mre11p nuclease activity is dispensable for telomeric rapid deletion
TRANSCRIPT

DNA Repair 4 (2005) 994–1005
Mre11p nuclease activity is dispensable for telomeric rapid deletion
Bridget Williams, Mrinal K. Bhattacharyya, Arthur J. Lustig∗
Department of Biochemistry, Tulane University Medical School and Tulane Cancer Center, 1430 Tulane Avenue, New Orleans, LA 70112, USA
Received 18 January 2005; received in revised form 28 April 2005; accepted 29 April 2005Available online 28 June 2005
Abstract
Telomeric rapid deletion (TRD) is an intrachromatid recombination process that truncates over-elongated telomeres to the geneticallydetermined average telomere length. We have proposed that TRD is initiated by invasion of the 3′ G-rich overhang into centromere-proximaltelomere sequence, forming an intermediate that leads to excision of the distal telomere tract. TRD efficiency is dependent on Mre11p andRad50p, two members of the widely conserved Mre11p/Rad50p/Xrs2p (MRX) complex. To investigate the role of Mre11p in TRD, weconducted a structure/function analysis by testing the TRD rate and precision of mutations within known functional domains. We analyzed12 alleles that disrupt different Mre11p activities. Surprisingly, mutations in essential residues of the nuclease domain do not inhibit TRD,e o1p in anM tion of DNAb g domainsd efect in theM of TRD.R elomeresd so exhibitst mplex thati©
K
1
pesslc
ett
gthsbyrac-eeasttents
tatelyosis,
ansed in
1d
ffectively ruling out nuclease activity as the source of the Mre11p requirement. Interestingly, loss of Exo1p alone or loss of Exre11 nuclease deficient background does not eliminate TRD, suggesting the presence of an additional nuclease. Second, deleinding sites A (residues 410–420) and B (residues 644–692) actually enhances the TRD rate. Even deletion of both DNA bindinoes not abrogate TRD, although its kinetics and precision are variable. This suggests altered DNA binding or a conformational dRX complex may affect the rate of TRD product formation and indicates that the DNA binding sites formally act as repressorsemarkably, the H213Y allele (nuclease motif IV) confers an extraordinarily rapid kinetics, with the vast majority of elongated teleted imprecisely in a single round of subculturing. In striking contrast, the P162S allele that confers dissolution of the complex al
he null phenotype. These data suggest that Mre11p can act as a positive and negative regulator of TRD in context of the MRX cos essential for TRD.
2005 Elsevier B.V. All rights reserved.
eywords:Mre11p; MRX complex; TRD; Recombination; Telomeres
. Introduction
The maintenance of telomeres, the protein–DNA com-lexes present at the termini of eukaryotic chromosomes, isssential for complete replication and capping of chromo-ome ends that are vital processes in maintaining genomictability. Indeed, loss of telomeric G + T simple sequenceength or structure is a key step in oncogenesis, cellular senes-ence, and apoptosis (reviewed in refs.[1–3]).
Telomere length is maintained through a tightly regulatedquilibrium between mechanisms that shorten and lengthen
elomere tracts. One of these processes, telomeric rapid dele-ion (TRD), is the single-stepRAD52-dependent intrachro-
∗ Corresponding author. Tel.: +1 504 988 3688; fax: +1 504 988 2739.E-mail address:[email protected] (A.J. Lustig).
matid deletion of elongated telomeres to wild type len[4,5], with the size of the TRD products being governedthe size of the majority of telomeres in an as yet unchaterized trans-acting process[4]. This latter property may bmediated through the anchoring and/or clustering of ytelomeres at the nuclear periphery[6]. Interestingly, efficienTRD requires both Mre11p and Rad50p, two componof the MRX complex. Null mutations of eitherMRE11orRAD50dramatically impair TRD, withmre11null allelesdisplaying the most severe phenotypes[5].
TRD occurs at a rate of 3× 10−3 events/cell division aan individual telomere, a rate that increases approxim40-fold in a comparable process present early in meieffectively resetting telomeres to wild type size[7]. TRDalso appears to influence telomere dynamics in hum[8,9]. Abrupt telomere deletion events have been observ
568-7864/$ – see front matter © 2005 Elsevier B.V. All rights reserved.oi:10.1016/j.dnarep.2005.04.016

B. Williams et al. / DNA Repair 4 (2005) 994–1005 995
numerous contexts including pre-senescent fibroblasts, cellsthat follow an alternative recombinational pathway of telom-ere addition (ALT), and in mutants of the human end bindingprotein hPOT1[9]. More recent studies have provided defini-tive proof for the excision of t-loops from human telomeresin cell culture[8].
T-loops form as a consequence of 3′ strand invasion ofthe chromosome terminus into duplex repeats forming aD-loop after pairing and are likely to be involved in endprotection[2,10–14]. Physical studies from our laboratory[4,5] suggested that TRD is mediated through invasion ofthe 3′ terminus into basal sequences, resulting in a transient‘t-loop-like’ intermediate that in yeast, is rapidly resolvedinto a truncated telomere and a linear or circular byproductby recombination. Interestingly, telomeric circles have beenrecently detected in yeast and human cells[8,15–17].
Mre11p is functional at the telomere in multiple aspectsof telomere size control and in the mitotic telomere/telomererecombination pathway that is responsible for post-senescentsurvivors in telomerase negative cells[18–21]. Mre11pappears to act in part as an upstream DNA damage sensorfor the ATM/ATR orthologues, Tel1p and Mec1p, both ofwhich are capable of telomere binding[22–24]. Indeed, Tel1pis dependent on the MRX complex for association in G1phase (Hector and Runge, personal communication). Termi-nal DNA damage may be ‘sensed’ as aberrant long overhangso m-e 1a telye ments[ 11pi
tivityi sme ow-e dent3 cedDh n-j berM ,i -ao nec-e thesis[ mericn ner-a 1, 3t rasea -r NAd
m-p to
telomeres where NBS1 and, presumably MRN, interacts withthe telomere binding protein TRF2[37]. TRF2 has also beenshown to bind at t-loop junctions in vitro[38]. Indeed a largeassembly of factors normally ascribed to DNA damage detec-tion and repair are likely to be present at t-loops in vivo. Theseinclude the BLM and WRN helicases, p53, and DNA-PKcs(the catalytic subunit of Ku not present in yeast)[39–47].These data suggest that DNA repair factors play a uniquerole to stabilize telomeres through association with t-loops[48].
In the current study, we present a comprehensivestructure–function analysis of Mre11p in TRD. Unexpect-edly, we find the nuclease activity of Mre11p is dispensablefor TRD, ruling out one of the major models for the actionof Mre11p. We present data suggesting that Mre11p domainshave both positive and negative functions that govern TRDrate and precision in context of the MRX complex and doesnot depend on the exonuclease Exo1p.
2. Materials and methods
2.1. Strains, plasmids, and media
YPD supplemented with 30 mg/mL adenine (YPAD) andSC media were prepared by standard methods. All strainsuH
a I-S odC n-iw entso ineS them oft ukiO two-s n ofTi laI-RC theM ed.
c-t 303aw Aaa ly;g db U( oU I-H -s
r, alternatively, blunt ends that fail to properly load telorase[25]. Consistent with this possibility, bothmre11mecnd tel1mec1mutations confer senescence and ultimand-to-end fusions and gross chromosomal rearrange
20,26]. These data also suggest that this function of Mres mediated through Tel1p.
Mre11p also functions as a downstream nuclease acnduced by the Tel1p and Mec1p pathways[27]. Mre11p ha
ultiple nuclease activities including 3′–5′ ss- and dsDNAxonuclease and ssDNA endonuclease activity in vitro. Hver, none of these activities can explain the MRX depen′ overhangs observed in vivo at meiotic and mitotic induNA double strand breaks (DSBs)[28,29]. In vitro, Rad50pelicase activity is necessary for 5′–3′ processing and, in co
unction with Mre11p ssDNA endonuclease activity, mayesponsible for 3′ overhang formation (reviewed in ref.[27]).re11p is redundant with at least one 5′–3′ nuclease, Exo1p
n most phases of the cell cycle[30–34]. However, in G2rrested cells, Mre11p is specifically required for 5′ resectionbserved at HO-induced telomere-proximal DSBs that isssary for telomerase-mediated de novo telomere syn
35]. These data suggest that Mre11 may act as a telouclease in G2 phase. While Mre11p is required for geting short 3′ telomere overhangs that are observed in G′
ail formation in late S phase that is necessary for telomectivity does not require Mre11p[36]. However, the interelationship of nuclease activity, telomere binding, and Damage signaling remain complex.
In vertebrates, the MRE11–RAD50–NBS1 (MRN) colex (NBS1 is the human ortholog of Xrs2p) localizes
sed in this study are isogenic to W303a (Mat a leu2-3, 112IS3 ade2-1 trp1 ura3-1).A yeast integrating vector carrying themre11P162S
llele [49–51] was constructed by ligation of the BamHmaI fragment of YCpM11(ts) (courtesy of Mahmohamankhah) into pRS304[52]. Integrating vectors contai
ng themre11D56N [53] andmre11H125N [33,53] allelesere constructed by insertion of the KpnI-SacI fragmf pSM444 and pSM438, respectively (courtesy of Lorraymington), into pRS304. A similar vector carryingre11�410-420[54] allele was constructed by insertion
he BamHI-XhoI fragment of pHO115 (courtesy of Hideygawa) into pRS304. These plasmids were used fortep gene replacement in W303a with counterselectioRP1by 5-FAA (5-flouroanthranilic acid)[55]. A �560-692
ntegrating vector was constructed by insertion of the CsaI fragment of pGADM11 (1-559)[56] (gift of Mahmoodhamankhah) into pRS304, followed by integration atRE11locus so that only the�560-692 allele was expressmre11D16Aandmre11�644-692 strains were constru
ed by conventional one-step gene replacement in With the BamHI-HindIII fragments of pTZgm11-D16nd pTZgm11-�C49, respectively (mre11D16A::URA3nd mre11�644-692::URA3 in pTZ19R, respectiveifts of Munenori Furuse).mre11H213Y was constructey integration of the BamHI fragment of pTAK-152mre11H213Y::URA3 in pUC18; courtesy of Takehiksui).mre11�A�B was made by replacement of the PflMindIII fragment of strainmre11�410-420 with the correponding fragment of pTZgm11�C49.
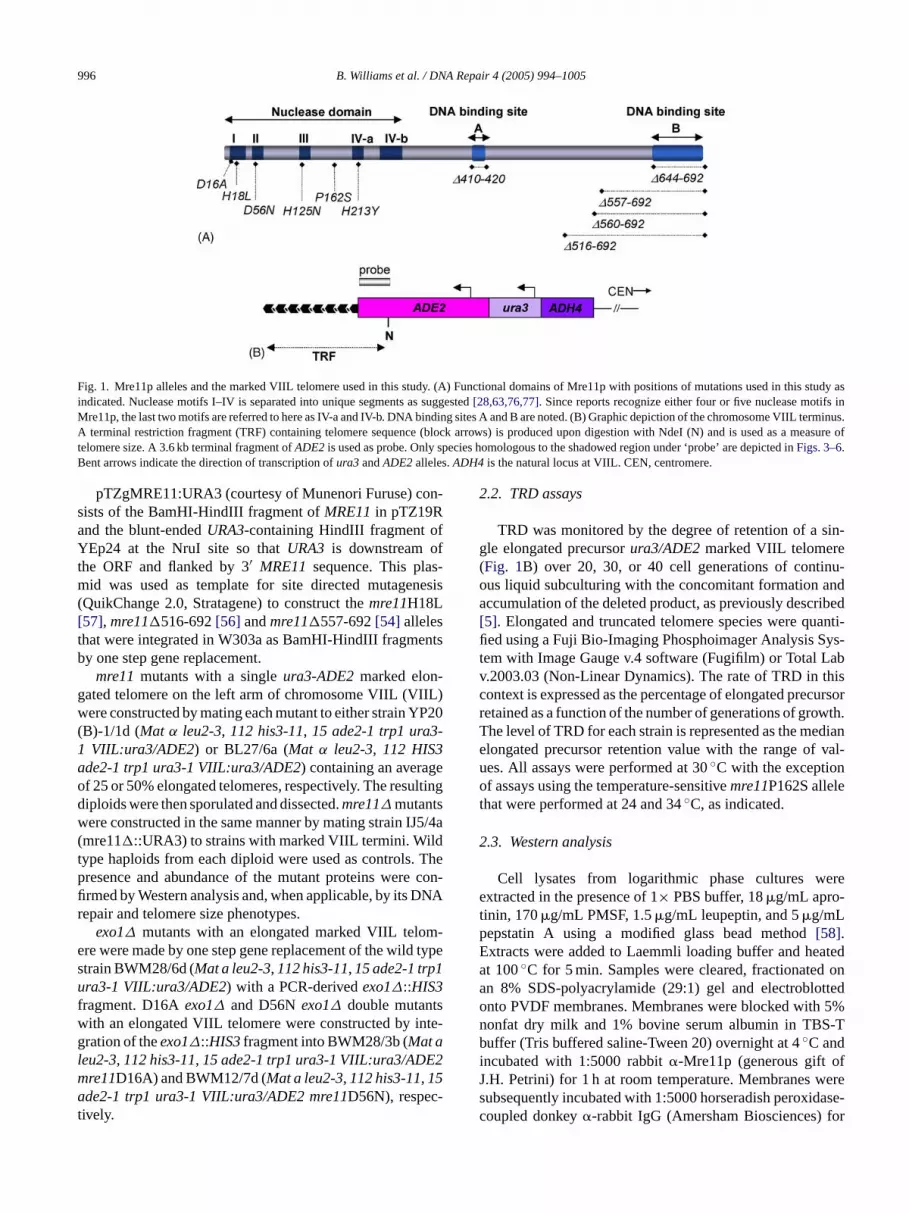
996 B. Williams et al. / DNA Repair 4 (2005) 994–1005
Fig. 1. Mre11p alleles and the marked VIIL telomere used in this study. (A) Functional domains of Mre11p with positions of mutations used in this study asindicated. Nuclease motifs I–IV is separated into unique segments as suggested[28,63,76,77]. Since reports recognize either four or five nuclease motifs inMre11p, the last two motifs are referred to here as IV-a and IV-b. DNA binding sites A and B are noted. (B) Graphic depiction of the chromosome VIIL terminus.A terminal restriction fragment (TRF) containing telomere sequence (block arrows) is produced upon digestion with NdeI (N) and is used as a measure oftelomere size. A 3.6 kb terminal fragment ofADE2is used as probe. Only species homologous to the shadowed region under ‘probe’ are depicted inFigs. 3–6.Bent arrows indicate the direction of transcription ofura3andADE2alleles.ADH4 is the natural locus at VIIL. CEN, centromere.
pTZgMRE11:URA3 (courtesy of Munenori Furuse) con-sists of the BamHI-HindIII fragment ofMRE11in pTZ19Rand the blunt-endedURA3-containing HindIII fragment ofYEp24 at the NruI site so thatURA3 is downstream ofthe ORF and flanked by 3′ MRE11 sequence. This plas-mid was used as template for site directed mutagenesis(QuikChange 2.0, Stratagene) to construct themre11H18L[57],mre11�516-692[56] andmre11�557-692[54] allelesthat were integrated in W303a as BamHI-HindIII fragmentsby one step gene replacement.mre11mutants with a singleura3-ADE2marked elon-
gated telomere on the left arm of chromosome VIIL (VIIL)were constructed by mating each mutant to either strain YP20(B)-1/1d (Mat α leu2-3, 112 his3-11, 15 ade2-1 trp1 ura3-1 VIIL:ura3/ADE2) or BL27/6a (Mat α leu2-3, 112 HIS3ade2-1 trp1 ura3-1 VIIL:ura3/ADE2) containing an averageof 25 or 50% elongated telomeres, respectively. The resultingdiploids were then sporulated and dissected.mre11∆ mutantswere constructed in the same manner by mating strain IJ5/4a(mre11�::URA3) to strains with marked VIIL termini. Wildtype haploids from each diploid were used as controls. Thepresence and abundance of the mutant proteins were con-firmed by Western analysis and, when applicable, by its DNArepair and telomere size phenotypes.exo1∆ mutants with an elongated marked VIIL telom-
ere were made by one step gene replacement of the wild typesuf sw nte-gl 2ma -t
2.2. TRD assays
TRD was monitored by the degree of retention of a sin-gle elongated precursorura3/ADE2marked VIIL telomere(Fig. 1B) over 20, 30, or 40 cell generations of continu-ous liquid subculturing with the concomitant formation andaccumulation of the deleted product, as previously described[5]. Elongated and truncated telomere species were quanti-fied using a Fuji Bio-Imaging Phosphoimager Analysis Sys-tem with Image Gauge v.4 software (Fugifilm) or Total Labv.2003.03 (Non-Linear Dynamics). The rate of TRD in thiscontext is expressed as the percentage of elongated precursorretained as a function of the number of generations of growth.The level of TRD for each strain is represented as the medianelongated precursor retention value with the range of val-ues. All assays were performed at 30◦C with the exceptionof assays using the temperature-sensitivemre11P162S allelethat were performed at 24 and 34◦C, as indicated.
2.3. Western analysis
Cell lysates from logarithmic phase cultures wereextracted in the presence of 1× PBS buffer, 18�g/mL apro-tinin, 170�g/mL PMSF, 1.5�g/mL leupeptin, and 5�g/mLpepstatin A using a modified glass bead method[58].Extracts were added to Laemmli loading buffer and heateda ona ttedo 5%n -Tbi fJ eres dase-c for
train BWM28/6d (Mat a leu2-3,112 his3-11,15 ade2-1 trp1ra3-1 VIIL:ura3/ADE2) with a PCR-derivedexo1∆::HIS3
ragment. D16Aexo1∆ and D56Nexo1∆ double mutantith an elongated VIIL telomere were constructed by iration of theexo1∆::HIS3fragment into BWM28/3b (Mat aeu2-3, 112 his3-11, 15 ade2-1 trp1 ura3-1 VIIL:ura3/ADEre11D16A) and BWM12/7d (Mat a leu2-3,112 his3-11,15de2-1 trp1 ura3-1 VIIL:ura3/ADE2 mre11D56N), respec
ively.
t 100◦C for 5 min. Samples were cleared, fractionatedn 8% SDS-polyacrylamide (29:1) gel and electroblonto PVDF membranes. Membranes were blocked withonfat dry milk and 1% bovine serum albumin in TBSuffer (Tris buffered saline-Tween 20) overnight at 4◦C and
ncubated with 1:5000 rabbit�-Mre11p (generous gift o.H. Petrini) for 1 h at room temperature. Membranes wubsequently incubated with 1:5000 horseradish peroxioupled donkey�-rabbit IgG (Amersham Biosciences)

B. Williams et al. / DNA Repair 4 (2005) 994–1005 997
1 h at room temperature. Antibody-protein complexes weredetected using the ECL Western blotting detection kit (Amer-sham Biosciences).
The abundance of each Mre11p allele was normalized tothe abundance of Sir2p. To this end, PVDF membranes werestripped of bound antibodies by incubation in 100 mM�-mercaptoethanol, 2% SDS, and 63 mM Tris–HCl, pH 6.7 for1 h at 50◦C. Membranes were then incubated 1 h at roomtemperature with 1:1000 rabbit�-Sir2p (provided by SusanGasser), followed by incubation with 1:7000 horseradishperoxidase-coupled donkey�-rabbit IgG and ECL detec-tion. Protein abundance was quantified using TotalLabv.2003.03.
2.4. Statistical analysis
The Wilcoxon–Mann–Whitney Rank Sum Test was usedto determine the degree of overlap between distributionsof wild type and mutant elongated precursor retention val-ues using KaleidaGraph 3.6 (KaleidaGraph Inc.). Signifi-cance was determined byP-values≤ 0.05. This test wasalso used to determine the significance of the difference intelomere heterogeneity between wild type andmre11mutantalleles.
3
3
os-p cle-aM -s ndu ctingi ty ofM ye fm pro-c ucedm tsa toticD
arei em hodie I),Hl tentwo t toDt int ug-g ysis
[63,64]. All of the nuclease-deficient alleles exhibit DNArepair defects. In contrast, telomere shortening is variableand allele-dependent (Fig. 2A), as previously observed[28,33,53,54,56,57,61,62].
TRD assays were conducted in strains carrying the D16A,H18L, D56N, and H125N alleles and a single elongatedura3-ADE2-marked VIIL telomere (Fig. 1B). Each allele wasproduced at levels within approximately two-fold of wildtype (Fig. 2B) and abundance did not correlate with the TRDphenotype (Fig. 2B). Telomeres were analyzed by South-ern hybridization and TRD was quantified by the fractionalretention of the elongated precursor[5] (Fig. 3; Table 1).Strikingly, the retention of precursor of each of the fouralleles after 20, 30, or 40 generations of liquid subcultur-ing did not differ significantly from wild type (Table 1). Ineach case, TRD precision was maintained, with elongatedtelomeres returning to the size of the majority of telomeres(Figs. 2A and 3). These results suggest the Mre11p nucle-ase domain does not play an essential role in TRD rate orprecision.
The H213Y allele displayed a unique phenotype. Thisallele conferred a rapid initial increase in TRD rate that con-verted 60% of the precursor to the deleted product (Fig. 3),significantly greater than wild type initial rates (P< 0.001,Table 1). In addition, the presence of deleted telomeres withheterogeneous size distributions is present in higher fre-q to ar isedi
3
hinM on-s uesa g ofa -t ughe fe inv
ticD itroi tion-sa them pre-c farg ns ofl
yl-t 4-6 ivitya -t n
. Results
.1. Analysis of nuclease motifs
Mre11p is characterized by four highly conserved phhodiesterase motifs that are required for the multiple nuse activities observed in vitro (Fig. 1A, [23]). Althoughre11p exonuclease processing in vitro (3′–5′) is the oppo
ite polarity of 5′–3′ resections required in vivo, stranwinding by a Rad50p-associated helicase activity a
n concert with the single-stranded endonuclease activire11p is able to produce 5′ extensions in vitro and ma
xplain the apparent polarity in vivo[59,60]. The presence outations in the nuclease domain abrogates nucleolytic
essing in vitro and processing at spontaneous and indeiotic DSBs in vivo[28,53,54,57,61]. Nuclease mutanlso display a partial loss of nucleolytic processing at miSBs[62].To test whether the Mre11p nuclease activities
nvolved in TRD, we analyzed TRD inmre11missensutants of conserved residues in each nuclease phospsterase motif: D16A and H18L (motif I), D56N (motif I125N (motif III) and H213Y (motif IV-A) (Fig. 1A). The
oss of in vitro nuclease activity in these alleles is consisith structural data from thePyrococcus furiosisMre11prtholog. In this organism, residues spatially equivalen16, H18, D56, and H213 coordinate Mn2+ ions within
he active site[63]. In contrast, the H125 equivalenthe P. furiosis enzyme has an active site geometry sesting involvement in phosphodiester bond hydrol
-
uency than wild type (P= 0.009,Fig. 3,Table 2), reminiscenf the behavior ofykumutations in TRD[65]. These dataise the possibility that telomere protection is compromn this allele.
.2. Mre11p DNA binding domains act to repress TRD
Two DNA binding domains have been identified witre11p: DNA binding site A (amino acids 407–421) c
isting of a cluster of positively charged amino acid residnd DNA binding site B (amino acids 644–692) consistincluster of negatively charged residues[54,61]. This charac
eristic clustering of charged residues is maintained throvolution despite divergence in sequence[61]. Absence oither DNA binding site leads to reduced DNA bindingitro [54,61].
Deletion of site A confers a specific deficiency in meioSB processing[54]. Nuclease activity as assayed in v
s also abrogated in this allele, suggesting a close relahip between DNA binding and nucleolytic activity[54]. Tossay the involvement of DNA binding site A in TRD,re11�410-420 allele was assayed for TRD rate and
ision. Cells lacking the site display a rate of deletionreater than the wild type rate after 30 and 40 generatio
iquid subculture (P< 0.001) (Fig. 4; Table 1).A similar analysis was carried out for the carbox
erminal DNA binding site B of Mre11p (amino acids 6492). While deletion of this region retains nuclease actnd MMS-induced resistance[61],mre11�644-692 is defec
ive in meiotic DSB formation[61]. This allele also confers a
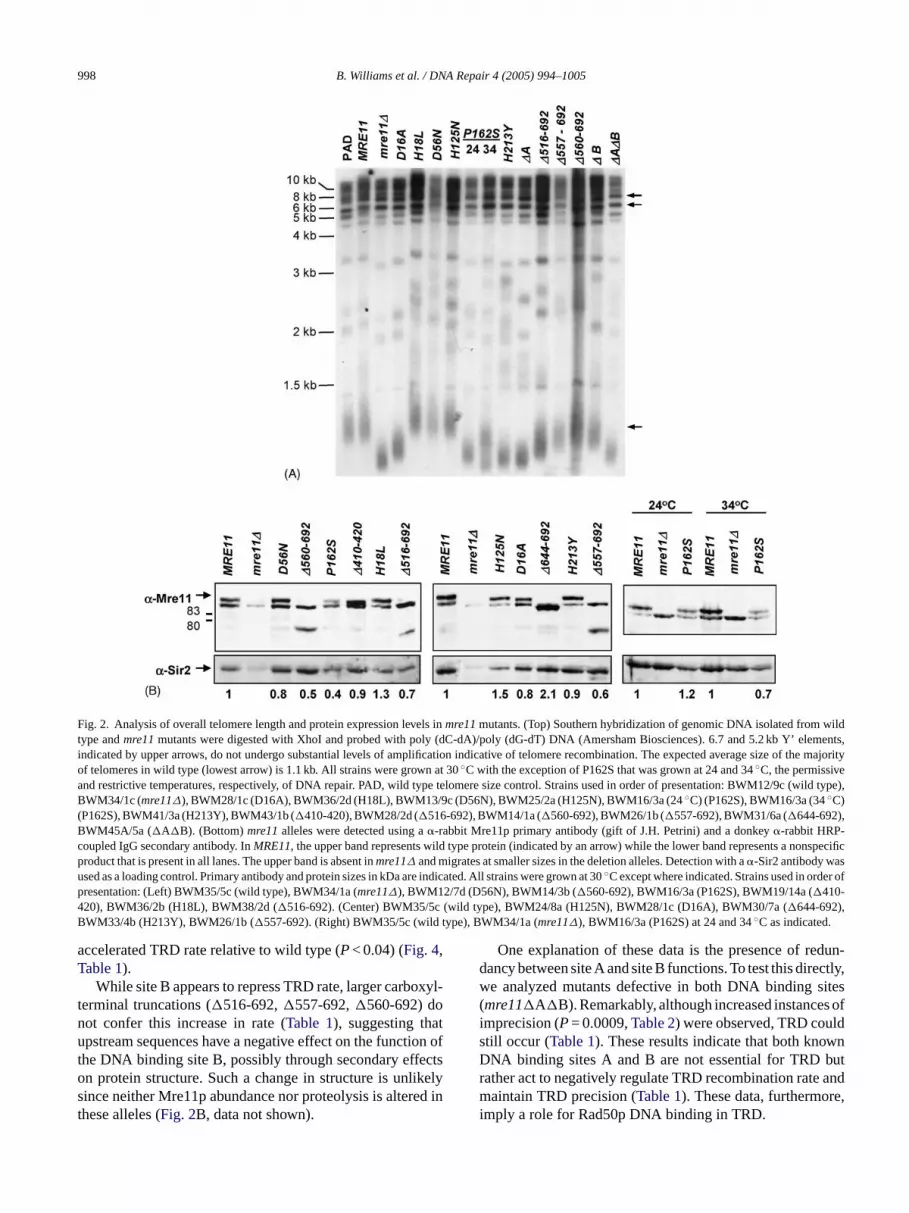
998 B. Williams et al. / DNA Repair 4 (2005) 994–1005
Fig. 2. Analysis of overall telomere length and protein expression levels inmre11mutants. (Top) Southern hybridization of genomic DNA isolated from wildtype andmre11mutants were digested with XhoI and probed with poly (dC-dA)/poly (dG-dT) DNA (Amersham Biosciences). 6.7 and 5.2 kb Y’ elements,indicated by upper arrows, do not undergo substantial levels of amplification indicative of telomere recombination. The expected average size of themajorityof telomeres in wild type (lowest arrow) is 1.1 kb. All strains were grown at 30◦C with the exception of P162S that was grown at 24 and 34◦C, the permissiveand restrictive temperatures, respectively, of DNA repair. PAD, wild type telomere size control. Strains used in order of presentation: BWM12/9c (wild type),BWM34/1c (mre11∆), BWM28/1c (D16A), BWM36/2d (H18L), BWM13/9c (D56N), BWM25/2a (H125N), BWM16/3a (24◦C) (P162S), BWM16/3a (34◦C)(P162S), BWM41/3a (H213Y), BWM43/1b (�410-420), BWM28/2d (�516-692), BWM14/1a (�560-692), BWM26/1b (�557-692), BWM31/6a (�644-692),BWM45A/5a (�A�B). (Bottom)mre11alleles were detected using a�-rabbit Mre11p primary antibody (gift of J.H. Petrini) and a donkey�-rabbit HRP-coupled IgG secondary antibody. InMRE11, the upper band represents wild type protein (indicated by an arrow) while the lower band represents a nonspecificproduct that is present in all lanes. The upper band is absent inmre11∆ and migrates at smaller sizes in the deletion alleles. Detection with a�-Sir2 antibody wasused as a loading control. Primary antibody and protein sizes in kDa are indicated. All strains were grown at 30◦C except where indicated. Strains used in order ofpresentation: (Left) BWM35/5c (wild type), BWM34/1a (mre11∆), BWM12/7d (D56N), BWM14/3b (�560-692), BWM16/3a (P162S), BWM19/14a (�410-420), BWM36/2b (H18L), BWM38/2d (�516-692). (Center) BWM35/5c (wild type), BWM24/8a (H125N), BWM28/1c (D16A), BWM30/7a (�644-692),BWM33/4b (H213Y), BWM26/1b (�557-692). (Right) BWM35/5c (wild type), BWM34/1a (mre11∆), BWM16/3a (P162S) at 24 and 34◦C as indicated.
accelerated TRD rate relative to wild type (P< 0.04) (Fig. 4,Table 1).
While site B appears to repress TRD rate, larger carboxyl-terminal truncations (�516-692,�557-692,�560-692) donot confer this increase in rate (Table 1), suggesting thatupstream sequences have a negative effect on the function ofthe DNA binding site B, possibly through secondary effectson protein structure. Such a change in structure is unlikelysince neither Mre11p abundance nor proteolysis is altered inthese alleles (Fig. 2B, data not shown).
One explanation of these data is the presence of redun-dancy between site A and site B functions. To test this directly,we analyzed mutants defective in both DNA binding sites(mre11�A�B). Remarkably, although increased instances ofimprecision (P= 0.0009,Table 2) were observed, TRD couldstill occur (Table 1). These results indicate that both knownDNA binding sites A and B are not essential for TRD butrather act to negatively regulate TRD recombination rate andmaintain TRD precision (Table 1). These data, furthermore,imply a role for Rad50p DNA binding in TRD.
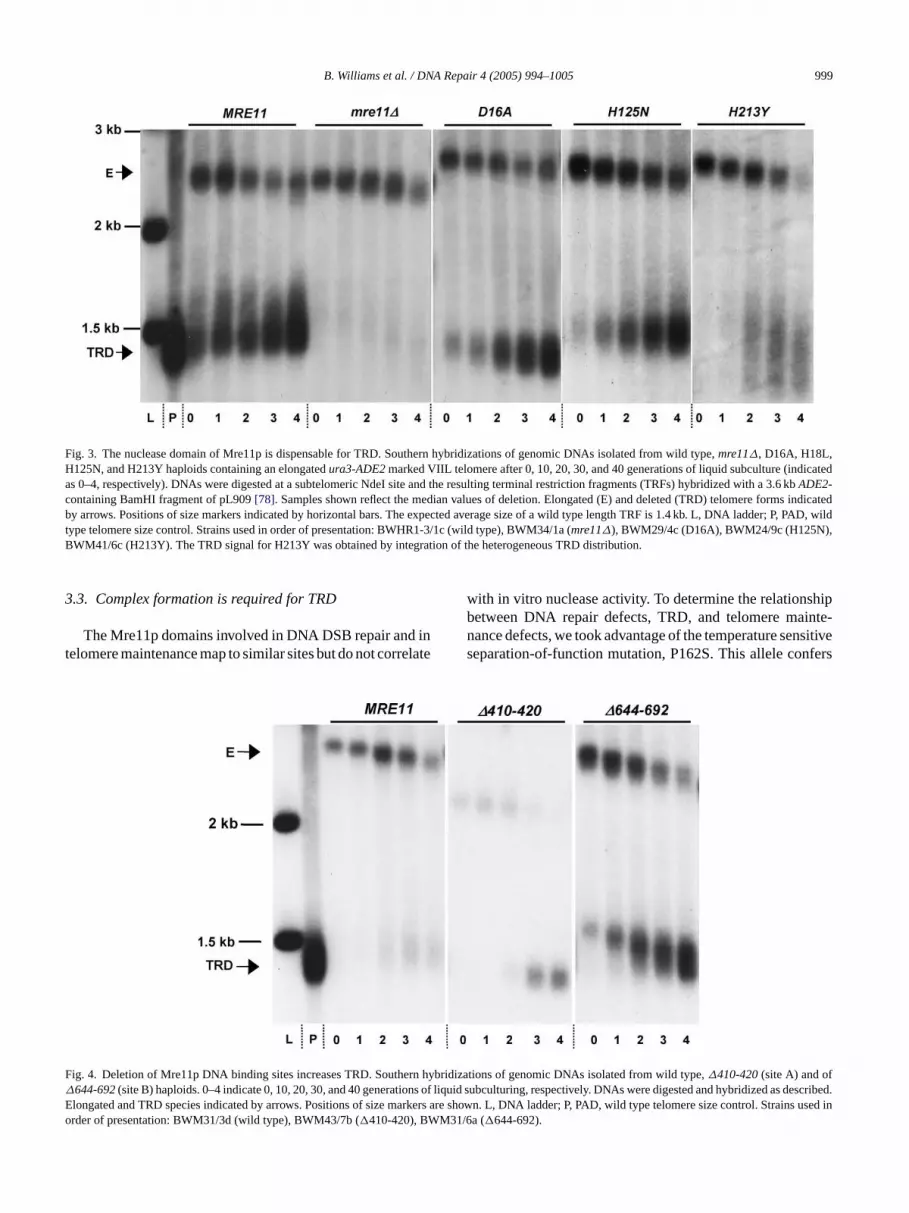
B. Williams et al. / DNA Repair 4 (2005) 994–1005 999
Fig. 3. The nuclease domain of Mre11p is dispensable for TRD. Southern hybridizations of genomic DNAs isolated from wild type,mre11∆, D16A, H18L,H125N, and H213Y haploids containing an elongatedura3-ADE2marked VIIL telomere after 0, 10, 20, 30, and 40 generations of liquid subculture (indicatedas 0–4, respectively). DNAs were digested at a subtelomeric NdeI site and the resulting terminal restriction fragments (TRFs) hybridized with a 3.6 kbADE2-containing BamHI fragment of pL909[78]. Samples shown reflect the median values of deletion. Elongated (E) and deleted (TRD) telomere forms indicatedby arrows. Positions of size markers indicated by horizontal bars. The expected average size of a wild type length TRF is 1.4 kb. L, DNA ladder; P, PAD, wildtype telomere size control. Strains used in order of presentation: BWHR1-3/1c (wild type), BWM34/1a (mre11∆), BWM29/4c (D16A), BWM24/9c (H125N),BWM41/6c (H213Y). The TRD signal for H213Y was obtained by integration of the heterogeneous TRD distribution.
3.3. Complex formation is required for TRD
The Mre11p domains involved in DNA DSB repair and intelomere maintenance map to similar sites but do not correlate
with in vitro nuclease activity. To determine the relationshipbetween DNA repair defects, TRD, and telomere mainte-nance defects, we took advantage of the temperature sensitiveseparation-of-function mutation, P162S. This allele confers
F rn hyb f∆ ns of li describeE rkers a ains useo WM31
ig. 4. Deletion of Mre11p DNA binding sites increases TRD. Southe644-692(site B) haploids. 0–4 indicate 0, 10, 20, 30, and 40 generatiolongated and TRD species indicated by arrows. Positions of size marder of presentation: BWM31/3d (wild type), BWM43/7b (�410-420), B
ridizations of genomic DNAs isolated from wild type,∆410-420(site A) and oquid subculturing, respectively. DNAs were digested and hybridized asd.re shown. L, DNA ladder; P, PAD, wild type telomere size control. Strd in
/6a (�644-692).

1000 B. Williams et al. / DNA Repair 4 (2005) 994–1005
Table 1Elongated telomere medians ofmre11mutants
Strain Mediana elongated VIIL precursor retention values (%)b nc
20 generations 30 generations 40 generations
MRE11d 80 68 48 75[43–100]e [26–97] [10–78]
mre11�f >90 (0.002)g >80 (0.0009) >72 (<0.0001) 28[86–97] [75–87] [66–87]
D16A 83 75 63 12[44–96] [29–85] [16–73]
H18L 70 58 44 13[47–100] [38–94] [25–69]
D56N 75 53 44 9[46–85] [47–77] [30–55]
H125N 71 52 42 13[43–95] [38–91] [25–80]
P162S >94 (0.014) >86 (0.0007) >82 (0.0002) 18[75–97] [59–99] [57–92]
H213Y 45 (<0.0001) 30 (<0.0001) 22 (0.002) 11[23–73] [11–64] [9–60]
�410-420 67 26(<0.0001) 8 (<0.0001) 11[7–97] [3–93] [1–58]
∆516-692 86 62 58 10[61–91] [47–75] [40–69]
∆557-692 70 55 41 10[40–99] [39–87] [29–81]
∆560-692 80 63 44 14[50–90] [32–82] [26–81]
�644-692 67 (0.037) 51 (0.025) 37 (0.032) 16[51–85] [37–76] [26–53]
�A �B 49 (<0.0001) 34 (0.0009) 8 (0.001) 9[31–68] [9–97] [1–62]
a The median of each distribution is indicated in bold.b Elongated VIIL precursor retention expressed as the ratio of elongated VIIL precursor retained after 20, 30, or 40 generations of growth in liquid culture
divided by the ratio of elongated VIIL precursor present in the initial pre-culture multiplied by 100.c n refers to sample size.d The distributions of elongated VIIL precursor retention values from wild type strains with 12.5 and 25% elongated telomeres on average were compared
using the Wilcoxon–Mann–Whitney Rank Sum test. The distributions were determined not to be significantly different from each other; therefore, the retentionvalues of strains with 12.5 and 25% elongated telomeres on average were pooled, when applicable, for each allele.
e The range of TRD values in each distribution is indicated in brackets.f Values represent under-estimates of median retention rates in these alleles since TRD product formation sharply decreases in the majority of samples and
ambiguous species not representative of TRD are produced.g P-values of each mutant distribution (indicated in parenthesis) for samples significantly deviating from wild type was determined using the
Wilcoxon–Mann–Whitney Rank Sum test. Mutants exhibiting a significant change are in bold.
near-wild-type levels of MMS resistance at the restrictivetemperature but shortened telomeres and disrupted homod-imerization and Rad50p interaction at both permissive andrestrictive temperatures[49–51]. Interestingly, we found thatthe TRD rate is reduced tomre11� levels inmre11P162Scells at both permissive and restrictive temperatures (Fig. 5;Table 1). Since other alleles that cause partial DNA repairdefects and defective telomere maintenance do not affectTRD rate or precision (i.e., H18L, D16A, D56N, H125N),the P162S data is most consistent with distinct roles for com-
plex formation and Mre11p function within the context of thecomplex.
3.4. Double mutants in the EXO1 and MRE11 genessuggest the presence of a novel nuclease involved in TRD
Yeast Exonuclease 1 has been shown in several contextsto act redundantly with Mre11p[66]. To test whether Exo1pnuclease activity is required for TRD, we analyzed loss ofExo1p in D16A and D56N nuclease-deficient alleles that

B. Williams et al. / DNA Repair 4 (2005) 994–1005 1001
Fig. 5. MRX complex formation is required for TRD. (A) Southern hybridizations of genomic DNAs isolated from wild type and P162S haploids containingan elongated marked VIIL terminus. 0–4 indicate 0, 10, 20, 30, and 40 generations of liquid subculturing, respectively. DNAs were digested and Southernshybridized as described for other mutants. Elongated and TRD species indicated by arrows. Positions of size markers as indicated. L, DNA ladder; P, PAD,wild type telomere size control. Strains used in order of presentation: BWM17/1d (wild type), BWM17/2c (P162S), BWM17/4a (P162S).
were proficient in TRD. These alleles were chosen since theyappear to fall into two classes of nuclease deficient alleleswith D16A displaying the most severe DNA damage pheno-types. Recent studies have demonstrated that different classesof DSB-mediated recombination are influenced in these twoalleles, even though the molecular basis remains unknown[67].
We observed no effect of Exo1p loss on TRD accumu-lation in D16A cells (Fig. 6) indicating that a nuclease in
Table 2Significance of heterogeneous TRD product distributions inmre11mutants
Allele Heterogeneous TRD product distributions
Median sizerangea (bp)
Size rangesb
(bp)P-valuesc nd
MRE11 350 [200–480] 20D16A 420 [380–560] 0.0343 3H18L 320 [270–370] 2H125N 360 [250–470] 2H213Y 440 [280–550] 0.0096 13�410-420 370 [200–530] 18�516-692 320 [260–520] 8�557-692 280 NA 1�560-692 330 [280–380] 4�644-692 360 [320–500] 7�A �B 470 [410–580] 0.0009 6
a The typical size range of TRD products in a single sample is∼150 bp.s was
d nted inb
t, thed e dis-ts utantst
addition to Exo1p or Mre11p is likely to play a role inTRD. Curiously, D56Nexo1∆ mutants display a repro-ducible biphasic pattern with repressed TRD followed bya massive accumulation after three rounds of subculturing(Fig. 6), suggesting that under some circumstances Exo1pmay play a direct role in TRD. Whether this is related tothe different repair activities inmre11D16Aandmre11D56Nmutants remains unknown.
4. Discussion
4.1. The nuclease activity of Mre11p is dispensable forTRD
The mutation of residues essential for Mre11p nucleaseactivity maintained or increased the rate of TRD, indicatingthat Mre11p nucleolytic terminal processing is dispensablefor TRD. This is a somewhat unexpected result given thatMre11p encodes multiple nuclease activities that may be dif-ferentially altered by each allele. H213Y is unique in that anincreased, but imprecise, spontaneous rate of mitotic recom-bination is observed in the context of themre11H213Y/L225Idouble mutant (mre11-58) [28], consistent with the acceler-ated rates of TRD in the single mutant allele. The mechanismof action of this stimulation remains unknown.
dox.Aa da e,E -t ction[ iono
b The size range of heterogeneous TRD products at 40 generationetermined. The most narrow and most broad size ranges are represerackets.c To determine if the degree of TRD heterogeneity was significanistribution of size ranges for each allele was compared to the wild typ
ribution using the Wilcoxon–Mann–Whitney Rank Sum test.P-values whenignificant as determined from each test is presented with values. Mhat result in a significant result are shown in bold.d n, number of samples with heterogeneous TRD products.
The role of Mre11p nuclease activity remains a paras noted, Mre11p does not contain an intrinsic 5′–3′ nucleasectivity that is required to form the 3′ overhang at HO-inducend meiotic DSBs in vivo[68,69]. An alternative nucleasxo1p, has the polarity expected for 3′ overhang produc
ion and appears to play a major role in telomere rese32–34]. InS. pombe, Exo1p is responsible for the generatf the constitutive elongated telomeric ssDNA 3′ overhangs in

1002 B. Williams et al. / DNA Repair 4 (2005) 994–1005
Fig. 6. Exo1p is not required for TRD in Mre11p nuclease deficient strains. (A) Genomic DNAs were isolated frommre11∆, exo1∆, D16Aexo1∆, and D56Nexo1∆ haploids containing an elongated marked VIIL terminus. DNA samples representing 0, 10, 20, and 30 generations of liquid subculturing (indicatedas 0–3, respectively) are shown. DNA isolations and Southern hybridizations were performed as described. Elongated (E) and TRD species are indicated.Strains used in order of presentation: BWM34/3c (mre11∆), MBE1-4 (exo1∆), MBE2-3 (D16Aexo1∆), MBE3-1 (D56Nexo1∆). Median elongated precursorretention over 20, 30, and 40 generations of growth were determined as:exo1∆-100, 100, 86%; D16Aexo1∆-40, 23, 14%; D56Nexo1∆-90, 69, 53%.
yku70cells at the restrictive temperature and incdc13-1cells,independent of Mre11p[70]. Similarly,S. pombetelomeric3′ overhangs are still formed in the nuclease deficientrad32-D25A allele, corresponding tomre11D16A, in the absenceof the major telomere binding protein Taz1[70]. Nonethe-less, the shorter single stranded telomeric overhangs that areformed in G1 and early S phases ofS. cerevisiaeare Mre11p-dependent[36]. Our data suggest that Exo1p nuclease activityis also not required for TRD; therefore, it is likely that othernucleases redundant with Exo1p are involved. However, weshould not rule out a role for Exo1p in differentmre11alle-les. Other exonucleases, such as the postulated ExoXp, havealso been proposed to be involved in 3′ overhang forma-tion at the telomere and may be responsible for Mre11p andExo1p nuclease independent TRD[71,72]. Our results do notconflict with data demonstrating that MRX-dependent pro-cessing is required for de novo telomere synthesis at telomericHO-induced DSBs in G2/M[35] because it remains unclearwhether this represents a signaling or nucleolytic activity.
4.2. The DNA binding domains of Mre11p repress TRD
Our results indicate that both DNA binding domains ofMre11p (sites A and B) are functional repressors of TRD. Wefound that deletion of either DNA binding site substantiallyincreases the TRD rate relative to wild type. The specificityo rgerC phe-n
ings thed leteds dings ing
domains could normally protect the termini against aberrantdeletion.
Three possible mechanisms may explain these data. First,residual DNA binding may be present at a hitherto uncharac-terized site. Second, DNA binding could be mediated throughanother member of the MRX complex, such as the well char-acterized Rad50p DNA binding domain[63,73]. It wouldnot be surprising to find the behavior of TRD to be alteredunder such conditions through altered precision of strandinvasion and/or resolution. Interestingly, H213Y, anotherallele exhibiting greater TRD product heterogeneity, has beenpreviously associated with compromised association withRad50p[56]. Third, deletion of the DNA binding domain(s)may produce a unique complex with distinct properties thatinfluence TRD, e.g. by increasing small tract homologousrecombination[74,75]. Finally, the heretical viewpoint thatDNA binding is not required for TRD, albeit unlikely, wouldsuggest recruitment of the MRX complex by other telomerebinding proteins. We cannot formally rule out this possibilityat the present time.
4.3. Does the MRX complex form the ‘matrix’ forMre11p function in TRD?
Our results bear on the issue of relative roles of Mre11pand the MRX complex in TRD. The absence of TRD activityi plexa ega-t mt lf is ap elem lit-t thec
f this process is further underscored by the failure of la-terminal deletions to recapitulate the site B deletionotype.
Remarkably, the loss of both known sites of DNA bindtill does not abrogate TRD. One notable difference inouble mutant is in the degree of heterogeneity of depecies, suggesting that loss of both Mre11p DNA binites may alter TRD function. In this sense, the DNA bind
n P162S is likely to be mediated through loss of comssociation. In addition, the wide range of positive and n
ive phenotypes inmre11alleles that substantially differ frohe null phenotype suggests that complex formation itsererequisite for Mre11p function. In this view, the null allay be an indication of a loss of the complex, but say
le about the function of Mre11p when present withinomplex.

B. Williams et al. / DNA Repair 4 (2005) 994–1005 1003
Hence, the MRX complex is likely to be essential at leastin part by providing a matrix for specific positive and neg-ative regulation by Mre11p and Rad50p in TRD that wehave revealed using themre11missense and deletion alle-les. Regardless of the specific mechanism, the dissection ofMre11p has already led to unique insights into its role withinthe MRX complex that was not anticipated by the phenotypeof the null allele.
Acknowledgments
We thank the following people for plasmids used in thisstudy: Lorraine Symington, Hideyuki Ogawa, MahmoodChamankhah, Munenori Furuse, and Takehiko Usui. Wethank John Petrini for his gift of�-Mre11 antibody and JamesHaber for a yeast strain harboring theexo1∆::HIS3 allele.We also thank E.B. Hoffman, J. Frederick, and S. Klotzerfor their continued advice. This work was supported by NIHgrant GM-069943 (to AJL), NSF grant MCB-0084460 (toAJL), NRSA award GM-020603 (to BW) and a Tulane Can-cer Center Matching Funds grant.
References
. 65
e 21
tion003)
l in
m--
eretion,151
ng of
com-, Cell
from923.
[ chi,plex
[ in acad.
[ ops
[ az1and
72.[ Rev.
[15] L. Tomaska, M.J. McEachern, J. Nosek, Alternatives to telomerase:keeping linear chromosomes via telomeric circles, FEBS Lett. 567(2004) 142–146.
[16] A.J. Cesare, J.D. Griffith, Telomeric DNA in ALT cells is character-ized by free telomeric circles and heterogeneous t-loops, Mol. CellBiol. 24 (2004) 9948–9957.
[17] J. Nosek, A. Rycovska, A.M. Makhov, J.D. Griffith, L. Tomaska,Amplification of telomeric arrays via rolling-circle mechanism, J.Biol. Chem. 280 (2005) 10840–10845.
[18] S.J. Boulton, S.P. Jackson, Components of the Ku-dependentnon-homologous end-joining pathway are involved in telomericlength maintenance and telomeric silencing, EMBO J. 17 (1998)1819–1828.
[19] C.I. Nugent, G. Bosco, L.O. Ross, S.K. Evans, A.P. Salinger, J.K.Moore, J.E. Haber, V. Lundblad, Telomere maintenance is dependenton activities required for end repair of double-strand breaks, Curr.Biol. 8 (1998) 657–660.
[20] K.B. Ritchie, T.D. Petes, The Mre11p/Rad50p/Xrs2p complex andthe Tel1p function in a single pathway for telomere maintenance inyeast, Genetics 155 (2000) 475–479.
[21] Q. Chen, A. Ijpma, C.W. Greider, Two survivor pathways that allowgrowth in the absence of telomerase are generated by distinct telom-ere recombination events, Mol. Cell Biol. 21 (2001) 1819–1827.
[22] H. Takata, Y. Kanoh, N. Gunge, K. Shirahige, A. Matsuura, Recipro-cal association of the budding yeast ATM-related proteins Tel1 andMec1 with telomeres in vivo, Mol. Cell 14 (2004) 515–522.
[23] D. D’Amours, S.P. Jackson, The Mre11 complex: at the crossroadsof DNA repair and checkpoint signalling, Nat. Rev. Mol. Cell Biol.3 (2002) 317–327.
[24] T. Naito, A. Matsuura, F. Ishikawa, Circular chromosome formationin a fission yeast mutant defective in two ATM homologues, Nat.
[ 11--
[ ousns in
[ re11tancest.)
[ ess-osis,
[ tionshing5.
[ iredaks,
[ e11aling
[ if-d50,nent
[ s ofin
[ le-e
[ forCurr.
[1] C.W. Greider, Telomere length regulation, Annu. Rev. Biochem(1996) 337–365.
[2] T. de Lange, Protection of mammalian telomeres, Oncogen(2002) 532–540.
[3] D.M. Feldser, J.A. Hackett, C.W. Greider, Telomere dysfuncand the initiation of genome instability, Nat. Rev. Cancer 3 (2623–627.
[4] B. Li, A.J. Lustig, A novel mechanism for telomere size controSaccharomyces cerevisiae, Genes Dev. 10 (1996) 1310–1326.
[5] M. Bucholc, Y. Park, A.J. Lustig, Intrachromatid excision of teloeric DNA as a mechanism for telomere size control inSaccharomyces cerevisiae, Mol. Cell Biol. 21 (2001) 6559–6573.
[6] E. Trelles-Sticken, M.E. Dresser, H. Scherthan, Meiotic telomprotein Ndj1p is required for meiosis-specific telomere distribubouquet formation and efficient homologue pairing, J. Cell Biol.(2000) 95–106.
[7] I. Joseph, D. Jia, A.J. Lustig, Ndj1-dependent epigenetic resettitelomere size in yeast meiosis, Curr. Biol. 15 (2005) 231–237.
[8] R.C. Wang, A. Smogorzewska, T. de Lange, Homologous rebination generates T-loop-sized deletions at human telomeres119 (2004) 355–368.
[9] A.J. Lustig, Clues to catastrophic telomere loss in mammalsyeast telomere rapid deletion, Nat. Rev. Genet. 9 (2003) 916–
10] J.D. Griffith, L. Comeau, S. Rosenfield, R.M. Stansel, A. BianH. Moss, T. de Lange, Mammalian telomeres end in a large duloop, Cell 97 (1999) 503–514.
11] K.G. Murti, D.M. Prescott, Telomeres of polytene chromosomesciliated protozoan terminate in duplex DNA loops, Proc. Natl. ASci. U.S.A. 96 (1999) 14436–14439.
12] J.L. Munoz-Jordan, G.A. Cross, T. de Lange, J.D. Griffith, T-loat trypanosome telomeres, EMBO J. 20 (2001) 579–588.
13] L. Tomaska, S. Willcox, J. Slezakova, J. Nosek, J.D. Griffith, Tbinding to a fission yeast model telomere: formation of t-loopshigher order structures, J. Biol. Chem. 279 (2004) 50764–507
14] T. de Lange, Opinion: T-loops and the origin of telomeres, Nat.Mol. Cell Biol. 5 (2004) 323–329.
Genet. 20 (1998) 203–206.25] Y. Tsukamoto, A.K. Taggart, V.A. Zakian, The role of the Mre
Rad50-Xrs2 complex in telomerase-mediated lengthening ofSaccharomyces cerevisiaetelomeres, Curr. Biol. 11 (2001) 1328–1335.
26] K. Myung, A. Datta, R.D. Kolodner, Suppression of spontanechromosomal rearrangements by S phase checkpoint functioSaccharomyces cerevisiae, Cell 104 (2001) 397–408.
27] T.H. Stracker, J.W. Theunissen, M. Morales, J.H. Petrini, The Mcomplex and the metabolism of chromosome breaks: the imporof communicating and holding things together, DNA Repair (Am3 (2004) 845–854.
28] H. Tsubouchi, H. Ogawa, A novel mre11 mutation impairs procing of double-strand breaks of DNA during both mitosis and meiMol. Cell Biol. 18 (1998) 260–268.
29] E.L. Ivanov, N. Sugawara, C.I. White, F. Fabre, J.E. Haber, Mutain XRS2 and RAD50 delay but do not prevent mating-type switcin Saccharomyces cerevisiae, Mol. Cell. Biol. 14 (1994) 3414–342
30] B. Llorente, L.S. Symington, The Mre11 nuclease is not requfor 5′ to 3′ resection at multiple HO-induced double-strand breMol. Cell Biol. 24 (2004) 9682–9694.
31] D. Nakada, Y. Hirano, K. Sugimoto, Requirement of the Mrcomplex and exonuclease 1 for activation of the Mec1 signpathway, Mol. Cell Biol. 24 (2004) 10016–10025.
32] L.K. Lewis, G. Karthikeyan, J.W. Westmoreland, M.A. Resnick, Dferential suppression of DNA repair deficiencies of Yeast ramre11 and xrs2 mutants by EXO1 and TLC1 (the RNA compoof telomerase), Genetics 160 (2002) 49–62.
33] S. Moreau, E.A. Morgan, L.S. Symington, Overlapping functiontheSaccharomyces cerevisiaeMre11, Exo1 and Rad27 nucleasesDNA metabolism, Genetics 159 (2001) 1423–1433.
34] H. Tsubouchi, H. Ogawa, Exo1 roles for repair of DNA doubstrand breaks and meiotic crossing over inSaccharomyces cerevisia,Mol. Biol. Cell 11 (2000) 2221–2233.
35] S.J. Diede, D.E. Gottschling, Exonuclease activity is requiredsequence addition and Cdc13p loading at a de novo telomere,Biol. 11 (2001) 1336–1340.

1004 B. Williams et al. / DNA Repair 4 (2005) 994–1005
[36] M. Larrivee, C. LeBel, R.J. Wellinger, The generation of properconstitutive G-tails on yeast telomeres is dependent on the MRXcomplex, Genes Dev. 18 (2004) 1391–1396.
[37] X.D. Zhu, B. Kuster, M. Mann, J.H. Petrini, T. de Lange, Cell-cycle-regulated association of RAD50/MRE11/NBS1 with TRF2 andhuman telomeres, Nat. Genet. 25 (2000) 347–352.
[38] R.M. Stansel, T. de Lange, J.D. Griffith, T-loop assembly in vitroinvolves binding of TRF2 near the 3′ telomeric overhang, EMBO J.20 (2001) 5532–5540.
[39] P.L. Opresko, C. von Kobbe, J.P. Laine, J. Harrigan, I.D. Hickson,V.A. Bohr, Telomere-binding protein TRF2 binds to and stimulatesthe Werner and Bloom syndrome helicases, J. Biol. Chem. 277(2002) 41110–41119.
[40] D.J. Stavropoulos, P.S. Bradshaw, X. Li, I. Pasic, K. Truong, M.Ikura, M. Ungrin, M.S. Meyn, The Bloom syndrome helicase BLMinteracts with TRF2 in ALT cells and promotes telomeric DNA syn-thesis, Hum. Mol. Genet. 11 (2002) 3135–3144.
[41] P.L. Opresko, M. Otterlei, J. Graakjaer, P. Bruheim, L. Dawut, S.Kolvraa, A. May, M.M. Seidman, V.A. Bohr, The Werner syndromehelicase and exonuclease cooperate to resolve telomeric D Loopsin a manner regulated by TRF1 and TRF2, Mol. Cell 14 (2004)763–774.
[42] Z.R. Razak, R.J. Varkonyi, M. Kulp-McEliece, C. Caslini, J.R. Testa,M.E. Murphy, D. Broccoli, p53 differentially inhibits cell growthdepending on the mechanism of telomere maintenance, Mol. CellBiol. 24 (2004) 5967–5977.
[43] F. d’Adda di Fagagna, M.P. Hande, W.M. Tong, D. Roth, P.M. Lans-dorp, Z.Q. Wang, S.P. Jackson, Effects of DNA nonhomologousend-joining factors on telomere length and chromosomal stabilityin mammalian cells, Curr. Biol. 11 (2001) 1192–1196.
[44] D. Gilley, H. Tanaka, M.P. Hande, A. Kurimasa, G.C. Li, M.ing,
[ hn-areNatl.
[ .A.nde-over-
[ ericem.
[ per,lom-plex,
[ nese
[ cter-r.
[ e-and.
[ hosts
[ ity ofing,99)
[ wa,ing
[55] J.H. Toyn, P.L. Gunyuzlu, W.H. White, L.A. Thompson, G.F. Hol-lis, A counterselection for the tryptophan pathway in yeast: 5-fluoroanthranilic acid resistance, Yeast 16 (2000) 553–560.
[56] M. Chamankhah, W. Xiao, Formation of the yeast Mre11-Rad50-Xrs2 complex is correlated with DNA repair and telomere mainte-nance, Nucleic Acids Res. 27 (1999) 2072–2079.
[57] D.A. Bressan, H.A. Olivares, B.E. Nelms, J.H. Petrini, Alterationof N-terminal phosphoesterase signature motifs inactivatesSaccha-romyces cerevisiaeMre11, Genetics 2 (1998) 591–600.
[58] J.A. Printen, G.F. Sprague Jr., Protein–protein interactions in theyeast pheromone response pathway: Ste5p interacts with all membersof the MAP kinase cascade, Genetics 138 (1994) 609–619.
[59] K.M. Trujillo, S.S. Yuan, E.Y. Lee, P. Sung, Nuclease activi-ties in a complex of human recombination and DNA repair fac-tors Rad50, Mre11, and p95, J. Biol. Chem. 273 (1998) 21447–21450.
[60] T.T. Paull, M. Gellert, Nbs1 potentiates ATP-driven DNA unwindingand endonuclease cleavage by the Mre11/Rad50 complex, GenesDev. 13 (1999) 1276–1288.
[61] M. Furuse, Y. Nagase, H. Tsubouchi, K. Murakami-Murofushi, T.Shibata, K. Ohta, Distinct roles of two separable in vitro activitiesof yeast Mre11 in mitotic and meiotic recombination, EMBO J. 17(1998) 6412–6425.
[62] S.E. Lee, D.A. Bressan, J.H. Petrini, J.E. Haber, Complementationbetween N-terminalSaccharomyces cerevisiaemre11 alleles in DNArepair and telomere length maintenance, DNA Repair (Amst.) 1(2002) 27–40.
[63] K.P. Hopfner, A. Karcher, L. Craig, T.T. Woo, J.P. Carney, J.A.Tainer, Structural biochemistry and interaction architecture of theDNA double-strand break repair Mre11 nuclease and Rad50-ATPase,Cell 105 (2001) 473–485.
[ .H.Car-cids
[ merand
[ om-2003)
[ .A.aenet-
[ mat-)
[ -trand91)
[ H.i-
d theble-86–
[ atheck-Dev.
[ ionae
[ ures
Oshimura, D.J. Chen, DNA-PKcs is critical for telomere cappProc. Natl. Acad. Sci. U.S.A. 98 (2001) 15084–15088.
45] S.M. Bailey, J. Meyne, D.J. Chen, A. Kurimasa, G.C. Li, B.E. Leert, E.H. Goodwin, DNA double-strand break repair proteinsrequired to cap the ends of mammalian chromosomes, Proc.Acad. Sci. U.S.A. 96 (1999) 14899–14904.
46] E. Samper, F.A. Goytisolo, P. Slijepcevic, P.P. van Buul, MBlasco, Mammalian Ku86 protein prevents telomeric fusions ipendently of the length of TTAGGG repeats and the G-strandhang, EMBO Rep. 1 (2000) 244–252.
47] R.M. Stansel, D. Subramanian, J.D. Griffith, p53 binds telomsingle strand overhangs and t-loop junctions in vitro, J. Biol. Ch277 (2002) 11625–11628.
48] J.Z. Ye, D. Hockemeyer, A.N. Krutchinsky, D. Loayza, S.M. HooB.T. Chait, T. de Lange, POT1-interacting protein PIP1: a teere length regulator that recruits POT1 to the TIN2/TRF1 comGenes. Dev. 18 (2004) 1649–1654.
49] M. Ajimura, S.H. Leem, H. Ogawa, Identification of new gerequired for meiotic recombination inSaccharomyces cerevisia,Genetics 133 (1993) 51–66.
50] M. Chamankhah, W. Xiao, Molecular cloning and genetic charaization of theSaccharomyces cerevisiaeNGS1/MRE11 gene, CurGenet. 34 (1998) 368–374.
51] M. Chamankhah, T. Fontanie, W. Xiao, TheSaccharomyces cervisiae mre11(ts) allele confers a separation of DNA repairtelomere maintenance functions, Genetics 155 (2000) 569–576
52] R.S. Sikorski, P. Hieter, A system of shuttle vectors and yeaststrains designed for efficient manipulation of DNA inSaccharomycecerevisiae, Genetics 122 (1989) 19–27.
53] S. Moreau, J.R. Ferguson, L.S. Symington, The nuclease activMre11 is required for meiosis but not for mating type switchend joining, or telomere maintenance, Mol. Cell Biol. 19 (19556–566.
54] T. Usui, T. Ohta, H. Oshiumi, J. Tomizawa, H. Ogawa, T. OgaComplex formation and functional versatility of Mre11 of buddyeast in recombination, Cell 95 (1998) 705–716.
64] L.M. Arthur, K. Gustausson, K.P. Hopfner, C.T. Carson, TStracker, A. Karcher, D. Felton, M.D. Weitzman, J. Tainer, J.P.ney, Structural and functional analysis of Mre11-3, Nucleic ARes. 32 (2004) 1886–1893.
65] R.M. Polotnianka, J. Li, A.J. Lustig, The yeast Ku heterodiis essential for protection of the telomere against nucleolyticrecombinational activities, Curr. Biol. 8 (1998) 831–834.
66] D. Lydall, Hiding at the ends of yeast chromosomes: teleres, nucleases and checkpoint pathways, J. Cell Sci. 116 (4057–4065.
67] L.K. Lewis, F. Storici, S. Van Komen, S. Calero, P. Sung, MResnick, Role of the nuclease activity ofSaccharomyces cerevisiMre11 in repair of DNA double-strand breaks in mitotic cells, Geics 166 (2004) 1701–1713.
68] C.I. White, J.E. Haber, Intermediates of recombination duringing type switching inSaccharomyces cerevisiae, EMBO J. 9 (1990663–673.
69] H. Sun, D. Treco, J.W. Szostak, Extensive 3′-overhanging, singlestranded DNA associated with the meiosis-specific double-sbreaks at the ARG4 recombination initiation site, Cell 64 (191155–1161.
70] K. Tomita, A. Matsuura, T. Caspari, A.M. Carr, Y. Akamatsu,Iwasaki, K. Mizuno, K. Ohta, M. Uritani, T. Ushimaru, K. Yoshnaga, M. Ueno, Competition between the Rad50 complex anKu heterodimer reveals a role for Exo1 in processing doustrand breaks but not telomeres, Mol. Cell Biol. 23 (2003) 515197.
71] L. Maringele, D. Lydall, EXO1-dependent single-stranded DNAtelomeres activates subsets of DNA damage and spindle cpoint pathways in budding yeast yku70Delta mutants, Genes16 (2002) 1919–1933.
72] X. Jia, T. Weinert, D. Lydall, Mec1 and Rad53 inhibit formatof single-stranded DNA at telomeres ofSaccharomyces cerevisicdc13-1 mutants, Genetics 166 (2004) 753–764.
73] D.E. Anderson, K.M. Trujillo, P. Sung, H.P. Erickson, Structof the Rad50× Mre11 DNA repair complex fromSaccharomyce

B. Williams et al. / DNA Repair 4 (2005) 994–1005 1005
cerevisiae by electron microscopy, J. Biol. Chem. 276 (2001)37027–37033.
[74] G. Ira, J.E. Haber, Characterization of RAD51-independent break-induced replication that acts preferentially with short homologoussequences, Mol. Cell Biol. 22 (2002) 6384–6392.
[75] G.M. Manthey, A.M. Bailis, Multiple pathways promote short-sequence recombination inSaccharomyces cerevisiae, Mol. Cell Biol.22 (2002) 5347–5356.
[76] G.J. Sharples, D.R. Leach, Structural and functional similaritiesbetween the SbcCD proteins ofEscherichia coliand the RAD50
and MRE11 (RAD32) recombination and repair proteins of yeast,Mol. Microbiol. 17 (1995) 1215–1217.
[77] S. Zhuo, J.C. Clemens, R.L. Stone, J.E. Dixon, Mutational analy-sis of a Ser/Thr phosphatase. Identification of residues important inphosphoesterase substrate binding and catalysis, J. Biol. Chem. 269(1994) 26234–26238.
[78] D.E. Gottschling, O.M. Aparicio, B.L. Billington, V.A. Zakian, Posi-tion effect atS. cerevisiaetelomeres: reversible repression of Pol IItranscription, Cell 63 (1990) 751–762.