age-related changes in circadian rhythm of serotonin synthesis in ring doves: effects of increased...
TRANSCRIPT

Age-related changes in circadian rhythm of serotonin synthesis in ring
doves: Effects of increased tryptophan ingestion
Celia Garau, Sara Aparicio, Ruben V. Rial, Marıa C. Nicolau, Susana Esteban *
Laboratori de Neurofisiologia, Departament de Biologia Fonamental i Ciences de la Salut, Universitat de les Illes Balears, E-07122 Palma de Mallorca, Spain
Received 26 July 2005; received in revised form 15 September 2005; accepted 27 September 2005
Available online 3 November 2005
Abstract
Alterations in the function of the hypothalamic suprachiasmatic nucleus (SCN) with age have been reported. As serotonin is an important
regulator of the circadian clock located in SCN, this work studied the changes produced in the synthesis of serotonin with age using the
accumulation of 5-HTP after decarboxylase inhibition as a measure of serotonin synthesis in the brain in vivo, in young and old ring doves at the
onset of lights-on and lights-off. A diurnal cycle in tryptophan hydroxylation was observed in young animals, with an increased daylight synthesis
and metabolism of 5-HT in hippocampus, neostriatum and hypothalamus. A single dose of melatonin (1 mg/kg, i.p., 1 h) at lighttime produced an
inhibitory effect on the synthesis of 5-HT. In contrast, differences in 5-HT synthesis and metabolism between day and night dissappeared in old
animals indicating an absence of a circadian rhythm in 5-HT synthesis and metabolism. The administration of L-tryptophan (240 mg/kg, i.p.)
strongly increased the 5-HT synthesis in young animals only during lights-off time while it increased in old ones irrespective of the administration
time. These results suggest that the supplemental administration of tryptophan might aid to improve the descent in 5-HT that normally occurs, as
animals get old.
q 2005 Elsevier Inc. All rights reserved.
Keywords: Serotonin synthesis; Aging; Tryptophan; Avian rhythms; Melatonin, Ring doves
1. Introduction
Daily rhythms in vertebrate physiology are generated and
maintained by the biological clock located in the hypothalamic
suprachiasmatic nucleus (SCN) (Moore and Eichler, 1972;
Stephan and Zucker, 1972). Serotonin (5-HT) is an important
regulator of the circadian clock located in the SCN. This clock
is synchronized by photic and non-photic signals. Light, the
principal synchronizer, is received by the retinal ganglion cells
and transmitted directly to the SCN through the retinohy-
pothalamic tract (Moore and Eichler, 1972), and indirectly
through the geniculohypothalamic tract (Card and Moore,
1989). Non-photic signals arrive to the SCN by a direct
serotonergic pathway from mesencephalic raphe nuclei mainly
originated from the median raphe nucleus (MRN) (Azmitia and
0531-5565/$ - see front matter q 2005 Elsevier Inc. All rights reserved.
doi:10.1016/j.exger.2005.09.010
* Corresponding author. Address: Valdes, Laboratori de Fisiologia,
Departament de Biologia Fonamental i Ciences de la Salut, Universitat de
les Illes Balears, Ctra. Valldemossa Km 7,5 E-07122 Palma de Mallorca, Spain.
Tel.: C34 971 173145; fax: C34 971 173184.
E-mail address: [email protected] (S. Esteban).
Segal, 1978; Hay-Schmidt et al., 2003) and secondly from the
dorsal raphe nucleus (DRN) (Kawano et al., 1996), also they
arrive indirectly by serotonergic projections from the dorsal
raphe nucleus to the thalamic intergeniculate leaflets which
project to the SCN (Azmitia and Segal, 1978).
Many aspects of circadian function change with age,
including changes in phase relationship of rhythms to the
environmental time signals, reduced sensitivity of the circadian
pacemaker to time cues, decreased amplitude of the circadian
rhythms and increased sleep fragmentation (Czeisler et al.,
1992; Myers and Badia, 1995; Penev et al., 1995; Scarbrough
et al., 1997; Turek et al., 1995; Van Reeth et al., 1992; Zee
et al., 1992). The neural mechanisms causing the age-related
changes in circadian timing system are far from being
elucidated. Some alterations in the function of the SCN have
been identified (Krajnak et al., 1998; Moore and Eichler, 1972).
An interesting candidate to explain some of the age-related
changes in the SCN is 5-HT. It is known that 5-HT takes part in
the regulation of some parameters of circadian timing system
which change with age as in the circadian rhythm of wheel
running or in the modulation of phase shifts (Cutrera et al.,
1994; Penev et al., 1995, 1997). In addition, age-related
changes in 5-HT receptors have been observed in the DRN
Experimental Gerontology 41 (2006) 40–48
www.elsevier.com/locate/expgero

C. Garau et al. / Experimental Gerontology 41 (2006) 40–48 41
(Duncan et al., 1999) and the SCN (Duncan et al., 2000).
Numerous relationships have been demonstrated between the
5-HT system and the SCN. For instance, electrical stimulation
of the DRN and MRN in hamsters evoked 5-HT release in the
SCN, an effect which was blocked by systemic injection of 5-
HT antagonists (Glass et al., 2003). The release of 5-HT in the
SCN was rhythmic and correlated with circadian variations in
the levels of the limiting enzyme in 5-HT synthesis, tryptophan
hydroxylase (Barassin et al., 2002). However, in vivo changes
in the circadian 5-HT synthesis during aging have not been
well studied in spite of plenty of evidence that 5-HT acts as an
inhibitory transmitter that modulates the responses of the
circadian clock to light.
The aim of this work is to study the circadian pattern of 5-HT
synthesis measuring the activity of tryptophan hydroxylase, the
most commonly assay to monitor 5-HT synthesis in vivo
(Carlsson and Lindqvist, 1973). The study has been performed in
three different brain areas of ring doves (Streptopelia risoria):
hypothalamus which contains the homologue of the mammalian
SCN, hippocampus and striatum, two brain regions which
receive a high amount of 5-HT innervations. The effects of the
administration of L-tryptophan, the amino acid precursor of
5-HT synthesis, in young and old ring doves have been studied
too, to analyze possible improvement of the age related changes
that could occur in 5-HT synthesis and metabolism. The ring
dove was chosen as experimental animal because their circadian
characteristics, diurnal and monocyclic, are similar to those of
human beings and also because of their rather well known age
related changes in the melatonin secretion rhythm (Terron et al.,
2002).
2. Material and methods
2.1. Animals
Young (y6 months, 130 g, nZ20) and old (O8 years,
170 g, nZ22) Streptopelia risoria ring doves, have been used.
The animals were individually housed under controlled
environmental conditions (22 8C; 70% humidity), kept under
a 12/12 h light/dark cycle (lights on at 08.00 h daily), with
standard bird seed food and tap water ad libidum.
2.2. Drug treatments of animals
Ring doves received a single intraperitoneally (i.p.)
administration of saline (1 ml/kg) or L-tryptophan
(240 mg/kg, 1 ml/kg) either at the beginning of lights period
(08.00 h) or dark period (20.00 h). After a lapse of 15 min, the
animals received 3-hydroxybenzylhydrazine HCl (NSD 1015,
100 mg/kg, 1 ml/kg, i.p.) (see below) and after a new lapse of
45 min they were sacrificed. In another experiment, ring doves
received a single dose of melatonin (1 mg/kg, 1 ml/kg, i.p.) at
lights on to analyze the effect of melatonin on 5-HT synthesis.
Unless otherwise stated, all used drugs and reagents were
obtained from Sigma chemical, St Louis Mo. The study
followed the ‘principles of laboratory animal care’ (NIH
publication No. 85-23, revised 1996) and was performed
according to the guidelines of the ethical committee of the
Universitat de les Illes Balears.
2.3. Synthesis of serotonin: tryptophan hydroxylase activity
To test the changes in the synthesis of 5-HT after the vehicle
or tryptophan administration, the in vivo activity of tryptophan
hydroxylase (tryptophan-5-monoxygenase; EC 1.14.16.4), was
assessed. The tryptophan hydroxylase is rate-limiting enzyme
for the synthesis of 5-HT and was determined by measuring the
accumulation of 5-hydroxytryptophan (5-HTP) within 45 min
after inhibition of the aromatic L-amino acid decarboxylase
(EC 4.1.1.28) by a maximally effective dose of NSD 1015
(100 mg/kg, i.p.) (Carlsson and Lindqvist, 1973). The 5-HTP-
accumulation method is the most commonly used assay system
to monitor the in vivo rate of tryptophan hydroxylation in the
brain. The synthesis of 5-HTP was measured in three brain
regions: hippocampus, striatum and hypothalamus.
2.4. Brain samples and chromatographic analyses
The animals were killed by decapitation and their brain was
quickly removed and dissected on an ice-cold plate into
hippocampus, hypothalamus and neostriatum (striatum),
following the anatomical guidelines of the Avian Nomencla-
ture Forum (2004), and then were stored at K80 8C for further
analysis. The brain regions were weighed, placed individually
into cold tubes containing 1 ml of 0.4 M HClO4, 0.01%
K2EDTA and 0.1% Na2S2O5 and homogenized with an Ultra-
Turrax homogenizer (type Tp 18/10). The homogenate was
centrifuged at 40,000 g for 15 min at 4 8C. The resulting
supernatant was filtered through 0.45 mm syringe filters
(Spartan-3, Aldrich Chemical, Milwaukee, Wis., USA) and
aliquots (20–40 ml) were injected into the HPLC system for
determination of 5-HTP (the precursor in 5-HT synthesis),
5-HT and its metabolite, the 5-hydroxyindoleacetic acid
(5-HIAA) as described previously (see Fig. 1 for representative
chromatograms). A Spherisorb S3 ODS1 C18 reversed-phase
column (3 mm particle size range, 4.6 mm!10 cm) coupled to
a Tracer ODS2 C18 (2–5 mm particle size range) pre-column
(Teknokroma) were used. The mobile phase consisted of 0.1 M
KH2PO4; 2.1 mM octane sulphonic acid; 0.1 mM K2EDTA,
2 mM NaCl and 12% (vol/vol) methanol (pH 2.7–2.8, adjusted
with 85% H3PO4) and was pumped at a flow rate of 0.8 ml/min
with a Waters M-510 solvent delivery system. The compounds
were detected electrochemically by means of a cell with a
glassy carbon-working electrode with an applied potential of
C0.75 V against an in situ Ag/AgCl reference electrode
(ISAAC; Waters Concorde Electrochemical detector). The
current produced was monitored using an interphase Waters
bus SAT/IN Module connected to a computer. The concen-
trations of 5-HTP, 5-HIAA and 5-HT in a given sample were
calculated by interpolating the corresponding peak height into
a parallel standard curve using the software Millennium32
(Waters).
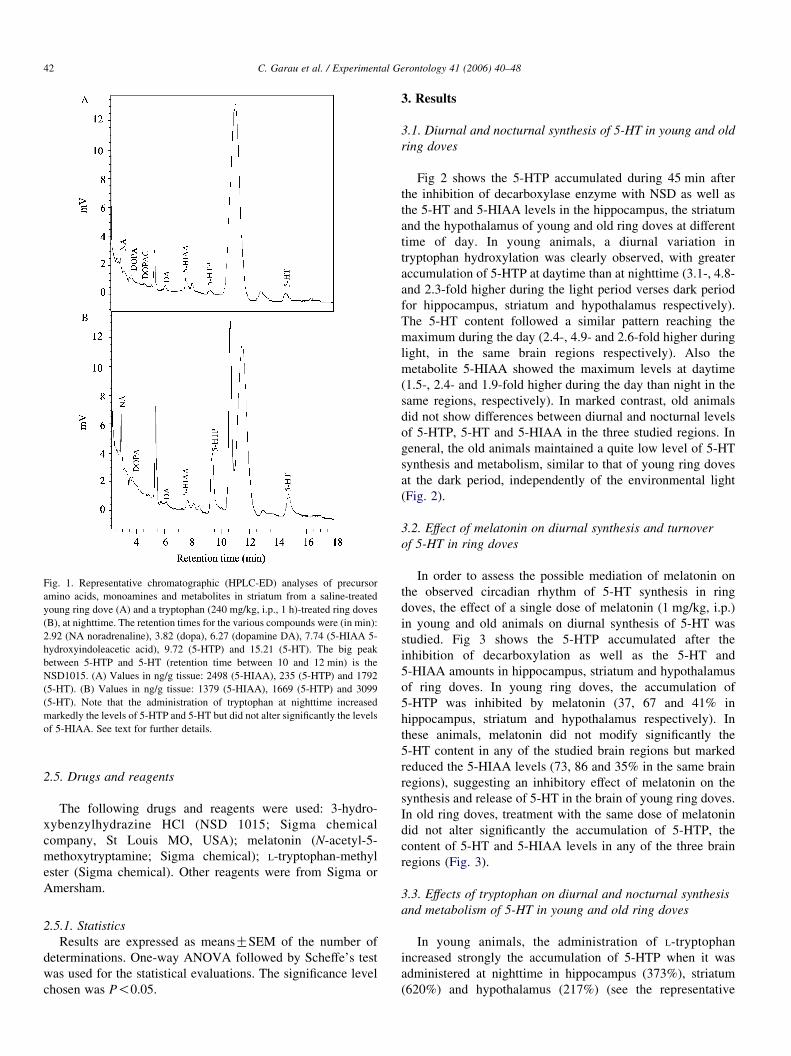
Fig. 1. Representative chromatographic (HPLC-ED) analyses of precursor
amino acids, monoamines and metabolites in striatum from a saline-treated
young ring dove (A) and a tryptophan (240 mg/kg, i.p., 1 h)-treated ring doves
(B), at nighttime. The retention times for the various compounds were (in min):
2.92 (NA noradrenaline), 3.82 (dopa), 6.27 (dopamine DA), 7.74 (5-HIAA 5-
hydroxyindoleacetic acid), 9.72 (5-HTP) and 15.21 (5-HT). The big peak
between 5-HTP and 5-HT (retention time between 10 and 12 min) is the
NSD1015. (A) Values in ng/g tissue: 2498 (5-HIAA), 235 (5-HTP) and 1792
(5-HT). (B) Values in ng/g tissue: 1379 (5-HIAA), 1669 (5-HTP) and 3099
(5-HT). Note that the administration of tryptophan at nighttime increased
markedly the levels of 5-HTP and 5-HT but did not alter significantly the levels
of 5-HIAA. See text for further details.
C. Garau et al. / Experimental Gerontology 41 (2006) 40–4842
2.5. Drugs and reagents
The following drugs and reagents were used: 3-hydro-
xybenzylhydrazine HCl (NSD 1015; Sigma chemical
company, St Louis MO, USA); melatonin (N-acetyl-5-
methoxytryptamine; Sigma chemical); L-tryptophan-methyl
ester (Sigma chemical). Other reagents were from Sigma or
Amersham.
2.5.1. Statistics
Results are expressed as meansGSEM of the number of
determinations. One-way ANOVA followed by Scheffe’s test
was used for the statistical evaluations. The significance level
chosen was P!0.05.
3. Results
3.1. Diurnal and nocturnal synthesis of 5-HT in young and old
ring doves
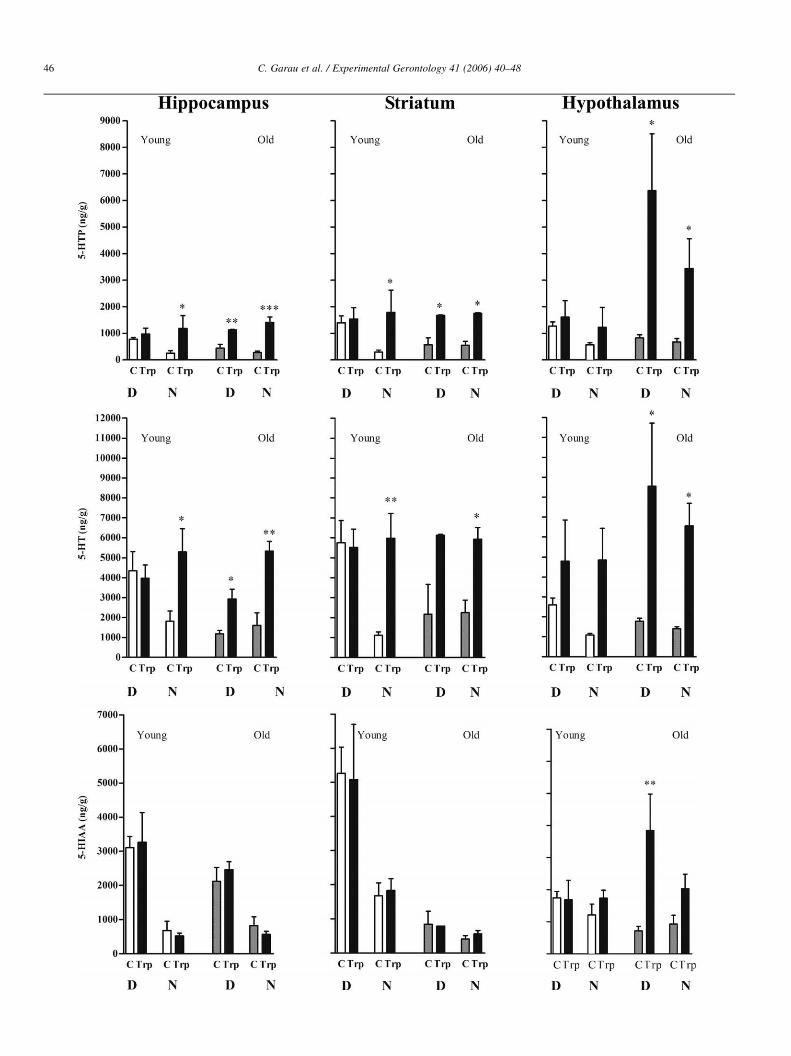
Fig 2 shows the 5-HTP accumulated during 45 min after
the inhibition of decarboxylase enzyme with NSD as well as
the 5-HT and 5-HIAA levels in the hippocampus, the striatum
and the hypothalamus of young and old ring doves at different
time of day. In young animals, a diurnal variation in
tryptophan hydroxylation was clearly observed, with greater
accumulation of 5-HTP at daytime than at nighttime (3.1-, 4.8-
and 2.3-fold higher during the light period verses dark period
for hippocampus, striatum and hypothalamus respectively).
The 5-HT content followed a similar pattern reaching the
maximum during the day (2.4-, 4.9- and 2.6-fold higher during
light, in the same brain regions respectively). Also the
metabolite 5-HIAA showed the maximum levels at daytime
(1.5-, 2.4- and 1.9-fold higher during the day than night in the
same regions, respectively). In marked contrast, old animals
did not show differences between diurnal and nocturnal levels
of 5-HTP, 5-HT and 5-HIAA in the three studied regions. In
general, the old animals maintained a quite low level of 5-HT
synthesis and metabolism, similar to that of young ring doves
at the dark period, independently of the environmental light
(Fig. 2).
3.2. Effect of melatonin on diurnal synthesis and turnover
of 5-HT in ring doves
In order to assess the possible mediation of melatonin on
the observed circadian rhythm of 5-HT synthesis in ring
doves, the effect of a single dose of melatonin (1 mg/kg, i.p.)
in young and old animals on diurnal synthesis of 5-HT was
studied. Fig 3 shows the 5-HTP accumulated after the
inhibition of decarboxylation as well as the 5-HT and
5-HIAA amounts in hippocampus, striatum and hypothalamus
of ring doves. In young ring doves, the accumulation of
5-HTP was inhibited by melatonin (37, 67 and 41% in
hippocampus, striatum and hypothalamus respectively). In
these animals, melatonin did not modify significantly the
5-HT content in any of the studied brain regions but marked
reduced the 5-HIAA levels (73, 86 and 35% in the same brain
regions), suggesting an inhibitory effect of melatonin on the
synthesis and release of 5-HT in the brain of young ring doves.
In old ring doves, treatment with the same dose of melatonin
did not alter significantly the accumulation of 5-HTP, the
content of 5-HT and 5-HIAA levels in any of the three brain
regions (Fig. 3).
3.3. Effects of tryptophan on diurnal and nocturnal synthesis
and metabolism of 5-HT in young and old ring doves
In young animals, the administration of L-tryptophan
increased strongly the accumulation of 5-HTP when it was
administered at nighttime in hippocampus (373%), striatum
(620%) and hypothalamus (217%) (see the representative

Fig. 2. Diurnal and nocturnal synthesis and metabolism of 5-HT in young (light bars) and old (dark bars) animals, at the onset of daytime (8.0 h) or nighttime (20.0 h)
in the hippocampus, striatum and hypothalamus. Animals received NSD1015 45 min before the sacrifice. Bars represent meansGSEM derived from 4 to 5 animals
of 5-HTP accumulated along 45 min, 5-HT and the metabolite 5-HIAA, in ng/g of wet tissue. One way ANOVA followed by Scheffe’s test was used for statistical
evaluation *P!0.05; **P!0.01; ***P!0.001 when compared with the diurnal young group.
C. Garau et al. / Experimental Gerontology 41 (2006) 40–48 43
chromatogram corresponding to striatum in Fig. 1), but no
significantly changes were observed after the lighttime
administration of tryptophan (24, 10 and 26% in the same
brain regions) (Fig. 4). These results indicate that, under control
conditions, the limiting enzyme of 5-HT synthesis—tryptophan
hydroxylase—was saturated by its substrate, at day but not at

Fig. 3. Acute effect (1 h) of melatonin (1 mg/kg, i.p.) (dark bars) on the accumulation of 5-HTP, 5-HT and 5-HIAA of young and old ring doves at daytime (8.0 h), in
hippocampus, striatum and hypothalamus. Ring doves received saline (light bars) or melatonin (dark bars) 15 before NSD1015 and were sacrificed after another
45 min. Bars represent meansGSEM derived from four animals of 5-HTP accumulated along 45 min, 5-HT and the metabolite 5-HIAA, in ng/g of wet tissue. One
way ANOVA followed by Scheffe’s test was used for statistical evaluation *P!0.05, **P!0.01, ***P!0.001 when compared with the corresponding saline
control group.
C. Garau et al. / Experimental Gerontology 41 (2006) 40–4844

C. Garau et al. / Experimental Gerontology 41 (2006) 40–48 45
night. In the same way, L-tryptophan significantly elevated the
content of 5-HT in all brain regions only when it was
administered at nighttime (190, 412 and 126% in hippo-
campus, striatum and hypothalamus, respectively), but did not
modify 5-HT content after lighttime administration (Fig. 4).
Interestingly, the metabolism of 5-HT, reflected by the
5-HIAA levels, was not modified after the administration of
L-tryptophan neither at day nor at night in any brain region
(Fig. 4), suggesting that the increased 5-HT synthesis
observed after nighttime L-tryptophan administration could
have been used for melatonin synthesis. In marked contrast
with the young animals, the administration of L-tryptophan to
old ones increased the 5-HTP accumulation, both at day (158,
196 and 680% in hippocampus, striatum and hypothalamus,
respectively) and at night (402, 218 and 408%, respectively),
indicating that, in old animals under control conditions, the
tryptophan hydroxylase enzyme was not saturated by its
substrate either at day and at night (Fig. 4). L-tryptophan also
significantly increased the contents of 5-HT in all brain
regions when it was administered at day (148, 181 and 379%,
respectively) and at night (231, 164 and 368%, respectively)
(Fig. 3). The 5-HIAA levels only were consistently increased
after diurnal administrarion of L-tryptophan in the hypothala-
mus (439%) (Fig. 4).
4. Discussion
5-HT is a neurotransmitter involved in many functions
throughout the brain, as the synchronisation of the circadian
clock located in the suprachiasmatic nucleus (SCN) and its
fundamental role as precursor in the melatonin synthesis.
Ageing brings many changes in the function of the circadian
timing system, so the present work found changes in the
rhythmic 5-HT synthesis with age, and studied the effects of
tryptophan supply on the circadian 5-HT synthesis rhythm.
A diurnal cycle in tryptophan hydroxylation was clearly
observed in young animals, with a major daytime synthesis and
metabolism of 5-HT. In line with the present results, a
circadian dependency of 5-HT release, with higher levels
during light phase was also observed in hippocampal (Monnet,
2002) and hypothalamic slices of rats (Blier et al., 1989). The
tryptophan hydroxylase protein levels detected by immunoau-
toradiography also showed circadian changes in rats (Malek
et al., 2004). Variation of 5-HT responses throughout the day
has been also described (Klein and Moore, 1979; Martin,
1991). In addition, abundant evidences indicate that melatonin
synchronizes various circadian neural and hypothalamic
endocrine processes (Dubocovich, 1995). However, the
participation of melatonin in the circadian rhythm of 5-HT
release has been questioned from in vitro studies in the
hypothalamus of rats (Cardinali et al., 1975). In fact, melatonin
Fig. 4. Acute effects (1 h) of tryptophan (240 mg/kg, i.p.) (black bars) on the accumul
of daytime (8.0 h) and nighttime (20.0 h), in hippocampus, striatum and hypothalam
before NSD1015 and were sacrificed after another 45 min. Tryptophan-treated an
derived from 4 to 5 animals of 5-HTP accumulated along 45 min, 5-HT and the met
test was used for statistical evaluation *P!0.05, **P!0.01 when compared with
reduced the spontaneous 5-HT release in slices of rat
hippocampus (Monnet, 2002) during the dark phase. Trying
to asses the possible mediation of melatonin on the observed
circadian rhythm of 5-HT synthesis in young ring doves, the
effect of a single dose of melatonin on diurnal synthesis of
5-HT was studied in the present work revealing an inhibitory
effect of melatonin on the synthesis and release of brain 5-HT.
Moreover, it was found that melatonin reduced the synthesis
and metabolism of 5-HT down to nocturnal levels, suggesting a
physiological role of melatonin in the 5-HT circadian synthesis
and release. Also agreeing with our results, a marked reduction
of 5-HT content in the rat hypothalamus was observed after the
administration of pharmacological doses of melatonin during
the light period (Lin and Chuang, 2002). Different experimen-
tal approaches in vitro (Monnet, 2002; Cardinali et al., 1975)
and in vivo (Miguez et al., 1994) showed an inhibitory effect of
melatonin on 5-HT release during the dark but not during the
light phase. Pinealectomy in rats (Miguez et al., 1995) and in
birds (Cassone et al., 1983) also modified the release of 5-HT
confirming the modulatory role of melatonin. In this context,
the present study demonstrated that the circadian rhythm of
5-HT synthesis in vivo in different brain regions of ring doves
was dependent on the presence of melatonin. It is well known
that the secretion of this hormone by the pineal gland during
the night synchronizes several processes (Dubocovich, 1995;
Monnet, 2002) including the 5-HT synthesis in ring doves,
as observed in the present work. Although the mechanism
mediating the action of melatonin on 5-HT synthesis remains
unknown, it has been reported that some effects of melatonin
were antagonized by the 5-HT1A antagonist pindolol (Lin
and Chuang, 2002). In this sense, it is well known that
the activation of 5-HT1A receptors inhibits 5-HT synthesis and
release (Adell et al., 1993; Esteban et al., 1999).
In marked contrast to young animals, the absence of a
circadian rhythm in 5-HT synthesis and metabolism was
observed in the old animals. A deficient effect of melatonin on
diurnal synthesis and metabolism of 5-HT in old ring doves in
comparison to young ones was also observed. These results are
consistent with the deteriorated melatonin rhythm during aging
(Reiter, 1992) that likewise weakens and become desynchro-
nized other circadian rhythms (Reiter, 1995). In ring doves of
the same age groups that the studied in the present work, the
circadian rhythm of serum melatonin showed a clear biphasic
cycle in young ring doves but absence of circadian rhythm was
observed in old animals (Terron et al., 2002). In addition, it is
known that the number and the sensitivity of melatonin
receptors decline with age (Gauer et al., 1998; Zhadanova,
2005). It has been observed numerous age dependent
physiological changes in brain 5-HT. For instance, age related
reductions in the binding of 5-HT receptors have been found in
the brain of humans (Rosier et al., 1996; Wang et al., 1995) and
ation of 5-HTP, 5-HT and 5-HIAA of young and old ring doves both at the onset
us. Control young (light bars) and old (dark bars) ring doves received saline 15
imals received tryptophan 15 min before NSD. Bars represent meansGSEM
abolite 5-HIAA, in ng/g of wet tissue. One way ANOVA followed by Scheffe’s
the corresponding saline control group.
"
C. Garau et al. / Experimental Gerontology 41 (2006) 40–4846

C. Garau et al. / Experimental Gerontology 41 (2006) 40–48 47
importantly in the SCN of rodents (Duncan et al., 2000) where in
addition the spontaneous firing of neurons showed a day/night
rhythm in young but not in old mice (Nygard et al., 2005).
Another important trait of the aging is the reduction in the
amplitude of the diurnal rhythms. In this context, our results
indicate that the diurnal 5-HT synthesis and metabolism were
strongly reduced in old animals compared with the observed in
young ring doves. Studies in humans showed a decrease of
serum/plasma tryptophan concentration related with aging and
associated with an enhanced indoleamine (2,3)-dioxygenase
(IDO) activity, which degrades tryptophan to form kynurenine
derivatives (Frick et al., 2004). In agreement, old ring doves
showed a significant decrease in the plasma melatonin levels
compared with young animals (Terrron et al., 2002) a result
which was already reported in other animal species (Turek
et al., 1999).
On the other hand, the rate of pineal melatonin synthesis is
dependent on the 5-HT levels. It also has been reported that
both tryptophan administration and a high plasma ratio of
tryptophan/neutral amino acids increases the availability of
brain tryptophan and consequently the 5-HT levels (Esteban
et al., 2004; Fernstrom and Wurtman, 1971). At night, when the
synthesis of melatonin is activated, the increased 5-HT
stimulated melatonin production (Esteban et al., 2004; Hajak
et al., 1991). On the contrary, the blocker of 5-HT synthesis
parachlorophenylalanine (PCPA), also inhibits the melatonin
release (Miguez et al., 1997). In order to examine the role of
substrate supply for the rhythmic synthesis of 5-HT, tryptophan
was administered at the beginning of either light or dark phases
to the different age groups of ring doves. The administration of
L-tryptophan to young animals, strongly increased the synthesis
of 5-HT only at nighttime, probably approaching the substrate
saturation of tryptophan hydroxylase and indicating that, under
control conditions, the limiting enzyme of 5-HT synthesis,
tryptophan hydroxylase, is saturated by its tryptophan substrate
at day but not at night. As the hydroxylation of tryptophan is
the rate-limiting step in the synthesis of 5-HT, tryptophan
hydroxylase determines the physiological concentration of
5-HT in vivo. Interestingly, the metabolism of 5-HT reflected
on the 5-HIAA levels was not modified after L-tryptophan
injection neither at day nor at night, suggesting that the
increased synthesis of 5-HT at night could have been used to
the synthesis of melatonin, which is known that fundamentally
occurs during night in mammals (Borjigin et al., 1995) and
birds (Bernard et al., 1997).
In marked contrast with young animals, the administration
of L-tryptophan to old animals increased 5-HT synthesis,
irrespective of the time of administration. This suggests that,
under control conditions, tryptophan hydroxylase was always
far from being saturated by its tryptophan substrate. Moreover,
the 5-HIAA levels were strongly increased in hypothalamus
after L-tryptophan ingestion, but only during lighttime. This
indicates an increase in 5-HT metabolims at daytime but not at
nightime and suggests again an increased use of the 5-HT for
melatonin synthesis.
Taken together, the results confirm that the synthesis of
5-HT and probably melatonin can be modulated by tryptophan
ingestion. The available 5-HT and melatonin results to be
dependent, first on an adequate dietary supply of tryptophan,
and second on the balance between 5-HT’s use as a
neurotransmitter and its availability as precursor for melatonin
synthesis, which is deeply dependent on the environmental
light. If the decrease in 5-HT and melatonin which normally
occurs as animals age could be prevented, perhaps some
complaints of aging could also be delayed. In this aspect, the
supplemental administration of tryptophan might aid to
improve some age-related degenerative conditions.
Acknowledgements
This investigation was supported by DGICYT Grant BFI
2002-04583-C02-029. The authors wish to thank the gently
technical assistance of David Moranta in this work. The two
first authors contributed equally to this work.
References
Adell, A., Carceller, A., Artigas, F., 1993. In vivo brain dialysis study of the
somatodendritic release of serotonin in the Raphe nuclei of the rat: effects
of 8-hydroxy-2-(di-n-propylamino)tetralin. J. Neurochem. 60, 1673–1681.
Azmitia, E.C., Segal, M., 1978. An autoradiographic analysis of the differential
ascending projections of the dorsal and median raphe nuclei in, the rat.
J. Comp. Neurol. 79, 641–667.
Barassin, S., Raison, S., Saboureau, M., Bienvenu, C., Maitre, M., Malan, A.,
Pevet, P., 2002. Circadian tryptophan hydroxylase levels and
serotonin release in the suprachiasmatic nucleus of the rat. Eur.
J. Neurosci. 15, 833–840.
Bernard, M., Iuvone, P.M., Cassone, V.M., Roseboom, P.H., Coon, S.L.,
Klein, D.C., 1997. Avian melatonin synthesis: photic and circadian
regulation of serotonin N-acetyltransferase mRNA in the chicken pineal
gland and retina. J. Neurochem. 68, 213–224.
Blier, P., Galzin, A.M., Langer, S.Z., 1989. Diurnal variation in the function of
serotonin terminals in the rat hypothalamus. J. Neurochem. 52, 453–459.
Borjigin, J., Wang, M.M., Snyder, S.H., 1995. Diurnal variation in mRNA
encoding serotonin N-acetyltransferase in pineal gland. Nature 378, 783–
785.
Card, J.P., Moore, R.Y., 1989. Organization of lateral geniculate-hypothalamic
connections in the rat. J. Comp. Neurol. 284, 135–147.
Cardinali, D.P., Nagle, C.A., Freire, F., Rosner, J.M., 1975. Effects of
melatonin on neurotransmitter uptake and release by synaptosome-rich
homogenates of the rat hypothalamus. Neuroendocrinology 18, 72–85.
Carlsson, A., Lindqvist, M., 1973. In-vivo measurements of tryptophan
and tyrosine hydroxylase activities in mouse brain. J. Neural. Transm. 34,
79–91.
Cassone, V.M., Lane, R.F., Menaker, M., 1983. Daily rhythms of serotonin
metabolism in the medial hypothalamus of the chicken: effects of
pinealectomy and exogenous melatonin. Brain Res. 289, 129–134.
Cutrera, R.A., Ouarour, A., Pevet, P., 1994. Effects of the 5-HT1a receptor
agonist 8-OH-DPAT and other non-photic stimuli on the circadian rhythm
of wheel-running activity in hamsters under different constant conditions.
Neurosci. Lett. 172, 27–30.
Czeisler, C.A., Dumont, M., Duffy, J.F., Steinberg, J.D., Richardson, G.S.,
Brown, E.N., Sanchez, R., Rios, C.D., Ronda, J.M., 1992. Association of
sleep-wake habits in older people with changes in output of circadian
pacemaker. Lancet 340, 933–936.
Dubocovich, M.L., 1995. Melatonin receptors: are there multiple subtypes?
Trends Pharmacol. Sci. 16, 50–56.
Duncan, M.J., Short, J., Wheeler, D.L., 1999. Comparison of the effects of
aging on 5-HT7 and 5-HT1A receptors in discrete regions of the circadian
timing system in hamsters. Brain Res. 829, 39–45.

C. Garau et al. / Experimental Gerontology 41 (2006) 40–4848
Duncan, M.J., Crafton, C.J., Wheeler, D.L., 2000. Aging regulates 5-HT(1B)
receptors and serotonin reuptake sites in the SCN. Brain Res. 856, 213–219.
Esteban, S., Llado, J., Sastre-Coll, A., Garcia-Sevilla, J.A., 1999. Activation
and desensitization by cyclic antidepressant drugs of alpha2-autoreceptors,
alpha2-heteroreceptors and 5-HT1A-autoreceptors regulating monamine
synthesis in the rat brain in vivo. Naunyn Schmiedeberg’s Arch. Pharmacol.
360, 135–143.
Esteban, S., Nicolaus, C., Garmundi, A., Rial, R.V., Rodriguez, A.B., Ortega,
E., Ibars, C.B., 2004. Effect of orally administered L-tryptophan on
serotonin, melatonin, and the innate immune response in the rat. Mol. Cell.
Biochem. 267, 39–46.
Fernstrom, J.D., Wurtman, R.J., 1971. Brain serotonin content: physiological
dependence on plasma tryptophan levels. Science 173, 149–152.
Frick, B., Schroecksnadel, K., Neurauter, G., Leblhuber, F., Fuchs, D., 2004.
Increasing production of homocysteine and neopterin and degradation of
tryptophan with older age. Clin. Biochem. 37, 684–687.
Gauer, F., Schuster, C., Poirel, V.J., Pevet, P., Masson-Pevet, M., 1998.
Cloning experiments and developmental expression of both melatonin
receptor Mel1A mRNA and melatonin binding sites in the Syrian hamster
suprachiasmatic nuclei. Brain Res. Mol. Brain Res. 60, 193–202.
Glass, D.J., Grossman, G.H., Farnbauch, L., Di Nardo, L., 2003. Midbrain
raphe modulation of nonphotic circadian clock resetting and 5-HT release
in the mammalian suprachiasmatic nucleus. J. Neurosci. 23, 7451–7460.
Hajak, G., Huether, G., Blanke, J., Blomer, M., Freyer, C., Poeggeler, B.,
Reimer, A., Rodenbeck, A., Schulz-Varszegi, M., Ruther, E., 1991. The
influence of intravenous L-tryptophan on plasma melatonin and sleep in
men. Pharmacopsychiatry 24, 17–20.
Hay-Schmidt, A., Vrang, N., Larsen, P.J., Mikkelsen, J.D., 2003. Projections
from the raphe nuclei to the suprachiasmatic nucleus of the rat. J. Chem.
Neuroanat. 25, 293–310.
Kawano, H., Decker, K., Reuss, S., 1996. Is there a direct retina-raphe-
suprachiasmatic nucleus pathway in the rat? Neurosci. Lett. 212, 143–146.
Klein, D.C., Moore, R.Y., 1979. Pineal N-acetyltransferase and hydroxyindole-
O-methyltransferase: control by the retinohypothalamic tract and the
suprachiasmatic nucleus. Brain Res. 174, 245–262.
Krajnak, K., Kashon, M.L., Rosewell, K.L., Wise, P.M., 1998. Aging alters the
rhythmic expression of vasoactive intestinal polypeptide mRNA but not
arginine vasopressin mRNA in the suprachiasmatic nuclei of female rats.
J. Neurosci. 18, 4767–4774.
Lin, M.T., Chuang, J.I., 2002. Melatonin potentiates 5-HT(1A) receptor
activation in rat hypothalamus and results in hypothermia. J. Pineal Res. 33,
14–19.
Malek, Z.S., Pevet, P., Raison, S., 2004. Circadian change in tryptophan
hydroxylase protein levels within the rat intergeniculate leaflets and raphe
nuclei. Neuroscience 125, 749–758.
Martin, K.F., 1991. Rhythms in neurotransmitter turnover: focus on the
serotonergic system. Pharmacol. Ther. 51, 421–429.
Miguez, J.M., Martin, F.J., Aldegunde, M., 1994. Effects of single doses and
daily melatonin treatments on serotonin metabolism in rat brain regions.
J. Pineal Res. 17, 170–176.
Miguez, J.M., Martin, F.J., Aldegunde, M., 1995. Effects of pinealectomy and
melatonin treatments on serotonin uptake and release from synaptosomes of
rat hypothalamic regions. Neurochem. Res. 20, 1127–1132.
Miguez, J.M., Simonneaux, V., Pevet, P., 1997. The role of the intracellular and
extracellular serotonin in the regulation of melatonin production in rat
pinealocytes. J. Pineal Res. 23, 63–71.
Monnet, F.P., 2002. Melatonin modulates [3h]serotonin release in the
rat hippocampus: effects of circadian rhythm. J. Neuroendocrinol. 14,
194–199.
Moore, R.Y., Eichler, V.B., 1972. Loss of a circadian adrenal
corticosterone rhythm following suprachiasmatic lesions in the rat.
Brain Res. 42, 201–206.
Myers, B.L., Badia, P., 1995. Changes in circadian rhythms and sleep quality
with aging: mechanisms and interventions. Neurosci. Biobehav. Rev. 19,
553–571.
Nygard, M., Hill, R.H., Wikstrom, M.A., Kristensson, K., 2005. Age-related
changes in electrophysiological properties of the mouse suprachiasmatic
nucleus in vitro. Brain Res. Bull. 65, 149–154.
Penev, P.D., Zee, P.C., Wallen, E.P., Turek, F.W., 1995. Aging alters the
phase-resetting properties of a serotonin agonist on hamster circadian
rhythmicity. Am. J. Physiol. 268, R293–R298.
Penev, P.D., Zee, P.C., Turek, F.W., 1997. Quantitative analysis of the age-
related fragmentation of hamster 24-h activity rhythms. Am. J. Physiol.
273, R2132–R2137.
Reiter, R.J., 1992. The ageing pineal gland and its physiological consequences.
Bioessays 14, 169–175.
Reiter, R.J., 1995. The pineal gland and melatonin in relation to aging: a
summary of the theories and of the data. Exp. Gerontol. 30, 199–212.
Rosier, A., Dupont, F., Peuskens, J., Bormans, G., Vandenberghe, R., Maes,
M., de Groot, T., Schiepers, C., Verbruggen, A., Mortelmans, L., 1996.
Visualization of loss of 5-HT2A receptors with age in healthy volunteers
using (18F)altanserin and positron emission tomographic imaging.
Psychiatry Res. 68, 11–22.
Scarbrough, K., Losee-Olson, S., Wallen, E.P., Turek, F.W., 1997. Aging and
photoperiod affect entrainment and quantitative aspects of locomotor
behavior in Syrian hamsters. Am. J. Physiol. 272, R1219–R1225.
Stephan, F.K., Zucker, I., 1972. Circadian rhythms in drinking behavior and
locomotor activity of rats are eliminated by hypothalamic lesions. Proc.
Natl Acad. Sci. USA 69, 1583–1586.
Terron, M.P., Cubero, J., Marchena, J.M., Barriga, C., Rodrıguez, A.B., 2002.
Melatonin and aging: in vitro effect of young and mature ring dove
physiological concentrations of melatonin on the phagocytic function of
heterophils from old ring dove. Exp. Gerontol. 37, 421–426.
Turek, F.W., Penev, P., Zhang, Y., Van Reeth, O., Zee, P., 1995. Effects of age
on the circadian system. Neurosci. Biobehav. Rev. 19, 53–58.
Turek, F.W., Zee, P., Van Reeth, O., 1999. Melatonin and aging. Adv. Exp.
Med. Biol. 460, 435–440.
Van Reeth, O., Zhang, Y., Zee, P.C., Turek, F.W., 1992. Aging alters feedback
effects of the activity-rest cycle on the circadian clock. Am. J. Physiol. 263,
R981–R986.
Wang, G.J., Volkow, N.D., Logan, J., Fowler, J.S., Schlyer, D., MacGregor,
R.R., Hitzemann, R.J., Gur, R.C., Wolf, A.P., 1995. Evaluation of age-
related changes in serotonin 5HT2 and dopamine D2 receptor availability in
healthy human subjects. Life Sci. 56, PL249–PL253.
Zee, P.C., Rosenberg, R.S., Turek, F.W., 1992. Effects of aging on entrainment
and rate of resynchronization of circadian locomotor activity. Am.
J. Physiol. 263, R1099–R1103.
Zhadanova, I.V., 2005. Melatonin as a hypnotic: Pro. Sleep Med. Rev. 9, 51–65.