activating mutations of n-wasp alter shigella pathogenesis
TRANSCRIPT

Biochemical and Biophysical Research Communications 384 (2009) 284–289
Contents lists available at ScienceDirect
Biochemical and Biophysical Research Communications
journal homepage: www.elsevier .com/locate /ybbrc
Activating mutations of N-WASP alter Shigella pathogenesis
David A. Adamovich a,e, Fumihiko Nakamura b,c, Austen Worth d, Siobhan Burns d, Adrian J. Thrasher d,John H. Hartwig b,c, Scott B. Snapper a,b,*
a Gastrointestinal Unit, Massachusetts General Hospital, Boston, MA 02114, USAb Department of Medicine, Harvard Medical School, Boston, MA 02115, USAc Hematology Division, Brigham & Women’s Hospital, Boston, MA 02115, USAd Molecular Immunology Unit, Institute of Child Health, University College London, London, WC1N 1EH, UKe Biological and Biomedical Sciences Graduate Program, Harvard Medical School, Boston, MA 02115, USA
a r t i c l e i n f o
Article history:Received 2 April 2009Available online 18 April 2009
Keywords:N-WASPShigellaActin tailsBacterial pathogenesisAutophagyActin polymerization
0006-291X/$ - see front matter � 2009 Elsevier Inc. Adoi:10.1016/j.bbrc.2009.04.050
* Corresponding author. Address: GastrointestinalHospital, Boston, MA 02114, USA
E-mail address: [email protected] (S.B. S
a b s t r a c t
The pathogenesis of Shigella requires binding to the host protein N-WASP. To examine the roles of struc-tural conformation and phospho-regulation of N-WASP during Shigella pathogenesis, mutant N-WASPconstructs predicted to result in a constitutively open conformation (L229P and L232P) or either a phos-pho-mimicking (Y253E) or phospho-disruptive (Y253F) structure were constructed. Pyrene actin assaysdemonstrated that the N-WASP L229P and L232P constructs are constitutively active. Despite theincrease in actin polymerization seen in vitro, cell lines expressing N-WASP L229P and L232P supportedshorter actin tails when infected with Shigella. Shigella actin tails were unchanged in cells expressing N-WASP phospho-regulation mutant proteins. Shigella invasion, intracellular, and intercellular motilitywere not altered in cells expressing N-WASP L229P or L232P. However, plaque numbers were increasedin cells expressing N-WASP L229P and L232P. These data demonstrate that N-WASP structural conforma-tion is an important regulator of Shigella pathogenesis in distinct segments of its lifecycle.
� 2009 Elsevier Inc. All rights reserved.
Introduction Steps controlling activation and regulation of N-WASP follow-
Shigella flexneri is a bacterial pathogen of the human intestinethat usurps control of the actin cytoskeletal machinery for its path-ogenesis [1]. Intracellular Shigella movement is facilitated bydirecting host cell actin polymerization exclusively at one pole ofthe bacteria – a process known as actin-based motility [1]. Theforce generated by the polymerizing actin is sufficient to propelShigella through the cytoplasm and into neighboring cells [1].
Shigella regulates the host actin cytoskeleton through its inter-actions with the host protein N-WASP [1]. N-WASP is a memberof the WASP family of cytoskeletal regulators, which interact withvarious signaling molecules to transduce Arp2/3-mediated actinpolymerization [2]. In its native conformation, intramolecularforces hold WASP and N-WASP in an auto-inhibited closed, hair-pin-like structure [3,4]. Binding of activated Cdc42 to N-WASP dis-rupts this closed conformation, presumably opening up themolecule with a release of auto-inhibition, permitting binding tothe Arp2/3 complex and inducing actin polymerization [2]. Oncein an open conformation, N-WASP can also be modulated by phos-pho-regulation. Phosphorylation of tyrosine residue Y253 of N-WASP leads to enhanced actin polymerization activity in vivo andin vitro [5].
ll rights reserved.
Unit, Massachusetts General
napper).
ing Shigella IcsA binding are largely unknown. Cdc42 is not abso-lutely required for Shigella-mediated actin-based motility [6]. Therole of IcsA and other bacterial/host molecules in the relief ofN-WASP auto-inhibition is largely unknown. Moreover, the roleof phospho-regulation of N-WASP during Shigella-mediated actin-based motility remains controversial. An early study failed toobserve phosphorylated tyrosine residues located at the site ofShigella-mediated actin polymerization [7]. However, recent datasuggests that the phosphorylation status of N-WASP may play animportant role in the ability of Shigella to form actin tails [8].
Here, we sought to assess the role of N-WASP regulationthrough structural conformation or phosphorylation on Shigella-mediated actin-based motility by generating specific mutationsin N-WASP predicted to regulate structural conformation andphosphorylation, and assessing their behavior during infection.
Materials and methods
Bacterial strains and cell culture. Shigella flexneri serotype 2a strain2457T was used for all studies and grown on tryptic soy broth (TSB)with agar containing Congo red as described [17]. Individual red col-onies were grown overnight in liquid TSB and then diluted andgrown in fresh media until mid-log phase on the day of infection.N-WASP�/� mouse embryonic fibroblasts [17] were maintained in

D.A. Adamovich et al. / Biochemical and Biophysical Research Communications 384 (2009) 284–289 285
Dulbecco’s modified Eagle’s medium (DMEM), supplemented with10% bovine calf serum (with iron supplement) and L-glutamine.
Generation of N-WASP mutations. Point mutations in rat N-WASPwere generated using an overlapping PCR strategy with WT rat N-WASP as a template. The FLAG peptide DNA sequence (50-atggacta-caaggacgacgatgacaag-30) was inserted before the stop codon at theC-terminal end of N-WASP. All constructs were sequenced to verifypresence of single point mutations.
Actin polymerization assay. Pyrene actin assays were performedas previously described [23]. Pyrene actin, Cdc42, and the Arp2/3complex were purified and Cdc42 was loaded with GTPcS. Assayswere performed by adding 2 lM pyrene actin to a protein mixtureof 22.4 nM Arp2/3 complex, 500 nM Cdc42 GTPcS, and 100 nM N-WASP. For 20 min the fluorescence change was measured at386 nm with excitation at 366 nm using a luminescence spectrom-eter (LS50B, Perkin Elmer Life Sciences).
Actin tail length assay. Exponential phase bacteria were centri-fuged at 1000 rpm for 15 min onto a semi-confluent monolayer ofcells seeded on a 20 � 20 mm glass coverslip at an MOI of 100. Cellswere incubated for 30 min at 37 �C with 5% CO2. Cells were washedthree times with PBS, and then incubated for 2 h in media containing50 lg/ml gentamicin. Cells were washed three times with PBS, thenfixed with 3.7% paraformaldehyde in PBS for 10 min and stored in 1%BSA in PBS. Following staining, images of 10 different cells per con-dition per experiment were taken and tail lengths were measuredusing ImageJ software (Open Source Software). Statistics were per-formed using the Mann–Whitney Test.
Immunofluorescence microscopy. Fixed cells were permeabilizedwith 0.2% Triton X-100 in PBS for 5 min and washed three times with1% BSA in PBS. N-WASP was stained with mouseaFLAG antibody (SIG-MA catalog #F1804) at 1:1000 for 1 h, followed by either FITC-conju-gated or Cy3-conjugated donkey anti-mouse antibody at 1:200 for1 h. F-actin was stained with rhodamine-labeled or FITC-conjugatedphalloidin (1 lg/ml in 1% BSA in PBS; Molecular Probes) for 1 h.DNA was stained with DAPI (300 nM; Molecular Probes) for 10 min.
Intracellular motility assay. Exponential phase bacteria express-ing GFP were centrifuged at 1000 rpm for 15 min onto a semi-con-fluent monolayer of cells seeded on a glass-bottom 35 mm dish atan MOI of 100. Cells were incubated for 30 min at 37 �C with 5%CO2. Cells were washed three times with PBS, and incubated inmedia containing 50 lg/ml gentamicin. After 1 h, the cells wereimaged on a microscope (IX-70; Olympus Optical) with an auto-mated stage using a 100� objective. Images were taken in boththe phase and GFP channels every 10 s for 15 min. Using Slidebooksoftware (Intelligent Imaging Innovations, Denver, CO), the GFPchannel was analyzed to track bacterial movement. Only bacteriamoving in at least 10 consecutive frames were counted.
Shigella invasion. Exponential phase bacteria were centrifuged at1000 rpm for 15 min onto a semi-confluent monolayer of cellsseeded on a 20 � 20 mm glass coverslip at an MOI of 100. Cellswere incubated for 60 min at 37 �C with 5% CO2. Cells were washedthree times with PBS, then fixed with 3.7% paraformaldehyde inPBS for 10 min and stored in 1% BSA in PBS. Cells were stained asdescribed above, and counted microscopically (AX-70; OlympusOptical). Invasion rate is calculated as the number of cells contain-ing internalized bacteria divided by the total number of cellscounted multiplied by 100. At least one hundred cells from two dif-ferent coverslips were counted for each experiment.
Results
Point mutations in N-WASP predicted to alter either structuralconformation or phospho-regulation result in constitutive activation
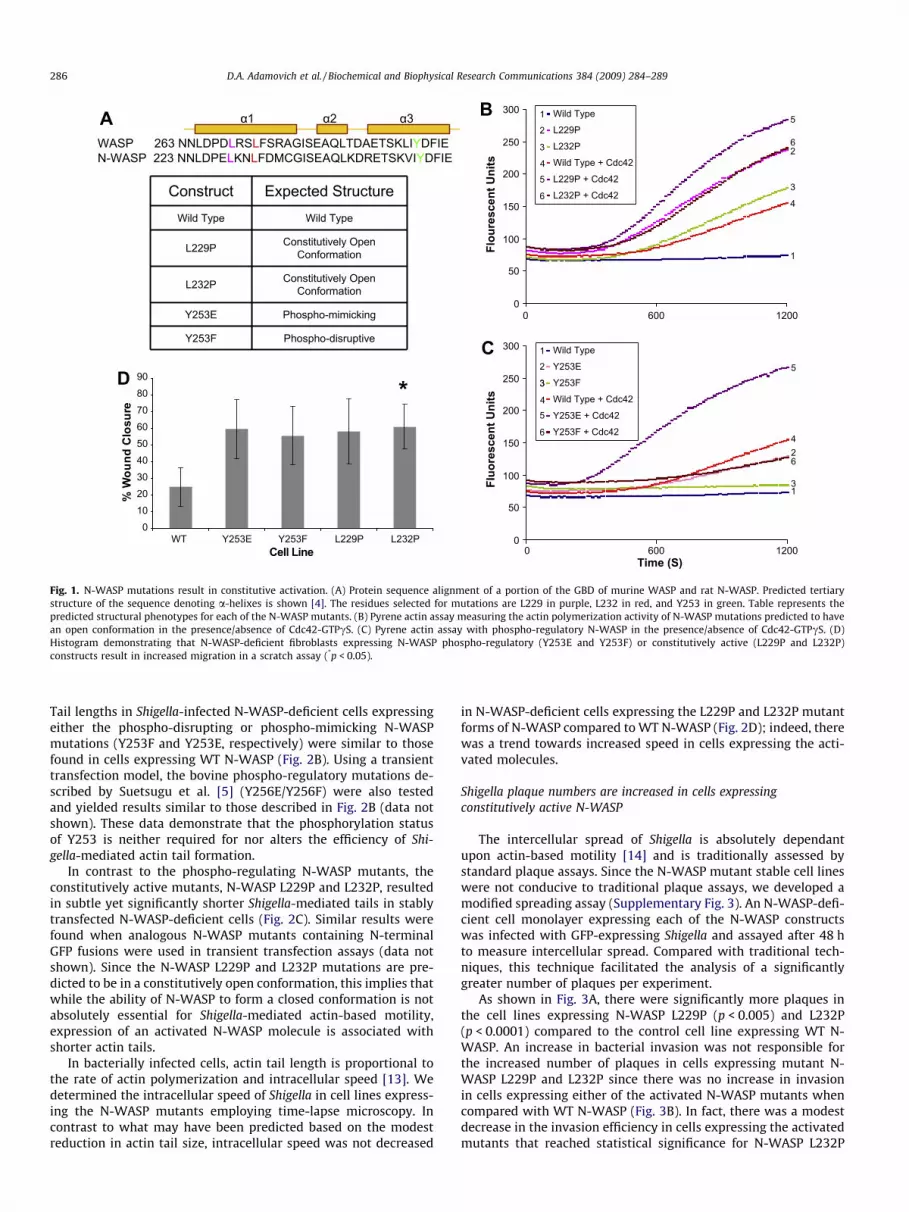
A series of point mutations were generated that were predictedto alter either N-WASP structural conformation or phospho-regula-
tion (Fig. 1A). A leucine to proline mutation in the homolog humanWASP (L270P) leads to a change in the structure of WASP resultingin its open conformation and constitutive activation [9]. We pre-dicted that a homologous mutation in N-WASP (L232P) would alsolead to a constitutively open and active conformation given theconserved location in the a-helix (Fig. 1A). Similarly, we predictedthat a L229P mutation in N-WASP, located �1 turn away from L232in the critical a-helix, would lead to a constitutively open and ac-tive conformation of N-WASP (Fig. 1A). Point mutations in N-WASPthat alter the phospho-regulation of N-WASP and cytoskeletal pro-cesses have previously been described by Suetsugu et al. [5]. Wegenerated both the phospho-mimicking tyrosine to glutamic acid(Y253E) and the phospho-disruptive tyrosine to phenylalanine(Y253F) mutations in N-WASP (Fig. 1A).
To assess whether the N-WASP L229P and L232P mutations al-ter baseline actin polymerization activity, we employed a pyreneactin assay. Wild type (WT) N-WASP when combined with Arp2/3 and pyrene actin does not lead to significant actin polymerization(Fig. 1B) [10]. However, when activated Cdc42 is added to N-WASP,Arp2/3 and pyrene actin, a significant increase in actin polymeriza-tion was observed (Fig. 1B). N-WASP L229P and L232P, in the ab-sence of activated Cdc42, polymerizes actin at rates similar orgreater than polymerization rates achieved with WT N-WASP inthe presence of activated Cdc42 (Fig. 1B). Moreover, this constitu-tive activation of N-WASP L229P and N-WASP L232P can be furtherincreased by the addition of activated Cdc42 (Fig. 1B). Similar re-sults demonstrating constitutive activation of L229P and L232Pwere also obtained employing a modified bead assay (Supplemen-tary Fig. 1) [11].
The pyrene actin assay was also employed to assess the activityof the N-WASP constructs containing phospho-regulatory muta-tions. Similar to results obtained with an in vitro bead assay usingWASP Y291E [12], the homologous N-WASP Y253E exhibited anenhanced basal level of actin polymerization compared to WT N-WASP (Fig. 1C). This enhanced basal level of activation of N-WASPY253E was also sensitive to Cdc42 stimulation (Fig. 1C). In contrast,the phospho-disruptive N-WASP Y253F does not exhibit an en-hanced level of actin polymerization compared to WT N-WASP,but was sensitive to stimulation by activated Cdc42 (Fig. 1C).
N-WASP-deficient cell lines expressing N-WASP activation mutationshave enhanced migration
To assess, in vivo, the role of each of the N-WASP structural con-formation and phospho-regulation mutants on a processes that re-quires dynamic modulation of the actin cytoskeleton (e.g., cellmotility), we generated stable cell lines where each mutant con-struct, or WT N-WASP, was expressed in N-WASP-deficient cells.The effect of each of the constructs on cell motility was determinedemploying a standard scratch assay. Cell lines expressing each ofthe structural conformation and phospho-regulation mutantsclosed wounds faster than the control cell line expressing WT N-WASP (Supplementary Fig. 2 and Fig. 1D).
Shigella-mediated actin tail formation does not absolutely requireregulation of N-WASP structural conformation or phospho-regulation
To assess Shigella’s ability to utilize each of the mutant N-WASPconstructs for actin-based motility, Shigella infections were per-formed in N-WASP-deficient cell lines that stably expressed eachof the N-WASP mutants. Each cell line permitted bacterial invasionand supported Shigella-mediated actin tail formation, with N-WASP localizing appropriately to a bacterial pole (Fig. 2A).
We measured the length of actin tails in Shigella-infected cellsexpressing each of the various N-WASP mutants to determinethe efficiency by which Shigella utilized the N-WASP constructs.
Construct Expected StructureWild Type Wild Type
L229P Constitutively Open Conformation
L232P Constitutively Open Conformation
Y253E Phospho-mimicking
Y253F Phospho-disruptive
0
50
100
150
200
250
300
0 600 1200
0 600 1200
Flou
resc
ent U
nits
Wild Type
L229P
L232P
Wild Type + Cdc42
L229P + Cdc42
L232P + Cdc42
0
50
100
150
200
250
300
Time (S)
Fluo
resc
ent U
nits
Wild Type
Y253E
Y253F
Wild Type + Cdc42
Y253E + Cdc42
Y253F + Cdc42
WASP 263 NNLDPDLRSLFSRAGISEAQLTDAETSKLIYDFIEN-WASP 223 NNLDPELKNLFDMCGISEAQLKDRETSKVIYDFIE
α1 α2 α3
0102030405060708090
WT Y253E Y253F L229P L232PCell Line
% W
ound
Clo
sure
*
1
1
2
3
45
6
35
3
4
62
3
4
6
1
2
5
1
26
5
4
3
Fig. 1. N-WASP mutations result in constitutive activation. (A) Protein sequence alignment of a portion of the GBD of murine WASP and rat N-WASP. Predicted tertiarystructure of the sequence denoting a-helixes is shown [4]. The residues selected for mutations are L229 in purple, L232 in red, and Y253 in green. Table represents thepredicted structural phenotypes for each of the N-WASP mutants. (B) Pyrene actin assay measuring the actin polymerization activity of N-WASP mutations predicted to havean open conformation in the presence/absence of Cdc42-GTPcS. (C) Pyrene actin assay with phospho-regulatory N-WASP in the presence/absence of Cdc42-GTPcS. (D)Histogram demonstrating that N-WASP-deficient fibroblasts expressing N-WASP phospho-regulatory (Y253E and Y253F) or constitutively active (L229P and L232P)constructs result in increased migration in a scratch assay (*p < 0.05).
286 D.A. Adamovich et al. / Biochemical and Biophysical Research Communications 384 (2009) 284–289
Tail lengths in Shigella-infected N-WASP-deficient cells expressingeither the phospho-disrupting or phospho-mimicking N-WASPmutations (Y253F and Y253E, respectively) were similar to thosefound in cells expressing WT N-WASP (Fig. 2B). Using a transienttransfection model, the bovine phospho-regulatory mutations de-scribed by Suetsugu et al. [5] (Y256E/Y256F) were also testedand yielded results similar to those described in Fig. 2B (data notshown). These data demonstrate that the phosphorylation statusof Y253 is neither required for nor alters the efficiency of Shi-gella-mediated actin tail formation.
In contrast to the phospho-regulating N-WASP mutants, theconstitutively active mutants, N-WASP L229P and L232P, resultedin subtle yet significantly shorter Shigella-mediated tails in stablytransfected N-WASP-deficient cells (Fig. 2C). Similar results werefound when analogous N-WASP mutants containing N-terminalGFP fusions were used in transient transfection assays (data notshown). Since the N-WASP L229P and L232P mutations are pre-dicted to be in a constitutively open conformation, this implies thatwhile the ability of N-WASP to form a closed conformation is notabsolutely essential for Shigella-mediated actin-based motility,expression of an activated N-WASP molecule is associated withshorter actin tails.
In bacterially infected cells, actin tail length is proportional tothe rate of actin polymerization and intracellular speed [13]. Wedetermined the intracellular speed of Shigella in cell lines express-ing the N-WASP mutants employing time-lapse microscopy. Incontrast to what may have been predicted based on the modestreduction in actin tail size, intracellular speed was not decreased
in N-WASP-deficient cells expressing the L229P and L232P mutantforms of N-WASP compared to WT N-WASP (Fig. 2D); indeed, therewas a trend towards increased speed in cells expressing the acti-vated molecules.
Shigella plaque numbers are increased in cells expressingconstitutively active N-WASP
The intercellular spread of Shigella is absolutely dependantupon actin-based motility [14] and is traditionally assessed bystandard plaque assays. Since the N-WASP mutant stable cell lineswere not conducive to traditional plaque assays, we developed amodified spreading assay (Supplementary Fig. 3). An N-WASP-defi-cient cell monolayer expressing each of the N-WASP constructswas infected with GFP-expressing Shigella and assayed after 48 hto measure intercellular spread. Compared with traditional tech-niques, this technique facilitated the analysis of a significantlygreater number of plaques per experiment.
As shown in Fig. 3A, there were significantly more plaques inthe cell lines expressing N-WASP L229P (p < 0.005) and L232P(p < 0.0001) compared to the control cell line expressing WT N-WASP. An increase in bacterial invasion was not responsible forthe increased number of plaques in cells expressing mutant N-WASP L229P and L232P since there was no increase in invasionin cells expressing either of the activated N-WASP mutants whencompared with WT N-WASP (Fig. 3B). In fact, there was a modestdecrease in the invasion efficiency in cells expressing the activatedmutants that reached statistical significance for N-WASP L232P

Wild
TypeL229P
Y253E
L232PNo
N-WASP
Y253F
A
B C D
0
WT
Y253
E
Y253
F
WT
L229
P
L232
P
WT
WT
L229
P
L232
P
1
Rel
ativ
e Ta
ll Le
ngth
2
3
0
1
Rel
ativ
e Ta
ll Le
ngth
2
3
0
1
Avg
spe
ed (u
m/m
in)
2
3
Fig. 2. N-WASP regulatory mutants localize appropriately and support Shigella-mediated actin-based motility. (A) Stable cell lines expressing each of the N-WASP constructswere infected with Shigella. Cells were stained for DNA (blue), N-WASP (green), and actin (red). Insets show polar N-WASP localization (arrow heads) and actin tail formation(arrows). (B) Tails formed in cells expressing N-WASP Y253E and Y253F are similar in length to those expressed in cells expressing WT N-WASP. (C) Tails formed in cellsexpressing N-WASP L229P and L232P are shorter in length than those expressed in cells expressing WT N-WASP. *p < 0.05 (Mann–Whitney test). (D) Time lapse microscopywas used to quantify the velocity of Shigella undergoing actin-based motility. Average speed of Shigella in cells expressing WT N-WASP, L229P, or L232P is shown.
D.A. Adamovich et al. / Biochemical and Biophysical Research Communications 384 (2009) 284–289 287
(Fig. 3B; p = 0.03). However, despite the increase in the number ofplaques, there was no difference in the diameter of plaques formedin the cell lines expressing L229P and L232P compared to the cellline expressing WT N-WASP (Fig. 3C), indicating that the rate ofintercellular spread is not altered by expression of activated N-WASP.
Discussion
Intracellular pathogens exploit diverse mechanisms to modu-late the host actin cytoskeleton during infection [1]. Shigella IcsAbinds and activates host-encoded N-WASP resulting in activationof Arp-2/3 dependent actin assembly [15,16]. The processes thatregulate N-WASP activation during Shigella infection are largelyunknown. We have examined whether the actin-based motilityof Shigella requires active modulation of the structure and phos-phorylation of N-WASP during infection.
We demonstrate that N-WASP L232P mutation, homologous tothe WASP L270P mutation implicated in X-linked neutropenia [9],results in the constitutive activation of N-WASP. In addition, a pro-line mutation in a neighboring leucine residue can also lead to con-stitutive N-WASP activation. These studies reproduce previouslypublished results demonstrating that the N-WASP Y253E mutationhas an enhanced intrinsic actin polymerization activity compared
to WT N-WASP, and that the N-WASP Y253F mutation behavessimilar to WT N-WASP in the pyrene actin assay [5]. Activationby the structural conformation (L229P and L232P) and phospho-mimicking (Y253E) N-WASP mutations could be further enhancedin vitro by the addition of Cdc42 – suggesting that these mutationsmay not result in complete disruption of auto-inhibition in vivo.
We have demonstrated that Shigella-mediated actin assemblyoccurs independent of N-WASP phosphorylation at Y253 – a sitepreviously shown to be required for modulation of the actin cyto-skeleton during neurite extension [5]. Moreover, the efficiency ofactin tail formation does not seem to require this form of phos-pho-regulation since tail lengths were similar in cells expressingWT N-WASP or either the phospho-mimicking (Y253E) or phos-pho-disruptive (Y253F) N-WASP. This work contrasts with anotherstudy that showed that phosphorylation of Y253 in N-WASP regu-lates the length of Shigella-induced actin tails [8]. Shorter actin tailswere observed upon Shigella infection when a phospho-disruptiveN-WASP Y253F mutant was expressed in N-WASP-deficient cellsemploying transient transfection techniques [8]. In these studies,inhibition of tyrosine phosphorylation using the Abl/Arg kinaseinhibitor Gleevac also inhibited tail length [8]. We observed no af-fect of the phospho-disruptive mutation on Shigella-mediated actinassembly employing either stable cell lines (Fig. 3B) or in transienttransfection assays (data not shown). Moreover, Gleevac had no af-

NWASP KO NWASP L229PWild Type NWASP
0 242 4600
Wt
L229
P
L232
P
Wt
L229
P
L232
PWt
L229
P
L232
P
Rel
ativ
e In
vasi
on R
ate
Rel
ativ
e Pl
aque
Dia
met
er
Rel
ativ
e Pl
aque
Num
bers
0.5
1.5
2.5
2
1
0
0.2
0.4
0.6
0.8
1.2
1
0
0.2
0.4
0.6
0.8
1.2
1.4
1.6
1
* *
*
Fig. 3. Markedly increased plaque numbers, but not invasion rate or intercellular spread, in cells expressing constitutively active N-WASP. (A) Increased number of plaquesformed in cell lines expressing constitutively active N-WASP L229 and L2332. Representative monolayers imaged 48 h after infection with GFP-expressing Shigella. Greensignal represents Shigella plaques (inset indicates number of plaques in monolayer). Histogram on right is quantification of three independent experiments. (B) Shigellainvasion rates are compared in stable cell lines expressing WT, L229 or L2332 N-WASP constructs. Results shown are average of three independent experiments (*p < 0.05). (C)The average size of plaques formed in each cell line expressing mutant N-WASP constructs was not significantly different from cell lines expressing WT N-WASP. Combinedresults of three individual experiments are shown. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
288 D.A. Adamovich et al. / Biochemical and Biophysical Research Communications 384 (2009) 284–289
fect on Shigella-mediated actin tail formation in our studies (datanot shown). Although we have no obvious explanation for the dif-ferences in results obtained in each of these studies – the N-WASP-deficient cell lines in each of the studies were obtained from differ-ent mouse lines [17,18].
Our data indicate that a constitutively active N-WASP mutantprotein can readily support Shigella-mediated actin assemblyand movement. In contrast to bacterial speed, the size of Shigellatails was modestly reduced in cells expressing constitutivelyactive N-WASP L299P or L232P as compared to cells expressingWT N-WASP. Leung et al. did not observe a difference inShigella-mediated actin tails in N-WASP-deficient cells expressinghuman N-WASP L235P (homologous to N-WASP L232P) comparedto cells expressing WT human N-WASP [19]. Given the shorter taillengths without concomitant reductions in bacterial speed associ-ated with these mutations, our data is consistent with a model bywhich activated N-WASP results in an increase in actin polymer-ization yet may have a more pronounced direct or indirect roleon actin depolymerization [20]. Our data showing that N-WASPL229P or L232 led to increased actin polymerization and increasedmigration of cells in vitro gives support to this model.
The increased number of plaques formed in the cell monolayersexpressing N-WASP L229P and L232P as compared to thoseexpressing WT N-WASP suggests that a change in the activationof N-WASP through modulation of its resting conformation altersShigella pathogenesis. An altered rate of bacterial invasion couldnot explain the differences in plaque numbers since the rate of bac-terial invasion was similar among the different cell lines (Fig. 3B).Moreover, mechanisms that enhance the rate of intercellularspreading could not explain the observation of increased plaquenumbers because there was no difference in the diameter of theplaques formed in each of the cell lines (Fig. 3C). An increase in pla-
que numbers may reflect an alteration in the rate by which Shigellais cleared following invasion from the extracellular environmentwhen an activated N-WASP is present in host cells. In this regard,Shigella uptake into autophagosomes has been shown to be depen-dent upon the presence of IcsA [21] and the process of autophagy isdependent upon Arp2/3 activation [22].
Acknowledgment
This work was supported by NIH 5R01 AI052354 and 5P01HL59561 to S.B.S.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.bbrc.2009.04.050.
References
[1] E. Gouin, M.D. Welch, P. Cossart, Actin-based motility of intracellularpathogens, Curr. Opin. Microbiol. 8 (2005) 35–45.
[2] T. Takenawa, S. Suetsugu, The WASP–WAVE protein network: connecting themembrane to the cytoskeleton, Nat. Rev. Mol. Cell Biol. 8 (2007) 37–48.
[3] H. Miki, T. Takenawa, Direct binding of the verprolin-homology domain in N-WASP to actin is essential for cytoskeletal reorganization, Biochem. Biophys.Res. Commun. 243 (1998) 73–78.
[4] A.S. Kim, L.T. Kakalis, N. Abdul-Manan, G.A. Liu, M.K. Rosen, Autoinhibition andactivation mechanisms of the Wiskott-Aldrich syndrome protein, Nature 404(2000) 151–158.
[5] S. Suetsugu, M. Hattori, H. Miki, T. Tezuka, T. Yamamoto, K. Mikoshiba, T.Takenawa, Sustained activation of N-WASP through phosphorylation isessential for neurite extension, Dev. Cell 3 (2002) 645–658.
[6] T. Shibata, F. Takeshima, F. Chen, F.W. Alt, S.B. Snapper, Cdc42 facilitates invasionbut not the actin-based motility of Shigella, Curr. Biol. 12 (2002) 341–345.

D.A. Adamovich et al. / Biochemical and Biophysical Research Communications 384 (2009) 284–289 289
[7] F. Frischknecht, V. Moreau, S. Rottger, S. Gonfloni, I. Reckmann, G. Superti-Furga, M. Way, Actin-based motility of vaccinia virus mimics receptor tyrosinekinase signalling, Nature 401 (1999) 926–929.
[8] E.A. Burton, T.N. Oliver, A.M. Pendergast, Abl kinases regulate actin comet tailelongation via an N-WASP-dependent pathway, Mol. Cell. Biol. 25 (2005)8834–8843.
[9] K. Devriendt, A.S. Kim, G. Mathijs, S.G. Frints, M. Schwartz, J.J. Van Den Oord,G.E. Verhoef, M.A. Boogaerts, J.P. Fryns, D. You, M.K. Rosen, P. Vandenberghe,Constitutively activating mutation in WASP causes X-linked severe congenitalneutropenia, Nat. Genet. 27 (2001) 313–317.
[10] H.Y. Ho, R. Rohatgi, A.M. Lebensohn, M.W. Kirschner, In vitro reconstitution ofcdc42-mediated actin assembly using purified components, Methods Enzymol.406 (2006) 174–190.
[11] P.J. Ancliff, M.P. Blundell, G.O. Cory, Y. Calle, A. Worth, H. Kempski, S. Burns,G.E. Jones, J. Sinclair, C. Kinnon, I.M. Hann, R.E. Gale, D.C. Linch, A.J. Thrasher,Two novel activating mutations in the Wiskott-Aldrich syndrome proteinresult in congenital neutropenia, Blood 108 (2006) 2182–2189.
[12] G.O. Cory, R. Garg, R. Cramer, A.J. Ridley, Phosphorylation of tyrosine 291enhances the ability of WASp to stimulate actin polymerization andfilopodium formation. Wiskott-Aldrich Syndrome protein, J. Biol. Chem. 277(2002) 45115–45121.
[13] J.A. Theriot, T.J. Mitchison, L.G. Tilney, D.A. Portnoy, The rate of actin-basedmotility of intracellular Listeria monocytogenes equals the rate of actinpolymerization, Nature 357 (1992) 257–260.
[14] S. Makino, C. Sasakawa, K. Kamata, T. Kurata, M. Yoshikawa, A geneticdeterminant required for continuous reinfection of adjacent cells on largeplasmid in S. flexneri 2a, Cell 46 (1986) 551–555.
[15] C. Egile, T.P. Loisel, V. Laurent, R. Li, D. Pantaloni, P.J. Sansonetti, M.F. Carlier,Activation of the CDC42 effector N-WASP by the Shigella flexneri IcsA protein
promotes actin nucleation by Arp2/3 complex and bacterial actin-basedmotility, J. Cell Biol. 146 (1999) 1319–1332.
[16] T. Suzuki, H. Miki, T. Takenawa, C. Sasakawa, Neural Wiskott-Aldrichsyndrome protein is implicated in the actin-based motility of Shigellaflexneri, EMBO J. 17 (1998) 2767–2776.
[17] S.B. Snapper, F. Takeshima, I. Anton, C.H. Liu, S.M. Thomas, D. Nguyen, D.Dudley, H. Fraser, D. Purich, M. Lopez-Ilasaca, C. Klein, L. Davidson, R. Bronson,R.C. Mulligan, F. Southwick, R. Geha, M.B. Goldberg, F.S. Rosen, J.H. Hartwig,F.W. Alt, N-WASP deficiency reveals distinct pathways for cell surfaceprojections and microbial actin-based motility, Nat. Cell Biol. 3 (2001) 897–904.
[18] S. Lommel, S. Benesch, K. Rottner, T. Franz, J. Wehland, R. Kuhn, Actin pedestalformation by enteropathogenic Escherichia coli and intracellular motility ofShigella flexneri are abolished in N-WASP-defective cells, EMBO Rep. 2 (2001)850–857.
[19] Y. Leung, S. Ally, M.B. Goldberg, Bacterial actin assembly requires toca-1 torelieve N-wasp autoinhibition, Cell Host Microbe 3 (2008) 39–47.
[20] M.F. Carlier, V. Laurent, J. Santolini, R. Melki, D. Didry, G.X. Xia, Y. Hong, N.H.Chua, D. Pantaloni, Actin depolymerizing factor (ADF/cofilin) enhances the rateof filament turnover: implication in actin-based motility, J. Cell Biol. 136(1997) 1307–1322.
[21] M. Ogawa, T. Yoshimori, T. Suzuki, H. Sagara, N. Mizushima, C. Sasakawa,Escape of intracellular Shigella from autophagy, Science 307 (2005) 727–731.
[22] I. Monastyrska, C. He, J. Geng, A.D. Hoppe, Z. Li, D.J. Klionsky, Arp2links autophagic machinery with the actin cytoskeleton, Mol. Biol. Cell(2008).
[23] L. Blanchoin, K.J. Amann, H.N. Higgs, J.B. Marchand, D.A. Kaiser, T.D. Pollard,Direct observation of dendritic actin filament networks nucleated by Arp2/3complex and WASP/Scar proteins, Nature 404 (2000) 1007–1011.