adaptation in visuomanual tracking depends on intact proprioception
TRANSCRIPT

Adaptation in Visuomanual TrackingDepends on lntact Proprioception
Olivier GuedonGabriel GauthierUnité Mixte de RechercheMouvement et PerceptionFaculté des Sciences du SportUniversité de la Méditerranée
ABSTRACT. The role of arm proprioception in motor learningwas investigated in experiments in which, by moving the arm, sub-jects followed the motion of a target displayed on a monitorscreen. Adaptive capabilities were tested in visuomanual trackingtasks following alterations in the relationship between the observ-er's actual arm movement and visual feedback of the arm move-ment given by a cursor motion on the screen. Tracking perform-ance and adaptive changes, measured in terms of spatiotemporalerror, tracking trajectory curvature, and spatial gain, were com-pared in 7 control subjects (CSs) and in 1 deafferented subject(DS). CSs adapted appropriately to altered visuomanual relation-ships; those changes were present in trials immediately afterrestoration of normal scaling. In contrast, although the DS modi-fied his tracking strategy from trial to trial according to the alteredconditions, he did not show plastic changes in internal visuoman-ual scaling. Like the results of prismatic adaptation experiments,the present results suggest that arm proprioception contributes tothe plastic changes that follow alterations in the scaling of visuo-manual gain.
Key words: adaptation, deafferentation, motor control, oculoman-ual tracking, proprioception, visuomanual scaling
he way individuals learn the spatiotemporal relation-ship between the central command of arm movement
and its effects, as perceived through vision or propriocep-tion, is still not fully understood. Our purpose in the presentstudy was to investigate the learning of scaled movementsof the arm in relation to a moving visual target when therelationship between the visual target and arm movement ischanged. That situation is of interest because we often per-form arm movements that have consequences beyond inter-
Jonathan ColeClinical Neurological SciencesUniversity of SouthamptonSouthampton General HospitalUnited Kingdom
Journal of Motor Behavior, 1998, Vol.30, No.3, 234-248
Jean-Louis VercherJean BlouinUnité Mixte de RechercheMouvement et PerceptionFaculté des Sciences du SportUniversité de la Méditerranée
for instance, the intent of the driver is not to make a circu-lar motion with the arms but to achieve a smooth change inthe trajectory of the car. When controlling a computermouse, the operator's intention is to move a cursor on ascreen, and he is less concerned with the precise movementof the hand or the mouse itself. Such tasks involve visu-al-motor transformations perceived, in part, through visualfeedback (Jeannerod, 1991; Soechting, Lacquaniti, &Terzuolo, 1986). Burnod et al. (1992) have addressed thequestion of where and how in the brain such visual-motortransformations occur. Those transformations must belearned, although it is clear that some are more easilylearned than others (Droulez & Cornilleau, 1986).
If the relationship between the mouse and cursor motionis homomorphic, the operator becomes more efficient thanif the mouse's anterior-posterior or forward-backward scal-ing is unequal (Bedford, 1994). Hand drawing with mirror-reversed visual feedback (Lajoie et al.,1992) and pointingwith optical prisms (Haris, 1963; Lacknet 1914; Redding& Wallace, 1988) that produce a mismatch between visualfeedback of the hand and the target and the position of thearm perceived through proprioception are the tasks thathave been studied in some experiments. The positions of thehand and target are likely to be represented as maps coded
Correspondence address: Olivier Guedon, UMR Mouvement etPerception, Faculté des Sciences du Sport, Université de laMéditerranée C.P 910, 163 avenue de Luminy, 13288 MarseilleC e dex 9, Franc e. E - mail addre s s : g ue don @ Iap s. univ -mr s.frpersonal space. When moving tlie steering wheel of a car,
234

within visual, proprioceptive, and motor spaces. Normally,there is correspondence between visual and motor spaces;
that is, the visual and motor representations match eachother (Bedford, 1994). Although the process of visuomanu-al rearrangement following visual alteration with opticaldevices such as prisms is not fully understood, there is nowagreement that reorganization involves both visual and pro-prioceptive information (Harris, 1963; Lackner, 1974) andthat adaptation also has two separable mechanisms: anadaptive coordination, which is a highJevel process thatcan be strategically controlled by attentional processes, and
a realignment, which is a low-level process automaticallyactivated with a spatial discordance (Redding, Rader, &Lucas, L992). Propnoception seems to be involved in bothcontrol and adaptive control of human movements.
Proprioception and On-Line Control of Movement
The role of afferent information in the control of thelength, speed, and force of a movement has been investigat-ed by a number of researchers (Bossom & Ommaya, 1968;Cole & Sedgwick, 1992; Gauthier & Mussa Ivaldi, 1988;Goldberger, 1980). Polit and Bizzi (1978) suggested thatproprioception plays a greater role in updating and adjust-ing central programs subserving learned motor patternsthan in the servo control of movements. Loss of proprio-ception leads to compensation over time, however, and
other sensory cues (particularly vision) are used (Goldberg-er, 1980) and alternative cognitive strategies are developed(Cole, 1995). That compensation is only partially effective,however. Whereas deafferented subjects (DSs) demonstratenormal or minimally affected muscle strength (Rothwell etal., 1982), their motor performance is disturbed in variousways, especially when visual control is prevented (Bard,Fleury Teasdale, Paillard, & Nougier, 1995; Blouin, Gau-thier, Vercher, & Cole, 1996).
Studies of DSs' wrist movements have shown that pos-tural maintenance and movement accuracy are dependenton visual control (Miall, Haggard, & Cole, 1995; Nougier etal., 1996; Sanes, Mauritz, Dalakas, & Evarts, 1985). Thosestudies have shown that, without vision, the limb drifts inrandom directions, and during intended movements the tra-jectory and thç movement's end point are perturbed. Thoseauthors concluded that whereas deafferentation does notprevent movement initiation, it is essential both for finecontrol of movement and for postural maintenance. Otherimpairments in movements in DSs include instability in theproduction of handwriting ellipses without vision (Teasdale
et a1.,1993), errors in the extent and direction of aimed armmovements, and a lack of compensation for inertia differ-ence (inertia anisotropy) in different movement directions(Ghez, Gordon, & Ghilardi, 1995; Gordon, Ghilardi, Coop-er, & Ghez, 1994; Gordon, Ghilardi, & Ghez, 1995).
Proprioception and Adaptive Control of Movement
In the earlier mentioned studies of deafferented humanbeings and animals, the role of proprioception in the on-line
September 1998, Vol.30, No.3
Proprioception in 2D Visuomanual Adaptation
conffol of movement has been investigated. Fewer experi-ments with DSs have addressed the question of the role ofproprioception in the longer term control of movement andleaming processes. Taub and Goldberg (1974) srudied theadaptive changes resulting from prism-induced displacementin a hand reaching task. They observed that adaptation wasgreater in a group of deafferented monkeys than in controlsand concluded that the adaptive changes involved were madeeasier in the deafferented group because of the absence ofconflict between visual and kinesthetic feedback to motorcommand. Bossom and Ommaya (1968) also showed adap-
tation to prismatic deviation in surgically deafferented mon-keys. They concluded that recalibration of visuomanual scal-ing occurred through altered efferent motor commands. Thatinterpretation was not supported by Bard et al. (1995), how-ever, who observed in a prismatic adaptation task carried outby 2 DSs that 1 DS adapted to the level of control subjects(CSs), whereas the other DS failed to adapt at all. The DSwho adapted had intact neck proprioception, whereas theother DS did nol The authors therefore concluded that adap-tation requires neck propriocepton, which allows transfer ofspatial information from the eye-head to the hand-head ref-erence frame, thereby enabling adjustments in the visuoman-ual relationship to take place.
The results of those experiments do not completely clar-ify the role of proprioception in prismatic adaptation and inother forms of motor learning, however, in part because ofthe principal measures of adaptation that were used in thosestudies. In most studies, researchers have determined adap-tation by comparing the results of tests in which subjectsgenerally performed without vision before and after expo-sure to an altered visuomanual relationship. In measuringadaptation, however, analyses of subjects' movements dur-ing exposure to an alteration would also be informative,because one could see how the subject improves his initialperformance. In the present experiment, we reexamined therole of arm proprioception in adaptive control of movementby altering the visuomanual scaling in a task dependent onvisual feedback for accuracy. That experimental paradigmallowed us not only to measure adaptation of the subjectsbut also to follow their adaptive process by quantifying theinduced adaptive changes occurring during the alteration.We deduced the role of proprioception in that adaptation bycomparing the level of adaptive changes between CSs and asubject deprived of proprioception (DS).
MethodSubjects
Seven CSs (23-53 years old) and 1 DS (43 years old)were tested. The DS suffered an acute, purely sensory neu-ropathy when he was 19 years old. Neurophysiological testshave confirmed the loss of large myelinated fibers below theneck. For further description of the DS's neuropathy, see
Cole and Paillard (1995) and Cole (1995). All subjects gave
written consent to participate in the Local Ethics Commit-tee's approved protocols.

O. Guedon, G. Gauthier, J. Cole, J.-L. Vercher, & J. Blouin
Experimental Setup
In the experimental paradigm used (Figure 1), the subject'sarm tracked targets presented on a videographic screen. Thesubject was seated in front of a stand, facing the screen. Weadjusted the height of the upper horizontal board of the standto just above the subject's shoulder level to allow free armmoton underneath and yet prevent vision of the arm (and therest of the body). The screen was positioned vertically I m infront of the subject. The target was a green dot,2.5 mm indiameter, and the tracking cursor was a white cross, 4 mm x4 mm. We monitored two-dimensional (horizontal and verti-cal) hand motion by means of an infrared device (Hamamat-su; 0.2-mm spatial resolution). We adjusted the amplitude ofthe output signals driving the cursor on the screen to ensure,in the normal condition (see the following), a one-to-one met-rical relationship, that is, so that a given continuous displace-ment of the hand in any direction produced a similar dis-placement (i.e., same magnitude and direction) of the cursoron the screen.
Tracking Instructions
Subjects were instructed to follow with their preferredarm the motion of a target displayed on the screen. The sub-ject's hand movement occurred in a frontoparallel plane.When first exposed to the setup, the subjects were shownthe coupling between the motion of the hand-carried in-
frared emitter and the motion of the cursor on the screen.They were instructed to move the hand and, hence, the cur-sor so that the cursor coincided temporally and spatallywith the moving-dot target.
Adaptation Conditions
We used a circular frajectory (15-cm diameter) drawn in5 s to test adaptation. The target always started to move up-ward from the left, at equatorial level. Each trial consisted ofa single circular trajectory that always started from the leftcardinal position. The various test and training tracking trialsduring a typical adaptation session are listed in Table 1. Forthe four experimental conditons, a session was composed ofseveral series. We exposed the subjects to a first series of 5
trials with visual feedback of the movement (VCL) and a nor-mal visuomanual relationship to determine the VCL baselinetracking performance. Another series of 5 trials was then per-formed without visual feedback of the hand movement(VOL) as a reference for preadaptive exposure. Without par-ticular notice, the horizontal or the vertical visuomanual rela-tionship, or both, were then increased by a factor of 2.5, and
a series of 50 VCL tracking trials was then performed. A2.5-times increase in the visuomanual relationship means that ahand movement of 1 cm produces a cursor movement of 2.5cm. At the end of the adaptation series, 10 VOL ffials wereperformed, which allowed us to measure the effect of train-
o: Target
+ : Arm-DrivenCursor
lnfraredEmitter
lnfraredCamera
FIGURE 1. Schematic representation of the experimental apparatus. The observer was seat-ed facing a graphic screen on which a target moved along a circular path in 5 s. Vision of theann was prevented by a horizontal board fixed at shoulder level. The observer's arm steeringmotion (with the arm extended) occurred in a frontoparallel plane. Two-dimensional handmotion was monitored with an infrared device whose sensitivity ensured a one-to-one metri-cal relationship between arm motion and the resulting cursor motion on the screen. See alsoTable 1 for horizontal and vertical sensitivities along the four experimental sessions.
f--\[\
<*--/
236 Journal of Motor Behavior

TABLE 1
Test and Training Tracking Trials During a TypicalAdaptation Session: Horizontal and Vertical Sen-sitivities Along the Four Experimental Sessions
Sensitivitydirection
Trial number
l-5 6-10 il-60 6t-70 71-80 8r-90VCL VOL VCL VOL VCL VCL
No alteration
HorizontalVertical
lX1XIlX1XI
1X1X1X2.5X
Horiz.ontal alteration
HorizontalVertical
1X2.5X2.5 1
lXtX11
2.5 I2.5 I
Vertical alteration
HorizontalVertical
I2.5
Horizontal and vertical alteration
HorizontallX2.5XVerticallX2.5X
Note. Hor. = horizontal; Ver. = vertical; VCL = tracking conditionwith visual feedback of the hand movement; VOL = tracking con-dition without visual feedback of the hand movement. An X in theVOL series means that the sensitivity had no meaning becausetracking occurred without vision of the cursor.
ing. In the VOL condition, because the subject did not see thecursor, the visuomanual alteration had no meaning. We thencarried out further training to compensate for any decrease inadaptation that might have occuned during those previous l0VOL trials. For that training, subjects performed 10 trials ofVCL in the altered condition. We ran a final series of 10 VCLtrials with a normal visuomanual relationship to evaluate thepostadaptation affect and the time course of the return to thepreadaptation state. Throughout the sessions, subjects wereverbally encouraged to minimize tracking enor.
All subjects underwent four experimental conditions, start-ing with the one that included no alteration in the visuoman-ual relationship (control condition). We used that condition as
a control to evaluate response stability over a series of 50 tri-als. After that session, we tested subjects with all of the threealtered tracking conditions-horizontal, vertical, or com-bined horizontal and vertical changes of sensitivity-pseudo-randornly selected to minimize cumulative training effects. Aminimum rest period of 2lu was allowed between sessions,and most subjects completed the experiment over 2 or 3 days.
The data reported here refer exclusively to experimentscarried out with a 2.5-times visuomanual sensitivity increasefactor. However, in side experiments, we also tested 0.5-, 2-,
September 1998, Vol. 30, No. 3
Proprioception in 2D Visuomanual Adaptation
and 3-times increase factors, which showed similar results,
with adaptive changes proportional to the alteration.
Data Recording and Analysis
Vertical and horizontal components ofboth the target and
the hand trajectories recorded during test and training trialswere sampled at the rate of 100 Hz and saved on disk foroff-line analyses. Each performance during VCL trials was
measured in terms of spatial tracking error and trajectorycurvature. Using factors that are defined later as horizontaland vertical gains, we measured the adaptation.
Tracking error. For each trial, we evaluated the trackingerror by calculating the cumulative sum of the length of theeffor vectors that joined at the same sampled time the posi-tion of the target and the position of the cursor (Magenes,
Vercher, & Gauthier, 1992; Yiviani, Campadelli, & Mou-noud, 1987). That criterion takes into account both temporalerror (the cursor may be on the target path but not at the righttime) and spatial error (the cursor may be off the target path).That tracking effor, expressed in millimeters, is linearlydependent on sample rate, which remained constant (100Hz) for all conditions and all subjects. A spatial error wascalculated as the absolute value of the area between targetand hand paths (so that differences directed in the oppositedirection did not cancel out).
Cursor-to-target lag. T\e lag between target and cursorwas calculated according to the following rule: For a par-ticular position of the target at a time /,, the cursor-to-targetdistance when the cursor varied from time (n - I s) to time(1, + I s) was computed. That distance was minimum at atime /", and the lag was evaluated as the difference (t, - t,).Note that if h < t" < 4+ 1, then the target was leading; andif r, - 1 1 t" 1 tt, then the cursor was leading.
Curvature. Because the target trajectory curvature had a
constant value as a result of its circular nature, all curvaturechanges above or below that of the target can be attributedto tracking errors, colrections, or both. That was particular-ly relevant when the DS's trajectories were described,because those appeared to be made of more or less linearsegments (low curvature) with sharp changes of direction(high curvature). We found the curvature radius for eachpoint of the path by calculating the radius of a circular arcfitting the two extreme points and the center points of aseries of 41 consecutive points of the trajectory. We count-ed the number of peaks (greater than twice the target radius)and troughs (less than half the target radius) to obtain a rel-evant criterion of the smoothness of the trajectory.
Visual tracking gain. The visual tracking gain wasdefined by the horizontal and vertical extents of two rectan-gles, one containing the tracking response and the othercontaining the target trajectory. We computed the veftical orhorizontal dimensions of the rectangles from a Fourieranalysis of the vertical and horizontal components of thetrajectory and found the ratio of the magnitude of the firstharmonic of the cursor signal and the magnitude of the firstharmonic of the target signal. That method can be applied to
O. Guedon, G. Gauthier, J. Cole, J.-L. Vercher, & J. Blouin
unclosed cursor trajectories such as those that were gener-ally observed in VOL trials. Average gains were calculatedas the mean of five single consecutive trials.
Normalized gains. Tlte normalized gain in a directionwas defined by the ratio between the gain in the VOL seriesbefore and after the adaptation series.
Adaptation curves. The time course of the adaptation ingain after training with the altered visuomanual relationshipswas plotted as a function of the number of trials. Using a
power function,flx) = CIXF, we derived a mathematical fit.
Statistics
The various criteria selected to assess the performance ofthe subjects vr'ere averaged over the trials in each tracking
condition and over the 7 CSs. Statistical comparison (Stu-dent f test) was carried out between the overall average fromthe CSs and the average from the DS.
Results
In the analysis, arm tracking responses were separatedinto spatial and temporal errors with respect to the targettrajectory. Therefore subjects' performances are illustratedin both of those domains. Because all CSs behaved in a sim-ilar manner and yielded similar results, we selected spatialand spatiotemporal recordings from only 1 subject to illus-trate the overall behavior. However, the tracking error andthe changes of gain following the adaptive period are pre-sented in Table 2 for all subjects, including the DS.
TABLE 2Mean Values t Standard Deviations of Tracking Error, Spatial Error, Horizontal and Vertical Amplitudes,
Cursor-to-Target Lags, Number of Peaksn and Number of Troughs for all CSs and the DS
Subject no.
Measure CSI CS2 cs3 CS4 CS5 cs6 CS 7 CS Mean DS
Mean tracking error over 40 VCL trtals (mm)
MSD
4,9771.578
3,393762
4,174844
4,381 5,446 4,821 1,402551
4,085 t2,942r,688 4,863908 1,792 1,018
Mean spatial error over 20 VCL trials (mm2)
MSD
3,2t51,429
)'7))991
985 t,244284 436
t,t45 1,506 1,765562
1,797r,076
3,57rr,454357 420
Mean horizontal amplitude (mm)
MSD
150.34.2
r45.63.9
152.1 153.6 151.5 151.84.5
155.44.5
15r.5 152.84.8 9.32.8 4.t 3.0
Mean vertical amplitude (mm)
MSD
153.06.7
150.15.8
150.4 t49.0 152.6 t52.76.5
151.93.6
15t.4 148.45.3 t2.93.3 4.8 4.5
MSD
t2,8724,745
Mean tracking enor oyer 30 VOL trtak @m)8,514 13,855 16,846 18,022 12,346 16,740 r4,t7t 23,8M
3,262 4,803 5,8702.rr5 4.446 4,122 4,340 2,892
Mean lag between target and cursor over 50 trials (ms)
MSD
-10430
46 -18 46 --68 -122 4222
-:7240
-191t7729 1l 26 t7 36
Mean number of peaks over 20 VCL trialsMSD
4.01.0
8.12.0
4.81.3
3.9 3.8 7.32.0
3.92.3
5.12 8.502.26 2.061.0 1.6
Mean number of troughs over 20 VCL trialsMSD
0.80.6
3.61.6
1.9t.4
0.30.6
0.40.7
3.21.3
1.81.0
1.68 4.301.60 1 .03
Nore. CS = control subject; DS = deafferented subject. VCL = tracking condition with visual feedback of the hand movements; VOL = tracking con-dition without visual feedback of the hand movement.
238 Journal of Motor Benavror

DS
/---\
,\-/
FIGURE 2. Single and superimposed VCL tracking responses from a DS (A and C) and 1
CS (B and D). Superimposition of 20 target and arm-steered cursor trajectories emphasizesdifferences in the spatial tracking error between the DS and the CS in terms of both overallamplitude and nature of the error. The DS tended to follow by using straight lines connectedwith curvilinear segments. VOL tracking resulted in large variations in shape and error in theresponse of the DS (E) as compared with that of a CS (F).
CS
ri \ \
\
t- /
I
l
Visual Closed-Loop TFackingThe tracking error between the target and the cursor trajec-
tory in the VCL conditon was morphologically fairly similarin the DS and the CSs (Figure 2A and B). There was, howev-er, one clear difference between the CSs and the DS: Quan-tification of the amplitudes of the tracking error showed theDS to be far less accurate overall and more variable. In Thble2,the average data for the the CSs and the DS and the corre-
September 1998, Vol. 30, No. 3
Proprioception in 2D Visuomanual Adaptation
sponding standard deviations (40 trials) are presented. Theaverage tracking enor,t(7) = 17.3,p <.001, as well as the
variability, t(7) =22.2, O < .001, were larger in the DS than inthe CSs. A corresponding difference was also observed be-
tween the CSs and the DS in ûerms of spatial error, as shownin Table 2. The DS's average spatal error was twice as largeas that calculated over the 7 CSs, t(7) = 5.5, p < .0I.
A qualitative difference between the CSs and the DS

O. Guedon, G. Gauthier, J. Cole, J.-L. Vercher, & J. Blouin
can also be observed in Figure 2A and B. The CSs' cursortrajectories were smooth despite some small intermittentcorrections. The DS's trajectories were less smooth, withthe appearance of quasi-linear segments between majortrajectory corrections. The spatial pattern observed in theDS appeared as if vertical and horizontal displacementswere executed in sequence rather than simultaneously,resulting in a segmented trajectory (see also Figure 2C andD). In the graphs, a much greater spread of trajectories inthe DS than in the CSs is also shown. The data found in
Table 2 show that the mean horizontal and vertical excur-sion amplitudes were not significantly different betweenthe DS and the CSs; ls(7) = 0.58 and2.l8, ps >.05, forhorizontal and vertical amplitudes, respectively. However,the variability of horizontal and vertical amplitudes wassignificantly different, rs(7) - 8.86 and 5.76, ps < .01, forhorizontal and vertical amplitudes, respectively.
A quantitative description of the DS's tracking undervisual feedback control was provided by the average num-ber of peaks and troughs of the curvature function reported
li
II
ti
I
Ji
iI
il
,l:
i-\
iI
1.
i
1
IIj
I
TL
l'
iII
\{
I
DS.VCL
DS.VOL
CS-VCL'F'- - "1" '*,CS-VOL
5cm
L_,"
FIGURE 3. Recording of horizontal (X) and vertical (Y) components of target and cursor ffa-jectories in VOL and VCL tracking for the DS and a CS in the normal visuomanual relatonship.
240 Journal of Motor Behavior
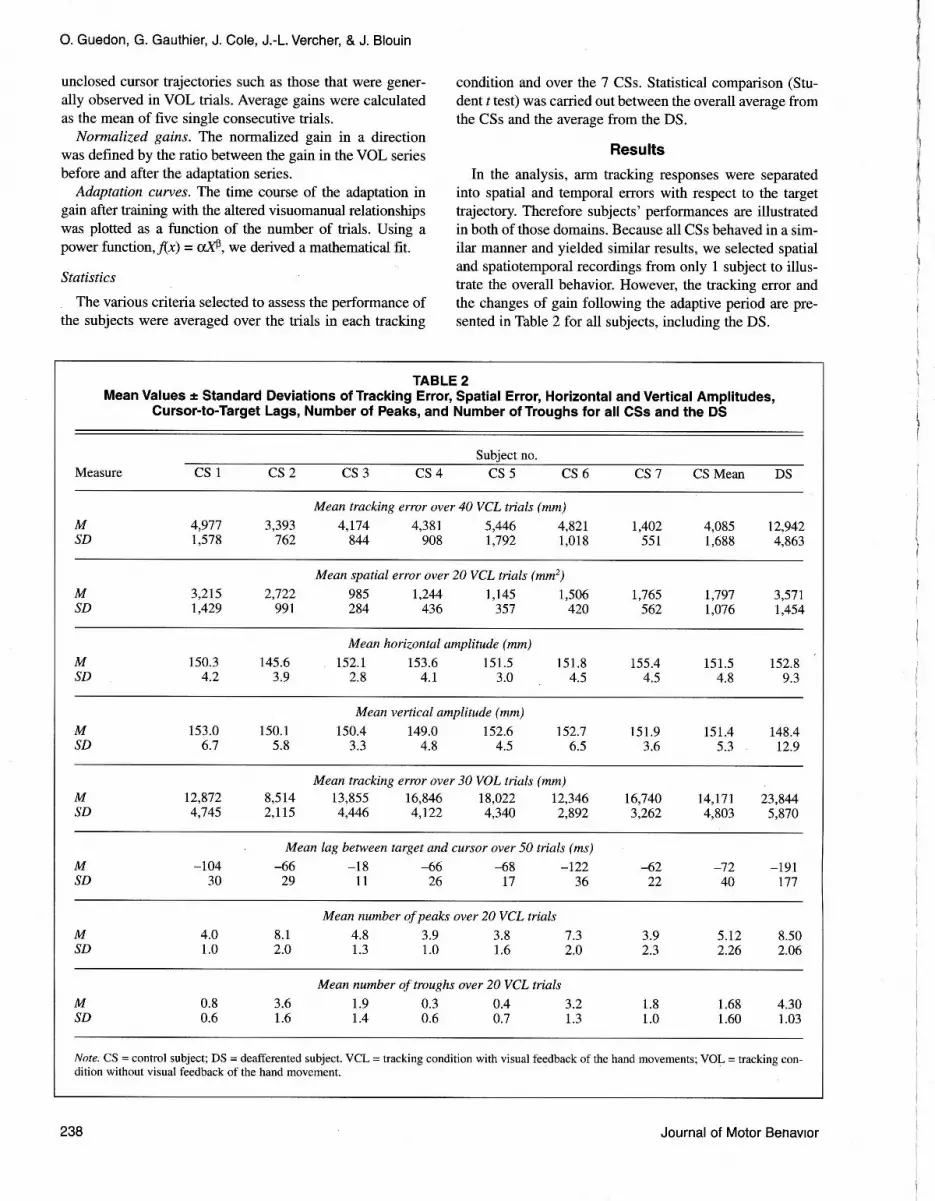
in Table 2. The DS's average value of peaks (8.5 + 2.06),calculated over 20 trajectories, was larger than the corre-sponding CSs' values (5.12 + 2.26). For the troughs, rheDS's average value was 4.30 + 1.03 and corresponding CSs'values were 1.68 + 1.60. Those differences were statistical-ly significant: t(7) = 4.94, p < .01, for peaks; and r(7) = 5.34,p < .01, for troughs.
Time recordings for horizontal and vertical componentsof target and response motions are shown in Figure 3. In therepresentative single trial shown in Figure 3, the DS'srecordings (DS-VOL) showed larger errors than the CSs,(CS-VOL). No systematic analysis was carried out to quan-tify the initial tracking latency (that analysis was only mod-erately informative because the protocol used led to targetmotion that was fairly predictable). Nevertheless, analysisof the time recordings showed that the initial tracking laten-cy was similar in the CSs and in the DS; therefore, their dif-ferences in mean tracking error resulted not from a differ-ence in initial tracking latency but rather from thepersistence of a lag all along the DS's trials. Indeed, thetime records for the DS (DS-VCL) showed a constant phaseshift between target and cursor; whereas, for the CSs (CS-VCL), the initial latency was progressively compensatedfor. To clarify that point, we calculated the cursor-to-targetlag every 250 ms from the beginning of the trial to the 4th s(the last second was not considered because the targetstopped at 4.5 s). In Figure 4 are plotted two curves repre-senting the evolution of the target-to-cursor lag: The firstcurve corresponds to a mean lag over 140 CS trials (20 tri-als for each of the 7 CSs), and the other corresponds to a
September 1998, Vol.30, No.3
Proprioception in 2D Visuomanual Adaptation
mean lag over 2O DS trials. The curves indicate that CSsrapidly compensated for the initial delay, whereas the DSnever achieved such compensation during the entire trial(the mean lag was greater for the DS than for CSs), r(7) =5.77, p < .01.
Visual Open-Loop Tlacking
The tracking error between actual target and cursor ffa-jectories was larger in VOL than in VCL tracking in all sub-jects: (7) = 29.I,p < .001, in CSs; and t(7) - 8.3, p < .001,in the DS. In addition, the rracking path usually did nor rer-minate on the starting point (Figures 2E and F and Table 2).The difference in the tracking error amplitude betweenVCLand VOL was larger in the DS than in rhe CSs, t(7) = 9.6,p < .001. As the series of trials progressed, the distortion ofthe DS's trajectories increased to the point where some ofthe trials could not be analyzed because the amplitudeexceeded the recording space.
Adaptation to Visuomanual Scaling Change:Preexposure Effects
All subjects, including the DS, became immediatelyaware of the alteration in the visuomanual relationshipwhen the sensitivities of the horizontal, vertical, or bothdirections were suddenly changed; and all subjects reactedappropriately to the gain change. In most adaptation exper-iments, emphasis is placed on the changes seen after theexposure period. As an index of adaptation, in prism adap-tation experiments target localization errors are usuallymeasured after exposure in a visually open-loop condition,
^ 0.5Io)-5 o.+(D
Pg 0.3
sE o.z(ûI
0.1
FIGURE 4. Average tracking lag as a function of time for the DS and the cSs. The lag wascalculated as the time difference between the target and the hand-driven cursor position ât thesampling frequency of 100 Hz and was reproduced for clarity on the curve every 250 ms.
Time (s)
241

O. Guedon, G. Gauthier, J. Cole, J.-L. Vercher, & J. Blouin
PRE.ADAPTATION PER.ADAPTATION POST-ADAPTATION
DS A
CS
FIGURE 5. Selected responses in the DS and I CS during an adaptive session to a 2.5-timeschange of the arm-to-target sensitivity in the horizontal direction, illustrating typical trajec-tories in VCL (4, G) and VOL (8, H) tracking. The first VCL tracking trial followlng the iud-den change in arm-to-target relationship was severely affected in the CS (I) as compared withthe DS (C). At the end of the 50-trial training period in the altered condition, thè trackingresponse was considerably improved in the cS (J), and to a lesser extent in the DS (D). Thedecrease of horizontal gain appeared in the first VOL trial following training in the CS (K)and the DS (E). A posteffect is noticeable in the CS (L) and the DS (F) after the visuoman-ual relationship was returned to normal.
although measurements are sometimes also made duringexposure. In the present article, the adaptive changes toaltered visual scaling both during and after exposure interms of tracking effors and gain changes are described.
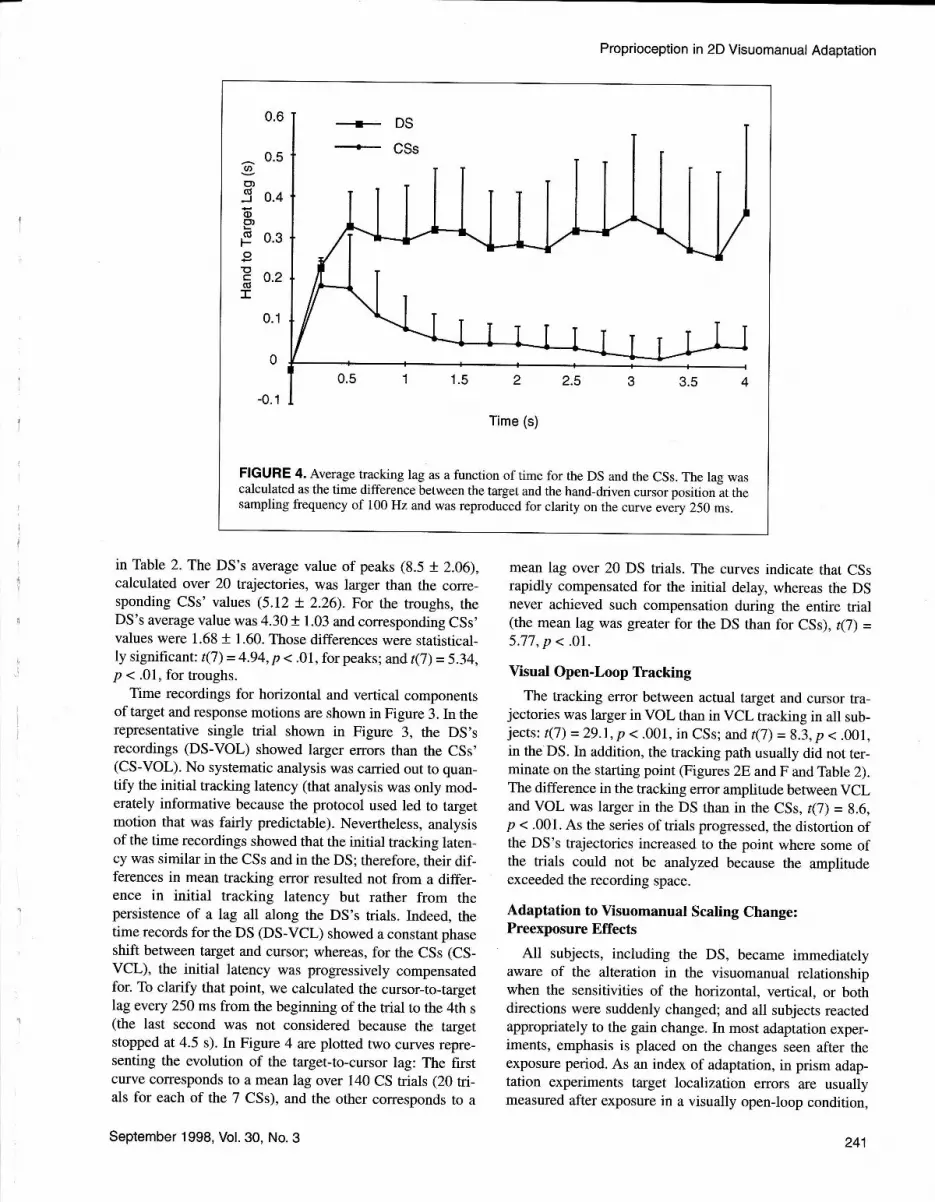
The unaltered condition (preadaptation) yielded the typi-cal trajectories shown in Figure 5A and G in VCL and Fig-ure 58 and H in VOL for the DS and the CSs, respectively.It is worth noting that, with an alteration in gain, precisetracking along a circular path at screen level required thesubject to produce an elliptical hand trajectory with a long-to-short axis ratio of 2.5.In Figure 5I, the frst trial of a CSis shown. If the hand path was reduced along the horizontaldirection, the pursuit error in that direction was not fully
242
compensated for before the end of this first trajectory. InCSs, as the number of trials increased under visual feed-back, the spatial error decreased rapidly. After l0 trials, thetracking trajectory had recovered a fairly circular shape.After 50 trials, CSs had compensated fully for the alteration(Figure 5J). The rate of adaptation, as observed from track-ing curves during the VCL trials, varied somewhat from CSto CS. In Figure 6C is shown the time course of the adap-tive changes in terms of tracking error (modeled by a powerfunction; see the Method section) of all 7 CSs. The rapidimprovement in performance, as a function of the numberof trials, is shown in detail for I CS in Figure 68.
In the first trial with the visuomanual alteration of gain
l
I
t,
Journal of Motor Behavior
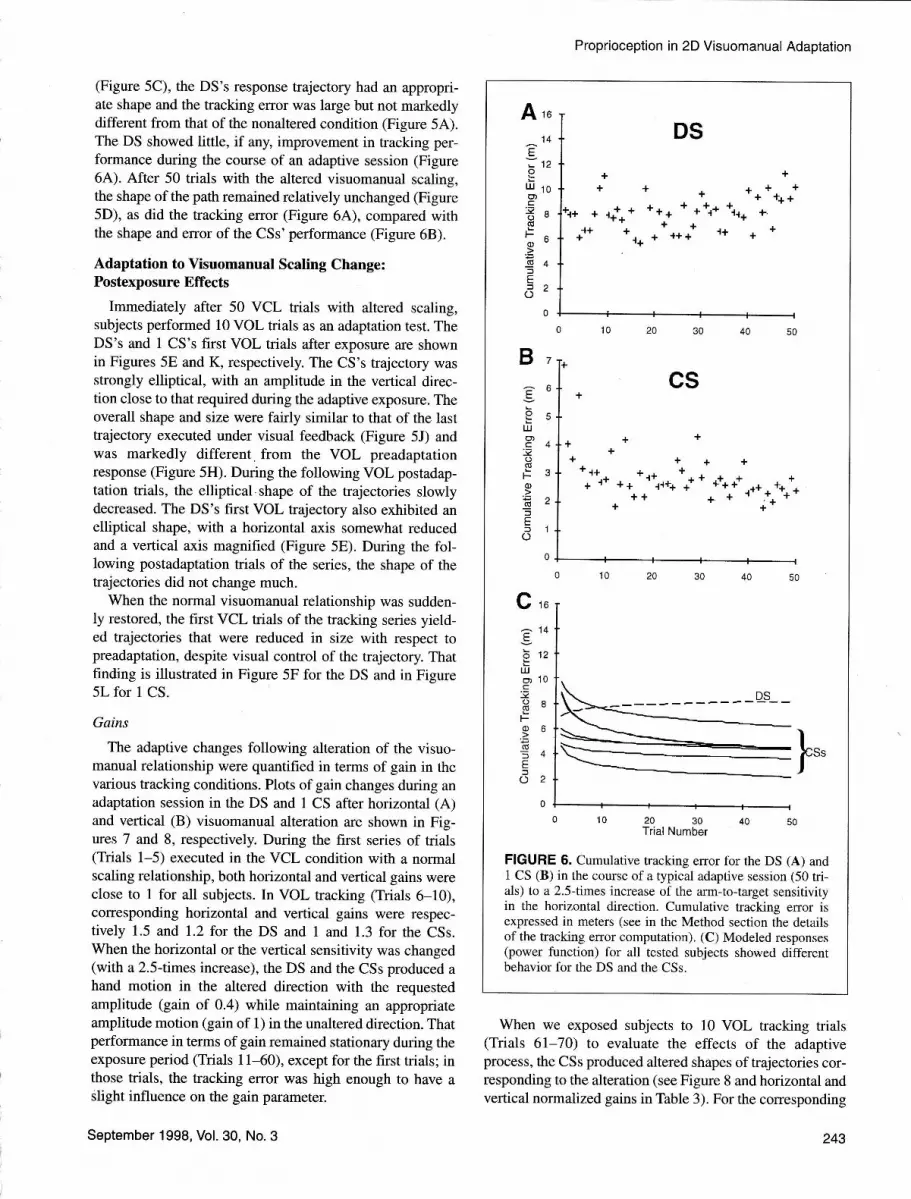
(Figure 5C), the DS's response trajectory had an appropri-ate shape and the tracking error was large but not markedlydifferent from that of the nonaltered condition (Figure 5A).The DS showed little, if any, improvement in tracking per-formance during the course of an adaptive session (Figure6A). After 50 trials with the altered visuomanual scaling,the shape of the path remained relatively unchanged (Figure5D), as did the tracking error (Figure 6A), compared withthe shape and error of the CSs' performance (Figure 68).
Adaptation to Visuomanual Scaling Change:Postexposure Effects
Immediately after 50 VCL trials with altered scaling,subjects performed 10 VOL trials as an adaptation test. TheDS's and 1 CS's first VOL trials after exposure are shownin Figures 5E and K, respectively. The CS's trajectory wasstrongly elliptical, with an amplitude in the vertical direc-tion close to that required during the adaptive exposure. Theoverall shape and size were fairly similar to that of the lasttrajectory executed under visual feedback (Figure 5J) andwas markedly different. from the VOL preadaptationresponse (Figure 5H). During the following VOL postadap-tation trials, the elliptical shape of the trajectories slowlydecreased. The DS's first VOL trajectory also exhibited anelliptical shape, with a horizontal axis somewhat reducedand a vertical axis magnified (Figure 5E). During the fol-lowing postadaptation trials of the series, the shape of thetrajectories did not change much.
When the normal visuomanual relationship was sudden-ly restored, the first VCL trials of the tracking series yield-ed trajectories that were reduced in size with respect topreadaptation, despite visual control of the trajectory. Thatfinding is illustrated in Figure 5F for the DS and in Figure5L for I CS.
Gains
The adaptive changes following alteration of the visuo-manual relationship were quantified in terms of gain in thevarious tracking conditions. Plots of gain changes during anadaptation session in the DS and 1 CS after horizontat (A)and vertical (B) visuomanual alteration are shown in Fig-ures 7 and 8, respectively. During the first series of trials(Trials 1-5) executed in the VCL condition with a normalscaling relationship, both horizontal and vertical gains wereclose to 1 for all subjects. In VOL tracking (Trials 6*10),corresponding horizontal and vertical gains were respec-tively 1.5 and 1.2 for the DS and I and 1.3 for the CSs.When the horizontal or the vertical sensitivity was changed(with a 2.5-times increase), the DS and the CSs produced ahand motion in the altered direction with the requestedamplitude (gain of 0.4) while maintaining an appropriateamplitude motion (gain of 1) in the unaltered direction. Thatperformance in terms of gain remained stationary during theexposure period (Trials 11-60), except for the first trials; inthose trials, the tracking effor was high enough to have aslight influence on the gain parameter.
September 1998, Vol.30, No.3
Proprioception in 2D Visuomanual Adaptation
When we exposed subjects to 10 VOL tracking trials(Trials 6l-70) to evaluate the effects of the adaptiveprocess, the CSs produced altered shapes of trajectories cor-responding to the alteration (see Figure 8 and horizontal andvertical normalized gains in Table 3). For the corresponding
++*
+ *r* *ll+
+++
Aru14
E
-12u10of,aL
o"S4fE=2o-
0
=ôt
9sulo
=o(6È3Q)
Èôz
=Eo
;14È
btzLllor 10ç=Oo6vFo6(g
=4E5o2o
++++ +++
+
B7
Ctu
+
tt++ {* ++
+
+ +
++'
*t1.**1* *++ ***.**a.l***
+'
F.,
Trial Number
FIGURE 6. Cumulative tracking error for the DS (A) and1 CS (B) in the course of a typical adaptive session (50 tri-als) to a 2.5{imes increase of the arm-to-target sensitivityin the horizontal direction. Cumulative tracking error isexpressed in meters (see in the Method section the detailsof the tracking error computation). (C) Modeled responses(power function) for all tested subjects showed differentbehavior for the DS and the CSs.
243

DS
A r't
1.4
1.2.ç,0.8
0.6
0.4
o.2
0
30 40 50
Trial Number
El r.e
1.4
c'6 1
0.8
0.6
o.4
0.2
0
30 40 50
Trial Number
0 5 10VCL VOL'1 x
VCL2.5 x
70VOL VCL
2.5 xVCL1x
FIGURE 7. Time course of horizontal and vertical gainchanges during adaptive exposure to a2.5x increase of thehorizontal (A) and vertical (B) arm-to-target sensitivity(one trial = one 5-s circular trajectory) in the DS. Gain in adirection is defined as the ratio between amplitude of thearm signal first harmonic and the amplitude of the targetsignal in that direction.
O. Guedon, G. Gauthier, J. Cole, J.-L. Vercher, & J. Blouin
trials, the DS modified the shape of the trajectory accordingto the alteration (see normalized gains of Table 3); but, con-trary to expectation, in response to an increase of verticalsensitivity (Figure 7B), the horizontal gain increased. Thedata from all subjects in the three conditions of gain in-crease (horizontal, vertical, and horizontal plus vertical) and
with the nonaltered condition as control (see the Methodsection) are summarized in Table 3. The normalized gains
shown in Table 3 (see the Method section) allowed us toevaluate the changes occurring in the subjects. The DS and
the CSs showed no significantly different behavior in the
nonaltered condition, t(7) = 1.51, p > .1, and t(7) = 1.59,p > .1, respectively; and in the horizontal altered conditions,t(7) = 2.14, p > .05, and (7) = 1. 13, p > .2, respectively. Incontrast, when the visuomanual scaling was altered verti-cally or in both directions, the DS's gain values differed sig-nificantly from those of CSs. For vertical alteration, the
DS's horizontal gain was higher than were those of the CSs,
244
t(7) = 9 .25, p < .0I. For simultaneous horizontal and verti-cal alterations, the DS's horizontal and vertical gains were
higher than the CSs', (7) = l0 and 10.8, p < .01.
The last series of trials (Irials 71-80) was performed in aVCL condition once the normal target-to-hand relationship
had been reestablished. Following the horizontal alteraton,subjects returned to a vertical and horizontal gain of 1 withinone or two trials. Another difference between the CSs and the
DS was observed in VOL trials (Trials 6l-:70) immediatelyafter the end of the training period. Unlike the gain of the DS,
the CSs' gain in the altered direction increased slowly, trialafter trial, toward the preexposure value.
Tracking Error
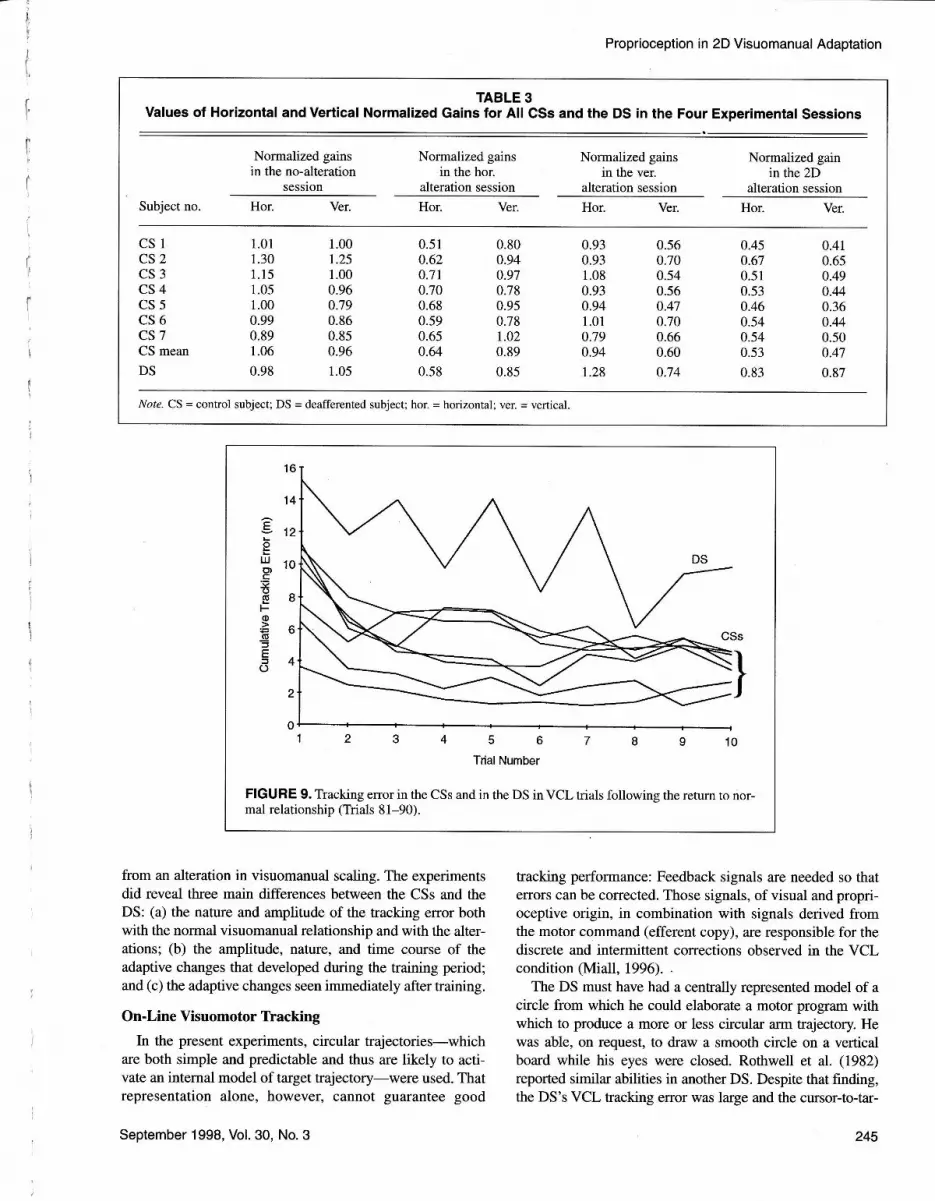
When the normal visuomanual relationship was restored(Trials 81-90), in contrast to the curves of the DS, the CSs'
curves of tracking error fairly decreased toward the preex-
posure level (Figure 9).
Discussion
The experimental design allowed us to determine the
influence ofproprioception in the adaptive changes resulting
CS
A 1.6
1.4
1.2
1
0.8
0.6
0.4
0.2
0
1.4
1.2
1
0.8
0.6
o.4
0.2
0
B
30 40 50
Trial Number
0 5 10
VC L VOL1x
VCL2.5x
70 80 90
VOL VCL VCL2-5x 1x
FIGURE 8. Time course of horizontal and vertical gainchanges (power function) for all tested subjects showed dif-ferent behavior for the DS and the CSs.
Journal of Motor Behavior
Proprioception in 2D Visuomanual Adaptation
I
TABLE 3Values of Horizontal and Vertical Normalized Gains for All CSs and the DS in the Four Experimental Sessions
Normalized gainsin the no-alteration
session
Normalized gainsin the hor.
alteration session
Normalized gainsin the ver.
alteration session
Normalized gainin the 2D
alteration session
Subject no. Hor, Ver. Hor. Ver. Ver.
cs1cs2cs3CS4cs5CS6CS7CS mean
DS
r.0t1.301.151.051.000.990.891.06
0.98
1.001.251.000.960.790.860.850.96
1.05
0.510.620.710.700.680.590.650.64
0.58
0.800.940.970.780.950.781.020.89
0.85
0.930.931.080.930.941.010.790.94
1.28
0.560.700.540.560.470.700.660.60
o.74
0.450.670.510.530.460.540.540.53
0.83
0.410.650.490.440.360.440.500.47
0.87
Nore. CS = control subject; DS = deafferented subject; hor. = horizontal; ver. = vertical.
E
oUJo,go(tFo(s
Ëo
16
14
12
10
I
6
4
2
0
Trial Number
FIGURE 9. Tracking error in the CSs and in the DS in VCL trials following the return to nor-mal relationship (Trials 81-90).
)
from an alteration in visuomanual scaling. The experimentsdid reveal three main differences between the CSs and theDS: (a) the nature and amplitude of the tracking error bothwith the normal visuomanual relationship and with the alter-ations; (b) the amplitude, nature, and time course of theadaptive changes that developed during the training period;and (c) the adaptive changes seen immediately after training.
On-Line Visuomotor Thacking
In the present experiments, circular trajectories-whichare both simple and predictable and thus are likely to acti-vate an internal model of target trajectory-were used. Thatrepresentation alone, however, cannot guarantee good
September 1998, Vol.30, No.3
tracking performance: Feedback signals are needed so thatelrors can be corrected. Those signals, of visual and propri-oceptive origin, in combination with signals derived fromthe motor command (efferent copy), are responsible for thediscrete and intermittent corrections observed in the VCLcondition (Miall, 1996). .
The DS must have had a centrally represented model of a
circle from which he could elaborate a motor program withwhich to produce a more or less circular arm trajectory. Hewas able, on request, to draw a smooth circle on a verticalboard while his eyes were closed. Rothwell et al. (1982)reported similar abilities in another DS. Despite that finding,the DS's VCL tracking error was large and the cursor-to-tar-
245

O. Guedon, G. Gauthier, J. Cole, J.-L. Vercher, & J. Blouin
get lag remained constant (300 ms) in the present study, sug-gesting that the DS relied almost exclusively on on-line visu-al feedback control. That feedback has both a long latencyand a large lag, however, leading to large errors and theapparent low-frequency segmentation in the trajectory.
Without vision, the DS produced trajectories that werelarger than the target, suggesting that the DS's internal rep-resentation was not very accurate and that, in the CSs atleast, part of the gain control was proprioceptive in origin. Asecond difference between CSs' and the DS's VOL trackingwas that whereas CSs produced trajectories that were rela-tively circular, the DS produced vertically oriented ellipses,with the major part of the distortion affecting the lower halfof the trajectory. Because the starting position of each trialwas located to the left of the circle, that larger error mayreflect the fact that position error accumulated with timealong the trajectory. That errôr was not seen in CSs in ourexperiment, although Bock and Eckmiller (1986) did reportsimilar cumulating error. To elucidate that point, we ran aside experiment in which the DS tracked a target that start-ed along a circular path from one of the cardinal positions(top, bottom, right, lef|. In VOL rrials, tracking effor accu-mulated along the trajectory for all starting points and direc-tions, giving further support to the important role of propri-oception as source of feedback for control of armmovement, especially when vision is not available.
Effects of Changing the Visuomanual Relationship
During the adaptation phase, the CSs' performance wasaltered during the first l0 or so trials. In contrast, theresponse of the DS to the visuomanual alteration hadalready stabilized by the end of the lst trial, and, subse-quently, his tracking performance did not improve marked-ly with training in any of the conditions. A similar resultwas seen in VCL trials when, after the exposure to anincrease of gain, the normal relationship was restored. TheDS immediately modified his tracking behavior to take intoaccount the returl to the normal condition, whereas CSsresisted the return to the normal condition for several trials.The finding that the CSs experienced difficulty in recover-ing normal control supports the hypothesis that a plasticchange in visuomanual control takes place as a consequenceof the conflict between visual and proprioceptive informa-tion that results from the scaling change.
Supporl for the idea that visuoproprioceptive conflictleads to adaptive changes comes from recent studies onlearning that have shown that adaptation in a natural or arti-ficial neural system can occur only when there are conflictsbetween actual and expected behavior during the learrringphases (Barto & Gullapalli, 1992; Bullock, Grossberg, &Guenther, 1993). Kawato, Uno, Isobe, and Suzuki (1988)developed a model of the control and learning of move-ments in which the inverse dynamics of the commandedsystem is learned. In the model, the torque error is used as
a teacher for the network. Those studies emphasized therole of effors or conflicts in generating adaptation.
246
An example of apparently improved on-line performancewhen conflict is absent has been found in the minor-draw-ing experiments of Lajoie et" al. (1992). In those experi-ments, a deafferented subject was able to trace a Star ofDavid with mirror-reversed feedback, whereas the CSsfound it very diffrcult. The authors interpreted the difficultyas revealing the effect of conflict between visual and kinet-ic information. In the present experiment, fewer conflictswere present for the DS when the visuomanual relationshipwas changed (the main remaining conflict was betweenefference copy and vision) and adaptive changes developedto a much lesser extent than in the CSs.
Adaptive Changes
Following alterations in the visuomanual scaling, CSsadapted by decreasing the arm-to-target gain in the appro-priate direction (or directions, when vertical and horizontalam-to-target sensitivities were simultaneously affected).All those alterations led also to changes in the DS's visuo-manual scaling, but each alteration was responded to in aparticular way.
The DS's adaptation to an increase of horizontal sensitiv-ity provided results similar to those of the CSs. However, hisadaptation may reflect not a plastic alteration but the natureof the experimental paradigm and his cognitive response toit. The DS's VOL trajectories were kidney shaped; that is,they were larger in their lower halves, reflecting cumulativeelror as the movement progressed. Right at the end of a tra-jectory howeveE the DS turned inward, toward the center of.the circle, to approximately attain his starting position,which may have been a learned "cardinal" position (Miall etal., 1995). That presumably cognitive strategy had the effectof reducing horizontal error in a manner that might be con-fused with a plastic alteration. That was made apparent bythe way in which the effect was independent of adaptation.
When the alteration consisted of an increase of verticalsensitivity, the DS did not reduce the vertical amplitude ofhis movement but instead increased his horizontal ampli-tude. Although all of the subjects detected a change ofvisuomanual relationship when the alteration was applied,some CSs, when questioned after the experiment, respond-ed that they sensed an alteration but attributed it to thewrong direction (i.e., subjects could not tell whether thesensitivity was increased in the attended direction ordecreased in the other). That erroneous cognitive evaluationof the alteration may explain the abnormal increase of hor-izontal gain as a result of the DS's adaptation when he expe-rienced an increase of vertical gain.
When the alteration was an increase of sensitivity in bothdirections, the DS reduced his movements in both direc-tions, but to a lesser extent than did the CSs. The fact thatthe CSs appeared to adapt more rapidly to the increase insensivity in both directions than to the other alterations sug-gests that that it is easier for them to adapt to that change.
One may wonder why changes in visuomanual scalingwere present in the DS after the adaptation phase if no
l
l
Journal of Motor Behavior

!i
improvement in performance during the adaptation phasewas observed. Those results may be explained by consider-ing that following a sudden change in visuomanual sensi-tivity, adaptation occurs in several phases (Redding et al.,1992). In a first phase, highJevel neural processes areinvolved, producing a cognitive response to the alterationthat is achieved with attentional cost. In a second phase, byproprioceptive recalibration a transfer from highJevel tolow-level neural processes occurs. For a deafferented sub-ject, only the first phase of adaptation can occur. That limi-tation could explain the lack of performance improvementof the DS during the adaptive exposure to the alteration andwhy some adaptation took place because of a cognitiveprocess. An explanation for the rapid adjustment of the DSto the alteration may be that the DS has better visuomanualcontrol than the CSs because he can never delegate hismotor response to low-level processes involving proprio-ception. In the absence of proprioception, voluntary motorresponses are always under cognitive control through visu-al feedback. Observations from the DS's everyday activityand from other DSs prove that point (Cole, 1995). In thepresent task, like the DS in Lajoie's (1992) experiment, theDS must have used a cognitive strategy to control his armmovements and employed vision as the sole source of sen-sory feedback.
Conclusions
The present results allow one to conclude that in a circle-drawing task, CSs develop plastic changes similar to thoseobserved in subjects during pointing tasks in which prismsor visual vestibular conflicts affecting the vestibular ocularreflex gain are used. Subjects seem to use proprioceptiveinput to adaptively modify the calibration of the visuoman-ual tracking system in response to an alteration of the visuo-manual relationship. That calibration is updated when con-flicts between proprioception and vision arise. In theabsence of proprioception, the mechanism of rearrangementdoes not operate, even if some contrasting changes can beobserved, because of cognitive conffol and subjects' use oftrial-by-trial visual feedback to maintain accuracy.
ACKNOWLEDGMENTS
The authors wish to express their gratitude to the deafferentedsubject and to all the control subjects for their participation in thisstudy. This experiment was supported by grants from the CentreNational de la Recherche Scientifique, Unité Recherche Associée372,La Fondation pour la Recherche Médicale, La Fondation deFrance, and from the Human Frontier Science Program (HFSP).We thank Paul Cordo and an anonymous reviewer for their helpfulcomments on an earlier version of this article.
REFERENCES
Bard, C., Fleury, M., Teasdale, N., Paillard, J., & Nougier, V.(1995). Contribution of proprioception for calibrating andupdating the motor space. Canadian Journal of Physiology andPharmacology, 7 3, 246-254.
Barto, A. G., & Gullapalli, V. (1992). Neural networks and adap-tive control. NPB Technical Report No.6, Amherst, MA.
September 1998, Vol.30, No.3
Proprioception in 2D Visuomanual Adaptation
Bedford, F. L. (1994). Of computer mice and man. Cahiers de Psy-chologie Cognitive/Cunent Psychology of Cognition, 13,405426.
Blouin, J., Gauthier, G. M., Vercher,J.-L., & Cole, J. D. (1996).Contribution of retinal and extraretinal signals for defining thetarget goal of reaching movements in normal subjects and adeafferented patient. Experimental Brain Research, 109,148*153.
Bock, O., & Eckmiller, R. (1986). Goal directed movements inabsence of visual guidance: Evidence for amplitude rather thanposition control. Expeimental Brain Research, 62, 451458.
Bossom, J., & Ommaya, A. (1968). A visuo-motor adaptation toprismatic transformation of the retinal image in monkeys withbilateral dorsal rhizotomy. Brain, 91, 16l-172.
Bullock, D., Grossberg, S., & Guenther, F. H. (1993). A self orga-nizing neural model of motor equivalent reaching and tool useby multijoint arm. Journal of Cognitive Neuroscience, 5,408-435.
Burnod, Y., Grandguitlaume, P., Otto, I., Donzzi, B., Guigon, E.,Ferraina, S., Johnson, P., & Caminiti, R. (1992). Visuo-motortransformations underlying rum movements towards visual tar-gets: A neural network model of cerebral cortical operations.Journal of Neuroscience, I 2, 1435-1453.
Clower, D. M., Hoffman, J. M., Votow, J. R., Faber, T. L., Woods,R. P., & Alexander, G. E. (1996). Role of parietal cortex in thecalibration of visually guided reaching. Nature, 383, 618421.
Cole, J. (1995). Pride and a daily marathon. Boston, MA: M.LT.Press.
Cole, J. D., & Paillard, J. (1995). Living without touch and periph-eral information about body position and movement: Studieswith deafferented subjects. In J. L. Bermudez, A. Marcel, & N.Eilan (Eds.), The body and the sefi London: M.I.T. Press.
Cole, J. D., & Sedgwick, E. M. (1992). The perceptions of forceand of movement in a man without large myelinated sensoryafferents below the neck. Journal of Phy s iolo gy, 44g, 503-5 15.
Droulez, J., & Cornilleau, V. (1986). Adaptive changes in percep-tual responses and visuomanual coordination during exposureto visual metrical distortion. V.sion Research, 26, 1783-1792.
Gauthier, G. M., & Mussa Ivaldi, F. (1988). Oculo-manual track-ing of visual targets in monkeys. Role of the arm afferent infor-mation in the control of arm and eye movements. ExperimentalBrain Research, 73, 138-154.
Ghez, C., Gordon, J., & Ghilardi, M. F. (1995). Impairments ofreaching movements in patients without proprioception. II.Effects of visual information on accuracy. Joumal of Neuro-phy siolo gy, 7 3, 361-37 2.
Goldberger, M. E. (1980). Motor recovery after lesions. Trends inNeuros cienc e, 3, 288-29 I
Gordon, J., Ghilardi, M. F., Cooper, S. E., & Ghez, C. (1994).Accuracy of planar reaching movements. II. Systematic extenterrors resulting from inertial anisotropy. Experimental BrainResearch, 99, ll2-130..
Gordon, J., Ghilardi, M. F., & Ghez, C. (1995). Impairments ofreaching movements in patients without proprioception. I. Spa-tial errors. Journal of N europlry siolo gy, 7 3, 347 -360.
Harris, C. S. (1963). Adaptation to displaced vision: Visual, motor,or proprioceptive change? Science, 1 40, 812-814.
Jeannerod, M. (1991). The interaction ofvisual and proprioceptivecues in controlling reaching movements. In D. R. Humphrey &H. J. Freund, (F,ds.), Motor control: Concepts and issues ((pp.277 -29I). New York: Wiley.
Kawato, M., Uno, Y., Isobe, M., & Suzuki, R. (1988, April). Hier-archical neural network model for voluntary movement withapplications to robotics. IEEE Control System Magazine, S-15.
Lackner, J. R. (1974). Adaptation to displaced vision: Role of pro-prioception. Perceptual and Motor Skills, 38, 125l-1256.
Lajoie, Y., Paillard, J., Teasdale, N., Bard, C., Fleury M., Forget,
247

rO. Guedon, G. Gauthier, J. Cole, J.-L. Vercher, & J. Blouin
R., & Lamarre,Y. (1992). Mirror drawing in a deafferentedpatient and control subjects: Visuoproprioceptive conflict. Ner-rology, 42, 1 104-1 106.
Magenes, G., Vercher, J. L, & Gauthier, G. M. (1992). Handmovements strategies in telecontrolled motion along 2D trajec_toies. IEEE Transactions on System, Man and Cybàrnetics,- 22,242-257.
Miall, R. C. (1996). Task-dependenr changes in visual feedbackcoltrgl: A frequency analysis of human manual trackjng. Jour_rnl of Motor Behaviot; 28, 125-135.
Miall, R. C., Haggard, P. N., & Cole, J. D. (1995). Evidence of alimited visuo-motor memory used in programming wrist move_ments. Expe rimental Brain Re s earch, t 07, 267 -ZgO.
Nougier, V., Bard, C., Fleury, M., Teasdale, N., Cole, J., Forget, R.,Paillard, J., & Lamarre, Y. (1996). Control of single-joint move-ments in deafferented patients: Evidence for amplitude codingrather than position control. Experimental Brain Research, 16,473482.
Polit, A., & Bizzi, E. (1978). hocesses controlling arm move_ments in monkeys. Science, 201, 1245-1237.
Redding, G. M., Rader, S. D., & Lucas, D. R. (1992). Cognitiveload and prism adaptation. Journal of Motor Behaviôa 24,238-245.
Redding, G. M., & Wallace, B. (1988). Adaptive mechanisms inperceptual-motor coordination: Components of prism adapta_
tion. Journal of Motor Behavior, 20,242-254.Rothwell, J. C., Traub, M. M., Day, B. L., Obeso, J. A., Thomas,
P. K., & Marsden, C. D. (1982). Manual motor performance ina deafferented man. Brain, 105, 515-542.
Sanes, J. N., Mauritz, K., Dalakas, M., & Evarts, E. (19g5). Motorcontrol in humans with large fiber sensory neuropathy. HumanNeurobiology, 4, 101-114.
Soechûng, J. F., Lacquaniti, F., & Terzuolo, C. (1986). Coordina-tion of arm movements in three-dimensional space: Sensorimo-tor mapping during drawing movements. Neuroscience, 17,295-3t1.
Thub, E., & Goldberg, I. A. (1974). Use of sensory recombinationand somatosensory deafferentation techniques in the investiga-tion of sensory-motor integratron. Perception, 3, 3g34}g. -
Teasdale, N., Forget, R., Bard, C., paillard, J., Fleury, M., &Lamarre, Y. (1993). The role of proprioceptive information forthe production of isometric forces and for handwriting tasks.Acta Psychologica, 82, l7g-lgl.
Viviani, P., Campadelli, P., & Mounoud, p. (1987). Visuomanualpursuit tracking of human two-dimensional movements. "/otr_nal of Expertmental Physiology, 13, 62-:7 B.
Submitted April 2, 1996Revised June 4, 1997
Second reyision December 23, 1997
248 Journal of Motor Behavior