglong-term potentiation (Link et al., 1995; Lyford etal., 1995), administration of cocaine, amphetamine(Fosnaugh et al., 1995; Tan et al., 2000), and dopa-mine (Berke et al., 1998; Pei et al., 2000) were all ableto induce an increase in arg 3.1 expression in severalbrain areas. A peculiarity of arg 3.1 is that its mRNAselectively localizes in activated dendritic segments(Steward et al., 1998: Wallace et al., 1998), suggest-ing that the Arg 3.1 protein may be synthesized indendrites at activated postsynaptic sites. In the OB,the expression of arg 3.1 mRNA is increased in neu-rons selectively activated by an odorant, and the Arg3.1 protein is localized in spine-like processes in theexternal plexiform layer and in granule cell bodiesand dendrites, supporting the hypothesis that changesin expression of this effector gene may underlie someof the morphological effects of sensory experienceseen in this system (Guthrie et al., 2000). Finally,using a gustatory stimulus, we showed previously thatthe expression of c-fos and of arg 3.1 is triggered bythe novelty of the stimulus, and that it is sensitive topreliminary familiarization with the same stimulus(Montag-Sallaz et al., 1999). In summary, these datasuggested c-fos and arg 3.1 as good tools to studymolecular mechanisms underlying synaptic plasticityinvolved in familiarization with an olfactory stimulus.
To test the hypothesis that the decrease in M/T cellresponsiveness following a preceding familiarizationwith an odorant may be related to a particular form of
synaptic plasticity, we examined the effect of a pre-ceding odor familiarization on c-fos and arg 3.1mRNA expression in the OB and different brain areasin response to an odorant, and we analyzed the odor-specificity of this effect.
METHODS
Our experiments were carried out in accordance with theEuropean Communities Council directives of 24 November1986 (86/609/EEC).
Animals and Exposure to Odorants
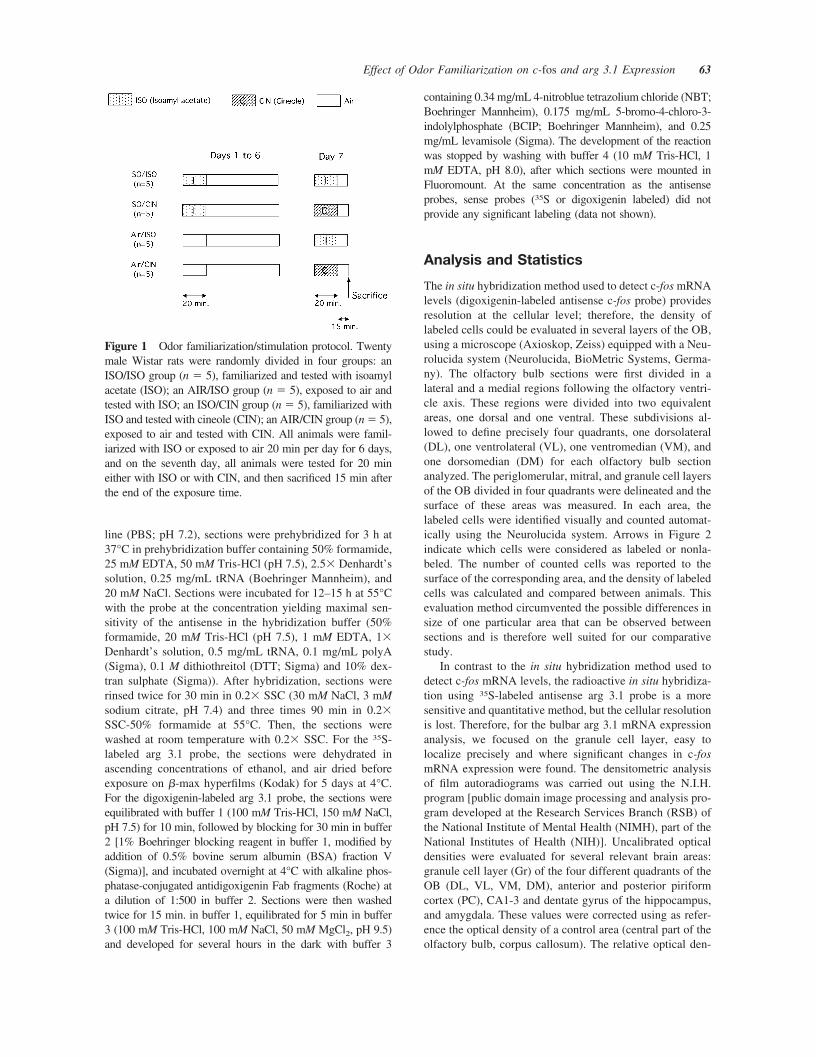
Experiments were performed on 20 male Wistar rats (IFFA-Credo) weighing 250–300 g at the first day of exposure.They were housed in our facilities under standard conditions(12 h:12 h inverted light:dark cycle with light off at 0600;21 � 2°C room temperature, free access to food and water).For exposure, rats were taken from their home cage andplaced individually for 20 min per day in a 50 � 50-cmwhite plastic cage opened to room air where they wereallowed to explore freely. Half of the animals (n � 10)received a transient odor experience with one odor only(ISO, isoamyl acetate, Sigma-Aldrich, L’Isle d’Abeau) over6 consecutive days, which corresponds to the time taken foracquisition of classical associative olfactory tasks (Freemanand Schneider, 1982). For the animals of this group, thelitter of the exposure cage was previously odorized by threedrops of the undiluted compound in one corner. As control,the second half of the rats were similarly handled, but noodorant was given in the exposure cage during the 6 daysfamiliarization period. On the seventh day, five familiarizedand five control rats were tested with ISO (ISO/ISO n � 5,AIR/ISO n � 5, respectively; Fig. 1). In parallel, fivefamiliarized and five control rats were tested with cineole(CIN, Sigma-Aldrich, L’Isle d’Abeau), an odorant differentfrom the ISO used for familiarization (ISO/CIN n � 5,AIR/CIN n � 5, respectively; Fig. 1). On this last experi-mental day, 15 min after the end of the 20-min stimulation,all rats were deeply anesthetized (equithesine IP, 1 mL/300g weight) and sacrificed by decapitation.
In Situ Hybridization
Levels of c-fos and arg 3.1 mRNAs were examined in tissuesections using in situ hybridization of digoxigenin- and35S-labeled antisense cRNA probes, respectively. Sense andantisense c-fos and arg 3.1 probes were generated as de-scribed (Montag-Sallaz et al., 1999). Brain tissue was pro-cessed for in situ hybridization as described previously(Montag-Sallaz et al., 1999). Briefly, brains were removedand quickly frozen by immersion in isopentane (�40°C).Frontal sections (14 �m thick) were cut in a cryostat at�15°C and collected on silane-coated glass slides. Afterfixation in 4% paraformaldehyde in phosphate-buffered sa-
62 Montag-Sallaz and Buonviso
line (PBS; pH 7.2), sections were prehybridized for 3 h at37°C in prehybridization buffer containing 50% formamide,25 mM EDTA, 50 mM Tris-HCl (pH 7.5), 2.5� Denhardt’ssolution, 0.25 mg/mL tRNA (Boehringer Mannheim), and20 mM NaCl. Sections were incubated for 12–15 h at 55°Cwith the probe at the concentration yielding maximal sen-sitivity of the antisense in the hybridization buffer (50%formamide, 20 mM Tris-HCl (pH 7.5), 1 mM EDTA, 1�Denhardt’s solution, 0.5 mg/mL tRNA, 0.1 mg/mL polyA(Sigma), 0.1 M dithiothreitol (DTT; Sigma) and 10% dex-tran sulphate (Sigma)). After hybridization, sections wererinsed twice for 30 min in 0.2� SSC (30 mM NaCl, 3 mMsodium citrate, pH 7.4) and three times 90 min in 0.2�SSC-50% formamide at 55°C. Then, the sections werewashed at room temperature with 0.2� SSC. For the 35S-labeled arg 3.1 probe, the sections were dehydrated inascending concentrations of ethanol, and air dried beforeexposure on �-max hyperfilms (Kodak) for 5 days at 4°C.For the digoxigenin-labeled arg 3.1 probe, the sections wereequilibrated with buffer 1 (100 mM Tris-HCl, 150 mM NaCl,pH 7.5) for 10 min, followed by blocking for 30 min in buffer2 [1% Boehringer blocking reagent in buffer 1, modified byaddition of 0.5% bovine serum albumin (BSA) fraction V(Sigma)], and incubated overnight at 4°C with alkaline phos-phatase-conjugated antidigoxigenin Fab fragments (Roche) ata dilution of 1:500 in buffer 2. Sections were then washedtwice for 15 min. in buffer 1, equilibrated for 5 min in buffer3 (100 mM Tris-HCl, 100 mM NaCl, 50 mM MgCl2, pH 9.5)and developed for several hours in the dark with buffer 3
containing 0.34 mg/mL 4-nitroblue tetrazolium chloride (NBT;Boehringer Mannheim), 0.175 mg/mL 5-bromo-4-chloro-3-indolylphosphate (BCIP; Boehringer Mannheim), and 0.25mg/mL levamisole (Sigma). The development of the reactionwas stopped by washing with buffer 4 (10 mM Tris-HCl, 1mM EDTA, pH 8.0), after which sections were mounted inFluoromount. At the same concentration as the antisenseprobes, sense probes (35S or digoxigenin labeled) did notprovide any significant labeling (data not shown).
Analysis and Statistics
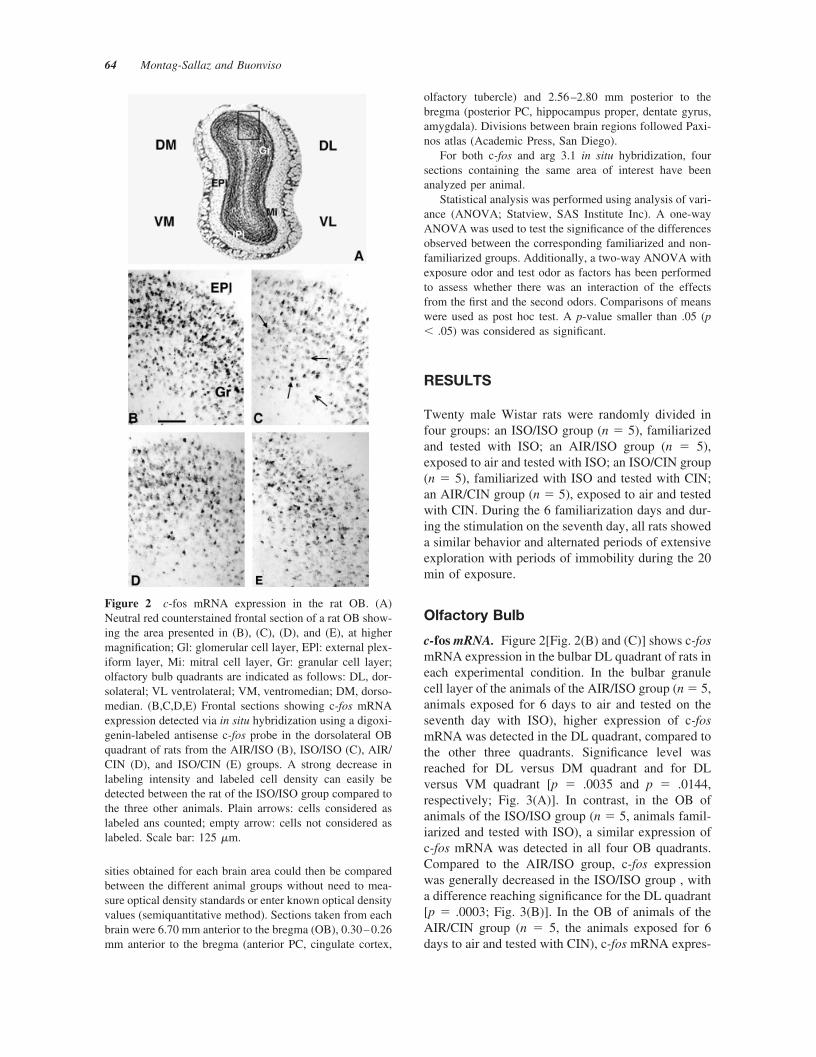
The in situ hybridization method used to detect c-fos mRNAlevels (digoxigenin-labeled antisense c-fos probe) providesresolution at the cellular level; therefore, the density oflabeled cells could be evaluated in several layers of the OB,using a microscope (Axioskop, Zeiss) equipped with a Neu-rolucida system (Neurolucida, BioMetric Systems, Germa-ny). The olfactory bulb sections were first divided in alateral and a medial regions following the olfactory ventri-cle axis. These regions were divided into two equivalentareas, one dorsal and one ventral. These subdivisions al-lowed to define precisely four quadrants, one dorsolateral(DL), one ventrolateral (VL), one ventromedian (VM), andone dorsomedian (DM) for each olfactory bulb sectionanalyzed. The periglomerular, mitral, and granule cell layersof the OB divided in four quadrants were delineated and thesurface of these areas was measured. In each area, thelabeled cells were identified visually and counted automat-ically using the Neurolucida system. Arrows in Figure 2indicate which cells were considered as labeled or nonla-beled. The number of counted cells was reported to thesurface of the corresponding area, and the density of labeledcells was calculated and compared between animals. Thisevaluation method circumvented the possible differences insize of one particular area that can be observed betweensections and is therefore well suited for our comparativestudy.
In contrast to the in situ hybridization method used todetect c-fos mRNA levels, the radioactive in situ hybridiza-tion using 35S-labeled antisense arg 3.1 probe is a moresensitive and quantitative method, but the cellular resolutionis lost. Therefore, for the bulbar arg 3.1 mRNA expressionanalysis, we focused on the granule cell layer, easy tolocalize precisely and where significant changes in c-fosmRNA expression were found. The densitometric analysisof film autoradiograms was carried out using the N.I.H.program [public domain image processing and analysis pro-gram developed at the Research Services Branch (RSB) ofthe National Institute of Mental Health (NIMH), part of theNational Institutes of Health (NIH)]. Uncalibrated opticaldensities were evaluated for several relevant brain areas:granule cell layer (Gr) of the four different quadrants of theOB (DL, VL, VM, DM), anterior and posterior piriformcortex (PC), CA1-3 and dentate gyrus of the hippocampus,and amygdala. These values were corrected using as refer-ence the optical density of a control area (central part of theolfactory bulb, corpus callosum). The relative optical den-
Figure 1 Odor familiarization/stimulation protocol. Twentymale Wistar rats were randomly divided in four groups: anISO/ISO group (n � 5), familiarized and tested with isoamylacetate (ISO); an AIR/ISO group (n � 5), exposed to air andtested with ISO; an ISO/CIN group (n � 5), familiarized withISO and tested with cineole (CIN); an AIR/CIN group (n � 5),exposed to air and tested with CIN. All animals were famil-iarized with ISO or exposed to air 20 min per day for 6 days,and on the seventh day, all animals were tested for 20 mineither with ISO or with CIN, and then sacrificed 15 min afterthe end of the exposure time.
Effect of Odor Familiarization on c-fos and arg 3.1 Expression 63
sities obtained for each brain area could then be comparedbetween the different animal groups without need to mea-sure optical density standards or enter known optical densityvalues (semiquantitative method). Sections taken from eachbrain were 6.70 mm anterior to the bregma (OB), 0.30–0.26mm anterior to the bregma (anterior PC, cingulate cortex,
olfactory tubercle) and 2.56–2.80 mm posterior to thebregma (posterior PC, hippocampus proper, dentate gyrus,amygdala). Divisions between brain regions followed Paxi-nos atlas (Academic Press, San Diego).
For both c-fos and arg 3.1 in situ hybridization, foursections containing the same area of interest have beenanalyzed per animal.
Statistical analysis was performed using analysis of vari-ance (ANOVA; Statview, SAS Institute Inc). A one-wayANOVA was used to test the significance of the differencesobserved between the corresponding familiarized and non-familiarized groups. Additionally, a two-way ANOVA withexposure odor and test odor as factors has been performedto assess whether there was an interaction of the effectsfrom the first and the second odors. Comparisons of meanswere used as post hoc test. A p-value smaller than .05 (p� .05) was considered as significant.
RESULTS
Twenty male Wistar rats were randomly divided infour groups: an ISO/ISO group (n � 5), familiarizedand tested with ISO; an AIR/ISO group (n � 5),exposed to air and tested with ISO; an ISO/CIN group(n � 5), familiarized with ISO and tested with CIN;an AIR/CIN group (n � 5), exposed to air and testedwith CIN. During the 6 familiarization days and dur-ing the stimulation on the seventh day, all rats showeda similar behavior and alternated periods of extensiveexploration with periods of immobility during the 20min of exposure.
Olfactory Bulb
c-fos mRNA. Figure 2[Fig. 2(B) and (C)] shows c-fosmRNA expression in the bulbar DL quadrant of rats ineach experimental condition. In the bulbar granulecell layer of the animals of the AIR/ISO group (n � 5,animals exposed for 6 days to air and tested on theseventh day with ISO), higher expression of c-fosmRNA was detected in the DL quadrant, compared tothe other three quadrants. Significance level wasreached for DL versus DM quadrant and for DLversus VM quadrant [p � .0035 and p � .0144,respectively; Fig. 3(A)]. In contrast, in the OB ofanimals of the ISO/ISO group (n � 5, animals famil-iarized and tested with ISO), a similar expression ofc-fos mRNA was detected in all four OB quadrants.Compared to the AIR/ISO group, c-fos expressionwas generally decreased in the ISO/ISO group , witha difference reaching significance for the DL quadrant[p � .0003; Fig. 3(B)]. In the OB of animals of theAIR/CIN group (n � 5, the animals exposed for 6days to air and tested with CIN), c-fos mRNA expres-
Figure 2 c-fos mRNA expression in the rat OB. (A)Neutral red counterstained frontal section of a rat OB show-ing the area presented in (B), (C), (D), and (E), at highermagnification; Gl: glomerular cell layer, EPl: external plex-iform layer, Mi: mitral cell layer, Gr: granular cell layer;olfactory bulb quadrants are indicated as follows: DL, dor-solateral; VL ventrolateral; VM, ventromedian; DM, dorso-median. (B,C,D,E) Frontal sections showing c-fos mRNAexpression detected via in situ hybridization using a digoxi-genin-labeled antisense c-fos probe in the dorsolateral OBquadrant of rats from the AIR/ISO (B), ISO/ISO (C), AIR/CIN (D), and ISO/CIN (E) groups. A strong decrease inlabeling intensity and labeled cell density can easily bedetected between the rat of the ISO/ISO group compared tothe three other animals. Plain arrows: cells considered aslabeled ans counted; empty arrow: cells not considered aslabeled. Scale bar: 125 �m.
64 Montag-Sallaz and Buonviso
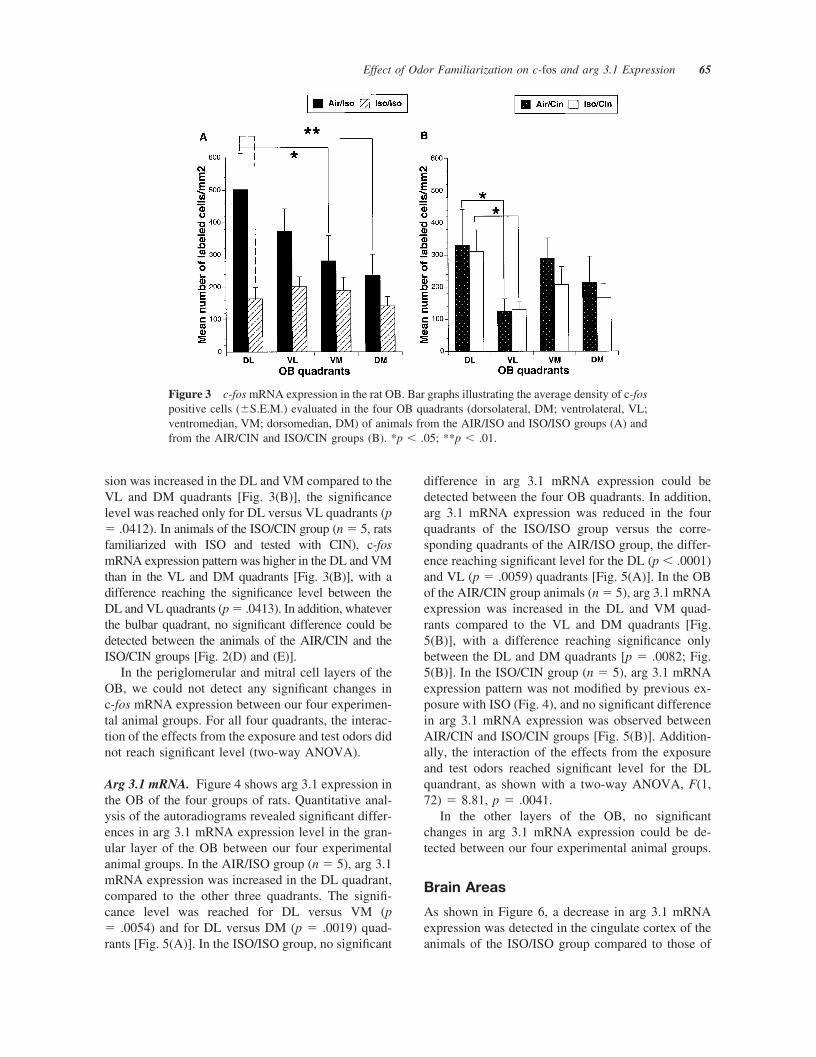
sion was increased in the DL and VM compared to theVL and DM quadrants [Fig. 3(B)], the significancelevel was reached only for DL versus VL quadrants (p� .0412). In animals of the ISO/CIN group (n � 5, ratsfamiliarized with ISO and tested with CIN), c-fosmRNA expression pattern was higher in the DL and VMthan in the VL and DM quadrants [Fig. 3(B)], with adifference reaching the significance level between theDL and VL quadrants (p � .0413). In addition, whateverthe bulbar quadrant, no significant difference could bedetected between the animals of the AIR/CIN and theISO/CIN groups [Fig. 2(D) and (E)].
In the periglomerular and mitral cell layers of theOB, we could not detect any significant changes inc-fos mRNA expression between our four experimen-tal animal groups. For all four quadrants, the interac-tion of the effects from the exposure and test odors didnot reach significant level (two-way ANOVA).
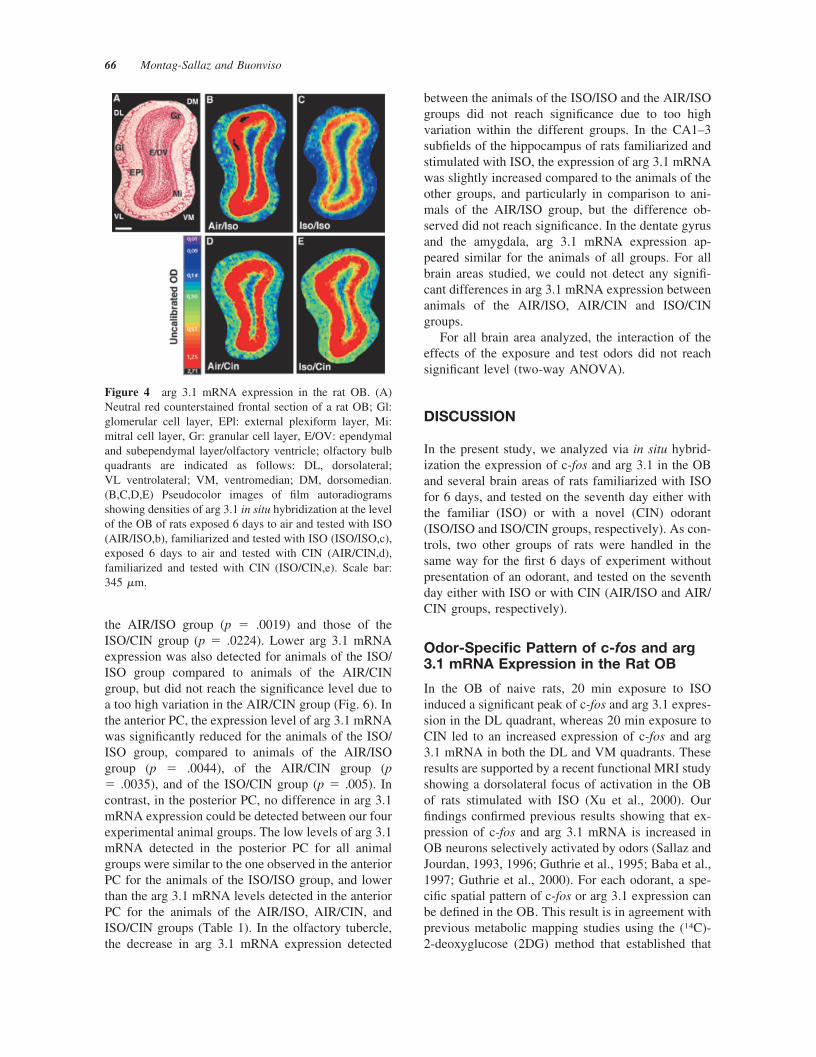
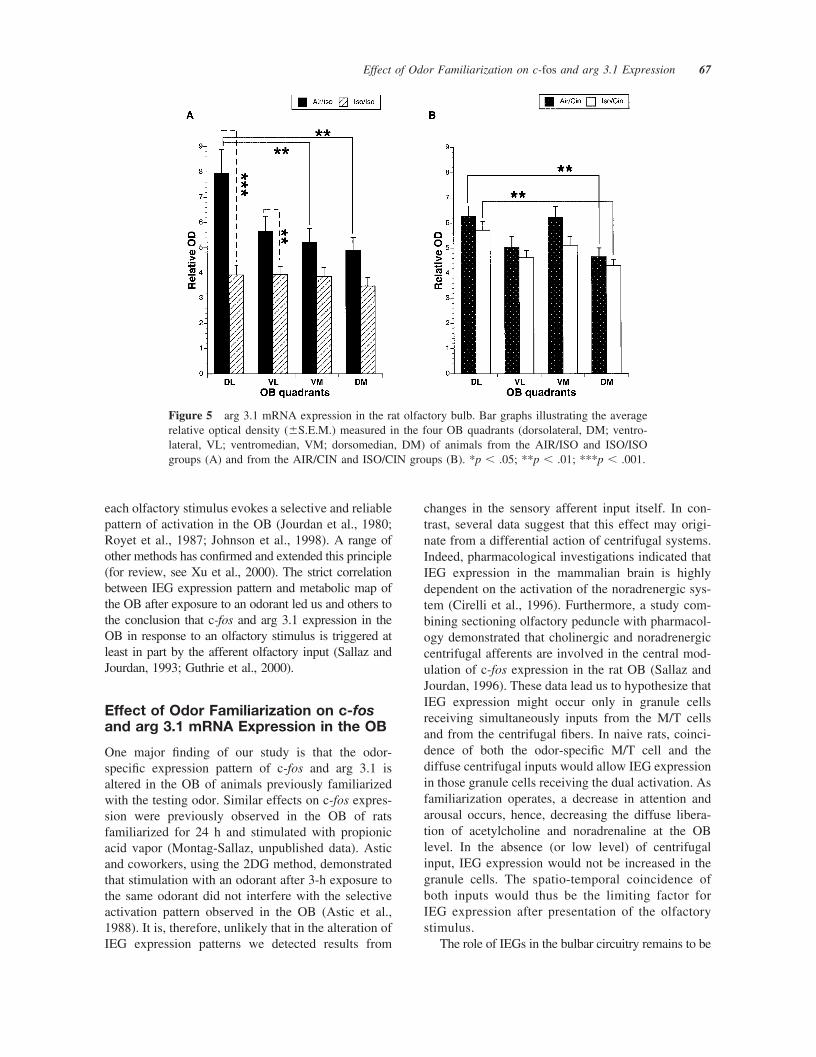
Arg 3.1 mRNA. Figure 4 shows arg 3.1 expression inthe OB of the four groups of rats. Quantitative anal-ysis of the autoradiograms revealed significant differ-ences in arg 3.1 mRNA expression level in the gran-ular layer of the OB between our four experimentalanimal groups. In the AIR/ISO group (n � 5), arg 3.1mRNA expression was increased in the DL quadrant,compared to the other three quadrants. The signifi-cance level was reached for DL versus VM (p� .0054) and for DL versus DM (p � .0019) quad-rants [Fig. 5(A)]. In the ISO/ISO group, no significant
difference in arg 3.1 mRNA expression could bedetected between the four OB quadrants. In addition,arg 3.1 mRNA expression was reduced in the fourquadrants of the ISO/ISO group versus the corre-sponding quadrants of the AIR/ISO group, the differ-ence reaching significant level for the DL (p � .0001)and VL (p � .0059) quadrants [Fig. 5(A)]. In the OBof the AIR/CIN group animals (n � 5), arg 3.1 mRNAexpression was increased in the DL and VM quad-rants compared to the VL and DM quadrants [Fig.5(B)], with a difference reaching significance onlybetween the DL and DM quadrants [p � .0082; Fig.5(B)]. In the ISO/CIN group (n � 5), arg 3.1 mRNAexpression pattern was not modified by previous ex-posure with ISO (Fig. 4), and no significant differencein arg 3.1 mRNA expression was observed betweenAIR/CIN and ISO/CIN groups [Fig. 5(B)]. Addition-ally, the interaction of the effects from the exposureand test odors reached significant level for the DLquandrant, as shown with a two-way ANOVA, F(1,72) � 8.81, p � .0041.
In the other layers of the OB, no significantchanges in arg 3.1 mRNA expression could be de-tected between our four experimental animal groups.
Brain Areas
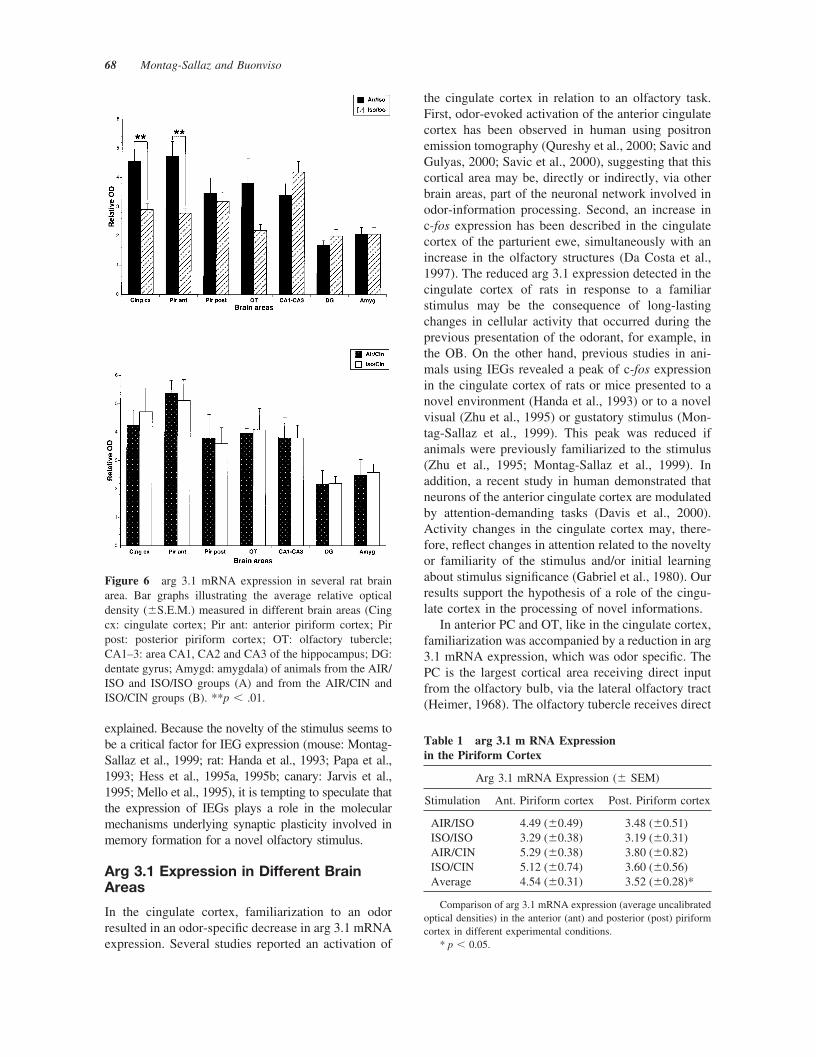
As shown in Figure 6, a decrease in arg 3.1 mRNAexpression was detected in the cingulate cortex of theanimals of the ISO/ISO group compared to those of
Figure 3 c-fos mRNA expression in the rat OB. Bar graphs illustrating the average density of c-fospositive cells (�S.E.M.) evaluated in the four OB quadrants (dorsolateral, DM; ventrolateral, VL;ventromedian, VM; dorsomedian, DM) of animals from the AIR/ISO and ISO/ISO groups (A) andfrom the AIR/CIN and ISO/CIN groups (B). *p � .05; **p � .01.
Effect of Odor Familiarization on c-fos and arg 3.1 Expression 65
the AIR/ISO group (p � .0019) and those of theISO/CIN group (p � .0224). Lower arg 3.1 mRNAexpression was also detected for animals of the ISO/ISO group compared to animals of the AIR/CINgroup, but did not reach the significance level due toa too high variation in the AIR/CIN group (Fig. 6). Inthe anterior PC, the expression level of arg 3.1 mRNAwas significantly reduced for the animals of the ISO/ISO group, compared to animals of the AIR/ISOgroup (p � .0044), of the AIR/CIN group (p� .0035), and of the ISO/CIN group (p � .005). Incontrast, in the posterior PC, no difference in arg 3.1mRNA expression could be detected between our fourexperimental animal groups. The low levels of arg 3.1mRNA detected in the posterior PC for all animalgroups were similar to the one observed in the anteriorPC for the animals of the ISO/ISO group, and lowerthan the arg 3.1 mRNA levels detected in the anteriorPC for the animals of the AIR/ISO, AIR/CIN, andISO/CIN groups (Table 1). In the olfactory tubercle,the decrease in arg 3.1 mRNA expression detected
between the animals of the ISO/ISO and the AIR/ISOgroups did not reach significance due to too highvariation within the different groups. In the CA1–3subfields of the hippocampus of rats familiarized andstimulated with ISO, the expression of arg 3.1 mRNAwas slightly increased compared to the animals of theother groups, and particularly in comparison to ani-mals of the AIR/ISO group, but the difference ob-served did not reach significance. In the dentate gyrusand the amygdala, arg 3.1 mRNA expression ap-peared similar for the animals of all groups. For allbrain areas studied, we could not detect any signifi-cant differences in arg 3.1 mRNA expression betweenanimals of the AIR/ISO, AIR/CIN and ISO/CINgroups.
For all brain area analyzed, the interaction of theeffects of the exposure and test odors did not reachsignificant level (two-way ANOVA).
DISCUSSION
In the present study, we analyzed via in situ hybrid-ization the expression of c-fos and arg 3.1 in the OBand several brain areas of rats familiarized with ISOfor 6 days, and tested on the seventh day either withthe familiar (ISO) or with a novel (CIN) odorant(ISO/ISO and ISO/CIN groups, respectively). As con-trols, two other groups of rats were handled in thesame way for the first 6 days of experiment withoutpresentation of an odorant, and tested on the seventhday either with ISO or with CIN (AIR/ISO and AIR/CIN groups, respectively).
Odor-Specific Pattern of c-fos and arg3.1 mRNA Expression in the Rat OB
In the OB of naive rats, 20 min exposure to ISOinduced a significant peak of c-fos and arg 3.1 expres-sion in the DL quadrant, whereas 20 min exposure toCIN led to an increased expression of c-fos and arg3.1 mRNA in both the DL and VM quadrants. Theseresults are supported by a recent functional MRI studyshowing a dorsolateral focus of activation in the OBof rats stimulated with ISO (Xu et al., 2000). Ourfindings confirmed previous results showing that ex-pression of c-fos and arg 3.1 mRNA is increased inOB neurons selectively activated by odors (Sallaz andJourdan, 1993, 1996; Guthrie et al., 1995; Baba et al.,1997; Guthrie et al., 2000). For each odorant, a spe-cific spatial pattern of c-fos or arg 3.1 expression canbe defined in the OB. This result is in agreement withprevious metabolic mapping studies using the (14C)-2-deoxyglucose (2DG) method that established that
Figure 4 arg 3.1 mRNA expression in the rat OB. (A)Neutral red counterstained frontal section of a rat OB; Gl:glomerular cell layer, EPl: external plexiform layer, Mi:mitral cell layer, Gr: granular cell layer, E/OV: ependymaland subependymal layer/olfactory ventricle; olfactory bulbquadrants are indicated as follows: DL, dorsolateral;VL ventrolateral; VM, ventromedian; DM, dorsomedian.(B,C,D,E) Pseudocolor images of film autoradiogramsshowing densities of arg 3.1 in situ hybridization at the levelof the OB of rats exposed 6 days to air and tested with ISO(AIR/ISO,b), familiarized and tested with ISO (ISO/ISO,c),exposed 6 days to air and tested with CIN (AIR/CIN,d),familiarized and tested with CIN (ISO/CIN,e). Scale bar:345 �m.
66 Montag-Sallaz and Buonviso
each olfactory stimulus evokes a selective and reliablepattern of activation in the OB (Jourdan et al., 1980;Royet et al., 1987; Johnson et al., 1998). A range ofother methods has confirmed and extended this principle(for review, see Xu et al., 2000). The strict correlationbetween IEG expression pattern and metabolic map ofthe OB after exposure to an odorant led us and others tothe conclusion that c-fos and arg 3.1 expression in theOB in response to an olfactory stimulus is triggered atleast in part by the afferent olfactory input (Sallaz andJourdan, 1993; Guthrie et al., 2000).
Effect of Odor Familiarization on c-fosand arg 3.1 mRNA Expression in the OB
One major finding of our study is that the odor-specific expression pattern of c-fos and arg 3.1 isaltered in the OB of animals previously familiarizedwith the testing odor. Similar effects on c-fos expres-sion were previously observed in the OB of ratsfamiliarized for 24 h and stimulated with propionicacid vapor (Montag-Sallaz, unpublished data). Asticand coworkers, using the 2DG method, demonstratedthat stimulation with an odorant after 3-h exposure tothe same odorant did not interfere with the selectiveactivation pattern observed in the OB (Astic et al.,1988). It is, therefore, unlikely that in the alteration ofIEG expression patterns we detected results from
changes in the sensory afferent input itself. In con-trast, several data suggest that this effect may origi-nate from a differential action of centrifugal systems.Indeed, pharmacological investigations indicated thatIEG expression in the mammalian brain is highlydependent on the activation of the noradrenergic sys-tem (Cirelli et al., 1996). Furthermore, a study com-bining sectioning olfactory peduncle with pharmacol-ogy demonstrated that cholinergic and noradrenergiccentrifugal afferents are involved in the central mod-ulation of c-fos expression in the rat OB (Sallaz andJourdan, 1996). These data lead us to hypothesize thatIEG expression might occur only in granule cellsreceiving simultaneously inputs from the M/T cellsand from the centrifugal fibers. In naive rats, coinci-dence of both the odor-specific M/T cell and thediffuse centrifugal inputs would allow IEG expressionin those granule cells receiving the dual activation. Asfamiliarization operates, a decrease in attention andarousal occurs, hence, decreasing the diffuse libera-tion of acetylcholine and noradrenaline at the OBlevel. In the absence (or low level) of centrifugalinput, IEG expression would not be increased in thegranule cells. The spatio-temporal coincidence ofboth inputs would thus be the limiting factor forIEG expression after presentation of the olfactorystimulus.
The role of IEGs in the bulbar circuitry remains to be
Figure 5 arg 3.1 mRNA expression in the rat olfactory bulb. Bar graphs illustrating the averagerelative optical density (�S.E.M.) measured in the four OB quadrants (dorsolateral, DM; ventro-lateral, VL; ventromedian, VM; dorsomedian, DM) of animals from the AIR/ISO and ISO/ISOgroups (A) and from the AIR/CIN and ISO/CIN groups (B). *p � .05; **p � .01; ***p � .001.
Effect of Odor Familiarization on c-fos and arg 3.1 Expression 67
explained. Because the novelty of the stimulus seems tobe a critical factor for IEG expression (mouse: Montag-Sallaz et al., 1999; rat: Handa et al., 1993; Papa et al.,1993; Hess et al., 1995a, 1995b; canary: Jarvis et al.,1995; Mello et al., 1995), it is tempting to speculate thatthe expression of IEGs plays a role in the molecularmechanisms underlying synaptic plasticity involved inmemory formation for a novel olfactory stimulus.
Arg 3.1 Expression in Different BrainAreas
In the cingulate cortex, familiarization to an odorresulted in an odor-specific decrease in arg 3.1 mRNAexpression. Several studies reported an activation of
the cingulate cortex in relation to an olfactory task.First, odor-evoked activation of the anterior cingulatecortex has been observed in human using positronemission tomography (Qureshy et al., 2000; Savic andGulyas, 2000; Savic et al., 2000), suggesting that thiscortical area may be, directly or indirectly, via otherbrain areas, part of the neuronal network involved inodor-information processing. Second, an increase inc-fos expression has been described in the cingulatecortex of the parturient ewe, simultaneously with anincrease in the olfactory structures (Da Costa et al.,1997). The reduced arg 3.1 expression detected in thecingulate cortex of rats in response to a familiarstimulus may be the consequence of long-lastingchanges in cellular activity that occurred during theprevious presentation of the odorant, for example, inthe OB. On the other hand, previous studies in ani-mals using IEGs revealed a peak of c-fos expressionin the cingulate cortex of rats or mice presented to anovel environment (Handa et al., 1993) or to a novelvisual (Zhu et al., 1995) or gustatory stimulus (Mon-tag-Sallaz et al., 1999). This peak was reduced ifanimals were previously familiarized to the stimulus(Zhu et al., 1995; Montag-Sallaz et al., 1999). Inaddition, a recent study in human demonstrated thatneurons of the anterior cingulate cortex are modulatedby attention-demanding tasks (Davis et al., 2000).Activity changes in the cingulate cortex may, there-fore, reflect changes in attention related to the noveltyor familiarity of the stimulus and/or initial learningabout stimulus significance (Gabriel et al., 1980). Ourresults support the hypothesis of a role of the cingu-late cortex in the processing of novel informations.
In anterior PC and OT, like in the cingulate cortex,familiarization was accompanied by a reduction in arg3.1 mRNA expression, which was odor specific. ThePC is the largest cortical area receiving direct inputfrom the olfactory bulb, via the lateral olfactory tract(Heimer, 1968). The olfactory tubercle receives direct
Figure 6 arg 3.1 mRNA expression in several rat brainarea. Bar graphs illustrating the average relative opticaldensity (�S.E.M.) measured in different brain areas (Cingcx: cingulate cortex; Pir ant: anterior piriform cortex; Pirpost: posterior piriform cortex; OT: olfactory tubercle;CA1–3: area CA1, CA2 and CA3 of the hippocampus; DG:dentate gyrus; Amygd: amygdala) of animals from the AIR/ISO and ISO/ISO groups (A) and from the AIR/CIN andISO/CIN groups (B). **p � .01.
Table 1 arg 3.1 m RNA Expressionin the Piriform Cortex
Arg 3.1 mRNA Expression (� SEM)
Stimulation Ant. Piriform cortex Post. Piriform cortex
AIR/ISO 4.49 (�0.49) 3.48 (�0.51)ISO/ISO 3.29 (�0.38) 3.19 (�0.31)AIR/CIN 5.29 (�0.38) 3.80 (�0.82)ISO/CIN 5.12 (�0.74) 3.60 (�0.56)Average 4.54 (�0.31) 3.52 (�0.28)*
Comparison of arg 3.1 mRNA expression (average uncalibratedoptical densities) in the anterior (ant) and posterior (post) piriformcortex in different experimental conditions.
* p � 0.05.
68 Montag-Sallaz and Buonviso

afferent inputs from the OB (Van Hoesen et al.,1976). Like proposed for the cingulate cortex, themodification in arg 3.1 expression detected in anteriorPC and OT could be the consequence of long-lastingchanges in bulbar neuron activity occurring during thefamiliarization to the odorant and leading to a modi-fied neuronal input to these structures. This does notexclude the possibility that these structures, as the cin-gulate cortex, could be part of an olfactory novelty-encoding network. Another possibility is that pyramidalcells of these regions would be silenced by the exposurebecause Wilson (1998, 2000) reports a rapid odor habit-uation of anterior piriform cortex responses.
In contrast to the anterior PC, the posterior part ofthe PC remained not affected by familiarization. An-atomical differences between anterior and posteriorparts of PC may at least in part explain why an effectof familiarization was detected exclusively in the an-terior PC. First, afferent fibers are more numerous inthe anterior than in the posterior part of the PC(Haberly, 1985). Second, the PC returns projections tothe OB via centrifugal fibers mainly originating fromits anterior part, whereas the posterior part gives riseonly to sparse projections (Haberly and Price, 1978).From the functional point of view, the anterior andposterior parts of the PC also differ (Lynch andGranger, 1989; Litaudon and Cattareli, 1995, 1996;Litaudon et al., 1997; Mouly et al., 1997; Rosin et al.,1999). The anterior PC may be mainly involved inodor detection and categorization, whereas the poste-rior PC could be particularly involved in olfactorymemory (Hasselmo, 1995). Finally, it is likely that theanterior PC participates in an olfactory novelty-en-coding network or in the processing of simple taskswhile the posterior part of the PC would be ratherengaged in the processing of more complex tasks notrequired in our experimental protocol.
A significant effect of the familiarization on arg 3.1expression has been detected neither in the hippocam-pus (CA1–3 and DG) nor in the amygdala, althoughboth structures receive olfactory projections and con-nect several olfactory cortical areas (Haberly, 1985;Biella and Curtis, 2000). Indeed, the dentate gyrusparticipates in olfactory information processing(Heale and Vanderwolf, 1994; Heale et al., 1994;Heale and Vanderwolf, 1995) while the hippocampusproper is implicated in novelty detection (animal:Hess et al., 1995a,b; human: Tulving et al., 1994;Knight, 1996). Using c-fos expression, Gall et al.(1998) reported CA1 activation in rats exploring anovel open field and rats overtrained to the same openfield and responding to odors. However, a lack ofnovelty-specific changes in IEG expression (Zhu etal., 1995; Montag-Sallaz et al., 1999) and in 2DG
uptake (Hamrick et al., 1993) in the hippocampus hasbeen already reported using visual, gustatory, andolfactory stimuli, respectively. Similarly, severalstudies in human showed an activation of the amyg-dala after presentation of an olfactory stimulus (Savicet al., 2000; Savic and Gulyas, 2000). Although dif-ferences in c-fos expression were detected in theamygdala of animals stimulated with a novel gusta-tory stimulus, no effect was observed in arg 3.1mRNA expression. In summary, these data suggestthat either the familiarization effect on arg 3.1 mRNAexpression in the hippocampus and amygdala re-mained below detection level or that these structuresare not involved in the odor familiarization processrequiring changes at the genomic level.
Comparison with ElectrophysiologicalData
Our previous electrophysiological recordings of M/Tcells revealed that the familiarization specificity de-pended on the odor used for exposure: the morepowerful an odor was in activating control cells, themore nonspecific the decrease in M/T cell reactivitywas (Buonviso and Chaput, 2000). The nonspecificityof the effects probably originates from the large over-lap of the molecular features contained in the familiarodor (ISO) and those contained in the other test odors.This assumption is strongly confirmed by the recentreport of Wilson (2000) showing a strong crosshabitu-ation of M/T cell responses to an odor set. In contrast,the present results demonstrate that the effects of odorfamiliarization on the expression patterns of c-fos andarg 3.1 in the bulbar granular cells are odor specific.Both results are compliant if we consider the hypoth-esis of coincident inputs outlined above and in Figure7. When an odor 1 (Od1) is presented to naive rats, thecorresponding M/T cells are activated, some respond-ing specifically to Od1 (Od1-M/T) and some othersresponding to both Od1 and Od2 (Od1�Od2-M/T),due to overlapping molecular features between Od1and Od2. In addition, centrifugal systems are activebecause of the novelty of the stimulus, leading toan expression of IEGs in granule cells connected tothe Od1-M/T (Od1-Gr) and to the Od1�Od2-M/T(Od1�Od2-Gr) ]Fig. 7(A)]. The same effect can beexpected after presentation of Od2 to naive rats, re-sulting in the activation of a different but overlappingpattern of M/T cells (Od2-M/T and Od1�Od2-M/T)and expression of IEGs in the corresponding granulecells (Od2-Gr and Od1�Od2-Gr) [Fig. 7(A)]. Whenthe odor used for familiarization is presented for thetest (Od1/Od1), centrifugal systems are silenced, andonly a limited number of M/T cells are responsive,
Effect of Odor Familiarization on c-fos and arg 3.1 Expression 69

both factors leading to a decreased probability ofinput coincidence onto granule cells necessary forIEGs to be expressed [Fig. 7(B)]. When Od1 is usedfor the familiarization and a novel odor (Od2) for thetest, centrifugal systems are active, the M/T cellsprocessing both Od1 and Od2 features (Od1�Od2-M/T) remain silent, while those processing the mo-lecular features specific to Od2 (Od2-M/T) show a
normal activity pattern. Therefore, IEG expressionwill be triggered selectively in the correspondinggranule cells (Od2-Gr). The number of granule cellsexpressing IEGs may be slightly reduced compared toAIR/Od2, but is still sufficient for the odor-specificpattern of IEGs to be maintained. This hypothesiscould explain the odor specificity of the familiariza-tion we detected on arg 3.1 and c-fos expression in thegranule cells. The relative specificity of the familiar-ization at the M/T cell level could reflect the degree ofoverlap of the activation maps evoked at the bulbarlevel by the odorants used for exposure and test.
CONCLUSION
Our results confirm that the expression pattern of c-fosand arg 3.1 in the granule cell layer of the OB is odorspecific, and demonstrate that this specificity is al-tered by previous familiarization with the odorantused as stimulus. We also demonstrated that the odorspecificity of the familiarization can be detected in theanterior PC and the cingulate cortex, which is part ofthe cerebral novelty encoding network. These resultsstrongly suggest that c-fos and arg 3.1 may play a rolein the molecular mechanisms underlying synapticplasticity involved in memory formation for a newolfactory stimulus and confirm the usefulness of c-fosand arg 3.1 as markers for detection of novelty/famil-iarization processes detection. This study comple-ments our previous electrophysiological data and pro-poses an hypothesis for the cellular mechanismsunderlying the familiarization process in the olfactorysystem.
We thank Dr. Dirk Montag for providing the c-fos andarg 3.1 antisense cRNA probes.
REFERENCES
Astic L, Saucier D, Jourdan F, Holley A. 1988. Functionalactivaty of the rat olfactory bulb related to differentduration of odor exposure. Chem Senses 13:333–343.
Baba K, Ikeda M, Houtani T, Nakagawa H, Ueyama T, SatoK, Sakuma S, Yamashita T, Tsukahara Y, Sugimoto T.1997. Odour exposure reveals non-uniform expressionprofiles of c-Jun protein in rat olfactory bulb neurones.Brain Res 774:142–148.
Berke JD, Paltzki RF, Aronson GJ, Hyman SE, Gerfen CR.1998. A complex program of striatal gene expressioninduced by dopaminergic stimulation. J Neurosci 18:5301–5310.
Biella G, De Curtis M. 2000. Olfactory inputs activate the
Figure 7 Familiarization and IEGs expression in the ol-factory bulb. (A) The presentation of Odor 1 (Od1) to naiverats activates M/T cells responding specifically to Od1(Od1-M/T) and others responding to both Od1 and Odor 2(Od2) (Od1�Od2-M/T); in addition, centrifugal systemsare turned on because of the novelty of the stimulus, leadingto an expression of IEGs in granule cells connected to theOD1-M/T (Od1-Gr) and to the Od1�Od2-M/T (Od1�Od2-Gr). The presentation of Od2 to naive rats results in theactivation of a different but overlapping pattern of M/T cells(Od2-M/T and Od1�Od2-M/T) and expression of IEGs inthe corresponding granule cells (Od2-Gr and Od1�Od2-Gr). (B) Od1 is presented for the test after familiarizationwith the same odor (Od1/Od1). Centrifugal systems areturned off, and only a limited number of M/T cells (Od1-M/T and Od1�Od2-M/T) are responsive, resulting in a verylimited IEG expression in the Od1-Gr and Od1�Od2-Grand an alteration of the odor-specific pattern of IEG expres-sion. (C) Od2 is presented for the test after familiarizationwith Od1 (Od1/Od2). Centrifugal systems are turned on,and the M/T cells processing both Od1 and Od2 features(Od1�Od2-M/T) remain silent while those processing themolecular features specific to Od2 (Od2-M/T) show a nor-mal activity pattern. Consequentely, IEG expression will betriggered selectively in the corresponding granule cells(Od2-Gr).
70 Montag-Sallaz and Buonviso

medial entorhinal cortex via the hippocampus. J Neuro-physiol 83:1924–1931.
Buonviso N, Chaput MA. 2000. Olfactory experience de-creases responsiveness of the olfactory bulb in the adultrat. Neuroscience 95:325–332.
Buonviso N, Gervais R, Chalansonnet M, Chaput M. 1998.Short-lasting exposure to one odour decreases generalreactivity in the olfactory bulb of adult rats. Eur J Neu-rosci 10:2472–2475.
Cirelli C, Pompeiano M, Tononi G. 1996. Neuronal geneexpression in the waking state: a role for the locus coer-uleus. Science 274:1211–1215.
Da Costa AP, Broad KD, Kendrick KM. 1997. Olfactorymemory and maternal behaviour-induced changes in c-fos and zif/268 mRNA expression in the sheep. Mol BrainRes 46:63–76.
Davis KD, Hutchison WD, Lozano AM, Tasker RR,Dostrovski JO. 2000. Human anterior cingulate cortexneurons modulated by attention-demanding tasks. J Neu-rophysiol 83:3575–3577.
Fosnaugh JS, Bhat RV, Yamagata K, Worley P, BarabanJM. 1995. Activation of arg, a putative “effector” Imme-diate early gene, by cocaine in rat brain. J Neurochem64:2377–2380.
Freeman WJ, Schneider W. 1982. Changes in spatial pat-terns of rabbit olfactory EEG with conditioning to odors.Psychophysiology 19:44–56.
Gabriel M, Foster K, Orona E, Saltwick SE, Stanton M.1980. Limbic encoding of stimulus significance. InProgress in psychobiology and physiological psychology.Sprague JM, Epstein AN, editors.
Gall CM, Hess US, Lynch G. 1998. Mapping brain net-works engaged by, and changed by, learning. NeurobiolLearn Mem 70:14–36.
Gervais R, Holley A, Keverne B. 1988. The importance ofcentral noradrenergic influences on the olfactory bulb inthe processing of learned olfactory cues. Chem Senses13:3–12.
Guthrie K, Rayhanabad J, Kuhl D, Gall C. 2000. Odorsregulate Arc expression in neuronal ensembles engagedin odor processing. Neuroreport 11:1809–1813.
Guthrie KM, Nguyen T, Gall CM. 1995. Insulin-like growthfactor-1 mRNA is increased in deafferented hippocam-pus: spatiotemporal correspondence of a trophic eventwith axon sprouting. J Comp Neurol 352:147–160.
Haberly LB. 1985. Neuronal circuitry in olfactory cortex:anatomy and functional implications. Chem Senses 10:219–238.
Haberly LB, Price JL. 1978. Association and commissurlsystems of the olfactory cortex of the rat. I—Systemsoriginating in the piriform cortex ans adjacent areas.J Comp Neurol 178:711–740.
Hamrick WD, Wilson DA, Sullivan RM. 1993. Neuronalcorrelates of memory for odor detection conditioning inadult rats. Neurosci Lett 163:36–40.
Handa RJ, Nunley KM, Bollnow MR. 1993. Induction ofc-fos mRNA in the brain and anterior pituitary gland bya novel environment. Neuroreport 4:1079–1082.
Hasselmo ME. 1995. Neuromodulation and cortical func-tion: modeling the physiological basis of behavior. BehavBrain Res 67:1–27.
Heale VR, Vanderwolf CH. 1994. Dentate gyrus and olfac-tory bulb responses to olfactory nd noxious stimulation inurethane anaesthetized rats. Brain Res 652:235–242.
Heale VR, Vanderwolf CH. 1995. Scopolamine blocks ol-faction-induced fast waves but not olfactory evoked po-tentials in the dentate gyrus. Behav Brain Res 68:57–64.
Heale VR, Vanderwolf CH, Kavaliers M. 1994. Compo-nents of weasel and fox odors elicit fast wave bursts inthe dentate gyrus of rats. Behav Brain Res 63:159–165.
Heimer L. 1968. Synaptic sidtribution of centripetal andcentrifugal nerve fibers in the olfactory system in the rat.J Anat 103:413–432.
Herdegen T, Leah JD. 1998. Inducible and constitutivetranscription factors in the mammalian nervous system:control of gene expression by Jun, Fos and Krox, andCREB/ATF proteins. Brain Res Brain Res Rev 28:370–490.
Hess US, Lynch G, Gall CM. 1995a. Changes in c-fosmRNA expression in rat brain during odor discriminationlearning: differential involvement of hippocampal sub-fields CA1 and CA3. J Neurosci 15:4786–4795.
Hess US, Lynch G, Gall CM. 1995b. Regional patterns ofc-fos mRNA expression in rat hippocampus followingexploration of a novel environment versus performanceof a well-learned discrimination. J Neurosci 15:7796–7809.
Jarvis ED, Mallo C, Nottebohm F. 1995. Associative learn-ing and stimulus novelty influence the song-induced ex-pression of an Immediate Early Gene in the Canaryforebrain. Learn Mem 2:62–80.
Johnson BA, Woo CC, Leon M. 1998. Spatial coding ofodorant features in the glomerular layer of the rat olfac-tory bulb. J Comp Neurol 393:457–471.
Jourdan F, Duveau A, Astic L, Holley A. 1980. Spatialdistribution of 14C 2-deoxyglucose uptake in the olfac-tory bulbs of rats stimulated with two different odours.Brain Res 188:139–154.
Kaczmarek L, Nikolajew E. 1990. c-fos protooncogene ex-pression and neuronal plasticity. Acta Neurobiol Exp(Warsz) 50:173–179.
Knight RT. 1996. Contribution of human hippocampal re-gion to novelty detection. Nature 383:256–259.
Link W, Konietzko U, Kauselmann G, Krug M, SchwankeB, Frey U, Kuhl D. 1995. Somatodendritic expression ofan immediate early gene is regulated by synaptic activity.Proc Natl Acad Sci USA 92:5734–5738.
Litaudon P, Cattarelli M. 1995. Piriform cortex late activityrevealed functional spatial heterogeneity. Neuroreport6:1377–1380.
Litaudon P, Cattarelli M. 1996. Olfactory bulb repetitivestimulations reveal non-homogeneous distribution of theinhibitory processes in the rat piriform cortex. Eur J Neu-rosci 8:21–29.
Litaudon P, Datiche F, Cattarelli M. 1997. Optical recording
Effect of Odor Familiarization on c-fos and arg 3.1 Expression 71

of the rat piriform cortex activity. Prog Neurobiol 52:485–510.
Lyford G, Yamagata K, Kaufmann W, Barnes C, Sanders L,Copeland N, Gilbert D, Jenkins N, Lanahan A, Worley P.1995. Arc, a growth factor and activity-regulated gene,encodes a novel cytoskeleton-associated protein that isenriched in neuronal dendrites. Neuron 14:433–445.
Lynch G, Granger R. 1989. Simulation and analysis of asimple cortical network. Psychol Learn Motiv 23:205–241.
Mello C, Nottebohm F, Clayton D. 1995. Repeated expo-sure to one song leads to a rapid and persistent decline inan Immediate Early Gene’s response to that song in ZebraFinch Telencephalon. J Neurosci 15:6919–6925.
Montag-Sallaz M, Welzl H, Kuhl D, Montag D, SchachnerM. 1999. Novelty-induced increased expression of theimmediate early genes c-fos and arg 3.1 in the mousebrain. J Neurobiol 38:234–246.
Mouly AM, Elaagouby A, Ravel N. 1995. A study of theeffects of noradrenaline in the rat olfactory bulb usingevoked field potential response. Brain Res 681:47–57.
Papa M, Pellicano MP, Welzl H, Sadile AG. 1993. Distrib-uted changes in c-fos and c-jun immunoreactivity in therat brain associated with arousal and habituation to nov-elty. Brain Res Bull 32:509–515.
Pei Q, Lewis L, Sprakes ME, Jones EJ, Grahame-Smith DG,Zetterstrom TSC. 2000. Serotoninergic regulation ofmRNA expression of Arc, an immediate early gene se-lectively localized at neuronal dendrites. Neuropharma-cology 39:463–470.
Qureshy A, Kawashima R, Imran MB, Sugiura M, Goto R,Okada K, Inoue K, Itoh M, Schormann T, Zilles K,Fukuda H. 2000. Functional mapping of human brain inolfactory processing: a PET study. J Neurophysiol 84:1656–1666.
Ravel N, Vigouroux M, Elaagouby A, Gervais R. 1992.scopolamine impairs delayed matching in an olfactorytask in rats. Psychopharmacology (Berlin) 109:439–443.
Rosin JF, Datiche F, Cattarell M. 1999. Modulation of thepiriform cortex activity by the basal forebrain: an opticalrecording study in the rat. Brain Res 820:105–111.
Royet JP, Sicard G, Souchier C, Jourdan F. 1987. Specific-ity of spatial patterns of glomerular activation in themouse olfactory bulb: computer-assisted image analysisof 2-deoxyglucose autoradiograms. Brain Res 417:1–11.
Sallaz M, Jourdan F. 1993. c-fos expression and 2-deoxy-glucose uptake in the olfactory bulb of odour-stimulatedawake rats. Neuroreport 4:55–58.
Sallaz M, Jourdan F. 1996. Odour-induced c-fos expressionin the rat olfactory bulb: involvement of centrifugal af-ferents. Brain Res 721:66–75.
Savic I, Gulyas B. 2000. PET shows that odors are pro-cessed both ipsilaterally and contralaterally to the stimu-lated nostril. Neuroreport 11:2861–2866.
Savic I, Gulyas B, Larsson M, Roland P. 2000. Olfactoryfunctions are mediated by parallel and hierarchical pro-cessing. Neuron 26:735–745.
Silva AJ, Giese KP. 1994. Plastic genes are in! Curr OpinNeurobiol 4:413–420.
Steward O, Wallace CS, Lyford G, Worley PF. 1998. Syn-aptic activation causes the mRNA for the IEG Arc tolocalize selectively near activated postsynaptic sites ondendrites. Neuron 21:741–751.
Tan A, Moratalla R, Lyford GL, Worley P, Graybiel AM.2000. The activity-regulated cytoskeletal-associated pro-tein Arc is expressed in different stiosome-matrix pat-terns following exposure to amphetamine and cocaine.J Neurochem 74:2074–2078.
Tischmeyer W, Grimm R. 1999. Activation of immediateearly genes and memory formation. Cell Mol Life Sci55:564–574.
Tulving E, Markowitsch HJ, Kapur S, Habib R, Houle S.1994. Novelty encoding networks in the human brain:positron emission tomography data. Neuroreport 5:2525–2528.
Van Hoesen GW, Mesulam MM, Haaxma R. 1976. Tem-poral cortical projections to the olfactory tubercle in therhesus monkey. Brain Res 109:375–381.
Wallace CS, Lyford G, Worley PF, Steward O. 1998. Dif-ferential intracellular sorting of IEG mRNAs depends onsignals in the mRNA sequence. J Neurosci 18:26–35.
Wilson DA. 1998. Habituation of odor responses in the ratanterior piriform cortex. J Neurophysiol 79:1425–1440.
Wilson DA. 2000. Comparison of odor receptive field plas-ticity in the rat olfactory bulb and anterior piriform cor-tex. J Neurophysiol 84:3036–3042.
Xu F, Kida I, Hyder F, Shulman RG. 2000. Assessment anddiscrimination of odor stimuli in rat olfactory bulb bydynamic functional MRI. Proc Natl Acad Sci USA 97:10601–10606.
Zhu H, Wu F, Schacher S. 1995. Changes in expression anddistribution of Aplysia cell adhesion molecules can influ-ence synapse formation and elimination in vitro. J Neu-rosci 15:4173–4183.
72 Montag-Sallaz and Buonviso