acetylated nucleosome assembly on telomeric dnas
TRANSCRIPT

Biophysical Chemistry 104(2003) 381–392
0301-4622/03/$ - see front matter� 2003 Elsevier Science B.V. All rights reserved.doi:10.1016/S0301-4622Ž03.00028-0
Acetylated nucleosome assembly on telomeric DNAs
Stefano Cacchione , Jose Luis Rodrıguez , Rosella Mechelli , Luis Franco , Maria Savino *a b a b a,´ ´
Dipartimento di Genetica e Biologia Molecolare, Fondazione Istituto Pasteur-Fondazione Cenci Bolognetti,a
Universita di Roma ‘La Sapienza’, Piazzale A. Moro 5, 00185 Roma, Italy`Departamento de Bioquımica y Biologıa Molecular, Universidad de Valencia, 46100 Burjassot, Valencia, Spainb ´ ´ ` `
Received 31 October 2002; received in revised form 20 January 2003; accepted 20 January 2003
Abstract
The role of histone N-terminal domains on the thermodynamic stability of nucleosomes assembled on severaldifferent telomeric DNAs as well as on ‘average’ sequence DNA and on strong nucleosome positioning sequences,has been studied by competitive reconstitution. We find that histone tails hyperacetylation favors nucleosomeformation, in a similar extent for all the examined sequences. On the contrary, removal of histone terminal domainsby selective trypsinization causes a decrease of nucleosome stability which is smaller for telomeres compared to theother sequences examined, suggesting that telomeric sequences have only minor interactions with histone tails.Micrococcal nuclease kinetics shows enhanced accessibility of acetylated nucleosomes formed both on telomeric and‘average’ sequence DNAs. These results suggest a more complex role for histone acetylation than the decrease ofelectrostatic interactions between DNA and histones.� 2003 Elsevier Science B.V. All rights reserved.
Keywords: Telomeres; Nucleosome; Histone acetylation; Thermodynamic stability; Nucleosome positioning
1. Introduction
Telomeres, the specialized structures located atthe ends of eukaryotic chromosomes, generallyconsist of tandemly repeated units 5–8 bp long,rich in GT on the 39 ending strandsw1,2x. In highereukaryotes, telomeres are organized in regularnucleosomal arrays with unusually short repeatlength w3–6x. In yeast telomeres, nucleosomesseem to be absentw7x, whereas inTetrahymenatermophila a nucleosome organization is presentin 3–10% of telomeres, suggesting that the differ-
*Corresponding author. Tel.:q39-06-49912238; fax:q39-06-4440812.
E-mail address: [email protected](M. Savino).
ence in telomeric chromatin between higher andlower eukaryotes could be quantitative, rather thanqualitativew8x.We previously found that several telomeric
DNAs from different organisms form the lessstable nucleosomes among the DNA sequences sofar studiedw9–11x. The values of free energy ofnucleosome formation of telomeric sequences withrespect to an ‘average’ sequence DNA, obtainedby the competitive reconstitution methodw12x,range fromq1.80 toq0.40 kcalymol of nucleo-some. These values are in fairly good agreementwith those obtained adopting a theoretical methodrecently developed by De Santis and co-workersw11,13–15x, which derives the free energy of
382 S. Cacchione et al. / Biophysical Chemistry 104 (2003) 381–392
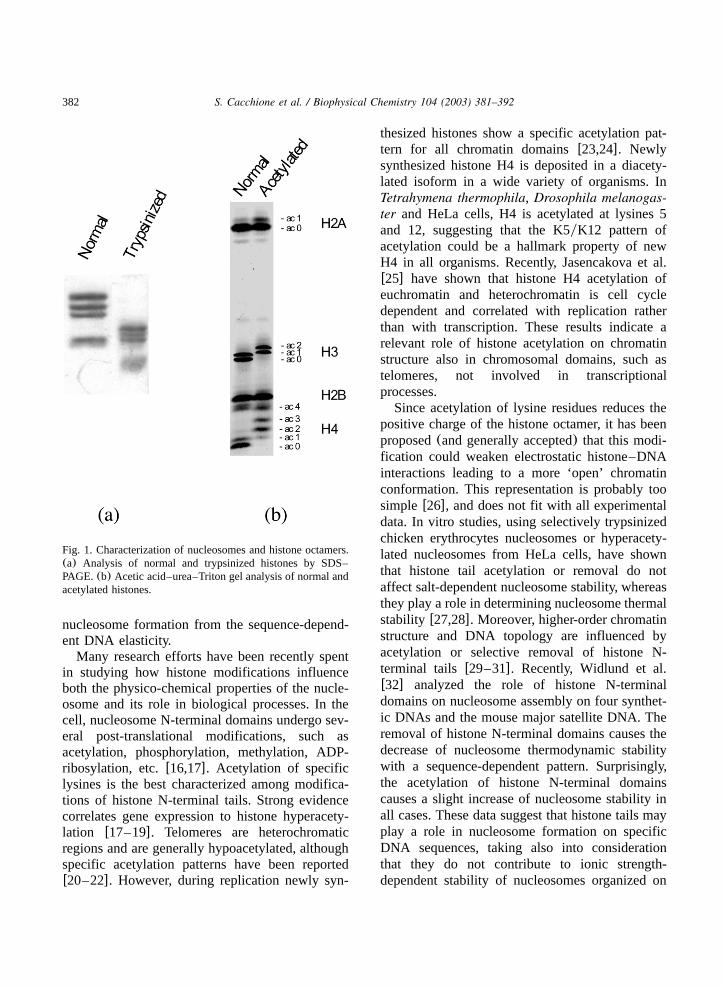
Fig. 1. Characterization of nucleosomes and histone octamers.(a) Analysis of normal and trypsinized histones by SDS–PAGE.(b) Acetic acid–urea–Triton gel analysis of normal andacetylated histones.
nucleosome formation from the sequence-depend-ent DNA elasticity.Many research efforts have been recently spent
in studying how histone modifications influenceboth the physico-chemical properties of the nucle-osome and its role in biological processes. In thecell, nucleosome N-terminal domains undergo sev-eral post-translational modifications, such asacetylation, phosphorylation, methylation, ADP-ribosylation, etc.w16,17x. Acetylation of specificlysines is the best characterized among modifica-tions of histone N-terminal tails. Strong evidencecorrelates gene expression to histone hyperacety-lation w17–19x. Telomeres are heterochromaticregions and are generally hypoacetylated, althoughspecific acetylation patterns have been reportedw20–22x. However, during replication newly syn-
thesized histones show a specific acetylation pat-tern for all chromatin domainsw23,24x. Newlysynthesized histone H4 is deposited in a diacety-lated isoform in a wide variety of organisms. InTetrahymena thermophila, Drosophila melanogas-ter and HeLa cells, H4 is acetylated at lysines 5and 12, suggesting that the K5yK12 pattern ofacetylation could be a hallmark property of newH4 in all organisms. Recently, Jasencakova et al.w25x have shown that histone H4 acetylation ofeuchromatin and heterochromatin is cell cycledependent and correlated with replication ratherthan with transcription. These results indicate arelevant role of histone acetylation on chromatinstructure also in chromosomal domains, such astelomeres, not involved in transcriptionalprocesses.Since acetylation of lysine residues reduces the
positive charge of the histone octamer, it has beenproposed(and generally accepted) that this modi-fication could weaken electrostatic histone–DNAinteractions leading to a more ‘open’ chromatinconformation. This representation is probably toosimple w26x, and does not fit with all experimentaldata. In vitro studies, using selectively trypsinizedchicken erythrocytes nucleosomes or hyperacety-lated nucleosomes from HeLa cells, have shownthat histone tail acetylation or removal do notaffect salt-dependent nucleosome stability, whereasthey play a role in determining nucleosome thermalstability w27,28x. Moreover, higher-order chromatinstructure and DNA topology are influenced byacetylation or selective removal of histone N-terminal tails w29–31x. Recently, Widlund et al.w32x analyzed the role of histone N-terminaldomains on nucleosome assembly on four synthet-ic DNAs and the mouse major satellite DNA. Theremoval of histone N-terminal domains causes thedecrease of nucleosome thermodynamic stabilitywith a sequence-dependent pattern. Surprisingly,the acetylation of histone N-terminal domainscauses a slight increase of nucleosome stability inall cases. These data suggest that histone tails mayplay a role in nucleosome formation on specificDNA sequences, taking also into considerationthat they do not contribute to ionic strength-dependent stability of nucleosomes organized on

383S. Cacchione et al. / Biophysical Chemistry 104 (2003) 381–392
heterogeneous sequence genomic DNA(bulknucleosomes) w27,28x.In this study we analyze the influence of histone
N-terminal domains on thermodynamic stability oftelomeric nucleosomes compared to nucleosomesformed on DNA sequences with high affinity forthe histone octamer such as 5S rDNA and TGpentamer w12x, and on the sequence TAND-1,which has the same free energy of nucleosomeformation as ‘average’ sequence DNAw11x.Telomeric sequences are straight, due to the regularrepetition of short nucleotide sequences out ofphase with the B-DNA helical period. They foldaround the histone octamer in a sequence-depend-ent way with lower affinity with respect to bulknucleosomes. Therefore, they represent an interest-ing structurally homogeneous set of sequences tostudy the role of histone N-terminal domains onnucleosome thermodynamic stability. We find thathyperacetylation favors nucleosome formation, toan extent that is substantially similar in all exam-ined sequences. On the contrary, the enzymaticremoval of the histone tails causes a decrease ofnucleosome stability, which is smaller in the caseof telomeres. In apparent contrast with the resultson thermodynamic stability, micrococcal nuclease(MNase) digestion studies show an increasedaccessibility of the enzyme to hyperacetylatednucleosomes.
2. Materials and methods
2.1. DNA and histone octamer preparation
TAND-1 was a gift from A. Travers. Thesequence TG was a gift from D. Crothers. Esa1pypRSET plasmid was kindly provided by C.D.Allis. Telomeric DNAs and TAND-1 were pre-pared as previously describedw8,10x. Kluy-veromyces lactis telomeric DNA(GGTATGTGGTGTACGGATTTGATTA) was6
multimerized as previously describedw9x andcloned in theSmaI site of pBluescript II KS(y).5S rDNAs were prepared by partially digesting theplasmid pPOL 208–12w33x containing 12 copiesof 5S rRNA gene fromLytechinus variegatus (agift from R. Sendra) with AvaI. Fragments corre-sponding to the monomer and the dimer bands of
5S rDNA were gel extracted and cloned in theEcoRV site of pSTBlue1(Novagen) after fillingin protruding ends. H1-free polynucleosomal par-ticles were prepared from chicken erythrocytes aspreviously describedw9x. Purified histone octamerswere obtained from polynucleosomes by hydrox-yapatite chromatography. Trypsinized nucleosomeswere obtained by digesting chicken erythrocytesnucleosomes with immobilized trypsin accordingto Ausio et al.w28x. The extent of trypsin digestionwas assessed on standard SDS–18% polyacryla-mide gel.Recombinant enzyme rEsa1p was expressed in
Escherichia coli BL21-DE3 cells and purified byaffinity chromatography on Ni –agarose as2q
described by Smith et al.w34x. A stoichiometricquantity (500 mg) of chicken erythrocyte corehistones was mixed in a final reaction volume of2.5 ml, with approximately10mg of recombinantenzyme, in the presence of 100 mM acetyl-CoA(or absence in case of the controls), and proteasesinhibitors w1 mM PMSF and 10mM trans-epoxy-succinyl-L-leucylamido-(4-guanidino)butanex. Themixture was incubated at 258C for 2 h and thereaction was stopped by 0.25 M HCl. After cen-trifugation at 10 000=g for 10 min, histones wererecovered in the supernatant and the core histonesoctamers were reconstituted by consecutive dialy-ses following the procedure of Marvin et al.w35x.The extent of histone acetylation was checked byacetic acid–urea–Triton polyacrylamide gel elec-trophoresisw36x (Fig. 1).
2.2. Competitive reconstitutions
When normal or selectively trypsinized polyn-ucleosomes were used as the histone donor, recon-stitutions were carried out by the salt dilutionmethodw12x, as described by Filesi et al.w11x. Forreconstitutions starting from histone octamers, 2mg of purified histone octamer(hypo- or hypera-cetylated) was mixed with various amounts ofsonicated calf thymus DNA and 30 ng of radio-labeled DNA in 10ml of dilution buffer w10 mMTris–HCl (pH 8), 1 mM EDTA, 0.1% Nonidet-P40, 0.1 mgyml BSAx plus 1.0 M NaCl, or in 15ml of dilution buffer plus 0.65 M NaCl. Samplesat 1.0 M NaCl were incubated for 1 h at 258C,
384 S. Cacchione et al. / Biophysical Chemistry 104 (2003) 381–392
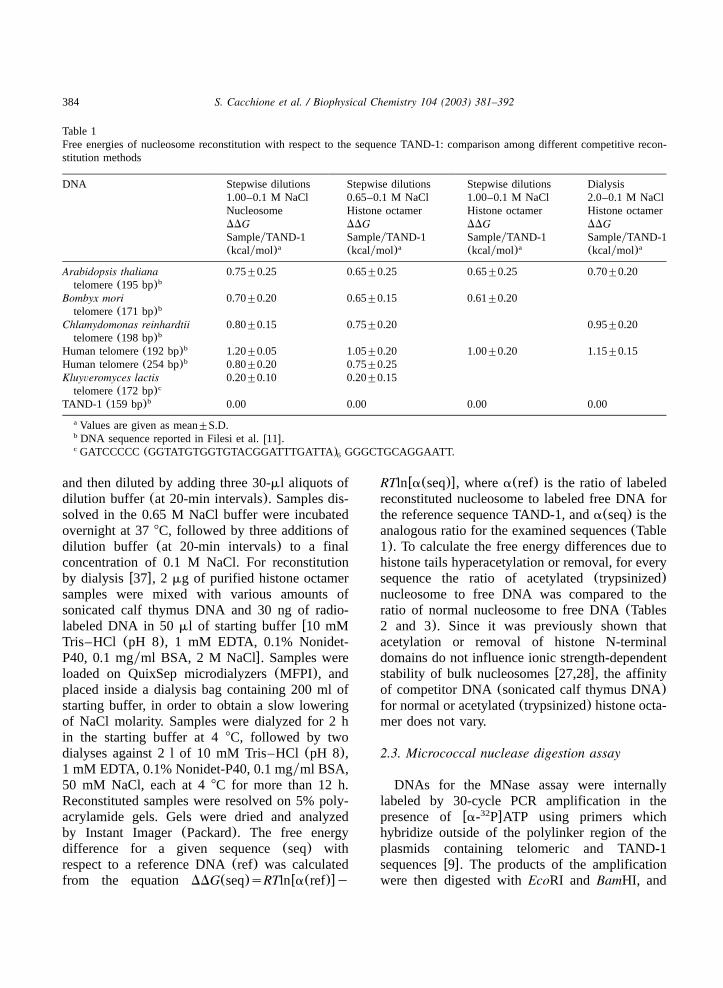
Table 1Free energies of nucleosome reconstitution with respect to the sequence TAND-1: comparison among different competitive recon-stitution methods
DNA Stepwise dilutions Stepwise dilutions Stepwise dilutions Dialysis1.00–0.1 M NaCl 0.65–0.1 M NaCl 1.00–0.1 M NaCl 2.0–0.1 M NaClNucleosome Histone octamer Histone octamer Histone octamerDDG DDG DDG DDGSampleyTAND-1 SampleyTAND-1 SampleyTAND-1 SampleyTAND-1(kcalymol)a (kcalymol)a (kcalymol)a (kcalymol)a
Arabidopsis thaliana 0.75"0.25 0.65"0.25 0.65"0.25 0.70"0.20telomere(195 bp)b
Bombyx mori 0.70"0.20 0.65"0.15 0.61"0.20telomere(171 bp)b
Chlamydomonas reinhardtii 0.80"0.15 0.75"0.20 0.95"0.20telomere(198 bp)b
Human telomere(192 bp)b 1.20"0.05 1.05"0.20 1.00"0.20 1.15"0.15Human telomere(254 bp)b 0.80"0.20 0.75"0.25Kluyveromyces lactis 0.20"0.10 0.20"0.15telomere(172 bp)c
TAND-1 (159 bp)b 0.00 0.00 0.00 0.00
Values are given as mean"S.D.a
DNA sequence reported in Filesi et al.w11x.b
GATCCCCC(GGTATGTGGTGTACGGATTTGATTA) GGGCTGCAGGAATT.c6
and then diluted by adding three 30-ml aliquots ofdilution buffer (at 20-min intervals). Samples dis-solved in the 0.65 M NaCl buffer were incubatedovernight at 378C, followed by three additions ofdilution buffer (at 20-min intervals) to a finalconcentration of 0.1 M NaCl. For reconstitutionby dialysisw37x, 2 mg of purified histone octamersamples were mixed with various amounts ofsonicated calf thymus DNA and 30 ng of radio-labeled DNA in 50ml of starting bufferw10 mMTris–HCl (pH 8), 1 mM EDTA, 0.1% Nonidet-P40, 0.1 mgyml BSA, 2 M NaClx. Samples wereloaded on QuixSep microdialyzers(MFPI), andplaced inside a dialysis bag containing 200 ml ofstarting buffer, in order to obtain a slow loweringof NaCl molarity. Samples were dialyzed for 2 hin the starting buffer at 48C, followed by twodialyses against 2 l of 10 mM Tris–HCl (pH 8),1 mM EDTA, 0.1% Nonidet-P40, 0.1 mgyml BSA,50 mM NaCl, each at 48C for more than 12 h.Reconstituted samples were resolved on 5% poly-acrylamide gels. Gels were dried and analyzedby Instant Imager(Packard). The free energydifference for a given sequence(seq) withrespect to a reference DNA(ref) was calculatedfrom the equation DDG(seq)sRTlnwa(ref)xy
RTlnwa(seq)x, wherea(ref) is the ratio of labeledreconstituted nucleosome to labeled free DNA forthe reference sequence TAND-1, anda(seq) is theanalogous ratio for the examined sequences(Table1). To calculate the free energy differences due tohistone tails hyperacetylation or removal, for everysequence the ratio of acetylated(trypsinized)nucleosome to free DNA was compared to theratio of normal nucleosome to free DNA(Tables2 and 3). Since it was previously shown thatacetylation or removal of histone N-terminaldomains do not influence ionic strength-dependentstability of bulk nucleosomesw27,28x, the affinityof competitor DNA(sonicated calf thymus DNA)for normal or acetylated(trypsinized) histone octa-mer does not vary.
2.3. Micrococcal nuclease digestion assay
DNAs for the MNase assay were internallylabeled by 30-cycle PCR amplification in thepresence of wa- PxATP using primers which32
hybridize outside of the polylinker region of theplasmids containing telomeric and TAND-1sequencesw9x. The products of the amplificationwere then digested withEcoRI and BamHI, and
385S. Cacchione et al. / Biophysical Chemistry 104 (2003) 381–392
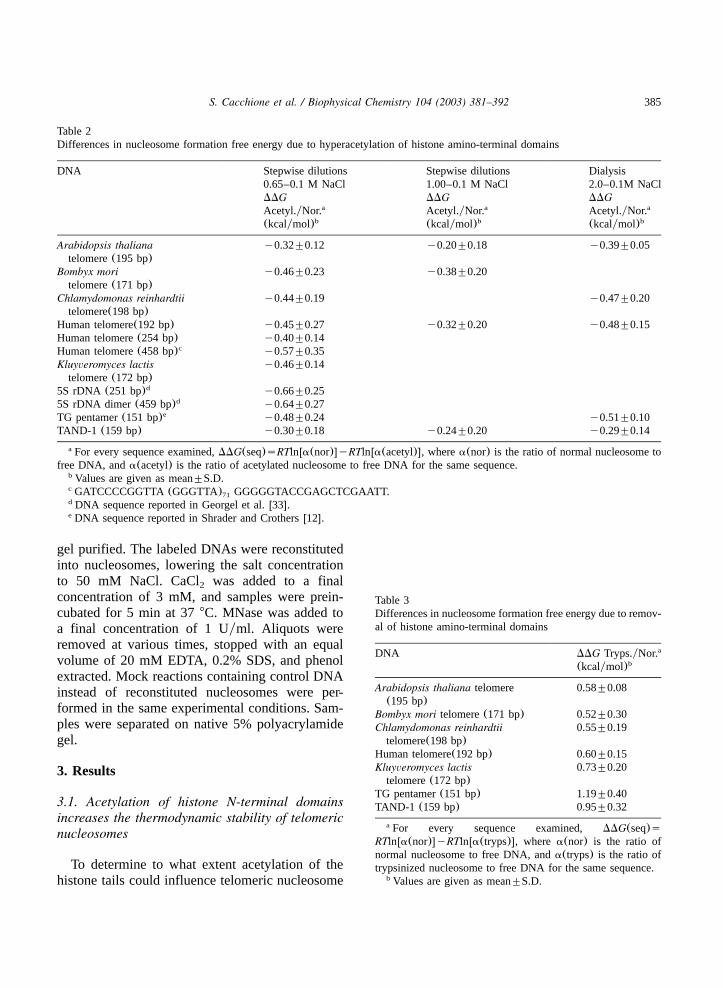
Table 2Differences in nucleosome formation free energy due to hyperacetylation of histone amino-terminal domains
DNA Stepwise dilutions Stepwise dilutions Dialysis0.65–0.1 M NaCl 1.00–0.1 M NaCl 2.0–0.1M NaClDDG DDG DDGAcetyl.yNor.a Acetyl.yNor.a Acetyl.yNor.a
(kcalymol)b (kcalymol)b (kcalymol)b
Arabidopsis thaliana y0.32"0.12 y0.20"0.18 y0.39"0.05telomere(195 bp)
Bombyx mori y0.46"0.23 y0.38"0.20telomere(171 bp)
Chlamydomonas reinhardtii y0.44"0.19 y0.47"0.20telomere(198 bp)
Human telomere(192 bp) y0.45"0.27 y0.32"0.20 y0.48"0.15Human telomere(254 bp) y0.40"0.14Human telomere(458 bp)c y0.57"0.35Kluyveromyces lactis y0.46"0.14telomere(172 bp)
5S rDNA (251 bp)d y0.66"0.255S rDNA dimer(459 bp)d y0.64"0.27TG pentamer(151 bp)e y0.48"0.24 y0.51"0.10TAND-1 (159 bp) y0.30"0.18 y0.24"0.20 y0.29"0.14
For every sequence examined,DDG(seq)sRTlnwa(nor)xyRTlnwa(acetyl)x, wherea(nor) is the ratio of normal nucleosome toa
free DNA, anda(acetyl) is the ratio of acetylated nucleosome to free DNA for the same sequence.Values are given as mean"S.D.b
GATCCCCGGTTA(GGGTTA) GGGGGTACCGAGCTCGAATT.c71
DNA sequence reported in Georgel et al.w33x.d
DNA sequence reported in Shrader and Crothersw12x.e
Table 3Differences in nucleosome formation free energy due to remov-al of histone amino-terminal domains
DNA DDG Tryps.yNor.a
(kcalymol)b
Arabidopsis thaliana telomere 0.58"0.08(195 bp)
Bombyx mori telomere(171 bp) 0.52"0.30Chlamydomonas reinhardtii 0.55"0.19telomere(198 bp)
Human telomere(192 bp) 0.60"0.15Kluyveromyces lactis 0.73"0.20telomere(172 bp)
TG pentamer(151 bp) 1.19"0.40TAND-1 (159 bp) 0.95"0.32
For every sequence examined, DDG(seq)sa
RTlnwa(nor)xyRTlnwa(tryps)x, wherea(nor) is the ratio ofnormal nucleosome to free DNA, anda(tryps) is the ratio oftrypsinized nucleosome to free DNA for the same sequence.
Values are given as mean"S.D.b
gel purified. The labeled DNAs were reconstitutedinto nucleosomes, lowering the salt concentrationto 50 mM NaCl. CaCl was added to a final2
concentration of 3 mM, and samples were prein-cubated for 5 min at 378C. MNase was added toa final concentration of 1 Uyml. Aliquots wereremoved at various times, stopped with an equalvolume of 20 mM EDTA, 0.2% SDS, and phenolextracted. Mock reactions containing control DNAinstead of reconstituted nucleosomes were per-formed in the same experimental conditions. Sam-ples were separated on native 5% polyacrylamidegel.
3. Results
3.1. Acetylation of histone N-terminal domainsincreases the thermodynamic stability of telomericnucleosomes
To determine to what extent acetylation of thehistone tails could influence telomeric nucleosome
386 S. Cacchione et al. / Biophysical Chemistry 104 (2003) 381–392
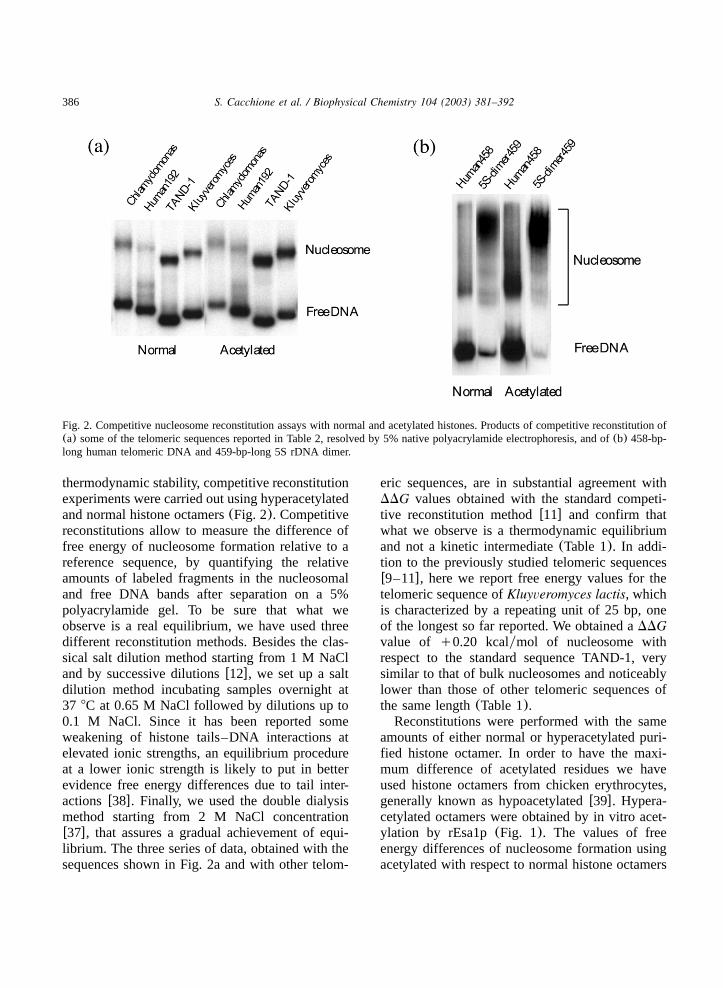
Fig. 2. Competitive nucleosome reconstitution assays with normal and acetylated histones. Products of competitive reconstitution of(a) some of the telomeric sequences reported in Table 2, resolved by 5% native polyacrylamide electrophoresis, and of(b) 458-bp-long human telomeric DNA and 459-bp-long 5S rDNA dimer.
thermodynamic stability, competitive reconstitutionexperiments were carried out using hyperacetylatedand normal histone octamers(Fig. 2). Competitivereconstitutions allow to measure the difference offree energy of nucleosome formation relative to areference sequence, by quantifying the relativeamounts of labeled fragments in the nucleosomaland free DNA bands after separation on a 5%polyacrylamide gel. To be sure that what weobserve is a real equilibrium, we have used threedifferent reconstitution methods. Besides the clas-sical salt dilution method starting from 1 M NaCland by successive dilutionsw12x, we set up a saltdilution method incubating samples overnight at37 8C at 0.65 M NaCl followed by dilutions up to0.1 M NaCl. Since it has been reported someweakening of histone tails–DNA interactions atelevated ionic strengths, an equilibrium procedureat a lower ionic strength is likely to put in betterevidence free energy differences due to tail inter-actionsw38x. Finally, we used the double dialysismethod starting from 2 M NaCl concentrationw37x, that assures a gradual achievement of equi-librium. The three series of data, obtained with thesequences shown in Fig. 2a and with other telom-
eric sequences, are in substantial agreement withDDG values obtained with the standard competi-tive reconstitution methodw11x and confirm thatwhat we observe is a thermodynamic equilibriumand not a kinetic intermediate(Table 1). In addi-tion to the previously studied telomeric sequencesw9–11x, here we report free energy values for thetelomeric sequence ofKluyveromyces lactis, whichis characterized by a repeating unit of 25 bp, oneof the longest so far reported. We obtained aDDGvalue of q0.20 kcalymol of nucleosome withrespect to the standard sequence TAND-1, verysimilar to that of bulk nucleosomes and noticeablylower than those of other telomeric sequences ofthe same length(Table 1).Reconstitutions were performed with the same
amounts of either normal or hyperacetylated puri-fied histone octamer. In order to have the maxi-mum difference of acetylated residues we haveused histone octamers from chicken erythrocytes,generally known as hypoacetylatedw39x. Hypera-cetylated octamers were obtained by in vitro acet-ylation by rEsa1p(Fig. 1). The values of freeenergy differences of nucleosome formation usingacetylated with respect to normal histone octamers
387S. Cacchione et al. / Biophysical Chemistry 104 (2003) 381–392
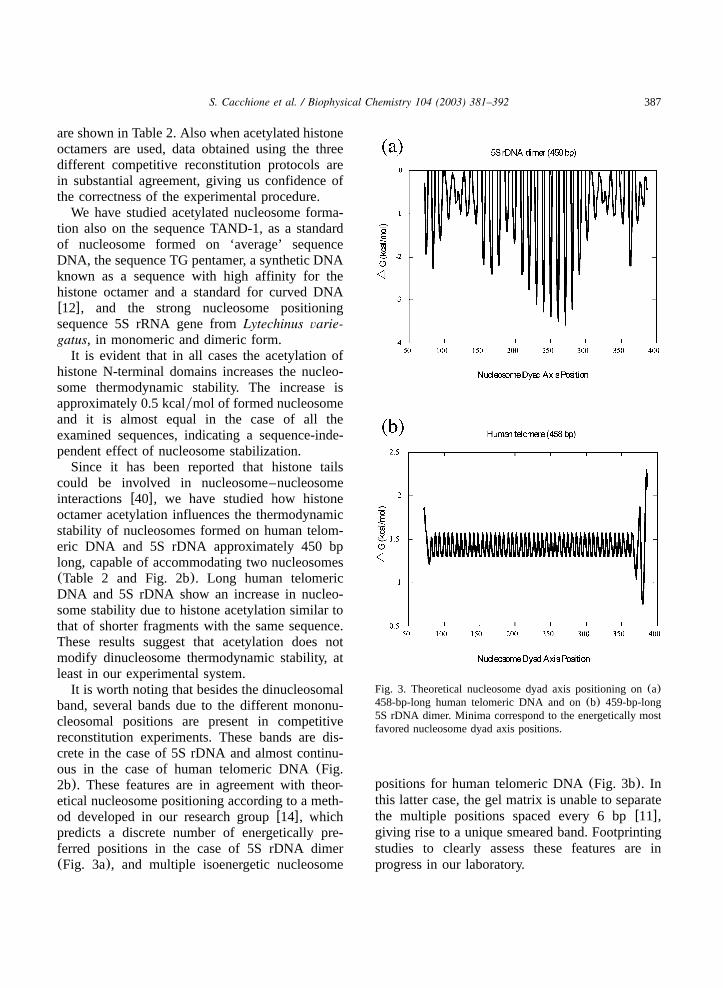
Fig. 3. Theoretical nucleosome dyad axis positioning on(a)458-bp-long human telomeric DNA and on(b) 459-bp-long5S rDNA dimer. Minima correspond to the energetically mostfavored nucleosome dyad axis positions.
are shown in Table 2. Also when acetylated histoneoctamers are used, data obtained using the threedifferent competitive reconstitution protocols arein substantial agreement, giving us confidence ofthe correctness of the experimental procedure.We have studied acetylated nucleosome forma-
tion also on the sequence TAND-1, as a standardof nucleosome formed on ‘average’ sequenceDNA, the sequence TG pentamer, a synthetic DNAknown as a sequence with high affinity for thehistone octamer and a standard for curved DNAw12x, and the strong nucleosome positioningsequence 5S rRNA gene fromLytechinus varie-gatus, in monomeric and dimeric form.It is evident that in all cases the acetylation of
histone N-terminal domains increases the nucleo-some thermodynamic stability. The increase isapproximately 0.5 kcalymol of formed nucleosomeand it is almost equal in the case of all theexamined sequences, indicating a sequence-inde-pendent effect of nucleosome stabilization.Since it has been reported that histone tails
could be involved in nucleosome–nucleosomeinteractions w40x, we have studied how histoneoctamer acetylation influences the thermodynamicstability of nucleosomes formed on human telom-eric DNA and 5S rDNA approximately 450 bplong, capable of accommodating two nucleosomes(Table 2 and Fig. 2b). Long human telomericDNA and 5S rDNA show an increase in nucleo-some stability due to histone acetylation similar tothat of shorter fragments with the same sequence.These results suggest that acetylation does notmodify dinucleosome thermodynamic stability, atleast in our experimental system.It is worth noting that besides the dinucleosomal
band, several bands due to the different mononu-cleosomal positions are present in competitivereconstitution experiments. These bands are dis-crete in the case of 5S rDNA and almost continu-ous in the case of human telomeric DNA(Fig.2b). These features are in agreement with theor-etical nucleosome positioning according to a meth-od developed in our research groupw14x, whichpredicts a discrete number of energetically pre-ferred positions in the case of 5S rDNA dimer(Fig. 3a), and multiple isoenergetic nucleosome
positions for human telomeric DNA(Fig. 3b). Inthis latter case, the gel matrix is unable to separatethe multiple positions spaced every 6 bpw11x,giving rise to a unique smeared band. Footprintingstudies to clearly assess these features are inprogress in our laboratory.

388 S. Cacchione et al. / Biophysical Chemistry 104 (2003) 381–392
Fig. 4. Polyacrylamide gel electrophoresis of competitive nucleosome reconstitution experiments with normal and trypsinizedhistones.
3.2. Removal of histone N-terminal domainsdecreases the thermodynamic stability of telomericnucleosomes
The results obtained in the case of hyperacety-lated nucleosomes suggest that the decrease ofelectrostatic interactions between DNA and histoneside chains does not necessarily give rise to lessstable nucleosomes. Since nucleosomes lackinghistone N-terminal domains have been often con-sidered as a suitable model for hyperacetylatedhistones, we carried out competitive reconstitutionexperiments using selectively trypsinized nucleo-somes in comparison with normal nucleosomesfrom chicken erythrocytes(Fig. 4, Table 3). Wemeasured the free energies of nucleosome forma-tion on the telomeric sequences, together with thesequence TAND-1, and the sequence TG pentamer.Tail removal causes a decrease of nucleosomestability for all the examined sequences, but thedecrease is significatively smaller for telomericsequences(approx. 0.5 kcalymol of nucleosome)with respect to average sequence and curved DNAs
(approx. 1 kcalymol of nucleosome). These resultsindicate that the interactions of histone tails withtelomeric sequences are less strong than withTAND-1 and TG pentamer.
3.3. Acetylated nucleosomes show higher accessi-bility to MNase digestion than normal nucleosomes
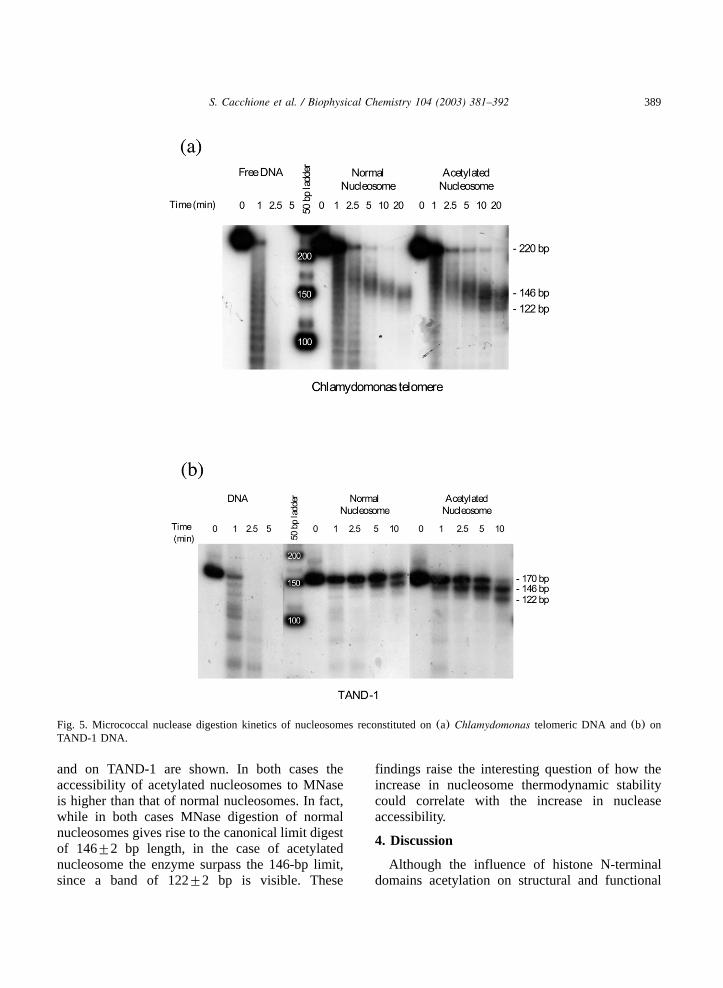
MNase was used as a probe of acetylated nucle-osome structure. We have previously found thatMNase and DNAse I digestion patterns of telom-eric nucleosomes are typical of canonical nucleo-somesw9,10x. In fact MNase digestion gives riseto a digest limit corresponding to 146"2 bp, whileDNAse I footprinting is characterized by thecanonical 10 base periodicity. The finding thatnucleosome organized onto hyperacetylated his-tone octamers are thermodynamically more stablethan those organized onto normal histone octamerssuggests that this histone modification should notincrease nucleosome accessibility to MNase. InFig. 5 MNase digestion kinetics of nucleosomesreconstituted onChlamydomonas telomeric DNA
389S. Cacchione et al. / Biophysical Chemistry 104 (2003) 381–392
Fig. 5. Micrococcal nuclease digestion kinetics of nucleosomes reconstituted on(a) Chlamydomonas telomeric DNA and(b) onTAND-1 DNA.
and on TAND-1 are shown. In both cases theaccessibility of acetylated nucleosomes to MNaseis higher than that of normal nucleosomes. In fact,while in both cases MNase digestion of normalnucleosomes gives rise to the canonical limit digestof 146"2 bp length, in the case of acetylatednucleosome the enzyme surpass the 146-bp limit,since a band of 122"2 bp is visible. These
findings raise the interesting question of how theincrease in nucleosome thermodynamic stabilitycould correlate with the increase in nucleaseaccessibility.
4. Discussion
Although the influence of histone N-terminaldomains acetylation on structural and functional

390 S. Cacchione et al. / Biophysical Chemistry 104 (2003) 381–392
properties of nucleosomes has been extensivelystudied, acetylated nucleosome structural and func-tional features have yet to be adequately estab-lished w26x. In this research, we studied the effectof histone acetylation and DNA length on thethermodynamic stability of nucleosomes formedon a set of several different telomeric sequences,on DNAs with high affinity for the histone octamer(5S rDNA and TG pentamer), and on the sequenceTAND-1, having the same free energy of nucleo-some formation as bulk DNA. We have previouslyshown that telomeric DNAs are straight and organ-ize nucleosomes with the lowest stability amongall the DNA sequences so far studiedw10,11x.We find that acetylation increases the stability
of nucleosomes formed on all the examinedsequences by approximately 0.5 kcalymol, makingthe assembly of nucleosome twice more probablewith acetylated than with normal histones(seeTable 2). These findings suggest the possibilitythat in nucleosome assembly histone acetylation,besides favoring the interactions with assemblyfactors such as CAFw23x, could have an intrinsicrole in making more probable the nucleosomeformation. These features would be relevant inheterochromatic as well as in euchromaticdomains. Since these findings could be consideredin apparent contrast with the hypothesis that oneof the main effects of acetylation should be thedecrease of electrostatic interactions between his-tones and DNA, we have studied the influence ofthe removal of histone N-terminal on nucleosomethermodynamic stability. The removal of histoneamino terminal domains causes a decrease ofnucleosome thermodynamic stability for all theexamined sequences. However, telomeric sequenc-es are less affected by tail removal than ‘average’nucleosomal DNA(TAND-1) and curved DNA(TG pentamer). These results indicate that telom-eric sequences should interact less tightly withhistone tails than the other examined DNAsequences, and suggest that the extent of tailinteractions is sequence-specific.The enhanced stability of acetylated nucleo-
somes is in apparent contrast with the view ofmore ‘open’ conformation with respect to normalnucleosomes. Since evidences on facilitated bind-ing of specific proteins to DNA organized on
acetylated nucleosomes is still very controversialw26x, we studied the accessibility of nucleosomalDNA to MNase, a probe with low sequence-specificity that preferably digests internucleosomalDNA. MNase digestion kinetics shows anenhanced accessibility of nucleosomal DNA whenthe histone octamer is hyperacetylated, both in thecase of telomeric and ‘average’ sequence DNA,with a protection from digestion of approximately120 bp instead of the canonical 146 bp.At first sight our results might appear as contra-
dictory, because we have found that hyperacetylat-ed nucleosomes are thermodynamically morestable than hypoacetylated nucleosomes, and, atthe same time, they show an enhanced accessibilityto MNase as well as to other probesw41x. However,the recent resolution of the nucleosome structureat 2.8 Aw42x and at 1.9 Aw43x suggests interesting˚ ˚remarks. Although several histone N-terminal res-idues interact with nucleosomal DNA, acetylatablelysines seem not involved in histone–DNA con-tacts w43x. Consequently, histone acetylation islikely not to cause any binding loss between N-terminal tails and nucleosomal DNA. Theincreased stability of acetylated nucleosomes couldderive from more favorable local interactions orfrom conformational changes influencing the over-all nucleosome structure. It has been reported thathistone acetylation is able to reduce the linkingnumber change per nucleosome in reconstitutedminichromosomesw44x. More recently Ausio andco-workers showed that acetylation of histonesresults in a significant increase of thea-helicalcontent of their tailsw45x. This increase couldcause an increased hydrophobicity of the tailsallowing better interactions with DNA grooves.Moreover, it has been proposed that the increaseof thea-helical content could result in shorteningof histone N-terminal domainsw45x. This lastobservation could contribute to explain theenhanced accessibility of hyperacetylated nucleo-somes to enzymatic probes, since from the nucle-osome structure at 1.9 A resolutionw43x it emerges˚that the terminal 13 bp of the DNA superhelixinteracts with H3 N-terminal tail. However, it islikely that a definite answer to this issue couldcome only from the resolution of the structure of

391S. Cacchione et al. / Biophysical Chemistry 104 (2003) 381–392
an acetylated nucleosome by crystallograficanalysis.
Acknowledgments
Thanks are due to P. De Santis and A. Scipionifor many helpful discussions. This work was finan-cially supported by the ItalyySpain program AzioniintegrateyAcciones Integradas 1999, and by COF-IN MURST 2001y2003.
References
w1x E.H. Blackburn, C.W. Greider, Telomeres, Cold SpringHarbor Laboratory Press, Cold Spring Harbor, NewYork, 1995.
w2x C.W. Greider, Telomeres length regulation, Annu. Rev.Biochem. 65(1996) 337–365.
w3x V.L. Makarov, S. Lejnine, J. Bedoyan, J.P. Langmore,Nucleosomal organization of telomere-specific chroma-tin in rat, Cell 73(1993) 775–787.
w4x H. Tommerup, A. Dousmanis, T. De Lange, Unusualchromatin in human telomeres, Mol. Cell. Biol. 14(1994) 5777–5785.
w5x S. Lejnine, V.L. Makarov, J.P. Langmore, Conservednucleoprotein structure at the ends of vertebrate andinvertebrate chromosomes, Proc. Natl. Acad. Sci. U.S.A.92 (1995) 2393–2397.
w6x J. Fajkus, J. Kovarik, R. Kralovics, M. Bezdek, Organ-ization of telomeric and subtelomeric chromatin in thehigher plantNicotiana tabacum, Mol. Gen. Genet. 247(1995) 633–638.
w7x J.H. Wright, D.E. Gottschling, V.A. Zakian, Saccharo-myces telomeres assume a non-nucleosomal chromatinstructure, Genes Dev. 6(1992) 197–210.
w8x P. Cohen, E. Blackburn, Two types of telomeric chro-matin in Tetrahymena thermophila, J. Mol. Biol. 280(1998) 327–344.
w9x S. Cacchione, M.A. Cerone, M. Savino, In vitro lowpropensity to form nucleosomes of four telomericsequences, FEBS Lett. 400(1997) 37–41.
w10x L. Rossetti, S. Cacchione, M. Fua, M. Savino, Nucleo-`some assembly on telomeric sequences, Biochemistry37 (1998) 6727–6737.
w11x I. Filesi, S. Cacchione, P. De Santis, L. Rossetti, M.Savino, The main role of the sequence-dependent DNAelasticity in determining the free energy of nucleosomeformation on telomeric DNA, Biophys. Chem. 83(2000) 223–237.
w12x T.E. Shrader, D.M. Crothers, Artificial nucleosome posi-tioning sequences, Proc. Natl. Acad. Sci. U.S.A. 86(1989) 7418–7422.
w13x C. Anselmi, G. Bocchinfuso, P. De Santis, M. Fua, M.`Savino, A. Scipioni, Dual role of DNA intrinsic curva-
ture and flexibility in determining nucleosome stability,J. Mol. Biol. 286(1999) 1293–1301.
w14x C. Anselmi, P. De Santis, R. Paparcone, M. Savino, A.Scipioni, From the sequence to the superstructural prop-erties of DNAs, Biophys. Chem. 95(2002) 23–47.
w15x C. Anselmi, G. Bocchinfuso, P. De Santis, M. Savino,A. Scipioni, A theoretical model for the prediction ofsequence-dependent nucleosome thermodynamic stabil-ity, Biophys. J. 79(2000) 601–613.
w16x K.E. van Holde, Chromatin, Springer Verlag, New York,1988.
w17x B.D. Strahl, C.D. Allis, The language of covalenthistone modifications, Nature 403(2000) 41–45.
w18x G. Lopez-Rodas, G. Brosch, E.I. Georgieva, R. Sendra,L. Franco, P. Loidl, Histone deacetylase. A key enzymefor the binding of regulatory proteins to chromatin,FEBS Lett. 317(1993) 175–180.
w19x M. Grunstein, Histone acetylation in chromatin structureand transcription, Nature 389(1997) 349–352.
w20x L.P. O’Neill, B.M. Turner, Histone H4 acetylation dis-tinguishes coding regions of the human genome fromheterochromatin in a differentiation-dependent but tran-scription-independent manner, EMBO J. 14(1995)3946–3957.
w21x B.M. Turner, A.J. Birley, J. Lavender, Histone H4isoforms acetylated at specific lysine residues defineindividual chromosomes and chromatin domains inDro-sophila polytene nuclei, Cell 69(1992) 375–384.
w22x M. Braunstein, R.E. Sobel, C.D. Allis, B.M. Turner,J.R. Broach, Efficient transcriptional silencing inSac-charomyces cerevisiae requires a heterochromatin his-tone acetylation pattern, Mol. Cell. Biol. 16(1996)4349–4356.
w23x R.E. Sobel, R.G. Cooke, C.A. Perry, A.T. Annunziato,C.D. Allis, Conservation of deposition-related acetyla-tion sites in newly synthesized histones H3 and H4,Proc. Natl. Acad. Sci. U.S.A. 92(1995) 1237–1241.
w24x A. Verreault, De novo nucleosome assembly: new piecesin an old puzzle, Genes Dev. 14(2000) 1430–1438.
w25x Z. Jasencakova, A. Meister, J. Walter, B.M. Turner, I.Schubert, Histone H4 acetylation of euchromatin andheterochromatin is cell cycle dependent and correlatedwith replication rather than with transcription, Plant Cell12 (2000) 2087–2100.
w26x J. Ausio, D.W. Abbott, X. Wang, S.C. Moore, Histonevariants and histone modifications: a structural perspec-tive, Biochem. Cell Biol. 79(2001) 693–708.
w27x J. Ausio, K.E. van Holde, Histone hyperacetylation: itseffect on nucleosome conformation and stability, Bio-chemistry 22(1986) 1421–1428.
w28x J. Ausio, F. Dong, K.E. van Holde, Use of selectivelytrypsinized nucleosome core particles to analyze therole of the histone tails in the stabilization of thenucleosome, J. Mol. Biol. 206(1989) 451–463.
w29x M. Garcia-Ramirez, F. Dong, J. Ausio, Role of thehistone ‘tails’ in the folding of oligonucleosomes deplet-

392 S. Cacchione et al. / Biophysical Chemistry 104 (2003) 381–392
ed of histone H1, J. Biol. Chem. 267(1992)19587–19595.
w30x M. Garcia-Ramirez, C. Rocchini, J. Ausio, Modulationof chromatin folding by histone acetylation, J. Biol.Chem. 270(1995) 17923–17928.
w31x P. De Santis, B. Kropp, L. Leoni, B. Sampaolese, M.Savino, Influence of DNA superstructural features andhistones aminoterminal domains on mononucleosomeand dinucleosome positioning, Biophys. Chem. 62(1996) 47–61.
w32x H.R. Widlund, J.M. Vitolo, C. Thiriet, J.J. Hayes, DNAsequence-dependent contributions of core histone tailsto nucleosome stability: differential effects of acetyla-tion and proteolytic tail removal, Biochemistry 39(2000) 3835–3841.
w33x P. Georgel, B. Demeler, C. Terpening, M.R. Paule, K.E.van Holde, Binding of the RNA polymerase I transcrip-tion complex to its promoter can modify positioning ofdownstream nucleosomes assembled in vitro, J. Biol.Chem. 268(1993) 1947–1954.
w34x E.R. Smith, A. Eisen, W. Gu, et al., ESA1 is a histoneacetyltransferase that is essential for growth in yeast,Proc. Natl. Acad. Sci. U.S.A. 95(1998) 3561–3565.
w35x K.W. Marvin, P. Yau, E.M. Bradbury, Isolation andcharacterization of acetylated histones H3 and H4 andtheir assembly into nucleosomes, J. Biol. Chem. 265(1990) 19839–19847.
w36x W.M. Bonner, M.H. West, J.D. Stedman, Two-dimen-sional gel analysis of histones in acid extracts of nuclei,cells, and tissues, Eur. J. Biochem. 109(1980) 17–23.
w37x A. Thastrom, P.T. Lowary, H.R. Widlund, H. Cao, M.¨˚Kubista, J. Widom, Sequence motifs and free energies
of selected natural and non-natural nucleosome posi-tioning DNA sequences, J. Mol. Biol. 288(1999)213–229.
w38x H.R. Drew, Can one measure the free energy of bindingof the histone octamer to different DNA sequences bysalt-dependent reconstitution?, J. Mol. Biol. 219(1991)391–392.
w39x D.-E. Zhang, D.A. Nelson, Histone acetylation in chick-en erythrocytes. Rates of acetylation and evidence thathistones in both active and potentially active chromatinare rapidly modified, Biochem. J. 250(1988) 233–240.
w40x T.E. Shrader, D.M. Crothers, Effects of DNA sequenceand histone–histone interactions on nucleosome place-ment, J. Mol. Biol. 216(1990) 69–84.
w41x J.D. Anderson, P.T. Lowary, J. Widom, Effects ofhistone acetylation on the equilibrium accessibility ofnucleosomal DNA target sites, J. Mol. Biol. 307(2001)977–985.
w42x K. Luger, A.W. Mader, R.K. Richmond, D.F. Sargent,T.J. Richmond, X-Ray structure of the nucleosome coreparticle at 2.8 A resolution, Nature 389(1997) 251–259.˚
w43x C.A. Davey, D.F. Sargent, K. Luger, A.W. Mader, T.J.Richmond, Solvent mediated interactions in the structureof the nucleosome core particle at 1.9 A resolution, J.˚Mol. Biol. 319 (2002) 1097–1113.
w44x V.G. Norton, B.S. Imai, P. Yau, E.M. Bradbury, Histoneacetylation reduces nucleosome core particle linkingnumber change, Cell 57(1989) 449–457.
w45x X. Wang, S.C. Moore, M. Laszckzack, J. Ausio, Acet-ylation increases the alpha-helical content of the histonetails of the nucleosome, J. Biol. Chem. 275(2000)35013–35020.