a synaptic mechanism underlying the behavioral abnormalities induced by manganese intoxication
TRANSCRIPT

S
pt(
Neurobiology of Disease 8, 419–432 (2001)doi:10.1006/nbdi.2000.0379, available online at http://www.idealibrary.com on
A
A Synaptic Mechanism Underlying the BehavioralAbnormalities Induced by ManganeseIntoxication
Paolo Calabresi,* ,† Martine Ammassari-Teule,‡ Paolo Gubellini,* ,§
Giuseppe Sancesario,* Maria Morello,* Diego Centonze,* ,†
Girolama A. Marfia,* Emilia Saulle,* Enrica Passino,‡
Barbara Picconi,* ,† and Giorgio Bernardi* ,†
*Clinica Neurologica, Dipartemente Neuroscienze, Universita’ di Roma Tor Vergata, Via diTor Vergata 135, Rome 00133 Italy; †I.R.C.C.S., Ospedale Santa Lucia, Rome, Italy;‡Istituto di Psicobiologia e Psicofarmacologia; and §Istituto di Medicina
perimentale, C.N.R., Rome, Italy
Received June 27, 2000; revised November 22, 2000; accepted December 12, 2000;published online May 3, 2001
In the present study we have characterized a rat model of manganese (Mn) intoxication leading tobehavioral disinhibition in the absence of major motor alterations. These behavioral changes wereassociated with significantly increased brain Mn levels but were uncoupled to anatomical lesions ofthe striatum or to morphological and cytochemical changes of the nigrostriatal dopaminergic path-way. The analysis of this model at cellular level showed an enhanced dopaminergic inhibitory controlof the corticostriatal excitatory transmission via presynaptic D2-like dopamine (DA) receptors in slicesobtained from Mn-treated rats. Conversely, the use of agonists acting on presynaptic purinergic,muscarinic, and glutamatergic metabotropic receptors revealed a normal sensitivity. Moreover, mem-brane responses recorded from single dopaminergic neurons following activation of D2 DA autore-ceptors were also unchanged following Mn intoxication. Thus, our findings indicate a selectiveinvolvement of the D2-like DA receptors located on glutamatergic corticostriatal terminals in thispathological condition and suggest that the behavioral symptoms described in the “early” clinicalphase of manganism may be caused by an abnormal dopaminergic inhibitory control on corticostriatalinputs. The identification of the synaptic mechanism underlying the “early” phase of Mn intoxicationmight have a critical importance to understand the causes of the progression of this pathologicalcondition towards an “established” phase characterized by motor abnormalities and anatomicallesions of the basal ganglia. © 2001 Academic Press
Key Words: basal ganglia; dopamine receptors; learning; movement disorders; parkinsonism; stri-atum.
tacaC
hafr
INTRODUCTION
Manganese (Mn) is an essential element for biologicfunctions, but excessive exposure is toxic. Chronic Mnencephalopathy was first recognized among workersengaged in the grinding of manganese ores (Couper,1837; Cotzias, 1958; Gorell et al., 1997). Cases of Mn
oisoning have also been reported in families exposedo this metal through contaminated well-waterKawamura et al., 1941). Mn is used in the manufac-
0969-9961/01 $35.00Copyright © 2001 by Academic Press
ll rights of reproduction in any form reserved. 419
ure of dry batteries and this metal is also contained inwidely used organochemical fungicide. In some
ountries, an organic compound containing Mn isdded to gasoline as an antiknock agent (Saric, 1986;ooper, 1984).As a result of the frequent industrial accidents Mn
as been labeled by the World Health Organization asn environmental toxic factor that induces brain dys-unction. Moreover, Mn intoxication has also beeneported in patients with chronic liver failure (Hauser

(elatttM
ic
420 Calabresi et al.
A
et al., 1994) and in children receiving long-term par-enteral nutrition (Fell et al., 1996).
Chronic Mn intoxication was once thought to pro-duce clinical features mimicking idiopathic Parkin-son’s disease (PD), but a close examination of theneurologic manifestations and of the pathology hasrevealed important differences between the two dis-orders (Barbeau, 1984; Calne et al., 1994). The mostimportant among these differences is the lack of clin-ical response to levodopa in manganism (Calne et al.,1994; Olanow et al., 1996).
Interestingly, Rodier and Cotzias reported thatmanganism involves two domains of effects, emo-tional and motor, that were considered as differentphases of an apparently progressive syndrome. Theearly phase was called “manganese madness” andwas characterized by emotional lability, mania, distur-bance of sleep, and eating and sexual disturbances(Rodier, 1955; Cotzias, 1958) but few, and subtle, mo-tor effects. A later phase, which Rodier called “estab-lished” phase, was dominated by motor sequelae suchas bradykinesia, rigidity, and dystonia. These motoreffects have been clearly attributed to an abnormalbasal ganglia functioning (Rodier, 1955; Cotzias, 1958;Mena et al., 1969; Barbeau et al., 1976; Barbeau, 1984;Calne et al., 1994; Huang et al., 1998).
Magnetic resonance imaging of the brain may revealalterations in the globus pallidus, striatum, and in themidbrain (Calne et al., 1994). Positron emission tomog-raphy reveals normal presynaptic and postsynapticdopaminergic function (Shinotoh et al., 1995, 1997).These findings lead to hypothesize that the pathologyis downstream the dopaminergic projection. Thus, theprimary site of neurological damage has been postu-lated to be the globus pallidus (Calne et al., 1994;Shinotoh et al., 1997). However, data obtained fromclinical observations do not fully account for the bio-chemical changes measured in the nigrostriatal systemin various animal models of Mn intoxication. In par-ticular, most of these experimental studies showedaltered striatal DA concentration after Mn intoxication(Newland, 1999). In addition, the physiological activ-ity of this system following Mn-intoxication has neverbeen investigated at cellular level.
For this reason, in the present study we haveutilized a rat model of Mn intoxication to analyse: (i)various behavioral parameters related to emotionalstates and learning processes, (ii) possible morpho-logical changes of the nigrostriatal system, and (iii)synaptic and cellular properties of rat striatal andnigral neurons and their response to dopamine ago-nists.
Copyright © 2001 by Academic Pressll rights of reproduction in any form reserved.
METHOD
Chronic Treatment of the Animals
One hundred male Wistar rats, weighing 80 6 10 gabout 20 days age), were divided into two groups ofqual numbers. All the animals were housed in stain-ess steel cages in a conditioned room, where regularlternate cycles of 12 h light and darkness were main-ained. Animals were supplied with pellet diet ad libi-um (Morini, Bologna, Italy). The rats of group I (Mn-reated) were given free access to drinking solutions of
nCl2 (20 mg/ml of water) in place of their drinkingwater. The group II (control) were given the same dietand sodium chloride in their drinking solution. Theaverage consumption of drinking solution was similarat different time intervals. The treatment lasted for 10weeks and then the animals were either sacrificed forbiochemical, morphological, and electrophysiologicalexperiments or were analyzed for behavioral activity.The average final weight was 312 6 63 g for controlrats and 319 6 66 g for Mn-treated animals (P . 0.05).
Measurement of Manganese Levels in Brainand Liver
Samples of homogenates of either brain areas orlivers were digested for 1 week at room temperatureby addition of four volumes of 65% HNO3. Mn contentwas estimated either in Mn-treated rats (n 5 10) or incontrol animals (n 5 10) by an atomic absorptionspectrophotometer (Perkin–Elmer, 2100), equippedwith a graphite furnace with platform. Mn standardfor atomic absorption was purchased from Carlo Erba(Milano, Italy). Data were expressed as nmol/mg pro-tein. Protein content was estimated with bovine serumalbumin as a standard as previously reported (Lowry,1951).
Behavioral Experiments
In order to investigate behavioral alterations consec-utive to exposure to Mn, control (n 5 10), and Mn-treated rats (n 5 10) at the beginning of the experi-ments were first tested for their locomotor activity andtheir reactivity to object novelty in an open field, thenfor spatial memory in an eight-arm radial maze (Am-massari-Teule et al., 1994). Locomotor activity wasmeasured in a quadratic open field made of greyplastic material. The floor (100 3 100 cm) was dividednto 25 squares by white lines and delimited by 40-m-high walls. Each rat was put on the center of the

421Synaptic Mechanisms of Manganese Intoxication
open field and retained 15 s in a transparent cylinder.After the removal of the cylinder, behavioral record-ings were made for 5 min by mean of a keyboardconnected to a PC. Number of crossed squares andnumber of rearing responses (against the walls or not)were scored. After 5 min elapsed, an object—a plexi-glass cube with holes irregularly distributed on eachside—was placed on a central square and the timespent in contact with the novel object was measured.The rat was then removed from the open field, thedefecation boluses were counted and both the openfield and the object were cleaned with a solution madeof water and alcohol (10%). Rats were tested for threeconsecutive days at 2:00 p.m. Each variable was ana-lyzed with a two-way ANOVA (treatment 3 sessions).
After a rest period of one week, eight rats wererandomly extracted from the control and the treatedgroups and tested for the spatial memory in an eight-arm radial maze. They were caged in pairs with foodand water ad libitum. Three days before the pretrainingbegan, food was available only 1 h per day in order tobring the weight of each rat at about 90% of the initiallevel. Rats were tested in a maze made of grey plasticmaterial and consisting of eight identical paths (65 312 cm) radiating from an octogonal platform. Anopaque food cup was put at the distal end of each arm.The maze was elevated at 60 cm above the floor,maintained in a constant orientation throughout theexperiment, and placed in a sound proof cabin whichhad several conspicuous extra maze cues. During pre-training, each rat was placed on the starting platformand allowed both to explore the paths and to consumefood scattered in the whole maze for 10 min. Thispretraining procedure was repeated for 3 consecutivedays. The actual training procedure started on thefourth day. Before each trial all the paths were baitedwith a 1-g piece of purina chow placed in each foodcup. The animals ran a trial per day during 8 consec-utive days. On each trial, the rats were placed on thecentral platform and allowed to make eight choices. Atrial was correct when the rat visited the eight differ-ent paths only once, in a free order. The followingvariables were recorded: number of correct choices,time spent to complete the trial, and number of 45°angle turns displayed during the sequence of runs.Each variable was analyzed with a two-way ANOVA(treatment 3 trials).
Morphological Experiments
Control (n 5 10) and Mn-treated (n 5 10) rats weredeeply anesthetized with chloral hydrate (400 mg/kg)
and fixed by transcardiac perfusion with 4% bufferedparaformaldheyde (pH 7.4) after a brief rinse withsaline and heparin (0.1%) at room temperature. Thebrains were removed, kept in cold fixative for 2 h andstored in PB 0.1 M with 30% sucrose for 48 h, frozen,and then sectioned in a cryostat at 40-mm thickness.
Brain sections through the forebrain and the mid-brain were either stained with cresyl violet to evaluategeneral tissue morphology or processed immunohis-tochemically for the visualization of glial fibrillaryacid proteins (GFAP) and tyrosine hydroxylase (TH).Variations of GFAP immunoreactivity is thought to bea sensitive index of astrocyte reaction to any damageoccurring in brain tissue, while TH is a key enzyme inthe synthesis of DA and specifically marks dopami-nergic neurons and nerve fibers in the nigrostriatalregion. For morphological studies, sections weremounted on gelatin-chromallum-coated slices, stainedwith cresyl violet, deydrated in graded alcohol, andexamined using a light microscope (Zeiss). For immu-nohistochemical studies, free-floating sections wereplaced for 30 min at room temperature in a solutioncontaining an equal volume of PBS and 3% hydrogenperoxide to eliminate endogenous peroxidase activity.After three rinses of 10 min each, the sections werepreincubated for 30 min in a solution containing 10%horse serum and 0.1% of Triton X-100 in PBS. Theywere then incubated overnight at 4°C in a solutioncontaining 10% horse serum plus antimonoclonalGFAP, dilution 1:400 (Sigma, Milan Italy) or anti-monoclonal TH, dilution 1:100 (Incstar, DBA Milan,Italy). After three rinses of 10 min each in PBS, thesections were incubated for 1 h at room temperature inthe biotinilated secondary antibody and then incu-bated with avidin-biotin-peroxidase complex diluted1:100 at room temperature (Kit ABC, Vector, DBA,Milan, Italy). The sections were washed twice in PBSand once in 0.05 M Tris buffer (pH 7.6). The boundperoxidase was revealed by placing the sections in amedium containing 0.05% and 393-diaminobenzidinetetrahydrochloride (Sigma, Milan, Italy) in 0.05 M Trisbuffered (pH 7.6) at room temperature. The sectionswere mounted on gelatin-chromallum-coated sliceand examined with a light microscope.
Two independent observers evaluated in a blind fash-ion the morphological characteristics and the relativenumbers of cells, neurons and astrocytes, in the striatumand substantia nigra using a light microscope (Zeiss)equipped with a 25X objective. Morphometric analysiswas performed by an expert operator blind to the phar-macological treatment received by each animal, using animage analyser system (Quantimet 500-Leica). The num-
Copyright © 2001 by Academic PressAll rights of reproduction in any form reserved.

bcgii9dcN
sK
lamicassqacfaqpl(
422 Calabresi et al.
A
ber of striatal neurons was counted in four alternateNissl-stained sections, involving the whole cross-sec-tional area of the striatum (1.7 mm anterior to 1.0 poste-rior to bregma) and expressed as mean 6 SD per unittest area (3 3 105 mm2). The number of astrocytes and therelative extent of their processes was counted in threeconsecutive sections, involving the whole cross-sectionalarea of the striatum (1.7 mm anterior to 1.0 posterior tobregma). To determine the abundance of TH-positiveneurons in the substantia nigra (between 5.2 mm behindand 6.0 behind bregma), and of TH-positive nerve fibersin the striatum (1.7 mm anterior to 1.0 posterior to bregma),the relative intensity of TH immunoreactive productagainst the background staining in the regions of interestwas evaluated by an image analysis system Quantimet970 (Cambridge Instruments). Student’s t test was usedto evaluate differences between control and experimen-tal animals. Data are presented as mean 6 SD; when P ,0.05, differences were considered significant.
Electrophysiological Experiments fromCorticostriatal and Nigral Slices
Forty control rats and forty Mn-treated animalswere used. The preparation and maintenance of coro-nal slices have been described previously for bothcorticostriatal slices and nigral slices (Kita et al., 1984;Calabresi et al., 1990, 1992a, 1993, 1998a). Briefly, slices(200–300 mm) were prepared from tissue blocks of the
rain with the use of a vibratome. Eight to twelveorticostriatal slices were usually obtained from a sin-le rat brain. Single slice was transferred to a record-
ng chamber and submerged in a continuously flow-ng Krebs solution (35°C, 2–3 ml/min) gassed with5% O2–5% CO2. Complete replacement of the me-ium in the chamber took approximately 90 s. Theomposition of the control solution was (in mM): 126aCl, 2.5 KCl, 1.2 MgCl2, 1.2 NaH2PO4, 2.4 CaCl2, 11
Glucose, 25 NaHCO3. For intracellular recordingsharp electrodes were used; they were filled with 2 MCl (30–60 MV). An Axoclamp 2A amplifier was used
TABLE 1
Manganese Levels in the Brain and Liver of Control and Chronic M
Basal ganglia
Control 0.0415 6 0.0047 0.0Chronic manganese 0.1260 6 0.0288 0.1
Note. Values are expressed as means 6 SD of nmol Mn/mg prosignificantly higher than those measured in control animals (P , 0
Copyright © 2001 by Academic Pressll rights of reproduction in any form reserved.
for recordings either in current-clamp or in voltage-clamp mode. In single-electrode voltage-clamp modethe switching frequency was 3 kHz. The headstagesignal was continuously monitored on a separate os-cilloscope. Traces were displayed on an oscilloscopeand stored on a digital system. For synaptic stimula-tion, bipolar electrodes were used. These stimulatingelectrodes were located in the white matter betweenthe cortex and the striatum in order to activate corti-costriatal glutamatergic fibres. Stimuli were simplerectangular pulses, typically 100 ms and 8–30 V. Syn-aptic potentials were measured by averaging re-sponses to four or eight stimuli.
Values given in the text and in the figures aremean 6 SEM of changes in the respective cell popu-ations. Wilcoxon’s test or Student’s t test (for pairednd unpaired observations) were used to compare theeans and ANOVA was used when multiple compar-
sons were made against a single control group. Theharacteristics of action potentials and of current-volt-ge curves in different experimental conditions weretudied by utilizing a fast chart recorder and a digitalystem (Nicolet System 400: Benchtop Waveform Ac-uisition System and Axon Axoscope 7.0). Drugs werepplied by dissolving them to the desired final con-entration in the saline and by switching the perfusionrom control saline to drug-containing saline. D-2-mino-5-phosphonovalerate (APV), 6-cyano-7-nitro-uinoxaline-2,3-dione (CNQX), and l-serine-O-phos-hate (L-SOP) were from Tocris. Adenosine, bicucul-
ine, dopamine (DA), and glutamate were from SigmaItaly). Oxotremorine was from RBI (U.S.A.).
RESULTS
Measurements of Manganese Levels in BrainAreas and Liver
As shown in Table 1, chronic Mn-treatment induceda significant (P , 0.01) increase in Mn levels in basal
ese-Treated Rats
x Cerebellum Liver
.0071 0.0445 6 0.0039 0.0505 6 0.0144
.0234 0.1720 6 0.0593 0.5002 6 0.0805
ll values in tissues from chronic manganese-treated animals weredent’s t test).
angan
Corte
373 6 0134 6 0
tein. A.01, Stu
4
423Synaptic Mechanisms of Manganese Intoxication
ganglia, cortex, cerebellum, and liver in comparisonwith the values obtained from control rats. Note thatin the various brain areas the Mn concentration in-creased of about three times following this treatmentwhile in the liver the Mn concentration in treatedanimals was about 10 times higher than in controlanimals.
Locomotor Activity and Reactivity to ObjectNovelty
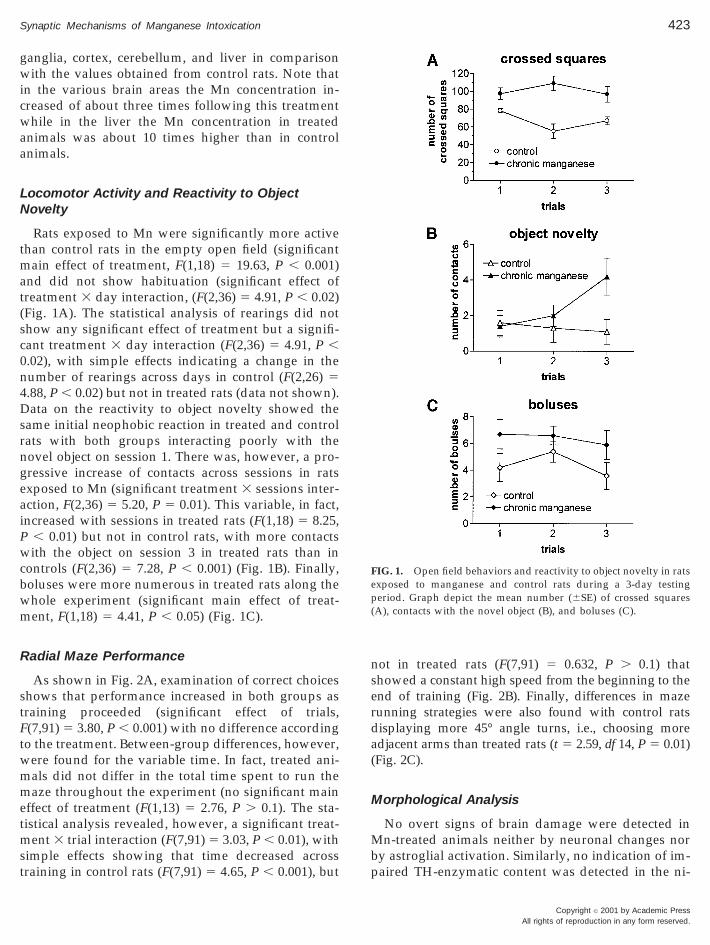
Rats exposed to Mn were significantly more activethan control rats in the empty open field (significantmain effect of treatment, F(1,18) 5 19.63, P , 0.001)and did not show habituation (significant effect oftreatment 3 day interaction, (F(2,36) 5 4.91, P , 0.02)(Fig. 1A). The statistical analysis of rearings did notshow any significant effect of treatment but a signifi-cant treatment 3 day interaction (F(2,36) 5 4.91, P ,0.02), with simple effects indicating a change in thenumber of rearings across days in control (F(2,26) 5.88, P , 0.02) but not in treated rats (data not shown).
Data on the reactivity to object novelty showed thesame initial neophobic reaction in treated and controlrats with both groups interacting poorly with thenovel object on session 1. There was, however, a pro-gressive increase of contacts across sessions in ratsexposed to Mn (significant treatment 3 sessions inter-action, F(2,36) 5 5.20, P 5 0.01). This variable, in fact,increased with sessions in treated rats (F(1,18) 5 8.25,P , 0.01) but not in control rats, with more contactswith the object on session 3 in treated rats than incontrols (F(2,36) 5 7.28, P , 0.001) (Fig. 1B). Finally,boluses were more numerous in treated rats along thewhole experiment (significant main effect of treat-ment, F(1,18) 5 4.41, P , 0.05) (Fig. 1C).
Radial Maze Performance
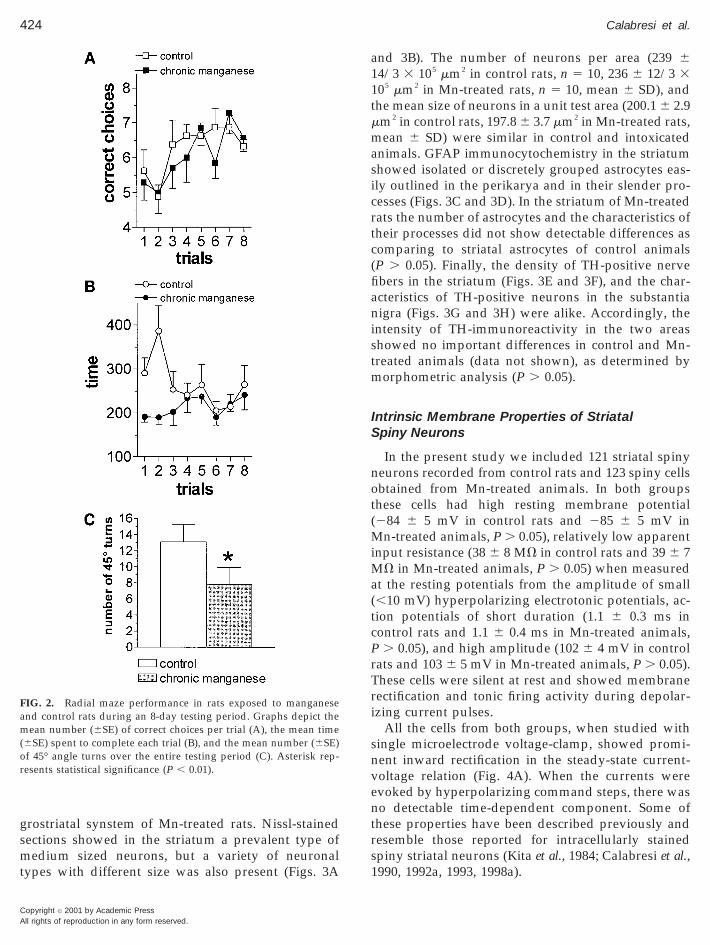
As shown in Fig. 2A, examination of correct choicesshows that performance increased in both groups astraining proceeded (significant effect of trials,F(7,91) 5 3.80, P , 0.001) with no difference accordingto the treatment. Between-group differences, however,were found for the variable time. In fact, treated ani-mals did not differ in the total time spent to run themaze throughout the experiment (no significant maineffect of treatment (F(1,13) 5 2.76, P . 0.1). The sta-tistical analysis revealed, however, a significant treat-ment 3 trial interaction (F(7,91) 5 3.03, P , 0.01), withsimple effects showing that time decreased acrosstraining in control rats (F(7,91) 5 4.65, P , 0.001), but
not in treated rats (F(7,91) 5 0.632, P . 0.1) thatshowed a constant high speed from the beginning to theend of training (Fig. 2B). Finally, differences in mazerunning strategies were also found with control ratsdisplaying more 45° angle turns, i.e., choosing moreadjacent arms than treated rats (t 5 2.59, df 14, P 5 0.01)(Fig. 2C).
Morphological Analysis
No overt signs of brain damage were detected inMn-treated animals neither by neuronal changes norby astroglial activation. Similarly, no indication of im-paired TH-enzymatic content was detected in the ni-
FIG. 1. Open field behaviors and reactivity to object novelty in ratsexposed to manganese and control rats during a 3-day testingperiod. Graph depict the mean number (6SE) of crossed squares(A), contacts with the novel object (B), and boluses (C).
Copyright © 2001 by Academic PressAll rights of reproduction in any form reserved.
424 Calabresi et al.
A
grostriatal synstem of Mn-treated rats. Nissl-stainedsections showed in the striatum a prevalent type ofmedium sized neurons, but a variety of neuronaltypes with different size was also present (Figs. 3A
FIG. 2. Radial maze performance in rats exposed to manganeseand control rats during an 8-day testing period. Graphs depict themean number (6SE) of correct choices per trial (A), the mean time(6SE) spent to complete each trial (B), and the mean number (6SE)of 45° angle turns over the entire testing period (C). Asterisk rep-resents statistical significance (P , 0.01).
Copyright © 2001 by Academic Pressll rights of reproduction in any form reserved.
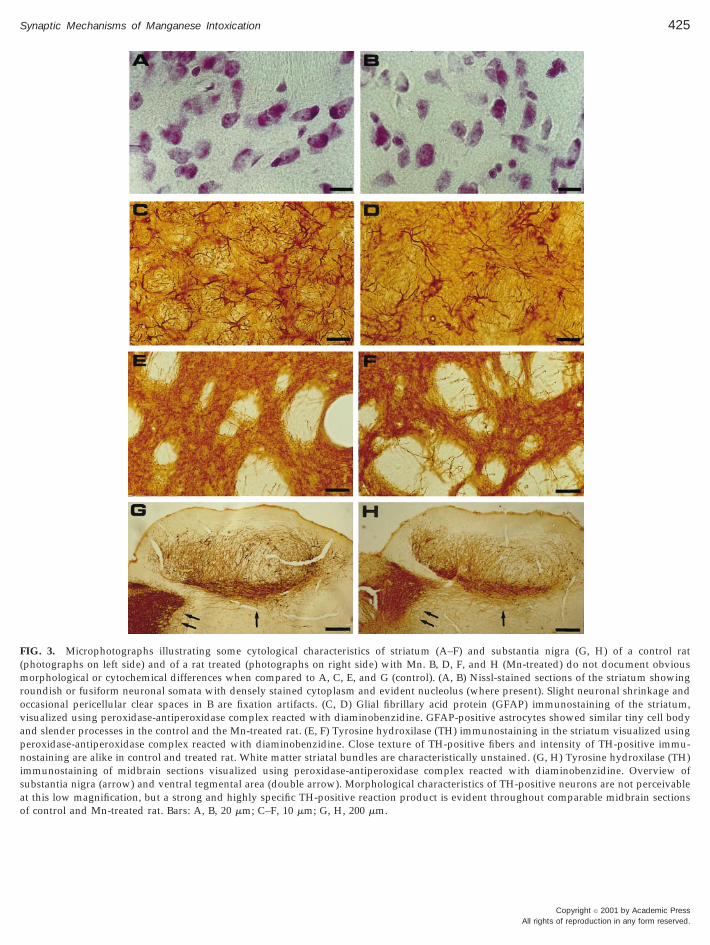
and 3B). The number of neurons per area (239 614/3 3 105 mm2 in control rats, n 5 10, 236 6 12/3 3105 mm2 in Mn-treated rats, n 5 10, mean 6 SD), andthe mean size of neurons in a unit test area (200.1 6 2.9mm2 in control rats, 197.8 6 3.7 mm2 in Mn-treated rats,mean 6 SD) were similar in control and intoxicatedanimals. GFAP immunocytochemistry in the striatumshowed isolated or discretely grouped astrocytes eas-ily outlined in the perikarya and in their slender pro-cesses (Figs. 3C and 3D). In the striatum of Mn-treatedrats the number of astrocytes and the characteristics oftheir processes did not show detectable differences ascomparing to striatal astrocytes of control animals(P . 0.05). Finally, the density of TH-positive nervefibers in the striatum (Figs. 3E and 3F), and the char-acteristics of TH-positive neurons in the substantianigra (Figs. 3G and 3H) were alike. Accordingly, theintensity of TH-immunoreactivity in the two areasshowed no important differences in control and Mn-treated animals (data not shown), as determined bymorphometric analysis (P . 0.05).
Intrinsic Membrane Properties of StriatalSpiny Neurons
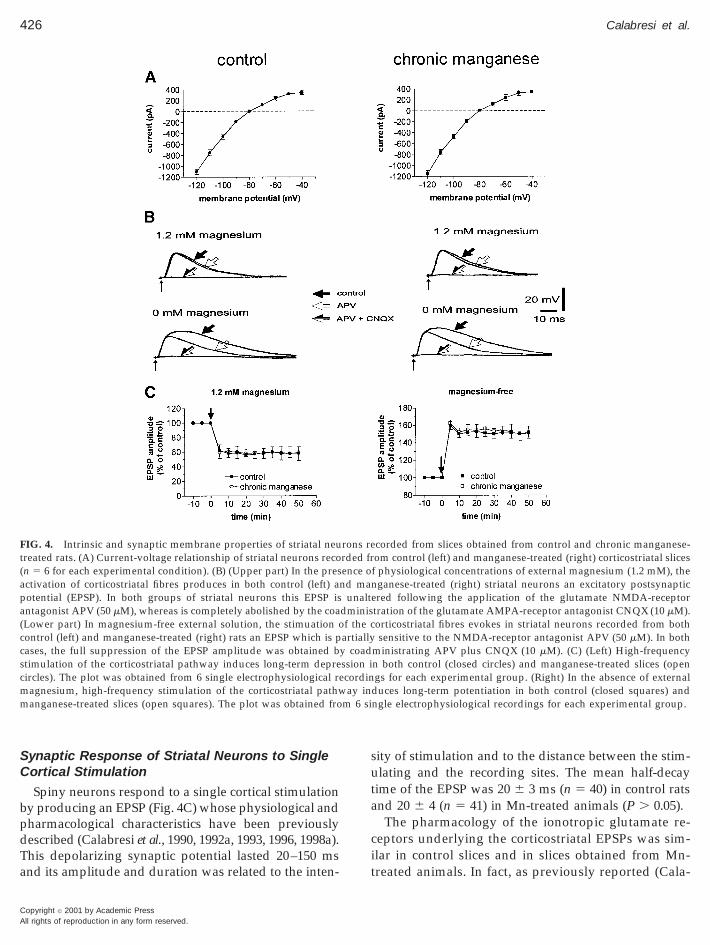
In the present study we included 121 striatal spinyneurons recorded from control rats and 123 spiny cellsobtained from Mn-treated animals. In both groupsthese cells had high resting membrane potential(284 6 5 mV in control rats and 285 6 5 mV inMn-treated animals, P . 0.05), relatively low apparentinput resistance (38 6 8 MV in control rats and 39 6 7MV in Mn-treated animals, P . 0.05) when measuredat the resting potentials from the amplitude of small(,10 mV) hyperpolarizing electrotonic potentials, ac-tion potentials of short duration (1.1 6 0.3 ms incontrol rats and 1.1 6 0.4 ms in Mn-treated animals,P . 0.05), and high amplitude (102 6 4 mV in controlrats and 103 6 5 mV in Mn-treated animals, P . 0.05).These cells were silent at rest and showed membranerectification and tonic firing activity during depolar-izing current pulses.
All the cells from both groups, when studied withsingle microelectrode voltage-clamp, showed promi-nent inward rectification in the steady-state current-voltage relation (Fig. 4A). When the currents wereevoked by hyperpolarizing command steps, there wasno detectable time-dependent component. Some ofthese properties have been described previously andresemble those reported for intracellularly stainedspiny striatal neurons (Kita et al., 1984; Calabresi et al.,1990, 1992a, 1993, 1998a).
425Synaptic Mechanisms of Manganese Intoxication
FIG. 3. Microphotographs illustrating some cytological characteristics of striatum (A–F) and substantia nigra (G, H) of a control rat(photographs on left side) and of a rat treated (photographs on right side) with Mn. B, D, F, and H (Mn-treated) do not document obviousmorphological or cytochemical differences when compared to A, C, E, and G (control). (A, B) Nissl-stained sections of the striatum showingroundish or fusiform neuronal somata with densely stained cytoplasm and evident nucleolus (where present). Slight neuronal shrinkage andoccasional pericellular clear spaces in B are fixation artifacts. (C, D) Glial fibrillary acid protein (GFAP) immunostaining of the striatum,visualized using peroxidase-antiperoxidase complex reacted with diaminobenzidine. GFAP-positive astrocytes showed similar tiny cell bodyand slender processes in the control and the Mn-treated rat. (E, F) Tyrosine hydroxilase (TH) immunostaining in the striatum visualized usingperoxidase-antiperoxidase complex reacted with diaminobenzidine. Close texture of TH-positive fibers and intensity of TH-positive immu-nostaining are alike in control and treated rat. White matter striatal bundles are characteristically unstained. (G, H) Tyrosine hydroxilase (TH)immunostaining of midbrain sections visualized using peroxidase-antiperoxidase complex reacted with diaminobenzidine. Overview ofsubstantia nigra (arrow) and ventral tegmental area (double arrow). Morphological characteristics of TH-positive neurons are not perceivableat this low magnification, but a strong and highly specific TH-positive reaction product is evident throughout comparable midbrain sectionsof control and Mn-treated rat. Bars: A, B, 20 mm; C–F, 10 mm; G, H, 200 mm.
Copyright © 2001 by Academic PressAll rights of reproduction in any form reserved.
(
(c
426 Calabresi et al.
A
Synaptic Response of Striatal Neurons to SingleCortical Stimulation
Spiny neurons respond to a single cortical stimulationby producing an EPSP (Fig. 4C) whose physiological andpharmacological characteristics have been previouslydescribed (Calabresi et al., 1990, 1992a, 1993, 1996, 1998a).This depolarizing synaptic potential lasted 20–150 msand its amplitude and duration was related to the inten-
FIG. 4. Intrinsic and synaptic membrane properties of striatal neutreated rats. (A) Current-voltage relationship of striatal neurons recon 5 6 for each experimental condition). (B) (Upper part) In the pres
activation of corticostriatal fibres produces in both control (left) apotential (EPSP). In both groups of striatal neurons this EPSP isantagonist APV (50 mM), whereas is completely abolished by the coaLower part) In magnesium-free external solution, the stimuation oontrol (left) and manganese-treated (right) rats an EPSP which is p
cases, the full suppression of the EPSP amplitude was obtained bstimulation of the corticostriatal pathway induces long-term deprecircles). The plot was obtained from 6 single electrophysiological remagnesium, high-frequency stimulation of the corticostriatal pathwmanganese-treated slices (open squares). The plot was obtained fro
Copyright © 2001 by Academic Pressll rights of reproduction in any form reserved.
sity of stimulation and to the distance between the stim-ulating and the recording sites. The mean half-decaytime of the EPSP was 20 6 3 ms (n 5 40) in control ratsand 20 6 4 (n 5 41) in Mn-treated animals (P . 0.05).
The pharmacology of the ionotropic glutamate re-ceptors underlying the corticostriatal EPSPs was sim-ilar in control slices and in slices obtained from Mn-treated animals. In fact, as previously reported (Cala-
corded from slices obtained from control and chronic manganese-om control (left) and manganese-treated (right) corticostriatal slicesf physiological concentrations of external magnesium (1.2 mM), thenganese-treated (right) striatal neurons an excitatory postsynapticered following the application of the glutamate NMDA-receptortration of the glutamate AMPA-receptor antagonist CNQX (10 mM).orticostriatal fibres evokes in striatal neurons recorded from bothsensitive to the NMDA-receptor antagonist APV (50 mM). In both
ministrating APV plus CNQX (10 mM). (C) (Left) High-frequencyn both control (closed circles) and manganese-treated slices (opengs for each experimental group. (Right) In the absence of external
duces long-term potentiation in both control (closed squares) andngle electrophysiological recordings for each experimental group.
rons rerded frence ond ma
unaltdminisf the cartiallyy coadssion icordinay in
m 6 si

fpwgvc
g
tsai
(WccdsapgTlnllttg
EC
ndti
astraciFtp1dr0
qiam
427Synaptic Mechanisms of Manganese Intoxication
bresi et al., 1996), the subthreshold EPSPs recordedrom spiny neurons of both groups in the presence ofhysiological concentrations of magnesium (1.2 mM)ere not affected by 50 mM APV (n 5 5 for each
roup), a NMDA glutamate receptor antagonist. Con-ersely, EPSPs were almost completely abolished byoadministration of 50 mM APV plus 10 mM CNQX, an
AMPA glutamate receptor antagonist (n 5 5 for eachroup) (Fig. 4B, upper row).In the absence of external magnesium the corticos-
riatal EPSP recorded from both groups of animalshowed a significant APV-sensitive component. Themplitude and duration of this component was similarn both groups of animals (n 5 5 for each group, P .
0.05; Fig. 4B, lower row), suggesting that neither theAMPA nor the NMDA-mediated component of thecorticostriatal EPSP were altered by Mn treatment.
Synaptic Plasticity Evoked in Striatal Neurons byRepetitive Corticostriatal Stimulation
We also investigated the possible effect of chronicMn treatment on two opposite forms of striatal syn-aptic plasticity evoked by tetanic stimulation of corti-costriatal pathway in vitro. As previously reportedCalabresi et al., 1992a, 1996; Lovinger et al., 1993;
alsh, 1993) in the presence of physiological externaloncentration of magnesium tetanic stimulation oforticostriatal afferents (3 trains, 100 Hz frequency, 3-suration, 20-s interval) induced a long-term depres-ion (LTD) of EPSP amplitude in both control slicesnd slices obtained from Mn-treated animals. The am-litude and the duration of LTD were similar in bothroups (n 5 6 for each group, P . 0.05) (Fig. 4C, left).he same protocol of repetitive corticostriatal stimu-
ation, when delivered in the absence of external mag-esium to deinactivate NMDA receptors, induced
ong-term potentiation (LTP) of EPSP amplitude (Ca-abresi et al., 1996). Also in this case the amplitude andhe duration of this long-term synaptic change werehe same in both groups of animals (n 5 6 for eachroup, P . 0.05) (Fig. 4C, right).
ffect of D2 Dopamine Receptor Agonists onorticostriatal Synaptic Transmission
Although the experiments analyzing the TH-immu-oreactivity in the substantia nigra and in the striatumid not reveal any significant morphological change of
he nigrostriatal dopaminergic pathway following Mnntoxication, it is still possible that a functional alter-
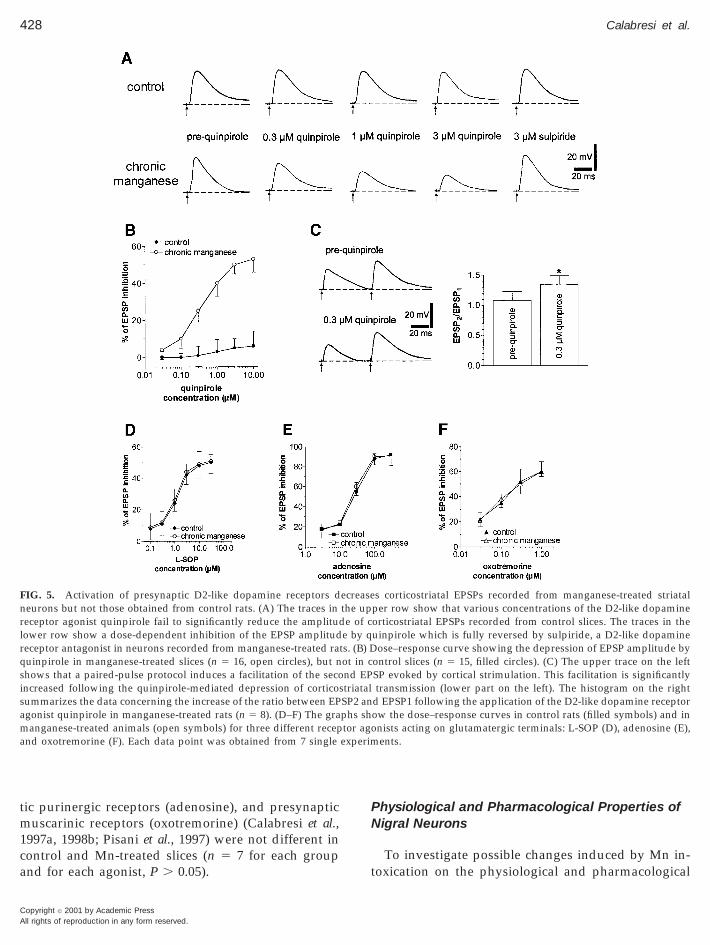
tion of the dopaminergic system occurs within thetriatum. This potential alteration might account forhe behavioral abnormalities observed in Mn-treatedats. For this reason, we have investigated whether thectivation of D2-like DA receptors by quinpiroleauses different effects on corticostriatal transmissionn Mn-treated rats and in control animals. As shown inigs. 5A and 5B, application of this D2-like DA recep-or agonist induced minor changes on the EPSP am-litude recorded from control striatal neurons (n 55). Conversely, quinpirole induced a significant andose-dependent decrease of EPSP amplitude in neu-ons recorded from Mn-treated rats (n 5 16, P ,.001). This inhibitory effect was fully reversed by 3
mM sulpiride, a D2-like DA receptor antagonist (n 5 8,P , 0.01). Moreover, the effect of quinpirole on theEPSP amplitude was coupled neither to significantchanges of membrane potential (n 5 16, P . 0.05) norto alteration of input resistance of the recorded cells(n 5 16, P . 0.05).
In order to investigate whether this pharmacologi-cal change observed in Mn-treated rats involved pre-or postsynaptic effects, we measured synaptic re-sponses to a pair of stimuli before and during theapplication of quinpirole. In these experiments theinterstimulus interval was 50–60 ms. Paired-pulsemodification of neurotransmission has been studiedextensively and is attributed to a presynaptic changein release probability (Manabe et al., 1993; Schultz etal., 1994). An increase in the ratio of the second pulseresponse to the first pulse response (EPSP2/EPSP1)indicates a decrease in release probability. The de-crease in transmitter release probability is consistentwith the observation that manipulations depressingtransmitter release usually increase the magnitude ofthis ratio also at corticostriatal synapses (Calabresi etal., 1997a). As shown in Fig. 5C, the decrease of theEPSP amplitude induced by quinpirole in Mn-treatedrats was coupled with a significant (n 5 8, P , 0.05)increase of paired pulse-facilitation as expected by aprevalent presynaptic site of action.
In agreement with this hypothesis we also foundthat in slices obtained from treated rats the membranedepolarizations induced by exogenous glutamate (1mM, 0.5 s of focal application by a micropipette, pres-sure ejection) were not significantly altered by 1–3 mM
uinpirole (n 5 6, P . 0.05, data not shown). Interest-ngly, the sensitivity of corticostriatal EPSPs to thepplications of other agonists acting on presynapticetabotropic glutamate receptors (L-SOP), presynap-
Copyright © 2001 by Academic PressAll rights of reproduction in any form reserved.
ca
PN
t
nrlrqsisama
428 Calabresi et al.
A
tic purinergic receptors (adenosine), and presynapticmuscarinic receptors (oxotremorine) (Calabresi et al.,1997a, 1998b; Pisani et al., 1997) were not different inontrol and Mn-treated slices (n 5 7 for each groupnd for each agonist, P . 0.05).
FIG. 5. Activation of presynaptic D2-like dopamine receptors deurons but not those obtained from control rats. (A) The traces ineceptor agonist quinpirole fail to significantly reduce the amplitudower row show a dose-dependent inhibition of the EPSP amplitudeceptor antagonist in neurons recorded from manganese-treated rauinpirole in manganese-treated slices (n 5 16, open circles), but nhows that a paired-pulse protocol induces a facilitation of the seconcreased following the quinpirole-mediated depression of corticoummarizes the data concerning the increase of the ratio between EPgonist quinpirole in manganese-treated rats (n 5 8). (D–F) The graanganese-treated animals (open symbols) for three different recep
nd oxotremorine (F). Each data point was obtained from 7 single
Copyright © 2001 by Academic Pressll rights of reproduction in any form reserved.
hysiological and Pharmacological Properties ofigral Neurons
To investigate possible changes induced by Mn in-oxication on the physiological and pharmacological
s corticostriatal EPSPs recorded from manganese-treated striatalper row show that various concentrations of the D2-like dopamineorticostriatal EPSPs recorded from control slices. The traces in theuinpirole which is fully reversed by sulpiride, a D2-like dopamine
ose–response curve showing the depression of EPSP amplitude byontrol slices (n 5 15, filled circles). (C) The upper trace on the leftSP evoked by cortical strimulation. This facilitation is significantlytransmission (lower part on the left). The histogram on the right
d EPSP1 following the application of the D2-like dopamine receptorow the dose–response curves in control rats (filled symbols) and innists acting on glutamatergic terminals: L-SOP (D), adenosine (E),ents.
ecreasethe upe of c
e by qts. (B) Dot in cnd EP
striatalSP2 anphs shtor agoexperim

tp
(itd
429Synaptic Mechanisms of Manganese Intoxication
properties of single nigral neurons, we intracellularlyrecorded putative dopaminergic neurons from controlslices (n 5 15) and from slices obtained from treatedrats (n 5 18). The active and passive membrane prop-erties of these neurons recorded from both groupscould not be distinguished and were similar to thosepreviously described for presumed substantia nigraDA containing cells (Lacey et al., 1987). These cellsfired action potentials spontaneously at a rate of 2.2 60.3 Hz in control rats and of 2.1 6 0.2 in Mn treatedanimals (P . 0.05). The mean peak input resistancewas 167 6 15 MV in control rats and 166 6 14 MV inreated animals as measured within 200 ms of a hy-erpolarizing pulse (P . 0.05). In both groups a time-
dependent inward rectification was observed with thehyperpolarization of the membrane (Lacey et al., 1987).
A characteristic feature of these cells is the hyper-polarizing response to DA. This response involves D2DA autoreceptors and it is caused by an increasedpotassium conductance (Lacey et al., 1987; Mercuri etal., 1989, 1997). In order to test whether Mn treatmentaltered D2-mediated neuronal responses to applica-tion of exogenous DA, we studied the membranechanges induced by various concentrations of DA incontrol slices and in slices obtained from treated ani-mals. As shown in Fig. 6, DA inhibited spontaneousfiring discharge and induced a dose-related hyperpo-larization. The amplitude and the duration of the DA-induced hyperpolarizations were similar in the twogroups (n 5 10 for each group, P . 0.05), indicatingthat the Mn intoxication alters the sensitivity of D2-like receptors controlling the release of glutamate butnot the function of nigral D2 autoreceptors involved inthe regulation of DA release.
DISCUSSION
Mn Intoxication Causes a Disinhibitory BehavioralSyndrome in the Absence of Major MotorAlterations and Learning Deficits: A Model for the“Early” Clinical Phase of Manganism?
Classical clinical studies clearly indicate that thepsychiatric symptoms may appear in the “early”phase of manganism in the absence of motor effects(Rodier, 1955; Cotzias, 1958). Conversely, the latterdisorders are more frequently reported in the “estab-lished” phase. Motor effects have been clearly attrib-uted to a basal ganglia dysfunction while the possibleinvolvement of other motor systems, such as cerebel-lum or cortex, has been ruled out (Rodier, 1955; Cot-
zias, 1958; Mena et al., 1969; Barbeau et al., 1976; Bar-beau, 1984; Calne et al., 1994).
The anatomical and physiological mechanisms un-derlying the “early” emotional disorders are morecomplex to investigate and for this reason they re-mained largely unknown. Interestingly, a critical re-view of studies dealing with nonhuman primatesclearly shows that subtle features of behavior are af-fected in ways that cannot be explained by the classi-cal hypothesis of the anatomical lesions of basal gan-glia structures. In fact, no gross impairment of thesestructures was reported in several studies (Newland,1999). Our study has clearly addressed this issue dem-onstrating, in Mn-treated rats, the occurrence of acomplex behavioral syndrome in the absence of sig-nificant striatal neuronal loss and gliosis. We found, infact, that rats exposed to Mn were significantly moreactive than control animals in the empty open fieldand did not show habituation. Defective habituationwas also observed when rearings were measured. The
FIG. 6. Chronic manganese treatment does not affect the mem-brane responses to dopamine in nigral neurons. (A) The histogramsshow that dopamine hyperpolarizes nigral neurons recorded fromboth control animals (left) and manganese-treated rats (right) in adose-dependent manner. (B) The electrophysiological traces showthat 30 mM dopamine hyperpolarizes to a similar extent both controlleft) and manganese-treated nigral neurons (right). In both exper-mental groups this effect blocks the spontaneous firing activity ofhe neurons and is reversible at the washout of the agonist. Theotted line indicates the extimated RMP of the cells (255 mV). Note
that the full action potential amplitude has not been captured by thechart record.
Copyright © 2001 by Academic PressAll rights of reproduction in any form reserved.

1rwmdoname
DDGA
obt(
wtsoMp1dtlcCcgccracwegs
itftiie
p
430 Calabresi et al.
A
reactivity to object novelty showed a progressive in-crease of contacts across sessions in rats exposed toMn and boluses were more numerous in treated ani-mals along the whole experiment. This pattern of be-havioral alterations associating increased activity, de-fects in habituation, reduced neophobia, and intensedefecation strongly suggests an hyperactive behavior.An altered behavior in Mn-treated rats was also sug-gested by the constant high radial maze-runningspeed together with the limited number of 45° angleturns. Interestingly, radial maze performance analysisshowed that correct choices increased as training pro-ceeded in both controls and Mn-treated rats suggest-ing that learning procedures and spatial memory abil-ities were not impaired following manganism. It istherefore apparent that rats exposed to Mn do notdiffer from controls in choice accuracy, a variablemore specifically estimating the cognitive aspects ofthe radial maze task.
This behavioral finding is in good agreement withthe electrophysiological observation that the twoforms of striatal synaptic plasticity (LTD and LTP)were unaltered in slices from Mn-treated rats. Thesecellular events, in fact, have been considered possiblesynaptic substrates for some forms of learning andmemory (Calabresi et al., 1996, 1997b).
It is also worth noting that an intact dopaminergicprojection is required for both striatal LTD (Calabresiet al., 1992a) and LTP (Centonze et al., 1999) as well asfor various learning procedures involving the basalganglia (Sahakian et al., 1988; Brown and Marsden,991; Dubois et al., 1991). Thus, the anatomical integ-ity of the nigrostriatal pathway in Mn-treated ratsell fits with the behavioral findings suggesting intactemory abilities and with the electrophysiological
ata describing normal synaptic plasticity. Moreover,ur morphological observation of a normal TH-immu-oreactivity also confirms previous works describingn intact dopaminergic projection in Mn-intoxicatedonkeys (Olanow et al., 1996) and humans (Shinotoh
t al., 1997).
oes a Selective Dysfunction of D2-Likeopamine Receptors Controlling the Striatallutamate Release Account for Behaviouralbnormalities in Manganism?
D2 receptors are pre- and postsynaptically locatedn striatal spiny neurons, and their activation seems toe involved in the control of two main forms of cor-icostriatal synaptic plasticity such as LTD and LTPCalabresi et al., 1997b). Since neither LTD nor LTP
Copyright © 2001 by Academic Pressll rights of reproduction in any form reserved.
ere altered following Mn intoxication we can arguehat D2 DA receptors required for these forms ofynaptic plasticity do not play a role in the generationf the behavioral effects caused by this treatment.oreover, D2 DA autoreceptors located on nigral do-
aminergic neurons (Lacey et al., 1987; Mercuri et al.,989, 1997) do not seem to play a role in the behavioralisinhibition following manganism because the ampli-
ude and the time-course of the membrane hyperpo-arization induced by DA in these cells were un-hanged in slices obtained from Mn-intoxicated rats.onversely, it seems that D2-like receptors located on
orticostriatal terminals and controlling the release oflutamate within the striatum (Maura et al., 1988) areritically important in the Mn-induced behavioralhanges. In fact, in Mn-treated rats, unlike in controlats, activation of D2-like DA receptors reduces themplitude of glutamate-mediated EPSPs evoked byortical stimulation. Since this effect was not coupledith significant changes of intrinsic membrane prop-
rties and with modifications of exogenously appliedlutamate, we can hypothesize that this action is pre-ynaptically mediated.
A further support to the idea that the quinpirole-nduced decrease of the EPSP amplitude in Mn-reated rats is presynaptically mediated originatesrom the experiments utilizing the paired-pulse pro-ocol. These experiments, in fact, have shown that thenhibitory effect of quinpirole is associated with anncreased paired-pulse facilitation, which is consid-red an index of presynaptic activity (Manabe et al.,
1993; Schultz et al., 1994; Calabresi et al., 1997a). Inter-estingly, we have observed similar quinpirole-in-duced presynaptic effects at corticostriatal terminals ineither 6-hydroxy-dopamine-denervated rats (Cala-bresi et al., 1993) or animals exposed to chronic halo-
eridol treatment (Calabresi et al., 1992b). Both theseexperimental conditions may induce supersensitivityof D2-like DA receptors as a consequence of the long-term impairment of the dopaminergic system (Saave-dra et al., 1978; Mac Lennan et al., 1988). Thus, theabnormal effect of quinpirole in these two conditionsmight be caused by an adaptive receptor change.
At this point, however, a simple question can beraised. Why do Mn-treated rats not show evident mor-phological or functional alterations of the dopaminer-gic system but reveal an abnormal sensitivity of theD2-like DA receptors controlling glutamate release?At present, we do not have a conclusive explanationfor this event. We can hypothesize that chronic Mn-treatment does not induce gross anatomical changesof the nigrostriatal projection but it might alter the

t
B
B
C
C
C
C
C
C
C
C
C
C
C
C
C
C
C
431Synaptic Mechanisms of Manganese Intoxication
release and/or the life-time of DA within the striatalextracellular space leading to an altered expression ofthis selective class of D2-like DA receptors. Alterna-tively, it is possible that the increased response follow-ing the activation of D2-like DA receptors located oncorticostriatal terminals is a compensatory mechanismto counteract an excessive release of glutamate in-duced by Mn-intoxication. This latter explanationseems to suggest the hypothesis that the motor andbehavioral signs induced by manganism result froman excitotoxic mechanism (Brouillet et al., 1993).
It should be noted that in manganism the increasedinhibitory control on the glutamatergic transmissionselectively affects D2-like DA receptors. In fact, Mn-treatment does not alter the sensitivity of other recep-tors also involved in the inhibitory control of gluta-mate release within the striatum such as A1 adenosinereceptors (Calabresi et al., 1997a), group III metabo-ropic glutamate receptors (Pisani et al., 1997), and
M2-like muscarinic receptors (Calabresi et al., 1998b).
Conclusion
In our model we hypothesize that cortical glutama-tergic inputs to the striatum are greatly reduced by anaugmented activity of presynatic D2-like DA receptorslocated on corticostriatal axon terminals. This abnor-mal dopaminergic influence may relieve the basal gan-glia from the cortical control leading to the observedbehavioral disinhibition. Interestingly, an abnormalinhibitory dopaminergic control on corticostriatal glu-tamatergic inputs has been previously proposed toplay a critical role in schizophrenia (Carlsson & Carls-son, 1990). Further studies are required to investigatehow the synaptic changes detected at the early stage ofmanganism can lead to the anatomical abnormalitiesunderlying the motor signs observed in the estab-lished phase of Mn intoxication.
ACKNOWLEDGMENTS
We thank M. Tolu for technical assistance. This study was sup-ported by a grant from Ministero del Lavoro (Italy) to G.B. (RicercaNo. 629) and by a grant from Telethon Project to P.C. (No. E.729).
REFERENCES
Ammassari-Teule, M., Fagioli, S., & Rossi-Arnaud, C. (1994) Radialmaze performance and open field behaviors in aged C57BL/6mice: Evidence for preserved cognitive abilities during senes-cence. Physiol. Behav. 55, 341–345.
Barbeau, A., Inoue, N., & Cloutier, T. (1976) Role of manganese indystonia. Adv. Neurol. 14, 339–352.
Barbeau, A. (1984) Manganese and extrapiramida disorders. Neuro-toxicology 1, 13–36.
rouillet, E. T., Shinovu, L., McGarvey, U., Hochberg, F., & Beal,M. F. (1993) Manganese injection into the rat striatum producesexcitotoxic lesions by impairing energy metabolism. Exp. Neurol.120, 89–94.
rown, R. J., & Marsden, C. D. (1991) Dual task performance andprocessing resourches in normal subjects and patients with Par-kinson’s disease. Brain 114, 215–231.
alabresi, P., Mercuri, N. B., & Bernardi, G. (1990) Synaptic andintrinsic control of membrane excitability of neostriatal neurons.II. An in vitro analysis. J. Neurophysiol. 63, 663–675.
alabresi, P., Maj, R., Pisani, A., Mercuri, N. B., & Bernardi, G.(1992a) Long-term synaptic depression in the striatum: Physio-logical and pharmacological characterization. J. Neurosci. 12,4224–4233.
alabresi, P., De Murtas, M., Mercuri, N. B., & Bernardi, G. (1992b)Chronic neuroleptic treatment: D2 dopamine receptors supersen-sitivity and striatal glutamatergic transmission. Ann. Neurol. 31,366–373.
alabresi, P., Mercuri, N. B., Sancesario, G., & Bernardi, G. (1993)Electrophysiology of dopamine-denervated striatal neurons: Im-plications for Parkinson’s disease. Brain 116, 433–452.
alabresi, P., Pisani, A., Mercuri, N. B., & Bernardi, G. (1996) Thecorticostriatal projection: From synaptic plasticity to dysfunctionsof the basal ganglia. Trends Neurosci. 19, 19–24.
alabresi, P., Centonze, D., Pisani, A., & Bernardi, G. (1997a) En-dogenous adenosine mediates mediates the presynaptic inhibi-tion induced by aglycemia at corticostriatal synapses. J. Neurosci.17, 4509–4516.
alabresi, P., Saiardi, A., Pisani, A., Baik, J-H., Centonze, D., Mer-curi, N. B., Bernardi, G., & Borrelli, E. (1997b) Abnormal synapticplasticity in the striatum of mice lacking dopamine D2 receptors.J. Neurosci. 17, 4536–4544.
alabresi, P., Centonze, D., Pisani, A., Sancesario, G., Gubellini, P.,Marfia, G. A., & Bernardi, G. (1998a) Striatal spiny neurons andcholinergic interneurons express differential ionotropic glutama-tergic responses and vulnerability: Implications for ischemia andHuntington’s disease. Ann. Neurol. 43, 586–597.
alabresi, P., Centonze, D., Pisani, A., & Bernardi, G. (1998b) Block-ade of M2-like muscarinic receptors enhances long-term potenti-ation at corticostriatal synapses. Eur. J. Neurosci. 10, 3020–3023.
alne, D. B., Chu, N. S., Huang, C. C., Lu, C. S., & Olanow, W. (1994)Manganism and idiopathic parkinsonism: similarities and differ-encies. Neurology 44, 1483–1486.
arlsson, M., & Carlsson, A. (1990) Interactions between glutama-tergic and monoaminergic systems within the basal ganglia—Implications for schizophrenia and Parkinson’s disease. TrendsNeurosci. 13, 272–276.
entonze, D., Gubellini, P., Picconi, B., Giacomini, P., Calabresi, P.,& Bernardi, G. (1999) Unilateral dopamine denervation blockscorticostriatal LTP. J. Neurophysiol. 82, 3575–3579.
ooper, W. C. (1984) The health implications of increased manga-nese in the environment resulting from the combustion of fueladditives: A review of the literature. J. Toxicol. Environ. Health 14,23–46.
otzias, J. C. (1958) Manganese in health and disease. Physiol. Rev.38, 503–532.
ouper, J. (1837) On the effects of black oxide of manganese wheninhaled into the lungs. Brit. Ann. Med. Pharmacol. 1, 41–42.
Copyright © 2001 by Academic PressAll rights of reproduction in any form reserved.

L
M
M
M
M
M
M
N
O
P
R
S
S
S
S
S
S
W
432 Calabresi et al.
A
Dubois, B., Boller, F., Pillon, B., & Agid, Y. (1991) Cognitive deficitsin Parkinson’s disease. In: Handbook of Neuropsychology (F. Bollerand J. Grafman, Eds.), Vol. 5, pp. 195–240. Elsevier, Amsterdam.
Fell, J. M. E., Reynolds, A. P., Meadows, N., Khan, K., Long, S. G.,Quaghebeur, G., Taylor, W. J., & Milla, P. J. (1996) Manganesetoxicity in children receiving long-term parenteral nutrition. Lan-cet 347, 1218–1221.
Gorell, J. M., Johnson, C. C., Rybicki, B. A., Peterson, E. L., Kortsha,G. X., Brown, G. G., & Richardson, R. J. (1997) Occupationalexposures to metals as risk factors for Parkinson’s disease. Neu-rology 48, 650–658.
Hauser, R. A., Zesiewicz, T. A., Rosemurgy, A. S., Martinez, C., &Olanow, C. W. (1994) Manganese intoxication and chronic liverfailure. Ann. Neurol. 36, 871–875.
Huang, C. C., Chu, N. S., Lu, C. S., Chen, R. S., & Calne, D. B. (1998)Long-term progression in chronic manganism. Neurology 50, 698–700.
Kawamura, R., Ikuta, H., Fukuzumi, S., Yamada, R., Tsubaki, S.,Kodama, T., & Kurata, S. (1941) Intoxication by manganese in wellwater. Kitasato Arch. Exp. Med. 18, 145–169.
Kita, T., Kita, H., & Kitai, S. T. (1984) Passive electrical membraneproperties of rat neostriatal neurons in an in vitro slice prepara-tion. Brain Res. 300, 129–139.
Lacey, M. G., Mercuri, N. B., & North, R. A. (1987) Dopamine acts onD2 receptors to increase potassium conductance in neurones ofthe rat substantia nigra zona compacta. J. Physiol. 392, 397–416.
Lovinger, D. M., Tyler, E. C., & Merrit, A. (1993) Short- and long-term synaptic depression in the neostriatum. J. Neurophysiol. 70,1937–1949.
owry, O. H., Rosebrough, N. J., Farr, A. L., & Randall, R. J. (1951)Protein measurements with the folin-phenol reagents. J. Biol.Chem. 193, 265–275.ac Lennan, J., Atmadja, S., Lee, N., & Fibiger, H. C. (1988) Chronichaloperidol administration increases the density of D2 dopaminereceptors in the medial prefrontal cortex of the rat. Psychopharma-cology 95, 255–257.anabe, T., Wyllie, D. J. I., & Nicoll, R. A. (1993) Modulation ofsynaptic transmission and long-term potentiation: Effects onpaired pulse facilitation and EPSC variance in the CA1 region ofthe hippocampus. J. Neurophysiol. 70, 1451–1459.aura, G., Giardi, A., & Raiteri, M. (1988) Release-regulating D2dopamine receptors are located on striatal glutamatergic nerveterminals. J. Pharmacol. Exp. Ther. 247, 680–684.ena, I., Horiuchi, K., Burke, K., & Cotzias, G. C. (1969) Chronicmanganese poisoning. Neurology 19, 1000–1036.
Copyright © 2001 by Academic Pressll rights of reproduction in any form reserved.
ercuri, N. B., Calabresi, P., & Bernardi, G. (1989) Physiology andpharmacology of dopamine D2-receptors: Their implications indopamine-substitutive therapy for Parkinson’s disease. Neurology39, 1106–1108.ercuri, N. B., Saiardi, A., Bonci, A., Picetti, R., Calabresi, P., Ber-nardi, G., & Borrelli, E. (1997) Loss of autoreceptor function indopaminergic neurons from dopamine D2 receptor-deficientmice. Neuroscience 79, 323–327.ewland, M. C. (1999) Animal model of manganese’s neurotoxicity.Neurotoxicology 20, 415–432.lanow, W., Good, P. F., Shinitoh, H., Hewitt, K. A., Vingerhoets, F.,Snow, B. J., Beal, M. F., Calne, D. B., & Perl, D. P. (1996) Manga-nese intoxication in the rhesus monkey: A clinical, imaging,pathologic, and biochemical study. Neurology 46, 492–498.
isani, A., Calabresi, P., Centonze, D., & Bernardi, G. (1997) Acti-vation of group III metabotropic glutamate receptors depressesglutamatergic transmission at corticostriatal synapse. Neurophar-macology 36, 845–851.
odier, J. (1955) Manganese poisoning in moroccan miners. Br. J.Ind. Med. 12, 21–35.
aavedra, J. M., Setler, P. E., & Kebabian, J. W. (1978) Biochemicalchanges accompanying unilateral 6-hydroxydopamine lesions inrat substantia nigra. Brain Res. 151, 339–352.
ahakian, B., Morris, R., Evenden, J., Heald, A., Levy, R., Philpot,M., & Robins, T. (1988) A comparative study of visuospatialmemory and learning in Alzheimer-type dementia and Parkin-son’s disease. Brain 111, 695–718.
aric, M. (1986) Manganese. In: Handbook on the Toxicology of Metals(I. Friberg, G. I. Nordberg, & V. Vouk, Eds.), 2nd ed, pp. 354–385.Elsevier, Amsterdam.
chulz, P. E., Cook, E. P., & Johnston, D. (1994) Changes in paired-pulse facilitation suggest presynaptic involvement in long-termpotentiation. J. Neurosci. 14, 5325–5337.
hinotoh, H., Snow, B. J., Hewitt, K. A., Pate, B. D., Doudet, D.,Nugent, R., Perl, D. P., Olanow, W., & Calne, D. B. (1995) MRI andPET studies of manganese-intoxicated monkeys. Neurology 45,1199–1204.
hinotoh, H., Snow, B. J., Chu, N-S., Huang, C-C., Lu, C-S., Lee, C.,Takahashi, H., & Calne, D. B. (1997) Presynaptic and postsynapticstriatal dopaminergic function in patients with manganese intox-ication: A positron emission tomography study. Neurology 48,1053–1056.alsh, J. P. (1993) Depression of excitatory input in rat striatalneurons. Brain Res. 608, 123–128.