a short history of eighty years of limnology - in spain - citeseerx
TRANSCRIPT







A SHORT HISTORY OF EIGHTY YEARS OF LIMNOLOGY IN SPAIN
S. Casado and C. Montes Laboratorio de Limnología. Departamento de Ecología. Universidad Autónoma de Madrid. 28049 Madrid. Spain.
Keywords: History of ecology, History of limnology, Laboratorio de Hidrobiología Española, Spain.
ABSTRACT
Historically both socioeconomical and ideological constraints have prevented a normal development of science in Spain. Ecological sciences such as limnology had in the past problems to attain an appropriate institutional framework, which only very recently has been created. In this context, special attention is deserved for early scientific projects in ecology such as that of Celso Arévalo, founder of the first center devoted to limnological research in Spain, the Laboratorio de Hidrobiología Española, in Valencia in 1912. Arévalo's approach was partially continued by his collaborator Luis Pardo at the Sección de Biología de las Aguas Continentales, in Madrid, which was one of the scientific centers asociated to the Corps of Forestry Engineers. After the Civil War of 1936 an important new period for limnology started with the works of Ramon Margalef, eventually resulting in a normalization of this science in Spain during the decades of the 1970's and 1980's.
INTRODUCTION
In 1912, eighty years ago, Celso Arévalo created in Valencia the Laboratorio Hidrobiológico, later called Labo- ratorio de Hidrobiología Española, which was the first center, and for many years the only one, devoted to limno- logical research in Spain. The word "limnology" reachs its centenary in 1992 and the international congress of limno- logy will be held in Spain this year. This anniversary is a good opportunity to look back to that early period of Spanish limnology, lasting until the Civil War of 1936, which to a large extent has remained unknown to most of the people involved in the development of limnology in Spain during the last two decades. Indeed, no attempt has been made to give a historical overview of Spanish limno- logy. The aim of this article is to provide a short account of this history, with special attention to the early times of limnological research, as an introduction to the currettt state of limnology in Spain, which is presented in detail in other contributions of this volutne.
Knowledge of previous periods of Spanish limrlology is bound to supply Liseful infurmation to present day resear- chers working in &he area of aquatic ecology in Spain, both on the historictii constraints that have affected its develop-
ment and on the conceptual background that explains our current approach in studying rivers, lakes and wetlands.
Economic, social and also ideological limitations have historically prevented a normal development of science in Spain as compared to other Western Europe countries, ecological science being no exception. In spite of the remar- kable process of modernization and improvement of scien- tific research that took place in Spain during the first third of twentieth century, ecology had a very limited reception in the small Spanish scientific community and suffered an almost complete lack of specialized centers in the institu- tional framework of science and research. The ecological approach was accepted by most Spanish naturalists of this period at a theoretical level but only occasionally was it used in specific scientific research. Lack of specialization resulted, with few though remarkable exceptions, in very poor standards in ecological research, a situation that remai- ned unchanged throughout the first half of the century. Only the early works of Ramon Margalef around 1945, devoted mostly to limnology of Iberian lakes and wetlands, involved a more ambitious approach to ecological research, as well as the first serious attempt to obtain a reasonable level of tatonomic and ecological knówledge of the species and communities of organisms of Spanish continental aquatic
Limnetica, 8 : 1-9 (1992) O Asociación Española de Limnologia, Madrid. Spain

ecosystems. Yet, the failures and achievements of the early period of Spanish limnology are worth examining in come detail, both as a part of the general picture of science in
Spain at the begining of the century and as a basis and histo- rica1 background to what is being done today in the same scientific and geographical area.
FIRST BIOLOGICAL AND GEOGRAPHICAL STUDIES ABOUT SPANISH CONTINENTAL WATERS
Prior to 191 2 there were no truly limnological research projects in Spain. Some scientifíc activities related with aquatic groups of organisms must be mentioned but even
those investigations were few and scattered in time. During
most of nineteenth century natural history went through a
critical, dark period in Spain. The Iberian peninsula became a favourite collecting ground for foreign naturalists, coming
from other European countries, who found here a largely unexplored and very rich natural territory, whereas national scientific production was extremely limited (GOMIS BLANCO, 1988).
Specific methodological requirements for collecting and identifying aquatic organisms prevented even more its
study'by Spanish naturalists. Only occasionally aquatic species of insects or plants and macroscopic algae appea- red in local or regional taxonomic inventories made by
Spanish zoologists and botanists. Yet, some aquatic groups with special appeal because of their morphological variety received some specific attention, such as freshwater
molluscs (GRAELLS, 1846; MACHO VELADO, 1878) or
diatoms. In fact, diatoms reached a certain popularity
among Spanish microscopists, the first record of a living diatom collected in Spain appearing as early as 1824 (AZPEITIA, 1908). But this scientific interest only focus-
sed on taxonomic and morphological aspects, without any relation with the study of aquatic habitats, as it can be infe- rred from the equal interest for fossil or living specimens shown by early Spanish diatomologists as Alfredo Truan
(1 833- 1890), Florentino Azpeitia (1 859-1934) or Ernesto
Caballero (1858-1935). Other groups of microscopic algae
received almost no attention during that time. Freshwater malacology was particularly developed in
Catalonia, with the works of Manuel de Chia (1856-1917), Artur Bofill ( 1 846- 1929) and Joan Baptista d' Aguilar-Amat ( 1 882- 1936). During the World War of 19 14 this Catalonian school of malacologists was joined by German specialist F.
Haas, who did a very valuable work in collaboration with local naturalists.
Ichthyology must also be mentioned because of some valuable works containing information about the fishes of
Spanish rivers and wetlands and related environmental observations, as that of CISTERNAS (1887) on the conti-
nental ichthyofauna of the Valencia region. In relation to
this it is interesting to note the early developments of aqui-
culture made by Mariano de la Paz Graells (1809-1898),
who wrote a whole book on the subject (GRAELLS, 1864) and created the first fish farming installation in Spain at La Granja, Segovia, in 1866. As will be seen later, ichthyology
and fish farming played an important role in the first stages of institutional development of limnology in Spain.
From a different point of view, other naturalists, such as geologist Salvador Calderón (1 85 1 - 19 1 l ) , became interes-
ted in the "steppic" features of Iberian landscape, including saline wetlands. Calderón, who studied the saline minerals of Spain, did geological research, postulated geochemical
explanations about the origin of these saline wetlands and
collected specimens of their peculiar biota (CALDERON, 1888, 1889). Botanist Eduardo Reyes Prósper (1860-1920) also directed his attention towards the steppic flora of
Central and Southern Spain regions, following the geobota-
nical interpretation of Iberian vegetation first proposed by WILLKOMM (1852). He published a monograph on the Charophyta of Spain, "particularly those growing in its step- pes" (REYES PROSPER, 1910), and a general description
of Spanish "steppic" flora, soils and wetlands (REYES PROSPER, 19 15).
Mountain lakes associated to glacial morphology also
attracted the attention of some naturalists. In 1912, an important year in the history of Spanish limnology for seve- ral reasons, two different scientific expeditions surveyed Sanabria lake, in Northwestern Spain. The first expedition was led by German geographer W. Halbfass, who took
detailed measurements of lake morphology and temperature and sampled phytoplankton. Some weeks later Spanish natu- ralist José Taboada made similar observations. The works
of TABOADA TUNDIDOR (1913) and HALBFASS
(1913), even though both dealt mainly with physical featu-
res, were the first truly limnological studies carried out in Spain to be published.
Nevertheless, the dominating feature of this early period was an overwhelming lack of original research and reliable information on aquatic biota and ecological features of Iberian continental waters. For example, at the end of nine- teenth century faunistic data of such an important group as

crustaceans hardly included more than half a dozen species scattered in a few localities (BUEN, 1887). Only in the
decade of 1950's, with the works of Margalef, a general knowledge of Spanish aquatic communities would be
reached. A number of associations were created precisely to tackle
these scientific gaps of Spanish natural history, such as the Sociedad Española de Historia Natural, the Institució Cata-
lana d1Hist6ria Natural or the Sociedad Aragonesa de Cien-
cias Naturales. The first and most important of this associa-
tions of naturalists was the Sociedad Española de Historia
Natural, founded in 1871. One of the major achievements of this group of scientists was the creation in 1886 of the Esta-
ción de Biología Marítima, in Santander, the first oceano-
graphical center established in Spain.
CELSO AREVALO AND THE LABORATORIO DE HIDROBIOLOGIA ESPANOLA
As a young graduate Celso Arévalo (1885-1944) obtai-
ned two scholarships for training periods at the Estación de
Biología Marítima in 1904 and 1905. In 1912, when Arévalo
became Professor of Natural History of the Instituto General
y Técnico de Valencia, a secondary school center, he deci- ded to establish a hydrobiological laboratory there. In doing this Arévalo was influenced not only by his training at a coastal biological station but also by his knowledge of
limnological stations placed at the shore of lakes in diffe-
rent European countries, which were taken as a model for
his project. The city of Valencia is located next to an impor-
tant coastal lagoon, named L'Albufera, and Arévalo, who
was an ambitious and gifted young scientist, saw the oppor- tunity to introduce his country to a kind of study that had
not been developed at that time in Spain. Arévalo's project was in fact the first ecologically oriented resarch programme to be initiated in Spain, a merit that has received almost no
recognition in recent times. Only MARGALEF (1983) and
CAMARASA (1 989) mention the work of Arévalo, though only regarding it with limited interest.
It must be recognized that the scientific results of the
hydrobiological laboratory were not as satisfactory as
Arévalo had first intended. Several reasons can be pointed
out to explain this partial failure when examining the history of the laboratory (AREVALO, 1914, 1929; PARDO, 1942, 1945, 1952). First, the lack of official and financial support, resulting in practica1 problems of al1 kinds. No full time senior research post was ever obtained for the laboratory. In
19 12, when Arévalo placed some scientific tools at a corri-
dor of the Instituto, the Laboratorio Hidrobiológico was just a personal project of his. In 19 14 the laboratory was offi- cially incorporated as a department of the Instituto General y Técnico de Valencia. In 19 16 Arévalo promoted the publi-
cation of a journal, Anales del Instituto General y Técnico
de Vulrncia, including a scientific series of "hydrobiological works", of which 33 were published up to 1928. Finally, in
19 17, due to the prestige gained by Arévalo during those
years, the government gave the laboratory the status of a national research center, with the name of Laboratorio de Hidrobiología Española. But no funds were delivered nor
suitable rooms and equipment were provided for the new
center. Another kind of problem seems to have been related with
the reluctance shown by the Spanish community of natura-
lists to accept an ecological research project such as Arévalo's. With so much yet to be done in order to complete the taxonomic and geographical knowledge of Iberian fauna
and flora it seemed that ecology was not a priority. In 1919 Arévalo moved to the Instituto del Cardenal
Cisneros, in Madrid, always as a Professor of Natural
History. Then, in the most important national research
center in the field of natural sciences, the Museo de Cien- cias Naturales, a new section devoted to hydrobiology was
created for him. The laboratory was mantained in Valencia as a field station associated to this new hydrobiological section of the museum. Luis Pardo (1897-1957), a disciple of Arévalo who had become his closest collaborator, was
designated keeper of the laboratory, with a status of assis-
tant researcher paid by the museum. In Madrid, another
assistant researcher, Manuel Ferrer Galdiano was also incor- porated into the hydrobiological section, suposedly to help
Arévalo in his ecological work. Instead, Ferrer Galdiano's wirk was devoted mostly to the taxonomy of freshwater
crustaceans and no financial support was given to Arévalo and neither to Pardo. Personal conflicts between Arévalo and other researchers at the Museo de Ciencias Naturales
could also have been influential in this situation, that ended
in 1932 when the hydrobiological section of the museum was finally removed. Earlier, in 1928, Pardo had moved to Madrid for another job and the laboratory in Valencia had
virtually ceased to exist. During the first years in Valencia Arévalo had concen-
trated on the study oí' plankton at L'Albufera lagoon (AREVALO, 19 16, 19 17), an endevour that required consi- derable effort in solving methodological and taxonomical probleins, considering that there was no previous experience

in Spain. Twenty years after F. A. Forel had first proposed the word limnology, but ten years before E. Naumann and
A. Thienemann created the SIL, Arévalo tried to fill the gap in Spanish science as far as limnology was concerned. Not only did he work in taxonomical and ecological studies but
he also made some theoretical and conceptual work, striving to establish a "hydrobiology", the word he chose (ARE-
VALO, 1921), with a conceptual framework that was fit for Spanish aquatic ecosystems, quite different from those of
Central Europe and Eastern North America, where limno- logy had been developed for the most part.
Whereas oceanography had received some support in Spain from governmental authorities (the Instituto Español de Oceanografía was founded in 1914, grouping several already existing coastal stations) small Spanish rivers and wetlands, with temporary and fluctuating regimes typical of semiarid climates, seemed of little economic and scientific interest to the politicians and leading scientists. Trying to get financia1 and institutional support for his project,
Arévalo claimed that, though not comparable in size or
volume to the lakes and rivers of other countries, Spanish continental waters had their own scientific and economic
values (AREVALO, 1921). A good knowledge of leading scientific languages, espe-
cially German, and contacts with most important limnologi- cal centers of Europe, some of which he visited personally in France, Belgium, Italy, Switzerland and Germany, made
it possible for Arévalo to have a sound and up to date know- ledge of the main trends of limnology of that period, being
himself particularly influenced by Thienemann's work. Arévalo attended several SIL congresses and was designated
SIL delegate in Spain. Another source of international contacts for Arévalo was the presence of foreign scientists in Spain seeking a safe place to stay during the First World War of 1914. Among them there were several researchers
who took advantage of Arévalo's laboratory in Valencia, as Swiss ichthyologist spccialized in eel A. Gandolfi, German malacologist F. Haas and, indirectly, German specialist in Hydracarina K. Viets. Part of their papers based on Spanish
specimens can be found among the "hydrobiological works" published in the Anales del Instituto General y Técnico de Valencia, the journal of the center where the laboratory was located.
Once in Madrid, integrated in such an important center as the Museo de Ciencias Naturales, Arévalo thought it was time to adopt a more ecological approach to his work, focu- sing on what he had always regarded as a central issue in limnology, temporal and spatial variation of plankton
communities, what he did studying a pond placed in a public park in Madrid (AREVALO, 1923a). But the isolated posi-
tion of Arévalo in the Spanish scientific community, and specially in the museum, was made evident when he attemp-
ted to organize a limnological survey of a lake distant from Madrid, Carucedo lake. The expedition to Carucedo lake, due to very limited financial and technical support, produced
poor scientific results (AREVALO, 1923b). This virtually meant the end of Arévalo's limnological project. A final product of that project was his book La vida en las aguas dulces (1929), the first general treatise on limnology to be
published in Spain, and, in spite of his popularizing appro-
ach, an interesting work that includes in a summarized form many of his observations on Spanish wetlands and lakes.
Meanwhile Luis Pardo had moved to Madrid to work as editor of the Boletin de Caza y Pesca, an official journal
devoted to hunting and fishing issues, where he published many bibliographical notes on aquatic ecosystems and their
biota. Pardo kept Arévalo's idea of promoting the ecological
study of Spanish continental waters and the assessment of their natural resources and he worked hard to support and
popularize this view throughout his life.
LUIS PARDO AND THE SECCION DE BIOLOGIA DE LAS AGUAS CONTINENTALES
The participation of Forestry Engineers in the history of
Spanish limnology is strongly related to applied ichthyo- logy. The direction of the first fish farming installation,
created in 1866 by zoologist Graells as we have already seen, was soon undertaken by Forestry Engineer Rafael
Breñosa (1835-1916). Since then management of fishing resources of continental waters was a duty officially assig- ned to the Corps of Forestry Engineers. In 1929 the Labo-
ratorio de la Fauna Forestal, Piscicultura y Ornitología, crea- ted in 1918 with the shorter name of Laboratorio de la Fauna Forestal Española, published the first issue of a journal enti-
tled Revista de Biología Forestal y Limnología, of which several issues appeared containig various papers on Spanish ichthyofauna. In 1931 this laboratory was incorporated into
the Instituto Forestal de Investigaciones y Experiencias, a center that grouped together al1 official forestry research
activities. A special section devoted to limnology was then created, called Sección de Biología de las Aguas Continen- tales (PARDO, 1949), soon joined by Luis Pardo, who brought with him his important specialized library as well as valuable files with geographical, faunistic and floristic

data resulting from Pardo's previous dedication to limnolo- gical studies.
The Sección de Biología de las Aguas Continentales, directed by Forestry Engineer Luis Vélaz de Medrano, got its own building as well as an experimental fish farming installation, both built during the years 1932 to 1934 close to the Manzanares river, in the outskirts of Madrid. The most important scientific result of this center was precisely a limnological monograph of the Manzanares river (VELAZ DE MEDRAN0 & UGARTE, 1933), in which physicoche- mica1 and biological data were used to explain the distribu- tion of fish communities along the stream. Several other papers, mainly dealing with ichthyology and fish farming subjects, were also published during this period, lasting until 1936, when the Spanish Civil War began.
With General Franco's troops close to the area where the center was placed, Vélaz de Medrano tried to move the section to a safer location, but local military authorities denied the permission. Eventually, the scientific and assis- tant staff was forced to leave the building and uncontrolled groups stole and completely destroyed the library, files and tools (PARDO, 1949). Because of this destruction it is diffi- cult to evaluate how great the scientific potential of this center would have been.
At any rate, the Civil War interrupted an important process of modernization of Spanish science, not only regarding natural sciences but most aspects of culture and knowledge. It also meant, in the last instance, the end of the limnological tradition initiated by Arévalo. In spite of the enthusiasm that Pardo showed repeatedly for Franco's new regime and the officially claimed new era of scientific progress, and even though some research was carried out in the Sección de Biología de las Aguas Continentales after 1939, when the war was finished, the reality is that original scientific work in limnology was almost non-existent, as well as in many other fields of Spanish science.
Paradoxically, some naturalists who had been working in oceanographical sciences shifted to limnology when, forced by the adverse results of the war, went into exile and settled in Latin America, especially in México (REYES NEVA- RES, 1982), a country rich in lakes. This is the case of Enri- que Rioja (1895-1963), a leading specialist in marine bent- hic groups when working in Spain, who later dealt also with continental aquatic organisms in México, where he was chief of the department of hydrobiology at México Univer- sity. Fernando de Buen (1895-1962) worked first as ichth- yologist at the Instituto Español de Oceanografía and was later involved in the development of Mexican limnology
since he worked for several years at the Estación Limnoló- gica de Pátzcuaro.
In Spain, Pardo published many articles, monographs and books mostly devoted to applied issues. The most interes- ting of these publications is a comprehensive monograph on L'Albufera (PARDO, 1942), the lagoon in which Spanish limnology had started thirty years before. In this monograph Pardo explicitly adopted the same descriptive approach that Forel had initiated with his study of lake Léman, but intro- ducing also human aspects, both cultural and economical, as an important part of the description of the ecosystem, giving a full account of the history of human populations in the area and their relationship with the natural resources of L'Albufera. In doing this he established a very interesting approach, related to a modern concern for human activities as an integral part of ecosystems, particularly regarding the role of local people in the traditional management of natural resources. But, at the same time, Pardo's descriptive work, based mostly on bibliographical data, lacked a truly ecolo- gical approach. Another valuable contribution of Pardo, again based almost entirely in bibliographical research, was his catalogue of Spanish "lakes" (PARDO 1948), in which he compiled very complete and useful information on lakes, ponds and wetlands, both natural and man made, that even today is still a main source of data, and often the only one when lost ecosystems are concerned, on Spanish lacustrine and palustrine heritage.
But it was a young naturalist from Barcelona named Ramon Margalef, at that time begining his career as a scien- tist, who would start a completely new phase, building the scientific basis for a regional limnology of Iberian conti- nental waters and thus making possible the later develop- ment of limnological studies in Spain. Yet, even though Margalef's work virtually meant a new birth of limnology in Spain, some cbnnections with the previous phase did exist. As an incidental example, one of Margalef's most important early limnological contributions (MARGALEF, 1955) was published precisely by the Instituto Forestal de Investigaciones y Experiencias.
Before briefly analyzing what could be called the modern period of limnology of Spain, intimately associated to Marga- lef, we must mention other scientists, trained in the years before the war, who faced difficult times for scientific acti- vity in Spain during their careers and yet made some valuable, even when modest, contributions. Geographer Juan Dantín (1881-1943) became interested in the "endorheic" character of semiarid regions of the Iberian peninsula, studying from a physiographical point of view many of the saline and fresh-

Figure 1. Scietitijt, nnd hcieiitific cciitei.\ ot ihc eorly period of Spniii\li Iiiiiiiol~gy. Lel'i lop. Celso Ai-Cvalo. Kiglii iop. rlie Laboi-atory of Hidro- biology in the lnstitlito Valencia. Lefr hottom. The laboratory of ecology i i i tlie Secci6n d e Biología de las Aguas Continentales. Right hottom, Luis Pardo.

water wetlands typical of these areas (DANTIN CERECEDA,
1929, 1930). Pedro González Guerrero (1902- 1984) was one the first Spanish specialists in microscopic algae of continen- tal waters and developed a considerable work on these groups
which can be found in his numerous publications (ALVAREZ
COBELAS & GALLARDO. 1985).
RAMON MARGALEF AND THE MODERN PERIOD OF LIMNOLOGY IN SPAIN
Ramon Margalef (1 9 19) becarne interested i i i thc study of
nature when he was very young. The Civil War prevented
the normal development of his scientific career, so it was not until the 1940's when Margalef was introduced to the scientific institutions in Barcelona and began publishing his
research on aquatic biota. His studies, devoted mostly to
marine and continental plankton. soon acquired an ecologi-
cal pcr\pective ~h;it Margalef developcd to i i \ fulle\t exiciic
1-12iii-e 2. l i ; ~ i i i t r i i 1I.ii~;ilc.l. I<~~i i i t lc . i 111 i l ic riio(lc~iii (>i,iicid 01 I i i i i i io- logy in Spaiii.
in the ensuing years, resulting iii his excellcnt work of synt- hesis Los or.,yutii.snios iridic.ridoi.e.s rri lo Linr~zolo,yí(~
(MARGALEF, 1955). This was the naturalist period of Margalef's work. in which he surveyed the aquatic ecosyst-
mes of the Iberian peninsula, building up a rcgiotial limno-
logy similar in its approach to those that were being devc-
loped in Central Europe. In 195 1 he had started to work at the Instituto de Investi-
gaciones Pesqueras, in Barcelona, which he later directed, and thus much of his rescarch was focused on marine
ecology. During this period Margalef also became interes- ted in theoretical ecology. His well known contributions in this field have had a very important international impact and
are far beyond the scope of this paper. Anyhow, Margalef rctained a deep interest in basic limnological research.
In 1967 Margalef became the first Professor of Ecology
and Director of the first Department of Ecology of Spanish
university. which was created in the Universitat de Barce-
lona. In this center a true limnological school was at last formed around Margalef. During the decade of the 1970's an important group of young researchers were trained in the
study of karstic and mountain lakes, coastal lagoons, rivers 2nd rrservoiss. These 5lirnnoIrtgista su.uhseqcienity f~ rmed new
research groups in other Spanish universities. Finally, an specialized journal also appeared, Oecolo,gicz uc~uutic,n, which was published in Barcelona since 1973, devoted fo limnology and oceanography grouped together under the
concept of aquatic ecology. The publication of Margalef's book Limtlologiu in 1983.
which completed his ecological Corpus which started in 1974
with E<,ologíu, nieans the maturity of Margalef as a limnolo- gist and of limnology as a firmly established science in Spain.
The creation of the Asociación Española de Limnología in 198 I also had an important significance in the normali-
zation process of this science in Spain. Up until this time, there had been numerous groups of people, mainly in zoology departments of different faculties, who worked independently from those ot' the Universitat de Barcelona
and in a more or less isolated and self-taught way on taxo- nomy of aquatic organisms and biological indication of
water quality. During the 1970's a certain degree of concern
about pollution in Spanish rivers grew among governmen- tal authorities and funds were provided for research in this
field. As a result some research groups appeared in different regions oí' Spaiii. The Asociación Española de Limnología connected these previously isolated efforts and with its scientific meetings and publications provided means for cotnmunication and exchange of ideas and experiences

among Spanish limnologists. Recently an official scientific center, the Centro de Investigaciones del Agua, in Arganda, Madrid, has been reactivated and hopefully it will contribute to the growth and development of Spanish limnology.
Today we can claim that limnology, as a basic science, is firmly established in Spain. An association exists with more than 300 members, an international scientific journal, Limne- cica, and a newsletter bulletin, Alquihla. Bibliographical checklists and identification keys of aquatic fauna and flora are published by the association and national congresses are held every two years. Limnology is taught in 6 different universities and an international course takes place periodi- cally in the Centro Internacional de Altos Estudios Agronó- micos, at Zaragoza. Finally, the 1992 edition of SIL congress will be held this year in Barcelona. In a way, this event can be regarded as international recognition of the work done during the last eighty years by limnologists in Spain.
CONCLUDING REMARKS
REFERENCES
As any other social or cultural process, limnological science in Spain has followed a historical path parallel to political and economical factors that, since nineteenth century, have condi- tioned strongly its development. In this regard, limnology shares some historical patterns with other sciences in Spain:
a). As a new scientific field limnology is introduced in Spain relatively soon after it was defined by Forel in 1892.
b). Later development of limnology is difficult because of lack of official support, being personal projects respon- sible for most of its limited achievements and resulting in a lack of continuity.
It is difficult to understand why a country where water is such a limited and therefore valuable resource has paid so
little attention to the scientific study of continental aquatic ecosystems. Instead an engineering approach has dominated the understanding and management of Spanish waters, regarded almost exclusively as something to be stored in reservoirs and transported by channels, while ecological aspects have been largely ignored, and to a certain extent still are, by official authorities.
ALVAREZ COBELAS, M. & T. GALLARDO. 1985. In memoriam. Pedro González Guerrero. Anales del Jardín
Botánico de Madrid, 42 (1): 3-7. AKEVALO. C. 1914. El Laboratorio Hidrobiológico del
Instituto de Valencia. Boletín de la Real Sociedad Espariola
de Historia Natural, 14: 338-348. AREVALO, C. 1916. Introducción al estudio de los
cladóceros del plankton de la Albufera de Valencia. Anales del Instituto General y Técnico de Valencia, 1.
AREVALO, C. 1917. Algunos rotíferos planktónicos de la Albufera de Valencia. Anales del Instituto General y Técnico de Valencia, 2.
AREVALO, C. 1921. Sobre el concepto preciso de la palabra "hidrobiología", en su sentido estricto. Boletín de la
Real Sociedad Española de Historia Natural, 2 1 : 171 -177. AREVALO, C. 1923a. Algunas consideraciones sobre la
variación temporal del plankton en aguas de Madrid. Boletín de la Real Sociedad Española de Historia Natural, 23: 94- 103.
AREVALO, C. 1923b. El lago de Carucedo. Memorias de
la Real Sociedad Española de Historia Natural, 11: 305-330. AREVALO, C. 1929. La vida en las aguas dulces. Labor.
Barcelona. AZPEITIA, F. 1908. La diatomología española a comien-
zos del siglo XX. Asociación Española para el Progreso de
las Ciencias. Congreso de Zaragoza IV (2): 5-320. BUEN, O. de. 1887. Materiales para la fauna carcinoló-
gica de España. Anales de la Sociedad Española de Historia Natural, 16: 405-434.
CALDERON, S. 1888. La salina de Fuente-Piedra. Anales de la Sociedad Española de Historia Natural, 17: 72-83.
CALDERON, S. 1889. Excursión á Fuente-Piedra (parte zoológica). Anales de la Sociedad Española de Historia Natural, 18, Actas: 57-59.
CAMARASA, J. M. 1989. Aspectes histbrics dels estu- dis ecolbgics als Pai'sos Catalans. In: Historia Natural dels
PaiSos Catalans (R. FOLCH 1 GUILLEN, ed.) 14. Sistemas naturals: 25-40. Fundació Enciclopedia Catalana. Barcelona.
CISTERNAS, R. 1887. Ensayo descriptivo de los pe-ces de agua dulce que habitan en la provincia de Valencia. Anales de la Sociedad Española de Historia Natural, 6: 69- 138.
DANTIN CERECEDA, J. 1929. Localización de las
ACKNOWLEDGEMENTS zonas endorreicas de España. Memorias de la Real Socie-
dad Española de Historia Natural, 15: 829-836. This article is part of a research project on the history of DANTIN CERECEDA, J . 1940. La aridez y el endo-
Spanish ecology partially supported by the Residencia de rreismo en España. El endorreismo bético. Estudios Geográ-
Estudiantes, Madrid. ficos, 1 : 75-1 17.

GOMIS BLANCO, A. 1988. Las ciencias naturales en España en el siglo XIX (1833-1874): morfología, fisiología y sistemática. Tesis doctoral, Facultad de Ciencias Biológi- cas, Universidad Complutense de Madrid. Madrid.
GRAELLS, M. P. 1846. Catálogo de los Moluscos Terrestres y de Agua Dulce observados en España. Imprenta Martínez. Madrid.
GRAELLS, M. P. 1864. Manual Practico de Piscicultura.
E. Aguado. Madrid. HALBFASS, W. 1913. Der Castañedasee, der grosste
Süsswassersee Spaniens, und seine Umgebung. Petersen Mitteilungen, 59: 306-3 12.
MACHO VELADO, J. 1878. Moluscos de agua dulce de Galicia, con observaciones sobre las especies y Socalida- des. Anales de la Sociedad Española de Historia Natural, 7: 235-248.
MARGALEF, R. 1955. Los organismos indicadores en la
Limnología. Instituto Forestal de Investigaciones y Expe- riencias. Madrid.
MARGALEF, R. 1974. Ecología. Omega. Barcelona. MARGALEF, R. 1983. Limnología. Omega. Barcelona. PARDO, L. 1942. La Albufera de Valencia. Estudio
limnografico, biológico, económico y antropológico. Insti- tuto Forestal de Investigaciones y Experiencias. Madrid.
PARDO, L. 1945. Nota necrológica. D. Celso Arévalo Carretero. Boletín de la Real Sociedad Española de Historia
Natural, 43: 187-198.
PARDO, L. 1948. Catálogo de los lagos de España. Insti- tuto Forestal de Investigaciones y Experiencias. Madrid.
PARDO, L. 1949. Breve noticia acerca de la Sección de Biología de las Aguas Continentales. Real Sociedad Española de Historia Natural. Tomo extraordinario publicado con mofivo del LXXV aniversario de sufundación. 1946: 201 -21 2.
PARDO, L. 1952. Para la historia de la Hidrobiología Española. El Laboratorio de Hidrobiología de Valencia. Boletín de la Real Sociedad Española de Historia Natural, 50: 407-415.
REYES NEVARES, S., dir. 1982. El exilio español en Mé,~ico. Fondo de Cultura Económica, Salvat Editores Mexicana. México, D. F.
REYES PROSPER, E. 1910. Las Car&tus de España, singular-mente las que crecen en sus estepas. Imprenta Artís- tica Española. Madrid.
REYES PROSPER, E. 1915. Las estepas de España y su ~!egetación. Real Casa. Madrid.
TABOADA TUNDIDOR, J . 1913. El lago de San Martín de Castañeda. Boletín de la Real Sociedad Española de Historia Natural, 13: 359-386.
VELAZ DE MEDRANO, L. & J. UGARTE. 1933. Estu- dio monografico del río Manzanares. Instituto Forestal de Investigaciones y Experiencias. Madrid.
WILLKOMM, H. M. 1852. Die Strand- und Steppenge- biete der iberischen Halbinsel und dere vegetution. Frie- drich Fleischer. Leizpig.

ECOLOGICAL ASPECTS OF WETLANDIGROUNDWATER RELATIONSHIPS IN SPAIN
F. González Bernáldez Departamento de Ecología. Universidad Autónoma de Madrid. 28049 Madrid. Spain.
Keywords: Aquifer discharge habitats, Groundwater flow, Biological indicator, Phreatophyte, Wetland conservation
ABSTRACT
Groundwater and sub-surficial water relationships are very important aspects of the ecological processes of Spanish wetlands.
Although only a part of these relationships is known and a relatively small number of wetland types and geographical aseas have
been investigated, results indicate that geohydrology should be an essential part of wetland characterization. Hypogenic and epigenic wetlands are quite different categories of ecosystems and it has been established that groundwater relationships are key factors in determining flooding, water regime and leve1 stability, hydrochemistry, basin origin and morphometry, turbidity,
eutrophy, plant and animal communities and susceptibility to impact. Much closer cooperation between geohydrologists and
wetland ecologists and better diffusion of ecological and hydrogeological knowledge among both groups of specialists are needed to cope with the important tasks of wetland management and conservation as well as to respond to new socioeconomic demands.
INTRODUCTION
Groundwater plays an important role in determining the ecological characteristics of many Spanish aquatic ecosys-
tems although these circumstances have only been conside- red recently. There are several reasons for this lack of atten- tion but the responsibility is shared by various professions.
Hydrogeologists have traditionally paid little attention to surficial phenomena involving wetland-aquifer relations-
hips. When they did show concern the aim of the project was usually to suppress loss of the groundwater reserve,
thus considering water flow to the wetland as an obstacle
and an economic problem to be solved.
Ecologists have also rarely been aware of the upward groundwater flows connected with many wetlands. They tended to believe that when water infiltrates deeply in the
ground it is definitively lost by the surficial ecosystems and were unaware ignored that it may flow back again to the surface with new chemical characteristics.
In Spain, the seminal paper of R. Llamas (1982) on the
neglected role of groundwater in wetland management and conservation has been very important in triggering research by various groups. This work has recently received more attention primarily due to the requirements of environmen- tal impact studies.
Perhaps the catastrophic destruction of the Las Tablas de Daimiel National Park and the Ojos del Guadiana asea due
to excessive groundwater withdrawal was the event that finally increased both government and public awareness of
the importante of groundwater systems in wetland conser- vation. This disastrous outcome had been accurately predic-
ted in various interna1 reports, although little attention was paid to them by the authorities.
In the years preceding the Spanish Civil War, however, the Spanish ecologist Emilio Huguet del Villar y Serratacó, correctly interpreted many hydric soils and wetlands as the
result of the action of upward moving groundwater flows
(GONZALEZ BERNALDEZ, 1987). These interpretations
based on extensive field experience were published at a time when the models establishing the principies of upward groundwater flow in unconfined aquifers (HUBBERT, 1940,
TOTH 1965) had not yet appeared. At the time, it was gene- rally believed that al1 the upward groundwater movements were due to "artesian" phenomena only possible in the
special case of a confined aquifer. Del Villas's explanations therefore had to rely on discredited mechanisms such as the early interpretation of "plutonism", thus partly explaining the scarce repercussion of his theories.
Twenty years ago, there was a similar lack of attention
to the effect of groundwater on wetlands in other coun-
L~mnetica, 8: 11-26 (1992) O Asociación Espafiola de Limnologia, Madrid. Spain

tries, although during the last two decades, research on groundwater-wetland relationships has been considerably developed in most advanced countries. One reason for this growing interest is also the generalization of Envi- ronmental Impact Statements that forced many geohy- drologists to tackle the complicated problems of ground- water influences on wetlands. This is the case of Canada, where the extraordinary progress of the applications of geohydrology to wetland conservation is mainly due to the Government's policy of financing private conserva- tionist groups for selecting experts who are asked to duplicate the developer's impact study. Excellent studies on wetland-groundwater relationships are presently carried out in the USA (WINTER, 1981) and in Holland (GROOTJANS, 1985).
GENERALITIES
The most important variables in wetland-groundwater relationships include the following:
- The nature of the aquifer or saturated material involved. - The substratum's permeability. - The groundwater flow characteristics: e.g., length and
residence time of the water. - The hydrochemistry and lithological characteristics of
the territory. - The relations between the different inputs: groundwa-
ter, meteoric water and exogenous water brought by inflo- wing water courses.
Groundwater inputs may be important enough to consti- tute significative components of the waterbody balance. In contrast, they may be negligible from the point of view of net water balance but be an important factor by preventing surface water infiltration as a result of the substrate satura- tion. Del ViHar (GONZALEZ BERNALDEZ, 1987) mentio- ned upward moving groundwater as the most effective cause of the so called "impeded drainage".
These inputs normally depend on the permeability of the geologic materials in which the waterbody is confined. The most important and concentrated inputs are generally found in karst environments or in heavily fissured ground where groundwater flows through fractures and joints. The bottom of waterbodies on coarse sand such as dune slacks is also highly permeable.
The nature of the waterbody bottom is very important in controlling permeability. Small lakes formed by dissolution in hard, relatively pure limestone are carpeted by silt or
"decalcification clays" flocculated by the influence of Ca ions. These waterbodies behave as "piezometric lakes" promptly transducing piezometric fluctuations of the aqui- fer with which they are connected ("torcas", Cuenca). In contrast, waterbodies located on detritic siliceous Miocene sediments in both Central Subplateaus are often affected by alkaline groundwater provoking clay dispersion and imper- meable bottoms that respond to piezometric fluctuations with a considerable time lag.
The same examples can illustrate the effects of ground- water hydrochemistry. Detritic siliceous Miocene sediments, poor in Ca, permit the observation of the progressive "ageing" process of groundwater when its flows become longer and residence time within the sediments increases. Groundwater becomes increasingly alkaline when moving towards the lowest parts of the aquifer, thus influencing the hydrochemistry, sediment flocculation and the kind of orga- nisms of the wetlands affected by its discharge (GONZA- LEZ BERNALDEZ et al., 1989). In contrast, areas in which Ca is dominant may induce rapid changes in infiltrated water chemistry and the evolution process is very short and barely noticeable (FREEZE & CHERRY, 1979).
The relationship between meteoric and groundwater inputs is also important. Very often, groundwater seepage to wetlands is a stabilization factor, regulating water level and ensuring flooding persistence even in areas characterized by the Mediterranean summer dry period. Water level fluctuations and flooding duration are related to the nature of aquatic biocenoses that can be used as indicators of average water regime and groundwater relationships (GONZALEZ BERNALDEZ et al., 1988). Maximum water level stability is achieved in wetlands receiving regional flows from large aquifers while the buffering effect may be incomplete (GONZA- LEZ BERNALDEZ e t al., 1992a) in other instances (connection with small volumes of saturated ground, low capacity detritic sediments such as aeolian sands or thin outwash deposits such as "rañas") .
The particularities of wetland-groundwater relationships are better considered in the framework of the following
typology.
WETLAND TYPOLOGY AND CHARACTERISTICS FROM THE POINT OF VIEW OF GROUNDWATER RELATIONSHIPS
The following tentative types can be considered:

Maritime wetlands Riverine wetlands and floodplains
Both in coastal salt marches intluenced by tides and in Mediterranean areas tlooded by autumn storms. the effects of upward continental groundwater flows are generally noti- ceable. Ground saturated by continental flows can lengthen
the flooding period, resulting in complex mixture patterns. Continental groundwater and sea water interactions are especially complex in the case of complicated fluviomarine
formations such as deltas (Delta del Ebro, Tarragona; Llobregat, Barcelona; Tinto and Odiel, Huelva, etc.).
Fluviomarine deltas
Natural os less modified deltas are made up of heteroge- neous territories in which either continental os maritime influences predominate. Tidal and brackish water influen-
ces dominate in the lowest, dista1 reaches of the delta while continental groundwater and watercourses are important in the highest parts. Most deltas were important aquifers on the surface of which areas of groundwater discharge, waterbo-
dies and channels alternated with aseas of raised ground and
levées. The upper parts were characterized by barely mine-
ralized water while the lowest reaches were normally heavily saline. Part of the lakes aiid lower channels were affected by tide water and a transition area of intermediate
salinity was located betwcen the maritime and continental zones of influence.
Before recent profound alteration, these effects were
clearly visible in the Doñana area (S.W. Spain). There was
sharp contrast between the less mineralized "lucios altos"
(waterbodies of the upper reaches), upper "caños" (chan- nels), dune slack ponds and the heavily mineralized "lucios
bajos" (waterbodies of the lower reaches). Hydrochemical
complexity was increased by the interaction between the water carried by exogenous water courses and channels, the inflowing groundwater and the brackish water from tides. Gradients of salinity and differences in water regime were very important in conditioning animal behaviour rhythms,
including migrations. It is known that a given level of sali-
nity and flooding in intermediate "lucios" was necessary for
geese to feed on bulrush rhizomes (Scirp~fs nzul-irinzus subsp. compuctus) and that seasonal connections between the diffe-
rent channels and waterbodies controlled fish migrations. Very often, complex sediment stratification in deltas, inclu- ding alternated permeable and impermeable layers, add more variable influences to this complexity by diversifying the origin and characteristics of the inflowing groundwater.
These wetlands have also been profoundly altered in most developed countries, especially by river regulation and in some cases by excesive groundwater extraction.
Large water courses flowing on very flat plains help to
flood extensive aseas but the main reason for braiding and
permanent flooding ("tablas") is the presence of diffuse groundwater discharges that ensure substratum impermea-
bility. The role of groundwater in these ecosystems was
clearly shown in the destruction of the Tablas de Daimiel
National Park (Ciudad Real) affected by excesive ground- water withdrawal. The resulting damage is attributed to the
loss of the aquifer influences that controlied the regime and
chemistry of the water and maintained an impervious, satu- rated substrate preventing the infiltration of surficial water.
The disturbance of the Tablas de Daimiel natural area star- ted in the 1960's with the irrigation of a rapidly increasing
area. Today most of the water used for the irrigation of the 150.000 ha. farming area comes from the 6 000 km2 aquifer that was formerly responsible for tlooding the Tablas. About
15.000 wells withdraw 600 cubic hectometer/year and have
lowered the upper limit of the saturated zone by as much as
30 metres. As a result, the flood-plain jointly made up of the Guadiana-Cigüela junction and the important groundwater systems has been profoundly altered and groundwater inflow
con~pletely suppressed. As the groundwater no longer satu- rates the Tablas bottom, the surface waters infiltrate deeply and do not form the extensive flooded aseas characteristic of this wetland. The former discharge wetland has now been
converted into a recharge area. Therefore, the costly attempts
to solve the problem by bringing water through the Tagus-
Segura channel also result in infiltration and in aquifer recharge. Ironically, this infiltrated water is finally pumped
into the wells and used for irrigation. In order to reduce water losses, the former exorheic
system that maintained an appreciable water flow has now been converted into a closed system by damming the outlet.
As a result, and also owing to the lack of the less minerali- zed groundwater inputs, salination has increased. Water
quality changes, turbidity increases and the depression of the saturated zone have al1 brought about important changes in vegetation. The Myr-ioplzillaceue, Potamogeton spp..
Charophytes and other submerged plants carpeting the ground are greatly diminished and the water-stress sensitive Cludiunz nrur-iscus is been steadily replaced by the more resistant Phl-agmites austrulis. These changes are affecting
animal communities, including the most important objecti-

ves of the conservation policy such as the red crested pochard Netra rlrfincl (LLAMAS, 1988).
Even much simpler ecosystems than the conventianal
floodplains such as the "vegas" of the Cigüela River have been recently destroyed in Spain. These "vega" areas located upstrearn of the Tablas de Daimiel were periodically flooded by the Cigüela with the important contribution of diffuse groundwater discharges that maintained a saturated ground. At present, only some vestiges remain such as the El Masegar and El Taray private estates (Toledo) where the flooded condition is ensured by artificial groundwater pumping. The absence of natural flooding by river water is due to the exca- vation and drainage of the Cigüela stream channel, even in its upper reaches, thus resulting in the loss of the natural values of the "vegas". This work carried out in the 1980's when al1 governments of the world were already aware of the impor- tance of wetland conservation is one of the most gratuitous
and harmful outrages against natural values in modern Spain.
Peririverine wetlands
Floodplains in their simplest form, floodable "vegas" are part of a continuum of riverine wetland types in which the role of groundwater is always important. Oxbow lakes ("madres viejas", "galachos") and backwaters or flooded
depressions between the natural levee and the terrace system ("derrames", "cilancos") are very much influenced by natu- ral groundwater systems. Oxbow lakes are flooded by groundwater from the same aquifer flowing into the usually
"gaining" stream. "Backwaters" and other peririverine lakes
originate during floods but they persist for longed periods due to the Iack of vertical drainage induced by upward verti- cal components of groundwater flow (examples: "madres
viejas" or oxbows of the Jarama river, near Madrid; "gala- chos" of the Ebro river, near Zaragoza).
Ecosystems related to river floods and influenced by groundwater were very important prior to river regulation (in Spain, for instance, they were the favoured locality of
the now extinct sedentary crane populations). Today the numerous grave1 and sand pits resulting from mining opera- tions on river terraces and flooded by groundwater from the
alluvial aquifer can be restored in order to i~nitate these lost
habitats (DEPARTAMENTO DE ECOLOGIA, 199 1 ).
Hidden seepages and seeps
In the hidden seepages. water from the saturated zone is evaporates directly to the atmosphere through soil micros-
tructures (capillaries, colloids, etc.) or plant conducting systems. But no liquid water accumulates on the surface as in other wetlands. The upper limit of the saturated zone
should be relatively close to the soil surface to permit this e~a~o t ransp i r a t ion process. The exact distance depends on soil texture and structure and on the size of the capillary
zone, extending to 3 meters or more under special circums- tances. Some plant species, however, are able to extract and
transpire groundwater from considerable depths. Mediterra- nean Tarnari.~ species have been observed pumping ground-
water at about 10 meters and depths of 20 to 30 meters have been recorded for some American Prosopis. Groundwater
influence may profoundly affect soil chemistry and profile morphology inducing hydric, saline or alkaline conditions (GONZALEZ BERNALDEZ & REY BENAYAS, 1992a) even if no liquid water accumulates on its surface.
Hidden seepages or seeps are important forms of aquifer
discharge in semiarid areas. giving rise to characteristic
landscape sectors and biocenoses. Seepages in the granite areas of the Sistema Central give, for instance, rise to the majority of wet meadows and peaty meadows ("chortales",
"tallos") on fractured and altered granite. These wet surfaces are essential for grazing by livestock during the summer dry period. Acid meadows (Nurdus stricta. Trifoliurn lueviga- f ~ t n ~ , Juncus squarrosus, etc.) correspond to short tlows and
short residence times of the groundwater. In contrast, meadows linked to longer flows, coming up from conside- rable depths and having long residence times are characteri- zed by slightly alkaline conditions and high pH (Trvolium resupinaturn, T . n?ii,helianurn, Descharnpsia cespirosu subsp. refi-acta, etc.). Alkaline meadows are always located at
lower altitudinal levels (lower pediments, granite outcrops in the plains) while acid meadows can be found everywhere (GONZALEZ BERNALDEZ, 1989b)
The interstitial meadows scattered through the vast cereal fields on both Castillian subplateaus ("prados entrepana- dos", "longares", "rodiles", "mangadas") are similar. They are due to evaporative discharges from the main aquifer at the botton of the thalwegs. They are often associated with
creeks or small waterbodies in the center of the meadow. In siliceous deposits they tend to be saline or alkaline when located at the lowest levels of the aquifer (thus correspon-
ding to the longest, regional flows).
Continental "saladares", areas where mineralized groundwater is evaporated and evaporite minerals accumu- lated on the surface or in the soil profile, normally corres- pond to "solonchak" soils (La Mancha, Monegros). Very often they are associated with more or less ephemeral saline

lakes (continental discharge "sebkhas" or "playas",
GONZALEZ BERNALDEZ, 1992). It has been observed
that the lowering of the water table leads to the transfor- mation of "playas" or "sebkhas" into non-floodable "sala- dar". Its salination depends on the distance of the soil
surface from the upper level of the saturated zone. The
vegetation cover reflects the rate of salinization, ground- . water fluctuation and hydrochemistry. There are, for instance, indicator plant differences between "soloniets"
areas (where sodium is the dominant cation) characterized
by a lower plant diversity and the absence of some species (for example, Lirnoni~lrn spp.) and the more biodiverse,
calcium-rich "solonchak". In areas with groundwater at a considerable depth, howe-
ver, different temporarily flooded "saladar"-similar or "sebkhaM-similar structures can be formed at the outlet or the confluence of wadis, "ramblas" or other runoff features.
The floodable territory thus behaves like a recharge area.
Under these circumstances, however, true salinization and
halophytes can only occur in the truly arid zone. Halophytes are often absent, as in the Argentinian "barriales".
Wetlands in detritic siliceous facies, sand and siliceous outwash deposits
The detritic facies of the continental Miocene in central
Spain offer many examples of these wetlands, although a large percentage of the variants connected with non-mine-
ralized groundwater (upper reaches of the aquifer) have been altered or destroyed by groundwater withdrawal from
the aquifer for irrigation purposes (GONZALEZ BERNAL-
DEZ & REY BENAYAS, 1992). The wetlands correspon- ding to the mineralized parts of the aquifer (including the interesting alkaline lakes of Coca, El Raso del Portillo near
Boecillo, Valladolid, or the Villafáfila lakes) have also been
subjected to serious disturbance by drainage attempts. As already indicated, the main difference from the
wetlands associated with limestone or mar1 (for instance
the "chemical" facies of the continental Miocene) is the process of groundwater evolution in silicate aquifers
leading to alkalinization, high pH (9 to 11 in come lakes), the presence of carbonate and the deposits of characteristic minerals such as "trona". But in addition to the wetlands
connected with the regional aquifer, a wide range of other geohydrological circumstances can be observed, including recharge waterbodies exclusively dependent on meteoric water (GONZALEZ BERNALDEZ et al. , 1988; BES- TEIRO, 1992).
15
Most wetlands on silicate detritic materials occupy ancient depressions of fluvial origin, but it has been obser-
ved that upward moving groundwater and wind can be agents of lake-basin formation. Mud volcanos or "soap- holes" (GONZALEZ BERNALDEZ et al . ) are able to
excavate depressions that can be extended by wind action. On the other hand, salination of the soil surface by ground-
water may provoke the abscnce of plant cover, thus expo- sing the soil to wind erosion. Clusters of small saline
waterbodies accompanied by sediment accumulation remo- ved by wind ("clay dunes" or "lunettes") on the leeward
side are characteristic of saline discharge areas (Raso de El Portillo, Valladolid; Lagunas de Villafáfila, Zamora).
They probably represent initial stages of lake-basins (REY BENAYAS et al. , 1989; GONZALEZ BERNALDEZ er al., 1987).
Wetlands associated with saturated zones in outwash deposits or "rañas" corresponding to small volume aquifers
are always characterized by short flows and non-minerali-
zed grounwater and have acidophilous vegetation belts (Nardus stricta, .lunc.us sqliari-osus, etc.). Outstanding examples are the Laguna de El Cristo, Laguna Cervera
(Salamanca), and the Laguna Delgada (Toledo), with less acidophilous margins.
Water bodies in granitic areas are associated with the previously mentioned hidden seepages and correspond to the same hydrochemical relationships. The most determinan1
factor is their connection with differently evolved ground-
water flows. They can vary from low mineralized water to hypersaline conditions such as the small ponds and springs
found near large faults or contacts with other geological materials (for instance Miocene arkoses as in El Salobral,Toledo). The circumstances of the heavy salination of some groundwater discharges near the edges of the
granite area are not clear. They probably correspond to the
upward trajectories of very old, deep water (HERRERA,
1987). In contrast, groundwater evolution in the center of the granitic area only leads to slight alkalinization and mine-
ralization. Wetlands located on aeolian sands are remarkable for
their high substratum permeability and their "piezometric" character. The small lakes on dune slacks in the Doñana area are very good examples. Water is poor in dissolved mine-
rals, leading to acid and dystrophic conditions indicated by characteristic communities. However, sodium enrichment can occur in some instantes (Laguna de Santa Olalla), probably due to groundwater flow interception by deposits containing soluble materials.

Wetlands in limestone areas
A common characteristic of wetlands on limestone is the
predominante of dissolution processes in the formation of lake basins. Aside from this factor, the ecological circums- tances of wetlands on limestone are highly diverse, depen- ding mainly on altitude and climate. The following cases
summarize the majos groundwater relationship patterns. Karst: Karstification leads to huge concentrated flows of
groundwater which are often responsible for permanent floo- ding even when outlets os emissaries are present. An exam-
ple of karst wetlands are the Ojos de Villaverde (Albacete). Water level stability is indicated by the presence of peaty soils with Cladiuwi mar.isc.us and a mosaic of other typical vegetation of more fluctuating conditions in the aseas less affected by the main discharge (Phi.agmites ausrrnlis and
Schoerzus nigric5an.v). The continuoiis water ílow ensured by the outlet and the abundance of Ca ions leads to high water transparency, allowing the growth of an extensive carpet of
Characeue. The same circumstances were typical of many wetlands in La Mancha characterized by flowing calcium-
bicarbonate water (Lagunas del Arquillo, Sanguijuela, etc.). Subcircular waterbodies are frequent in relatively pure
and hard limestone os in marls with a high calcium carbo- nate content from the Secondary period (Albacete, Cuenca, Aragón). Very often, grouridwater is an important agent in the origin of these cavities. Different modalities of suffosion
mechanisms may be combined with other factors (for
instance salinity and wind in the case of the steppe aseas
such as Los Monegros, Zaragoza). Groundwater influences in these small lakes can be quite
diverse. A typical case is the action of calcium-rich water in relatively pure limestone aseas contributing to the formation of a highly permeable, flocculated "decalcification clay" bottom and high transparency (example Las Melchoras complex in Albacete). The permeable bottom allows the
water table to fluctuate and even to expose the dry lake
bottom without salt os evaporite deposits. In mar1 containing evaporites, groundwater flows intercept
soluble materials, creatirig salinity conditions even for very
short groundwater flows and residence times. This is the case of many waterbodies in La Mancha and Aragon. Most of them are temporary, presenting the characteristics of a discharge "sebkha" with subaqeuous or interstitial evaporite formation during a part of the year (Lagunas de la Playa, Salineta, Peña Hueca, Tirez) but others are permanent due to their link with steady regional groundwater flows. This is the case of the hypersaline Laguna de Chiprana (Zaragoza).
Drairlage restriction by upward moving groundwater in fine textured deposits and other phenomena (such as repla- cement of monovalent cations by ageing water) often lead
to the formation of montmorillonite-rich vertisols (bujeos)
with typical associated impondments (Examples: The Lante- juela-Ballestera complex, Sevilla os the temporary "lagunas" in La Sagra asea, Toledo).
Wetlands in recharge areas and wetlands with no influence of groundwater
Several criteria can be used to differentiate waterbodies in discharge aseas receiving the influence of groundwater
flows from recharge waterbodies exclusively dependent on meteoric water (GONZALEZ BERNALDEZ et a l . , 1988;
GONZALEZ BERNALDEZ et al., 1 9 8 9 ~ ) . In the foremen-
tioned siliceous detritic aseas, discharge waterbodies tend to be less epheineral, to exhibit at least a belt of true phrea- tophytes, to have more mineralized water and salt efflores-
cences on their margins and to be associated with typical
discharge phenomena: mud volcaiios, "lunettes" and saline os alkaline soils ("saladares", "salgüeros"), etc.
A water balance o f t h e waterbody should, however, be carried out for more strict discrimination (BESTEIRO, 1992). The water regime of the wetland can be deduced
from piezometric measurements, precipitation and evapora- tion records, water levels and out- os in-flowing water
discharges. Although some of these measurements are diffi-
cult and not always very reliable, an approximate water balance is often achieved that can reveal the influence of
groundwater inputs. The water regime of a given wetland may be rather
complex owing to the participation of surface runoff, inflo- wing and outflowing watercourses, net inputs due to inflo- wing groundwater and losses by interrial seepage. Al1 these factors may act simultaneously, the lake can lose water by underground seepage in certain areas of its bottom and at the
same time receive net groundwater inputs through other aseas. The gaining os losing state can alternate in time depending on water level in the lake os on particularities of the saturated zone geometry (WINTER, 198 1).
In the semiarid Mediterranean region, wetlaiids receiving only meteoric water are always ephemeral. The extreme case of shallow rain pools with characteristic biocenoses adapted to short flooding periods are very important in dry regions.
The lack of connection with groundwater flows and the meteoric origin of the water may be suspected in the small
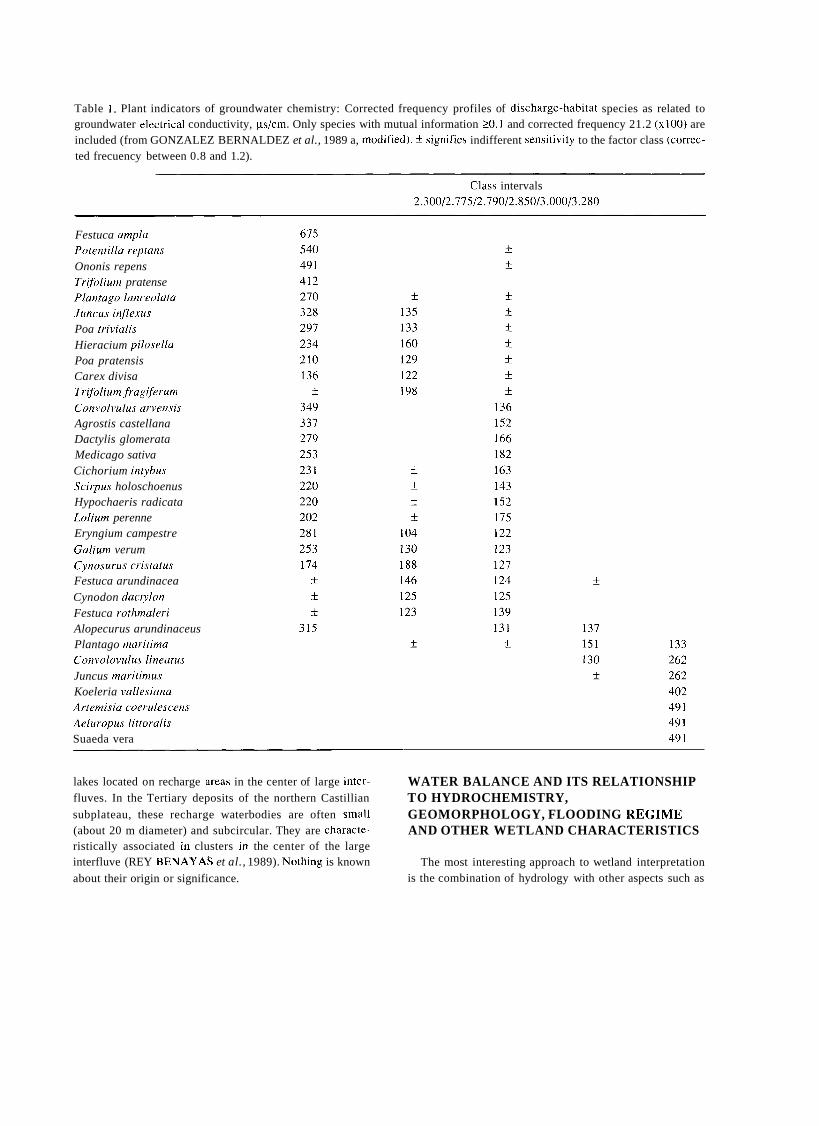
Table 1 . Plant indicators of groundwater chemistry: Corrected frequency profiles of discharge-habitat species as related to groundwater electrical conductivity, ps/cm. Only species with mutual information 20. 1 and corrected frequency 21.2 (x100) are included (from GONZALEZ BERNALDEZ et al., 1989 a, rnodified). f signifies indifferent sensitivity to the factor class (correc- ted frecuency between 0.8 and 1.2).
Class intervals 2.300/2.775/2.790/2.850/3.000/3.280
Festuca ampla Potentillu reptans Ononis repens Trifolium pratense Plantagu lanceolata Jlincus inflexus Poa rrivialis Hieracium pilosella Poa pratensis Carex divisa Trifolium frugiferum Convolvulus arvensis Agrostis castellana Dactylis glomerata Medicago sativa Cichorium intyhus Scirpus holoschoenus Hypochaeris radicata Lolium perenne Eryngium campestre Galium verum Cynosurus cristatus Festuca arundinacea Cynodon dactylon Festuca rothmaleri Alopecurus arundinaceus Plantago maritima Convolovulus lineatus Juncus maritimus Koeleria vallesiana Artemisia coerulescens Aeluropus littoralis Suaeda vera
lakes located on recharge areas in the center of large inter- fluves. In the Tertiary deposits of the northern Castillian
subplateau, these recharge waterbodies are often small (about 20 m diameter) and subcircular. They are characte- ristically associated in clusters in the center of the large interfluve (REY BENAYAS et al., 1989). Nothidg is known
about their origin or significance.
WATER BALANCE AND ITS RELATIONSHIP TO HYDROCHEMISTRY, GEOMORPHOLOGY, FLOODING REGIME AND OTHER WETLAND CHARACTERISTICS
The most interesting approach to wetland interpretation is the combination of hydrology with other aspects such as
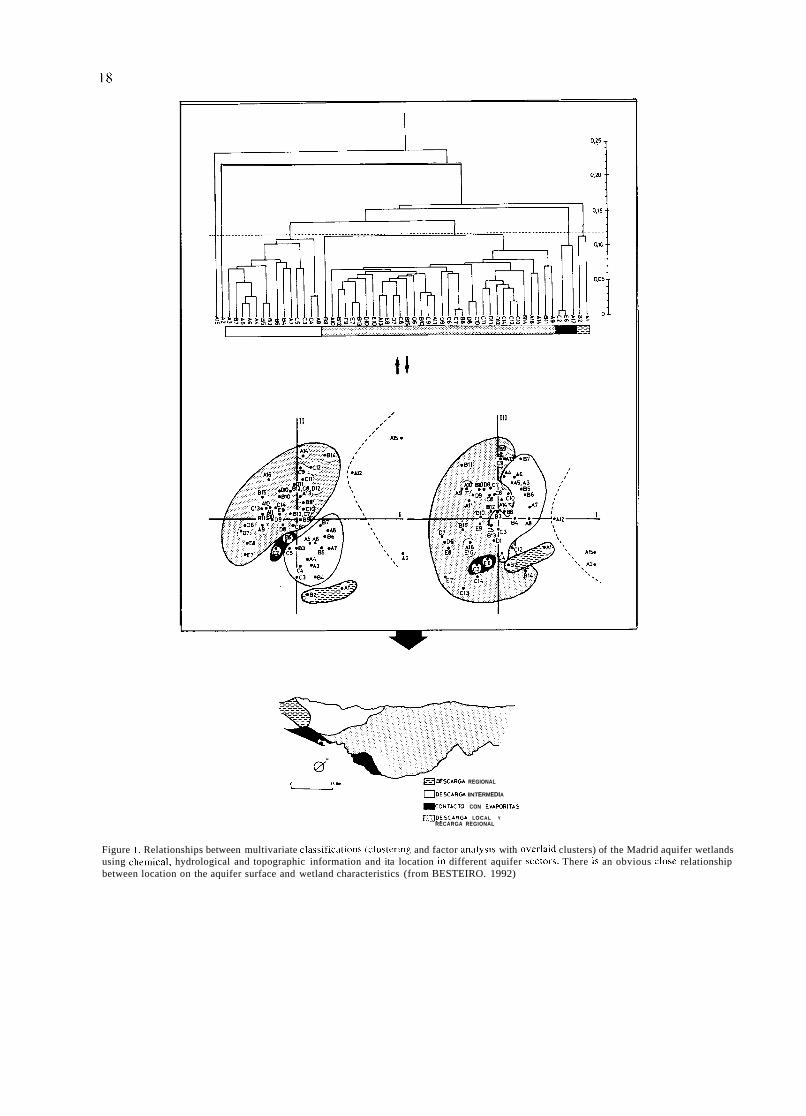
ESCARGA REGIONAL
ODESCAR~ INTERMEDIA
I I ~ o N T A C T O CON EVAFURITAS
DESCARGA LOCAL Y RECARGA REGIONAL
Figure l . Relationships between multivariate classifications (clustering and factor analysis with overlaid clusters) of the Madrid aquifer wetlands using chemical, hydrological and topographic information and ita location in different aquifer sectors. There is an obvious close relationship between location on the aquifer surface and wetland characteristics (from BESTEIRO. 1992)

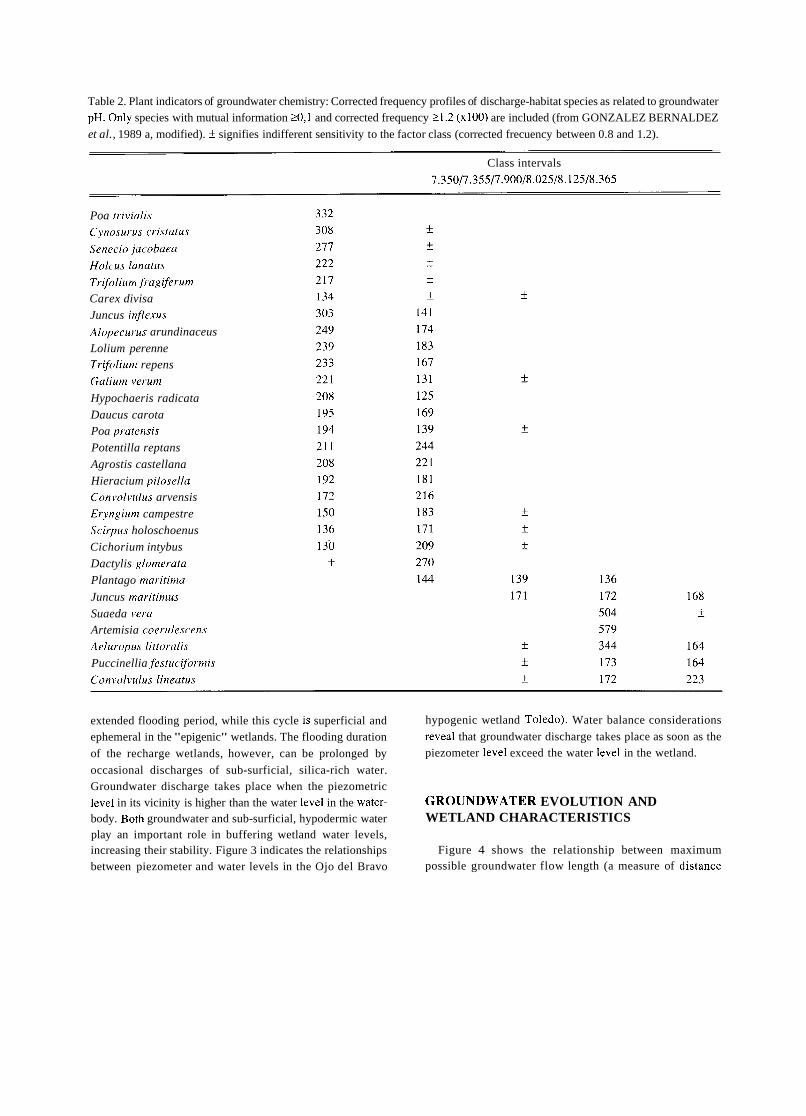
Table 2. Plant indicators of groundwater chemistry: Corrected frequency profiles of discharge-habitat species as related to groundwater pH. Only species with mutual information 20,l and corrected frequency 21.2 (x100) are included (from GONZALEZ BERNALDEZ
et al., 1989 a, modified). f signifies indifferent sensitivity to the factor class (corrected frecuency between 0.8 and 1.2).
Class intervals
7.350/7.355/7.900/8.025/8.125/8.365
Poa trivialis Cynosurus cristatus
Senecio jacohaea
Holcus lanatus Trijolium fragiferurn
Carex divisa
Juncus inflexus
Alopecurus arundinaceus
Lolium perenne Trifolium repens
Galium verum
Hypochaeris radicata
Daucus carota Poa pratensis
Potentilla reptans
Agrostis castellana
Hieracium pilosella Convolvulus arvensis
Eryngium campestre
Scirpus holoschoenus
Cichorium intybus
Dactylis glomerata
Plantago mar-itima
Juncus maritimus
Suaeda vera Artemisia coerulesceris
Aeluropus littoralis
Puccinellia ,festuciformis
Convoli~ulus lineatus
extended flooding period, while this cycle is superficial and hypogenic wetland Toledo). Water balance considerations
ephemeral in the "epigenic" wetlands. The flooding duration reveal that groundwater discharge takes place as soon as the
of the recharge wetlands, however, can be prolonged by piezometer leve1 exceed the water leve1 in the wetland.
occasional discharges of sub-surficial, silica-rich water. Groundwater discharge takes place when the piezometric leve1 in its vicinity is higher than the water leve1 in the water- GROUNDWATER EVOLUTION AND body. Both groundwater and sub-surficial, hypodermic water WETLAND CHARACTERISTICS play an important role in buffering wetland water levels, increasing their stability. Figure 3 indicates the relationships Figure 4 shows the relationship between maximum
between piezometer and water levels in the Ojo del Bravo possible groundwater flow length (a measure of distance
1 , , . , , , , , , , , , , . , , , , , ,L-Z.& E F M M A M y M y J J I J I A S O N D F M M y J JI
N . Y U f S I I E O I 2 1 L I 6 7 1 9 10 11 12 11 I I 95 * 17 18 (9 ?O
@ 010 D<L 91110 l i H AGUA
001T 010 OEL 811A10 )++ PISIOHFTRO 0 s
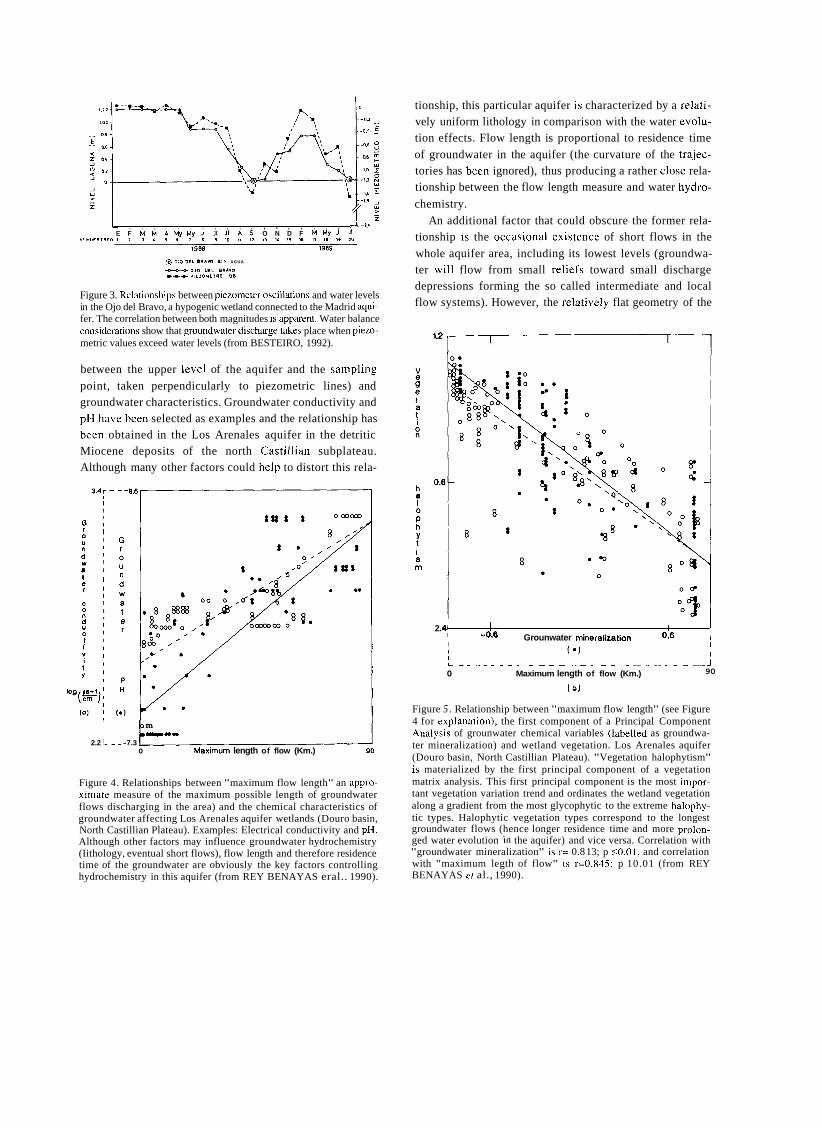
Figure 3. Relationships between piezometer oscillations and water levels in the Ojo del Bravo, a hypogenic wetland connected to the Madrid aqui- fer. The correlation between both magnitudes is apparent. Water balance considerations show that groundwater discharge takes place when piezo- metric values exceed water levels (from BESTEIRO, 1992).
between the upper leve1 of the aquifer and the sampling point, taken perpendicularly to piezometric lines) and groundwater characteristics. Groundwater conductivity and pH have been selected as examples and the relationship has been obtained in the Los Arenales aquifer in the detritic Miocene deposits of the north Castillian subplateau. Although many other factors could help to distort this rela-
1 m
I . . a 2.2 !. - - -7.3
o Maximum length of flow (Km.) 90
Figure 4. Relationships between "maximum flow length" an appro- ximate measure of the maximum possible length of groundwater flows discharging in the area) and the chemical characteristics of groundwater affecting Los Arenales aquifer wetlands (Douro basin, North Castillian Plateau). Examples: Electrical conductivity and pH. Although other factors may influence groundwater hydrochemistry (Iithology, eventual short flows), flow length and therefore residence time of the groundwater are obviously the key factors controlling hydrochemistry in this aquifer (from REY BENAYAS eral.. 1990).
tionship, this particular aquifer is characterized by a relati- vely uniform lithology in comparison with the water evolu- tion effects. Flow length is proportional to residence time of groundwater in the aquifer (the curvature of the trajec- tories has been ignored), thus producing a rather close rela- tionship between the flow length measure and water hydro-
chemistry. An additional factor that could obscure the former rela-
tionship is the occasional existence of short flows in the whole aquifer area, including its lowest levels (groundwa- ter will flow from small reliefs toward small discharge depressions forming the so called intermediate and local flow systems). However, the relatively flat geometry of the
2. I I Grounwater mineralization I
L - - - - - - - - - - - -- - - --.. - - - - - - - J o Maximum length of flow (Km.) 90
( 0 )
Figure 5 . Relationship between "maximum flow length" (see Figure 4 for explanation), the first component of a Principal Component Analysis of grounwater chemical variables (labelled as groundwa- ter mineralization) and wetland vegetation. Los Arenales aquifer (Douro basin, North Castillian Plateau). "Vegetation halophytism" is materialized by the first principal component of a vegetation matrix analysis. This first principal component is the most impor- tant vegetation variation trend and ordinates the wetland vegetation along a gradient from the most glycophytic to the extreme halophy- tic types. Halophytic vegetation types correspond to the longest groundwater flows (hence longer residence time and more prolon- ged water evolution in the aquifer) and vice versa. Correlation with "groundwater mineralization" is r= 0.8 13; p S0.01, and correlation with "maximum legth of flow" is r=0.845; p 10.01 (from REY BENAYAS er al., 1990).
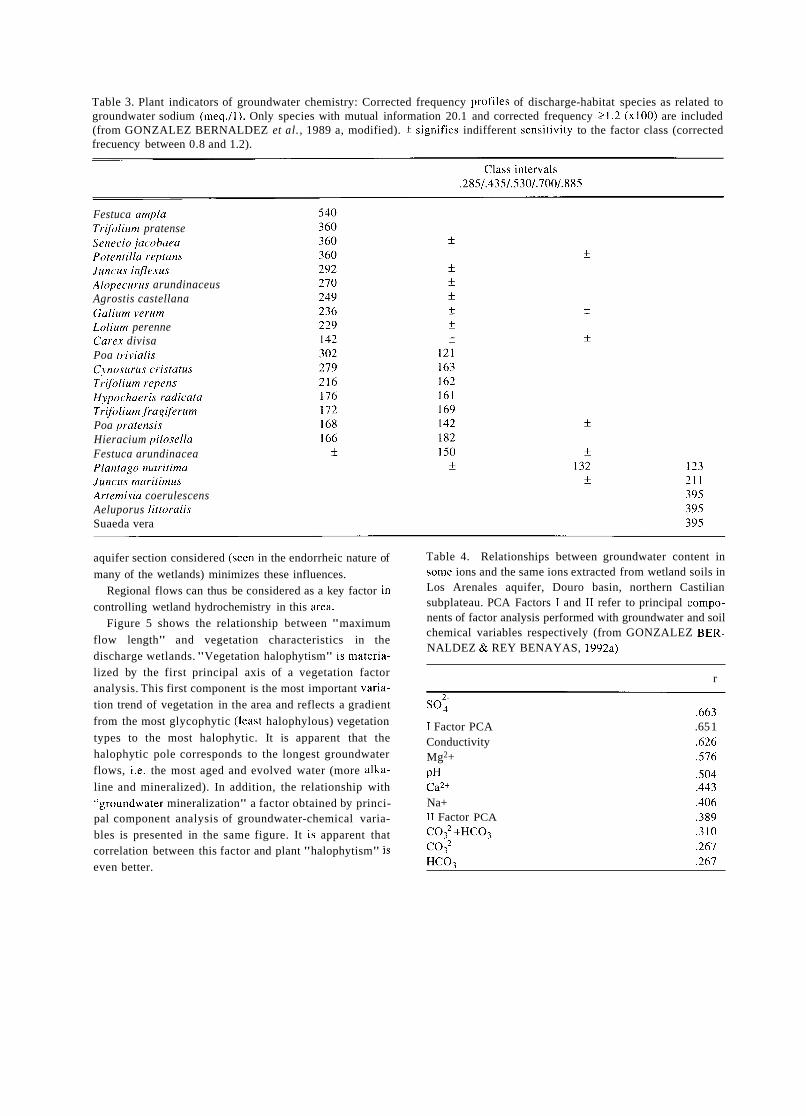
Table 3. Plant indicators of groundwater chemistry: Corrected frequency profiles of discharge-habitat species as related to groundwater sodium (meq.11). Only species with mutual information 20.1 and corrected frequency 21.2 (x100) are included (from GONZALEZ BERNALDEZ et al., 1989 a, modified). f signifies indifferent sensitivity to the factor class (corrected frecuency between 0.8 and 1.2).
Class intervals .285/.435/.530/.700/.885
-
Festuca arnpla TI-ifolium pratense Senecin jacobaea Potentilla reptans Junc~ls inflexus Alopecurus arundinaceus Agrostis castellana Galiurn i'erurn Lolium perenne Curex divisa Poa trivialis Cynosurus cristatus Trijioliurn repens Hypochaeris radicutu Trifoliurn fragiferum Poa pratensis Hieracium pilosella Festuca arundinacea Plantago rnaritirna Juncus rnaritirnus Artemisia coerulescens Aeluporus littoralis Suaeda vera
aquifer section considered (seen in the endorrheic nature of
many of the wetlands) minimizes these influences. Regional flows can thus be considered as a key factor in
controlling wetland hydrochemistry in this area. Figure 5 shows the relationship between "maximum
flow length" and vegetation characteristics in the discharge wetlands. "Vegetation halophytism" is materia- lized by the first principal axis of a vegetation factor analysis. This first component is the most important varia- tion trend of vegetation in the area and reflects a gradient
from the most glycophytic (least halophylous) vegetation
types to the most halophytic. It is apparent that the halophytic pole corresponds to the longest groundwater flows, ¡.e. the most aged and evolved water (more alka-
line and mineralized). In addition, the relationship with "groundwater mineralization" a factor obtained by princi- pal component analysis of groundwater-chemical varia- bles is presented in the same figure. It is apparent that correlation between this factor and plant "halophytism" is even better.
Table 4. Relationships between groundwater content in some ions and the same ions extracted from wetland soils in Los Arenales aquifer, Douro basin, northern Castilian subplateau. PCA Factors 1 and 11 refer to principal compo- nents of factor analysis performed with groundwater and soil chemical variables respectively (from GONZALEZ BER- NALDEZ & REY BENAYAS, 1992a)
r
so: .663
1 Factor PCA .65 1 Conductivity .626 Mg2+ .576
PH S 0 4 Ca2+ .443 Na+ ,406 11 Factor PCA .389 CO?+HCO~ ,310 CO," ,267 HCO; ,267
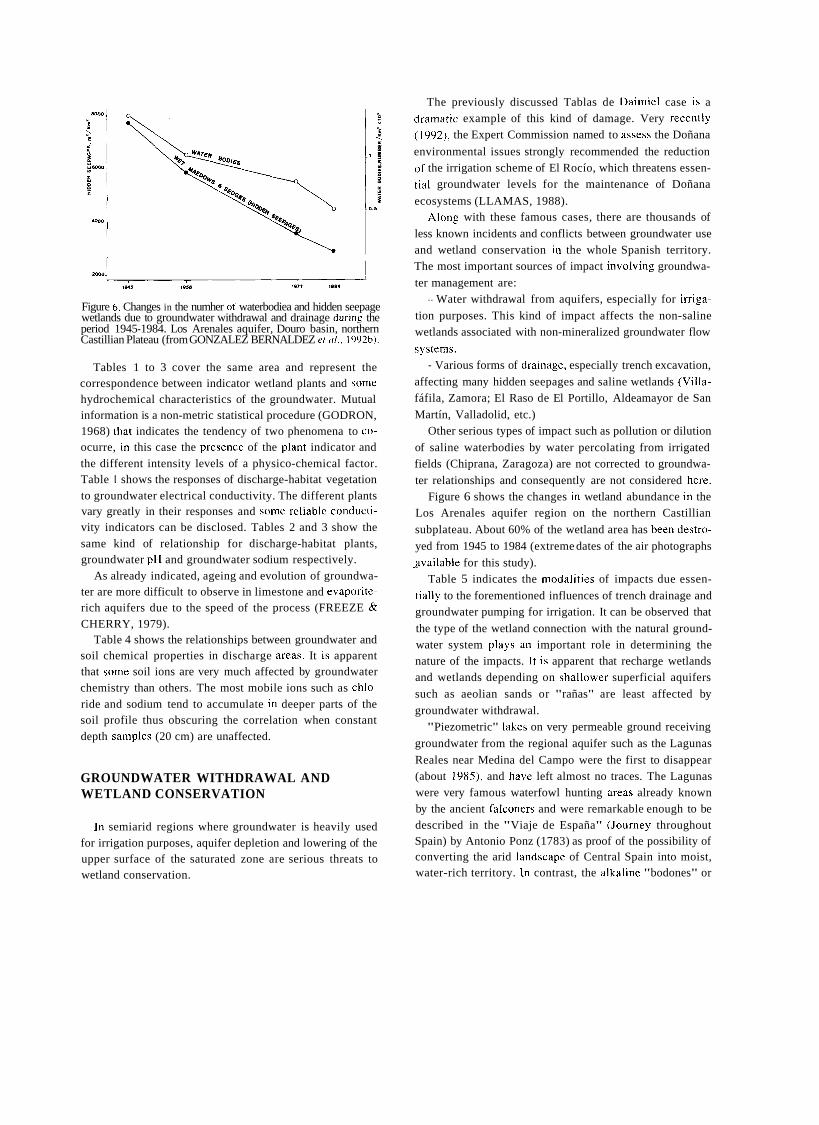
Figure 6. Changes in the numher oí' waterbodiea and hidden seepage wetlands due to groundwater withdrawal and drainage during the period 1945-1984. Los Arenales aquifer, Douro basin, northern Castillian Plateau (from GONZALEZ BERNALDEZ et al.. 1992b).
Tables 1 to 3 cover the same area and represent the correspondence between indicator wetland plants and some hydrochemical characteristics of the groundwater. Mutual information is a non-metric statistical procedure (GODRON, 1968) ihat indicates the tendency of two phenomena to co- ocurre, in this case the presence of the plant indicator and the different intensity levels of a physico-chemical factor. Table 1 shows the responses of discharge-habitat vegetation to groundwater electrical conductivity. The different plants vary greatly in their responses and some reliable conducti- vity indicators can be disclosed. Tables 2 and 3 show the same kind of relationship for discharge-habitat plants, groundwater pH and groundwater sodium respectively.
As already indicated, ageing and evolution of groundwa- ter are more difficult to observe in limestone and evaporite- rich aquifers due to the speed of the process (FREEZE & CHERRY, 1979).
Table 4 shows the relationships between groundwater and soil chemical properties in discharge areas. It is apparent that some soil ions are very much affected by groundwater chemistry than others. The most mobile ions such as chlo- ride and sodium tend to accumulate in deeper parts of the soil profile thus obscuring the correlation when constant depth samples (20 cm) are unaffected.
GROUNDWATER WITHDRAWAL AND WETLAND CONSERVATION
In semiarid regions where groundwater is heavily used for irrigation purposes, aquifer depletion and lowering of the upper surface of the saturated zone are serious threats to wetland conservation.
The previously discussed Tablas de Daimiel case is a dramatic example of this kind of damage. Very recently ( 1 992), the Expert Commission named to assess the Doñana
environmental issues strongly recommended the reduction of the irrigation scheme of El Rocío, which threatens essen- tia1 groundwater levels for the maintenance of Doñana
ecosystems (LLAMAS, 1988). Along with these famous cases, there are thousands of
less known incidents and conflicts between groundwater use and wetland conservation in the whole Spanish territory. The most important sources of impact involving groundwa-
ter management are: - Water withdrawal from aquifers, especially for irriga-
tion purposes. This kind of impact affects the non-saline wetlands associated with non-mineralized groundwater flow
systeins. - Various forms of drainage, especially trench excavation,
affecting many hidden seepages and saline wetlands (Villa- fáfila, Zamora; El Raso de El Portillo, Aldeamayor de San
Martín, Valladolid, etc.) Other serious types of impact such as pollution or dilution
of saline waterbodies by water percolating from irrigated fields (Chiprana, Zaragoza) are not corrected to groundwa-
ter relationships and consequently are not considered here. Figure 6 shows the changes in wetland abundance in the
Los Arenales aquifer region on the northern Castillian
subplateau. About 60% of the wetland area has been destro-
yed from 1945 to 1984 (extreme dates of the air photographs
available for this study). Table 5 indicates the modalities of impacts due essen-
tially to the forementioned influences of trench drainage and groundwater pumping for irrigation. It can be observed that
the type of the wetland connection with the natural ground- water system plays an important role in determining the nature of the impacts. It is apparent that recharge wetlands and wetlands depending on shallower superficial aquifers
such as aeolian sands or "rañas" are least affected by
groundwater withdrawal. "Piezometric" lakes on very permeable ground receiving
groundwater from the regional aquifer such as the Lagunas
Reales near Medina del Campo were the first to disappear (about 1985), and have left almost no traces. The Lagunas were very famous waterfowl hunting areas already known by the ancient falconers and were remarkable enough to be described in the "Viaje de España" (Journey throughout Spain) by Antonio Ponz (1783) as proof of the possibility of converting the arid landscape of Central Spain into moist, water-rich territory. In contrast, the alkaline "bodones" or

Table 5. Correspondence between wetland characteristics and types of impacts due to groundwater withdrawal and drainage. Los Arenales aquifer, Douro basin, northern Castillian subplateau (from GONZALEZ BERNALDEZ et al., 1992b).
"Hidden seepages" (wet meadows)
Waterbodies Nature of impacts and interferences
Wetland types based Geology and on relationshiops with topography
the main aquifer
1. Recharge wetlands High, relatively flat interfluves, detrital Miocene
Small, roughly circular, ephemeral water holes, often forming clusters. Very sharp borders without true phreatophytic belts
Not affected by groundwater withdrawal. Occasional tillage during droughts.
2. Wetlands not Related to Pliocene connected with the outwash deposits main aquifer
Small seepages in the contact with Miocene materials. Glycophytes
Generally ephemeral, sometimes controlled by autonomus small perched aquifers Glycophytes and oxyphytes
Not affected by groundwater withdrawal. Occasional drainage
id. Related to Quternary (aeolian) sand deposits
Small seepages in the contact with underlying Miocene materials. Glycophytes
Not affected by groundwater withdrawal
Very narrow, incise discharges, accopanied by elongated meadows Glycophytes
3.1. Wetlands On relatively steep connected with pediment slopes and local flow alluvial fans, coarse systems detrital Pliocene 3.1
springs and more or less ephemeral watercourses
Not affected by groundwater withdrawal (taking place in lower areas)
3.2. Wetlands On generally flatter connected with areas, Miocene, intermediate or relatively fine textures subregional flow systems
More diffuse, less narrow meadows. Groundwater slightly mineralized give rise to subhalophytes meadows. Characteristic plots mosaic. Soloniets
From tolerant to alkalinophytes, subhalophytes flooded areas and potholes
seepages: - top soil salinization, xerohalophytes - A horizon erosion by wind - potholes and small lakes become ephemeral retaining meteoric water due to impermeable solonetz horizon
On Miocene covered with shallow sand id. deposits or coarser textures
id. - Vegetation changes to therophy tes - Potholes and lakes dry out and do not recover
Miocene 3.3. Wetlands connected with regional flow systems
Extensive meadows with ameboid contours. Saline evaporating surfaces forming interstitial evaporites Halophytes
Small saline sloughs alternating with hummocks (silt dunes, "lunettes") Halophytes
- No groundwater withdrawal due to insuitability for agriculture - Extensive drainage works leading to vegetation changes (xerohalophytes)

"lavajos" with dispersed clay, rather impermeable bottom may still collect occasional meteoric water although they are no longer connected with the main aquifer. On the other hand, the sticky and dense soloniets soil horizons accom- panying them are usually an obstacle for ploughing or recla- mation.
Vast areas of Spain have been affected by groundwater extraction (both central subplateaus, La Mancha and western Andalusia are perhaps the most affected areas). Drainage water disposal is the main threat to the steppe lakes in Aragon. "Reclamation" and irrigated agriculture is univer- sal, permanent source of impact for al1 wetlands.
ACKNOWLEDGEMENTS
This research has been funded by CAYCIT project NAT. 90-0721 -C 02-0 1
REFERENCES
BESTEIRO, A.G. 1992. Limnología de las formaciones palustres situadas sobre el acuqero de Madrid; Análisis de relaciones entre aguas superficiales y subterráneas. Tesis doctoral, Facultad de Ciencias, Universidad Autónoma. Madrid.
DEPARTAMENTO DE ECOLOGIA, U.A.M. 1991. Restauración del territorio afectado por las operaciones de extracción de áridos en El Piul de Rivas (Madrid). Pioneer S.A. Madrid. Universidad Autónoma de Madrid.
FREEZE, R. A. & J.A. CHERRY. 1979. Groundwater. Prentice Hall. New Jersey.
GONZALEZ BERNALDEZ, F. 1987. Las zonas enchar- cables: El marco conceptual. En: Bases Cient@cas para la protección y Gesrión de los humedales en España. Real Academia de Ciencias Matemáticas, Físicas y Naturales. Madrid: 9-30.
GONZALEZ BERNALDEZ, F. 1988. Typology of wetlands of arid and semiarid regions. Proc. Int. Sympos. on the Hydrology of Wetlands in Semiarid and Arid Regions:
GONZALEZ BERNALDEZ, F . 1992. Los paisajes del agua: Terminología popular de los humedales (En prensa).
GONZALEZ BERNALDEZ, F., P. HERRERA, A. SASTRE, A., J.M. REY & R. VICENTE. 1987. Compara- ción preliminar de los ecosistemas de descarga de aguas
25
subterráneas en las cuencas del Duero y del Tajo. Hidroge- ología y Recursos Hidráulicos, 11: 19-39.
GONZALEZ BERNALDEZ, F., C. MONTES, P. HERRERA & A. SASTRE. 1988. Genetical typology of the Madrid aquifer wetlands. Proc. Intern. Sympos. on the Hydrology of Wetlands in Semiarid and Arid Regions:
GONZALEZ BERNALDEZ, F., J.M. REY, C. LEVAS- SOR & B. PECO. 1989a. Landscape ecology of uncultivated lowlands in central Spain. Landscape Ecology, 3(1): 3-18.
GONZALEZ BERNALDEZ, F., C. MONTES, A.G. BESTEIRO, P. HERRERA & C. PEREZ. 1989b. Los hume- dales del acuvero de Madrid. Canal de Isabel 11. Madrid.
GONZALEZ BERNALDEZ, F., B. PECO, C. LEVAS- SOR, A. LLORCA & J. COMPAN. 1989c. La dehesa d p Colmenar. Aula de Ecología. Ayuntamiento de Colmenar Viejo. Madrid.
GONZALEZ BERNALDEZ, F. & J.M. REY BENAYAS. 1992a. Geochemical relationships between groundwater and wetland soils and vegetation in central Spain. Geodermu (In press).
GONZALEZ BERNALDEZ, F., J.M. REY BENAYAS & A. MARTINEZ. 1992b. Typology of impacts produced by groundwater extraction. Journal of Hydrology (In press).
GODRON, M. 1968. Quelques applications de la notion de fréquence en écologie végétale. Oecologiu Plantarum, 3: 185-212.
GROOTJANS, A. P. 1985. Changes of groilndwater regime in wet meadows. Proefschrift ter vekrijging van het doktoraat. Rijksuniversiteit te Groningen.
HERRERA, P. 1987. Aspectos ecológicos de las aguas subterráneas en la facies arcósica de la cuenca de Madrid. Tesis doctoral. Universidad de Alcalá de Henares.
HUBBERT, M.K. 1940. The theory of groundwater movement. Jr. Geol., 48 : 785-944.
LLAMAS, M.R. 1982. Notas sobre peculiaridades de los sistemas hídricos de las zonas húmedas. En: Las Zonas Húmedas de Andalucía. Dirección General de Medio Ambiente, MOPU, Madrid: 77-85.
LLAMAS, M.R. 1988. Conflicts between wetland conser- vation and groundwater explotation: two case histories in Spain. Environmental Geol. Water Sci., l(3): 241-25 1.
REY BENAYAS, J.M. 1989. Ecosistemas de descarga de acuqeros de la cuenca del Duero. Tesis doctoral. Facultad de Ciencias. Universidad Autónoma. Madrid.
REY BENAYAS, J.M., C. PEREZ, F.G. BERNALDEZ & M. ZABALETA. 1989. Tipología y cartografía de los humedales de las cuencas del Duero y Tajo. Mediterranea. Sec. Biol. 12: 5-26.

REY BENAYAS, J.M., F.G. BERNALDEZ, C. LEVAS- TOTH, J. 1963. A theoretical analysis of groundwater flow
SOR & B. PECO. 1990. Vegetation of groundwater in small drainage basins. J. Geophys. Res., 68: 4795-4812. discharge sites in the Douro basin, central Spain. J. Vegeta- WINTER, T.C. 198 l . Effects of the water table configuration
tiotz Science, 1 : 461-466. in seepage through lake beds. Limnol. Ocenogr., 26: 925-934.

LIMNOLOGY IN THE PYRENEAN LAKES
J. Catalan 2, E. Ballesteros 2, L. Camarero ' 2, M. Felip ' and E. Gacia ' 1. Department of Ecology. University of Barcelona. Diagonal 645. 08028 Barcelona. Spain. 2. Institute of High-Mountain Research. University of Barcelona. Apartat 21. 25530 Vielha. Lleida. Spain. 3. Centre of Advanced Studies. CSIC. Carretera de St. Barbara sn. 17300 Blanes. Girona. Spain.
Keywords: High mountain lakes, Pyrenees, Biogeochemistry, Precipitation, Snow, Ice, Plankton, Aquatic macrophytes.
ABSTRACT
The Pyrenees hold more than 1,000 lakes, which provide valuahle sites for limnological research. Since 1983 the Institute of High-Mountain Research of the University of Barcelona is carrying out studies on biogeochemistry, plankton ecology, and physiology and ecology of macrophytes. Between November and December al1 lakes freeze and remain covered in ice and snow for 4 to 7 months, especial attention is paid to this winter period. This paper presents the main results obtained to date, and the
outlines of the ongoing studies.
INTRODUCTION
The Pyrenees hold the main freshwater lake district in Spain. Because of the severe weather and abrupt landscape, the Pyrenean lakes are still pristine aquatic ecosystems, which provide valuable sites for fundamental research and as reference for studies of man-influenced systems (MARGA- LEF, 1985; CATALAN, 1986; CATALAN, 1989b; CATA- LAN, in press). Yet these lakes cannot escape global and regional influences, so that they serve as sensors of global change, both past and future, for their sediments hold a record of the last 40,000 years (VILAPLANA et al., 1989).
DELEBECQUE & RITTERO (1898) provided the first data on the biota of lakes on the French slopes of the Pyre- nees. In the 20's some German naturalists contributed with invertebrate studies of Spanish valley5 (ARDNT, 1926; BOFILL & HAAS, 1920; VIETS, 1930). LOZANO REY (1935) in his study of fish in Spanish rivers included some data on Pyrenean lakes. MARGALEF (1948, 1949, 1952, 1953, 1956) extended our knowledge of the biota of Pyre- nean lakes, especially algae, and started an ecological appro- ach that was followed in the 70's by an extensive study of chemistry, phytoplankton and zooplankton (MARGALEF et al. 1975; VILASECA, 1978; MIRACLE, 1978a, 1978b; CAMPAS & VILASECA, 1979). Detailed studies of the primary production, zooplankton and benthos have only been carried out in the Neouvielle range, especially in the Porth-Bielh lake, as part of the International Biological
Program (CAPBLANCQ & LAVILLE, 1983). Since 1983, limnological studies have been carried out by a working group of the Institute of High-Mountain Research of the University of Barcelona (fig. 1) with the broad aim of understanding the patterns and causes of variability from the
atmospheric input to the sediment record through the lake ecosystem dynamics. In this paper we summarize the present state of knowledge on the lakes and describe ongoing research aims. Most information comes from the studies carried out in a deep cirque lake, Lake Redó, since 1984 (42' 38' 34" N, 0' 46' 1.3" E), a shallow valley lake, Lake Bacivér, since 1987 (42' 41' 46" N, O0 59' 1" E), and from a survey of 102 lakes carried out from July to Septem- ber 1987. This research was in the fields of physical and biogeochemical limnology, and plankton and macrophyte ecology; but much basic information is still lacking for the Pyrenean lakes, especially for benthos (CAPBLANCQ &
LAVILLE, 1983: CATALAN, 1989b).
PHYSICAL LIMNOLOGY
The lakes of the Pyrenees are deep for their modest surface area (fig. 2). As in other ranges with glacial erosion, the lakes can be broadly classified into cirque and valley lakes. Even then, most shallow valley lakes show dimictic behaviour, with mixing periods after thaw in June and during autumn from October to December (CATALAN et
Limnetica, 8: 27-38 (1992) O Asociación Española de Limnología, Madrid. Spain
Figure 1. Map of the area covered by the lirnnological studies of the Institute of High Mountain Research of the University of Barcelona. Circles represent lakes or groups of lakes covered in extensive regional studies. Stars show our rnain locations (Lake Baciver and Lake Redó) for process-oriented studies. Inverted triangles show precipitation sarnpling stations.
al., 1990). Deep lakes show sharp thermoclines during summer between 10 and 15 m deep. Especially interesting is mixing in the hypolimnion in lakes without summer inflow. In a vertical profile in Lake Redó, apparent diffusi- vities are higher between 5 to 10 m below the thermocline (CATALAN, 1988). Microstructure studies have shown that there is turbulent mixing in this zone although it is highly stratified, and this is related with significant boundary mixing close to the shore (IMBERGER, pers. comm.).
Until recently little information has been published on winter conditions. Between November and December al1 the lakes freeze and remain covered in ice and snow for 4 to 7 months. Some differences between these covers and those of the lakes at higher latitudes are remarkable (fig. 3). Except in very unusual years, there is a short period between lake freezing and accumulation of a large amount of snow. This determines that black ice thickness rarely exceeds 30 to 40 cm, whereas total cover thickness ranges between 2 and 5 m. Melting and refreezing of the snow and new snow- falls produces a complex layered structure in the cover, which is further modified by water flooding from the lake. Initially, the black ice supports the weight of the snow, but as this increases, water rises from below until a hydrostatic equilibrium is reached. In Lake Redó, this water accounts for more than 50% of the water equivalent of the cover and the amount is proportional to the accumulated precipitation (CATALAN, 1989a). This flooding of the cover with water from the lake deepens the base of the ice, often leading to scouring of the shallow aquatic macrophytes, and diverting stream flow through the snow cover above the ice, instead of
through the lake (CATALAN et al., 1990). The main hydro- logical event in the lakes is spring thaw. About 30 to 50% of the annual precipitation falls while the lakes are covered by ice, then most of this water circulates through the lake in about one month. For small lakes this means water renewal in the whole lake. Autumn vertical isothermy is longer than spring isotherrriy (CAPBLANCQ & LAVILLE, 1983; CATALAN, 1988) . The duration and intensity of the spring mixing may be closely related to the duration of the ice cover, so that we may expect summer productivity to be negatively correlated to winter precipitation.
There are a few data on annual heat budgets, which lie between 15,000 and 20,000 cal cm-2, from 25 to 50 % corresponding to heat for melting the snow and ice cover. During summer, surface temperature rarely exceeds 15OC, and only in very small ponds.
During the ice-free season, light penetration is very high, the photic zone always being deeper than the thermocline during the stratified period and usually reaching the bottom in lakes less than 20 m deep. When lakes freeze, the accu- mulation of snow on the surface of the lake reduces under- water irradiante to compensatory levels; this condition may last several months.
BIOGEOCHEMICAL LIMNOLOGY
Pyrenean lakes have severa1 features of interest to gene- ral bioyeochemistry: a) catchments are usually of non-sedi-
Area ha Figure 2. Plot of area v r i \ u \ i i i ~ i \ i i i r ~ i ~ i i i l e l ~ l i IOI I ' ) i c i i c ~ n lakes.
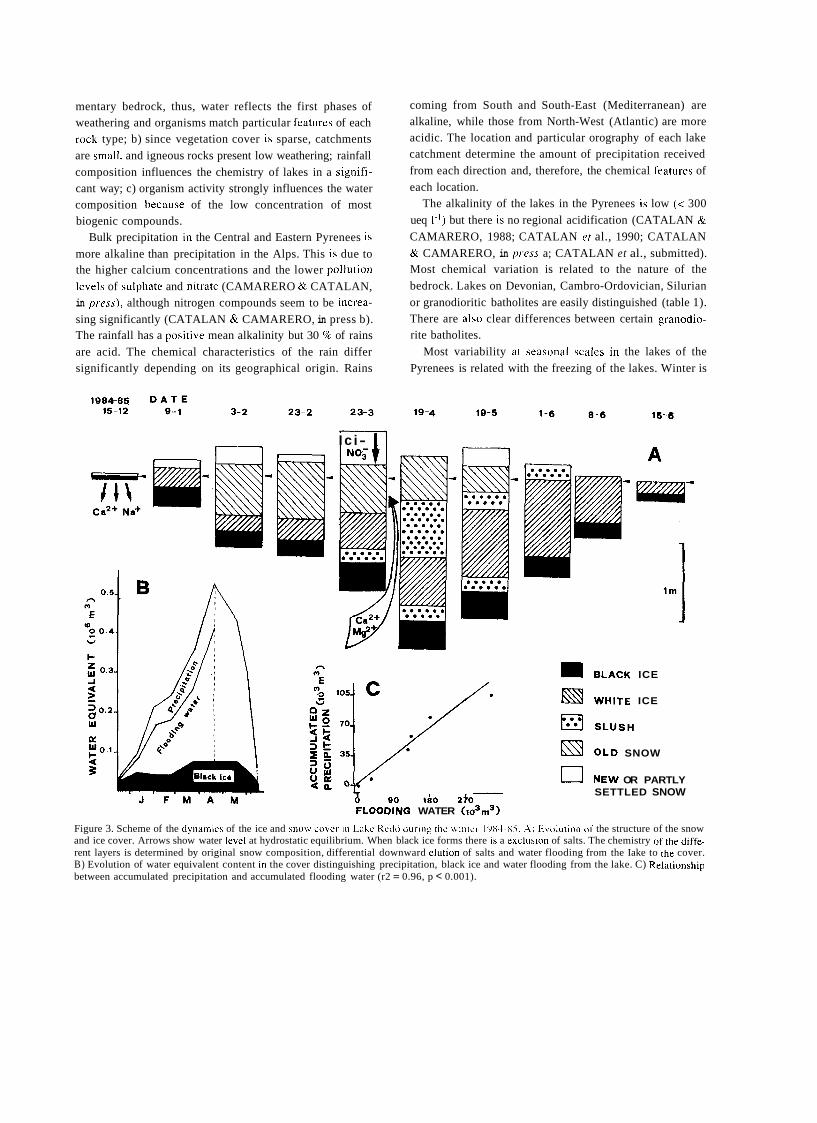
mentary bedrock, thus, water reflects the first phases of weathering and organisms match particular features of each rock type; b) since vegetation cover is sparse, catchments are small, and igneous rocks present low weathering; rainfall composition influences the chemistry of lakes in a signifi- cant way; c) organism activity strongly influences the water composition because of the low concentration of most biogenic compounds.
Bulk precipitation in the Central and Eastern Pyrenees is more alkaline than precipitation in the Alps. This is due to the higher calcium concentrations and the lower pollution levels of sulphate and nitrate (CAMARERO & CATALAN, in press), although nitrogen compounds seem to be increa- sing significantly (CATALAN & CAMARERO, in press b). The rainfall has a positive mean alkalinity but 30 O / o of rains are acid. The chemical characteristics of the rain differ significantly depending on its geographical origin. Rains
coming from South and South-East (Mediterranean) are alkaline, while those from North-West (Atlantic) are more acidic. The location and particular orography of each lake catchment determine the amount of precipitation received from each direction and, therefore, the chemical features of each location.
The alkalinity of the lakes in the Pyrenees is low (< 300 ueq 1.') but there is no regional acidification (CATALAN &
CAMARERO, 1988; CATALAN er al., 1990; CATALAN & CAMARERO, in pvess a; CATALAN et al., submitted). Most chemical variation is related to the nature of the bedrock. Lakes on Devonian, Cambro-Ordovician, Silurian or granodioritic batholites are easily distinguished (table 1). There are also clear differences between certain granodio- rite batholites.
Most variability at seasonal scales in the lakes of the Pyrenees is related with the freezing of the lakes. Winter is
I c i - I l
b 90 180 230 FLOODING WATER (ro3rn3)
BLACK ICE
m WHlTE ICE
SLUSH
OLD SNOW
O NEW OR PARTLY SETTLED SNOW
Figure 3. Scheme of the dynamics of the ice and snow covcr 111 LaLc Kcclo clui-1112 tiic \~iiitci. I')S4-X.í. .A) E\olutioti of the structure of the snow and ice cover. Arrows show water level at hydrostatic equilibrium. When black ice forms there is a exclusion of salts. The chemistry of the diffe- rent layers is determined by original snow composition, differential downward elution of salts and water flooding from the Iake to the cover. B) Evolution of water equivalent content in the cover distinguishing precipitation, black ice and water flooding from the lake. C) Relationship between accumulated precipitation and accumulated flooding water (r2 = 0.96, p < 0.001).
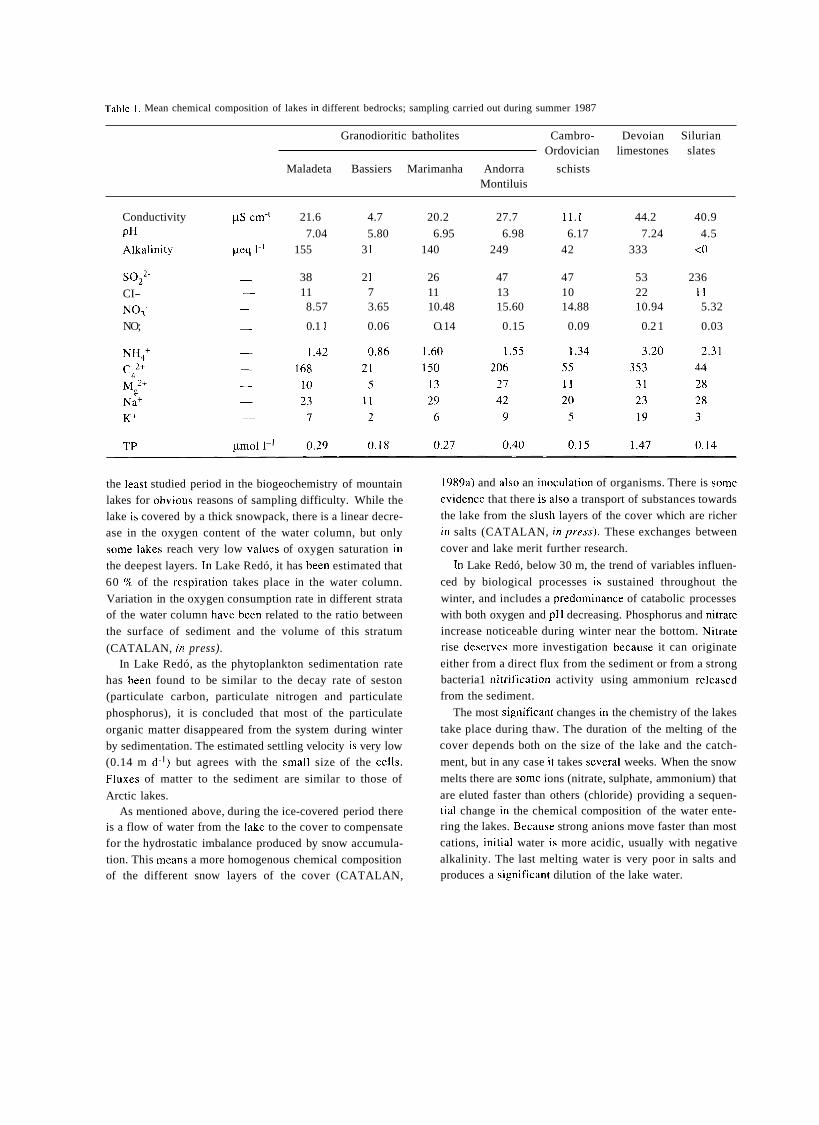
Table 1. Mean chemical composition of lakes in different bedrocks; sampling carried out during summer 1987
Granodioritic batholites Cambro- Devoian Silurian Ordovician limestones slates
Maladeta Bassiers Marimanha Andorra schists Montiluis
Conductivity pS cm-' 21.6 4.7 20.2 27.7 11.1 44.2 40.9
PH 7.04 5.80 6.95 6.98 6.17 7.24 4.5 Alkalinity peq 1.' 155 3 1 140 249 42 333 <O
so,2- - 3 8 2 1 26 47 47 53 236 CI- - 11 7 11 13 10 22 I I
NO; - 8.57 3.65 10.48 15.60 14.88 10.94 5.32
NO; - 0.1 1 0.06 O. 14 0.15 0.09 0.2 1 0.03
the least studied period in the biogeochemistry of mountain 1989a) and also an inoculation of organisms. There is some
lakes for obvious reasons of sampling difficulty. While the evidence that there is also a transport of substances towards
lake is covered by a thick snowpack, there is a linear decre- the lake from the slush layers of the cover which are richer
ase in the oxygen content of the water column, but only in salts (CATALAN, in pvess). These exchanges between
some lakes reach very low values of oxygen saturation in cover and lake merit further research.
the deepest layers. In Lake Redó, it has been estimated that In Lake Redó, below 30 m, the trend of variables influen-
60 % of the respiration takes place in the water column. ced by biological processes is sustained throughout the
Variation in the oxygen consumption rate in different strata winter, and includes a predominante of catabolic processes of the water column have been related to the ratio between with both oxygen and pH decreasing. Phosphorus and nitrate
the surface of sediment and the volume of this stratum increase noticeable during winter near the bottom. Nitrate
(CATALAN, in press). rise deserves more investigation because it can originate
In Lake Redó, as the phytoplankton sedimentation rate either from a direct flux from the sediment or from a strong
has been found to be similar to the decay rate of seston bacteria1 nitrification activity using ammonium released
(particulate carbon, particulate nitrogen and particulate from the sediment.
phosphorus), it is concluded that most of the particulate The most significan1 changes in the chemistry of the lakes
organic matter disappeared from the system during winter take place during thaw. The duration of the melting of the
by sedimentation. The estimated settling velocity is very low cover depends both on the size of the lake and the catch-
(0.14 m d.') but agrees with the small size of the cells. ment, but in any case it takes several weeks. When the snow
Fluxes of matter to the sediment are similar to those of melts there are some ions (nitrate, sulphate, ammonium) that
Arctic lakes. are eluted faster than others (chloride) providing a sequen- As mentioned above, during the ice-covered period there tia1 change in the chemical composition of the water ente-
is a flow of water from the lake to the cover to compensate ring the lakes. Because strong anions move faster than most
for the hydrostatic imbalance produced by snow accumula- cations, initial water is more acidic, usually with negative
tion. This means a more homogenous chemical composition alkalinity. The last melting water is very poor in salts and
of the different snow layers of the cover (CATALAN, produces a significan1 dilution of the lake water.

PLANKTON ECOLOGY
Small coccoid species and flagellates predominate in the phytoplankton of the Pyrenean lakes. Chrysophyceae are quite common (e.g. genera Chromulina, Ochr-omonas, Dinohryon, Bitrichia, Chrysolykos, Pseudokephyrion). and many of them show phagotrophy. Synurophyceae are much les5 common, no large population has been reported and there seems to be little specific variety in the Pyrenees. Diatoms appear during mixing periods in colonial forms (Melosira, Tahellaria), and some heliophilous Cyclotella subsist in the epilimnion during the summer stratification. Desmids are rare, although some very small Cosmarium can reach a rele- vant biomass in some lakes. Chlorococcales (Distyosphae- riun, Monoraphidium, Sphaerocystis, etc.) and species from other groups with similar organization (e.g., the chrysophyte Stichogloea) constitute most of the biomass during large periods of the year. Chlorophytes persist longer under ice than Chrysophytes. Cryptophytes are frequent throughout the year, sometimes peaking soon after melting. Small Dinoflagellates, some of them colourless, are also common throughout the year but larger forms (Per-idinium) are restricted to summer, and Ceratium has not been reported.
Small zooplankton is rather diverse throughout the year. Colourless flagellates (Oicomonas, Spumella, Monosiga, etc.) increase in biomass and diversity throughout winter. Ciliates show a great variety (Strumhiditam, Halteria, etc.), especially in late winter when unusual ciliates in the plank- ton can be found (Dileptus, Lacrymaria, etc.). Rotifers cons- titute from 2 to 10 % of summer zooplankton biomass, and
during winter when they can represent most of its biomass if Daphnia is absent. More than sixty species have been found in the Pyrenean lakes; the most common are Ascomorpha ecaudis, Asplanchna priodonta, Polyarthra spp. and the boreo-alpine species, Kellicottia longispina. This latter reaches its maximum biomass in mid winter, under ice
(Catalan, unpubl.). Large zooplankton consist of few species. Cyclops abys-
sorum can be found in most lakes, individuals survive during winter in a resting stage in the sediment, but a small percentage of copepodites persist in a free-swimming form. Daphnia longispinu is also very common, but with a less predictable seasonal pattern. Other planktonic Cladocera (Scapholeheris mucronata, Holopedium gihberum, Daphnia obtusa, D. pulicar-ia and Polyphemus pediculus) can occa- sionally be found. Diaptomids have a biogeographic interest in the Pyrenees because of the absence of the two most common species in the Alps. There are two large diapto- mids, Diaptomus cyaneus and D. castaneti, and two small diaptomids, Mixodiaptomus laciniatus and Eudiuptomus ~ulgaris . There is size segregation; the two large species or the two small species do not coexist in any lake (MIRA- CLE,1978a, 1978b).
As yet there are no measurements of bacteria1 activity
available for these lakes. However, indirect evidence, through changes in dissolved organic compounds, suggest significant activity even during the large periods when water temperature is below 3' C (CATALAN, in press).
According to the total phosphorus (TP) levels, more than 70 % of the lakes are oligotrophic (TP i 0.3 umol l.'), 25 %
1984 1985 Figure 4. lsopleth diagrairi fol- teniperature ("C. dashed lines) and chlorophyll (rng Chl rn~') in Lake Redó from June 1984 to Auguat 1985

are mesotrophic (0.3 < TP < I umol l.'), and only a few very shallow lakes, without a permanent outlet and rich in macrophytes and littoral vegetation, are eutrophic (TP > 3 umol 1.'). These lakes present much higher values of ammo- nium, nitrite, chloride and potassium, which relates the eutrophication to the presence of cattle (CATALAN et al.,
submitted). There is no significant statistical relation between any
single regional, morphometric or physiographic factor and mesotrophy. However a) lakes on Andorra-Montlluís grano-
dioritic batholite are al1 mesotrophic, although they include lakes at different altitude and with different morphometric characteristics. b) Although globally there is no correlation between TP and depth, the lakes with the highest TP values
inside each cirque or basin were usually shallow (< 5 m
deep). c) Most deep lakes with high TP are large and expe- rience level fluctuations because of hydroelectrical exploi- tation. Globally, however, there is no clear distinction either
in chemistry or in trophic status between lakes which have suffered modification to increase hydroelectrical power
resources and undisturbed lakes, except for specific extreme cases (CATALAN et al., submitted).
Seston composition reflects the scarcity of phosphorus,
Lake Redó presents C:N:P ratios about 345:34: 1 by atoms throughout the year. Productive periods are linked to the increase in phosphorus diffusion from the sediments by mixing in the water column (fig. 4). After a quick increase
Figure 5. Pl~oto:itlapcaiioii i i i L,iAi. Ki.clo c l ' > i c . ~ i ~ ~ i . \ ) i l u i III; 1 OSR-S9 winter. During the ice-free period (A) vertical mixing did not allow Ik differentiation, and phytoplankton was adapted to mean depth irradiance. During the ice-covered period with little snow (B) Ik variability followed light values, time of mixing was longer than time of adaptation. When more snow was accumulated in the cover (C), phytoplankton was not able to adapt its Ik to the very low irra- diance.
during these periods, chlorophyll decrease very slowly over long periods in deep lakes, either in the hypolimnion during summer, where a deep chlorophyll maximum appears, or
under ice during winter (CATALAN & CAMARERO, 1991). In situ recycling seems to play a significant role in maintaining the planktonic comillunity, although fertilizing
mechanisms related with boundary mixing and lateral intru-
sions rnay be also important. Mountain lakes are particularly interesting sites in which
to investigate scales of photoadaptation in oligotrophic systems. High transparency in summer combines with very low levels during winter below the snow and ice cover. For
instante, the transition from autumn overturn to mid winter in Lake Redó is a good illustration of the relationship between mixing and light regimes, and photosynthetic adap-
tations (CATALAN, 1991). During overturn, phytoplankton
distribution is rather homogeneous. Yet, there is a sigriifi- cant differentiation in a (the slope of the photosynthesis vs.
irradiance curves) and Pmax (maximum production at light
saturation) between surface and deep (> 1 5 m) layers. Howe-
ver Ik (Iower light intensity to reach Pmax, which is equal to Pmaxla), which requires a longer adaptation time because it represents a change in the size of the photosynthetic units
(HARDING et al., 1987) is quite similar for phytoplankton
populations at any depth, and its value corresponds to mid- depth light intensity (25 m). This means that during autumn overturn in Lake Redó the mixing time of phytoplankton is
shorter than the time of Ik adaptation and longer than the time of a adaptation, and Pmax changes parallel to a. During
overturn, the mixing time of heat has been estimated to be less than an hour (CATALAN, 1988). For phytoplankton it is likely to be longer, but less than a day. However, when
ice forms on the lake surface, turbulent mixing is markedly
reduced and, as a consequence, phytoplankton populations are subjected to a determinate light level during longer periods, then Ik adaptation to each level is possible (fig. 5). Finally, when snowpack is thick, available light falls to very low levels. Ik becomes very low but it does not reach the values of light in the water column and so it seems impossi- ble for these algae to adapt fully to such low light levels.
MACROPHYTE ECOLOGY
Nearly 65 O/c of the ca. 1000 lakes in the Pyrenees have submerged aquatic macrophytes. Species richness (fig. 6) of these lakes is higher than that of the lakes from other Euro- pean chains (BRAUN BLANQUET, 1948). Quillworts,
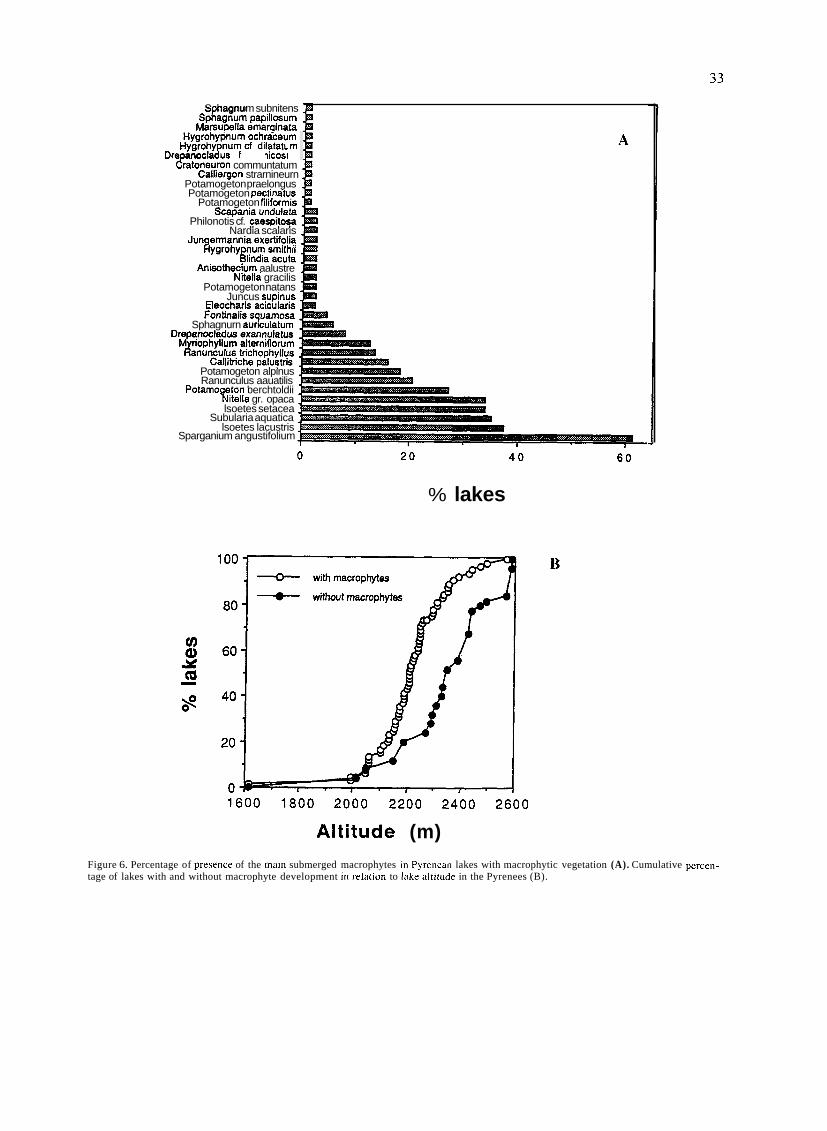
S hagnum subnitens .. sptagnum papiii?sum Marsuwlla emaralnata
Hygrohypnum ochrCceum Hygrohypnum cf. dilatatum
Drepanocladus cf. vermicosus Cratoneuron communtatum
Calliergon strarnineurn Potamogeton praelongus Potamogeton pectinahis
Potamogeton filiformis Scapanta undulata
Philonotis cf. caespitosa Nardia scalaris
Anisothecium aalustre Nitella gracilis
Potamogeton natans Juncus su~inus E
Eleocharis acicularis Fontinalis squamosa
Sphagnurn auriculaturn Dre anocladw exannulatus
hRriophyllum alterniflorurn 4anuncul"s trichophyll"~
Callitnche palustris Potamogeton alpinus Ranunculus aauatilis
Potarno eton berchtoldii t i t e l a gr. opaca lsoetes setacea
Subularia aquatica lsoetes lacustris
Sparganium angustifolium
% lakes
Altitude (m)
Figure 6. Percentage of presence of the main submerged macrophytes in Pyrenean lakes with macrophytic vegetation (A). Cumulative percen- tage of lakes with and without macrophyte development in relation to lake altitude in the Pyrenees (B).
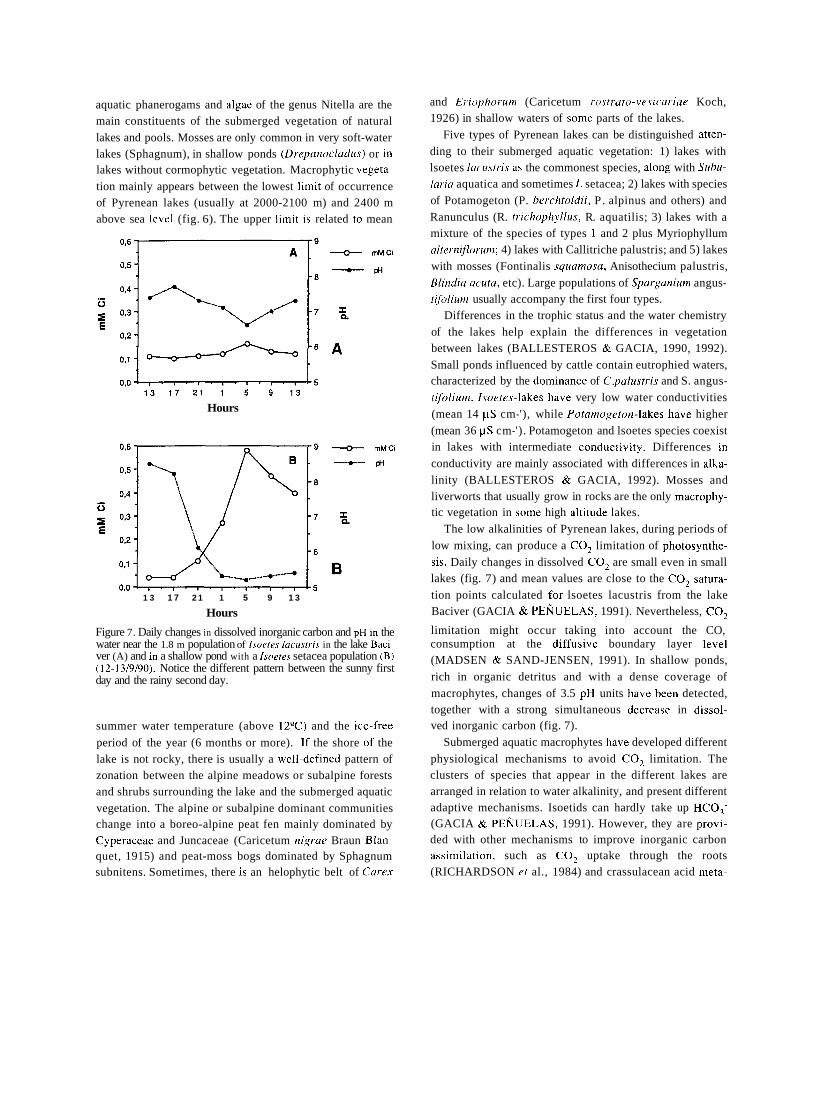
aquatic phanerogams and algae of the genus Nitella are the main constituents of the submerged vegetation of natural lakes and pools. Mosses are only common in very soft-water lakes (Sphagnum), in shallow ponds (Drepanocludus) or in lakes without cormophytic vegetation. Macrophytic vegeta- tion mainly appears between the lowest limit of occurrence of Pyrenean lakes (usually at 2000-2100 m) and 2400 m above sea level (fig. 6). The upper Iimit is related to mean
Hours
1 3 1 7 2 1 1 5 9 1 3
Hours
Figure 7. Daily changes in dissolved inorganic carbon and pH in the water near the 1.8 m population of lsoetes lacustris in the lake Baci- ver (A) and in a shallow pond with a lsoeres setacea population (B) (12-13/9/90), Notice the different pattern between the sunny first day and the rainy second day.
summer water temperature (above 12") and the ice-free
period of the year (6 months or more). If the shore of the lake is not rocky, there is usually a well-defined pattern of zonation between the alpine meadows or subalpine forests and shrubs surrounding the lake and the submerged aquatic vegetation. The alpine or subalpine dominant communities change into a boreo-alpine peat fen mainly dominated by Cyperaceae and Juncaceae (Caricetum nigrae Braun Blan- quet, 1915) and peat-moss bogs dominated by Sphagnum subnitens. Sometimes, there is an helophytic belt of Cares
and Eriophorum (Caricetum rostrato-vesicariue Koch, 1926) in shallow waters of some parts of the lakes.
Five types of Pyrenean lakes can be distinguished atten- ding to their submerged aquatic vegetation: 1) lakes with lsoetes Iuc~~stris as the commonest species, along with Suhu- laria aquatica and sometimes 1. setacea; 2) lakes with species of Potamogeton (P. berchtoldii, P. alpinus and others) and Ranunculus (R. trichophyllus, R. aquatilis; 3) lakes with a mixture of the species of types 1 and 2 plus Myriophyllum alterniflflor.um; 4) lakes with Callitriche palustris; and 5) lakes with mosses (Fontinalis squamosa, Anisothecium palustris, Blindia acutu, etc). Large populations of Sparganium angus- tif(jlium usually accompany the first four types.
Differences in the trophic status and the water chemistry of the lakes help explain the differences in vegetation between lakes (BALLESTEROS & GACIA, 1990, 1992). Small ponds influenced by cattle contain eutrophied waters, characterized by the dominance of C.palustris and S. angus- tifolium. Isoetes-lakes have very low water conductivities (mean 14 yS cm-'), while Potamogeton-lakes have higher (mean 36 pS cm-'). Potamogeton and lsoetes species coexist in lakes with intermediate conductivity. Differences in conductivity are mainly associated with differences in alka- linity (BALLESTEROS & GACIA, 1992). Mosses and liverworts that usually grow in rocks are the only macrophy- tic vegetation in some high altitude lakes.
The low alkalinities of Pyrenean lakes, during periods of low mixing, can produce a C0, limitation of photosynthe- sis. Daily changes in dissolved C 0 2 are small even in small lakes (fig. 7) and mean values are close to the C0 , satura- tion points calculated Sor lsoetes lacustris from the lake Baciver (GACIA & PEÑUELAS, 1991). Nevertheless, CO,
limitation might occur taking into account the CO, consumption at the diffusive boundary layer level (MADSEN & SAND-JENSEN, 1991). In shallow ponds, rich in organic detritus and with a dense coverage of macrophytes, changes of 3.5 pH units have been detected, together with a strong simultaneous decrease in dissol- ved inorganic carbon (fig. 7).
Submerged aquatic macrophytes have developed different physiological mechanisms to avoid C0, limitation. The clusters of species that appear in the different lakes are arranged in relation to water alkalinity, and present different adaptive mechanisms. Isoetids can hardly take up HC03- (GACIA & PENUELAS, 1991). However, they are provi- ded with other mechanisms to improve inorganic carbon assimilation, such as COZ uptake through the roots (RICHARDSON et al., 1984) and crassulacean acid meta-
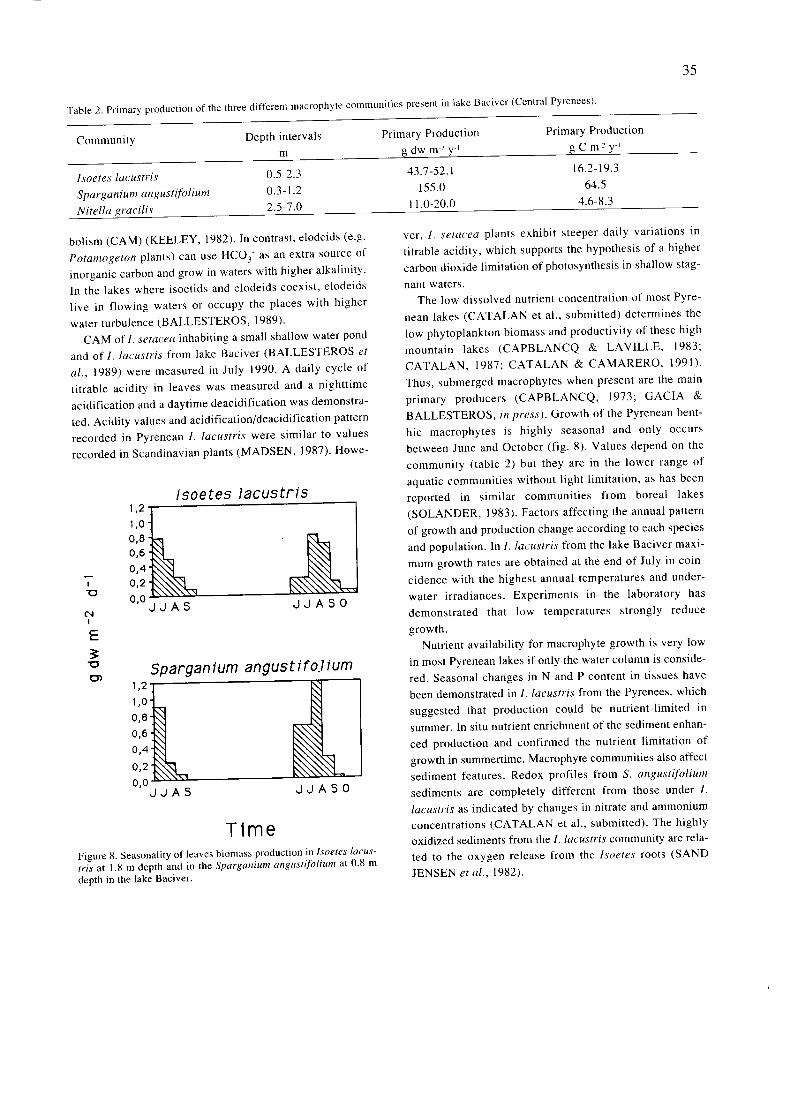

ONGOING RESEARCH REFERENCES
Research in Lake Redó and Lake Baciver is still being carried out. In lake Redó a dual approach to lake ecosystem is intended through the evaluation of carbon, nitrogen and phosphorus cycles together with the study of the life-histo- ries of the plankton organisms. Assuming hydrodynamics as main driving force of the changes in the lake system, we study how life mediates biogeochemical changes. Each season provides interesting aspects. The following aspects are under current research during winter: 1) inoculation of organisms from the lake to the snow and ice cover and biolo- gical activity in the slush layer; 2) changes in the function and structure of the plankton induced by the sudden freezing of the lake surface; 3) microbial succession during winter; and sediment-water interaction during winter inverse strati- fication. Summer investigation is concentrated in the study of the fertilizing and maintenance mechanisms of the deep chlorophyll maximum and production in the hypolimnion.
In Lake Baciver we are evaluating the ecological changes after the construction of a dam that has raised the water level by 7 m. Emphasis is placed on the effects on macrophyte
development and growth. Further investigation is being carried out on the relative importance of P and N in the sedi- ment interstitial waters in the limitation of the primary production of Isoetes primary production; the control of primary production in Potamogeton-lakes; factors contro- lling macrophyte distribution patterns; and detailed studies in the maintenance of CAM metabolism in emergent Isoe- tes setacea plants.
Future research includes palaeolimnology, studying chan- ges during the industrial period and, at longer time scales, during the last 40.000 years in connection with another rese- arch group in the Institute of High-Mountain Research (VILAPLANA et al., 1983; MONTSERRAT & VILA- PLANA, 1987; VILAPLANA et al., 1989; VILAPLANA &
BORDONAU, 1989).
ACKNOWLEDGMENTS
Many people have collaborated with us in the field, lab and data processing work during the last 7 years in a friendly atmosphere. We are especially in debt to those that helped in sampling in early studies when our expe- rience in high mountain was nil. Research has been funded by CIRIT, Caixa de Barcelona and CICYT (project NAT89-0943).
ARDNT, W., 1926. Spongilliden und Turbellarien aus dem nord-lichen und ostlichen Spanien. Senckenhergiana, 8: 22-30.
BALLESTEROS, E., 1989. Production of seaweeds in Northwestern Mediterranean marine communities: its rela- tion with environmental factors. Scient. Mar., 53: 357-364.
BALLESTEROS, E. & E. GACIA, 1990. Els macrofits aquatics dels estanys del Parc Nacional d'Aigües Tortes i Estany de Sant Maurici. In La investigacirj al Parc Nacional d'Aigües Tortes i Estany de Sant Maurici: 105- 121. Gene- ralitat de Catalunya. Lleida.
BALLESTEROS, E. & E. GACIA, 1992. Una nova asso- ciació de plantes aquatiques als Pirineus: el Ranunculo eradi- cati-Potametum alpini. Butll. Inst. Cat. Hist. Nat., 59: 82-88.
BALLESTEROS, E., E. GACIA & L. CAMARERO, 1989. Composition, distribution and biomass of benthic macrophyte communities from lake Baciver, a spanish alpine lake in the Central Pyrenees. Annals. Limnol., 25: 177- 184.
BRAUN BLANQUET, J., 1948. La végétation ulpine des
Pyrenées Orientales. CSIC. Barcelona. CAMARERO, L. & J . CATALAN, 1991. Horizontal
heterogeneity of phytoplankton in a small high mountain lake. Verh. Internat. Verein. Limnol., 24: 1005-1010.
CAMARERO, L. & J . CATALAN, (in press). Chemical composition of the rain in the Pyrenees. Atmospheric Envi- ronment.
CAMPAS, L. 1979. Els llacs i els estanys. In Elpatrimoni
natural d'Andorra (R. FOLCH, ed.). Ketres. Barcelona. CAMPAS, L. & J.M. VILASECA. 1979. Els llacs piri-
nencs. Quad. Ecol. Apl., 4: 25-36. CAPBLANCQ, J., 1973. Phytobenthos et productivité
primaire d'un lac de haute montagne dans les Pyrénées Centrales. Annals. Limnol., 9: 193-230.
CAPBLANCQ, J. & H. LAVILLE, 1983. Le lac de Port Bielh (Pyrénées): exemple de fonctionnement d'un écosystkme lacustre de haute montagne. In Prohlemes
d'Ecologie: Structure et fonctionnernent des écosystt?mes
limniques (M. LAMOTTE & F. BOURL-RE, eds.): 5 1-80. Masson. Paris.
CATALAN, J., 1986. Quan el Pirineu esdevé un labora- tori. Analisi del projecte d'estudi limnologic de l'estany Redó. Espais, 2: 29-33.
CATALAN, J. 1987. Limnologia de 1'Estany Redó (Piri- neu Central). El sistema pelagic d'un llac profund d'alta muntanya. Ph. D. Thesis. University of Barcelona. Barcelona.

CATALAN, J., 1988. Physical properties of the environ- ment relevant to the pelagic ecosystem dynamics of a deep high-mountain lake (Estany Redó, Central Pyrenees). Orco-
logia uquutica, 9:89- 123. CATALAN, J., 1989a. The winter cover of a high-moun-
tain mediterranean lake (Estany Redó, Pyrenees). Water
Resourc,es Research, 25: 5 19-527. CATALAN, J., 1989b. Els estanys del Pirineu. In HistC,-
ria Natural dels Puijos Catalans ( J. TERRADES, ed.) 14: 384-395. Enciclopedia Catalana. Barcelona.
CATALAN, J., 1991. The relationship between the func- tional anatomy of lakes and primary production. 0ec.ologia
uquarica, 10: 77-94. CATALAN, J., (in press). Evolution of dissolved and
particulate matter during the ice-covered period in a deep high-mountain lake. Crrn. J. Fish. aquar. Scien..
CATALAN, J.. E. BALLESTEROS, L. CAMARERO & E. GACIA, 1990. Aspectos generales de la limnología invernal de dos lagos pirenaicos morfológicamente muy distintos (Estany Redó y Estany Baciver). Scieritia Ge~.undensis, 16: 55-68.
CATALAN, J., E. BALLESTEROS, E. GACIA, A. PALAU & L. CAMARERO, (submitted). Chemical compo- sition of disturbed and undisturbed high-mountain lakes in the Pyrenees: a reference for acidified sites. M'ater research.
CATALAN, J. & L. CAMARERO, 1988. Determinación de los componentes del sistema carbónico-carbonatos de las aguas dulces mediante la titulación de Gran. Tecnología del
agua, 51: 66-75. CATALAN, J. & L. CAMARERO, 1991. Ergoclines and
biological processes in high-mountain lakes: similarities between the summer stratification and the ice-forming period in Lake Redó (Pyrenees). Verh. Internat. Verein. Limnol., 24: 101 1-1015.
CATALAN, J. & L. CAMARERO, (in press a). Limno- logical studies in the lakes of the Spanish Pyrenees (1984- 90). Memorie dello lstitlrto Italiano di Idrohiologia.
CATALAN, J. & L. CAMARERO, (in press b). Compo- sició química de la pluja al Parc Nacional d'Aigües Tortes i de 1'Estany de Sant Maurici. Acres cie les 2oties Jornades
sobre la Reserca al Purc Nacional d'Aigües Tortes i de
I'Estatzy de Sant Mau~.ici. CATALAN, J . , L. CAMARERO, E. GACIA, E.
BALLESTEROS & M. FELIP, (submitted). Nitrogen in the
Pyrenean Lakes. Hydrohiologia. CATALAN, J., L. CAMARERO, E. GACIA. E.
BALLESTEROS & A. PALAU. 1990. Avaluació del risc d'eutrofització i acidificació dels estanys del Pirineu. Caixa de Barceloiia. Barcelona.
CATALAN, J., L. CAMARERO & A. PALAU, 1990.
Composició química de I'aigua dels estanys del Parc Nacio- nal d'Aigües Tortes i Sant Maurici. La ini)estigació al Par-c Nacional d'Aigiies Tortes i de I'Estuny de Sanr Maurici: 90- 104. Generalitat de Catalunya. Lleida.
DELEBECQUE, A. & E. RITTERO. 1898. Sur quelques lacs des Pyrenees Orientales des Hautes et des Basses Pyre- nees. Comp. Rend. de I'Acad. des Scienc., 127.
GACIA, E. & E. BALLESTEROS, (in press). Leaf production in Isoetes luc~rstris L.: a critica1 assessment. Verh. int. Ver. Limnol., 24.
GACIA, E. & J. PENUELAS, 1991. Carbon assimilation of Isoetes 1ac.ustri.s L. from Pyrenean lakes. Photosyntherica,
25: 97-104.
HARDING, L. W. Jr., T. T. FISHER Jr. & M. A. TYLER, 1987. Adaptive response of photosynthesis in phytoplank- ton: specificity to time-scale of change in light. Biological
Oceanogr., 4: 403-437. KEELEY, J . E., 1982. Distribution of crassulacean acid
metabolism in the geiius Isoetes. Anrer. .I. Bor., 69: 254-257. LOZANO REY, L. 1935. Los peces fluviales de
España. Mem. R. Acad. Cienc.. Esac. Fisc. Nat., Ser. Cien. Nat. 55.
MADSEN, T. V., 1987. Interactions between interna1 and externa1 C 0 2 pools in the photosynthesis of the aquatic CAM plants Littorella ~lniflora (L.) Aschers. and Isoetes
lac~~rstris L.. New Phytol., 106: 35-50. MADSEN T.V. & K. SAND JENSEN, 199 1. Photosynthetic carbon assimilation in aquatic iiiacrophytes. Ayuat. Bot., 41: 5-40.
MARGALEF, R. 1948. Flora y fauna y comunidades bióticas de las aguas dulces del Pirineo de la Cerdaña. Monogr. Est. Esf. Pirenaicos, 1 1 : 1-226.
MARGALEF, R. 1949. La vida en los lagos de alta montaña de los Pirineos. Pirirzeos, 1 1 - 12: 5-30.
MARGALEF, R. 1952. La vida en las aguas dulces de Andorra. Prim. Congr. Inf . del Pirineo. [nst. Est. Pir., 69: 1-107.
MARGALEF, R. 1953. Algunos organismos interesantes de las aguas dulces de los Pirineos. Pirineos, 29: 407-420.
MARGALEF, R. 1956. Estudios hidrobiológicos en los valles de Boi (Prov. de Lérida). Acres 2eme Geners. Intern. Et. Pyi.eneennes, 87-108.
MARGALEF, R. 1985. La limnologia a Catalunya. Memor.ias de la Real Academia de Ciencias y Artes de Barc,elona, 46: 203-223.
MARGALEF, R.; L. CAMPAS; M.R. MIRACLE & S.M.
VILASECA. 1975. Introducción al estudio de los lagos pire- naicos. Nat~trulia Hispanicu, 4: 1-47.

MIRACLE, M.R. 1978a. Composición específica de las comunidades zooplanctónicas de 153 lagos de los Pirineos y su interés biogeográfico. Oecologia aquatica, 3: 167- 191.
MIRACLE, M.R. 1978b. Historical and ecological factors concurring in the distribution , biometry and fertility of planktonic crustaceans in Pyrenean Lakes. Verh. Internat.
Verein. Lirnnol., 20: 1657- 1663. MONTSERRAT, J. & J . M. VILAPLANA, 1987. The
paleoclimatic records of the Upper Pelistocene and Holo- cene in the Llauset valley. Pirineos, 129: 107- 1 13.
RICHARDSON, K., H. GRIFFITHS, M. L. REED, J. A. RAVEN & N. M. GRIFFITHS, 1984. Inorganic carbon assi- milation of the isoetids Isoetes lacushis L. and Lobelia dort-
manna L. Oecologia, 61: 115-121. SAND JENSEN, K., C. PRAHL & H. STOKHOLM,
1982. Oxygen release from roots of submerged aquatic macrophytes. Oikos, 39: 349-354.
SOLANDER, D., 1983. Biomass, production and nutrient content of the macrophytes in a natural and a fertilized subarctic lake. Acta Univ. Uppsala, 676: 1 - 17 1.
VIETZ, K. 1930. Zur kenntnis der hydracarinen-fauna von Spanien. Arch. Hydrobiol., 3 1 : 175-240, 359-446.
VILAPLANA, J. M. & J. BORDONAU, 1989. Dynami- que sédimentaire lacustre de marge glaciaire i le paléolac de Llestui (Noguera Ribagorcana - Vessant Sud des Pyrénées). Bull. A.F.E.Cl., 1984-4: 219-224.
VILAPLANA, J. M., J. MONTSERRAT & C. SCHLÜTER, 1989. Recent progress in Quaternari strati- graphy: The Lake Llauset sequence in the Spanish Pyre- nees. pp. 113-124. In Quaternary type sections, Imagi- nation or Reality? (C. SCHLUTER, ed.). Balkema. Rotterdam.
VILAPLANA, J. M., C. SCHLÜTER & A. VERDA- GUER, 1983. Sedimentology and stratigraphy of the Pleistocene sediments in lake Llauset (Southern Pyrenees, Spain): a first approach. Acta Geol. Hisp., 18: 235-248.
VILASECA, J.M. 1978. Fitoplancton de los lagos Pire- naicos. Memoria de Licenciatura. Universidad de Barcelona. Barcelona.

SOUTHERNMOST HIGH MOUNTAIN LAKES IN EUROPE (SIERRA NEVADA) AS REFERENCE SITES FOR POLLUTION AND CLIMATE CHANGE MONITORING.
R. M o r a l e s - B a q u e r o l , P. Carrillo', L. C r u z - P i z a r r o 2 and P. S a n c h e z - C a s t i l l o 3
1. Departamento de Biología Animal y Ecología, Fac. Ciencias, Universidad de Granada. Spain. 2. Instituto del Agua, Universidad de Granada. Spain. 3. Departamento de Biología Vegetal, Fac. Ciencias, Universidad de Granada. Spain.
Keywords: High mountain lakes, Regional limnology, Sierra Nevada, Spain.
ABSTRACT
This paper summarize, from a regional limnology point of view, some of the identifying characteristics of Sierra Nevada high mountain lakes, as a result of the studies carried out by the limnology group of the University of Granada since 1975. A overall narrow variability in the morphological, physical and chemical characteristics of this lakes seems to appear as a conse- quence of a smaller and more homogeneous distribution area than high mountain lakes of other mountain ranges (¡.e. the Alps) cover. The position of Sierra Nevada, far away from industrialized areas, keep their water bodies free of air pollution which reflects in the relatively high levels of alkalinity compared with other mountain lakes under acidification processes (Le. Norwe- gian ones). Likewise, their southern position makes the Sierra Nevada high mountain lakes ideal reference sites for climate change monitoring. The homogeneity in their abiotic environment is responsible of a limited biological diversity of their plank- tonic communities. The phytoplankton is dominated by nanoplanktonic species and the zooplankton is only composed by 20 roti- fer species, 8 cladocerans and 5 copepods. All these characteristics underline the interest of Sierra Nevada lakes as subject of scientific research both applied as in a purely theoretical plane.
INTRODUCTION
Small high mountain lakes, under severe physical condi- tions, have for long been recognized as unique inland waters ecosystems. Properties linked to a fluctuating climate, short growing seasons, low thermal stability, high irradiance and, usually, low nutrient levels, in turn due to small sized watersheds and weak human influence, are al1 responsible for their relatively simplified biological communities, thus being suitable sites to study the basic relationships between their components.
Their, at times, remote location can explain their knowledge to be far from desirable, and, because of their mostly glaciar origin, the bulk of the available informa- tion comes from septentrional and central mountains of the northern hemisphere where the glacial activity had its maximun impact (TONOLLI & TONOLLI, 1951;
PENNAK, 1955; THOMASSON, 1956: ANDERSON, 1974 ...). Toward southern latitudes, as higher altitudes are needed to observe such effects, areas suited to having
comparable lakes became much more scarce and isola- ted.
The Sierra Nevada (37W), is the most southerly inoun- tain system in Europe reaching altitudes higher than 3000 m, and embraces in its highest zone some 50 small water bodies, whose primary interest as reference sites in any comparison study concerning climatic factors and pollution background levels, reveals obvious.
Appart from some spot data coming from studies by LOFFLER (1 974) and GONZALEZ-GUERRERO (1 9 7 3 , an integrated research program was started by MARTINEZ (1975) at the limnology group of the University of Granada. In this paper we summarize some of the identifying characte- ristics of these lakes from a regional limnology point of view.
LlMITED VARIABILITY AMONG LAKES.
The glacial action in the Sierra Nevada was limited to the upper areas, close lo the crest lines, and so most of the
Llmnetica, 8: 3 9 4 7 (1992) O Asociación Española de Limnología, Madrid. Spain
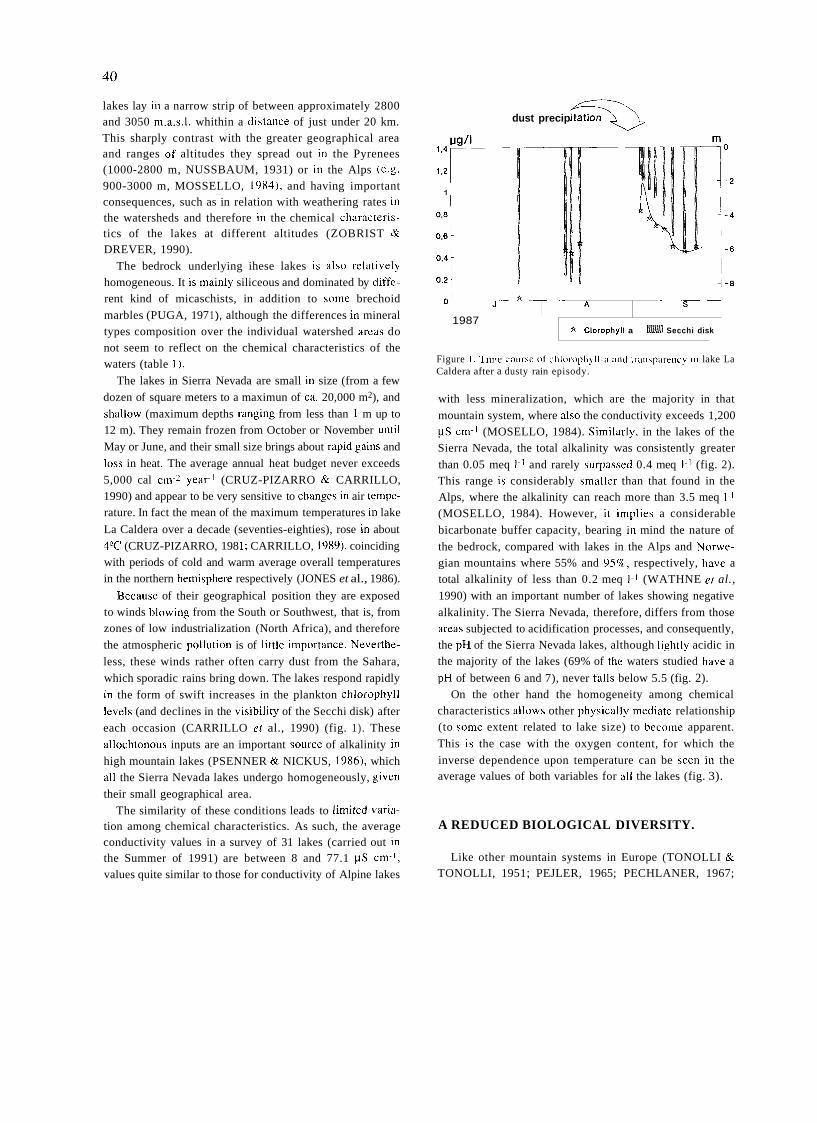
lakes lay in a narrow strip of between approximately 2800 and 3050 m.a.s.1. whithin a distance of just under 20 km. This sharply contrast with the greater geographical area and ranges of altitudes they spread out in the Pyrenees (1000-2800 m, NUSSBAUM, 1931) or in the Alps (e.g. 900-3000 m, MOSSELLO, 1984), and having important consequences, such as in relation with weathering rates in the watersheds and therefore in the chemical characteris- tics of the lakes at different altitudes (ZOBRIST &
DREVER, 1990).
The bedrock underlying ihese lakes is also relatively homogeneous. It is mainly siliceous and dominated by diffe- rent kind of micaschists, in addition to some brechoid
marbles (PUGA, 197 1 ), although the differences in mineral types composition over the individual watershed areas do not seem to reflect on the chemical characteristics of the waters (table 1).
The lakes in Sierra Nevada are small in size (from a few
dozen of square meters to a maximun of ea. 20,000 m2), and
shallow (maximum depths ranging from less than 1 m up to 12 m). They remain frozen from October or November until
May or June, and their small size brings about rapid gains and
loss in heat. The average annual heat budget never exceeds
5,000 cal cm-' year-1 (CRUZ-PIZARRO & CARRILLO, 1990) and appear to be very sensitive to changes in air tempe- rature. In fact the mean of the maximum temperatures in lake
La Caldera over a decade (seventies-eighties), rose in about
4" (CRUZ-PIZARRO, 198 1 ; CARRILLO, 1989), coinciding with periods of cold and warm average overall temperatures in the northern hemispliere respectively (JONES et al., 1986).
Because of their geographical position they are exposed to winds blowing from the South or Southwest, that is, from zones of low industrialization (North Africa), and therefore
the atmospheric pollution is of little importance. Neverthe-
less, these winds rather often carry dust from the Sahara,
which sporadic rains bring down. The lakes respond rapidly
in the form of swift increases in the plankton chlorophyll
levels (and declines in the visibility of the Secchi disk) after
each occasion (CARRILLO et al., 1990) (fig. 1) . These
allochtonous inputs are an important source of alkalinity in
high mountain lakes (PSENNER & NICKUS, 1986), which al1 the Sierra Nevada lakes undergo homogeneously, given
their small geographical area.
The similarity of these conditions leads to lirnited varia- tion among chemical characteristics. As such, the average conductivity values in a survey of 31 lakes (carried out in the Summer of 1991) are between 8 and 77.1 yS cm-l, values quite similar to those for conductivity of Alpine lakes
dust precipitation T
o'----- - A d 1987 * Clorophyll a m Secchi disk
Figure l. Tirne course ol' chloropliyll-a ai>d ti-aii\pa~.eiicy iii lake La Caldera after a dusty rain episody.
with less mineralization, which are the majority in that
mountain system, where also the conductivity exceeds 1,200 yS cm-' (MOSELLO, 1984). Similarly, in the lakes of the Sierra Nevada, the total alkalinity was consistently greater
than 0.05 meq 1- ' and rarely surpassed 0.4 meq 1-l (fig. 2). This range is considerably smaller than that found in the Alps, where the alkalinity can reach more than 3.5 meq 1- l
(MOSELLO, 1984). However, it implies a considerable
bicarbonate buffer capacity, bearing in mind the nature of
the bedrock, compared with lakes in the Alps and Norwe- gian mountains where 55% and 95%, respectively, have a total alkalinity of less than 0.2 meq 1-1 (WATHNE et al., 1990) with an important number of lakes showing negative
alkalinity. The Sierra Nevada, therefore, differs from those areas subjected to acidification processes, and consequently, the pH of the Sierra Nevada lakes, although lightly acidic in the majority of the lakes (69% of the waters studied have a
pH of between 6 and 7), never falls below 5.5 (fig. 2). On the other hand the homogeneity among chemical
characteristics allows other physically mediate relationship (to some extent related to lake size) to become apparent.
This is the case with the oxygen content, for which the
inverse dependence upon temperature can be seen in the average values of both variables for al1 the lakes (fig. 3).
A REDUCED BIOLOGICAL DIVERSITY.
Like other mountain systems in Europe (TONOLLI &
TONOLLI, 195 1 ; PEJLER, 1965; PECHLANER, 1967;
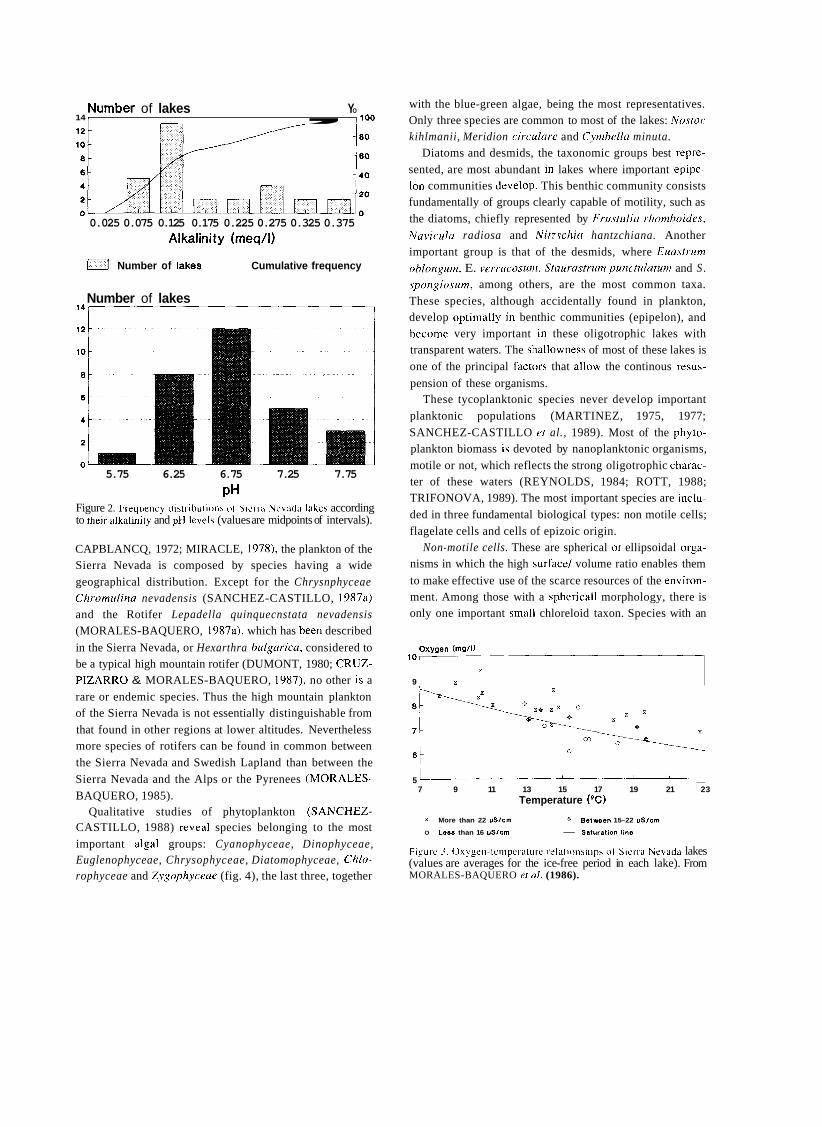
14 1 Number of lakes YO , 1100
0.025 0.075 0.125 0.175 0.225 0.275 0.325 0.375
Alkalinity (meq/l)
Number of lakes -- Cumulative frequency
Number of lakes
" 5.75 6.25 6.75 7.25 7.75
PH Figure 2. Frequeticy disiribuiioii\ (11 Sicil-~i hrvada lahes according to their alkalinity and pH levels (values are midpoints of intervals).
CAPBLANCQ, 1972; MIRACLE, 1978), the plankton of the Sierra Nevada is composed by species having a wide geographical distribution. Except for the Chrysnphyceae Chromulina nevadensis (SANCHEZ-CASTILLO, 1987a) and the Rotifer Lepadella quinquecnstata nevadensis (MORALES-BAQUERO, 1987a), which has been described
in the Sierra Nevada, or Hexarthra bulgarica, considered to be a typical high mountain rotifer (DUMONT, 1980; CRUZ- PIZARRO & MORALES-BAQUERO, 1987), no other is a rare or endemic species. Thus the high mountain plankton of the Sierra Nevada is not essentially distinguishable from that found in other regions at lower altitudes. Nevertheless more species of rotifers can be found in common between the Sierra Nevada and Swedish Lapland than between the Sierra Nevada and the Alps or the Pyrenees (MORALES- BAQUERO, 1985).
Qualitative studies of phytoplankton (SANCHEZ- CASTILLO, 1988) reveal species belonging to the most important alga1 groups: Cyanophyceae, Dinophyceae, Euglenophyceae, Chrysophyceae, Diatomophyceae, Chlo- rophyceae and Zygoph.vceae (fig. 4), the last three, together
with the blue-green algae, being the most representatives. Only three species are common to most of the lakes: Nostoc kihlmanii, Meridion circulare and Cymhrlla minuta.
Diatoms and desmids, the taxonomic groups best repre- sented, are most abundant in lakes where important epipe- Ion communities develop. This benthic community consists fundamentally of groups clearly capable of motility, such as the diatoms, chiefly represented by Frzlstulia rhomhoides,
Naiticula radiosa and Nitzschia hantzchiana. Another important group is that of the desmids, where Euustrum oblongum, E. i9errucosum, Staurastrum punctulutum and S . spongiosum, among others, are the most common taxa. These species, although accidentally found in plankton, develop optimally in benthic communities (epipelon), and become very important in these oligotrophic lakes with transparent waters. The shallowness of most of these lakes is one of the principal factors that allow the continous resus-
pension of these organisms. These tycoplanktonic species never develop important
planktonic populations (MARTINEZ, 1975, 1977; SANCHEZ-CASTILLO et al., 1989). Most of the phyto- plankton biomass is devoted by nanoplanktonic organisms, motile or not, which reflects the strong oligotrophic charac- ter of these waters (REYNOLDS, 1984; ROTT, 1988; TRIFONOVA, 1989). The most important species are inclu- ded in three fundamental biological types: non motile cells; flagelate cells and cells of epizoic origin.
Non-motile cells. These are spherical os ellipsoidal orga- nisms in which the high surfacel volume ratio enables them
to make effective use of the scarce resources of the environ- ment. Among those with a sphericall morphology, there is only one important small chloreloid taxon. Species with an
Oxygen (mgll) loo
&'
9 H
5 J 7 9 11 13 15 17 19 21 23
Temperature
X More than 22 pSlcm 9 Between 15-22 uSlcm
O Leas than 16 ~ S l c m - Saturation line
Fig~~l-e 2 . O A ) ~ ~ I ~ - I ~ I ~ ~ ~ C ~ ~ I I L I I . C relaiio~ish~p~ 0 1 S I C ~ ~ ; I hevada lakes (values are averages for the ice-free period in each lake). From MORALES-BAQUERO et al. (1986).
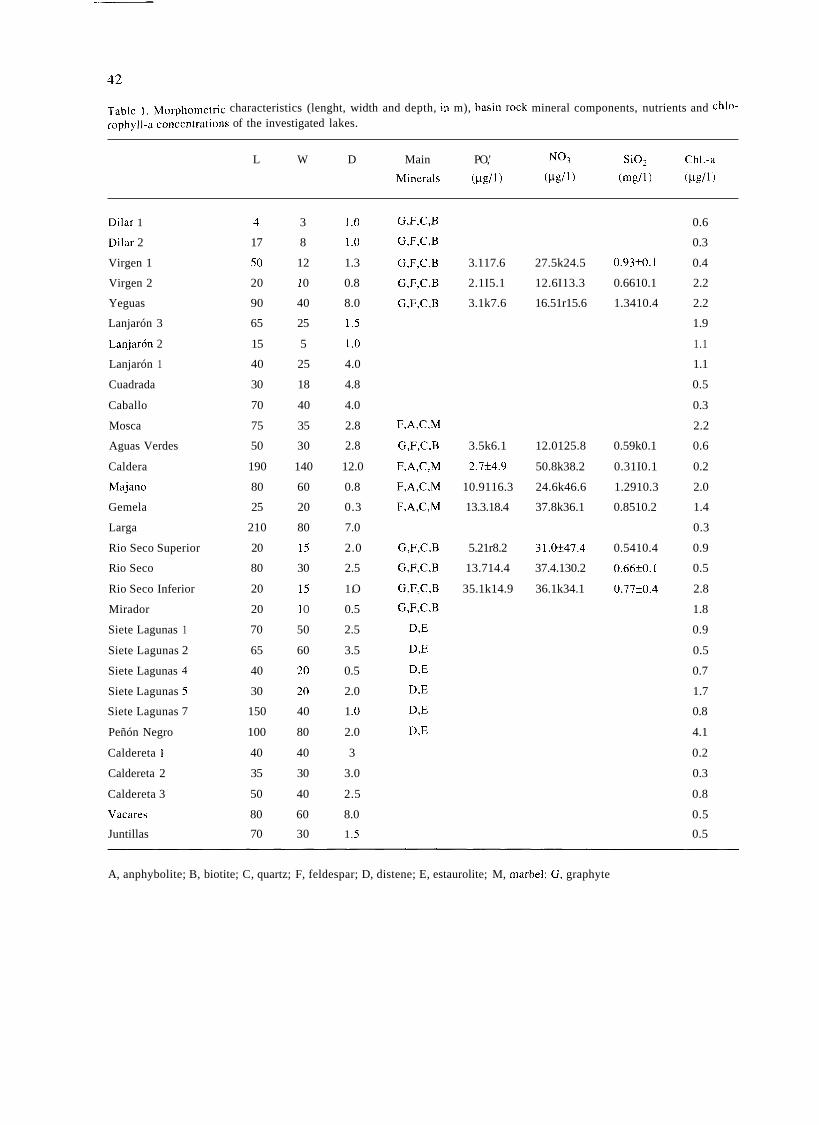
Table 1. Morphometric characteristics (lenght, width and depth, in m), basin rock mineral components, nutrients and chlo- rophy]]-a concentrations of the investigated lakes.
L W D Main PO,' NO, Si02 Chl.-a
Minerals (pg/ l ) (kg/ 1 ) (mg/l) ( ~ g / l )
Dilar 1 4 3 1 .O G,F,C,B 0.6
Dilar 2 17 8 1 .O G,F,C,B 0.3
Virgen 1 50 12 1.3 G,F,C,B 3.117.6 27.5k24.5 0.93k0.1 0.4
Virgen 2 20 1 0 0.8 G,F,C,B 2.1I5.1 12.6I13.3 0.6610.1 2.2
Yeguas 90 40 8.0 G,F,C,B 3.1k7.6 16.51r15.6 1.3410.4 2.2
Lanjarón 3 65 25 1 .S 1.9
Lanjarón 2 15 5 1 .O 1.1
Lanjarón 1 40 25 4.0 1.1
Cuadrada 30 18 4.8 0.5
Caballo 70 40 4.0 0.3
Mosca 75 3 5 2.8 F,A,C,M 2.2
Aguas Verdes 50 30 2.8 G,F,C,B 3.5k6.1 12.0125.8 0.59k0.1 0.6
Caldera 190 140 12.0 F,A,C,M 2.7I4.9 50.8k38.2 0.31I0.1 0.2
Majano 80 60 0.8 F,A,C,M 10.9116.3 24.6k46.6 1.2910.3 2.0
Gemela 25 20 0.3 F,A,C,M 13.3.18.4 37.8k36.1 0.8510.2 1.4
Larga 210 80 7.0 0.3
Rio Seco Superior 20 15 2.0 G,F,C,B 5.21r8.2 31.0k47.4 0.5410.4 0.9
Rio Seco 80 30 2.5 G,F,C,B 13.714.4 37.4.130.2 0.66I0.1 0.5
Rio Seco Inferior 20 15 1 .O G,F,C,B 35.1k14.9 36.1k34.1 0.77k0.4 2.8
Mirador 20 1 O 0.5 G,F,C,B 1.8
Siete Lagunas 1 70 50 2.5 D,E 0.9
Siete Lagunas 2 65 60 3.5 D,E 0.5
Siete Lagunas 4 40 20 0.5 D,E 0.7
Siete Lagunas 5 30 20 2.0 D,E 1.7
Siete Lagunas 7 150 40 1 .O D,E 0.8
Peñón Negro 100 80 2.0 D,E 4.1
Caldereta 1 40 40 3 0.2
Caldereta 2 35 30 3.0 0.3
Caldereta 3 50 40 2.5 0.8
Vacares 80 60 8.0 0.5
Juntillas 70 30 1 .S 0.5
A, anphybolite; B, biotite; C, quartz; F, feldespar; D, distene; E, estaurolite; M, marbel; G, graphyte
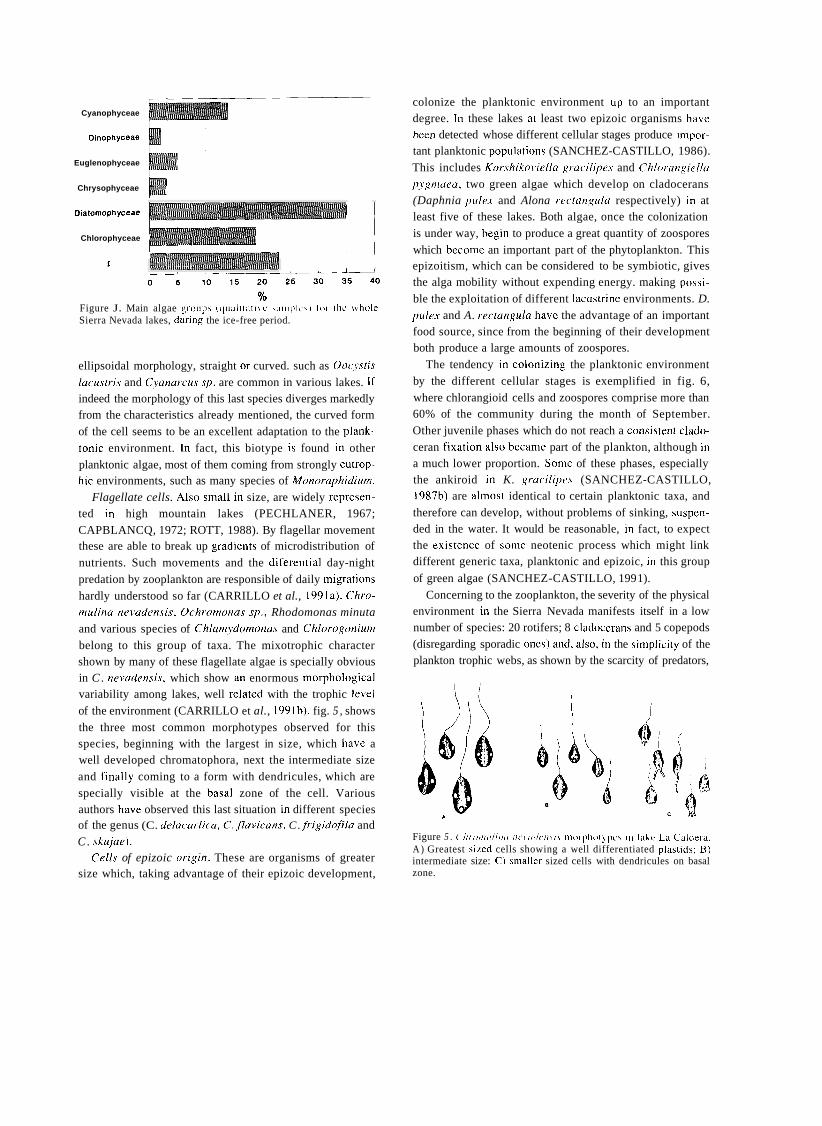
Cyanophyceae
Dinophyceae
Euglenophyceae
Chrysophyceae L Diatomophyceae
Chlorophyceae
Zygophyceae - - - - - - - -
'70 Figure J. Main algae gr«~ip\ (q~ialii;iii\c ~ . i i i i l ~ l c ~ i lo1 ilic \rli«le Sierra Nevada lakes, during the ice-free period.
ellipsoidal morphology, straight os curved. such as Oocystis
lacustr-is and Cyanar-cus sp. are common in various lakes. If indeed the morphology of this last species diverges markedly from the characteristics already mentioned, the curved form of the cell seems to be an excellent adaptation to the plank- tonic environment. In fact, this biotype is found in other planktonic algae, most of them coming from strongly eutrop- hic environments, such as many species of Monor-aphiclium.
Flagellate cells. Also small in size, are widely represen- ted in high mountain lakes (PECHLANER, 1967; CAPBLANCQ, 1972; ROTT, 1988). By flagellar movement these are able to break up gradients of microdistribution of nutrients. Such movements and the diferential day-night predation by zooplankton are responsible of daily migrations hardly understood so far (CARRILLO et al., 1991a). Chro- mulinu rrevaderzsis, Ochromonas sp., Rhodomonas minuta
and various species of Chlamydomonas and Chlorogonium
belong to this group of taxa. The mixotrophic character shown by many of these flagellate algae is specially obvious in C. neijadensis, which show an enormous morphological variability among lakes, well related with the trophic level of the environment (CARRILLO et al., 1991b). fig. 5 , shows the three most common morphotypes observed for this species, beginning with the largest in size, which have a well developed chromatophora, next the intermediate size and finally coming to a form with dendricules, which are specially visible at the basal zone of the cell. Various authors have observed this last situation in different species of the genus (C. delac~arlica, C.flavicans, C . fr-igidofila and C . skujae).
Cells of epizoic origin. These are organisms of greater size which, taking advantage of their epizoic development,
colonize the planktonic environment up to an important degree. In these lakes at least two epizoic organisms have been detected whose different cellular stages produce impor- tant planktonic populations (SANCHEZ-CASTILLO, 1986). This includes Kor-shikoviella gr-ac.ilipus and Chlor.crngiello
pjgniuea. two green algae which develop on cladocerans (Daphnia plcles and Alona r-e(.tangula respectively) in at least five of these lakes. Both algae, once the colonization is under way, begin to produce a great quantity of zoospores which become an important part of the phytoplankton. This epizoitism, which can be considered to be symbiotic, gives the alga mobility without expending energy. making possi- ble the exploitation of different lacustrine environments. D.
pulex and A. r-ectangula have the advantage of an important food source, since from the beginning of their development both produce a large amounts of zoospores.
The tendency in colonizing the planktonic environment by the different cellular stages is exemplified in fig. 6, where chlorangioid cells and zoospores comprise more than 60% of the community during the month of September. Other juvenile phases which do not reach a consistent clado- ceran fixation also became part of the plankton, although in a much lower proportion. Some of these phases, especially the ankiroid in K. gracilipes (SANCHEZ-CASTILLO, 1987b) are almost identical to certain planktonic taxa, and therefore can develop, without problems of sinking, susperi- ded in the water. It would be reasonable, in fact, to expect the existence of some neotenic process which might link different generic taxa, planktonic and epizoic, in this group
of green algae (SANCHEZ-CASTILLO, 199 1). Concerning to the zooplankton, the severity of the physical
environment in the Sierra Nevada manifests itself in a low number of species: 20 rotifers; 8 cladocerans and 5 copepods (disregarding sporadic ones) arid, also, in the simplicity of the plankton trophic webs, as shown by the scarcity of predators,
Figure 5 . C 111 O I I I I ~ ~ I I ~ C I I I C I III/(,II\I.\ 11io1 p l~o l ) ¡>e\ 111 l i ~ h e La Caldel-a, A ) Greatest s i ~ e d cells showing a well differentiated plastids; B j intermediate size: C j smaller sized cells with dendricules on basal zone.
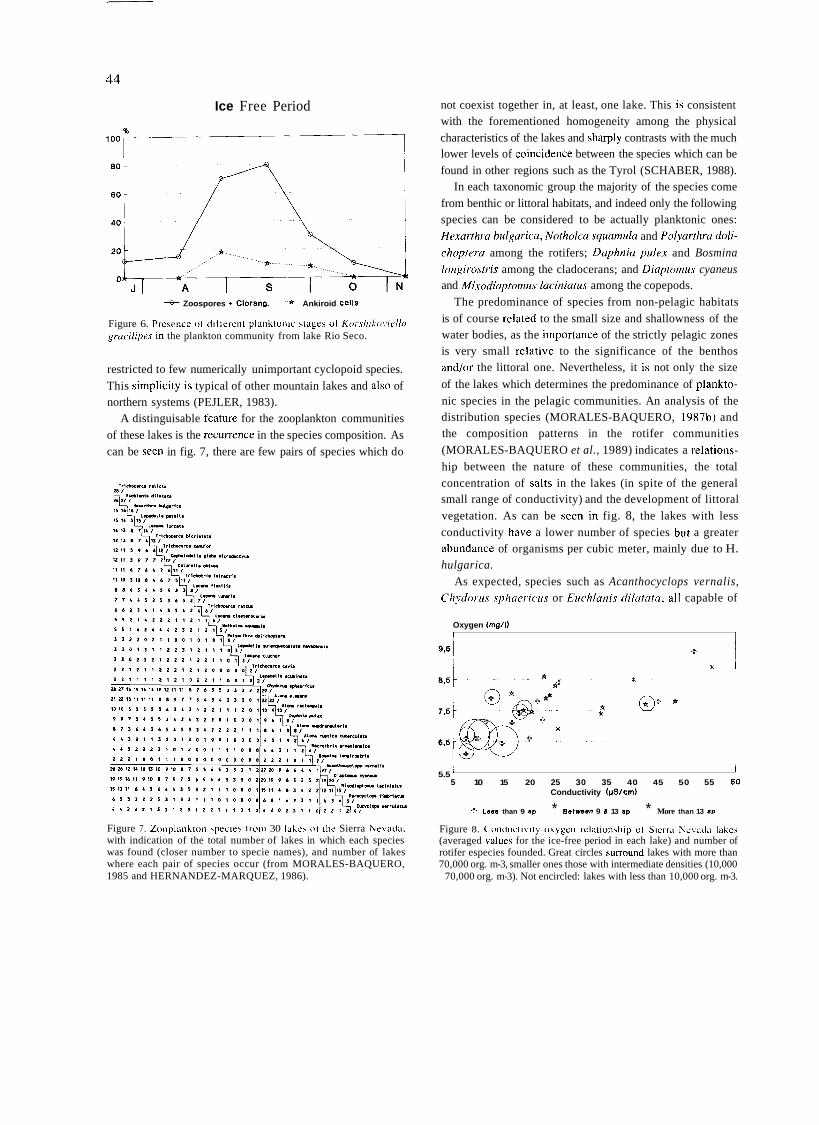
Ice Free Period
-3- Zoospores + Clorang. . * Ankiroid cells
Figure 6. Presence o1 dillercii~ plniiktoiiic atages ol Koi - . \ l~ iAo~~ i~~ / /a gracilipes in the plankton community from lake Rio Seco.
restricted to few numerically unimportant cyclopoid species. This simplicity is typical of other mountain lakes and also of northern systems (PEJLER, 1983).
A distinguisable feature for the zooplankton communities of these lakes is the recurrence in the species composition. As can be seen in fig. 7, there are few pairs of species which do
Figure 7. Zooplaiiktoii hpecieh iroiii 30 lahe\ ol tlie Sierra hevada, with indication of the total number of lakes in which each species was found (closer number to specie names), and number of lakes where each pair of species occur (from MORALES-BAQUERO, 1985 and HERNANDEZ-MARQUEZ, 1986).
not coexist together in, at least, one lake. This is consistent with the forementioned homogeneity among the physical characteristics of the lakes and sharply contrasts with the much lower levels of coincidence between the species which can be found in other regions such as the Tyrol (SCHABER, 1988).
In each taxonomic group the majority of the species come from benthic or littoral habitats, and indeed only the following species can be considered to be actually planktonic ones: Hexarthr,~ hulgauica, Notholca squamula and Polyarthra doli- choptera among the rotifers; Daphnia pule,r and Bosmina longirostris among the cladocerans; and Diaptomils cyaneus and Mi.xodiaptornus laciniatus among the copepods.
The predominance of species from non-pelagic habitats is of course related to the small size and shallowness of the water bodies, as the importance of the strictly pelagic zones is very small relative to the significance of the benthos and/or the littoral one. Nevertheless, it is not only the size
of the lakes which determines the predominance of plankto- nic species in the pelagic communities. An analysis of the distribution species (MORALES-BAQUERO, 1987b) and the composition patterns in the rotifer communities (MORALES-BAQUERO et al., 1989) indicates a relations- hip between the nature of these communities, the total concentration of salts in the lakes (in spite of the general small range of conductivity) and the development of littoral vegetation. As can be seen in fig. 8, the lakes with less conductivity have a lower number of species but a greater abundance of organisms per cubic meter, mainly due to H. hulgarica.
As expected, species such as Acanthocyclops vernalis, Chydorlrs sphriericus or Euchlanis dilutara, al1 capable of
Oxygen (rng/l)
l l
1
5.5 i
5 10 15 20 25 30 35 40 45 50 55 60 Conductivity íuS/cm)
+ Leas than 9 sp * Between 9 6 13 ap * More than 13 so
Figure 8. Coiiducticiiy-oxqgeii i-elatioiisliip o1 Sieii~i ;\ie\otla lahe!, (averaged values for the ice-free period in each lake) and number of rotifer especies founded. Great circles surround lakes with more than 70,000 org. m-3, smaller ones those with intermediate densities (10,000 - 70,000 org. m-3). Not encircled: lakes with less than 10,000 org. m-3.

exploiting both the pelagic and the littoral zones, are within those with the greater distribution pattern among lakes (in fact, occurring in more than 90 % of them), and offer an unique oportunity to analyze populations on well isolated and definite characteristics waters because of their compa- rable development squemes (only one abundance peak). In this way it has been possible to determine that the tempera- ture account for only a low percentage of the variability in the average size of the individuals from different E. dilatata populations (about 30%, MORALES-BAQUERO, 1989) and also that factors linked to the development of a popula- tion in a certain lake (food conditions, genetic isolation, etc.) play an importan1 role in determining an individual's size. Similarly the estimates of secundary production for this species in different lakes was quite variable (MORALES- BAQUERO et al., 1990); and, even more noticeable, the production rates (P/B for the growth season) covered prac- tically the entire range of values that have been described for al1 the rotifer species (fig. 9).
PIB mg dry wt (x 10E-4) 3 0 1 1 5
01 10 7 9 11 13 15 17 19 21 2 3 25
Temperature (9C)
+ PIB *. Individual weight
Figure 9. Individual mean weigth and growing season P/B ratio of Euchkznis dilatutu in relation wirh averaged ice-free period tempe- ratures of each lake.
Much less is known about the bacterial communities (CANTERAS & PEREZ, 1987) and on the benthic fauna of the lakes of the Sierra Nevada, for which data exist about the distribution of Quironomides (LAVILLE & VILCHEZ- QUERO, 1986), Coleoptera (SAINZ-CANTERO & ALBA- TERCEDOR, 199 1) and Nematodes (OCAÑA, in prep.) The benthos is therefore an environment that demands furt- her studies since its relative importance in the overall meta- bolism of these lakes could be very high. In addition, from a paleolimnological standpoint, the analysis of the sedi- ments promises interesting results after registering modifi- cations in the structure of communities associated with
global climatic changes to which these waters seem to be sensitive.
SUITABLE SITES.
In conclusion, from the point of view of the regional limnology, the high mountain lakes of the Sierra Nevada appear to be homogeneous waters easely characterizable and whith low levels of pollution. Therefore these lakes are useful both as a reference sites in analyzing acidification in Europe, and as sensors of global climatic changes. In anot- her sense, because of the narrow ranges of overall variation in lake characteristics are often covered by individual values, these lakes are ideals for comparative studies of processes determined by the specific parameters of each lake. This allows, among the other approaches, the study of not only factors which affect the distribution of species
occupying the lakes, but also of forces structuring their communities. Thus these waters, as subjects of research, can also generate interest on a purely theoretical aspect.
REFERENCES
ANDERSON, R. S., 1974. Crustacean plankton of 340 lakes and ponds in and near the National Parks of the Canadian Rocky Mountains. J. Fish. Res. Bd. Canada 31: 855-869.
CANTERAS, J.C. & L. PEREZ, 1987. The biochemical diversity of heterotrophic bacterial communities in high mountain lakes from Sierra Nevada. Acta Hydrobiol. 29(2): 137-147.
CAPBLANCQ, J., 1972. Phytoplankton et productivite primaire de quelques lacs d'altitude dans les Pyrenées. Ann. Limnol. 8(3): 23 1-32],
CARRILLO, P., 1989. Analisis de las interacciones trófi- cas en elplancton de un sistema oligotrófico. Ph. D. Thesis. Universidad de Granada.
CARRILLO, P., L. CRUZ-PIZARRO & R. MORALES- BAQUERO, 1990. Effects of unpredictable atmospheric allochthonous input on the light climate of an oligotrophic lake. Verh. Internat. Verein. Limnol. 24: 97-101.
CARRILLO, P., P. SANCHEZ-CASTILLO & L. CRUZ-PIZARRO, 199 1 a. Coincident zooplankton and phytoplankton diel migrations in a high mountain lake (La Caldera, Sierra Nevada, Spain). Arch. Hydrobiol. 122(1): 57-67.


SANCHEZ-CASTILLO, P., 1986. Estudio de las comu- nidades fitoplanctónicas de las lagunas de alta montaña de Sierra Nevada. Ph. D. Thesis. Universidad de Granada.
SANCHEZ-CASTILLO, P., 1987a. Un nuevo taxon del género Chromulina: C . nevadensis P.M. Sánchez sp. nov. CI-yptogamie-Algologie 8(4): 235-240.
SANCHEZ-CASTILLO, P., 1987b. Estudio del ciclo biológico de Kor-shikoviella gracilipes (Lambert)Silva (Chlorococcales, Chlorophyta). Phycologia, 26(4): 496-500.
SANCHEZ-CASTILLO P., 1988. Algas de las lagunas de alta montaña de Sierra Nevada (Granada, España). Acta Bot. Malacitana, 13: 21- 52.
SANCHEZ-CASTILLO, P., 199 1. Comentarios sobre la biología del fitoplancton. Alquihla, 19: 39-41.
SANCHEZ-CASTILLO, P. L. CRUZ-PIZARRO & P. CARRILLO, 1989. Caracterización del fitoplancton de las lagunas de alta montaña de Sierra Nevada (Granada, España) en relación con las características fisico-químicas del medio. Limnetica, 5: 37-50
SCHABER, P., 1988. Rotatorien und Crustaceen in Tiro- ler Hochgebirgseen. In: PSENNER, R., K. ARZET, A. BRUGGER, J. FRANZOI, F. HIESBERGER, W. HONSIG- ERLEN-BURG, F. HORNER, U. NICKUS, P. PFISTER, P. SCHABER & F. ZAPF; Versauerung von Hochgehirsseen in kristalline Einzugsgehieten Tirol und Karntens. Zustand,
47
Ursachen, Aushirkungen, Entwicklung. Wasserwirtsclzaft Wasservor-sorge, Forschungsarheiten. Bundesmin. f. Land- u. Forstwirtschaft, Wien: 209-258.
THOMASSON, K., 1956. Reflections on Artic and Alpine lakes. Oikos 7(1): 1 17- 143.
TONOLLI, L. & V. TONOLLI, 195 l . Osservazioni sulla biologia ed ecologia di 170 popolamenti zooplanctonici di laghi italiani di alta quota. Mem. 1st. Iral. Idrohiol. 653-136.
TRIFONOVA, I.S., 1989. Change in community struc- ture and productivity of phytoplankton as indicators of lake and reservoir eutrophication. Arch. Hydr-ohiol. Beih. 33: 363-371.
WATHNE, B.M., R. MOSELLO, A. HENNIKSEN & A. MARCHETTO, 1990. Comparison of chemical characteris- tics of mountain lakes in Norway and Italy. -1n: JOHAN- NESSEN, M., MOSELLO, R. & BARTH, H. (eds.): Acidi- fication processes in remote mountain lakes. Air Pollution Research Report 20. Commission of the European Commu- nities, Brussels, pp. 41-58.
ZOBRIST, J. & J.I. DREVER, 1990. Weathering proces-
ses in alpine watersheds sensitive to acidification. In: JOHANNESSEN, M., MOSELLO, R. & BARTH, H. (eds.): Acidification processes in remote mountain lakes. Air Pollu- tion Research Report 20. Commission of the European Communities, Brussels, pp. 149- 161.

THE SANABRIA LAKE. THE LARGEST NATURAL FRESHWATER LAKE IN SPAIN
J. C. vega1, C. de ~ 0 ~ 0 s ~ and J.J. ~ lda so ro ' 1. Calle Lago n V 0 , 49300 Puebla de Sanabria. Zamora. Spain. 2. Department of Ecology, Faculty of Biology, University of Salamanca. 37008 Salamanca. Spain
Keywords: Lake, Monomixis, Glaciarism, Oligotrophy, Cyanophyceae.
ABSTRACT
The Lake of Sanabria, at an altitude of 1000 m. above sea level, and a surface area of 3,187,500 m2 is of glacial origin. According to its physical and chemical factors this lake is oligotrophic, monomictic temperate, with a winter circulation at 8 T and thermal stratification in the summer. Its transparency is high (5-9 m., Secchi disk). The main nutrients are present at low concentrations, with a vertical distribution that is nearly homogeneous during winter circulation and with an orthograde trend during thermal stratification.
Its nutrient levels are not very closely related to greater abundances of phytoplanktonic algae. Chlorophyll levels are gene- rally low ( mean 2,2 pgr.1-'). Chroococcal Cyanophyceae and Chlorococcal Chlorophyceae are the best represented groups in the phytoplankton, specially during chlorophyll peaks. Other groups, such as Diatoms, Chrisophyceae and Criptophyceae are
scarce. Macrophytes are represented along the poorest ground shores by Isoetes i.,elata, M)iriophyllum alter-niflor-um and other species,
and in the richest ones by Equisetum fluviatile and Eleochur-is palinstris. Zooplankton are represented by 12 main Rotifer and Crustacean species that follow a stable pattern of succession. Their
composition is characteristic of an oligotrophic lake.
INTRODUCTION
Located in the NW of Spain (42%7'30nN, 06"3'00"W) between the provinces of Leon and Zamora at 1000 m. above the sea level, the Sanabria lake lies in the Tera river valley which has its sources in the mountain range of the Sierra Segundera and the Sierra Cabrera.(fig. 1)
It is a glaciar lake, situated in a valley locked by a frontal moraine; Despite the possibility of some overexcavation at its bottom, it can be classified as type 30 according to the Hutchinson's classification (1967). Its only tributary and emissary is the Tera river. In the drainage basin there are two more rivers that join the Tera just before the lake (table 1 , fig. 1). Al1 these rivers run over an acid rock substrate (gneiss and granodiorites) of low solubility, and are very poor in salts.
The population resident in the drainage basin is very small, living in two villages, and shows negative demo- graphic and socio-economic trends. However, the popula- tion increases sharply in the summer because of the impor- tant recreational use of the lake.
Liinnetica, 8: 49-57 (1992) O Asociación Española d e Lirnnologia. Madrid. Spain
In the past. little attention has been dedicated to the lake by the scientific community. The most interesting work has been that of TABOADA (1912) who compiled the first bathymetric maps. In that same year, HALFBASS collected plankton samples that were determined by BACHMAN (1913). This worker determined 12 species of phytoplank- ton, one of them new. Later, Margalef also visited the zone in 1952 and carried out an extensive work on the lake's flora and fauna (MARGALEF, 1955). Currently, different studies are under way, as yet unpublished.
The aim of the present work is to describe novel results obtai- ned between 1986 and 1989. These data were obtained follo- wing a limnological control program on the Sanabria lake and are expected to offer a better view of the dynamics of the lake.
POPULATION AND WATER USE
The resident population in the drainage area is now about 200 people, living in two villages. Both of then discharge

their untreated sewage into the area. This discharge is of A little importance, except in summer, when it increases
sharply owing to the vacational tourist influx. The agricul- tural use of the land has considerably decreased over the last two decades and artificial fertilizers are not used.
Hidroelectrical reservoirs control the water supply by the
three rivers that flow into the lake. This helps to maintain the water flow in the dry summers.(fig. 1).
GEOLOGY AND GEOMORPHOLOGY
The Sanabria lake occupies a glacial depression in the Tera valley (fig. 1) and is dammed by a frontal moraine. The basin is situated between the mountains of the Sierra Segun- C dera and the Sierra Cabrera, characterized by their acid rocks of metamorphic or plutonic origin. These formations
are composed by orthose- rich gneisses and some granodio- rites. The low water solubility of these rocks leads to a very low ionic content in al1 the waters that run through the area and also those contained in the Sanabria lake.
During the Würm glacial period, two main glaciar arms
descended from the Sierra Segundera highplain along the Tera and the Segundera and Cardena valley ensemble. Both of them were joined, forming a large glaciar arm in the lower part of Tera valley where the Sanabria lake is now
located. This glaciar pushed morainic matter to the end of
its course, damming the valley. Thus, the water filled the basin after the melting of the ices some 8,000 years ago. The only present lake effluent is the Tera river, that later joins
the Esla (A tributary of the Duero river).
In the heights of the Sierra Segundera and Cabrera there are more than 20 ponds of glaciar origin and numerous bog pools.
CLIMATE AND VEGETATION
Climate is of the wet mediterranean mountain type with
an annual mean temperature of 10.3" and a total cumula- tive yearly rainfall mean of 1,400 mm. The coldest month is February when temperatures reach a daily mean of 2.4". The highest temperatures are recorded in July and August
when they rise to a mean of 19.8T. In the drainage area vegetation is abundant and is mainly
constituted, as expected in this climate, by young Que1.cu.s pyrenaica woodland. In the highplains of this area it is cons- tituted by shrubs, pastures and bogs.
sanabria Lake
u Re-h - Riwrs
Figure 1. Geographical fratui-e5 aiid bathymery of the Sanabria lake. A: Location; B: Drainage area; C: Bathyrnetry.
PHYSICAL FACTORS
Temperature: The vertical distribution of the tempera-
ture of Sanabria Lake over several seasonal cycles (1986- 1990) shows that it is a monomictic lake with winter circu-
lation and summer stratification. The winter circulation corresponds to a homeothermy between 7 and 4-C, depen- ding on the year (Figs. 2 and 3). The surface layers begin to warm at the start of the spring, reaching 20-24" during the
summer. A thermocline is detected from February to December in some years, but it usually begins to appear in March and in the first week of December it disappears. The depth of the epilimnion reaches about 10 meters during the
summer. Hypolimnion temperatures range between 7 and 9" in the same season.
The dynamics of oligotrophic monomictic lakes depend on the winter water circulation that ensures the renewal of nutrients in the photic 1ayer.It also leads to oxygenation of the deep waters of the lake. Climatic conditions affect the strength and duration of the winter mixing, thus achieving a higher oxygen leve1 in the waters in the colder winters (fig.2).
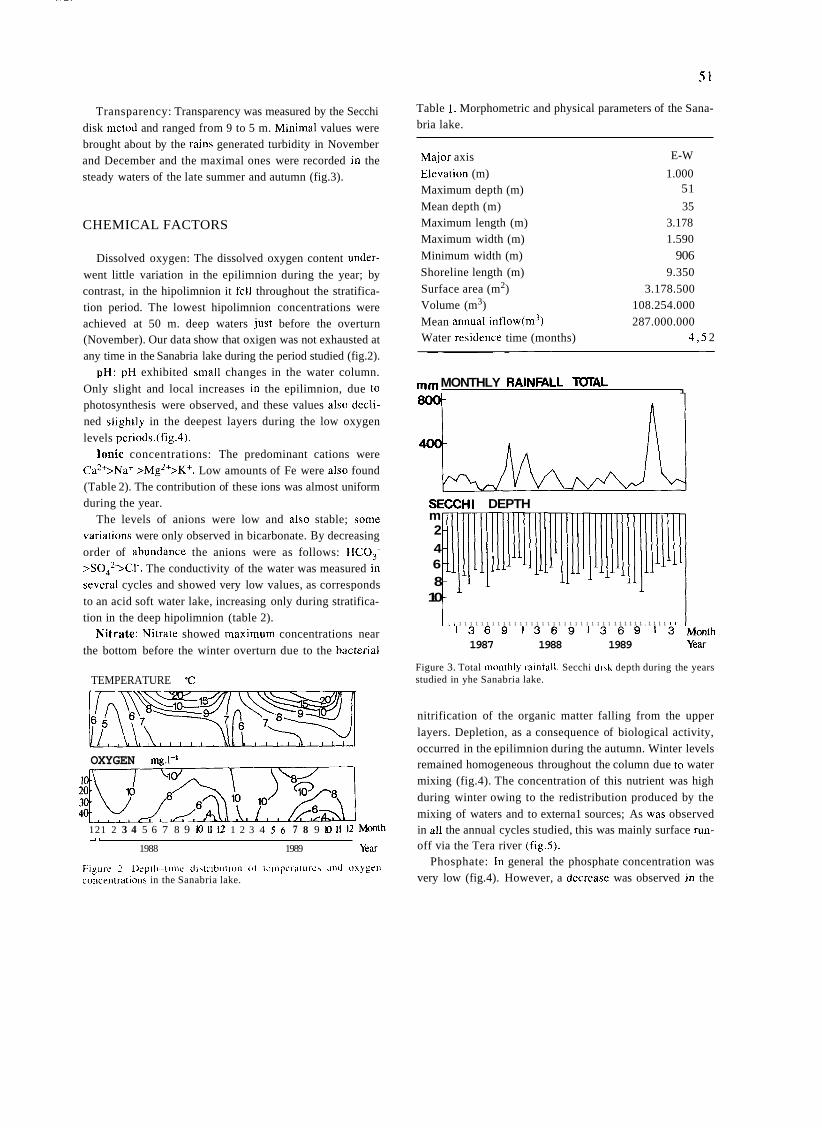
Transparency: Transparency was measured by the Secchi disk metod and ranged from 9 to 5 m. Minimal values were brought about by the rains generated turbidity in November and December and the maximal ones were recorded in the steady waters of the late summer and autumn (fig.3).
CHEMICAL FACTORS
Dissolved oxygen: The dissolved oxygen content under- went little variation in the epilimnion during the year; by contrast, in the hipolimnion it fe11 throughout the stratifica- tion period. The lowest hipolimnion concentrations were achieved at 50 m. deep waters just before the overturn (November). Our data show that oxigen was not exhausted at any time in the Sanabria lake during the period studied (fig.2).
pH: pH exhibited small changes in the water column. Only slight and local increases in the epilimnion, due to photosynthesis were observed, and these values also decli- ned slightly in the deepest layers during the low oxygen levels periods.(fig.4).
Ionic concentrations: The predominant cations were c a 2 + > ~ a + > M g 2 + > ~ + . Low amounts of Fe were also found (Table 2). The contribution of these ions was almost uniform during the year.
The levels of anions were low and also stable; some variations were only observed in bicarbonate. By decreasing order of abundance the anions were as follows: HC03- >SO,~>CI-. The conductivity of the water was measured in several cycles and showed very low values, as corresponds to an acid soft water lake, increasing only during stratifica- tion in the deep hipolimnion (table 2).
Nitrate: Nitrate showed maximum concentrations near the bottom before the winter overturn due to the bacteria1
TEMPERATURE %
OXYGEN mg.1-'
121 2 3 4 5 6 7 8 9 K)1112 1 2 3 4 5 6 7 8 9 101112Month 4 t
1988 1989 Year
Figure 2 . Dcpili-iiiiic tli\i~ibuiiuii ol tciiipei-ciiurc\ aild oxygeii concentrations in the Sanabria lake.
Table 1. Morphometric and physical parameters of the Sana- bria lake.
Major axis E-W
Elevation (m) 1.000 Maximum depth (m) 5 1
Mean depth (m) 3 5 Maximum length (m) 3.178 Maximum width (m) 1.590 Minimum width (m) 906 Shoreline length (m) 9.350 Surface area (m2) 3.178.500 Volume (m3) 108.254.000 Mean annual inflow(m3) 287.000.000 Water residente time (months) 4 , s 2
MONTHLY RAlNFALL m A L 1
SECCHI DEPTH m 2 4 6 8
10
1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 , 1 1 1 1 1 1 1
1 3 6 9 1 3 6 9 1 3 6 9 1 3 M o n t h 1987 1988 1989 Year
Figure 3. Total monthly rainfall. Secchi dihk depth during the years studied in yhe Sanabria lake.
nitrification of the organic matter falling from the upper layers. Depletion, as a consequence of biological activity, occurred in the epilimnion during the autumn. Winter levels remained homogeneous throughout the column due to water mixing (fig.4). The concentration of this nutrient was high during winter owing to the redistribution produced by the mixing of waters and to externa1 sources; As was observed in al1 the annual cycles studied, this was mainly surface run- off via the Tera river (fig.5).
Phosphate: In general the phosphate concentration was very low (fig.4). However, a decrease was observed in the
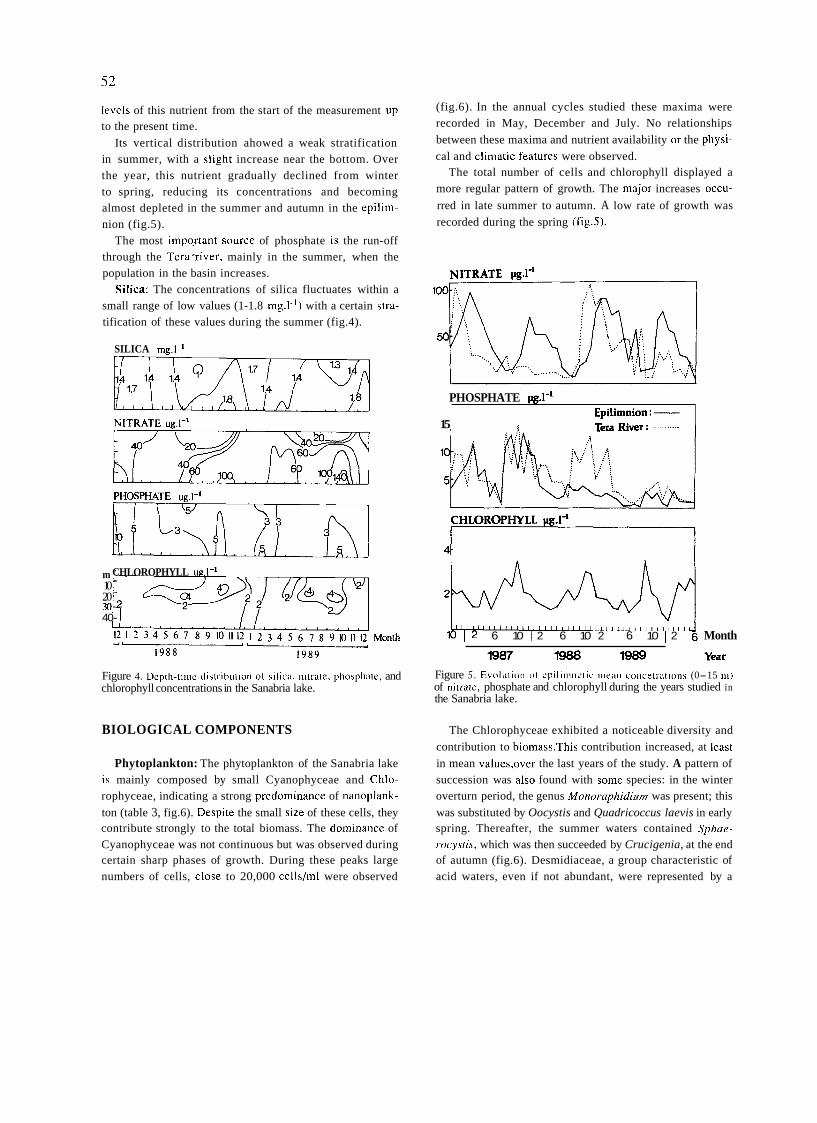
levels of this nutrient from the start of the measurement up to the present time.
Its vertical distribution ahowed a weak stratification in summer, with a slight increase near the bottom. Over the year, this nutrient gradually declined from winter to spring, reducing its concentrations and becoming almost depleted in the summer and autumn in the epilim- nion (fig.5).
The most impo.rtant source of phosphate is the run-off through the Tera-river, mainly in the summer, when the population in the basin increases.
Silica: The concentrations of silica fluctuates within a small range of low values (1-1.8 rng.1.') with a certain stra- tification of these values during the summer (fig.4).
SILICA rng.1-'
NITRATE ug.l-'
m - CHLOROPHYLL ug.1-' 10 - 20 - 302 40- 1
Figure 4. Depth-tiiiie distributioii of silicü, nitrate, phophate, and chlorophyll concentrations in the Sanabria lake.
BIOLOGICAL COMPONENTS
Phytoplankton: The phytoplankton of the Sanabria lake is mainly composed by small Cyanophyceae and Chlo- rophyceae, indicating a strong predominance of nanoplank-
ton (table 3, fig.6). Despite the small size of these cells, they contribute strongly to the total biomass. The dominance of Cyanophyceae was not continuous but was observed during certain sharp phases of growth. During these peaks large numbers of cells, close to 20,000 cells/ml were observed
(fig.6). In the annual cycles studied these maxima were recorded in May, December and July. No relationships
between these maxima and nutrient availability os the physi-
cal and climatic features were observed. The total number of cells and chlorophyll displayed a
more regular pattern of growth. The majos increases occu-
rred in late summer to autumn. A low rate of growth was
recorded during the spring (fig.5).
NITRATE pg.1-'
PHOSPHATE pg.1-' Epüirnnion :-
15 T~~~ w r : ......,....
I l l l l l l l l l l l l L I I I I I I I I I I I I I I I I I I , l l l l l l l l l l l
10 1 2 6 10 1 2 6 10 2 6 10 1 2 Month
1987 1988 1989 Year Figure 5 . Evolutioii ol epiliiii~it.tic ineaii coiiceu-atioiih (0-15 111)
of nitrate, phosphate and chlorophyll during the years studied in the Sanabria lake.
The Chlorophyceae exhibited a noticeable diversity and
contribution to biomass.This contribution increased, at least
in mean values,over the last years of the study. A pattern of succession was also found with some species: in the winter overturn period, the genus Monoraphidium was present; this
was substituted by Oocystis and Quadricoccus laevis in early spring. Thereafter, the summer waters contained Sphae-
rocystis, which was then succeeded by Crucigenia, at the end of autumn (fig.6). Desmidiaceae, a group characteristic of acid waters, even if not abundant, were represented by a
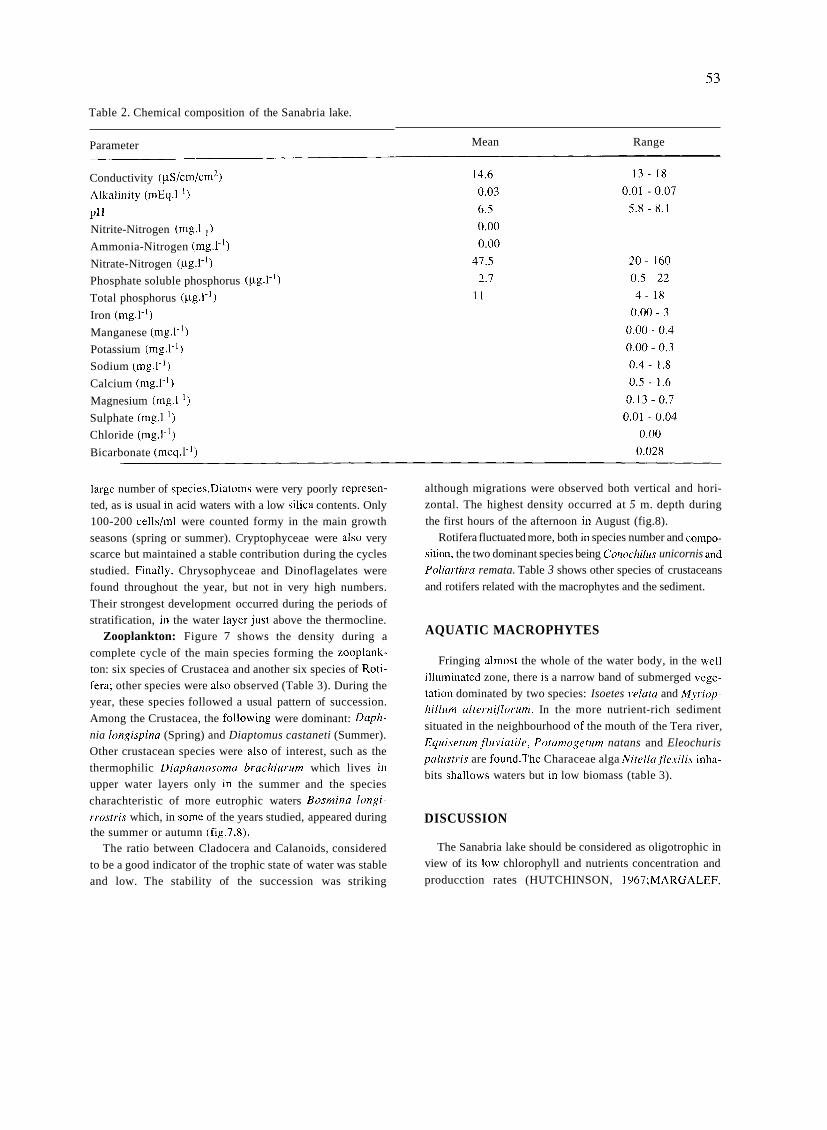
Table 2. Chemical composition of the Sanabria lake.
Parameter -- -
Conductivity ( y ~ / c r n / c m ~ )
Alkalinity ( m ~ ~ . l - ' )
PH Nitrite-Nitrogen (mg.1. ,)
Ammonia-Nitrogen (mg.1.')
Nitrate-Nitrogen (pg.1.')
Phosphate soluble phosphorus (yg.1.')
Total phosphorus (pg.l-')
Iron (mg.1.')
Manganese (rng.1.')
Potassium (mg.1.')
Sodium (mg.1-l)
Calcium (mg.1-')
Magnesium (mg.1 ')
Sulphate (mg.1.')
Chloride (mg.1.')
Bicarbonate (mq.1-l)
Mean Range
pp - -
large number of species.Diatoms were very poorly represen- ted, as is usual in acid waters with a low silica contents. Only 100-200 cells/ml were counted formy in the main growth
seasons (spring or summer). Cryptophyceae were also very scarce but maintained a stable contribution during the cycles
studied. Finally, Chrysophyceae and Dinoflagelates were
found throughout the year, but not in very high numbers.
Their strongest development occurred during the periods of stratification, in the water layer just above the thermocline.
Zooplankton: Figure 7 shows the density during a
complete cycle of the main species forming the zooplank- ton: six species of Crustacea and another six species of Roti- fera; other species were also observed (Table 3). During the year, these species followed a usual pattern of succession.
Among the Crustacea, the following were dominant: Daph-
nia longispina (Spring) and Diaptomus castaneti (Summer). Other crustacean species were also of interest, such as the
thermophilic Diaphanosoma brachiurum which lives in upper water layers only in the summer and the species
charachteristic of more eutrophic waters Bosmina longi- rrostris which, in some of the years studied, appeared during the summer or autumn (fig.73).
The ratio between Cladocera and Calanoids, considered
to be a good indicator of the trophic state of water was stable and low. The stability of the succession was striking
although migrations were observed both vertical and hori- zontal. The highest density occurred at 5 m. depth during
the first hours of the afternoon in August (fig.8). Rotifera fluctuated more, both in species number and compo-
sition, the two dominant species being Conochilus unicornis and Poliarthra remata. Table 3 shows other species of crustaceans
and rotifers related with the macrophytes and the sediment.
AQUATIC MACROPHYTES
Fringing almost the whole of the water body, in the well
illuminated zone, there is a narrow band of submerged vege- tation dominated by two species: Isoetes i,elatu and Myriop- hillum ulrerniflorum. In the more nutrient-rich sediment
situated in the neighbourhood of the mouth of the Tera river,
Equisetum flui'iatile, Potamo,qeti~m natans and Eleochuris pu1ustri.s are found.The Characeae alga Nitella,flexilis inha- bits shallows waters but in low biomass (table 3).
DISCUSSION
The Sanabria lake should be considered as oligotrophic in view of its low chlorophyll and nutrients concentration and
producction rates (HUTCHINSON, 1967;MARGALEF,

5 4
~ ~ b l ~ 3, ~i~~ of the main Species of phytoplankton, zooplankton fish and macrophytes identified in the Sanabria lake.
ALGAE CRUSTACEA
Cyanophyceae Copepodu Aphanotece sp. Diapton~lts castaneti ssp.castaneti
Coelosphaerium sp. Tropocyclops prasinus
Microcystis sp. Cyclops strenuiis
Merismopedia g l a i ~ a Macrocyclops alhid~rs
Pseudoanahaena catenata Eiicyclops serrulatus
Chlorophyceae Cludocera Botryococcus hraunii Daphnia longispina
Monoraphidium skujae Diaphanosoma hrachyurilm
M. komar-kovae Bosmina longirrostris
M. contortum Ceriodaphnia quadrangula
Quadrigula sp. Chidorils sphaericus Elakatotrir gelatinosa Alona qltadrangularis Ankistrodesmus fitsifoi.mis Alonella nana
Kirchneriela contorta Euricercus lamellatus Oocystis submarina Acroper~ts haipae O. lacustris Macrothry.~ hii.suticornis Qiradric~occu.~ luevis ROTIFERA Sphaerocystis sp. Keratella cochlearis Cr~ccigenia quadrata K. yuadrafa C . tetrapedia Trichocera similis Pediastrum tefras T . chatroni Scenedesmus sp. Lecane perplesa Staurastrum arktiskon Po1yarrhi.a remata S . aculeatum P. v~llgaris S. gracile Asplanchrra priodonta Staurodesmus jacirlijeri~m Conochilus unicornis S. glahrum Collotheca pelagica S. laevispinum Ascomor:fa ovalis Cosmarium contractum Filinia longiseta
Bacillariophyceae Pleosoma hudsoni Cyklotella glomerata Hexarthra intermedia C . stelligera FISH Melosira distans Salmo rrutta M . gl-anulata Barh~ts hocagei Tahellaria fenestrata Cohiris calderoni T . flocculosa Leuciscus cephalus Gomphonema constrictitm MACROPHYTES Diatoma hiemale Potamogeton natans Ceratoneis arcus Eleocharis palustris
Crysophyceae Eyi*iset~lm fl~tviatile Ochromonas sp. Isoetes velata Bitrichia ochridana Nitella flesilis Pseudopedinella gallica Antinoria agrostidea Mallomonas sp. Baldellia ranunculoides M. akrokomos Callitriche stagnalis Dinohryum sp. C . hamulata D. divergens Ranirncirlus peltatus D. crenulatum Jlrncus fluitans
Ci.yptophyceae Myriophillum alterniflorum Rhodomonas minuta Criptomonas erosa C . oilata
Dinophyceae Peridinium inconspicuum P . cinctum Gymnodinilfm sp Ceratium hirundinella
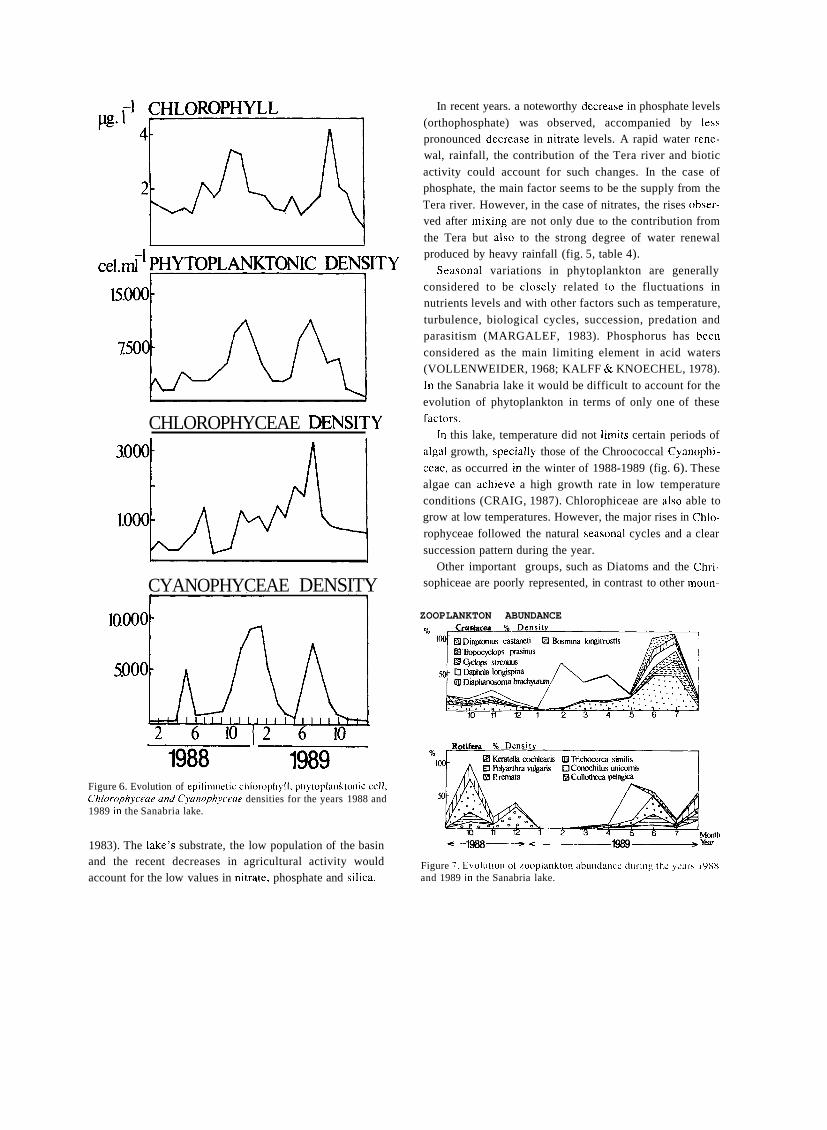
In recent years. a noteworthy decrease in phosphate levels (orthophosphate) was observed, accompanied by less
CHLOROPHYCEAE DENSITY
CYANOPHYCEAE DENSITY
Figure 6. Evolution of epilirntieiic cliloropliyll, pliyioplaiihronic cell, Chlorophycear und Cyonophy~,ear densities for the years 1988 and 1989 in the Sanabria lake.
1983). The lake's substrate, the low population of the basin and the recent decreases in agricultural activity would account for the low values in nitrate, phosphate and silica.
pronounced decrease in nitrate levels. A rapid water rene- wal, rainfall, the contribution of the Tera river and biotic activity could account for such changes. In the case of phosphate, the main factor seems to be the supply from the Tera river. However, in the case of nitrates, the rises obser- ved after mixing are not only due to the contribution from the Tera but also to the strong degree of water renewal produced by heavy rainfall (fig. 5, table 4).
Seasonal variations in phytoplankton are generally considered to be closely related to the fluctuations in nutrients levels and with other factors such as temperature, turbulence, biological cycles, succession, predation and parasitism (MARGALEF, 1983). Phosphorus has been considered as the main limiting element in acid waters (VOLLENWEIDER, 1968; KALFF & KNOECHEL, 1978). In the Sanabria lake it would be difficult to account for the evolution of phytoplankton in terms of only one of these factors.
In this lake, temperature did not lirnits certain periods of alga1 growth, specially those of the Chroococcal Cyanophi- ceae, as occurred in the winter of 1988-1989 (fig. 6). These algae can achjeve a high growth rate in low temperature conditions (CRAIG, 1987). Chlorophiceae are also able to grow at low temperatures. However, the major rises in Chlo- rophyceae followed the natural seasonal cycles and a clear succession pattern during the year.
Other important groups, such as Diatoms and the Chri- sophiceae are poorly represented, in contrast to other moun-
ZOOPLANKTON ABUNDANCE
I
Figure 7 . E\oluitoii o1 /ooplanktoii nbuiidatice dui-iiig the bears 1988 and 1989 in the Sanabria lake.
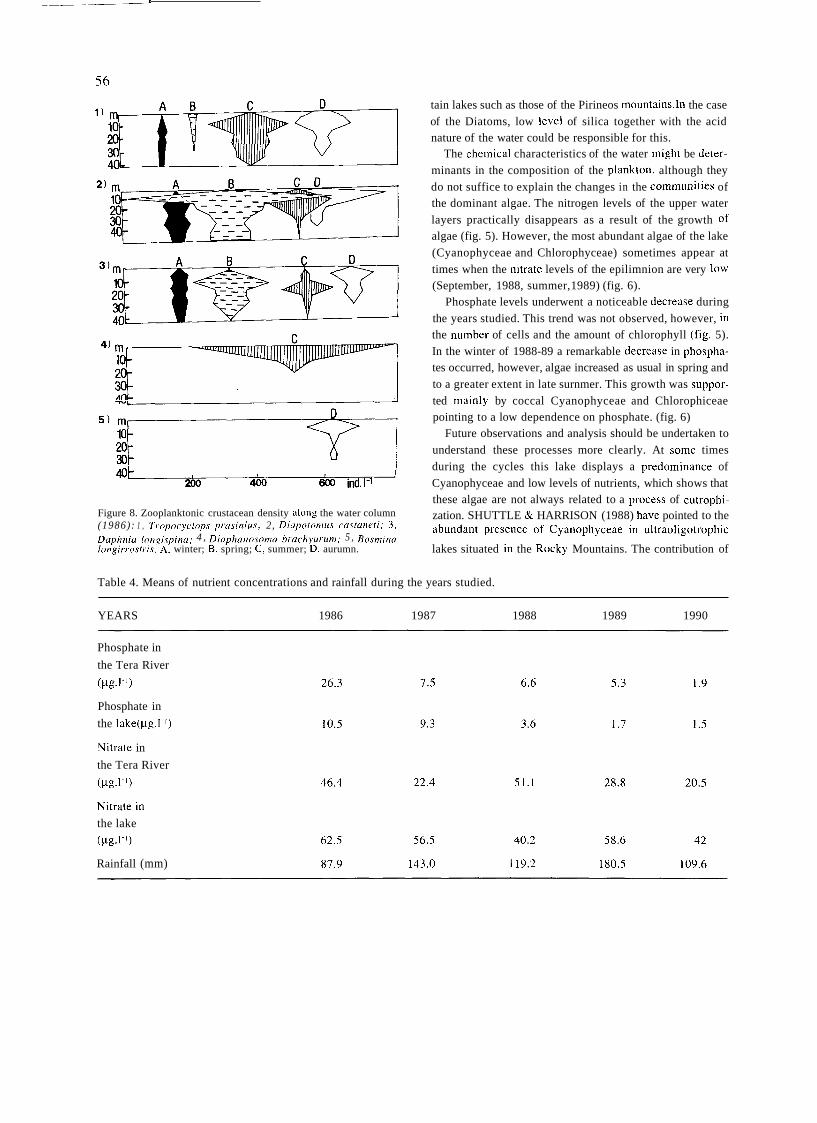
tain lakes such as those of the Pirineos mountains.In the case of the Diatoms, low leve1 of silica together with the acid nature of the water could be responsible for this.
The chemical characteristics of the water might be deter- minants in the composition of the plankton, although they - do not suffice to explain the changes in the communities of the dominant algae. The nitrogen levels of the upper water layers practically disappears as a result of the growth of algae (fig. 5). However, the most abundant algae of the lake (Cyanophyceae and Chlorophyceae) sometimes appear at times when the nitrate levels of the epilimnion are very low (September, 1988, summer, 1989) (fig. 6).
Phosphate levels underwent a noticeable decrease during the years studied. This trend was not observed, however, in the number of cells and the amount of chlorophyll (fig. 5). In the winter of 1988-89 a remarkable decrease in phospha- tes occurred, however, algae increased as usual in spring and to a greater extent in late surnmer. This growth was suppor- ted mainly by coccal Cyanophyceae and Chlorophiceae pointing to a low dependence on phosphate. (fig. 6)
Future observations and analysis should be undertaken to understand these processes more clearly. At some times during the cycles this lake displays a predominance of Cyanophyceae and low levels of nutrients, which shows that these algae are not always related to a process of eutrophi-
Figure 8. Zooplanktonic crustacean density along the water column zation. SHUTTLE & HARRISON (1988) have pointed to the (1986): 1 , Tropocyclnps pi.asinius; 2 , Diopntomus castaneti; 3 , Daphnja longjspjna; 4 , Djaphanos»ma brachyurum; 5 , Bosmjna abundant presence of C ~ a n o ~ h ~ c e a e in ultraO1igOtrO~hic longirrostris. A. winter; B, spring; C, summer; D, aurumn. lakes situated in the Rocky Mountains. The contribution of
Table 4. Means of nutrient concentrations and rainfall during the years studied.
YEARS 1986 1987 1988 1989 1990
Phosphate in
the Tera River
(pg.1-I)
Phosphate in
the lake(yg.1-')
Nitrate in
the Tera River
(pg.1-l)
Nitrate in
the lake
(yg.1-1)
Rainfall (mm)

the nanoplankton to the overall biomass of phytoplankton seems to be very important in oligotrophic lakes, despite the little attention that they have received (KALFF and KNOE- CHEL 1978). Their small size permits a better utilization of nutrients owing to the increase in the surface/volume ratio (KALFF & KNOECHEL, 1978; PAVONI, 1963; KALFF et al., 1975). Evidence exists to suggest that the Cyanophiceae accumulate phosphorus during the phases prior to growth peaks (SUTTLE & HARRISON, 1988). Other authors have speculated about their tendency to survive and thrive in waters with high contents in organic matter and the possibi- lity that this might be used by the algae (CRAIG, 1987; FOGG, 1973). In the Sanabria lake, high levels of organic matter could be accumulated, specially with the arrival of the autumn rains, which would contribute large amounts of particulate and non-particulate organic matter.
The Cyanophyceae are usually considered to be good
indicators of eutrophication; however this is not the case of the lake of Sanabria where their growth occurs at very low nutrient concentrations.In other cold oligotrophic and ultra- oligotrophic lakes, their importante has also been unders- cored. In the light of such data, we consider it important to bear in mind the complexity characterizing these oligotrop- hic lakes and that a review should be made of certain limno- logical concepts valid for more mineralized lakes but diffi- cult to implement for oligotrophic mountain lakes with
softer waters. The zooplankton follows normal trends, with stable
seasonal cycles, showing growth in spring of Daphniu longispina followed by two more thermophile species, Diap- tomus castaneti and Diaphanosoma hrachyurum. Finally, an autumn specie appears: Bosmina longirrostris (fig.7, 8).
The Rotifers are dominated by two species: Conochilus unicornis, in spring, and Polyarthl-a remata in autumn. These show more unstable cycles than the crustaceans (fig. 7).
The taxonomic composition of the zooplankton is consis- tent with that reported by several authors for oligotrophic lakes (TONOLLI, 1962; MC NAUGHT, 1975; EDMON-
SON, 1977).
ACKNOWLEDGEMENTS
Work supported by the Consejería de Medio Ambiente de la Junta de Castilla y Leon. Dirección del Parque Natural del Lago de Sanabria.
The authors wish to thanks to José María García Zorrilla for his important contribution to the study of Zooplankton,
and to the Director of the Parque Natural del Lago de Sana- bria, D. Guido Rodríguez de Lema Blanco for his uncondi- tional and continuous support in performing this study.
REFERENCES
BACHMAN,H. 1913: Plancktonproben aus Spanien, gesammet von Prof. Dr. Halbfas. Berichte des deustche Botanische Gesselbschaft 3 1: 183- 188.
CRAIG, S.R. 1987: The distribution and contribution of picoplancton to deep photosynthetic layers in some mero- mictic lakes. Acta Academiae Aboensis 47: 55-87.
EDMONSON, W.T. 1977: Lake Washington and the predic- tability of limnological events. Univ. of Upsala. August 21-27.
FOG, G.E. 1969: The Physiology of an alga1 nuisance. Proc. R. Soc. London Ser. B. 173: 175-189.
HALBFAS, W. 1913: Der Catanedasee, der groste Süswasersee Spaniens und seine Ungebung. Peterman Mitteilungen 59: 306-3 12.
HUTCHINSSON, G.H. 1967: A Treatise on Limnology. Willey. New York.
KALFF,J.K. & KNOECHEL, R. 1978: Phytoplankton and their dynamics in oligotrophic and eutrophic lakes. Ann. Rev.Eco1. Syst. 9: 475-495.
KALFF,J.K., KLING, H.J., HOLMGREN, S.H., & WELCH, H.E. 1975: Phytoplankton growth and biomass cycles.
MARGALEF, R. 1955: Comunidades dulceacuicolas del NW de España.Pub. Inst. Biol'. Apl. 21: 5-31.
MARGALEF, R. 1983:Limnologia. Omega. Barcelona. MAC NAUGHT, D.C. 1975: A hypothesis to explain the
succession from Calanoids to Cladocerans during eutrophi- cation. Verh.Internat. Verein. Limnol. 19: 724-731.
PAVONI, M. 1963: Die Bedentung des Nannoplanktons in Vergleich zum Netzplankton. Schweiz. Z. Hydrol. 25: 219-341.
SUTTLE, C.A. & HARRISON, P.J. 1988: Ammonium and Phosphate uptake rates N:P supply ratios and evidence for N and P limitation in some oligotrophic lakes. Limnol. Oceanogr.33: 186-202.
TABOADA, J. 1913: El Lago de San Martin de Casta- ñeda. Bol. Real Soc. Española Hist. Nat. 13: 960-986.
TONOLLI, V. 1961: Studio sulla dinamica dei popola- mento di un copepode (Eudiaptomus iulgaris, Scheneil) Mem. Inst. Ital.Idrobio1. 13:179-202.
VOLLENWEIDER, R.A. 1968: Scientific fundamentals of eutrophication of lakes and flowing waters with particu- lar references to nitrogen and phosphorus as factors in eutrophication.OCDE Directorate for scientific affairs. Paris.


BIOLOGICAL STUDIES OF SPANISH MEROMICTIC AND STRATIFIED KARSTIC LAKES
M. R. Miraclel, E. Vicente1 and C. Pedrós-Alió2
I Departments of Ecology and Microbiology, University of Valencia, Burjasot, Valencia, Spain. 2 Institut de Cikncies del Mar. CSIC, Barcelona. Spain.
Keywords: Karstic lakes, Meromictic, Stratification. Redoxcline, Oxycline, Anaerobic waters, Phytoplankton. Bacterioplankton, Ciliates, Zooplankton.
ABSTRACT
Surveys have been carried out in over 40 lakes in the Pre-Pyrenees, Cuenca mountains and Banyoles karstic areas in Spain.
About 18 of these lakes are deep enough to become stratified and have been studied in more detail. Here we discuss the verti- cal distribution and species composition of their planktonic communities, and their relationship to physico-chemical parameters. Meromictic and stratified karstic lakes are characterized by the large accumulations of organisms at the redoxcline. Sharp
gradients of oxygen and reduced compounds are found in this region as a consequence of the activities of the organisms. Diffe- rent organisms form abundance maxima at their preferred depths further contributing to the stratification. Special attention is given to these communities at the oxic-anoxic interface. Usually, rotifers accumulate in the microaerophilic zone. Algae may
be found at the zero oxygen depth. Phototrophic anaerobic bacteria are found underneath when enough light reaches the inter-
face. Different communities of ciliates appear in the aerobic waters, in the oxycline and in the anaerobic waters respectively.
INTRODUCTION
Karstic lakes originate by limestone and/or gypsum disso- lution. They have a peculiar morphometry, characterized by circular sinks with steep walls and a high relative depth (the
ratio of maximum depth to mean diameter, see table 1) .
These features, together with a high mineralization of the bottom waters due to their dissolution origins, favor the stra-
tification of the water layers. Permanent meromixis, howe-
ver, is a peculiar condition found only in a few lakes. Many karstic lakes with similar morphometry and substratum are not meromictic, such as those next to the locality of Cañada del Hoyo (Cuenca), where only Laguna de la Cruz is perma-
nently meromictic, out of a total of seven dissolution lakes.
Considering the whole karstic area in the mountains of Cuenca, where we have studied more than 35 dissolution
lakes and s~nall water bodies, only two (Laguna de la Cruz and Lake El Tobar) were found to be meromictic. Severa1 of these lakes can be seen in Plate l . Laguna de la Cruz presents a biogenic meromixis, due to enrichment of the monimolimnetic water in calcium, magnesium and iron bicarbonates. Lake El Tobar shows a crenogenic meromixis
where the monimolimnion is constituted mostly by a sodium
chloride brine (table 1). Another karstic area exists around Lake Banyoles (Girona). The main lake itself is a polje constituted by six main basins, but only one of them is
permanently meromictic. There are about 15 small lakes
surrounding the main lake and only one of them is mero-
mictic (Lake Vilar). Thus, we find a continuos spectrum of lakes with different degrees and schedules of stratification.
the most extreme and rase case being permanent meromixis. One of the most striking features of al1 these lakes is the
sharp stratification of organisms, which is determined by their own activity. This is common to both meromictic lakes and to those stratified during an important part of the year.
Actually, these stratified lakes were first studied for their
accumulation of sulfur photosynthetic bacteria at the oxic- anoxic boundary (ABELLA et a l . , 1980, 1981, 1985: GASOL et al.. 1990; GUERRERO & ABELLA, 1978: GUERRERO et al., 1978, 1980, 1985, 1987; MAS et a l . , 1990; PEDRÓS-ALIÓ et al., 1986; VICENTE & MIRA- CLE, 1984, 1988, 1991). Additional information can be found in GUERRERO & PEDRÓS-ALIÓ (1992). More general limnological studies were only carried out later on.
Limnetica, 8. 59-77 ( 1 992) O Asociación Española de Lirnnolopía, Madrid. Spain

Table 1. Representative values of several physico-chemical parameters in the different layers of Spanish karstic lakes during stratification. Lakes are arranged according to their mixing regime and to their sulfate content. Next to each lake, relative depth (zr), maximal depth (zmax), and geographical coordinates are provided. Abbreviations for geographical areas are: B, Barce- lona; CU, Cuenca; GI, Girona; HU, Huesca; L, Lleida; M, Mallorca; V, Valencia. Credit is given to appropriate references in the text and to C. AbellA and G. Moya personal communications.
Lake Z ( Z ) Conductivity El1 Alkalinity Sulfate Sulfide Phosphate Nitrate Ammonia mS cm-' mV meq 1-' mM mM pM PM PM
1. Meromictic lakes 1.1. Crenogenic
E1 Tobar (CU) 13 (19.5) 40°33'N/203'W Mixolimnion 0.6 +300
Oxic-anoxic interface 20 +270
Monimolimnion 200" +25
1.2. Biogenic 1.2.1. Low sulfate
Laguna de la Cruz (CU) 18 (24) 39°59'N/1052'W Mixolimnion 0.5 +375
Monimolimnion 1 .S +20
Montcortés (L)b 2.8 (30) 42"20'N/1 OO'E Mixolimnion 0.3 +200
Monimolimnion 0.3 ND
1.2.2. High sulfate Banyoles 111 (GI) 3 (30) 42"8'N/2"45'E
Mixolimnion 1.2 +275
Monimolimnion 2.2 -1 10
Vilar (Banyoles, GI) 2.3 (9) 42"8'N/2"45'E Mixolimnion 1.1 +130
Monimolimnion 1 .S - 175
2. Holomictic lakes 2.1. Low sulfate
Coromines (GI) 2.6 (5 ) 42"8'N/2"41'E Epilimnion 0.6 -80
Hypolimnion 0.8 -380
Negre 1 (GI) 6.5 (5.2) 42"8'N/2"41 'E Epilimnion 0.6 -100
Hypolimnion 0.8 -275
Laguna del Tejo (CU) 16 (32) 39"59'N/1°52'W Epilimnion 0.47 +150
Hypolimnion 0.5 1 -25
Lagunillo del Tejo (CU) 11 ( 1 1) 39"59'N/1 O52'W
Epilimnion 0.52 +300
Hypolimnion 0.64 +60
La Parra (CU) 11 (16) 39"59'N/I052'W Epilimnion 0.4 1 +300
Hypolimnion 0.32 +178
Table 1 (continuation)
Lake Z ( Z ) Conductivity Eh Alkalinity Sulfate
mS cm-' mV meq 1-1 mM Sulfide Phosphate
mM pM Nitrate Ammonia
PM FM
Las Cardenillas (CU) 9 (1 2) 39"59'N/1°52'W Epilimnion 0.4 +320 Hypolimnion 0.3 +90
Lagunillo de las Cardenillas 10 (6) 39"59'N/1°52'W
Epilimnion 1.28 +350 Hypolimnion 1.3 1 +350
2.2. High sulfate Cisó (Banyoles, GI) 9.2 (8) 42'8'N/2'45'E
Epilimnion 2.0 +130 Hypolimnion 1.6 -310
Nou (Banyoles, GI) 6.8 (7) 42'8'N/2'45'E
Epilimnion 1.8 +10 Hypolimnion 2.1 -225
Arcas-2 (CU) 28 (14.5) 39"59'N/2"8'W Epilimnion 2.4 +400 Hypolimnion 2.4 -100
Estanya (HU) 12.3 (20) 42"2'N/Oo32'E Epilimnion 1.9 +360 Hypolimnion 1 .S -35
3. Coastal lagoons Massona (GI)
Mixolimnion Monimolimnion
Remolar (B) Mixolimnion Monimolimnion
Cullera (V)C Mixolimnion Monimolimnion
Cibollar (M) Mixolimnion Monimolimnion
a The high conductivity is due to a sodium chloride brine. Typical chloride concentrations are 0.1, 500 and 6800 mM at the mixolimnion, interface and monimolimnion respectively.
b Meromictic in 1975-76, but mixed in 1979. Weromictic before a flood in 1982.
The syringes are filled simultaneously at the desired depth by by surface waves. The model shown in fig. 1B has 33 syringes the action of a vacuum pump connected to the device. A detai- in a one meter copper column, but other lengths and spacing led instantaneous image of the vertical distribution of the orga- among syringes can be used as desired. This systems works nisms can be obtained quickly and avoiding problems created only down to approximately 2 m before hydrostatic pressure

Non stretchable tape measure l n
Hose to -surface pump
Screwable connector
lnlet (1 cm high) -
Hose to surlace pump J Hose
20 rnl /*9 syringes
Copper tub 0.5 cm 0
Copper tube 2crn 0
Soldered copper cap
Figure 1 . Devices for sampling sharply stratified water columns. A) Modified double cone. B) Syringe system
fills the syringes spontaneously (at 3 to 4 m). For such depths,
air can be pumped into the system from the surface until it
reaches the desired depth. This may work, depending on the syringes used, down to 10 or 1.5 m. For better performance at
depth, however, a messenger triggered system such as that of BJ0RNSEN & NIELSEN (1991 ) is recommended.
The systems are lowered to the desired depth with outmost care not to disturb the fine stratification of organisms. A
good procedure consists of slowly lowering the device to the
desired depth, then very slowly and smoothly moving it side-
ways (syringes-side first in the case of the syringe system) to sample an undisturbed section of the water column.
In order to determine the exact depth of the peaks of
phototrophic organisms samples are taken at 5 cm intervals and filtered immediately through Whatman GFIC glass fiber filters. The color of the filters shows which is the dominant organism at each depth, and where the layers of algae and phototrophic bacteria start and end. Samples are then collec-
ted at the appropriate depths taking into account that accu- mulations of zooplankton are usually found a few cm above
the layers of algae and bacteria. Water from the pump can be diverted into a flow-through
chamber with electrodes for instantaneous determination of
oxygen, pH, and Eh (fig. 2). In al1 cases, samples have to be protected from direct sunlight and warm surface temperatures.
Since the bottom waters are generally anaerobic, samples must be distributed into appropriate bottles, tightly capped and impermeable to oxygen, and fixed immediately upon reaching the surface. When samples have to be fixed for counts of orga-
nisms, the fixative can be previously dispensed into appro- priate bottles and these are carefully filled (to avoid turbu-
lence) directly from the pump outlet. Care has to be taken to avoid spilling fixative. In the case of samples for reactive compounds such as oxygen or sulfide, and those which have to be incubated, the bottles are flushed with three times the bottle volume of water from the corresponding depth before filling

them to the brim and tightly capping them. In many cases, samples have to be obsemed immediately in vivo, to determine motility and general appearance OS the organisms.
Chemical measurements
a) Oxygen (Winkler method). To check for correct func- tioning of the oxygen electrodes, Winkler titrations should be done periodically. This should be done especially at the depths OS coexistence OS oxygen and sulfide. The modifica- tion OS INGVORSEN & J0RGENSEN (1 979) has to be used in order to avoid interferences from sulfide.
b) Sulfide. Sulfide can be measured in a number of ways. A titration with acid iodine can be performed after precipi- tating the sulfide with CdC12 or Zn acetate under alkaline conditions (GOLTERMAN et a l . , 1978). This method is extremely simple and has the advantage of using some of the same solutions as the Winkler titration, but it can only be used at relatively high sulfide concentrations. The methy- lene blue colorimetric method provides a good alternative, especially Sor low concentrations of sulfide. Samples can be immediately precipitated as in the previous case and analy- zed later in the laboratory (CLYNE, 1969; GOLTERMAN et al., 1978) or the reaction can be performed directly in the field and the color read later in the laboratory. Finally, samples with sulfide can be mixed in situ with a highly alka- line antioxidant buffer (to prevent both oxidation and escape of the gas) and later determined with a silver-sulfide elec- trode (BAUMANN, 1974).
C) Nutrients. Phosphate is measured by the molibdate blue method (STRICKLAND & PARSONS, 1972; GOLTER- MAN et al., 1978). Sulfide can interfere with this method and give artificially high values. Samples can be bubbled with N2 to purge them of sulfide before the analysis. Care, however, should be taken when interpreting results. For the remaining nutrients conventional methods can be used (for example those in STRICKLAND & PARSONS, 1972; GOLTERMAN et al., 1978). Samples for phosphate, nitrate, silicate, and DOC have to be filtered through Whatman GF/F filters (or similar filters) in situ, and biological acti- vity stopped immediately by addition of appropriate fixati- ves (chloroform, boric acid or mercuric chloride for phosp- hate, nitrate and DOC respectively). Samples for nitrite determinations must be fixed in sulfanilamide acid reagent for posterior analysis in the laboratory by the Giiess method (GOLTERMAN et al., 1978).
d) Iron compounds. Samples for iron determinations can be taken in BOD bottles to guarantee protection from
oxygen, and fixed with sulfuric acid (1% final concentra- tion). Ferrous iron is determined by the o-phenanthroline method. Total iron is determined by the same method after reduction of ferric to ferrous iron with ascorbic acid (GOLTERMAN et al . , 1978).
Enumeration of organisms and pigment quantification
The abundante of picoplankton organisms (bacteria and cyanobacteria) and OS small flagellates is determined by the acridine orange (either ZIMMERMANN & MEYER-REIL. 1974; or HOBBIE et al., 1977) or the DAPI epifluorescence methods (PORTER & FEIG, 1980), after fixation either with formalin or glutaraldehyde (2% final concentration buffered in Na-cacodylate, pH 6.8). The nanoplankton organisms
Temperature Screw probe
Flow-trhoug' charnber
Figure 2 . ¡-lo\&-iliiocigli c11:iiiibcr L I ~ C ~ 10 I I I ~ ; I \ L I I . C Eli. pH. oxygen and temperature itr s i r ~ r . Water from the conic sampler is directly pumpdd through the inlet and readings of the electrodes are recorded at the desired depth intervals. The Lipper portion of the drawing shows the lid from above.

(most phytoplankton and small ciliates) are usually fixed with Lugol and counted by the Utermohl technique (UTERMOHL, 1958) in a sedimentation chamber with an inverted micros-
cope. Many of these organisms can be counted in chambers
such as the Sedgwick-Rafter cell if their abundance is high enough or if a previous concentration step is performed. Roti-
fers and crustaceans usually have to be concentrated by filtra-
tion with nets of appropriate mesh size before counting them with an inverted microscope. These organisms are usually fixed with formalin to 4% final concentration.
Since many of the microorganisms at interfaces are
phototrophic, it is important to determine the abundance of
pigments. Chlorophylls, bacteriochlorophylls and carote- noids have taxonomic value and many times provide an easy-to-determine, synoptic view of the communities. Spec-
trophotometric determination of pigments is adequate for
routine purposes. The coincidente of several absorption maxima at the same wavelengths, however, creates a
problem when Chl a and Bchl c coexist. The chromatograp-
hic separation procedures are quite involved and time consu-
ming and, thus, HPLC becomes the method of choice when
characterizing a new system or performing detailed studies. We will summarize one spectrophotometric method and one
HPLC procedure we have used successfully. a) Spectrophotometric method. Known volumes from
each depth are filtered on small pore size glass fiber filters. Pigments are extracted by placing the filters in 5 m1 of 90%
acetone supersaturated with MgC03 in screw-capped tubes,
covered with aluminum foil to prevent degradation of
pigments through exposure to light. The extraction proceeds at 4" C, in the dark, overnight, after which the tubes are
centrifuged, part of the supernatant is carefully pipetted out
with a micropipette and placed in quartz cuvettes. Absorp-
tion spectra are determined in a spectrophotometer between 350 and 850 nm. Chl a concentration is calculated with the trichromatic formula of JEFFREY & HUMPHREY (1975)
with absorbance at 830 nm used to correct for background
absorption because of the presence of bacteriochlorophyll a , which absorbs at 775 nm in acetone extracts. Bacteriochlo- rophylls (BChl) are determined with the equations of
TAKAHASHI & ICHIMURA (1968), STANIER & SMITH
(1 960), and GLOE et al., (1975). b) HPLC method. Samples for HPLC analysis are
pumped directly on a filtering systerli with Whatman GF/F filters. The filters are then immediatelji frozen in dry ice and kept frozen until the analysis is carried out. The method of MANTOURA & LLEWELLYN (1983) can be used with minor modifications. HURLEY & WATRAS ( 1 99 1) and
KOHRTALS & STEENBERGEN (1985) have recently described HPLC protocols adequate for both algae and
phototrophic bacteria. The filters are ground with a teflon-
glass grinder in 90% acetone. Absorbance is measured at 440 nm , and fluorescente detection is used to aid in the identification of pigments. Peak areas are measured on the
absorbance trace. If a diode array is available, each peak can
be identified trough its absorption spectrum. For analysis, a 100-1 50 p1 mixture of 4: 1 pigment extract
in 90% acetone and an ion-pairing solution, is injected into
the chromatographic system. A gradient comprising some
steps is run: from 100% eluent A (a highly polar solvent) to 100% eluent B (a highly non polar solvent). Organic solvents must be HPLC grade, and has to be purified (for example with a Milli-Q systern, Millipore Corp.). Pigments are iden- tified by comparing their retention time against chromato-
graphic profiles of pure cultures of different microorganisms.
RESULTS AND DISCUSSION
The sysiems studied were the karstic lakes i11 Eastern Spain (see fig. 1 in GUERRERO & PEDRÓS-ALIÓ, 1992). The different water bodies have been classified according to
the mixing regime and the amount of dissolved sulfate in the water (table 1). We will examine the physicochemical stra- tification and the distribution of organisms in a few ecosys- tems chosen as representative of the different lake types in
table 1 .
Physico-chemical stratification
The most striking feature of these lakes is the existente of a multilayered planktonic community with a high abun- dance of organisms in a thin portion of the water column, coincident with the zonc of variation of the redox potential,
which may coiricide with a layer of oxygen and sulfide
coexistence (Lake Cisó in fig. 3) or may span a zone without either oxygen or sulfide (Laguna de la Cruz in fig. 3). The depth at which the redoxcline and the associated layers of organisms are found varies with the time ofthe year and the
vagaries of climate. In meromictic lakes the redoxcline coincides with thc
depth of meromixis when the mixolirnnion is not stratified. The depth of meromixis is characterized by an increase in temperature and conductivity, and only varies substantially under exceptional conditions (fig. 4). During summer ther- mal stratification, the redoxcline and thc associated micro-

Sulfide (mM) Oxygen (mgA) 0.00 0.25 050 0.75 100 -100 O 1W 200 3W 400 500
O 5 10 15 20 25 O 5 10 15 20 25
Conductiviiy (rnSIcrn) Temperature (OC) + Oxygen (mgll) 0
F I ~ U I C .:. \ C I . I I C , I I ~ I I \ I I I ~ ~ L I ~ I O I I 0 1 ~ ) l i ~ ~ i c o - c l i ~ i ~ ~ i c : ~ i l>~i~i~ii~cters in crenogenic meromictic Lake El Tobar (Auguat 1 1 , 199 l ) , biogenic meromictic, low sulfide, Laguna de la Cruz (August 27, 1987), and holomictic, high sulfide Lakes Cisó (June 1 1, 199 1) and Arcas-2 (August 28, 1987). Temperature (+), oxygen (O), conductivity (e) and Eh (O) have been plotted at the same scales in al1 panels except for Lake Cisó, where sulfide (O) is plotted instead of Eh, and where oxygen readings oH the scale have to be multiplied times 10 to obtain actual values. Only the upper three meters are shown in Lake Cisó to show the detail obtained with the syringe sampler used between 0.5 and 1.5 m. The cone sampler was used in the other lakes, where many data points have been omitted to clarify the drawings.
bial populations do not always coincide with the depth of
meromixis. In the more productive lakes studied, the redox-
cline rises as the season advances, and may reach the ther-
mocline towards the end of the summer. As the upper water
is progressively mixed down in the fall, the redoxcline is
pushed down with the thermocline until it reaches the depth
of meromixis again. Hardness of the water, which is an essential feature of
karstic lakea, often gives peculiar optical properties to the water. In Laguna de la Cruz, with alkalinities over 5 meq 1 - l ,
the blue-green water color turns Turkish blue and then
completely white in the last week of July or the first week of
August every year. This "whiting" is due to calcium carbo-
nate needle-shaped crystals, some forming aggregates. This
phenomenon occurs suddenly and persists only for a week.
Co-precipitation of phosphorous and decrease of light pene-
tration causes a reduction of phytoplankton abundance and
diversity. Secchi transparency during "whiting" in Laguna
de La Cruz is only half a meter. Usually after a week it reco-
vers back to the usual readings around 6 m. However,
because of calcium carbonate precipitation and decrease of
phytoplankton abundance, Secchi depth may reach 12 m
about two weeks after the "whiting" phenomenon.
The extinction coefficient by Iayers in Laguna de la Cruz
is high in the first meter layer due to its higher temperatu-
res, which favor C a c o 3 saturation, and to larger amounts
of particles. The extinction coefficient is low (mean of 0.2 m-l) at intermediate depths, and rises enormously at the
leve1 of the redoxcline, because of the high abundance of
microorganisms. In summer, light intensities from 1 to 10 yEinst. m-2 S - 1 (approximately 0.1 and 1 % of surface inci-
dent light respectively) reach these layers, where photosynt-
hetic activity is substantial and usually higher than in the
epilimnion.
Oxygen profiles in meromictic lakes are quite distinctive
(figs. 3 and 4). They are both extremely clinograde and posi-
tively heterograde. There is a permanently anoxic layer at
the bottom, segregated by a steep gradient. A very pronoun-
ced metalimnetic oxygen maximum is observed generally
during the spring, with supersaturated values reaching up to
200% (about 20 mg 1-', fig. 4). This oxygen maxima corres-
pond to maxima of photosynthesis by phytoplankton and,
thus, their exact depth changes with thermal stratification
and plankton activities. Oxygen is trapped in thc density
gradient at the thermocline, where diffusion is additionally
decreased by hydrostatic pressure and lack of turbulence. In
the epilimnion, gas solubility decreases with increasing
summer temperatures and diffuses to the atmosphere favo- red by turbulence. Thus the epilimnion has an homogeneous
oxygen concentration close to the saturation value.
At the depth where O2 disappears, a shift in redox poten-
tial takes place (fig. 3). The redoxcline, therefore, appears
slightly below the oxycline. Redox potential is lower at the
end of the summer. In sulfate-rich lakes such as Lakes Cisó
and Arcas-2 (fig. 3), high ratcs of sulfate reduction cause intense H2S release, and the redox potential is much more negative than in waters with low sulfate concentration. In such lakes there is usually a zone of oxygen and sulfide
coexistence.
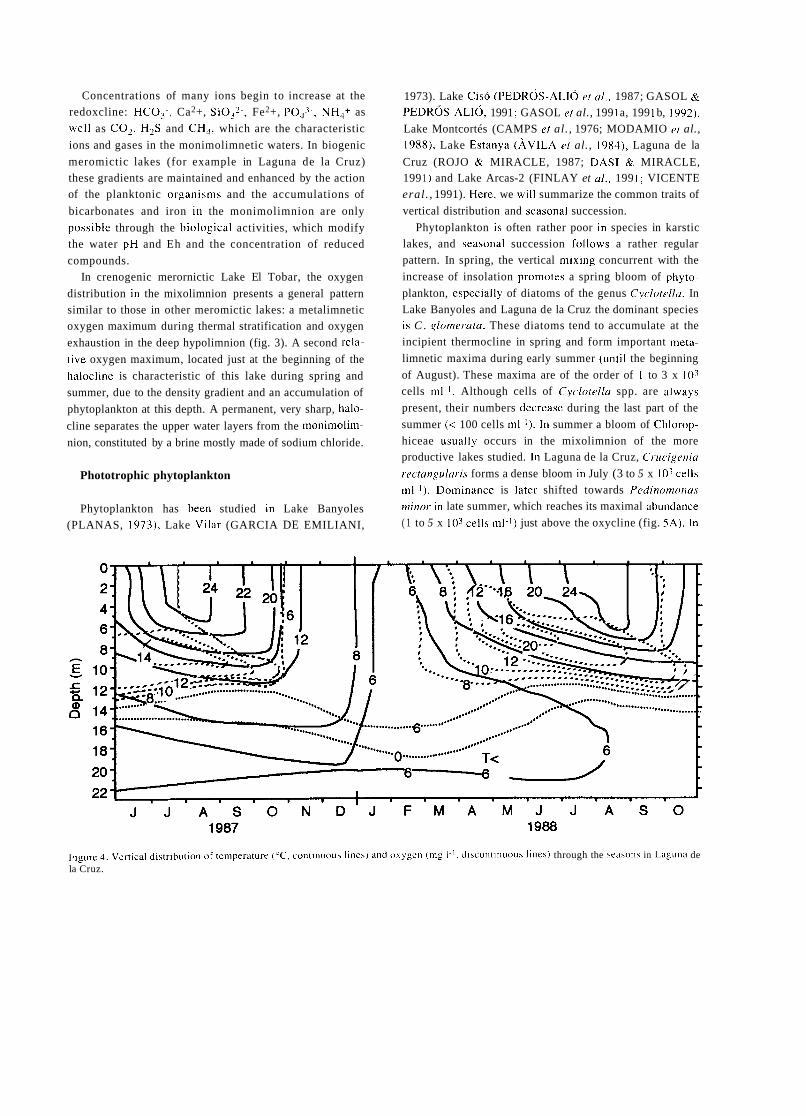
Concentrations of many ions begin to increase at the redoxcline: HC0,-, Ca2+, Si03'-, Fe2+, P043-, NH4+ as
well as CO,, H2S and CH,, which are the characteristic
ions and gases in the monimolimnetic waters. In biogenic
meromictic lakes (for example in Laguna de la Cruz) these gradients are maintained and enhanced by the action of the planktonic organisms and the accumulations of
bicarbonates and iron in the monimolimnion are only
possible through the biological activities, which modify the water pH and Eh and the concentration of reduced
compounds. In crenogenic merornictic Lake El Tobar, the oxygen
distribution in the mixolimnion presents a general pattern similar to those in other meromictic lakes: a metalimnetic oxygen maximum during thermal stratification and oxygen
exhaustion in the deep hypolimnion (fig. 3). A second rela-
tive oxygen maximum, located just at the beginning of the
halocline is characteristic of this lake during spring and
summer, due to the density gradient and an accumulation of
phytoplankton at this depth. A permanent, very sharp, halo-
cline separates the upper water layers from the monimolim-
nion, constituted by a brine mostly made of sodium chloride.
Phototrophic phytoplankton
Phytoplankton has been studied in Lake Banyoles (PLANAS, 1973), Lake Vilar (GARCIA DE EMILIANI,
1973). Lake Cisó (PEDRÓS-ALIÓ et al. , 1987; GASOL & PEDRÓS-ALIÓ, 1991 ; GASOL et al., 199 1 a, 199 1 b, 1992), Lake Montcortés (CAMPS et al., 1976; MODAMIO e f al.,
1988), Lake Estanya (AVILA et al., 1984), Laguna de la
Cruz (ROJO & MIRACLE, 1987; DAS1 & MIRACLE, 199 1 ) and Lake Arcas-2 (FINLAY et íil., 199 1 ; VICENTE eral. , 1991). Here, we will summarize the common traits of
vertical distribution and seasonal succession.
Phytoplankton is often rather poor in species in karstic lakes, and seasonal succession follows a rather regular pattern. In spring, the vertical mixing concurrent with the
increase of insolation promotes a spring bloom of phyto-
plankton, especially of diatoms of the genus Cyclotellu. In Lake Banyoles and Laguna de la Cruz the dominant species is C . glomei.ala. These diatoms tend to accumulate at the incipient thermocline in spring and form important meta-
limnetic maxima during early summer (until the beginning
of August). These maxima are of the order of 1 to 3 x 10' cells ml-l. Although cells of Cyclotella spp. are always
present, their numbers decrease during the last part of the
summer (< 100 cells ml-'). In summer a bloom of Chlorop- hiceae usually occurs in the mixolimnion of the more productive lakes studied. In Laguna de la Cruz, Cr-ucigenia
r-ec~tangu1ai~i.s forms a dense bloom in July (3 to 5 x 10ke l l s ml-1). Dominance is later shifted towards Pedinomoncis
mino]. in late summer, which reaches its maximal abundance
( 1 to 5 x 1 0 ~ e l l s ml-l) just above the oxycline (fig. SA). In
~i~~~~ 4, vertical distribution of tempei.üture (OC, contiiiuou\ lines) aritl oxygeii (iiig I k ' . rli\coritiriuous iines) through the sea\oiis in Lasuna de la Cruz.
Ternperature ("C)
O 10 20 30 O 800 1600 2400
Ternperature ("C)
O 10 20 30 O 2500 5000 O 250
o
2
4
.E. 6 5 Q
o" e
10
12
14 Polyarthra Filinia
l . , . ,
o 1 o 20
Oxygen (mg4
Figure 5. Vertical distribution of phytoplankton (A) and zooplankton (B) in Laguna de la Cruz on July 21, 1988. The three horizontal discon- tinuous lines indicate the oxygen concentrati&on isopleths of O, 1, and 2 mg l.', rcspectively. Temperature (continuous line) and oxygen (discon- tinuous line) profiles are also shown. A) Use the O to 2400 cells ml-' scale for striped kites and the Oto 600 cells ml-1 scale for the black kites. Crucigenia rectuiigularis and Rhodomonas lacustris are found throughout the mixolimnion. Peridinium, Pedinomonas minor, three species of Cryptomonas and the Chlorella-bearing ciliate Prorodon present deep maxima. Note that the maximal abundante of Cryptomonas and Proro- don is found at the depth of O mg 1.' oxygen concentration. B) Symbols are as in A). For Anuraeopsisfissa (striped kite) use the 0 to 5000 indi- vidual~ 1-' scale, and for the other rotifers (H. mira, K . quadruta, F. hofmanni, and Polyurthra sp., black kites) use the 0 to 500 individuals 1-1 scale. Note vertical separation of the two species of Anuraeopsis and the depth of the maxima of most rotifers around the 1 mg 1-1 oxygen concentration. (Data from DAS1 & MIRACLE, 1991 and ARMENGOL et al.. in press).

Lake Estanya the most important chlorophytes, are Planc- tonema lauterhorni (1 x 104 cells ml-l) and Chodatella quadrisetu (0,s x lo3 cells ml-l). In Lake Cisó, Crucigenia tetrapedia, Selenastrum capricornutum, Chlor-ella vulgaris and Hyaloruphidium contortum usually appear in the epilim- nion and later accumulate at the metalimnion.
Several species of Peridinium commonly appear in the
summer metalimnion. Some of these (P. inconspicuum) are more or less permanent. Peridinium species present relative maxima (around 100 cells ml-l) at the incipient spring ther- mocline and at the deep metalimnion in summer. Another dynoflagellate, Ceratium hirundinella, appears usually in the epilimnion during the summer.
Many of the algae tend to concentrate at the oxycline during stratification. ln Laguna de la Cruz, for example, total phytoplankton abundance is around 1000 cells ml-' in winter throughout the water column. Around 6000 cells ml-
can be found in the incipient thermocline in early spring and very low values through the rest of the vertical profile. During stratification, maxima appear at the oxycline with abundances between 5000 and 10000 cells ml-1 (fig. 5A). In meromictic Laguna de la Cruz, the community associated with the oxic/anoxic interface is mostly composed of three species of Cryptomonas: Cryptomonas erosa, C. phaseolus and C. ovohata, three of Trachellomonas, although only Trachellomonas hispida is common, three of Astasia and two of Euglena. The dominants are by far the species of Cryptomonas, which form rather dense and constant popu- lations during stratification (1 to 4 x 103 cells ml-1, fig. 5A). The peak of abundance is located at the depth of oxygen extinction which varies in Laguna de la Cruz between 18 m in winter and 15 to 13 m in summer (figs. 4 and 5A).
In Lake Cisó, more than 90% of the phytoplankton biomass is concentrated at the metalimnion during stratification (GASOL et al., 1992). In this lake the oxycline coincides with the thermocline. Some species grow both at the epi- and meta- limnion in early spnng and then disappear from the epilimnion in early summer, leaving residual metalimnetic peaks (GASOL & ~ D R Ó S - A L I Ó , 1991 ). Cryptomonas phaseolus, on the other hand, grows in the metalimnion and remains active at this depth throughout stratification, with abundances up to 105 cells ml-l. These development of oxyclinal popula- tion maxima of Ciyptomonas are a characteristic feature of most karstic lakes examined. Small chlorophyte flagellates such as Chlarrtyáomunas or Pedinomonas (fig. 5A) are commonly found above the layer of Cryptomonads.
In Lakes Montcortés (CAMPS et al., 1976), Estanya
(AVILA et al., 1984), Lagunillo del Tejo (VICENTE &
MIRACLE, 1984) and Arcas-:! (VICENTE et al.. 1991; FINLAY et al., 1991) Cryptomonas is found together with abundance maxima of Oscillatoriu. These Cryptomonas and Oscillatoriu populations must be adapted to low light condi- tions. Both have phycobilins as accessory pigments. These populations of C~.yptomonas seem to tolerate the presence of sulfide, at least during the night, while the cyanobacteria seem to resist sulfide for longer periods of time (VICENTE et al., 1991). In monomictic Lake Arcas-2, for instance, a dissolution lake on gypsum mar1 substrata, an anoxic hypo- limnion develops during summer stagnation, from March to October. The oxyclinal community is formed by a peak of Cllyptomonas, with abundances up to 17000 cells ml-1 in a
thin water layer less than half a meter thick. Abundant Osci- Ilatoria filaments (around 500 ml-') can be found together with the Cryptomonas layer. Layers of non filamentous cyanobacteria have been also described in the literature, such as Synechococi~us in several lakes (KOHRTALS & STEEN- BERGEN, 1985; CRAIG, 1987). In Spanish lakes, however, we have only found a maximum of the orange pigmented Gloeocapsa in Lake El Tobar, just above the halocline.
Oxyciinal peaks of flagellates are common in stratified lakes throughout the tyorld, for example Scourfieldia caeca in Tasmanian meromictic lakes (CROOME & TYLER, 1990; MIRACLE eral., 1991), Chi-ysosphaerella longispira in Canadian lakes (PIQK et al., 1984), or Cryptomonas in Finnish lakes (ARVOLA et al., 1991) and mountain lakes in central Europe (GERVAIS, 1991 ; ROTT, 1988), among many other examples. These oxyclinal communities differ from the mixolimnetic communities in several respeets. First, they are usually formed by only one or a few species. Second, they accumulate a tremendous amount of biomass. Third, they usually maintain a dynamic equilibrium throug- hout stratification, with low growth rates and reduced losses. Fourth, since predators seem inefficient at cropping this biomass, the carbon produced at the oxycline does not follow the traditional pathways to fishes and remains largely unexploited. Therefore, models of lake functioning, such as the PEG model of succession (SOMMER et al., 1986) or empirical models of phytoplankton abundance and produc- tion (DILLON & RIGLER, 1974) are bound to fail when applied to lakes with important oxyclinal communitiea.
Phototrophic bacterioplankton
The purple and green phototrophic sulfur bacteria are one of the best known features of meromictic and stratified lakes, because they build microstratified plates which are dense


day, but reaching high abundances in the whole vertical
profile during the night (up to 3 individuals I k l in Laguna de la Cruz and 8 in Lake Cisó). Chaohorus larvae are the only components of the benthic fauna over most of the sediment
surface.
a) Crustaceans. The most common crustacean community consists of two
cyclopoids: a small filter-feeder, generally Tropocyc1op.s
prasinus (in Lake Cisó Thermocyclops dyhowskii) and a
predator of the genus Cyc1op.s (most commonly Cyclops uhyssorum). These cyclopoids are generally present throug- hout the year with rather constant abundances. In the more
complex systems additional cyclopoids such as Dicr<yclop.\
hicuspidatus can be found. Diaptomids have been only found in three out of over 30
surveyed karstic permanent water bodies in the Pre-Pyre-
nean, Banyoles and Cuenca regions. Diaptomids, however,
are the most important crustacean components of temporary
karstic lakes. When present in a permanent lake, Diaptomids tend to be the dominant organisms in the epilimnion,
showing generally constant abundances through the year.
The most characteristic species is Arctodiuptomirs sc11inld.s (a
smaller and paler ecotype than that inhabiting saline lakes). The presence of this organism is in accordance with the high mineralization of the waters. A. salinus (a circumMediterra-
nean species) is found in Lake Banyoles and in Lake El
Tobar. The third lake with Diaptomids is the Pre-Pyrenean
Lake Basturs (Lleida), where Eudiaptomus padunus occurs,
(described from lakes of the Pre-Alpine region).
The most frequent cladoceran assemblage consists of
Duphnia longispina in autumn and spring and Diaphuno- soma hrachyurum in summer. During summer and autumn three smaller species are also commonly found: Ceriodaph-
nia (Ceriodaphnia quadrangula or C. reticulata), the more
littoral Bosmina longirostris and the neustonic Scapholehe- ris mucronutu.
b) Rotifers. The abundance and composition of the rotifer assemblage
changes markedly with time. Usually, around 20 species can be found coexisting in any one lake. Many of these constitute
couples of species belonging to the same genus, which replace each other in time os become segregated in the vertical profile.
Four distinct groups of species can be usually found in
summer (fig. 5B): ( 1 ) Epilimnetic species which are uniformly distributed through the epilimnion but are absent from the low oxygen hypolimnetic waters. Members of this group are Synchaeta pectinata, Asplanc.hna girodi, Hexarthra fennica and Polyurthra remata. (2) Species
present throughout the epilimnion, but showing a metalim-
netic maximum. This group includes He.var.thru n1ir.u. Kera- tellu c/~~udi.atu and Poljurthru i~~l1gur.i.s. (3) Species which are widespread during mixing, but concentrate in the
deepest microaerobic layers during the summer. Someti- mes, these species show a distribution with two peaks: one at the thermocline and another at the oxycline. Species within this group are Keratc~lla coc.hlearis, Kelli<~otia
longispina ( K . longisl,inn is only present in Lakes Mont-
cortés and Banyoles), Ariurcieopsis ,fis.sa and Polyar-thi.cr dolic,hopter-u. (4) Species which show a clear preference for the deep water layers. This group includes species which are exclusively found at the oxycline, such as Filiniu
hofmunrzi, Filinia longiscta lon,?isetu, and An~~raeops i s mirucleii (the latter only in Laguna de la Cruz, fig. 5B). Other members of this group are species found in the whole
vertical profile with an upper hypolimnetic maxima during
the spring, but limited to the oxycline during the summer,
such as Filiniu terminalis. The rotifer species which show a preference for the
oxycline are characteristically the same in most of the lakes st~idied. The genera Filiniu, PoI>~ur.thrrr, An~iruc~ol~.si.s and Kc~r.atellu are the more clearly associated with deep maxima (fig. 5B). Filinia and Anurueopsis may have a diet with a
large component consisting of bacteria and detritus. In Lake
Cisó, for example, A. fissa ha5 been experimentally shown to
feed on both phototrophic purple bacteria and chemotrophic
bacteria using fluorescently labeled bacteria as food. Polyul-thra, on the other hand, has been shown to feed on the
metalimnetic maximum of Crypton~onas phaseol~~s using radioactively labeled algae as food (MASSANA & PEDRÓS- ALIÓ, submitted). Pobarthra, thus, may preferentially feed on metalimnetic algae (Ctyptornonas) and the more polipha-
gous Krrcitella could select the smaller flagellates (Rhodo-
monas, chryso- and chloroflagellates) but also use detritus and bacteria. The photosynthetic bacteria, however, do not
seem to be a very important food source for rotifers, since the vertical distributions of predator and prey barely overlap.
Although the rotifers can grow under microaerophilic condi- tions. the phototrophic bacteria occupy the uppermost layer with sulfide, and this gas seems to prevent the presence of
metalimnetic rotifers. In Lake Cisó, the rotifer Mytilina
compressu has been found in the sulfide-rich hypolimnion (ALFONSO & MIRACLE, 1987), where it has been shown to feed on both purple phototrophic and chemotrophic bacte- ria (MASSANA & PEDRÓS-ALIÓ, unpublished).
The vertical distribution of pairs of congeneric species is fairly interesting and characteristic. Generally, there is
a stenoic species showing a very narrow and restricted occurrence at the oxycline, and a eurioic species. The latter may be differentiated into ecotypes and easily dominates over the .stenoic species when stratification conditions
change. The distribution of Filinia species has recently received considerable attention (RUTTNER-KOLISKO, 1989; HOF-MANN, 1982; SCHABER & SCHRIMPF, 1984). Careful taxonomic studies have revealed that the forms developing near the oxycline belong to a new species described as Filinia hofmanni (KOSTE, 1980). F .
hofmanni is almost exclusively found at the oxycline of lakes. In Lake Banyoles F. hofmanni was only found in the basins that become anoxic (mostly in meromictic basin 111). F. hofmanni has also been found in great numbers in Lake Arcas-2 and at lesser abundantes in Laguna de la Cruz, where it alternates dominance in the oxycline with A. miraclei and A. ,fissa in different years (fig. 5B). F. terminalis is more widely distributed, and presents oxycli- nal maxima slightly above those of F. hofmanni when both species co-occur.
The differential vertical distribution of Keratella species in stratified lakes is also well known (LARSSON, 1971; HOFMANN, 1985, 1987; RUTTNER-KOLISKO, 1975, 1977; MIKSCHI, 1989). Keratella hivmalis is frequently restricted to the oxycline layers, but it is not present in Spanish karstic lakes. Its role in the microaerophilic layers may be taken over by Keratella cochlearis (Lake Banyoles) or by Keratella quadrata (Lakes Montcortés, Cisó and Laguna de la Cruz). These species show a distribution with two peaks: one at the epi-metalimnion boundary and anot- her at the oxycline.
A similar case seems lo be true for Anurueopsis. A new species from Laguna de la Cruz, A. miruclei. has been recently described (KOSTE, 1991) as an exclusively oxycli- nal species (fig. 5B). This species coexists with the more widely distributed A. fissa, which can also present maxima at the oxycline (ARMENGOL et al., 1992). In Lake Banyo- les, where A. miraclei was not present, A. fissa showed a distribution with two peaks (MIRACLE, 1976; MIRACLE & ALFONSO, 1992). These different peaks could corres-
Figut-e h. Vertical cli\il~ibuiioii ol wiiie ciliates 111 Lahe Cisó (Junc 1 1 , 1991). The two hori~ontal discontinuous lines enclose the region of sulfide and oxygen coexistence. The double arrows labeled C and P indicate, respectively, the depths of the Cryptonionas phaseolus and purple phototrophic bacteria (Chromoti~irn minus and Arnoehohucter. M3) peaks of abundance. Note the four different scales for ciliates with diffe- rently shaded kites: Parumecium, Plu~iopylu and Cuenomorphu use the 0-3 ciliates ml-l scale; Stromhidium the 0-30 ciliates ml-l scale; Pi.oro- rlon, Coleps and Vorticella the 0-300 ciliates ml-' scale; and Cyclirii~~m the 0-3000 ciliates m l ' scale. (Unpublished data of R. Massana).

pond to different ecotypes within the same species. In stra- tified lakes, a subdivision of rotifer populations of a genus
into species and probably of a species into ecotypes clearly occurs. This differentiation matches the vertical environ- mental heterogeneity.
C) Ciliates. Ciliate distribution has been studied in Lakes Cisó (fig.
6) and Arcas-2. The ciliates of karstic lakes can be divided into three separate communities (DYER et al., 1986; FINLAY et al., 1991): 1) an epilimnetic assemblage of aero- bic, cosmopolitan ciliates such as Vorticella, Paramecium, Stentor, Stromhidium and others. This assemblage includes both algivorous and bacterivorous ciliates. 2) an oxyclinal
assemblage formed by sharply stratified maxima of algivo- rous ciliates with endosymbiotic Chlorella cells such as Coleps hirtus (ESTEVE et al., 1988) or Prorodon (see fig. 5A and fig. 6). And 3) an anaerobic assemblage constituted by sulfide-tolerant, bacterivorous ciliates such as Metopus es, Plagiopyla ovata or Caenomorpha medusula. Some of these anaerobic ciliates have endosymbiotic methanogenic bacteria (FINLAY et al., 1991). In addition, scuticociliates,
e.g. Cyclidium, are found either in the oxycline or the moni- molimnion. More details about the feeding activities of these ciliates can be found in GUERRERO & PEDRÓS-ALIÓ (1992) and FINLAY et al. (1991).
CONCLUSION
Karstic lakes present accumulations of organisms at depth.
The comparison of different kinds of meromictic and holo- mictic stratified lakes (fig. 3) has allowed to identify the redoxcline as the most important environmental factor for the organization of the community. In some cases the redoxcline may coincide with a thermocline, a chemocline or a halocline, but not necessarily. The organisms found in different lakes usually belong to the same genera and many times to the same species. Usually, one can find large abundances of copepod nauplii (especially T. prasinus), microaerophilic rotifers (Anuraeopsis, Filinia), flagellated algae (Cryptomonas) and in some lakes also cyanobacteria (Oscillatoria) at the oxic- anoxic boundary. Characteristic genera of ciliates are also associated with these interfaces (Coleps, Prorodon, and anae- robic ciliates), where purple and green sulfur bacteria form abundance maxima. The same components have been found in most lakes studied showing that this community is wides- pread and quite different from the epilimnetic community. Moreover, the deep living community persists throughout the
stratification period without much changes. These deep living communities, therefore, deserve more study before current models of trophic webs, nutrient cycling, and succession can be generalized to al1 aquatic communities.
ACKNOWLEDGMENTS
The work of M.R.M. and E.V. has been supported by DGICYT grant NT89-1124 and by the "Consejería de Agri- cultura" of the Castilla-La Mancha Autonomous Government. The work of C. P.A. has been supported by DGICYT grant PB87-0183 from the Spanish Ministry of Education and
Science.
REFERENCES
AB ELLA, C.A., 1980. Dinámica pohlacionul comparada de hacterias,fototr(~ica.s planctónicas. Ph.D. Thesis. Auto- nomous University of Barcelona.
ABELLA, C.A., E. MONTESINOS, & R. GUERRERO, 1980. Field studies on the competition between purple and green sulfur bacteria for available light. (lake Sisó, Spain). Deiielopments in Hydrohiology, 3: 173-1 8 1.
ABELLA, C.A., E. MONTESINOS & R. GUERRERO, 198 1. Dinamica i estratificació de poblacions bacterianes fototrbfiques en una cubeta meromíctica del llac de Banyo- les. Bull. Soc. Catalana Biol., 5-7: 11-27.
ABELLA, C.A., E. MONTESINOS & J. TURET, 1985. Colonization and dynamics of phototrophic bacteria in a recently formed lagoon in Banyoles karstic area (Girona, Spain). Scientia gerundensis, 10: 33-49.
ALFONSO, M.T. & M.R. MIRACLE, 1987. Variación temporal de las poblaciones zooplanctónicas de la laguna anóxica del Cisó (Gerona). Limneticu 3: 167-177.
ARMENGOL-DÍAZ, J., A. ESPARCIA, E. VICENTE & M.R. MIRACLE, 1992. Vertical distribution of planktonic rotifers in a karstic meromictic lake. Hydrohiologia, in press.
ARVOLA, L., A. OJALA, F. BARBOSA & S. 1. HEANEY, 1991. Migration behavior of three cryptophytes in relation to environmental gradients: an experimental approach. British Phycol. Jr . . 26: 361-373.
AVILA, A,, J.L. BURRELL, A. DOMINGO, E. FERNÁNDEZ, J. GODALL & J.M. LLOPART, 1984. Limno- logía del Lago Grande de Estanya. Oecol. aquat. 7: 3-24.
BAKER, A. L., K. K. BAKER & P. A. TYLER, 1985. A family of pneumatically-operated thin layer samplers for


cas en una laguna anaerobia de aguas sulfurosas. O c ~ o l . ayuat.. 3: 193-205.
GUERRERO. R.. C.A. ABELLA & M.R. MIRACLE, 1978. Spatial and temporal distribution of bacteria in a
meromictic karstic lake basin: relationship with physico- chemical parameters and zooplankton. Verh. Ii~tei . i~at. Verein. Limnol., 20: 2264-2274.
GUERRERO, R.. E. MONTESINOS, 1. ESTEVE & C.A. ABELLA, 1980. Physiological adaptation and growth of purple and green sulphur bacteria in a meromictic lake (Vil&) as compared to a holomictic lake (Cisó). Develop- rnentuls in Hydrohiology, 3: 16 1 - 17 1.
GUERRERO. R., E. MONTESINOS, C. PEDRÓS-ALIÓ, 1. ESTEVE, J. MAS, H. VAN GEMERDEN, P.A.G. HOFMAN & J.F. BAKKER, 1985. Phototrophic sulfur bacteria in two Spanish lakes: Vertical distribution and limi- ting factors. Limnol. Oceanogr., 30: 919-93 1.
GUERRERO, R., C. PEDRÓS-ALIÓ, 1. ESTEVE, J . MAS, D. CHASE & L. MARGULIS, 1986. Predatory prokaryotes: predation and prirnary consumption evolved in bacteria. Proc. Natl. Acad. Sci. USA, 83: 2138-2142.
GUERRERO, R., C. PEDRÓS-ALIÓ, I. ESTEVE & J . MAS, 1987. Communities of phototrophic sulfur bacteria in lakes of the Spanish Mediterranean region. Acta Acad. Ahoensis, 47: 125-151.
GUERRERO. R. & C. PEDRÓS-ALIÓ, 1992. The micro-
bial component in Spanish aquatic ecosystems. In C. Montes & C. Duarte (eds.) Limnology in Spain, Limnetica, 8.
HOBBIE, J.E., R.J. DALEY & S. JASPER, 1977. Use of Nuclepore filters for counting bacteria by epifluorescence microscopy. Appl. Environ. Microhiol., 33: 1225- 1228.
HOFMANN, W. 1982. On the coexistence of two pelagic Filinin species (Rotatoria) in Lake PluBsee. 1. Dynamics of abundance and dispersion. Arch. Hydrobiol., 95: 125- 137.
HOFMANN, W. 1985. Dynamics of vertical zooplankton community in the PluBsee: Cluster analysis. Verh. Internat. Verein. Limnol., 22: 2983-2986.
HOFMANN, W. 1987. Population dynamics of hypolim- netic rotifers in the PluBsee (North Germany). Hydrohiolo-
gis, 147: 197-201. HURLEY, J.P. & C.J. WATRAS, 1991. Identification of
bacteriochlorophylls in lakes via reverse-phase HPLC. Limnol. Oc,eanogr., 36: 307-3 14.
INGVORSEN, K. & B. B. JGRGENSEN, 1979. Combi- ned measuremerit of oxygen and sulfide in water samples. Limnol. O c e u n o ~ r . , 24: 390-393.
JEFFREY, S.W. & C.F. HUMPHREY, 1975. New spec- trophotometric equations for determining chlorophylls a , b ,
c , - and ('z - in higher plants, algae and natural phytoplankton. Biochern. Phjsiol. Pflanzen, 167: 19 1 - 194.
JORGENSEN, B.B., J.C. KUENEN & Y. COHEN, 1979. Microbial transformations of sulfur compounds in a stratified lake (Solar Lake, Sinai). Limnol. O(.canogr.,
28: 1075- 1093. KORTHALS, H.J. B: C.L.M. STEENBERGEN, 1985. Sepa-
ration and quantification of pigments from natural phototrophic microbial populations. FEMS Microbiol. Ecol., 3 1 : 177- 185.
KOSTE. W.. 1980. Uber zwei Plankton-Radertiertaxa Filiiiia arrsti.alensis n sp. und Filiniu kofrnanni n sp., init Bemerkungen zur Taxonomie des longiseta - terminalis
Gruppe. Genus Filinia Bory de St. Vincent, 1824, Familie Filiniidae Bastos 1959 (Ubeordnung Monogononta). Arch. Hydrohiol., 90: 230-256.
KOSTE, W., 1991. Aniiraeopsis miraclei, a new plankto- nic rotifer species in karstic lakes of Spain. Hydrohiologia,
209: 169- 173. LARSSON, P., 197 1. Vertical distribution of planktonic
rotifers in a meromictic lake. Noi .~ , . .J. Zool., 19: 47-75. MANTOURA, R.F.C. & C.A. LLEVELLYN, 1983. The
rapid determination of algal chlorophyll and carotenoid pigments and their breakdown products in natural waters by reverse-phase high-performance liquid chromatography. Anal. Chim. Acta, 15 1: 297-3 14.
MAS, J., C. PEDRÓS-ALIÓ & R. GUERRERO, 1 9 9 0 . 1 ~ ~
situ specific loss and growth rates of purple sulfur bacteria in Lake Cisó. FEMS Microbiol. Ecol., 73: 27 1-28 1.
MIKSCHI, E.. 1989. Rotifer distribution in relation to temperature and oxygen content. Hydrohiologiu, 18611 87: 209-214.
MIRACLE, M.R.. 1974. Niche structure in fresh water zooplankton. A principal components approach. Ecology,
55: 1306- 13 16. MIRACLE, M.R., 1975. Segregation of zooplankton
populations in several depressions within one lake basin. Verh. Iritel.nat. Verein. Limnol., 19: 1 140-1 149.
MIRACLE, M.R., 1976. Distribución en el espacio y en el tiempo de las especies del zooplancton del lago de Banyo- les. ICONA Monografías, Ministerio Agricultura y Pesca. 5: 1-270.
MIRACLE, M.R. & E. VICENTE. 1983. Vertical distri- bution and rotifer concentrations in the chemocline of mero- mictic lakes. Hydi-ohiologia, 104: 259-267.
MIRACLE, M.R. & E. VICENTE, 1985. Phytoplankton and photosynthetic sulphur bacteria production in the mero- mictic coastal lagoon of Cullera (Valencia, Spain). Verh.
Internat. Verein. Limnol., 22: 2214-2220.

MIRACLE, M.R. & M.T. ALFONSO, 1992. Rotifer vertical distribution in a meromictic basin of Lake Banyo- les (Spain). Hydrobiologia, (in press).
MIRACLE, M.R., E. VICENTE, R.L. CROOME & P.A. TYLER, 1991. Microbial microcosms of the chemocline of a meromictic lake in relation to changing levels of PAR. Verh. Internat. Verein. Limnol.. 24: 11 39-1 144.
MITCHELL, J. G. & J.A. FUHRMAN, 1989. Microdis- tribution of heterotrophic bacteria. Mar. Ecol. Prog. Ser.,
53: 141-148. MODAMIO, X., V. PÉREZ & F. SAMARRA, 1988.
Limnología del lago de Montcortks (ciclo 1978-79) (Pallars Jussh, Lleida). Oecol. aquat., 9: 9-17.
MONTESINOS, E., R. GUERRERO, C. ABELLA & 1. ESTEVE, 1983. Ecology and physiology of the competition for light between Chlorobium limicola and Chlorobium
phaeobacteroides in natural habitats. Appl. Environ. Micro- biol., 46: 1007-1016.
MOYA, G., G. RAMON, A. MARTINEZ TABERNER, V. FORTEZA, C. PICÓ, C. PONSELL, R. R O S S E L L ~ & M.A. SOBERATS, 1987. Limnology of a meromictic coas- tal lagoon, L'Estany Cibollar (Majorca, Balearic Islands). Limnetica, 3(2): 255-262.
PLANAS, M.D., 1973. Composición, ciclo y productivi- dad del fitoplancton del lago de Banyoles. Oecol. aquat., 1: 3-106.
PEDRÓS-ALIÓ, C., 1989. Towards an autecology of bacterioplankton. In: U. Sommer (ed.) Plankton Ecology. Succession in Plankton Communities. 297-336. Springer- Verlag, Berlin.
PEDRÓS-ALIÓ, C. & M.M. SALA, 1990. Microdistri- bution and diel vertical migration of flagellated vs. gas- vacuolate purple sulfur bacteria in a stratified water body. Limnol. Oceanogr., 35: 1637-1644.
PEDRÓS-ALIÓ, C., J.M. GASOL & R. GUERRERO, 1986. Microbial ecology of sulfurous lake Cisó. In: F. Megu- sur &M. Gantar (Eds.). Perspectives in Microhial Ecology: 638-643. Slovenian Soc. Microbiol, Ljubjana, Yugoslavia.
PEDRÓS-ALIÓ, C., J.M. GASOL & R. GUERRERO, 1987. On the ecology of a C~yptomonas phaseolus population forming a metalimnetic bloom in Lake Cisó, Spain: Annual distribution and loss factors. Limnol. Oceanogr., 32: 285-298.
PICK, F. R., C. NALEWAJKO & D. R. S. LEAN, 1984. The origin of a metalimnetic chrysophyte peak. Limnol. Oceanogr., 29: 125-134.
PORTER, K.G. & Y.S. FEIG, 1980. The use of DAPI for identification and enumeration of bacteria and blue-green algae. Limnol. Oceanogr., 25: 943-948.
RIERA, X G. & C.A. ABELLA, 1986. Dinhmica anual de les poblacions de bacteris fototrofics de la llacuna costanera de La Massona (Alt Empordh, Girona). Scientia gerunden- sis, 12: 35-48.
RIERA, X G. & C.A. ABELLA, 1991. Limnological cycle of the coastal lagoon La Massona (Girona, NE Spain). Origin, dynamics and influence of the sporadic seawater infalls on the meromixis. Verh. Internat. Verein. Limnol., 24: 1029-1031.
ROJO, C. & M.R. MIRACLE, 1987. Poblaciones fito- planctónicas de la Laguna de la Cruz (Cuenca), una laguna cárstica meromíctica. Actas IV Simp. Botanica Cryptoga- mica, Granada (Spain): 119-135.
ROJO, C. & M.R. MIRACLE, 1989. Phytoplankton fluc- tuations during an annual cycle in the coastal lagoon of Cullera (Spain). Int. Revue ges. Hydrobiol., 74: 179- 194.
ROTT, E., 1988. Some aspects of the seasonal distribution of flagellates in mountain lakes. Hydrobiologia, 161: 159- 170
RUTTNER-KOLISKO, A., 1975. The vertical distribu- tion of planktonic rotifers in a small Alpine lake with a sharp oxygen depletion (Lunzer Obersee). Verh. Internat.
Verein. Limnol., 19: 1286- 1294. RUTTNER-KOLISKO, A., 1977. Population dynamics of
rotifers as related to climatic conditions in Lunzer Obersee and Untersee. Arch. Hydrobiol., Beinh. Ergebn. Limnol., 8: 88-93.
RUTTNER-KOLISKO, A., 1989. Problems in taxonomy of rotifers, exemplified by the Filinia longiseta - terminalis
complex. Hydrobiologia, 1861187: 291-298. SCHABER, P. & A. SCHRIMPF, 1984. On morphology
and ecology of the Filinia terminalis - longiseta group (Rotatoria) in Bavarian and Tyrolean lakes. Arch. Hydro-
hiol., 101 : 247-257. SOMMER, U., Z. M. GLIWICZ, W. LAMPERT & A.
DUNCAN, 1986. The PEG-model of seasonal distribution of planktonic events in freshwaters. Arch. Hydrobiol., 106: 433-471.
SOROKIN, Y.I., 1970. Interrelation between sulphur and carbon turnover in meromictic lakes. Arch. Hydrobiol., 66: 391 -446.
STANIER, R.Y. & J.H.C. SMITH, 1960. The chlorophylls of green bacteria. Biochim. Biophys. Acta, 41: 478-484.
STRICKLAND, J. D. H. & T. R. PARSONS, 1972. A practica1 handbook of seawater analysis. Bull. Fish. Res.
Board Canada, 167. TAKAHASHI, M. & S . ICHIMURA, 1968. Vertical
distribution and organic matter production of phototrophic sulfur bacteria in Japanese lakes. Limnol. Oceanogr., 13: 644-655

uTERMOHL, H., 1958. Zur Vervollkommnung der
quantitativen Phytoplankton-Methodik. Mitt. Int. Verein. Limnol., 9: 1-38.
VICENTE, E., & M.R. MIRACLE, 1984. Distribution of photosynthetic organisms in a temporal stratified karstic pond near Cuenca (Spain). Verh. Internat. Verein. Limnol.,
22: 1504-1710. VICENTE, E., & M.R. MIRACLE, 1988. Physicochemi-
cal and microbial stratification in a meromictic karstic lake of Spain. Verh. Internat. Verein. Limnol., 23: 522-529.
VICENTE, E., M.A. RODRIGO, A. CAMACHO & M.R. MIRACLE, 1991. Phototrophic prokaryotes in a karstic sulphate lake. Verh. Internat. Verein. Limnol., 24: 998-1004.
ZIMMERMANN, R. & L. A. MEYER-REIL, 1974. A new method for fluorescence btaining of bacteria1 popula- tions on membrane filters. Kieler Meeresjbrch., 30: 24-27.

LAKE GALLOCANTA (ARAGON, NE. SPAIN), A PARADIGM OF FLUCTUATIONS AT DIFFERENT SCALES OF TIME.
F. A. Comín, X. Rodó and P. Comín Department of Ecology. University of Barcelona, Diagonal 645, 08028 Barcelona, Spain
Keywords: Saline lakes, Fluctuations, Time scales.
ABSTRACT
Physical, chemical and biological characteristics fluctuate in many saline lakes at different scales of time. They are very adequate for studies of the structure and functioning of the biological community because changes in the species composition and biomass occur extremely related to changes of the physical and chemical characteristics of the water, which, in turn, respond to climatic fluctuations.
In Lake Gallocanta, a saline lake in NE Spain, water level changed from a maximum depth of 2 m in 1977 to 0 m (in fact the water level was between 10 and 30 cm below the ground) in 1985, and reached again a relatively high depth of 140 cm in 1991. Seasonal fluctuations are also very clear. The chemical composition of the water is related to the water level. Seasonal fluc- tuations of total dissolved solids are inversely related to water volume in the lake. However, a more complex pattern of ionic composition and total dissolved solids is observed at a longer time scale (severa1 years). Salts are lost selectively by precipita- tion and eolic erosion during dry periods. The relative abundance of different ions changes between years.
Fluctuations of organisms are illustrated by changes in the relationship between number of cells and number of species of phytoplankton from a severa1 years. During periods of time of increasing water level-decreasing salinity the number of cells increases while the number of species decreases (relatively low K values). During periods of time of decreasing water level- increasing salinity, relatively high values of K are found. The same pattern is observed at different scales of time: comparing trends during a series of years and seasonal changes. At the ecosystem level, a similar view is proposed: the relationships between living and non-living compartments are also submitted to fluctuations at different scales of time. The number of links are lower and their intensity higher during low water level-high salinity periods than during high water level-low salinity periods. The number and intensity of the links between compartments must change seasonally, annually and at higher scales of time in accordance with the fluctuations of chemical and physical variables.
INTRODUCTION
Saline lakes provide excellent conditions for ecological studies of aquatic ecosystems. Their community structures and energy flux networks are simpler than in freshwater ecosystems (HAMMER, 1986). They also provide highly valuable natural resources to humans (WILLIAMS, 1981). In spite of these advantages and the fact that the volume of inland saline waters in the world is quite similar to the volume of freshwaters (VALLENTYNE, 1972), only recently have saline lakes received adequate attention from the international limnological community. Fortunately, limnological studies of saline lakes proliferated al1 around the world after the initiation of a series of Symposia on
Inland (Athalassic) Saline Lakes endorsed by SIL (WILLIAMS, 1981, HAMMER, 1983, MELACK, 1988, COMIN & NORTHCOTE, 1990, HURLBERT, in pr-ess).
Fluctuations in water level, as a consequence of meteo- rological changes, occur in saline lakes at different time scales. Consequently, changes in the biological communi- ties related to physical and chemical characteristics occur in saline lakes more often and more intensely than in freshwa- ter lakes. Adaptation to fluctuations rather than to ranges or precise values of physical and chemical variables must be acomplished by populations in order to survive (BROCK,
1986). Examples of saline lakes fluctuating at different frequencies and intensities range from lakes which only fill up ocasionally (Eyre, WILLIAMS & KOKKIN, 1988) to
Limnetica, 8: 79-86 (1992) O Asociación Española de Limnología, Madrid. Spain

permanent lakes (Pyramid, GALAT et al., 1990). Spanish saline lakes present a great variety of geomorphological characteristics and are located in different climatic regions (fig. 1 ) (COMIN & ALONSO, 1988). Water level fluctua- tions also occur in Spanish saline lakes with diverse frequency and intensity (fig. 2). Lake Gallocanta is an exam- ple of a lake in which fluctuations in the water level can be clearly observed at different scales of time (fig. 2). Conse- quently, the chemical characteristics of the water, the struc- ture of the biological community and, moreover, the func- tioning of the entire ecosystems also fluctuate at different time scales. Here, this view is developed reviewing data obtained in studies carried out irregularly during the last fourteen years as a way to propose long-term studies of ecosystems of this type.
Figure 1 . Location of i ~ l o b t of the Spanish saline lakes in different climatic regions. 1-Humid, 2-Semihumid, 3- Semiarid, 4-Arid. The asterisk indicates the location of Lake Gallocanta.
GEOGRAPHICAL SETTING
Lake Gallocanta is located at 987 m a.s.1. in the central part of the Iberian Range (400 50' N, 20 11' W), at the limit between semiarid and semihumid weather, with an average anuual precipitation of 440 mm which also undergoes wide seasonal and interannual fluctuations (COMIN et al., 1990). Lake Gallocanta represents the level of the water table at the hydrological terminus of a 520 km2 watershed. The present features of the watershed developed from a previously much wider area originated tectonically at the end of the Pleocene (HERNANDEZ & ARANEGUI, 1926) . During the Quater- nary period karstification and dissolution of triassic evapo-
rites gave rise to the present saline conditions of the lake (GRACIA, 1990).
Lake Gallocanta was long considered a closed endorheic area. Recent studies calculated indirectly a net export of
Gallocanta groundwater towards nearby watersheds. The groundwater export would account for between 5-30 % of
the total renewable groundwater (CRUZ et al., 1979, TRAGSA. 1988)
Time, yeors
Figure 2. Idealized types of above ground water leve1 fluctuations occurring in Spanish saline lakes during the last years: top, perma- nent saline lake; middle, saline lake which persisted inundated during one dry season; bottom, saline lake which remained dry during one wet season.
SALINITY VERSUS WATER LEVEL FLUCTUATIONS
The water level in Lake Gallocanta fluctuates at different scales of time (fig. 3) as a consequence of interannual chan- ges of precipitation. Seasonal fluctuations are overlapped by medium and long term fluctuations. At a time scale smaller than the seasonal the number of fluctuations increases, although it is clear that the changes in water level at these intervals of a few days are of the order of mm and cm. So, it is clear that there is an indirect relationship between inten- sity and frequency of water level changes, as for any distur- bance in any ecosystem (MARGALEF, 1991 ).
There is no direct relationship between annual precipita- tion and water level. Water level fluctuations are related to precipitation through complex interactions because of the way water must run through the watershed (above and below
Similar differences, although less intense, occur during
g I-' seasonal fluctuations of the water level. Precipitation of salts - a and accumulation in the sediments, which occurs with spatial > Q> +
heterogeneity, and wind deflation are responsible for the loss 300 u of large amounts of salts (COMIN et al., 1991). Redissolu-
L a tion of salts accumulating at the surface of the sediments and, C, 150 most important, washed up by water flowing through
g (bottom-up) the sediments (COMIN & FORES, 1990) incor- I porate salts from the solid to the dissolved phases.
Relationships of major ions dissolved in the water (chlo- Figure 3. Water level and total dissolved solids recoi-ded in Lake Gallocanta over time during a series of years. ride, sulphate, carbonate plus bicarbonate, sodium, magne-
sium, calcium, potassium) also show changes over time (table 1). Differences related to conservative changes (e.g.,
ground) and the high evaporation rates. A response time of linear relationship with chloride concentration) indicate the the water to rainfall between 0.3-2.4 years was calculated intensity of biogeochemical processes which can be impor- according to LANGBEIN (1961). This means that major tant in the lake waters (e.g.: differential precipitation, sulp- changes in the seasonal pattern of rainfall (or in the water hate reduction) (COMIN et al., 1990, 1991).
outflows) will cause marked lake level fluctuations after two years if the water level remained relatively stable for seve- ral years, and after four months if the water level fluctuated
during the previous years (COMIN et al., 1991). A general inverse relationship exists between water level
and salinity (fig. 3). However, fig. 4 gives a clearer picture of indicators of intense changes in the water characteristics after seasonal and longer fluctuations of the water volume. The comparison of concentrations at times of different years with identical water volume shows that the total amount of salts in aboveground lake waters decreases during dry periods (1983- 1986) and increases during refilling periods (1987-91).
POPULATION FLUCTUATIONS
Seasonal fluctuations in the biological community were extremely clear and intense in Lake Gallocanta during one year (1980-81) within the drying phase (fig. 3) (COMIN et al., 1983). Longer term fluctuations are still under study.
A quick and simple examination of the composition of a part of the biological community, the phytoplankton, at different years reveals important differences, both in speci- fic composition and abundance. A change in the structure of
Table 1. Total dissoved solids (g 1.') and Cl/S04 ratio (meqlmeq) in Lake Gallocanta during two years, before (1980-81) and just after (1986-87) a complete drought of the Lake.
1980 - 1981 1986 - 1987 TDS CI/S04 TDS CIISO~
November December February March April
M ay June July August September October
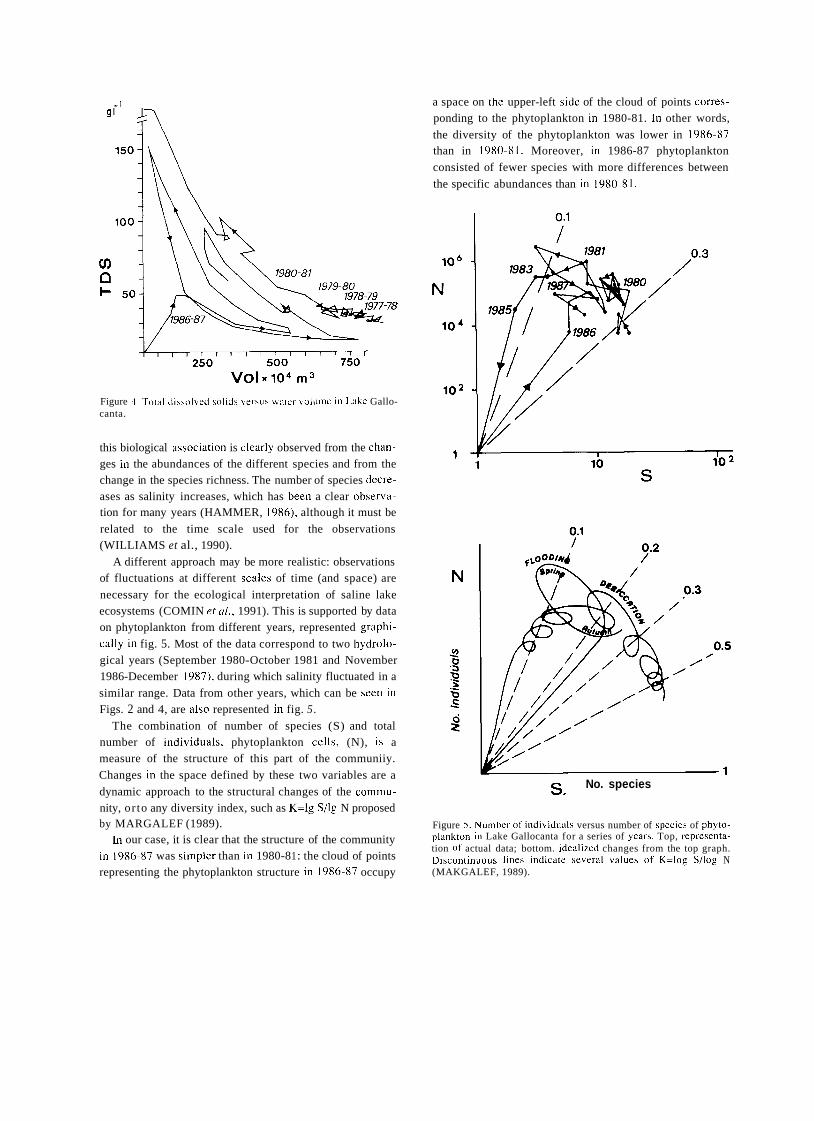
Figure 1. 'l'oial tli\aolved \olid\ versu\ water \oluiiic in Lake Gallo- canta.
this biological association is clearly observed from the chan- ges in the abundances of the different species and from the change in the species richness. The number of species decre- ases as salinity increases, which has been a clear observa- tion for many years (HAMMER, 1986), although it must be
related to the time scale used for the observations (WILLIAMS et al., 1990).
A different approach may be more realistic: observations of fluctuations at different scales of time (and space) are
necessary for the ecological interpretation of saline lake ecosystems (COMIN et d., 1991). This is supported by data on phytoplankton from different years, represented graphi-
cally in fig. 5. Most of the data correspond to two hydrolo- gical years (September 1980-October 1981 and November 1986-December 1987), during which salinity fluctuated in a
similar range. Data from other years, which can be seen in Figs. 2 and 4, are also represented in fig. 5.
The combination of number of species (S) and total number of individuals, phytoplankton cells, (N), is a measure of the structure of this part of the communiiy.
Changes in the space defined by these two variables are a
dynamic approach to the structural changes of the commu- nity, or to any diversity index, such as K=lg S/lg N proposed by MARGALEF (1989).
In our case, it is clear that the structure of the community in 1986-87 was simpler than in 1980-8 1 : the cloud of points representing the phytoplankton structure in 1986-87 occupy
a space on the upper-left side of the cloud of points corres- ponding to the phytoplankton in 1980-81. In other words,
the diversity of the phytoplankton was lower in 1986-87 than in 1980-8 1. Moreover, in 1986-87 phytoplankton
consisted of fewer species with more differences between
the specific abundances than in 1980-8 1.
S, No. species
Figure S . Nuinber of iiidividuals versus number of species of phyto- plankton in Lake Gallocanta for a series of yearb. Top, representa- tion oí' actual data; bottom. idealized changes from the top graph. Discontinuous lines indicatc several values of K=log Sllog N (MAKGALEF, 1989).

Fig. 5 also shows that the amplitude of the changes in the phytoplankton structure was wider in 1986-87 than in 1980- 8 1 . It corresponds to the characteristics of the community
after an intense perturbation like the dryness of the lake,
which was beginning its self organization, the early stages of ecological succession. In come way, this change of struc- ture during a fluctuation is the same type of dynamic change
a community follows through time (e.g.: seasonal cycles in
temperate ecosystems). In fig. 5, points corresponding to spring data lie on the ascendent left side and points corres- ponding to fall data lie in the descendent-right side of the
loops (MARGALEF, 1989). This corroborates the hypothe-
sis expressed before: each of the seasonal fluctuations in Lake Gallocanta follows a different trajectory. However, these seasonal trajectories are loops within a longer trajec- tory. The few available data of 1982 and 1985 pointed out in
fig. 5 allow us to form an image of the trajectory of the
structure of phytoplankton, which would correspond to the
fluctuation at longer time scale represented in fig. 2. Again, the biological structure fluctuates at different time scales in
Lake Gallocanta. We can easily imagine similar trajectories
for each fluctuation between dry periods, Lake Gallocanta has dried up four times this century (GUIRAL, 1981), or even at longer, geological, scale.
COMMUNITY FLUCTUATIONS
Like phytoplankton, every component of the ecosystem
fluctuates in Lake Gallocanta at different time scales. More- over, the relative importance of the relationships between the components of the biological community and of those
with abiotic factors also changes with time. So, fluctuations at different scales of time must also be shown by the energy
and trophic ecosystem networks. This is a suggestion that we will now discussed simply on the evidence of direct observations for a number of years and some data from other
saline lakes.
Let us first begin with a simple comparison of several differences that have been observed in Lake Gallocanta between a high water level-low salinity stage (e.g., 1977)
and a low water level-high salinity stage (e.g., 1981). Some
of these differences have been clearly explained by De DECKKER (1988), who pointed out three basic phases or groups of salt lakes according to their biological and sedi-
mentary facies: wet phases of ephemeral shallow lakes, dry lakes and deep water lakes. In Lake Gallocanta, during a high water level-low salinity stage, phytoplankton and
rooted submerged macrophytes are very important, both in bioniass arid dynamic roles, providing energy for other trop- hic levels. Zooplankton biomass is lower than phytoplank-
ton. Dense populations of macroinvertebrates live in asso-
ciation with the macrophytes and in the sediments. A great variety of birds are present in the lake reaching very high densities. E.g.: the number of aquatic birds observed during winter 1978-79 was 148,892 I23,410, while during winter
1980-81 it was 52,825 L 24,728 (n=5, census carried out
inonthly from October to February) (HERNANDEZ et al., 198 1). Rooted submerged macrophytes also develop high
biomass populations during high water level phases
(COMIN er al., in pr-ess), a balance for the entire ecosystem would give production over respiration, which would be predominantly aerobic. during high water level-low salinity stages.
During low water level periods, huge amounts of organic
matter accumulate in the lake bottom and shores . At low water level-high salinity stages, the biomass of consumers (e.g., zooplankton) is higher than the biomass of producers on which they feed (e.g., phytoplankton) for short periods
of time (one month) (GARCIA, 1991). However, conside- ring a longer period of time, the biomass ratio (phytoplank- ton/zooplankton) reaches an equilibrium close to 1, the lowest limit proposed by SHELDON er a1.(1977) for pelagic
systems. Enormous microbial mats developed in Lake
Gallocanta in summer 198 1 and huge amounts of decompo- sing filamentous algae accumulated on the shores and bottom and in the water column of Lake Gallocanta during
summer 1990 and 1991. Then respiration, mostly anaerobic, would overcome production during these stages. After this simplistic comparison it is necessary, following the point of
view expressed before, to address the changes in the rela- tionships between components of the ecosystem as fluctua-
tions at different scales of time, like those described for phytoplankton or salinity above. For example, it is clear that most of the diet of the bird populations in Lake Gallocanta consists of rooted submerged macrophytes. However, during
periods of macrophyte decline (1981), the proportion of the energy obtained by birds from a variety of insects living in the waters and seeds of terrestrial plants close to the shore-
line increases (GUIRAL, 1981). Moreover, the total nuniber
of birds decreases as macrophyte beds decline (HERNAN-
DEZ el al., 1981). Seasonal fluctuations in rnacrophytes are very clear. During
a series of years of low water level-high salinity (e.g.,1990- 91), two populations of rooted submerged macrophytes (Ruppia dr-epanensis and Lampr-othamnium l>upulosum ) grew

very quickly during winter-spring time, reached a relatively
low biomass and died, decomposing during summer while salinity was increasing. The interesting point is that the spatial distribution (studied in a transect perpendicular to the shore-
line) of the macrophyte populations was changing along the
growing season. Large shores appeared as the water level decreased. Drying-flooding cycles of these shores, which is relatively frequent at these low water level stages, stimulate
the microbial activity and enhance the mineralization of nutrients (AGARWAL et al., 197 1, HOWARD-WILLIAMS, 1972). The lake sediment became progressively anoxic as organic matter accumulated and decomposed on it and, suppo- sedly, the sediment-water exchanges were enhanced.
During years of high water level-low salinity, macrophyte populations also show seasonal fluctuations. However, dense stands reinain during summer, fall and winter. The atructure of the sediments remain with fewer changes
because the root system and relationships between sediment
and water column and between macrophytes and consumers are not submitted to such marked seasonal changes as during
low water level stages. Probably, the number of links (connections between
living and/or non living compartments of the system) decre- ases and their intensity (energy transfers) increases during low water level phases and viceversa during high water level
phases (fig. 6). Again, we suppose that these relationships
fluctuate at different scales of time in Lake Gallocanta. Evidently, the amount of research which is now waiting to be done is overwhelming.
WATER C W M N
Figure h. Schemaiic representatioii of the relationships between compartments of Lake Gallocanta at two different stages:top, low water level-high salinity; bottom, high water level-low salinity.
FUTURE PERSPECTIVES
The work presented here was carried out with the same
characteristics mentioned by STRAYER et al. (1986) for
other long-term studies. A serendipitous combination of dedicated scientists, the opportunity (site and ideas) and funding (fluctuating at different time scales) made it possi-
ble to sample Lake Gallocanta at irregular intervals. These efforts provided data which were very useful for partial interpretation of ecological processes. However, the number of ideas for future studies after each partial approach rose quickly. All the potential subjects of limnological research
in Lake Gallocanta had a common link: observations at different time scales are necessary for a correct interpreta- tion of phenomena.
In other words, Lake Gallocanta presents phenomena for which long-term studies are recognized as essential for a correct understanding. For example, for seven years (1977- 83) water level showed a net decreasing trend (fig. 2). During this period, a study lasting just one year would not
have shown this trend, nor some of its ecological conse- quences (e.g.: macrophyte and bird population changes).
This is a common feature of some saline lakes where it
has been observed that fluctuations of physical, chemical
and biological characteristics occur clearly defined at diffe- rerrt scales of time. The same types of slow process must occur in Lake Chilwa, where similar types of fluctuations were recorded (KALK et al., 1979).
Lake drying up is a rare event which has happened once in the 15 year series recorded here for Lake Gallocanta.

Sudden refilling of usually dry lakes (Lake Eyre, WILLIAMS & KOKKIN, 1988) or permanence of water al1 the year round in lakes which usually dry up every year (Lake Fuente de Piedra, GARCIA, 1991) are also examples of episodic phenomena. However, lakes of these types do not offer the wide range of fluctuations, both in frequency and intensity, which make a saline lake particularly suitable for long-term studies. Processes with high variability also require long-term periods of observations to describe al1 the variability range. This is the case of saline lakes where sali- nity and the biological community change over a wide range but they take several years to fluctuate within the entire range. Long-term series of data are also important to sort out the relative contribution of different factors involved in complex ecological phenomena (e.g.: temporal and spatial pattems of macrophyte distribution, COMIN et al., in press).
Once we are convinced of the necessity of long-term studies to understand saline lake ecosystems fluctuations, it is neces- sary to couple long-term data collecting with the formulation of ecological hypotheses (RISSER, 1991). This is necessary to justify the expenses of long-term studies. One needs to persuade to funding entities that the work will be useful and one should try to contribute to general ecological theory, or at least to test some ecological concepts. This is easy because many hypotheses arise soon after the first observations.
Saline lakes commonly act as useful recorders of climatic changes because many fossil remains are preserved in the sediments (De DECKKER, 1988). In addition, the high sedi- mentation rates of saline lakes favor reconstructions on long and short time scales, as the behaviour of these aquatic systems may be inferred during periods of climatic change. However, the biological remains are not well preserved in al1 the sediments because some chemical and geological processes alter their mineral components (De DECKKER, 1988). The key point is to find out the function to transfer from sedimentary records to changes in the past characte- ristics of the lake and of these with the hydrological regime. This has been successfully done by FRITZ (1990) with data from Lake Devils, North Dakota (USA) recorded for the last 100 years. She asserted "...saline lake sediments are probably unparalleled as tools for paleohydrologic and pale- oclimatic reconstruction in arid and semiarid regions", and many authors agree with her. From these types of study forecasts for future changes in saline lakes and wetlands can be inferred and, consequently, suggestions for water use and land management can be developed.
Ecological succession (changes in the community struc- ture through time, self-organization versus externa1 regula-
tion) and the controls of production in aquatic ecosystems (allocthonous versus autocthonous, planktonic vs. benthic, aerobic vs. anaerobic processes) are major subjects in Ecology. Many saline lakes provide excellent conditions to check regularities because: i- they are quite closed systems compared to other aquatic ecosystems and, consequently, boundaries can be better defined; ii- the shallowness of the water and the flatness of the bottom permit quite acceptable integrations at the ecosystem level. Moreover, because of these previous characteristics and the excellent coupling between hydrogeological and biological changes, the varia- bilities of different processes are well correlated or, at least, can be satisfactorily undertaken with little effort per unit of time, although, we insist, long-term records are necessary.
ACKNOWLEDGEMENTS
Thanks are given to Diputación General de Aragón (Servicio de Conservación del Medio Natural), which spon- sored most of tlie work presented here. One of the authors (X. R.) was in receipt of funds (F.P.I. grant) from the Spanish Ministry of Education and Science.
REFERENCES
AGARWAL, AS. , B.R. SINGH & Y. KANEHIRO, 197 1. Soil nitrogen and carbon mineralization as affected by drying-
rewetting cycles. Soil Sci. Soc. Amer. Proc. 35: 96-100. BROCK, M. A., 1986. Adaptation to fluctuations rather
than to extremes of environmental parameters. In: P. De Deckker & W.D. Williams (Eds.). Limnology in Austr-aliu: 13 1- 140. Dr. W. Junk Publishers. Dordrecht.
COMIN, F.A., M. ALONSO, P. LOPEZ & M. COME- LLES, 1983. Limnology of Gallocanta Lake, Aragón, NE. Spain. Hydrohiologia 105: 207- 221.
COMIN F.A. & M. ALONSO, 1988. Spanish salt lakes:
their chemistry and biota. Hydrohiologia 158: 237-245. COMIN, F.A. & T.G. NORTHCOTE (eds.), 1990. Suline
Lakes. Developments in Hydrohiology 59, 324 pp. COMIN, F.A. & E. FORÉS, 1990. Comprobación expe-
rimental de la importancia relativa de las vías superficial y subterránea de inundacibn en la liberación de sales de sedi- mentos desecados. Scientiu gerundensis 16: 69-77.
COMIN, F.A., R. JULIA, M.P. COMIN & F.PLANA, 1990. Hydrogeochemistry of Lake Gallocanta (Aragón, NE. Spain). Hydrohiologiu 197: 5 1-66.

COMIN, F.A., R. JULIA & P. COMIN, 1991. Fluctua- tions. the key aspect for ecological interpretation of saline lake ecosystems. Oecologia Aquuri<,cr 10: 127-135.
COMIN, F.A., X. RODO & M. MENENDEZ. (in press).
Spatial heterogenity of macrophytes in Lake Gallocanta. Hyrli.ohiolo~ía.
CRUZ, J., J.C. DOBLAS & M. GARRIDO, 1979. Hidro-
geología de la Laguna de Gallocanta. Actas 2 V i m p . Nac.
Hidro~eol . , Secc. IWidrogeol. , 4: 43 1-446. Asoc. Geol.
Esp. Minist. Obras Publ. DE DECKKER, P., 1988. Biological and sedimentary
facies of Australian salt lakes. In: Prileolimnology, aspects
c!f,fi-eshvt*ater paleor(~olo,qy and hiogeography (J. Gray, ed.): 237- 270. Elsevier Science Publ., Amsterdam.
FRITZ, S. C., 1990. Twentieth-century salinity and water
leve1 tluctuations in Devils Lake, North Dakota: Test of a diatom
based transfer function. Limnol. Oceanog~.. 35: 177 1- 178 1. GALAT, D.L., J.P. VERDIN & L.L. SIMS, 1990. Large
scale patterns of Nodularia spumigena blooms in Pyramid
Lake, Nevada, determined by Landsat imagery: 1972- 1986.
Hydrohiologia 197: 147- 164. GARCIA, C., 199 1. Estudio de un medio ac.uaric.o fluc-
t~iunte: La luguria atalusohalina de Fuente de Piedra (Mala,qa). (Unpublished Ph. D. Thesis). University of Málaga, 296 pp.
GRACIA, F.J., 1990. Geomorfología de la región de
Gallocanta (Unpublished Ph. D. Thesis). University of
Zaragoza, 660 pp. GUIRAL, J.J., 1981. Introducción al estudio de la alimen-
tación de las anátidas y fochas de la laguna de Galocanta.
In: E ~ t ~ i d i o de la hiocenosis de la loguna de galloc~urita y su cuenca. (PROYEX, S.A., ed.): 708-740. ICONA. Madrid.
HAMMER, U.T.(ed.), 1983. Saline Lakes. Developments
in Hydrohiology 105, 263 pp.
HAMMER, U.T., 1986. Saline Lake Ecosystems (f the
world. Dr. W. Junk Publ. Dordrecht. 616 pp. HERNANDEZ, E. & P. ARANEGUI, 1926. La laguna de
Gallocanta y la geología de sus alrededores. Bol. r. Soc. Esp.
Hisr. Nat. 26: 41 9-429. HERNANDEZ, F., M. BURGUETE & J.J. GUIRAL,
198 1. Estudio de la dinámica de las poblaciones de aves
acuáticas de la laguna de Gallocanta. In: Estudio de la
hiocenosis de la laguria de Gallocanta y s ~ i cuenca (PROYEX, S.A., ed.): 594-644. ICONA. Madrid.
HOWARD-WILLIAMS, C., 1972. Limnological studies in an african swamp: seasonal and spatial changes in the swamps of Lake Chilwa, Malawi. Arch. Hydrohiol. 70: 379-391.
HURLBERT, S. H. (ed.), (in press.). Suline Lukes. Deve-
lopnlents in Hydrohiology. KALK, M., A.J. MCLACHLAN & C. HOWARD-
WILLIAMS (eds.), 1979. Lake Chilwu, studies of change in
a tropical ecosystem. Monographiae Biologicae 35, 462 pp.
Dr. W. Junk Publ. Dordrecht.
LANGBEIN, W.B., 1961. Salinity and hydrology of closed lakes. USGS Prof. Paper 412, 220 pp.
MARGALEF, R., 1989. On diversity and conectivity as
historical expressions of ecosystems. Coenoses 4(3): 12 1 - 126. MARGALEF, R., 199 1. Teoria de los sistemas ecol6gi-
c.os. Estudi General 1, 290 pp. Publ. Univ. Barcelona.
MELACK, J.M.(ed.), 1988. Saline Lakes. Developments itz Hydrohiology 158, 316 pp.
SHELDON, R.W., W.H. SUTCLIFFE & M.J. PARAN- JAPE, 1977. Structure of pelagic food chain and relationship between plankton and fish production. J. Fish. Res. Bd. Can.
34:2344-2353.
STRAYER, D.J., GLITZENSTEIN, C.G. JONES, J. KOLASA, G.E. LIKENS, M.J. MCDONELL, G.G. PARKER &S.T.A. PICKETT, 1986. Long-term ecological
studies: an illustrated account of their design, operation, and importance to ecology. Occasional Puhlications 1 . Institute of Ecosystem Studies. New York.
TRAGSA, 1988. Estudio de los recursos hídricos de la Laguna de Gallocanta y acciones para su conservación.
ICONA, Minist. Agric. Pesca y Alim., 310 pp. VALLENTYNE, J.R., 1972. Effects of the demophoric
explosion on water and Man. In: The Environmental Future
(POLUNIN, N,, ed.): 406-414. The MacMillan Press.
London. 660 pp. WILLIAMS, W.D. (ed.), 198 1. Salt Lakes. De,,elopment.s
in Hydrohiology 5, 444 pp. WILLIAMS, W.D. & M.J. KOKKIN, 1988. The biogeo-
graphical affinities of the fauna in episodically filled salt lakes: A study of Lake Eyre South, Australia. Hydrobiologia 158:227-236.
WILLIAMS, W.D., A.J. BOULTON & R.G. TAAFFE,
1990. Salinity as a determinant of salt lake fauna: a question
of scale. Hydrohiologia 197: 257-266.

THE COASTAL LAGOON ALBUFERA D E VALENCIA: AN ECOSYSTEM UNDER STRESS
E. Vicente and M* R. Miracle Department of Microbiology and Ecology. Faculty of Biology, University of Valencia, E-46100 Burjassot. Valencia. Spain
Keywords: Eutrophication, Hypertrophic lagoons, Shallow lagoons, Coastal lagoons, Water chernistry, Phytoplankton. Zooplankton, Water management.
ABSTRACT
Recent studies (1980-88) on the Albufera of Valencia show that this shallow oligohaline coastal lagoon is an ecosystem under stress. Domestic and industrial waters loaded with phosphorus and ammonium reach the lagoon from the northern side. Nitrate-rich agricultural waters arrive mainly from the southern side. The estimated nutrient loadings are: Total P - 39 g m-2yr-1,
dissolved P = 14 g m-2yr-l. N-NH4+ = 74 g m-2yr-1 and N-(NO,-+NO;) = 81 g m-2yr-l. The stress produced in the Albufera by these P and N inputs is seen by the strikingly high values of chlorophyll concentration
and primary production, only comparable with the most hypertrophic lakes in the world. Chlorophyll u has an average value
around 300 pg 1-1, with a phytoplankton density of lo5 to loh ind ml-' corresponding to a biomass of 30-300 mg 1-1 (fresh weight).
The primary productivity, restricted to the lagoon surface, has been roughly estimated at 1.7 kg m-*yr-l. This phytoplankton
growth implies a nutrient consumption which results in very low nutrient concentrations in the lagoon water and the outlets. One half of the extraordinary biomass yield is then recycled and the other half exported to the sea (20 %,) and to the sediments (30 %).
Phytoplankton is generally constituted by filamentous cyanobacteria at 80 5%. The most frequent succession pattern of these
algae was a winter-spring dominance of P1anktothii.t- a,qrr~.dhii and a summer dominance of Ceirleiinema sp. Between their
maxima Pseudanahaena galeata peaks in spring and autumn. During the greater part of the year, zooplankton is almost exclusively constituted by Biac,hioriu.s arlgulai.i.s, a rather detriti-
vorous filter feeder, and its predator Acuntocyclops i~eirialis. The average annual zooplankton biomass, around 2 mg 1-1, is quite
high, but in relation to phytoplankton biomass it is very small (= 1 %).
INTRODUCTION
The Albufera of Valencia is a shallow coastal lagoon situated 12 km south of the town of Valencia, Spain (UTM 3 0 s YJ 2857).
This lacustrine system, Natural Park since 1986, was originated by the enclosure of a bay behind a beach bar 30 km in length. The bar sises at the mouth of the river Turia, its dista1 end being attached to the cliffs at the cape of Cullera. Studies realized on the continental shelf as well as calco-arenitic remains located in the bar suggest that the Holocene sediments lie on a previous Pleistocene bar. The Holocene bar has formed from sediments mainly carried by the river Turia, and it reaches a width of 500 to 1000 m. Two parallel dune systems, now covered by Mediterranean vegetation (Devesa del Sales), are developed on the bar. The
final closing of this beach bar occurred around 6000 B.P. (ROSSELLO, 1972; SANJAUME, 1985).
In Roman times, the primitive lagoon covered an area of 300 km2 according to coetaneous descriptions (Rufo Festo Avieno). Since then, the regression has been importaiit, yet it was in the second half of the 19th century when it was performed most drastically by filling 60 O/c of the lagoon then existent to gain land for rice cultivation (fig. 1 ) . Today the lagoon occupies an area of 23.2k0.1 km' (Land- sat-5). surrounded by 223 km2 of rice fields and a few small residual marshland aseas (CAVANILLES, 1795; ROSSELLO, 1972, 1976).
The Albufera, originally saltwater-filled, became progres- sively less saline when the closing process finished, leaving a lagoon independent of the sea. At the end of the 18th century. hydraulic works associated with the agricultural
Liinnciica, 8: 87-100 (1992)
O Asociación Española de Liriiiiologia. Madrid. Spain

.-a RICE AEIJS. 1761 --- ALBupeRb. 1761 - ALE- 1947 Um& 1970
Figure 1. Engraving of the Albufera ot Valencia at ihe eiid o l the 18th cciitui'y fi-otii CAVANILLES (179.5) (left). Outline of the rice fields in 1761 and the Albufera lagoon in 1761 and at present (1947, 1970) from ROSSELLO (1972) (right).
land uses not only increased freshwater intlow to the lagoon but also outflow through a channel which was opened as an artificial outlet to the sea, Pujol Vell. This channel, dug in 1762 and reconstructed several times after 1862, became definitely inoperative at the end of the 19th century (MOMBLANCH, 1960).
The increasing conversion of the old Albufera into rice fields put some of the natural water outlets, located at the southern side of the lagoon, out of service; so another two channels were opened during the first half of the 20th century (Pujol Nou and Perellonet). The 18th century brac- kish water lagoon has come to be today's oligohaline Albu- fera of Valencia. These changes in salinity were revealed by studies of sedimen) diatoms (MARGALEF & MIR, 1973) and mollusc fauna (ROBLES er al., 1985).
At present, water inflow takes place through some brooks and a series of channels dug for rice field irrigation. The lagoon is also freshwater-fed by springs located either within the lagoon or in the surrounding marshland. Water outlet goes through three main channels (Pujol Nou, Perellonet and Perelló) which link the lagoon with the sea (figs. 1 and 2), the flow being controlled by a system of sluicegates because the whole lagoon acts as a regulation reservoir in accordance with rice cultivation periods. This manipulation influences the annual cycle and the lagoon water level.
The lagoon and marshlands of the Albufera traditionally have been used for rice cultivation, fishing and hunting, but only recently the lagoon water has experienced severe chan- ges. In the 60ies the lagoon was clear to the bottom and magnificent subaquatic prairies could be seen from the surface. Human population in the drainage area was then slightly over 200,000 and industries did not number more than 500. However, in just one decade the population almost doubled in size while the industries multiplied by ten. The Albufera has been and still is the great depurative of al1 the sewage of the catchment area. Water enters the Albufera with high levels of nutrients and leaves it nutrient-free, although laden with cyanobacterial biomass.
Macrophytes have totally disappeared from the Albufera because of the extraordinary growth of plankton, which keeps off the light required for macrophyte photosynthesis. The lagoon now presents permanent blooms of non-N-fixing cyanobacteria.
Along with macrophytes, the interesting benthonic inver- tebrate fauna with severa1 endemics also has disappeared. The small endemic ciprinodont Valencia hispanica and the commercially appreciated sea bass species Dicentrarchus labras have vanished and eels are becoming scarce. Only Mugil cephalus and Cyprinus carpio are abundant at present. The important bird sanctuary that this environment

was once is only history now. Fifteen years ago, plankton formed a diverse community characteristically constituted by diatoms, although green algae also were important in summer. First data on chlorophyll a concentrations indicate that their annual mean was about 13 pg 1-1 in 1972, 25 pg 1.' in 1973 and 54 pg 1-l in 1974 (DAFAUCE 1975). Recently, however, the chlorophyll a concentrations in the lagoon water have increased to levels of up to 400 pg 1-1 and the phytoplankton has suffered the sudden general changes described in water quality deterioration, i.e., replacement of green algae by cyanobacteria in eutrophic lakes and compe- titive interaction between N-fixing and non-N-fixing cyano- bacteria in hypertrophic lakes.
Today the catchment area is heavily polluted; it includes more than 30 towns with a total of over 400,000 inhabitants and houses more than 4,000 industries of al1 types. Sewage and industrial discharges go directly into the brooks and imi- gation channels and on to the lagoon, where they produce high levels of eutrophication and seriously endanger the system.
Despite the extension and naturalistic importance of the Albufera, only a few limnological studies have been reali- zed so far. PARDO (1942) published a general monograph on the Natural History of the Albufera, summarizing the first limnological data. DAFAUCE (1975) reviews the results of a series of studies carried out during the years of 1972-74, laying emphasis on the eutrophication process which was urider way. The present paper aims at summari- zing some of the authors' recent studies on the characteris- tics of the Albufera and at offering a model of the functio- ning of this hypertrophic lagoon.
METHODS
The methodology used for obtaining the data of this paper is included in several works of the authors (SERRA et al., 1984; MIRACLE et al., 1984a, 1987; OLTRA & MIRACLE, 1984, 1992) and other publications of the scientific literature.
The water samples of the Albufera were taken at different depths by means of a 2.6 1 capacity Ruttner hydrographic bottle. Conductivity, temperature, pH, Eh and dissolved oxygen were measured "in situ" with the corresponding instruments. The major chemical parameters were determi- ned according to the methodology described by GOLTER- MAN et al. (1978). Water transparency was measured with a Secchi disk and direct measures of light penetration were taken with an underwater radiometer (PAR measurements).
The photosynthetic pigment content of the samples was evaluated spectrophotometrically, applying the equations proposed by LORENZEN (1967) and STRICKLAND & PARSONS (1972), once the algal cells had been retained in
Whatman GF/F filters and their pigments were extracted with a mixture of acetone 90% and DMSO in a 1 : 1 ratio (SHOAF & LIUM, 1976). Primary production was determined by adding 10 pCi of sodium bicarbonate I4C to transparent and opaque 125 ml bottles filled with lagoon water, which were incubated at the sampling site for one hour. After having filtered the water through Whatman GFIF filters, the carbon incorporated as particulate (POC) and dissolved (DOC) orga- nic compounds was counted in a scintillation counter (MAGUE et al., 1980; MIRACLE & VICENTE, 1985).
Lugol preserved samples of phytoplankton were observed according to the UTERMOHL (1958) technique for enume- ration.
To collect zooplankton, the contents of the 2.6 1 Ruttner bottle were filtered through a 50 pm Nytal mesh and preser- ved in 5 % formalin, to be counted afterwards with an inver- ted microscope at 100 or 200 X magnification.
The specific techniques for studying populations of aquatic bacteria can be looked up MIRACLE et a l . , (1992). Epifluorescence techniques revealing the auto- fluorescence of photosynthetic pigments (CRAIG, 1987) may be used in the counts for easily distinguishing fila- mentous or small cyanobacteria from morphologically similar organisms without chlorophyll (generally hetero- trophic bacteria).
RESULTS AND DISCUSSION
Water input
The Albufera lies at the end of a 917.1 km2 catchment area. Water flows to the Albufera from several brooks, only two of them - Massanassa and Beniparrell, which are also the most important ones - directly issuing into the lagoon in its northern part. Other running waters drain into several irrigation channels in the southerri part. Besides, the Albu- fera is supplied with groundwater springing from the surrounding marsh ("ullals") and from the bottom of the lagoon. Finally, apart from direct input by precipitation, the Albufera also is supplied with water coming from human use, i.e., irrigation water and sewage of the surrounding townships that flows through a complex system of irrigation channels. As regards irrigation water coming from the rivers Turia and Júcar, each year 321.86 Hm3 of water transferred from the Acequia Real del Júcar and 59.27 Hm3 ceded from the Acequia de Favara (river Turia) reach the Albufera or pass directly into the sea.
To evaluate the real amount of water flowing into the lagoon we have measured directly the flux in the 64 water courses (two brooks: Massanassa and Beniparrell, and 62

irrigation channels). For 1988, the total inflow to the Albu- fera was estimated to be 280 Hin3 yr-] (table 1). Despite the high number of inflow points, only a few of them are really important with regard to water input. These are particularly Overa and Dreta - which represent more than 40 % of the inflow - followed by ihe Carrera del Saler, the Alqueresía, the Fus channel and the brooks of Beniparrell rind Massa- nassa, their joint contribution to the total inflow being virtually 75 O/o (fig. 2). Considering the meteorological data of 1988, the Albufera presents a negative evaporation-preci- pitation balance, with a difference of 15 Hm3 of water evaporated and not compensated by precipitation on the surface of the lagoon. So the net water input is at least 265 H m Q r - ' , which corresponds to an average water renewal rate of ten times a year. However, large differences are observed with respect to the inflow in different sectors of tlie lagoon, making water turnover slow in extensive zones of the Albufera (once or twice a year in the northwestern
SEWAGE INPUT WATER INFLOW
Figure 2 . %oii;ll Iii~ici-ogciicii~ 01 ilic Alb~ilci :~ lago011 duc to tliffe- rences in water 1-encwal titne. Residence time (Rw). water inflow (Iw), total phosphorua input (P-t), suspended organic matter input (OM) and suspended mineral matter input (IM) into each zone are indicated. Below, pie diagrams showirig the contribution of the main northern (shaded sectors) and southern (unshaded sectors) water courses to the total water inflow and sewage input into the Albufera.
and western parts) while the sectors proximate to the outlet channels (northeast and southeast) may come up to a mean turnover time of 15 days (fig. 2). These mean values are subject to water manipulations derived from the cycle of rice cultivation. Consequently, renewal times are shorter al periods of important water flow (spring and autumn) and longer when water is being retained (winter and summer).
Hydrochemistry
Considering the typology obtained from a global study of the coastal Mediterranean wetlands of Spain (LOPEZ & TOMAS, 1989). the Albufera of Valencia would correspond to the more extreme range of the oligohaline and eutrophic type (fig. 3). The ionic proportions (in chemical equivalents) corres- pond to characteristics which are much closer to those of fresh- water than of marine water. Thus, the concentration of sulpha- tes is quite similar to that of chlorides and in many cases even higher. Ca++ is present in proportions which may equal Na+ in some instantes. Ca++ and Mg++ concentrations are normally very similar, although Ca++ presents slightly higher values in most cases. The Albufera waters correspond to the following relations: SO4= >< C1- > AIK and Na+ >< Ca++ >< Mg++ > K+.
This section summarizes the data from SERRA et al. 1984, MIRACLE et al., 1987, SORIA et al., 1987 and a technical report on the sewerage project of the Natural Park of the Albufera de Valencia (MIRACLE et ul., 1989).
Conductivity (fig. 4) has an annual cycle associated with human manipulation of the lagoon according with the rice farming cycle. There are two periods of low conductivity, which correspond to the opening of sluicegates and thus to flowing water. These periods lie in the months of March- April and September, dedicated respectively to the tilling and sowing and the h a r v e r t i n ~ of rice. In \irmrner 2nd \viriter
SO',- ALK Mg
Figure 3. Average ioriic coiiipo\iiioii ol tlir Albufera of Valencia waters as compared with other Spanish Mediterranean coastal wetlands (C) and inland waters (1). Points are defined by the equi- valent percentages of major anions and cations (H: hypersaline, P: polyhaline. M: mesohaline, 0 : oligohaline, F: freshwater). For coas- tal wetlands salinity categories (in ppt) are: C-O 5 5; C - M 5-18; C-P 18-40; C-H 2 40. Modified from LOPEZ & TOMAS (1989).
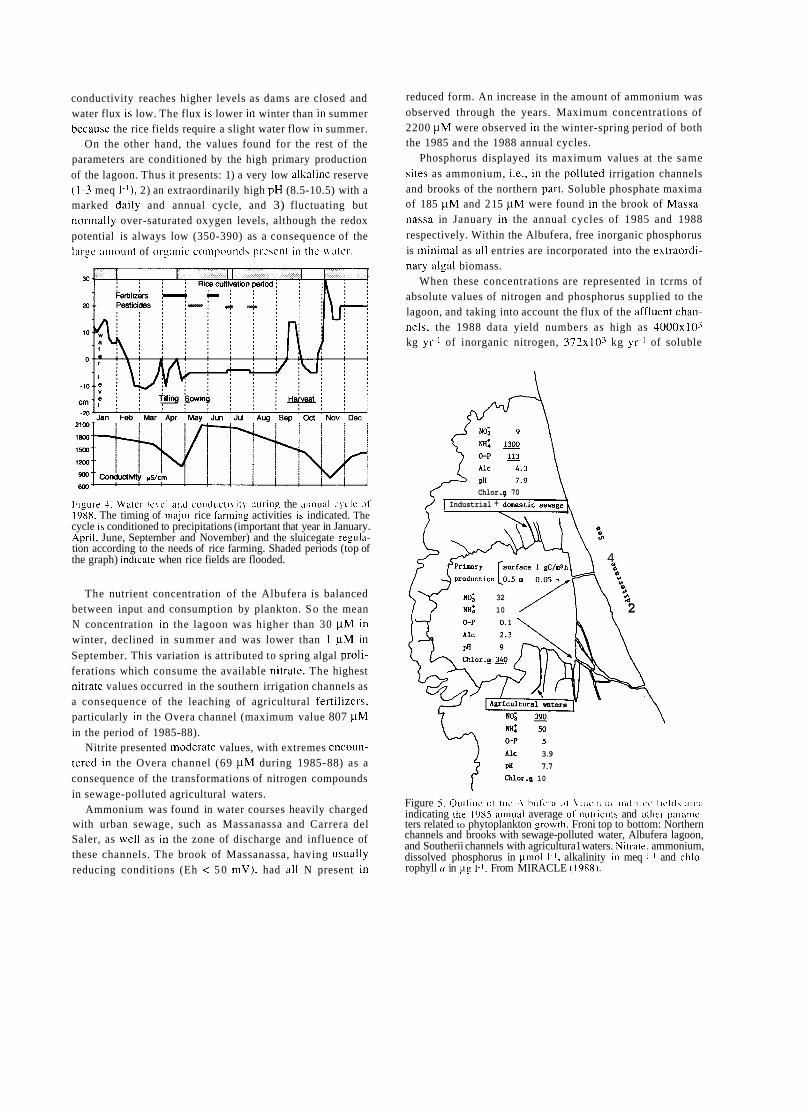
conductivity reaches higher levels as dams are closed and water flux is low. The flux is lower in winter than in summer because the rice fields require a slight water flow in summer.
On the other hand, the values found for the rest of the parameters are conditioned by the high primary production of the lagoon. Thus it presents: 1) a very low alkaline reserve (1-3 meq l.'), 2) an extraordinarily high pH (8.5-10.5) with a marked daily and annual cycle, and 3) fluctuating but normally over-saturated oxygen levels, although the redox potential is always low (350-390) as a consequence of the larre ninotiiii of o r ~ a n i c conipoiind\ prewiit i i i ihe 11 :ices.
Ftgure 1. Wniri- Ielel ancl ioiiclucii\ ity duriiig the niiti~ial c)clc 01' 1988. The timing of major rice farming activities is indicated. The cycle is conditioned to precipitations (important that year in January. April, June, September and November) and the sluicegate regula- tion according to the needs of rice farming. Shaded periods (top of the graph) indicate when rice fields are flooded.
The nutrient concentration of the Albufera is balanced between input and consumption by plankton. So the mean N concentration in the lagoon was higher than 30 p M in winter, declined in summer and was lower than 1 p M in September. This variation is attributed to spring algal proli- ferations which consume the available nitrate. The highest nitrate values occurred in the southern irrigation channels as a consequence of the leaching of agricultural fertilizers, particularly in the Overa channel (maximum value 807 p M in the period of 1985-88).
Nitrite presented moderate values, with extremes encoun- tered in the Overa channel (69 p M during 1985-88) as a consequence of the transformations of nitrogen compounds in sewage-polluted agricultural waters.
Ammonium was found in water courses heavily charged with urban sewage, such as Massanassa and Carrera del Saler, as well as in the zone of discharge and influence of these channels. The brook of Massanassa, having usually reducing conditions (Eh < 5 0 mV), had al1 N present in
reduced form. An increase in the amount of ammonium was observed through the years. Maximum concentrations of 2200 p M were observed in the winter-spring period of both the 1985 and the 1988 annual cycles.
Phosphorus displayed its maximum values at the same sites as ammonium, ¡.e., in the polluted irrigation channels and brooks of the northern part. Soluble phosphate maxima of 185 p M and 2 15 p M were found in the brook of Massa- nassa in January in the annual cycles of 1985 and 1988 respectively. Within the Albufera, free inorganic phosphorus is minimal as al1 entries are incorporated into the extraordi- nary algal biomass.
When these concentrations are represented in tcrms of absolute values of nitrogen and phosphorus supplied to the lagoon, and taking into account the flux of the afíluent chan- nels, the 1988 data yield numbers as high as 4000x10' kg yr-l of inorganic nitrogen, 372x10' kg yr-l of soluble
Chlor .a 70 Industrial + domestic seuag -
U
4
NO; 32
NH: 10 2
i Alc 3.9
PH 7.7
Chlor .a 10
Figure S. Ou~liiic 0 1 i l ic . .\lhiilci;i ol l.ilciici:i ,iiicl I I L . C 11~,1d\ aic~i indicating the 1985 aniiual average of tiutricnts and otlicr pararne- ters related to phytoplankton growth. Froni top to bottom: Northern channels and brooks with sewage-polluted water, Albufera lagoon, and Southerii channels with agricultura1 waters. Nitrate, ammonium, dissolved phosphorus in ymol l. ' , alkalinity in meq 1- ' and chlo- rophyll u in pg 1.'. From MIRACLE (1988).
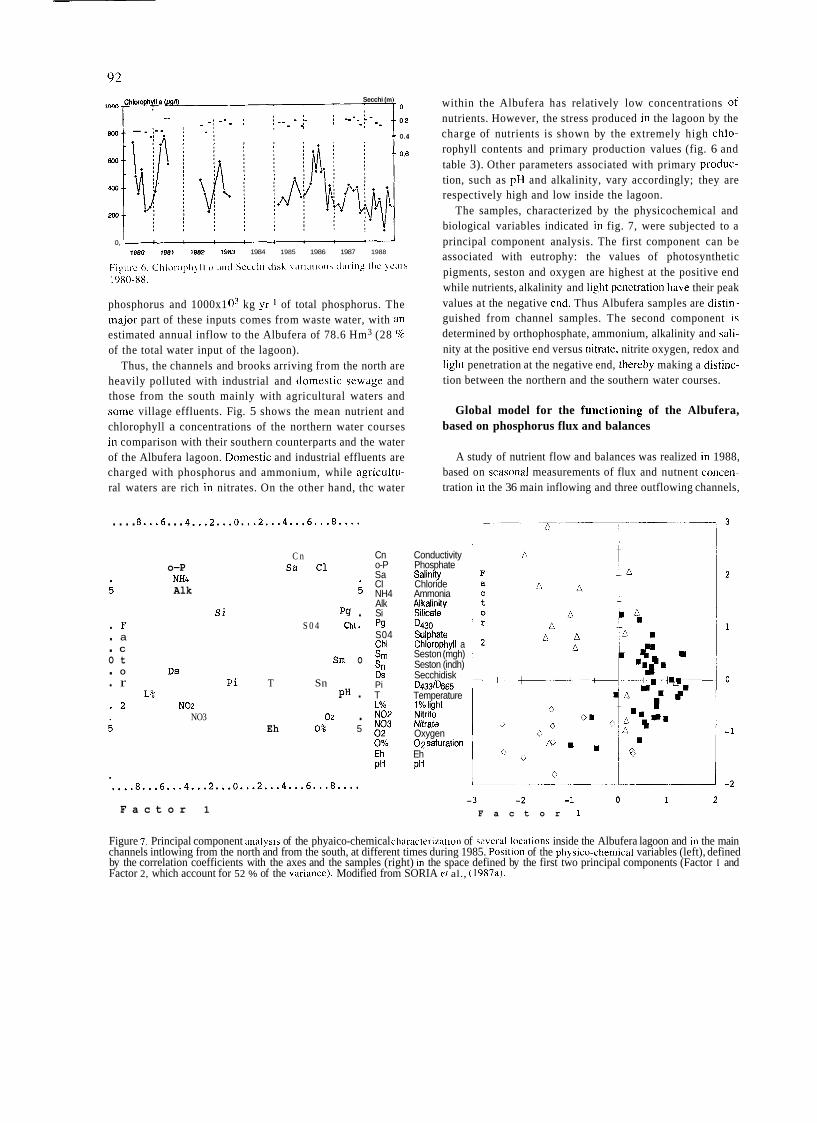
Secchi (m O
I -- I
b
I _ a _ - I I _ I - 0
8 W . . - . : - - ; . : , - 7
-- 0.4
- - 0.6
m --
o, 1980 1981 1882 1983 1984 1985 1986 1987 1988
phosphorus and 1000x1 0' kg yr-1 of total phosphorus. The major part of these inputs comes from waste water, with an estimated annual inflow to the Albufera of 78.6 Hm3 (28 % of the total water input of the lagoon).
Thus, the channels and brooks arriving from the north are heavily polluted with industrial and domestic sewage and those from the south mainly with agricultural waters and some village effluents. Fig. 5 shows the mean nutrient and chlorophyll a concentrations of the northern water courses in comparison with their southern counterparts and the water of the Albufera lagoon. Domestic and industrial effluents are charged with phosphorus and ammonium, while agricultu- ral waters are rich in nitrates. On the other hand, thc water
.... 8...6 ... 4...2...0...2...4...6...8....
C n o-P Sa C1
NH4 ; Alk ; Si pg .
. F S 0 4 Cht. . a . C
O t Sm O . O Ds . r Pi T Sn
L% PH . . 2 NO2
NO3 0 2 . ; Eh 0% 5
F a c t o r 1
Cn o-P Sa CI NH4 Alk Si pg S04 Chl Sm Sn Ds Pi T
within the Albufera has relatively low concentrations of nutrients. However, the stress produced in the lagoon by the charge of nutrients is shown by the extremely high chlo- rophyll contents and primary production values (fig. 6 and table 3). Other parameters associated with primary produc- tion, such as pH and alkalinity, vary accordingly; they are respectively high and low inside the lagoon.
The samples, characterized by the physicochemical and biological variables indicated in fig. 7, were subjected to a principal component analysis. The first component can be associated with eutrophy: the values of photosynthetic pigments, seston and oxygen are highest at the positive end while nutrients, alkalinity and light penetration have their peak values at the negative end. Thus Albufera samples are distin- guished from channel samples. The second component is determined by orthophosphate, ammonium, alkalinity and sali- nity at the positive end versus nitrate, nitrite oxygen, redox and liglit penetration at the negative end, thereby making a distinc- tion between the northern and the southern water courses.
Global model for the functioning of the Albufera, based on phosphorus flux and balances
A study of nutrient flow and balances was realized in 1988, based on seasonal measurements of flux and nutnent concen- tration in the 36 main inflowing and three outflowing channels,
Conductivity Phosphate Salinity Chloride Ammonia Alkalinity Silicaie D430 Sulphate Chlorophyll a Seston (mgh) Seston (indh) Secchidisk D4331D665 Temperature l%light Nitrite Nitrate Oxygen 02saturation Eh PH
F a c t o r 1
Figure 7. Principal component analysia of the phyaico-chemical characteri~ation of several locations inside the Albufera lagoon and in the main channels intlowing from the north and from the south, at different times during 1985. Position of the physico-chemical variables (left), defined by the correlation coefficients with the axes and the samples (right) in the space defined by the first two principal components (Factor 1 and Factor 2, which account for 52 % of the variance). Modified from SORIA et al., (1987a).

Table 1 . Global nutrient flux and pollution. lnorganic and organic N. dissolved and particulate P and sewage input. 28 % of
the total water inflow into the Albufera of Valencia are sewage water.
N-NH,' N-NO;+NNO< N-org P-POd3- P-part Sewage
INFLOW CHANNELS
OUTFLOW CHANNELS
as well as inside the Albufera (MIRACLE, VICENTE & SO- RIA, 1989). The data of this study have been used to elaborate a model of the functioning of this ecosystem (table 1 , fig. 8).
The annual nutrient inputs of the Albufera of Valencia are extremely high because the inflowing channels receive sewage water without treatment along their way. We esti- mate that the proportion of sewage in the water that enters the Albufera is 28 % of the total water inflow. This propor- tion has been calculated by measuring the P content at the exit of the sewage drains from villages around the lagoon; this value has been used as a reference for deducing the sewage content by the P concentration at the channel mouths in the Albufera (table 1, fig. 2).
The concentration of nutrients in the Albufera water is very low, as well as its inorganic nutrient output. lnorganic
N arid P outputs are respectively 10 and 50 times less than their corresponding inputs. The entrance of particulate P is mainly due to organic matter, while the outflow is mostly constituted by phytoplankton biomass (fig. 8).
Figure 8 shows the functioning of the Albufera of Valencia based on a phosphorus balance. Phosphorus enters the Albu- fera in soluble inorganic form o ras particles of organic matter with a low proportion of alga1 cells and leaves it mainly in form of phytoplankton biomass. Aquarium experiments using Albufera water and mud demonstrated that dissolved phosp- horus added to the water remains there, the rate of deposition in the sediments being extremely low. However, particulate phosphorus may be deposited in the sediment as not recycled organic matter. On the other hand, 100 times more biomass leaves the system than enters it. This is because the lagoon
OUTPUT
WI>JER COWTEWTS
B. Biomass, fresh weight O.M. Organic matter, fresh weight O-P Disolved Phosphorous P-P Particdated Phosphorous C Organic carbon
Lagoon volume 27 Hm3
Lagoon surface 23 Km2
Water and sediemnt contents in id kg The rest of values are rates in 1d kg yr-1
Figure 8. Model ot I'u~ictioiiiii~ o1 tlic i\llitilcia ol \ :ilciici~~
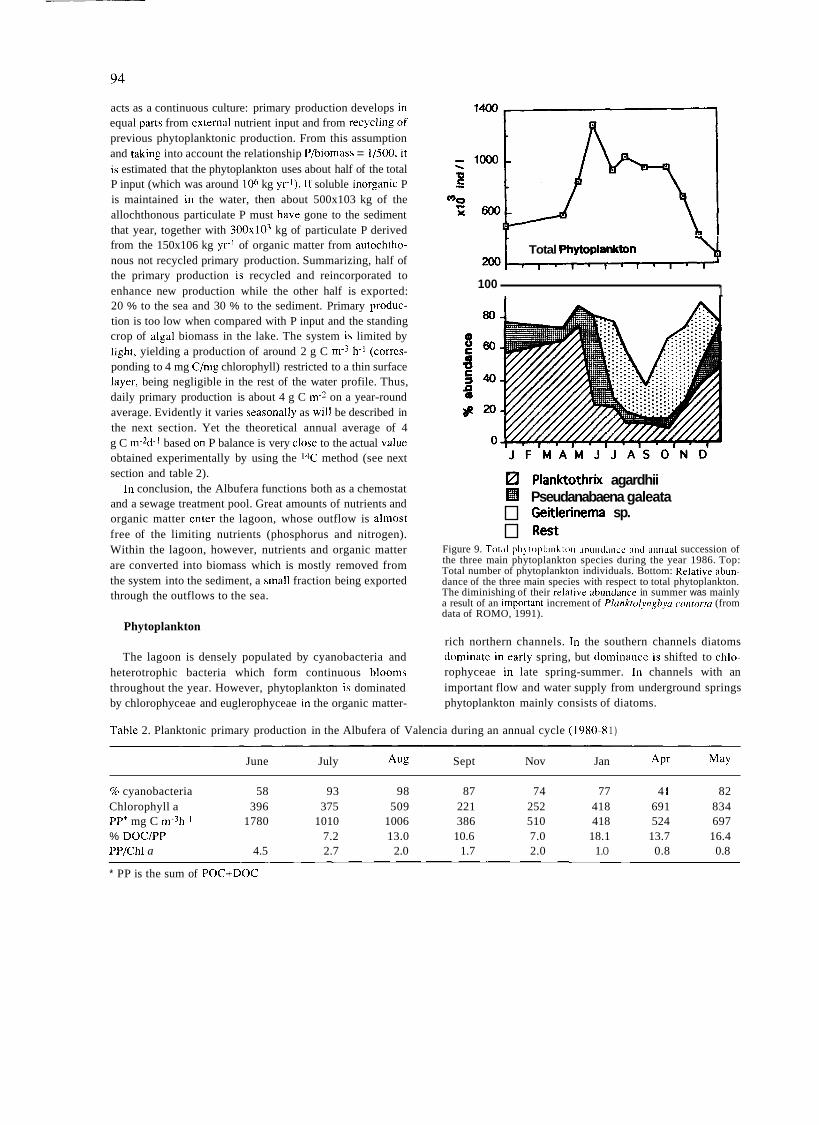
acts as a continuous culture: primary production develops in equal parts from externa1 nutrient input and from recycling of previous phytoplanktonic production. From this assumption and taking into account the relationship P/biomass = 1/500, it is estimated that the phytoplankton uses about half of the total P input (which was around 106 kg yr-l). If soluble inorganic P is maintained in the water, then about 500x103 kg of the allochthonous particulate P must have gone to the sediment that year, together with 300x103 kg of particulate P derived from the 150x106 kg yr-l of organic matter from autochtho- Total Phytoplankton nous not recycled primary production. Summarizing, half of
I
the primary production is recycled and reincorporated to enhance new production while the other half is exported:
100 1 20 % to the sea and 30 % to the sediment. Primary produc- tion is too low when compared with P input and the standing crop of alga1 biomass in the lake. The system is limited by light, yielding a production of around 2 g C m-3 h-1 (corres- ponding to 4 mg C/mg chlorophyll) restricted to a thin surface layer, being negligible in the rest of the water profile. Thus, daily primary production is about 4 g C m-2 on a year-round average. Evidently it varies seasonally as will be described in the next section. Yet the theoretical annual average of 4 g C m-2d-l based ori P balance is very close to the actual value obtained experimentally by using the I4C method (see next section and table 2).
In conclusion, the Albufera functions both as a chemostat and a sewage treatment pool. Great amounts of nutrients and organic matter enter the lagoon, whose outflow is almost free of the limiting nutrients (phosphorus and nitrogen). Within the lagoon, however, nutrients and organic matter are converted into biomass which is mostly removed from the system into the sediment, a small fraction being exported through the outflows to the sea.
Phytoplankton
The lagoon is densely populated by cyanobacteria and heterotrophic bacteria which form continuous blooms throughout the year. However, phytoplankton is dominated by chlorophyceae and euglerophyceae in the organic matter-
a Planktothrix agardhii Pseudanabaena galeata Geitlerinema sp. Rest
Figure 9. Tot~il pli) topl;iiibtoii .ihuiiil;iiici. ;iiiil aiinual succession of the three main phytoplankton species during the year 1986. Top: Total number of phytoplankton individuals. Bottom: Relative abun- dance of the three main species with respect to total phytoplankton. The diminishing of their relative abundance in summer was mainly a result of an important increment of P1anktoiyngbq.a contorta (from data of ROMO, 1991).
rich northern channels. In the southern channels diatoms dominate in early spring, but dominance is shifted to chlo- rophyceae in late spring-summer. In channels with an important flow and water supply from underground springs phytoplankton mainly consists of diatoms.
Table 2. Planktonic primary production in the Albufera of Valencia during an annual cycle (1980-8 1 )
June July Aug Sept Nov Jan A P ~ M ~ Y
% cyanobacteria 5 8 93 98 87 74 77 4 1 82 Chlorophyll a 396 375 509 22 1 252 418 69 1 834 PP* mg C m-3h-' 1780 1010 1006 386 510 418 524 697 % DOCIPP 7.2 13.0 10.6 7.0 18.1 13.7 16.4 PP/Chl a 4.5 2.7 2.0 1.7 2.0 1 .O 0.8 0.8 ---- -
* PP is the sum of POC+DOC

Tabie 3. Main phytoplankton species during 1980-88, indicating their frequency (Fr) with respect to the total number of indi-
vidual~, their occurrence (Occ) as % with respect to the total number of samples, and mean density when present (indfml).
Modified from ROMO (1991).
Species Fr Occ M e a n Species Fr Occ Mean
Cyanophyta Planktothrix agardhii 3 P.se~rdanuhaena grrleata 3 Geitlerinemu sp. 3 Arruhaerzopsis rlenkinii 2 Planktolynghya suhtilis 2 Planktolyngbya conrorta 2 Jaaginema cf. metaphyticum 2 Geitlerinema amphihium 1 Cylindrospermopsis rucihor~kii 1 Aphanothece clathrata 1 Microcystis flos-aquae 1 Merismopedia tenuissima 0sc.illatoria lanceaeformis Cholorophyta Microcystis incerta Monoraphidium grijjfithii Monoruphidiurrr corztot.tum Monoraphidium minut~tn Tetraedrom minimum Actinastrum hantzschii Chlamydomonus sp 1
Scenedesmzcs quadricaudu Scenedesmus acuminatus Scenedesmus acutus Scerzedesmus ohliquus Closteriopsis acic~tluris Chlorella vulgaris Dyctiosphuerium p ~ c l ~ hellzim Coelastrum micropor.um Choricystis rninor Bacillariophyceae Cyc'l»tellu meneghinianu Cyclotellu glomerata Nitzschia palea Nitzschia intermedia Nitzschia longissima Nitzschia gracilis Nitzschia acicularis Euglenophyceae Trachelomonus cf. \i'olvocina Cryptophyceae Cr.yptomonas erosa Rhodomonus lacustris
Freyuency values:3 >lo% 2 10 - 1% 1 I - O,l%
The phytoplankton density in the Albufera lagoon is 105 to 106 ind ml-', which corresponds to a biomass of 60-400 mg 1-' and chlorophyll concentrations of 200-600 ~g l.', with lninima during the periods of high water turnover rates (when sluicegates are opened) and maxima during the periods of stagnancy, with generally higher values in spring- summer due to the mentioned cycle in rice cultivation.
Studies on phytoplankton are only very recent (GARCIA et al., 1984; MIRACLE et al., 1984a, 1987; SORIA et al., 1987b; ROMO, 1991; ROMO & MIRACLE, in press). However, short lists of the principal phytoplankton species obtained from occasional net samples are presented in PARDO (1942) and BLANCO (1974), confirming the dramatic phytoplankton changes caused by the eutrophica- tion process. Phytoplankton has been shifting from a commu- nity dominated by diatoms and chlorophyceae (PARDO, 1942) to a eutrophic lake community still dominated by chlo- rophyceae (BLANCO, 1974) and on to the hypertrophic community of present times, absolutely dominated by fila
mentous cyanobacteria (plate 1, table 2). In the period of 1980-88 (ROMO, 1991), three dominant species accounted for more than 50 % of the total phytoplankton throughout the year, summing more than 75 % in many occasions: Plarzktothriir agardhii, Pseudanubaena galeata and Geitle- r.irzenza sp. They are virtually permanent in the plankton yet alternatively dominant according to the seasonal cycle. Planktothi-is agardhii (formerly Oscillatoria agardhii, fila- ment diameter ea. 8 pm), which primarily exhibited domi- nance throughout the winter plankton and during the last years (1986-88), is a large vacuolar oscillatorial blooming in early spring. This alga occasionally may produce floating accumulations distrihuted by wind action and water circula- tion. Pseudanuhaena galeata and Geitlerinema sp.(defined as Oscillatoria of the redekei group in our previous studies) are very thin oscillatorial species (filament diameter ca. 1-2 pm) (table 3).
These three species manifest a distinct annual succession (fig. 9). During the last years, a peak of P. agardhii appea-

I ' I ~ I I c l . hllcro~~llolcl~r:l~lll c1I ~\ll~~llci-:l (11 \ : l l ~ l l ~ l , l \ \ < l l ~ , l ~ ~ l l l l [ ~ l c \ (spring-summer 1985-86). A ) Arruhui~r7o~~~i.s (.ir1 l(/ur.i.\ (v i~~uola ted cyanobacteria with terminal heterocysts), B) P~e~rdutrohuenu ,ytrle- ata and Pla~zktotlrri.~ ugurdhii, C) Halisc~orn~nohar.ter, D ) Platrk- rothr-i.~ ugardhii, E ) Mor-ismopedia, P. ~alrrrta and Blasroc~a~r1i.s- Plancromyci~s. F) A. circiilar-is with acinetum, P. u~ardhii and Aphu- nothece clathr-ata, G) Mici.oc~~clus, P . galeuru, Blusrocuulis-P1~11c- tomycrs, H) Anculomicrohium, Huliscon7enohuctrr. P. galeutu.
red when the sluicegates were closed in April, followed by a peak of P. galeafa and then by a total dorniiiaiice of Geitlerinema throughout the summer, sometimes accompa- nied by other filamentous cyanobacteria such as L y n g h ~ a
spp (e.g. Lynghya contorna and Lynghya subtilis) and also by the characteristic tropical species Anahaenopsis c.ir-culci-
r-is, a recent colonizer (plate 1). In the autumn stagnancy period dominante of P. galeata was most common although the other two main species were also present. None of the just mentioned species had been cited before by PARDO (1942) or BLANCO (1974). Other phytoplankton groups are very scarce, chlorophyceae being second in importance with a maximum representation in spring, and diatoms (mainly Cyclotella and Nitzchia) seldom occurring and only with some incidente in late winter-early spring.
The growth of these cyanobacteria promotes a high primary production throughout the year, though restricted to the surface. The results of primary production diel experi- ments using the I4C and oxygen methods, carried out during the annual cycle of 1980-8 1, are shown in table 2. The inte- grated production in the water column varied from 2 g C m-*d-1 in winter to 7 g C m2d-1 in spring. Dark assimi- lation (DA) was also very high and fluctuated accordingly with a late spring maximum of 3 g C m-2d-1 (ca. 34 O/c of primary production) and a winter minimum of 0.2 g C m-2d-1 (ea. 1 I % of the primary production). The vertical distribu- tion of primary production and dark assimilation during a
diel cycle (30 May, 1980) is illustrated in fig. 10. The alter- native maxima of primary production (day at surface) and dark assimilation (night and day at bottom) are evident. I4C production experiments were repeated in October 1988, yiel- ding a high production for this time of the year, i.e., 1.3 g C m-'h-I at tlie surface, 0.07 g C m3h-I at 0.3 m and not detectable at 1 m of depth (measures taken in the afternoon).
Bacteria
Important populations of heterotrophic bacteria develop in the Albufera water due to its high content of dissolved orga- nic compounds (DOC), produce of algal excretions and allochthonous contributions. Other bacteria owe their presence to pollution of the lagoon with sewage water (Escherichia c,oli, Str(i)toc.oc.c~(~. Saln~or~el la , V ih i f o , etc.), studied by ALCAIDE cf (11. (1982); ALCAIDE & CARAY
PRIMARY PRODUCTION
DARK C Q UPTAKE mg C m-3 h-'
Figure 10. Diel cycle ot prriiinry pi-oductiori and C02 dark assimila- tion (experiments performed on May 29-30, 1980).
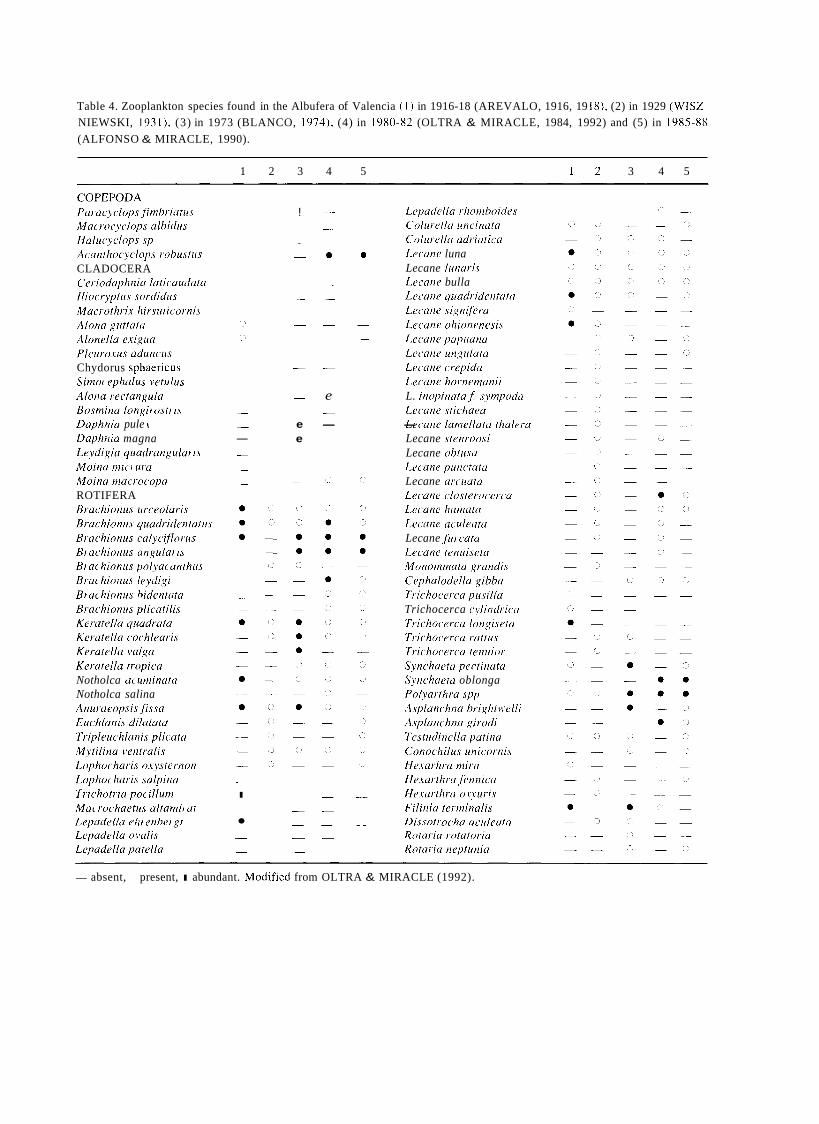
Table 4. Zooplankton species found in the Albufera of Valencia (1) in 1916-18 (AREVALO, 1916, 19 18), (2) in 1929 (WISZ- NIEWSKI, 1931), (3) in 1973 (BLANCO, 1974), (4) in 1980-82 (OLTRA & MIRACLE, 1984, 1992) and (5) in 1985-88
(ALFONSO & MIRACLE, 1990).
1 2 3 4 5 1 2 3 4 5
COPEPODA Paracyclops fimbr.iatils ! - Me~r,roc:,'r'lops ulbidirs - Hal~1c.yclops sp . -
Acanthocyclop.~ rohusrus - 0 0
CLADOCERA Ceriodrrphtiiu laric~uudata - - Iliocryptus .sor.didus - -
Macrothrix hir.sutic,ornis - - -
Lepudella r.liornboides Colur.ellu irnciriata Colurella adricrtic.~ Lecme luna Lecane 1unuri.s Leccrne bulla Lecune qrradridcntrrrrr Lecune signifera Lec,arie ohionenesis Leccrtze /~upuariu Lecarze rrn~irluta Lecarie crc.pida Lecune hoixemanii L. inopinatcr f: .~ym/~oda Lecme stichaea Lecar~c~ lan~ellata thalera Lecane .stenr.oosi Lecane obtusa Lecarie punc,tcrtu Lecane arc,uura Lc>cane closferoc~er.c,u Lecane hamata Lecme ac~~leatcr Lecane fifrc.ata Lecarie tenuiseta Monomn?atu gratidis Cephalodella gibba Tr.ichoc.erc~a pusillcr Trichocerca cylirzdric~ei Trichocerca 1otigi.seta Tr.ichoc~er(~a i.att14.s Trichocerc,cr telzrlior Synchaeta pectinatu Synchaeta oblonga Polyurrhra spp Asplanc.hncr brighti.c~lli A.sl~lutlc.ht~a girodi Te.sti~dirzellu pcrtiria Conochilris ~lnic,o~.ni.s Hescrrhra mircr He.ra1.thr.a fennic,cr Hesarthr.~ o.!-y uris Filirzia terminalis Bi.s~ot~.or.ha acrtl~uío Rofciria i.otcrtoi.icr Rotariu nepti~niu
Alona guttatrr Alonella exigua Ple~rroi~rs aduncus - - -
Chydorus sphaericus - - -
Slmoc ephalu~ vetirlu~ -
Alona i.ecfangula - e Bosminu longir octr rs - -
Daphnra pule i - e - - Daphniu magna - e Leydlgia quadrwngular 1.5 - -
Morna mrci rlra - Moinu macr.ocopa - ROTIFERA
Braclzionus angillar I\
BI ac hionils polyac anthil~ Brut hronus leydrgi Rr ac hronil~ bidentata - Brachionirs plic,atilis Ker.atellu quadr.utu Kerutella coch1ear.i~ Keratella i~al,qu Kerafella tropica Notholca acumincrta Notholca salina Anurueopsis fissa E~cc,hlarris dilatutu Tr.ipleuc~hlanis plicaru Mytilina verztralis Lopho(.haris osystei.rzon Lophoc harls salpina - - -
Tr.ichotria poc ~ l l ~ i m l - -
Mac rochaetir.r altamir ai - - - Lepadellu ehr errl7er gr - - -
Lepadellci ovulis - - -
Lepudella patella - -
- absent, present, l abundant. Modified from OLTRA & MIRACLE (1992).

(1984) MIRACLE et al. 1984b. Populations of planktonic heterotrophic bacteria not resulting from pollution are also important in the lagoon. Among these there are to be found: 1) gliding bacteria of the genera Fexihucter, Herpethosyp- hon, Achroonerna and Beggiatoa, 2 ) sheathed bacteria of the genus Haliscornenohacter, present in this lagoon because of their affinity to aquatic media with a high content of organic matter, 3) prosthecated and/or budding os appendaged bacte- ria of the genera Anculornic~rohium, Blastocczrtlis-Plan(,torny- ces, Phylomicrohi~lrn and Pasteuria, 4) besides these gene- rally infrequent forms, numerous flexuous bacteria of the genus Spil-or,kaetu, and 5) spiral and curved bacteria such as Spil-ill~tm and various species of the genera Mic~roryc1u.s and Brachyal-cus (Plate 1) (VICENTE & MIRACLE, 1988).
Zooplankton
At present, the zooplankton of the Albufera is constituted basically by two dominant species: the copepod Aunthocy- clops roh14stus and the rotifer Bruchionus angrtluris. Recent publications (OLTRA & MIRACLE, 1984, 1992; MIRACLE et al., 1988; ALFONSO & MIRACLE. 1990) show the distribution of zooplankton populations in different zones of the lagoon at different times of the year for the years 1980-88. From the quantitative analyses presented in these works it can be deduced that: (1) the zooplanktonic biomass of this lagoon is very high, on an average about 2 mg l.', corresponding roughly to 2 individuals/ml, and (2) the ratio of zooplankton to phytoplankton biomass is very low, of the order of 1 : 100; if the numbers of individuals are considered, the zooplankton/phytoplankton ratio oscillates around 2: 1,000,000. Both results are characteristic features of hyper- trophic lagoons dominated by filamentous cyanobacteria, which are not appropriate food for most zooplankton species.
The statistical study of Albufera zooplankton, from different sites and times of the year, during the annual cycle of 1982 (OLTRA & MIRACLE, 1984) points out the existente of several main associations os groups of species corresponding to seasonal succession: (a) a group consti- tuted by the permanent species Acanthoc~yc1op.s vernalis and the almost permanent Brachionus anguluris, (b) a second group, occurring at the end of summer, composed of Brachionus calyciflorus. Anul-aeopsisfis.sa, A.splun(.hna girodi and Moinu spp., and (c) a third group, occurring in autumn-winter, formed by Brachionus leydigi, Synchoeta tremula and Polyal-thra longir-eur1i.s. The importance of these groups varies greatly at the different sampling sites. Those corresponding to the northern margin of the lagoon show a predominance of rotifers over copepods and a much more marked succession in time than the rest. Furthermore, the sites stronger influenced by channel or brook flows are
í - ~ g ~ ~ r c l l . A ~ i ~ i u a l cbclc (11 llic I I I L I I I I / O O ~ I L I I ~ L I O I ~ \ ~ C C I C \ ~ I L I ~ I I I ~ 1980-81. Dcnsities (mcans of umples taken in vertical profiles at different hours) are represented by the width of the graphs accor- ding to the respective scales. ... < 1 ind. or egg pei- litre; -- 1-10 ind. or eggs per litre (from OLTRA & MIRACLE, 1992).
notably characterized, towards the end of summer, by the abovementioned second group of species with a dominance of B. c~ulyciflol-rls, which is almost inexistent in the rest of the lagoon. The same has been observed during the four annual cycles from 1985 to 1988 (ALFONSO & MIRA- CLE, 1990).
Comparison of these recent studies with former publica- tions on Albufera zooplankton makes evident that a drama- tic change in species composition has taken place. The present dominant species, A. rohustus and B. anguluris, were very rare at the beginning of the century (AREVALO, 19 16, 191 8; WISZNIEWSKI, 193 l ) , while species recorded then as the most abundant - such as Keratella quadruta, Fili- nia iei.minulis, Anurcreopsis fissa, Tric.hocerca longisetu, Ceriodaphnia luticaudatu and Sirnocephulzis vetulus - have disappeared os are much reduced (table 4).

In the quantitative study realized by BLANCO (1974, 1981), which corresponds to the years 1972-73, an impor- tant population of B. calyciflor~ls is distinguished in spring, attaining 450 ind 1.' in May, while K. q~4adratu is most outstanding in late summer and early autumn (1000 ind 1-1) and F. te~.minalis in winter and early spring (350 ind 1.'). With regard to the species most abundant at present, Blanco only mentions that B. at1gi~1ari.s and Polyarthra spp exhibit very low population densities with certain peaks (about 200 ind lb') strictly limited to come periods of the year. Blanco also describes a succession of cladoceran species, al1 of which have now almost disap- peared.
The comparison of these 1972-73 data with the annual zooplankton cycle in 1980-81 (fig. 11) confirms the drastic changes the lagoon has been suffering in recent years.
FINAL COMMENTS
The Albufera of Valencia, a shallow coastal lagoori surrounded by marshlands and formerly a complex ecosys- tem with a high biomass and species diversity, has been converted into "the great purification pool" of al1 the sewage waters of its watershed. At present, water enters the Albu- fera with high levels of nutrients and leaves it nutrient free, although laden with cyanobacterial biomass. This pheno- menon is illustrated in figs. 5 and 8.
Shallowness, turbulence and bioturbidity favour hetero- trophic metabolism in the water. The system is so stressed that nutrients and dissolved organic matter, released from the dying cells themselves, can short-cut and be used again immediately by the permanent bloom of autotrophic and mixotrophic organisms.
More than a quarter of this algal biomass, however, will not recycle but deposit as sediments and nearly another quarter will be transferred to the sea. Planktonic trophic chains have been much reduced and benthonic and littoral- benthonic communities greatly simplified. Biodiversity then has been strongly reduced, which is easily visualized in the higher organisms such as macrophytes and vertebrates. After years without apparent changes, catastrophic shifts occurred. Around the end of the sixties and beginning of the seventies the submersed macrophyte prairies vanished and planktonic communities drastically changed. Thus, the system now functions on an autotrophic (and mixotrophic) cyanobacteria and heterotrophic bacteria basis. Overcast after sunny periods, temperature changes or other factors could easily cause massive algal die-off and the delicate balance between production and respiration could be rever- sed producing a dystrophic crisis.
ACKNOWLEDGEMENTS
The authors wish to thank J.M. Soria for his aid in the preparation of the manuscript. They are also obliged to the City Council of Valencia for facilities given during the field work, and to CAICYT for a grant to M.R.M.
REFERENCES
ALCAIDE, E. & E. GARAY. 1984. R.-plasmid transfer in Salwionella spp isolated from wastewater ans sewage- contaminated surface waters. Appl. Environ. Microbiol. 48: 435-438.
ALCAIDE, E., J.P. MARTlNEZ & E. GARAY. 1982. Improved Salmor~ella recovery from moderato to highly polluted waters. .l.AppI. Buc~el-iol. 53: 143-1 46.
ALFONSO, M.T. & M.R. MIRACLE. 1990. Distribución espacial de las comunidades zooplanctónicas de la Albufera de Valencia. Scientia ,yeriinden.si.s 1 ó(2): 1 1-25.
AREVALO, C. 1916. Introducción al estudio de los cladóceros del plancton de la Albufera. Anul.lns.Gi~al.Tec. Valencia 1: 1-65.
AREVALO, C. 1 Y 18. Algunos rotíferos planctónicos de la Albufera de Valencia. Anal. Ins. Gral. Tec. Valencia 8: 1-74.
BIANCO, C. 198 1. Evolución planctónica en una zona en regresión. Guaita 8:29-33.
BLANCO, C. 1974. Estudio de la contaminución de la AlbiZfera de Valencia y de los efectos de dicha contamina- cicín sobre la.fa~4na y la flora del lago. Ph.D. thesis. Univer- sitat de Valencia, Valencia (Spain). 193 pp.
CAVANILLES, A.J. 1795. Obseri*uciones sobre la Histo- ria Natiri.al, Geog/.afía, Agricultura, Población y ,frutos del Reyno de Vu1enc.i~. Imprenta Real. Madrid (Spain).
CRAIG, S.R. 1987. The distribution and contribution of picoplankton to deep photosynthetic layers in some mero- mictic lakes. Acta Acad. Aboensis. 47: 55-8 1.
DAFAUCE, C. 1975. La Albufera de Valencia. Un estu- dio piloto. Monogruflas 4. ICONA, Madrid. 127 pp.
GARCIA, M.P., E. VICENTE & M.R. MIRACLE. 1984. Sucesión estaciona1 del fitoplancton de la Albufera de Valencia. Anales de B i o l o ~ í a M~rrcia 2 (Secc.Esp. 2 ) ; 91-100.
GOLTERMAN, H.L., R.S. CLYMO & M.A.M. OHNS- TAD, 1978. Methodsfir physi<ul and c,hernical unalysis of fresh waters. IBP Handbook n" 8. Blackwell Sci. Pub., Oxford, U.K.
LOPEZ, P. & X. TOMAS. 1989. Chemical composition of the small coastal lagoons of the Mediterranean Spanish littoral. In: Topics in marine hiology. Ros., J.D. (Ed.). Scient.Mar. 53(2-3):59 1-599.

LORENZEN, C.J. 1967. Determination of chlorophyll and pheopigments: Spectrophotometric equations. Limnol. Oceanogr.. 12: 343-348.
MAGUE, T.H., E. FRIBERG, D.J. HUGHES & 1. MORRIS. 1980. Extracellular release of carbon by marine phytoplankton: a physiological approach. Linznol. Ocea- nogr. 25: 262-279.
MARGALEF, R. & M. MIR. 1973. Indicadors de canvis de salinitat en els sediments de I'Albufera de Valencia. Treb.Soc.Cat.Bio1. 32: 11 1-1 17.
MIRACLE, M.R., 1988. Human impact on aquatic ecosystem: the "alga1 howl" (Cultural eutrophication). In: E~~i~ironmental cznd Pesticide Toxicib (Dalela, R.C., S. Kant & S. Vohsa, eds.). The Academy of Environmental Biology, India. Muzaffarnagar, India, 280 pp.
MIRACLE, M.R., M.P. GARCIA & E. VICENTE. 1 984a. Heterogeneidad espacial de las comunidades fitoplanctóni- cas de la Albufera de Valencia. Limnetica 1 : 20-3 1.
MIRACLE, M.R., M. SERRA, R. OLTRA & E. VICENTE. 1988. Differential distributions of Br.ac,hionu.s species in three coastal lagoons. Verh. Internat. Verein. Limnol. 23: 2006-20 1 5.
MIRACLE, M.R., J.M. SORIA, E. VICENTE & S. ROMO. 1987. Relaciones entre la luz, los pigmentos foto- sintéticos y el fitoplancton en la Albufera de Valencia, laguna litoral hipertrófica. Limnetica 3: 25-34.
MIRACLE, M.R. & E. VICENTE. 1985. Phytoplankton and photosynthetic sulphur bacteria production in the mero- mictic coastal lagoon of Cullera (Valencia, Spain). Verh. Internut.Verein.Limnol.22: 22 14-2220.
MIRACLE, M.R., E. VICENTE & E. GARAY. 1984b. L'Albufera de Valencia i la problematica de la contaminació de les aigües continentals costaneres. XII Congrés de Metges i Bi6logs de llengua catalana: 153-166.
MIRACLE, M.R., E. VICENTE & C. PEDROS-ALIO, 1992. Biological studies of Spanish meromictic and strati- fied karstic lakes. Limnetica 8.
MIRACLE, M.R., E. VICENTE & J.M. SORIA. 1989. Plan de saneanziento integral del Parque Natural de la Albu- ,fercl. Informe Técnico. COPUT - Generalitat Valenciana.
MOMBLANCH Y GONZALEZ, F.P. 1960. Histor.ia de lo Albufera de Valencia. Exmo. Ayuntamiento de Valencia.
OLTRA, R. & M.R. MIRACLE. 1984. Comunidades zooplanctónicas de la Albufera de Valencia. Limnetica 1: 51-61.
OLTRA, R. & M.R. MIRACLE. 1992. Seasonal succes- sion of zooplankton populations in the hypertrophic lagoon Albufera of Valencia (Spain). Arch.Hydrobio1. 124:187-204.
PARDO, L. 1942. La Alb~ffer-rr de Valencia. Biología de las aguas continentales 1I.Inst.Forest.Inv.Exp. Madrid ns 24. 263 pp.
ROBLES, F., M.A. COLLADO & V. BORREDA. 1985. Variaciones de la fauna de moluscos en la Albufera de Valencia: implicaciones paleogeográficas. In: Geomor:folo- gía litoral y cuater.nario. Homenaje a Juan Cuerda. Univer- sidad de Valencia: 123-133.
ROMO, S. 1991. Estudio delfitoplancton de la Albufera de Valencia, una laguna hipertrdfica y somera, entr-e 1980 y 1988. Ph.D thesis. Univ. de Valencia (Spain). 197 pp.
ROMO, S. & M.R. MIRACLE. Annual and seasonal periodicity of Planktothris agardhii, Pseudanabaena gulr- ata and Geitlerinema sp in a shallow hypertrophic lagoon, the Albufera of Valencia (Spain). Arch.Hydrobio1 (in press).
ROSELLO, V.M.; 1972. Los ríos Júcar y Turia en la génesis de la Albufera de Valencia. Cuad. de Geogr. 11: 7-25.
ROSSELLO, V.M. 1976. Evolution recente de I'Albufera de Valencia et ses environs. Mecliterranée 4: 19-30.
SANJAUME, E. 1985. Las costas valencianas. Sedimen- tologíu y morfología. Sección de Geografía. Universidad de Valencia, Valencia. 505 pp.
SERRA, M., M.R. MIRACLE & E. VICENTE. 1984. Interrelaciones entre los principales parámetros limnológi- cos de la Albufera de Valencia. Limnetica 1: 9- 19.
SHOAF, W.T. & B.N.. LIUM. 1976. Improved extraction of chlorophyll a and b from algae using dimethyl-sulpho- xide. Limnol. Oceanogr. 21: 926-928.
SORIA, J.M., M.R. MIRACLE & E. VICENTE. 1987a. Aporte de nutrientes y eutrofización de la Albufera de Valencia. Limnetica 3 : 227-242.
SORIA, J.M., M.R. MIRACLE & E. VICENTE. 1987b. La Albufera de Valencia: comunidades fitoplanctónicas en la campaña 1982-83. Act. VI Simp. Nac,.Bot.Cript.: 165.172.
STRICKLAND, J.D.H. & T.H. PARSONS. 1972. A prac- tical handbook of seawater analyses (2nd ed.) Bull.Fish.Res. Bd. Canada. 167 pp.
UTERMOHL, H. 1958. Zur Vervollkommnung der quan- titativen Phytoplankton-Methodik. Mitt.Int. Ver. Limnol. 9: 1-38.
VICENTE, E. & MIRACLE, M.R. 1988. Estructura y función de los procariotas en dos ecosistemas lagunares costeros: la Albufera de Valencia y el Estany de Cullera. Actas del Congreso de Biología Ambiental (11 Congreso Mundial Vusc.o) 1 : 79-107.
WISZNIEWSKI, J. 1931. Sur quelques rotiferes trouvés en Espagne. Arch.Hydrohio1. et Ichtiol., Varsovia 6 : 1-41.

RICEFIELDS, A LIMNOLOGICAL PERSPECTIVE
E. Forés and F. A. Comín Department of Ecology, University of Barcelona, Diagonal 645, 08028 Barcelona, Spain
Keywords: Ricefields, Microsuccession, Nitrogen, Phosphorus, Landscape, Global ecology.
ABSTRACT
Ricefields are extremely important ecosystems because they provide food for 40 % of the human populations and extend
over a large area (l50.106 ha), mostly in SE Asia although rice is cultivated between 50 ON and 40% al1 around the World.
Different aspects of the ecology of flooded ricefields are discussed in order to emphasize some unifying limnological perspec-
tives for ricefields. Rice paddies are agroecosystems which remain flooded during the growing season (April-September in Europe) and dry
during the rest of the year while they are prepared for the next cultivation period. In the ricefields of the Ebro Delta (N.E. Spain), the rice straw buried in the sediment contributes up to 33 and 8 % respectively of the total nitrogen and phosphorus
provided by the farmers. Chemical fertilizers are also applied at rates of 110 kg N ha-' and 20 kg P ha-'. The main nutrient
outputs are the rice plants (70% and 91% of total N and P, respectively), water outflows (15% and 3%, respectively) and bioge-
ochemical processes. Primary production exceeds total respiration in the ricefield water column during the first half of the cultivation period (from
flooding until rice tillering, about two months). During this period high oxygen concentrations (7-12 mg 1-1) and pH (8-9.3), and
low alkalinities (0.9-1.2 meq 1-1) are observed. Respiratiori by living organisms and accumulated organic matter exceeds primary
production during the second half of the cultivation period. Low oxygen concentrations (1.7-4.7 mg 1.') and pH (6-7.8). and high alkalinities (4-7.3 meq 1-1) are characteristic of this period. These environmental changes are paralleled by a community change
in the water column, from a dominance of free living organisms (phytoplankton, small filamentous algae, rotifers, cladocerans)
to that of macrophytic vegetation and detritivores.
Ricefields act as ecological filters because outflowing water contains less suspended and dissolved material than inflowing
waters. Total nitrogen and phosphorus retention efficiencies for al1 the cultivation period in the Ebro Delta ricefields
were,respectively, 35 and 75 %. Physical and biogeochemical processes are responsible for this filtering role of ricefields.
In areas where rice is cultivated extensively a change at landscape scale occurred, from relatively unpredictable natural areas
to controlled water flows and increased residence times. Establishment of a gradient of ecosystems at different organizational levels is recommended in order to alleviate impacts on connected aquatic ecosystems. Ricefields are also sites of considerable
gass (Coz and N20) emission to the atmosphere. Limnological research on ricefields should be enhanced in order to link theo-
retical aspects (ecosystem functioning) and applied aspects (food production, nature conservation).
INTRODUCTION
Rice cultivation is the most important agricultura1 acti- vity in the world, by its extension (150.106 ha) and the number of people whose basic food is rice (40% of the human population). Most of the rice is cultivated in SE Asia (90 5% of the world ricefield area) and in flooded lands (85% of the total ricefield area) (DE DATTA, 1986).
Temperature requirements for rice growth limits its
extension to between 53 ON and 40 O S (SWAMINATHAN,
1984). Rice is mostly cultivated in lands flooded to a depth of 15-25 cm. Rice development takes between 140 and 170 days. In Mediterranean countries the growing season runs from mid spring to early autumn. The characteristics of the soils in alluvial plains formed in the deltas of the large rivers (Nile, Danube, Po, Rhone, Ebro) are excellent for growing a
Limnetica, X: 1 0 1 - I O Y (19921
O Asociación Española de Limnología, Madrid. Spain


by the farnier (FORES & COMIN, 1988). In the ricefields in the Ebro Delta 110 kg ha-' of nitrogen and 20 kg ha-¡ of phosphorus are also applied as chemical fertilizers by the
farmers. These amounts, together the amount provided by
the straw, contitute 98 % of the total nitrogen and phospho- rus inputs to the ecosystem. Outputs are distributed between rice plants (70% N, 91% P), submerged macrophytes (1.4%
N, 5% P), water outflows (15% N, 3% P), and biogeoche-
mical processes (FORES & COMIN, 1989). Light and nutrients, are as in other ecosystems, the key
factors controlling the strong competition between popula-
tions occuring in the rice fields. Rice energy productivity (the amount of product obtained per unit of input energy) is
quite low in USA ricefields (0.082 kg MJ-') compared to other agricultura1 crops such as corn (0.18 kg MJ-1) and
sorghum (0.1 1 kgMJ-') (PIEROTTI et al., 1977). The rela-
tionship between any output factor (output energy, yield)
and any input factor (input energy, amount of used fertili- zer) is asymptotic (fig. 2), as in other agricultural systems
(PIMENTEL, 1974). Thus, ricefields are subject to the same
general ecological constraints as any other ecosystem. They
can be considered agroecosystems in virtue of the fact that humans control their functioning to obtain an agricultural
product.
.IZER PARTIAL
Y APPLICATION ENERGY
R ATE PRODUCTIVITY
LOW INTERMEDIATE y1 Xl
HlGH y2 1 x 2
y3 x3
FERTlLlZER ENERGY INPUTS Figui-e 2. A~)iiipioiic iclaiioiirliip hc.i\\c~.ii ilic. ~ i e l d 'iiitl riiergy input in agroecosystems showing the decreasing ratio of the effi- ciency (yield/energy input) as the inputs increase.
RICEFIELD COMMUNITY STRUCTURE
Ricefield are highly dynamic ecosystems where physical
and chemical characteristics of the water change during the cultivation period. These changes are caused by the increa-
sing complexity of the biological community during the growing season.
In spite of the overwhelming dominance of rice biomass
due to human effects, plankton (called heleoplankton in such a shallow water column) and floating and submerged macrophytes are apparent during the first phases of the rice growth, when the light is sufficient to permit photosyntesis
ir1 the water column. Phytoplankton develops abundant populations until the tillering phase. We have observed phytoplankton abundantes between 5,600 and 19,000 cell ml-1 in the Ebro Delta ricefields (mainly chlorophyceans and
diatoms), generally entering the ricefields with the irrigation
water. During the first phases of rice growth, the chemical characteristics of the water in the ricefields show clearly a dominance of production over respiration processes: high
values of dissolved oxygen (7- 12 mg.1-1) and pH (8-9.3) and
low values of alkalinity (0.95-1.2 meq.1-1 (PONT &
VAQUER, 1984: ANTONIETTI e? al, 1987; FORES & COMIN 1987). Rotifera and Cladocera are the dominant components of the heleoplankton (FERRARI et a1.,1984:
FORES & COMIN, 1986).
Phytoplankton growth decreases after the tillering phase. During this period, submerged vegetation develops dense populations: filamentous algae (Spvrogyra, Oedogonium,
Hvdrodyction), rooted macrophytes (Scirpus, Ncrjas, Zanni-
chelia, Potanzog~~ron), and Charu (VAQUER, 1984b, FORES & COMIN, 1986). Chara is the most abundant genus in the ricefields, and reaches maximum biomasses of
1,000 Kg ha-] in French ricefields (VAQUER, 1984). During the tillering phase the copepod Acunthocyclops g. i~erna1i.s displaces rotifers and cladocerans, forming the most
abundant zooplankton population (PONT, 1984, FORES & COMIN, 1986).
Rice reaches its maximum height and biomass in the reproductive and ripenning periods. Radiation intensities
reaching the upper part of the water column are less than
10% of the incident light at the top of the rice plants (fig. 3) (PONT & VAQUER, 1984). Submerged vegetation can develop between rice plants, if there is enough light. Ostra- coda, with a large number of extraeuropean species (Dole- i.oc.ypris sinensis, Stenoc.ypris malcolmsoni, Cypris subglo-
hosa,) becomes the most abundant heleoplankton group, with some summer copepoda as Thermocyc,lops dyhowsky or
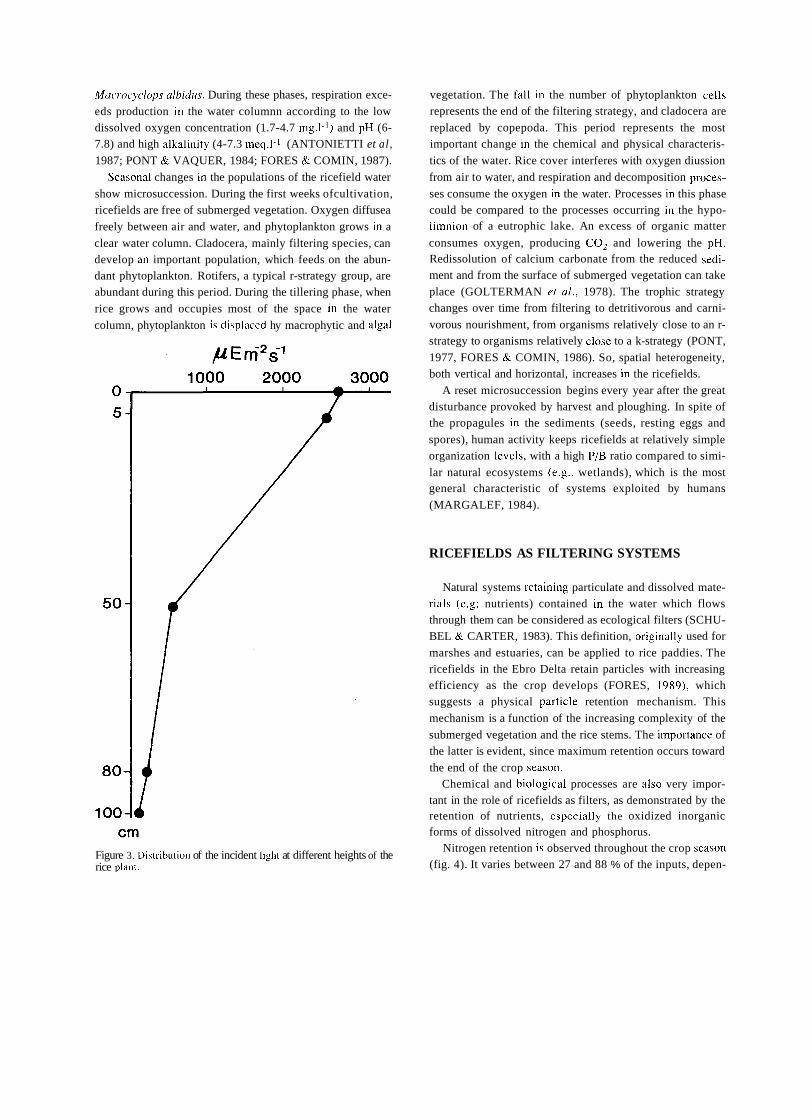
Macr-ocyc~lops ulbid~ls. During these phases, respiration exce-
eds production in the water columnn according to the low dissolved oxygen concentration (1.7-4.7 mg.1-I) and pH (6- 7.8) and high alkalinity (4-7.3 meq.1-1 (ANTONIETTI et al,
1987; PONT & VAQUER, 1984; FORES & COMIN, 1987).
Seasonal changes in the populations of the ricefield water
show microsuccession. During the first weeks ofcultivation, ricefields are free of submerged vegetation. Oxygen diffusea
freely between air and water, and phytoplankton grows in a
clear water column. Cladocera, mainly filtering species, can develop an important population, which feeds on the abun- dant phytoplankton. Rotifers, a typical r-strategy group, are abundant during this period. During the tillering phase, when
rice grows and occupies most of the space in the water
column, phytoplankton is di\placcd hy macrophytic and algal
Figure 3. Distribution of the incident light at different heights of the rice plant.
vegetation. The fall in the number of phytoplankton cells represents the end of the filtering strategy, and cladocera are
replaced by copepoda. This period represents the most important change in the chemical and physical characteris-
tics of the water. Rice cover interferes with oxygen diussion
from air to water, and respiration and decomposition proces-
ses consume the oxygen in the water. Processes in this phase could be compared to the processes occurring in the hypo- limnion of a eutrophic lake. An excess of organic matter
consumes oxygen, producing COZ and lowering the pH. Redissolution of calcium carbonate from the reduced sedi- ment and from the surface of submerged vegetation can take
place (GOLTERMAN rt al., 1978). The trophic strategy changes over time from filtering to detritivorous and carni-
vorous nourishment, from organisms relatively close to an r- strategy to organisms relatively close to a k-strategy (PONT,
1977, FORES & COMIN, 1986). So, spatial heterogeneity, both vertical and horizontal, increases in the ricefields.
A reset microsuccession begins every year after the great disturbance provoked by harvest and ploughing. In spite of the propagules in the sediments (seeds, resting eggs and
spores), human activity keeps ricefields at relatively simple organization levels, with a high P/B ratio compared to simi-
lar natural ecosystems (e.g., wetlands), which is the most general characteristic of systems exploited by humans
(MARGALEF, 1984).
RICEFIELDS AS FILTERING SYSTEMS
Natural systems retaining particulate and dissolved mate- rials (e.g: nutrients) contained in the water which flows through them can be considered as ecological filters (SCHU- BEL & CARTER, 1983). This definition, originally used for
marshes and estuaries, can be applied to rice paddies. The ricefields in the Ebro Delta retain particles with increasing efficiency as the crop develops (FORES, 1989), which suggests a physical particle retention mechanism. This
mechanism is a function of the increasing complexity of the
submerged vegetation and the rice stems. The importance of the latter is evident, since maximum retention occurs toward
the end of the crop season. Chemical and biological processes are also very impor-
tant in the role of ricefields as filters, as demonstrated by the retention of nutrients, especially the oxidized inorganic forms of dissolved nitrogen and phosphorus.
Nitrogen retention is observed throughout the crop season (fig. 4). It varies between 27 and 88 % of the inputs, depen-
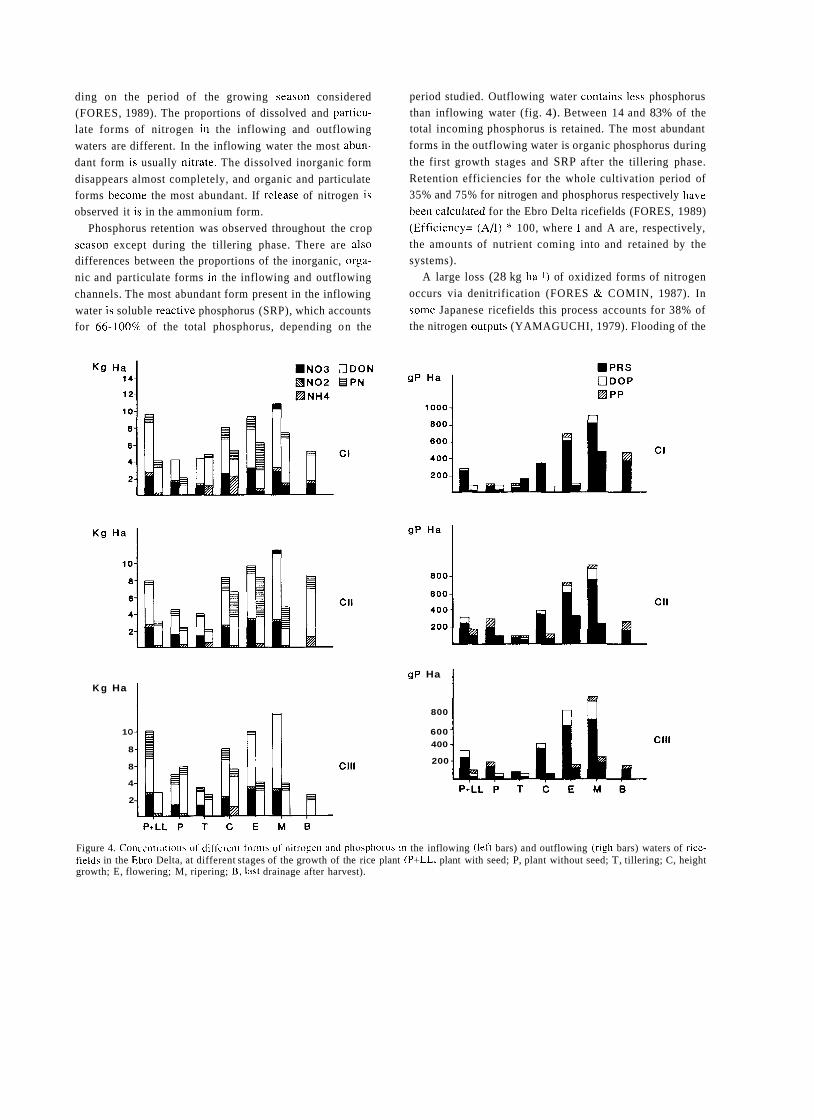
ding on the period of the growing season considered (FORES, 1989). The proportions of dissolved and particu- late forms of nitrogen in the inflowing and outflowing
waters are different. In the inflowing water the most abun-
dant form is usually nitrate. The dissolved inorganic form
disappears almost completely, and organic and particulate forms become the most abundant. If release of nitrogen is observed it is in the ammonium form.
Phosphorus retention was observed throughout the crop
season except during the tillering phase. There are also differences between the proportions of the inorganic, orga-
nic and particulate forms in the inflowing and outflowing
channels. The most abundant form present in the inflowing
water is soluble reactive phosphorus (SRP), which accounts for 66.100% of the total phosphorus, depending on the
period studied. Outflowing water contains less phosphorus than inflowing water (fig. 4). Between 14 and 83% of the total incoming phosphorus is retained. The most abundant
forms in the outflowing water is organic phosphorus during
the first growth stages and SRP after the tillering phase.
Retention efficiencies for the whole cultivation period of 35% and 75% for nitrogen and phosphorus respectively have
been calculated for the Ebro Delta ricefields (FORES, 1989)
(Efficiency= (A/I) * 100, where 1 and A are, respectively, the amounts of nutrient coming into and retained by the systems).
A large loss (28 kg ha- ') of oxidized forms of nitrogen
occurs via denitrification (FORES & COMIN, 1987). In
some Japanese ricefields this process accounts for 38% of the nitrogen outputs (YAMAGUCHI, 1979). Flooding of the
CII
gP Ha K g Ha
800
1 O 600
400 Clll 8
8 cl l l 200
4
2
Figure 4. Coiiceiiti~itioii\ ol' ditlei-e111 toriiis of iiiti-ogeii aiid plio\plioi.us in the inflowing (left bars) and outflowing (righ bars) waters of rice- fields in the Ebro Delta, at different stages of the growth of the rice plant (P+LL, plant with seed; P, plant without seed; T, tillering; C, height growth; E, flowering; M, ripering; B. last drainage after harvest).

soil results in the displacement of air spaces by water, and the dissolved oxygen present in the pore water is readily consumed during microbial respiration. Supply of oxygen to the flooded soil occurs by difussion through the overlying
water column, and transport through the stems of the rice
and other wetland plants to the roots, with subsequent diffu-
sion into the rizhosphere, which can support aerobic micro- bial populations. The rest of the soil is anaerobic. Microor- ganisms responsible for denitrification have facultative
anaerobic metabolism, but require nitrates, an oxidized form of nitrogen present in aerobic conditions. So, ricefields, with
aerobic-anaerobic interphases between sediment and water and the rizosphere, offer adequate conditions for denitrifi-
cation. The fertilizers used contain ammonium sulphate or urea as nitrogen source. Ammonium present in soil solution is subject to movement in two directions: upward into the
oxidized soil layer at the interface soil-water column and
downward the plant roots, where i t is nitrified by aerobic
microorganisms (fig. 5). Nitrates can reach the anaerobic zone where denitrifiers use them. The extent of nitrogen losses through nitrification-denitrification reactions in floo-
ded soils is dependent on the supply of ammonium to the
oxidized zones where nitrification potentially occurs. So, slow-release fertilizers can contribute to a decrease in the nitrogen losses due to this process, and increase the oxygen
demand in the root zone by increasing the organic content
of the soil (REDDY & PATRICK, 1986).
difference is that nutrient recycling in ricefields is accelera-
ted by man after the harvest. The incorporation of rice straw and fertilizers into the sediments maintains the ricefields at very high levels of productivity if enough water is availa-
ble. The system is maintained at the same stage year after
year with the energy invested by man. In contrast, natural wetlands processes occur more slowly and self-organization of the system produces successional changes in the commu- nity structure in the long -term.
RICEFIELDS AT LARGER SCALES
Ammonium from fertilizers, such as ammonium sulphate
and urea, can be volatilized as ammonia. The extent of vola- tilization depends mainly on the ammonium concentration. the pH and the urease activity of the flooding water; rates
as high as 2 1 C/o of the nitrogen added were recorded in Italy (PEDRAZZINI & TARSITANO, 1986).According to the characteristics of the Ebro Delta ricefields, this process could be an important quantitative loss of nitrogen during
the first phases of the cultivation period (FORES &
COMIN, 1986). Inorganic phosphorus precipitation can explain the obser-
ved retention of this element by the ricefields (FORES, 1989). Mediterranean ricefield sediment and water have a
high calcium concentration (MINZONI, 1986, FORES & GONZALEZ, 1988). Calcium and phosphorus can combine in different forms to give insoluble compounds that remain
in the sediments (GOLTERMAN, 1984). Chemical and physical characteristics of the flood water during the first
phases of the rice growth favor phosphorus precipitation, but the presence of an important anaerobic layer under the water-sediment interface may favor phosphorus release to
the water. Bacteria1 metabolism can explain a fraction of
this phosphorus release (MINZONI, 1986). So, ricefields, like many fresh- and salt-marshes, play a
role as filters Sor sediments and nutrients, which accumulate
mainly as organic matter in the soil. Rice biomass accumu-
lated during the growing season is similar to those of the most productive emergent macrophytes (e.g., Phragmites
communis, Typha latifolia, WETZEL, 1983). The main
Figure 5 . Scheniatic repreaciii~ii~oii o l pi-oceases al'fecting nitrogen transformations in the soot mne of rice.
Although ricefields are ecological filters, their outflows
still contain a huge amount of nutrients. They are responsi-
ble for accelerated eutrophication of the coastal lagoons in the Ebro Delta. Most of the rice in the world is cultivated in low lands, formerly or potentially wetlands. Land reclama- tion Sor extensive rice cultivation is a landscape transfor- mation which involves huge limnological changes.

Water must be supplied in huge amounts (between 104 and 50. 10, mm?ha-l), usually from rivers, in order to obtain high production. But water is commonly a scarce resource in the regions and during the time rice is cultivated. Compe- tition for water uses among direct human use, agriculture and industry arises in regions such as Catalonia as socio- economic development proceeds. Moreover, water is not readily available for use. A dense and complex network of irrigation and drainage channels is necessary. These new artificial aquatic ecosystems can be colonized by exotic populations and used for the propagation of species.
Channels also have their own metabolism. In the Ebro Delta drainage channels 25% of the total nitrogen inputs are lost along the network, while total phosphorus increases in the channel 50% (FORES, 1989). Nutrient changes in the channels depend on the flow rates and the type and amount of substrate and living organisms (EL HABR & GOLTER- MAN, 1987).
The hydrologic regime of coastal lagoons in the Ebro Delta is controlled by the water drained from the rice field area. The consequences are an inverse seasonal fluctuation of salinity compared to similar Mediterranean coastal lago- ons (COMIN et al., 1987, COMIN et al., in press) and intense eutrophication related rates of turnover of fresh and seawater (COMIN et al., 1990).
In the controversy between wetland conservation versus land reclamation for rice cultivation, a gradient of ecosystems at different organization levels (wetlands) is recommended instead of excessive water flow regulation as in the Ebro Delta, to alleviate impacts on connected aquatic ecosystems.
The extremely short water column and high organic matter content of sediments in ricefields favor the export to the atmosphere of gaseous forms of carbon, nitrogen an sulphur origiiiating in the predominantly anoxic conditions of ricefield sediments (PONNAMPERUMA, 1972, SAVANT, & DE DATTA, 1982). Thus, rice fields, like wetlands in general, are a source of gasses like CO,, CH,, NH,, N*, N20, SH,, which reach the atmosphere across the water column or through the rice plants (SCHUTZ et al.. 1989). Once in the atmosphere, these compounds and, after chehical transformations, their derivatives, contribute to global changes (climatic, acidification).
Some of the processes occur faster in ricefields than in other environments, e.g.: methane production (VALIELA, 1985). However, extrapolation of rates to rice fields in gene- ral is risky because of the variety of soils used for rice culti- vation (DE DATTA, 1986). One aspect is important because it is quite common: organic matter content is high in ricefield
sediments. Furthermore, the amount of chemical fertilizer used is increasing al1 around the world. These two factors contribute , in general, to enhance microbial activities.
So, limnological reseach of rice fields can contribute to an understanding of ecological processes which operate at different spatial scales. Moreover, linked to agronomic methods, they can contribute to a better production of a basic food, particularly in tropical countries. The link between theoretical and applied aspects of science is complete in the case of ricefield limnology.
ACKNOWLEDGMENTS
This work was supported by grants from EEC (EV4V- i 32-E) and CICYT ((NAT89-844).
REFERENCES
ANTONNIETI, R., P. VIAROLI & F. ANTONIETTI, 1987. Changes in the physical and chemical properties of flooded water and sediment in an experimental ricefield (Reggio Emilia,Italy). Hydi-oh. 144:83-88.
BARCELOJ., G. NICOLAS, B. SABATER & R. SANCHEZ. 1980. Fisiologia vegetal. Piramide Ed. Madrid.
750 pp. BERCZIK, A., 1972. Periodical changes in aspect of the
zoocenoses of Hungarian rice-fields. Verh. Internar. Verein. Limnol. 18: 1742- 1747.
BOLOS, O. & F. MASCLANS, 1975. La vegetación de los arrozales en la región mediterránea. Collect. Bo f .
4(3):415-434. BONETTO, C., F. MINZONI & H. L. GOLTERMAN,
1988. The nitrogen cycle in shallow water sediment system of ricefields. 2 Fractionation and bioavailability of organic nitrogen compounds. Hydrohiol. 159:203-2 10.
BURESH, R. J. & S. K. De DATTA, 1990. Denitrifica- tion losses from puddled rice soils in the tropics. Biol. Fertil. Soils. 9: 1-13.
CHINCHILLA, M. & F.A. COMIN, 1977. Contribució al coneixement dels crustacis del Delta de L'Ebre. Treb. Irlst.
Car. Hist. Na f . 8: 119- 144. COMIN, F.A. & E. FORES, 1989. Els arrossars. Historia
Natural dels Pai'sos Catalans 14:340-345. COMIN, F.A., M. MENENDEZ & E. FORES, 1987. Sali-
nidad y nutrientes en las lagunas costeras del Delta del Ebro. Limnetica 3: 1-8.

COMIN, F.A., M. MENENDEZ & J.R. LUCENA, 1990. Proposals for macrophyte restoration in eutrophic coastal lagoons. Hydrobiologia 200/201:427-436.
COMIN, F.A., E. FORES, M. MENENDEZ & V. ASTORGA, in press. Approach to the characterization of Mediterranean deltaic ecosystems at different scales. In: Ecology of the Po Delta. Italy, Sept. 1990.
De DATTA,S.K. 1986. Producción de arroz.Fundamen-
tos y prácticas. Limusa Ed. Mexico. 690pp. EL HABR, H. & H.L. GOLTERMAN, 1987. Input of
nutrients and suspended matter into the Golfe du Lion and The Camargue by the River Rh6ne. Rev. Sci. Eau 6 (4):393-402.
FERNANDO, C. H., 1977. Investigation on the aquatic fauna of tropical ricefields with special reference to South East Asia. Geo-Eco-Trop. 3:169-188.
FERNANDO, C. H., J. L. FURTADO & R. P. LIM, 1980. The ecology of ricefield with special refrence to the aquatic fauna. Trop. Ecol. & Develop. 19:943-95 1.
FERRARI I., A. de MARCHI, P. MENOZZI, F. MINZONI & F. PICCOLI. 1984. Heleopankton seasonal succession in an experimental ricefield in northern 1taly.Verh. Intern. Verein. Limnol. 22: 171 1-1716.
FERRER, X. & F.A. COMIN, 1979. Distribució i ecolo- gía dels macrofits submergits del Delta de L'Ebre. Butll. Inst. Cat. Hist. Nat. 44(3): 11 1-1 17.
FORES, E. 1989. Ricefields as filters. Arch. Hydrob. 116 (4): 517-527.
FORES, E. & F.A. COMIN. 1986. Características limno- lógicas de los arrozales del Delta del Ebro (N.E. España). Oecol. aquut. 8:39-45.
FORES, E. & F.A. COMIN. 1987. Chemical characteris- tics of the water in the ricefields of the Ebro delta (N.E. Spain). Arch. Hydrob. 11 l(1): 15-24.
FORES, E. & F.A. COMIN, 1989a. Balances de nitró- geno y fósforo en arrozales del Delta del Ebro. Bol. R. Soc. Esp. Hist. Nat. 84 (3-3):247-257.
FORES, E., M. MENENDEZ & F.A. COMIN, 1987. Contribución al conocimiento de crustáceos y rotíferos del Delta del Ebro. Misc. Zool. 10: 105-1 11.
FORES, E. & F.A. COMIN, 1988b. Liberación de C, N y P durante la descomposición de la paja del arroz. Medite- rranea 10: 13-18.
FORES,E. & F.A. COMIN. 1989b. Contenidos de nitró- geno, fósforo y carbono del arroz en diferentes estadios de crecimiento. Rev.Agroq.Tec. Alim. 29(4): 492-500.
FORES,E. & S. GONZALEZ. 1988. Heterogeneidad espacio-temporal del sedimentode los arrozales del Delta del Ebro. (N.E. España). Oecol. aquat. 9: 125- 135.
FORES, E., M. MENENDEZ & F.A. COMIN, 1988. Rice straw decomposition in rice-field soil. Plant and Soil 109: 145-146.
FOX, H.M., 1965. Ostracod crustacea from ricefields in Italy. Mem. Ist. Ital. Idrob. 18:205-214.
FRENEY, J. R., A. C. F. TREVITT, S. K. De DATTA, N. OBCEMEA & J.G. REAL, 1990. The interdependence of ammonia volatilization and denitrification as nitrogen loss processes in flooded rice fields in the Philippines. Biol. Fertil. Soils 9:3 1-36.
GARCIA,J.L. 1973. Séquence des produits formés au cours de la dénitrification dans les sols des rizikres du Sene- gal. Ann. Microb. 124:35 1-362.
GOLTERMAN, H. L., 1982. Preliminary observations on nutrient cycles in Scirpus and rice fields in The Camargue. In J.J. Symoens, S.S. Hooper & P. Compkre (eds.). Studies on aquatic vascular plants. Royal Botanical Sopciety of Belgium. Brussels, 200-201.
GOLTERMAN, H. L. 1984. Sediments, modifying and equilibrating factors in the chemistry of freshwater. Verh. Intern. Verein Limnol. 22:23-59.
GOLTERMAN H.L., R.S. CLYMO & M.A.M. OHNS- TAD. 1978. Method for physical and chemical analysis of fr-eshwater-s. IBP Handbook 8. Blakwell Ed. Oxford. Edin- bourgh. 214pp.
GREGORY, P.J., D.V. CRANFORD & M. McGOWAN. 1972. Nutrient relations of winter wheat.1. Accumulation and distribution of Na, K, Ca, Mg, P, S and N. J. Agric. Sci. Camb. 93:485-494.
MADONI, P., 1988. Distribution and seasonal succession of ciliated Protozoa in a ricefield ecosystem: A three-year study. Verh. Internat. Verein. Limnol. 23:1063-1067.
MARGALEF, R., 1984. Ecología. Ed. Omega, Barcelona, 95 1 págs.
MINZON1,F. 1986. Studio di processi legati ai cicli dell'azoto e del fosforo nell'ecosisterna risaia. Tesi dotto- rato di ricerca. Parma.Italia. l45pp.
MINZONI, F. & A. MORONI, 1987. Phosphorus balance of an experimental ricefield in the Po plain (Italy). Agric. Ecosis. & Evironm. 18:223-230.
MINZONI, F., C. BONETTO & H.L. GOLTERMAN, 1988. The nitrogen cycle in shallow water sediment system of ricefields. 1 The denitrification process. Hydrobiol. 159: 189-202.
MORONI, A., 1961. L'ecosistema de risaia. Ann. Fac. Agraria dell'U.C.S.C., 3:489-625.
MUKHAMEDIEV, A.M., 1960. Contribution to the hydrobiology of the ricefields of the Fergana Valley. Sci. Notes. Uzbek State Pedagogical Inst. Fergana (Biol.) 6:1-82.

PEDRAZZINI, F.R. & R. TARSITAN0.1986. Ammonia volatilization from flooded soil following urea application. Plant & Soil91:lOl-107.
PIEROTTI, A., A.C. KELLER & A.J. FRITSCH, 1977. Energy and food. CSPI Energy Series X. Centre for Scien- ces in the Public Interest. Washington D.C.
PONNANPERUMA, F.N., 1972. The chemistry of submerged soils. Adv. Agron. 24:29-96.
PONT,D. 1977. Structure et évolution saissonikre des populations de copépodes, cladoceres et ostracodes des rizieres de Camargue.Ann.Limno1. 13(1): 15-28.
PONT,D. 1984. Recherches quantitatives sur I'héléo- plancton des rizieres de Camargue. These Doct. Sci. Nat. Provknce (Marseille) 300 pp.
PONT,D. & A. VAQUER. 1984. Quelques caractéristi- ques physico-chimiques de l'eau de submersion des rizikres de camargue (France): r6le de la végétation inmergée et émergée. Ecol. Medit. lO(1-2): 133-145.
REDDY, K.R. & W.H.Jr. PATRICK. 1986. Denitrifica- tion losses in flooded ricefields.In: Nitrogen Economy of flooded rice soils. 99-116. S.DE DATTA & W.H. PATRICK Jr. Ed. Dordrecht. 186pp.
REDDY, K. R., W. H. PATRICK Jr. & C. W. LINDAU, 1989. Nitrification-denitrification at the plant root-sediment interface in wetlands. Limnol. Oceanogr. 34(6):1004-1013.
ROSSI, O., A. MORONI, P. BARONI & P. CARAVE- LLO, 1974. Annual evolution of the zooplankton diversity in twelve italian ricefields. Boll. Zool. 41,3.
SAVANT, N.K. & S.K. DE DATTA, 1982. Nitrogen trans- formations in wetland rice soils. Adv. Agron. 35:241-302.
SCHUBEL,R. & V.S. KENNEDY. 1983. The estuary as a filter: an introduction. In: The estuary a s a filter 1-15. KENNEDY Ed. Academic Press. Orlando. Florida.
SCHUBEL,R. & H.H. CARTER. 1983. The estuary as a filter for fine-grained suspended sediment. In: The estuary a s a filter. 81-105. KENNEDY Ed. Academic Press. Orlando. Florida.
SCHUTZ, H., W. SEILER & R. CONRAD, 1989. Proces- ses involved in formation and emission of methane in rice paddies. Biogeochem. 7:33-53.
SEILER, W., A. HOLZAPEL-PSCHORN & C. R. SCHARITE, 1984. Methane emmission from rice paddies. J. Atoms. Chem. 1:241-268.
SINGH, V.P., T.H. WICKHAM & I.T. CORPUZ. 1978. Nitrogen movement to Laguna Lake through drainage from ricefields. Internat. Conference on water Pollution Coun- tries. Bangkok. Thailand.
SWAMINATHAN, M.S. 1984. Arroz. 1nvest.Ciencia. 90:52-62.
VALIELA, I., 1985. Marine Ecological Processes. Sprin- ger-Verlag, N.Y., 543 pp.
VAQUER, A., 1984a. La production algale dans les rizik- res de Camargue pendant le pkriode de submersion. Verh. Intern. Ver. Limnol. 22:1615-1654.
VAQUER, A. 1984b. Biomasse et production des chara- cees dans les rizikres de Camargue (France) et leur impor- tance écologique. Oecol. Plant. 5(19):299-313.
VERGARA, B.S., 1985. Growth and development of the deep water rice plant. IRRI Res. Pap. Ser. 103. Internatio- nal Rice Research Institute, Los Baños. Laguna. Philippines,
38 PP. WETZEL, R.G., 1983. Limnology. Saunders College
Publ., Philadelphia, 767 págs. YAMAGUCH1,T. 1983. Biological nitrogen fixation in
flooded ricefield.In:Nitrogen and Rice 193-206.IRRI.Brady Ed. Philippines. 499pp.

PATTERNS OF VARIATION IN THE LIMNOLOGY OF SPANISH RESERVOIRS: A REGIONAL STUDY
J. L. Riera, D. Jaume, J. de Manuel, J. A. Morgui and J. Armengol Departament d'Ecologia, Universitat de Barcelona. Avgda. Diagonal, 645, 08028 Barcelona, Spain.
Keywords: Regional limnology, Multivariate analysis, Ionic composition, Nutrients, Seston, Plankton, Biogeography.
ABSTRACT
In this paper, an overview of the major patterns of variation in the limnology of Spanish reservoirs is provided, based on two regional studies carried out in 1972-76 and 1987-88. Geology and climate largely determined the ionic composition of Spanish reservoirs, whereas land uses in the cathment area seem to control nutrient concentrations. Nitrogen and phosphorus, however, presented an independent distribution. While the concentration of nitrate correlated with the mineral content of the water, phosphorus appeared to be associated with seston both during the mixing and the stratification period. Alga1 biomass was best correlated with phosphorus in summer, but the phosphorus-chlorophyll-a relationship was influenced by the alkalinity and the total nitrogen concentration of the water and, to a lesser extent, by the residence time of the water and the mean biomass of herbivorous crustaceans. Multivariate analysis of phytoplankton, rotifers and crustaceans abundance data consistently reve-
aled the significance of three factors of variation, namely, (1) the ionic composition of the water, (2) the nutrient content, and (3) the water residence time of reservoirs. Together with these ecological factors, historical processes of colonization help explain the geographic distribution of a number of taxa in the reservoirs of Spain, where other limnenic habitats are scarce. Because reservoir ecosystems are recent, colonization is still in progress, as is reflected by the expanded distribution of some
taxa in the 1987-88 survey with respect to the previous one.
INTRODUCTION
Spain is a country of low and variable water resources. Genuine lakes are almost absent: apart from Lake Sanabria, a glacial lake in NW Spain, and Lake Banyoles (NE Spain) and a few other smaller karstic lakes, freshwater bodies are only abundant in mountain ranges (Sierra Nevada, Pyrenees) where they originated from glacial activity. Small lagoons and temporal ponds, however, are profusely disseminated throughout the country (ALONSO, 1985). On the other hand, rivers, particularly in the extensive mediterranean areas of the Iberian Peninsula, show high fluctuations in discharge. While they are generally dry in summer, autum- nal storms occasionally cause them to flow in spates.
Either because of water shortages or because of hazar- dous excesses, management of water resources has always been a sorely felt necessity in Spain (PEREZ PICAZO &
LEMEUNIER, 1990). Some of the oldest reservoirs in the world that are still in use are found here: Cornalbo and Proserpina, both near the roman town of Mérida (Badajoz),
date from the 11 century, Almansa (Albacete) was first built
in the XIV century, and Tibi, among others, in the XVI century. Extensive construction of reservoirs, however, did not begin until this century, when technological develop- ments permitted it, and it has augmented steadily acompan- ying, and being stimulated by, economic progress (PEREZ PICAZO & LEMEUNIER, 1990). Today nearly 1,000 reser- voirs are in use in Spain, with a total capacity approaching 45-lo9 m3 (fig. 1). Apart from water supply, irrigation, and flood control, reservoir uses that predominate in the medi- terranean areas, many dams were built for hydropower gene- ration in mountainous areas and in the rainy northwestern region of the country.
Systematic research on Spanish reservoirs did not begin until the late sixties and early seventies, with the work of Vidal on the limnology of Sau Reservoir (NE Spain) (VIDAL 1969, 1973), and gained impetus after a regional study of one hundred reservoirs was undertaken by a rese- arch group from the Department of Ecology of the Univer- sity of Barcelona, under the direction of Ramon Margalef
Limnclica, 8: 11 1-123 (1992)
O Asociación Española de Limnología, Madrid. Spain
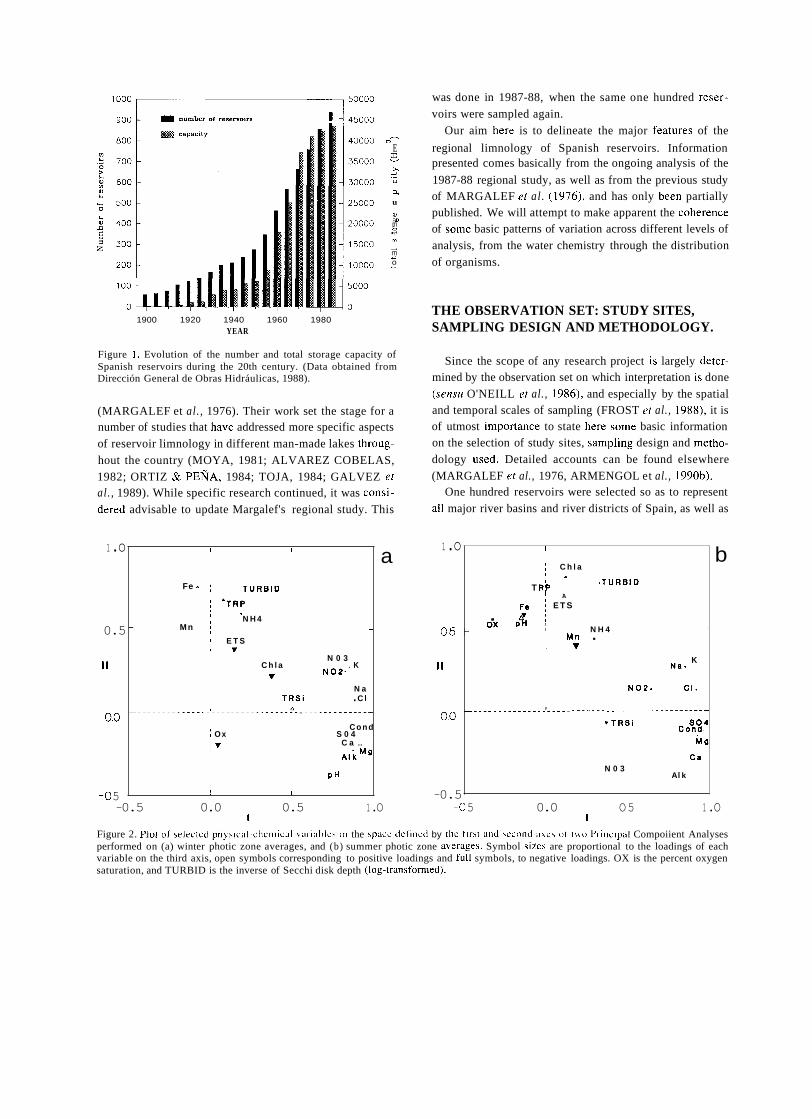
900 nurnber of reservoirs 1 4
1900 1920 1940 1960 1980 YEAR
was done in 1987-88, when the same one hundred reser- voirs were sampled again.
- Our aim here is to delineate the major features of the O
E regional limnology of Spanish reservoirs. Information 3:
presented comes basically from the ongoing analysis of the * .M
U 1987-88 regional study, as well as from the previous study P of MARGALEF et al. (1976), and has only been partially u
& published. We will attempt to make apparent the coherence m C o .e of some basic patterns of variation across different levels of "7 - analysis, from the water chemistry through the distribution m C o L-
of organisms.
THE OBSERVATION SET: STUDY SITES, SAMPLING DESIGN AND METHODOLOGY.
Figure 1. Evolution of the number and total storage capacity of Spanish reservoirs during the 20th century. (Data obtained from Since the scope of any research project is largely deter-
Dirección General de Obras Hidráulicas, 1988). mined by the observation set on which interpretation is done (sensu O'NEILL et al., 1986), and especially by the spatial
(MARGALEF et al., 1976). Their work set the stage for a and temporal scales of sampling (FROST et al., 1988), it is
number of studies that have addressed more specific aspects of utmost importance to state here some basic information
of reservoir limnology in different man-made lakes throug- on the selection of study sites, sampling design and metho-
hout the country (MOYA, 198 1; ALVAREZ COBELAS, dology used. Detailed accounts can be found elsewhere
1982; ORTIZ & PEÑA, 1984; TOJA, 1984; GALVEZ et (MARGALEF et al., 1976, ARMENGOL et al., 1990b).
al., 1989). While specific research continued, it was consi- One hundred reservoirs were selected so as to represent
dered advisable to update Margalef's regional study. This al1 major river basins and river districts of Spain, as well as
Figure 2. Plor of selectrd phq\ic;il-cliriiiicnl \ui-inblr\ i i i the spacc clctiiiecl by tlic tirst aild \ecoiici ;ixr\ ol i \ \ o I'riiicipal Compoiient Analyses performed on (a) winter photic zone averages, and (b) summer photic zone averages. Symbol sizes are proportional to the loadings of each variable on the third axis, open symbols corresponding to positive loadings and full symbols, to negative loadings. OX is the percent oxygen saturation, and TURBID is the inverse of Secchi disk depth (log-transformed).
b
-0.5 0.0 0.5 1 .O -0 5 0.0 O 5 1 .O I I
1 1
: C h l a ' A
T RF . T U R B I D
' A
F~ j E T S . f l l - OX PH N H 4
Mn . 1 7
N a . K
N 0 2 . C I .
v T R S i S 0 4 Cond.
M g
cs N 0 3
Al k
I I
1 .o a
0.5-
1 1
0.0
-0.5
1 .o
0.5
1 1
0.0
-0 5
1 1
Fe A ; T U R B I D
; ' T R P ; . N H 4
- M n j -
1 E T S i T
N 0 3 C h l a . K
N 0 2 . v N a
T R S i . C I -........-e---- .'.........-...... '. . -
C o n d l Ox S 0 4
C a .. : A I ~ " ' ~
P H
1 I
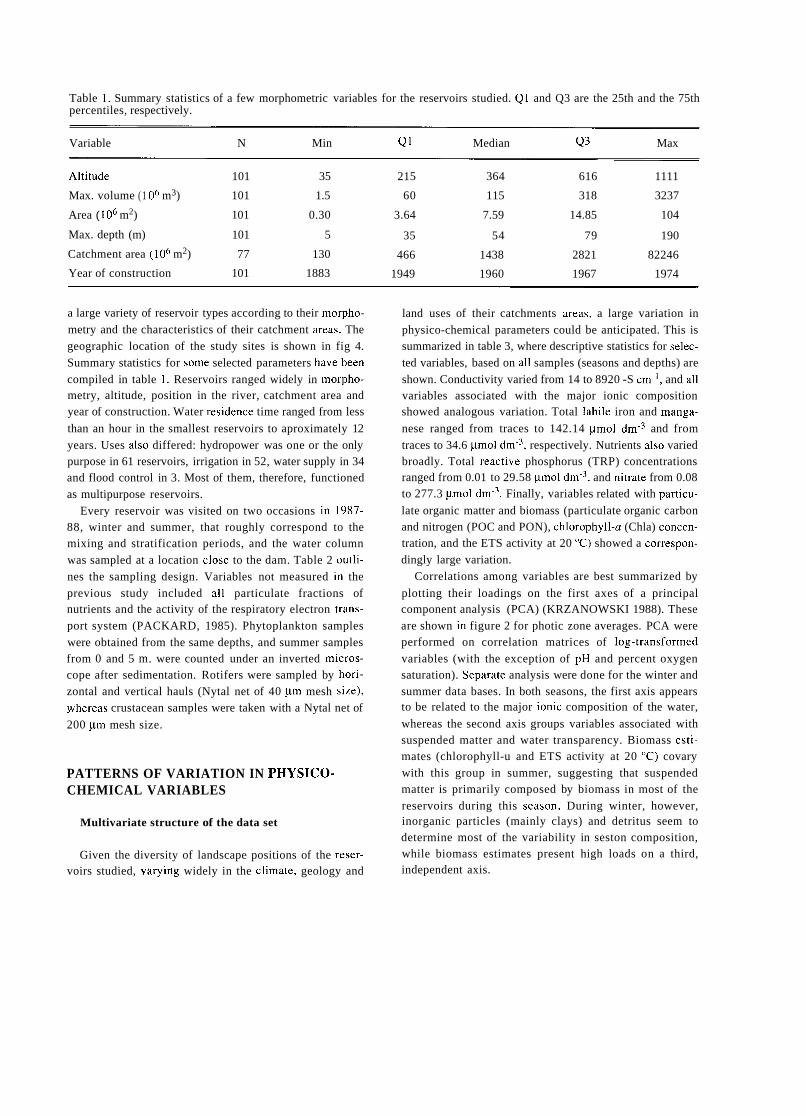
Table 1. Summary statistics of a few morphometric variables for the reservoirs studied. Q1 and Q3 are the 25th and the 75th percentiles, respectively.
Variable N Min Q 1 Median 4 3 Max
Altitude 101 3 5 215 364 616 1111
Max. volume ( 1 o6 m3) 101 1.5 60 115 318 3237
Area ( 1 O6 m2) 101 0.30 3.64 7.59 14.85 104
Max. depth (m) 101 5 35 54 79 190
Catchment area ( lo6 m2) 77 130 466 1438 2821 82246
Year of construction 101 1883 1949 1960 1967 1974
a large variety of reservoir types according to their morpho- land uses of their catchments areas, a large variation in metry and the characteristics of their catchment areas. The physico-chemical parameters could be anticipated. This is geographic location of the study sites is shown in fig 4. summarized in table 3, where descriptive statistics for selec- Summary statistics for some selected parameters have been ted variables, based on al1 samples (seasons and depths) are compiled in table 1. Reservoirs ranged widely in morpho- shown. Conductivity varied from 14 to 8920 -S cm-', and al1 metry, altitude, position in the river, catchment area and variables associated with the major ionic composition year of construction. Water residence time ranged from less showed analogous variation. Total labile iron and manga-
than an hour in the smallest reservoirs to aproximately 12 nese ranged from traces to 142.14 pmol dm-3 and from years. Uses also differed: hydropower was one or the only traces to 34.6 pmol dm-3, respectively. Nutrients also varied purpose in 61 reservoirs, irrigation in 52, water supply in 34 broadly. Total reactive phosphorus (TRP) concentrations and flood control in 3. Most of them, therefore, functioned ranged from 0.01 to 29.58 pmol dm-3, and nitrate from 0.08
as multipurpose reservoirs. to 277.3 pmol dm-3. Finally, variables related with particu-
Every reservoir was visited on two occasions in 1987- late organic matter and biomass (particulate organic carbon 88, winter and summer, that roughly correspond to the and nitrogen (POC and PON), chlorophyll-u (Chla) concen- mixing and stratification periods, and the water column tration, and the ETS activity at 20 "C) showed a correspon-
was sampled at a location close to the dam. Table 2 outli- dingly large variation.
nes the sampling design. Variables not measured in the Correlations among variables are best summarized by
previous study included al1 particulate fractions of plotting their loadings on the first axes of a principal nutrients and the activity of the respiratory electron trans- component analysis (PCA) (KRZANOWSKI 1988). These port system (PACKARD, 1985). Phytoplankton samples are shown in figure 2 for photic zone averages. PCA were were obtained from the same depths, and summer samples performed on correlation matrices of log-transformed from 0 and 5 m. were counted under an inverted micros- variables (with the exception of pH and percent oxygen cope after sedimentation. Rotifers were sampled by hori- saturation). Separate analysis were done for the winter and zontal and vertical hauls (Nytal net of 40 pm mesh size), summer data bases. In both seasons, the first axis appears whereas crustacean samples were taken with a Nytal net of to be related to the major ionic composition of the water,
200 pm mesh size. whereas the second axis groups variables associated with suspended matter and water transparency. Biomass esti- mates (chlorophyll-u and ETS activity at 20 T) covary with this group in summer, suggesting that suspended PATTERNS OF VARIATION IN PHYSICO-
CHEMICAL VARIABLES
Multivariate structure of the data set
Given the diversity of landscape positions of the reser- voirs studied, varyirig widely in the climate, geology and
matter is primarily composed by biomass in most of the reservoirs during this season. During winter, however, inorganic particles (mainly clays) and detritus seem to determine most of the variability in seston composition, while biomass estimates present high loads on a third, independent axis.
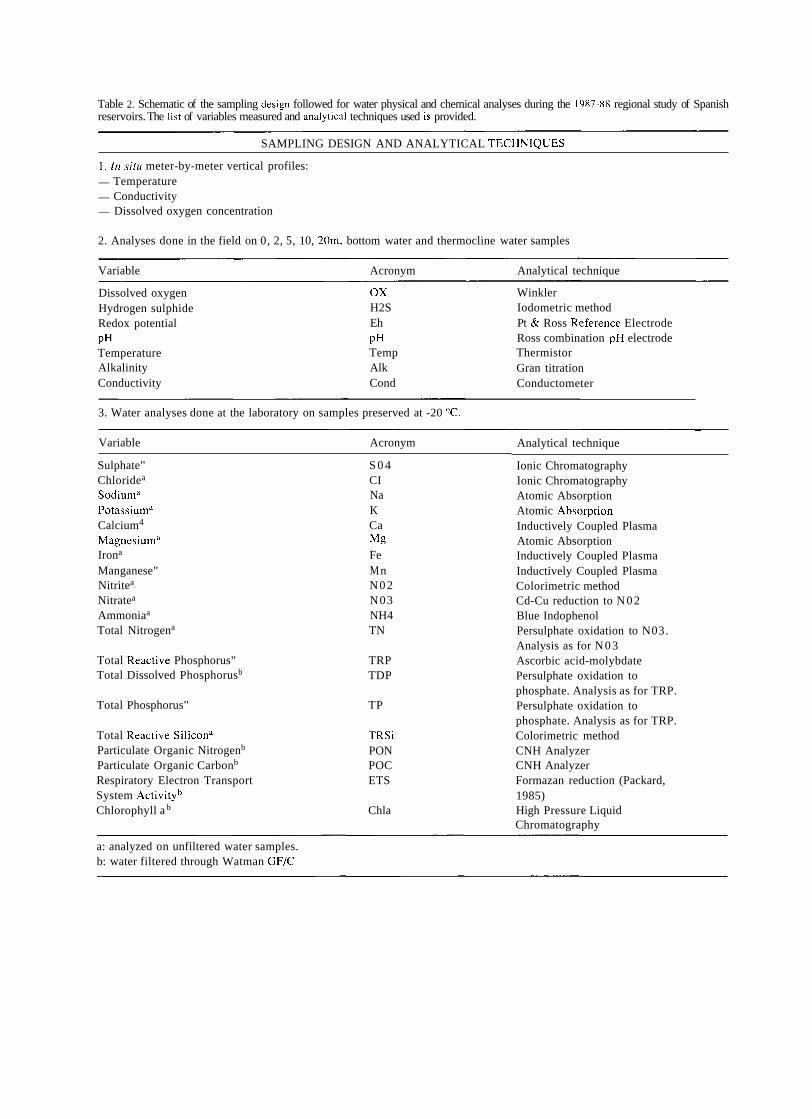
Table 2. Schematic of the sampling design followed for water physical and chemical analyses during the 1987-88 regional study of Spanish reservoirs. The list of variables measured and analytical techniques used is provided.
SAMPLING DESIGN AND ANALYTICAL TECHNIQUES
1. In situ meter-by-meter vertical profiles: - Temperature - Conductivity - Dissolved oxygen concentration
2. Analyses done in the field on 0, 2, 5, 10, 20m, bottom water and thermocline water samples
Variable Acronym Analytical technique
Dissolved oxygen Hydrogen sulphide Redox potential
PH Temperature Alkalinity Conductivity
OX H2S Eh
PH Temp Alk Cond
Winkler Iodometric method Pt & Ross Reference Electrode Ross combination pH electrode Thermistor Gran titration Conductometer
3. Water analyses done at the laboratory on samples preserved at -20 T.
Variable Acronym
Sulphate" S 0 4 Chloridea CI Sodiuma Na Potassiuma K Calcium4 Ca Magnesiuma Mg Irona Fe Manganese" M n Nitritea N 0 2 Nitratea N03 Ammoniaa NH4 Total Nitrogena TN
Total Reactive Phosphorus" Total Dissolved Phosphorusb
TRP TDP
Total Phosphorus" TP
Total Reactive Silicon" Particulate Organic Nitrogenb
Particulate Organic Carbonb
Respiratory Electron Transport System ~ c t i v i t ~ ' Chlorophyll a b
TRSi PON POC ETS
Chla
-
Analytical technique
Ionic Chromatography Ionic Chromatography Atomic Absorption Atomic Absorption Inductively Coupled Plasma Atomic Absorption Inductively Coupled Plasma Inductively Coupled Plasma Colorimetric method Cd-Cu reduction to N 0 2 Blue Indophenol Persulphate oxidation to N03. Analysis as for N 0 3 Ascorbic acid-molybdate Persulphate oxidation to phosphate. Analysis as for TRP. Persulphate oxidation to phosphate. Analysis as for TRP. Colorimetric method CNH Analyzer CNH Analyzer Formazan reduction (Packard, 1985) High Pressure Liquid Chromatography
a: analyzed on unfiltered water samples. b: water filtered through Watman GFIC
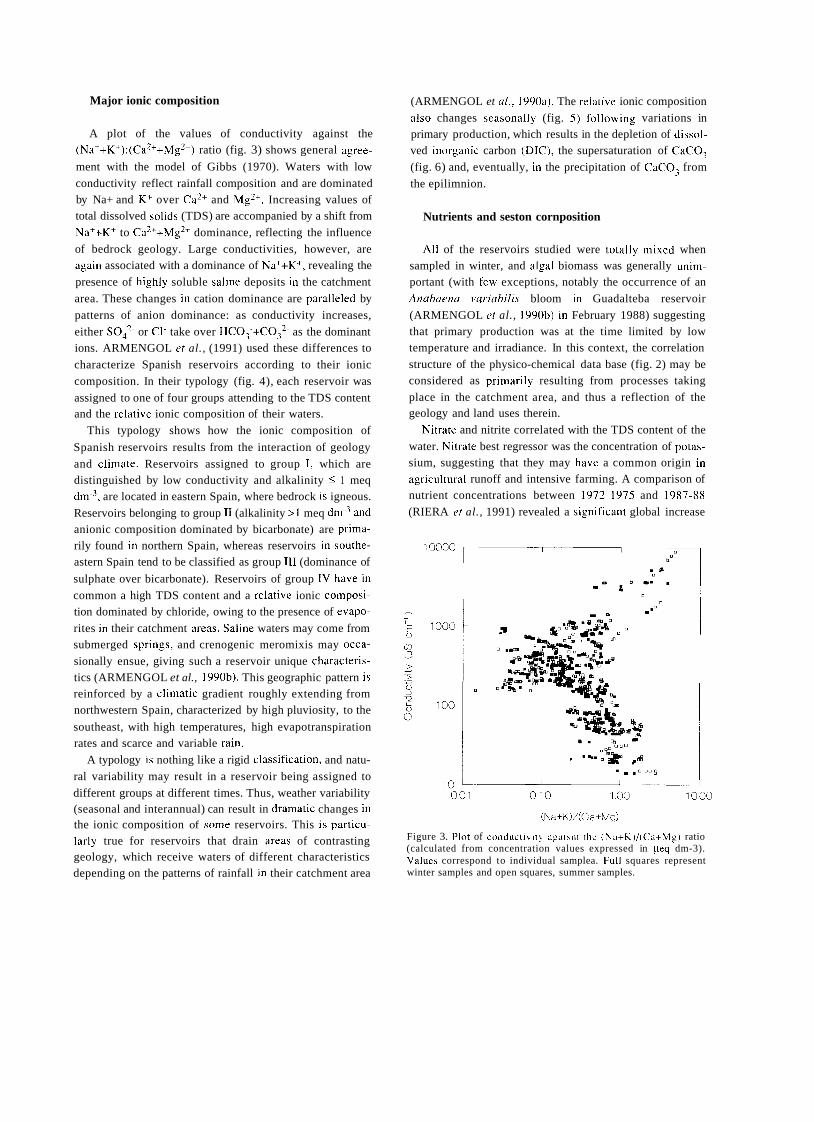
Major ionic composition
A plot of the values of conductivity against the ( N a + + ~ + ) : ( c a ~ + + ~ g * + ) ratio (fig. 3) shows general agree-
ment with the model of Gibbs (1970). Waters with low
conductivity reflect rainfall composition and are dominated by Na+ and K+ over ca2+ and Mg2+. Increasing values of total dissolved solids (TDS) are accompanied by a shift from
Na++Kf to C a 2 + + ~ g 2 + dominance, reflecting the influence
of bedrock geology. Large conductivities, however, are again associated with a dominance of Na++K+, revealing the
presence of highly soluble saline deposits in the catchment
area. These changes in cation dominance are paralleled by
patterns of anion dominance: as conductivity increases,
either SO,'. or C1- take over HCO~-+CO~*- as the dominant ions. ARMENGOL et al., (1991) used these differences to
characterize Spanish reservoirs according to their ionic
composition. In their typology (fig. 4), each reservoir was
assigned to one of four groups attending to the TDS content and the relative ionic composition of their waters.
This typology shows how the ionic composition of
Spanish reservoirs results from the interaction of geology
and climate. Reservoirs assigned to group 1, which are distinguished by low conductivity and alkalinity á 1 meq dm-3, are located in eastern Spain, where bedrock is igneous.
Reservoirs belonging to group 11 (alkalinity >1 meq d m - b n d anionic composition dominated by bicarbonate) are prima- rily found in northern Spain, whereas reservoirs in southe- astern Spain tend to be classified as group 111 (dominance of
sulphate over bicarbonate). Reservoirs of group IV have in
common a high TDS content and a relative ionic composi- tion dominated by chloride, owing to the presence of evapo-
rites in their catchment areas. Saline waters may come from submerged springs, and crenogenic meromixis may occa-
sionally ensue, giving such a reservoir unique characteris- tics (ARMENGOL et al., 1990b). This geographic pattern is reinforced by a climatic gradient roughly extending from northwestern Spain, characterized by high pluviosity, to the
southeast, with high temperatures, high evapotranspiration rates and scarce and variable rain.
A typology is nothing like a rigid classification, and natu-
ral variability may result in a reservoir being assigned to
different groups at different times. Thus, weather variability (seasonal and interannual) can result in dramatic changes in the ionic composition of some reservoirs. This is particu-
larly true for reservoirs that drain areas of contrasting geology, which receive waters of different characteristics depending on the patterns of rainfall in their catchment area
(ARMENGOL et al., 1990a). The relative ionic composition
also changes seasonally (fig. 5 ) following variations in primary production, which results in the depletion of dissol- ved inorganic carbon (DIC), the supersaturation of C a c o 3
(fig. 6) and, eventually, in the precipitation of C a c o 3 from
the epilimnion.
Nutrients and seston cornposition
All of the reservoirs studied were totally mixed when sampled in winter, and algal biomass was generally unim- portant (with few exceptions, notably the occurrence of an
Ancrhaena var-iahilis bloom in Guadalteba reservoir
(ARMENGOL et al., 1990b) in February 1988) suggesting that primary production was at the time limited by low temperature and irradiance. In this context, the correlation
structure of the physico-chemical data base (fig. 2) may be considered as primarily resulting from processes taking
place in the catchment area, and thus a reflection of the geology and land uses therein.
Nitrate and nitrite correlated with the TDS content of the
water. Nitrate best regressor was the concentration of potas- sium, suggesting that they may have a common origin in agricultura1 runoff and intensive farming. A comparison of nutrient concentrations between 1972-1975 and 1987-88
(RIERA et al., 1991) revealed a significant global increase
Figure 3. Plot of cotiducii\ 11) ;iga1\111 IIic (Ya+K)/(Ca+Mg) ratio (calculated from concentration values expressed in peq dm-3). Values correspond to individual samplea. Full squares represent winter samples and open squares, summer samples.
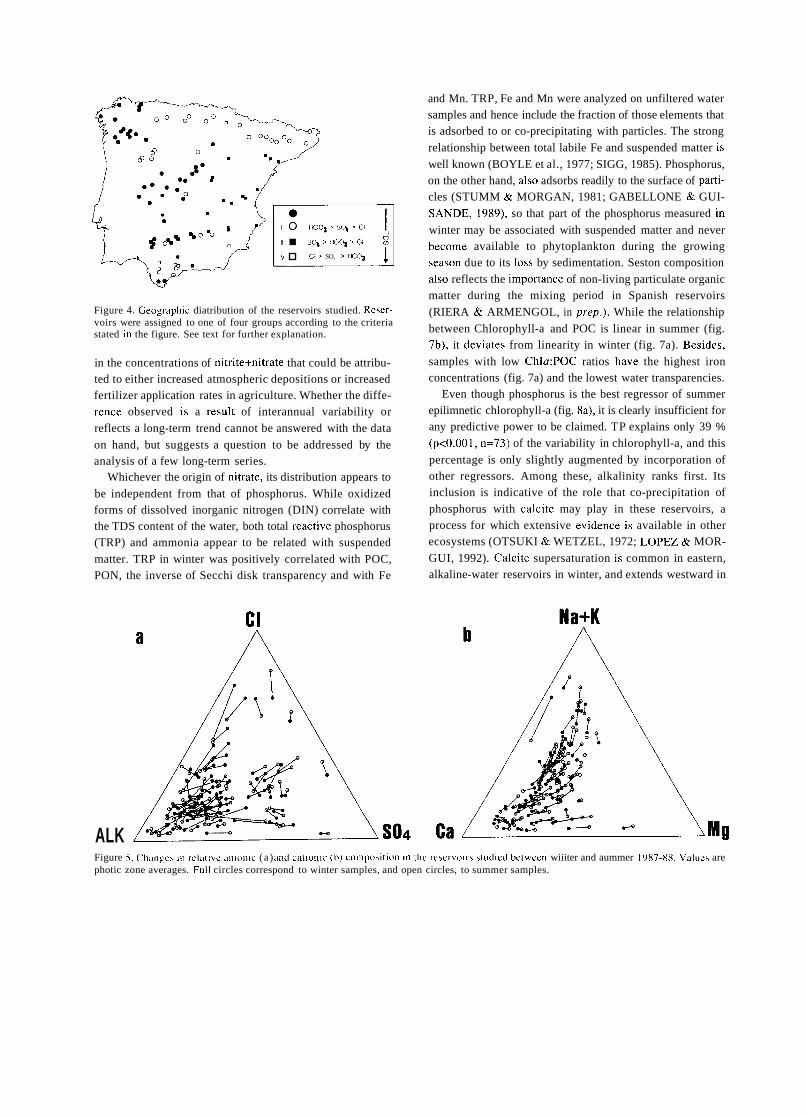
111 SO, > HC03 > Ci
V CI > SO, > HC03
Figure 4. Geographic diatribution of the reservoirs studied. Reser- voirs were assigned to one of four groups according to the criteria stated in the figure. See text for further explanation.
in the concentrations of nitrite+nitrate that could be attribu- ted to either increased atmospheric depositions or increased fertilizer application rates in agriculture. Whether the diffe- rence observed is a result of interannual variability or reflects a long-term trend cannot be answered with the data on hand, but suggests a question to be addressed by the analysis of a few long-term series.
Whichever the origin of nitrate, its distribution appears to be independent from that of phosphorus. While oxidized forms of dissolved inorganic nitrogen (DIN) correlate with the TDS content of the water, both total reactive phosphorus (TRP) and ammonia appear to be related with suspended matter. TRP in winter was positively correlated with POC,
and Mn. TRP, Fe and Mn were analyzed on unfiltered water samples and hence include the fraction of those elements that is adsorbed to or co-precipitating with particles. The strong relationship between total labile Fe and suspended matter is well known (BOYLE et al., 1977; SIGG, 1985). Phosphorus, on the other hand, also adsorbs readily to the surface of parti- cles (STUMM & MORGAN, 1981; GABELLONE & GUI- SANDE, 1989), so that part of the phosphorus measured in winter may be associated with suspended matter and never become available to phytoplankton during the growing season due to its loss by sedimentation. Seston composition also reflects the importance of non-living particulate organic matter during the mixing period in Spanish reservoirs (RIERA & ARMENGOL, in prep.). While the relationship between Chlorophyll-a and POC is linear in summer (fig. 7b), it deviates from linearity in winter (fig. 7a). Besides, samples with low Ch1a:POC ratios have the highest iron concentrations (fig. 7a) and the lowest water transparencies.
Even though phosphorus is the best regressor of summer epilimnetic chlorophyll-a (fig. 8a), it is clearly insufficient for any predictive power to be claimed. TP explains only 39 %
(p<0.001, n=73) of the variability in chlorophyll-a, and this percentage is only slightly augmented by incorporation of other regressors. Among these, alkalinity ranks first. Its inclusion is indicative of the role that co-precipitation of phosphorus with calcite may play in these reservoirs, a process for which extensive evidence is available in other ecosystems (OTSUKI & WETZEL, 1972; LOPEZ & MOR- GUI, 1992). Calcite supersaturation is common in eastern,
PON, the inverse of Secchi disk transparency and with Fe alkaline-water reservoirs in winter, and extends westward in
ALK Figure 5. Chaiigeh 111 r e l~ i i i~e aiiioiiic (a ) aiid catioiiic (b) conrpositioii i i i tlie 1-eservoii-S studied betweeii wiiiter and aummer 1987.88. Values are photic zone averages. Full circles correspond to winter samples, and open circles, to summer samples.

Figure 6. Distribution of phoiic Loiie avei-age5 ot' [he satui-aiioii iiidex loi. calciic tlui-~iig ( a ) u iiiicr aiitl (b) ~~iiiiiiici-. Dois represent undersatu- ration, whereas circles correspond to supersaturation, the size of the syrnbol being proportional to the saturation index.
Table 3. Summary statistics for selected physical and chemical variables. Values given were calculated on the entire data set, including a11 depths and seasons (rnixing and stratification).
Variable n min Q 1 median 4 3 max
Secchi disk depth (m) Temperature ("C) Dissolved oxygen (ppm)
Eh (PV) Conductivity (pS cm-') Alkalinity (peq dm3)
PH Sulphate (peq dm-3) Chloride (peq dm-3) Sodium (peq dm-3) Potassium (peq dm-3) Calcium (peq dm-3) Magnesium (peq dm-3) Iron (pmol dm-3) Manganese (cm01 dm-3) Nitrite (pmol dm-3) Nitrate (pmol dm-3) Ammonia (pmol dm-3) TN (pmol dm3) TRP (pmol dm-3) TP (pmol dm-3) TRSi (pmol dm-3) POC (prnol dm-3) PON (pmol d m ~ ~ ) ETS (p1 O, h-' dm-3) Chla (mg m-3)
0.1 1 4.7
0 -343.0
14 55
5.93 traces
21.3 2.55 3.78
14.75 12.07 traces traces
0.01 0.08 0.01 5.37 0.01 0.01 0.01 1.75 0.01
0 0.01
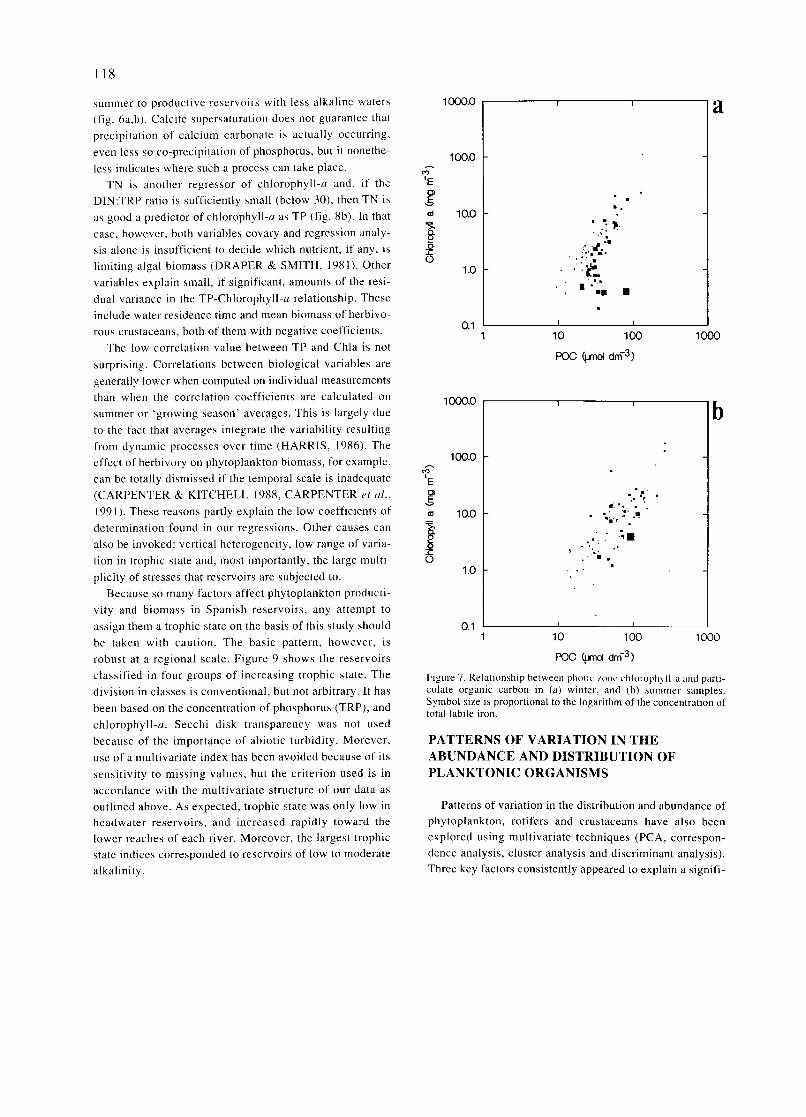
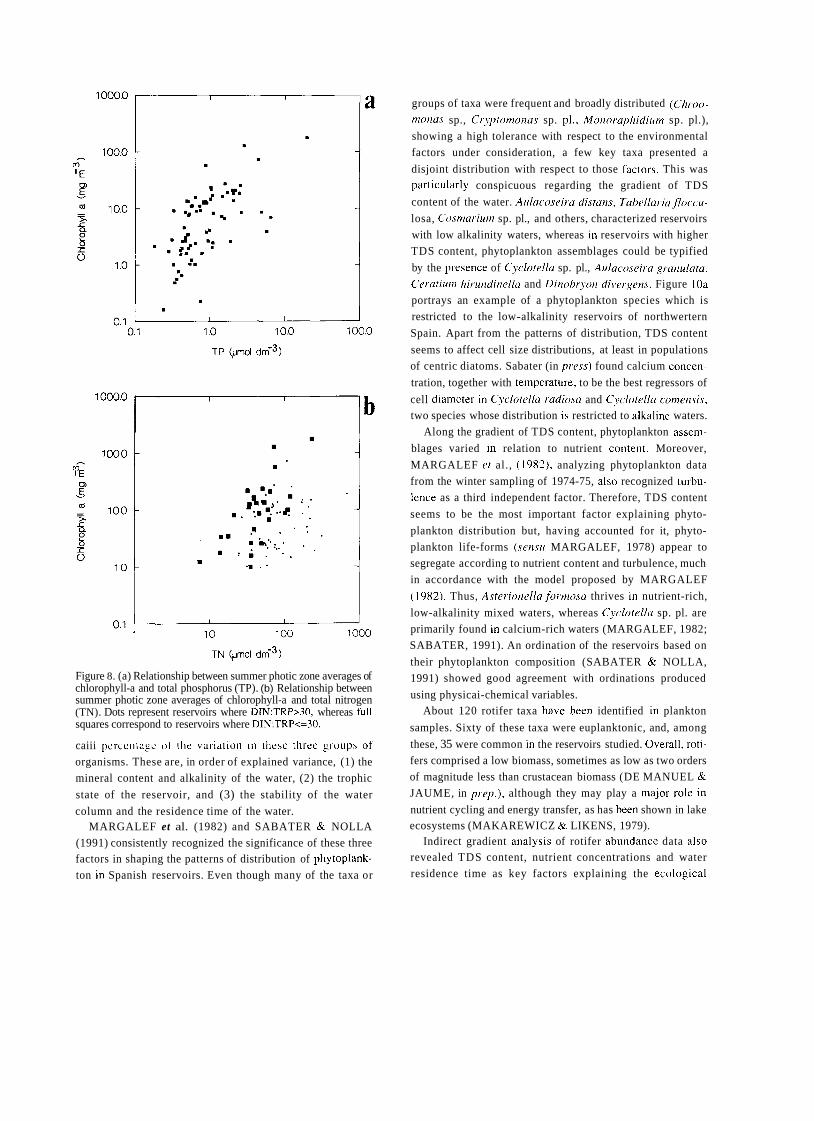
Figure 8. (a) Relationship between summer photic zone averages of chlorophyll-a and total phosphorus (TP). (b) Relationship between summer photic zone averages of chlorophyll-a and total nitrogen (TN). Dots represent reservoirs where DIN:TRP>30, whereas full squares correspond to reservoirs where DIN:TRP<=30.
caiii pci-cciiiagc ol thc variaiioii i i i ilicsc thi-ec groups of
organisms. These are, in order of explained variance, (1) the
mineral content and alkalinity of the water, (2) the trophic
state of the reservoir, and (3) the stability of the water
column and the residence time of the water. MARGALEF et al. (1982) and SABATER & NOLLA
(1991) consistently recognized the significance of these three factors in shaping the patterns of distribution of phytoplank-
ton in Spanish reservoirs. Even though many of the taxa or
groups of taxa were frequent and broadly distributed (Cht.oo-
rnonrrs sp., Cry17tornonas sp. pl., Monoraphidiurn sp. pl.),
showing a high tolerance with respect to the environmental
factors under consideration, a few key taxa presented a
disjoint distribution with respect to those factors. This was
particularly conspicuous regarding the gradient of TDS
content of the water. Aillacoseira distans, Tf~be/íarirrj7or~cu-
losa, Cosn~ariurn sp. pl., and others, characterized reservoirs
with low alkalinity waters, whereas in reservoirs with higher
TDS content, phytoplankton assemblages could be typified
by the presence of Cyclorella sp. pl., Aulucoseira granulara,
Ceratiurn kirundinella and Dinohr.youi diver.gens. Figure loa
portrays an example of a phytoplankton species which is
restricted to the low-alkalinity reservoirs of northwertern
Spain. Apart from the patterns of distribution, TDS content
seems to affect cell size distributions, at least in populations
of centric diatoms. Sabater (in press) found calcium concen-
tration, together with temperature, to be the best regressors of
cell diameter in Cyclotc4la i.adiosu and Qc.lotella cornen.si.s, two species whose distribution is restricted to alkaline waters.
Along the gradient of TDS content, phytoplankton assem-
blages varied in relation to nutrient content. Moreover,
MARGALEF et al., (1982), analyzing phytoplankton data
from the winter sampling of 1974-75, also recognized turbu-
lence as a third independent factor. Therefore, TDS content
seems to be the most important factor explaining phyto-
plankton distribution but, having accounted for it, phyto-
plankton life-forms (sensu MARGALEF, 1978) appear to
segregate according to nutrient content and turbulence, much
in accordance with the model proposed by MARGALEF
(1982). Thus, Astei.ionellcr forrnosa thrives in nutrient-rich,
low-alkalinity mixed waters, whereas Cyc'lotella sp. pl. are
primarily found in calcium-rich waters (MARGALEF, 1982;
SABATER, 1991). An ordination of the reservoirs based on
their phytoplankton composition (SABATER & NOLLA,
1991) showed good agreement with ordinations produced
using physicai-chemical variables. About 120 rotifer taxa have been identified in plankton
samples. Sixty of these taxa were euplanktonic, and, among
these, 35 were common in the reservoirs studied. Overall, roti-
fers comprised a low biomass, sometimes as low as two orders
of magnitude less than crustacean biomass (DE MANUEL &
JAUME, in pi.ep.), although they may play a major role in
nutrient cycling and energy transfer, as has been shown in lake ecosystems (MAKAREWICZ & LIKENS, 1979).
Indirect gradient analysis of rotifer abundance data also revealed TDS content, nutrient concentrations and water
residence time as key factors explaining the ecological

segregation of different taxa (DE MANUEL & ARMEN- GOL, in press). No distinct type communities could be found, but loosely formed assemblages, whose composition may vary rapidly in response to changes in environmental conditions, have been identified. Along the compound gradient of TDS content and aridity that extends from NW to SE Spain, different assemblages could be found, with distinct seasonal changes. Ascomorpha sp. pl., Gastropus stylifer, Synchaeta pectinata and Asplanchna sp. pl. were characteristic of alkaline reservoirs, while Ploesoma hudsoni, Collotheca sp. pl., Conochilus sp. pl. and Trico-
cerca cylindrica constituted an assemblage common in northwestern reservoirs. Moreover, water temperature played a significant role in relation with the geographic distribution of a few species. A number of warm stenother- mous species appeared to be restricted to southern reservoirs (e.g. Brachionus falcatus and Filinia opoliensis), while Keratella tropica (fig. 10b) has extended its geographic 1-iglii-c 10. ( L I J ~ i ~ ~ i i ~ ~ i ~ i i i i i ~ r ~ I I C I I ~ ~ O I I . ,i L ) ~ \ I I I I ~ ~ I , I L ~ C , I C i ~ \ i i - i i . i ~ d 111
Spain to northwestern reservoirs, which have waters with low alka- distributiOn in cOmparison with the 1972-1975 study linity and low TDS content. (b) Keratelia tropica (Brachionidae), a (GUISET 1977. DE MANUEL 199 1 j. warm stenothermous rotifer suecies that a ~ u e a r s mainlv in southern
. A
reservoirs. Scale bar, 50 pm. Scanning electron microscope photo- The fauna of eu~lanktonic crustaceans is scanty in the graphs at the Servei de Microscopia Electrbnica of the Univer- Iberian peninsula, mainly as a result of the scarcity of true sity of Barcelona.
Figure 9. Map showing the reservoirs studied classilietl i i i gi'olip\ o l iiicreasing trophic state. The trophic state index (TSI) was calculated from summer epilimnetic chlorophyll-a and TRP concentrations. For detaiis, see MORGUI et al., (1990).

limnetic habitats. Even though the extensive construction of reservoirs substantially increased the quantity of such habi- tats, only 28 euplanktonic species (15 cladocerans and 13
copepods) have been reported (ARMENGOL, 1978a), a number considerably lower than expected from studies in European lakes (MARGALEF, 1983). The fact that eight of these species had not been previously found in Spain (ARMENGOL, 1980), indicates the importance of reservoirs as new habitats for colonization. Ceriodaphnia cornuta, a species broadly dristrubuted throughout tropical and subtro-
pical areas around the world, has been found in two reser- voirs in southern Spain. On the other hand, cyclomorphic Daphnia (D. cucullata, D. galeata) appeared in northern reservoirs, possibly coming from Central-European lakes.
TDS content, trophic state and water residence time again appeared to explain a substantial proportion of the variance in crustacean data (ARMENGOL 1978b). Ecological segre- gation of species was often related to the compound gradient of mineralization and aridity. For example, the two species of Diaphanosoma (Ctenopoda, Sididae) found in Spanish reservoirs, D . brachyurum and D. mongolianum, show a clear allopatric distribution, the former being restricted to the north of the peninsula, and the latter to southern Spain (JAUME 1991). The residence time of water also affects crustaceans and, among al1 of the environmental factors considered, it is the best regressor of total crustacean biomass during the mixing period ( ~ ~ = 0 . 4 0 , p<0.001, n=60).
Our study of the regional limnology of Spanish reservoirs is a case in point. As we have seen, the landscape position of any particular reservoir, including the geology and land uses of its watershed, its position in the river and the climate of the region, largely determine the major ionic composition of its waters. Both the temporal and the spatial scales relevant at this level are large, and hence a very robust typology of Spanish reservoirs can be made. Nutrient concentrations are impor- tantly affected by living organisms, but the general pattems of variation at the regional scale are still robust, revealing the importance of processes taking place in the watershed.
At the scale of planktonic organisms, interactions taking place within the basin prevail over externa1 constraints. While the effect of the ionic composition and nutrient content of the waters, and ultimately that of landscape posi- tion and morphometry of reservoirs, still explain a signifi- cant proportion of the variation in the composition, abun- dance and geographical distribution of planktonic organisms, other processes operating at spacial-temporal scales dismis- sed by regional studies may have a much stronger effect.
ACKNOWLEDGEMENTS
Financia1 support has been provided by CICYT grant n" PB 85-0166 and sponsored by the Dirección General de Obras Hidráulicas of the Ministery of Public Affairs. J.L. Riera, D. Jaume and J.A. Morguí benefited from a FPI grant
to participate in this project.
CONCLUDING REMARKS REFERENCES
Interpretation of limnological data from studies at a regional scale is seriously affected by limitations at the spatial and temporal scales at which data acquisition is made. This problem is worsened by the fact that these
studies have to deal with different levels of analysis, each of which has its particular scales of variability. Only when these scales are matched by the scales of data acquisition is the recovery of information possible (ALLEN & STARR, 1982; FROST et al., 1989). Regional studies cannot, for this reason, efficiently recover the variace generated at short time scales or small spatial scales, and this variance is inevitably lost as uninformative noise. Countering this limitation, regional studies provide an opportunity to understand the hierarchical organization of limnological data and the interrelationships among the different levels of this hierarchy, let alone the massive body of descriptive data that they generate.
ALLEN, T.F.H. & T.B. STARR. 1982. Hierarchy. University of Chicago Press, Chicago.
ALONSO, M. 1985. Las lagunas de la España peninsu- '
lar: taxonomia, ecología y distribución de los cladóceros. Ph.D. dissertation, University of Barcelona, Spain.
ALVAREZ COVELAS, M. 1982. Una limnologia de la cuenca del embalse de El Vellón Madrid. Ph.D. dissertation, Univ. Complutense, Madrid.
ARMENGOL, J. 197th. Los crustáceos del plancton de los embalses españoles. Oecologia Aquatica,3:3-96.
ARMENGOL, J. 1978b. Zooplankton crustaceans in Spanish reservoirs. Verh. Internat. Verein. Limnol., 20: 1652- 1656.
ARMENGOL, J. 1980. Colonización de los embalses espa- ñoles por crustáceos planctónicos y evolución de la estructura de sus comunidades. Oecologia Ayuatica,4:45-70.

122
ARMENGOL, J., F. SABATER, J.L. RIERA, & J.A. MORGUI. 1990a. Annual & longitudinal changes in the envi- ronmental conditions in three consecutive reservoirs of the Guadiana river W. Spain. Areh. Hy(lrohio1. Beih., 33:679-687.
ARMENGOL J.. J. CATALAN, N. GABELLONE, D. JAUME, J. DE MANUEL, J.A. MORGUI, J. NOLLA, J. PEÑUELAS, M. REAL, J.L. RIERA, S. SABATER, F. SABATER, & J . TOJA. 1990b. A comparative limnogical study of the Guadalhorce reservoirs system Málaga, SE Spain. Sc,ientiu Ger.llndensis, 162:27-4 1.
ARMENGOL, J, J.L. RIERA, & J.A. MORGUI. 1991. Major ionic composition in the Spanish reservoirs. C7et.h. Internar. Verein. Limnol., 24: 1363: 1366.
BOYLE, E.A., J.M. EDMOND, & E.R. SHOLKOVITZ. 1977. The mechanism of iron removal in estuaries. Geochim. Cosrnochim. Acta 41 :13 13-1324.
CARPENTER, S.R. & J.F. KITCHELL. 1988. Consumer control of lake productivity. BioScirnce, 38:764-769.
CARPENTER, S.R., T.M. FROST. J.F. KITCHELL, T.K. KRATZ, D.W. SCHINDLER, J. SHEARER, W.G. SPRU- LES, M.J. VANNI, & A.P. ZIMMERMAN. 1991. Patterns of primary production & herbivory in 25 North American Lake Ecosystems. In: Coniparative crna-'ses ($ ec,osystrm.s J. Cole, G. Lovett, S. Findlay, eds.. Springer Verlag, New York, pp. 67-96.
DE MANUEL, J. 1991. Distribution of the Brachionidae Rotifera:Monogononta in Spanish reservoirs. Verh. Intei'nar. Vel.ein. Lirnnol., 24:2741-2744.
DE MANUEL, J. & J. ARMENGOL. itr pt.c7s.s. Rotifer assemblages: a contribution to the typology of Spanish reservoirs. Hydi.ohiologia.
DE MANUEL J. & D. JAUME. in prep. Zooplankton from reservoirs of the Guadalquivir river basin Spain.
DIRECCION GENERAL DE OBRAS HIDRAULICAS 1988. Inventario de Presas españolas, 1986. M.O.P.U. Madrid.
DRAPER, N. & H. SMITH. 1981. Applied Regression Analysis, 2nd Ed. Wiley, New York.
FROST, T.M., D.L. DEANGELIS, S.M. BARTELL, D.J.
HALL, & S.H. HURLBERT. 1988. Scale in the design &
interpretation of aquatic community research. In: Cornp1e.r interacíions in lake ecosystems S.R. Carpenter, ed.. Sprin- ger-Verlag, New York.
GABELLONE, N.A. & C. GUISANDE. 1989. Relations- hip between texture & fractions of inorganic phosphorus in the surface sediment of a reservoir. Aquatic S(,ience.s, 5 1 :306-3 16.
GIBBS, R.G. 1970. The mechanisms controlling world water chemistry. Science, 170: 1088-1090.
CALVEZ. J.A., F.X. NIELL, & J. LUCENA. 1989. Seston vertical flux model for an eutrophic reservoir. Arc.h. Hydrohiol. Beih., 335-1 8.
GUISET, A. 1977. General distribution of planktonic roti- fers in Spanish reservoirs. Arch. H.vdrohio1. Beih., 8:222-225.
HARRIS, G.P. 1986. Phytoplunkron Ecology: Structilre,
Furzction & Fluctuation. Chapman & Hall, London. JAUME, D. 199 1. The genus Diuphanosoma Ctenopoda:
Sididae in Spain. Hydrohiologia, 225:23-35. KRZANOWSKI. W.J. 1988. Principies of Mulri~~uria te
Analysis: A User's Perspectii,e. Clarendon Press, Oxford. LOPEZ, P. & J.A. MORGUI. 1992. Phosphate & calcium
carbonate saturation in a stratified coastal lagoon. Hydro- hiologia, 22855-63.
MAKAREWICZ, J.C. & G.E. LIKENS. 1979. Structure & function of the zooplankton of Mirror Lake. New Hamps- hire. Ecol. Monogr., 49: 109-127.
MARGALEF, R. 1978. Life-forms of phytoplankton as survival alternatives in an unstable environment. Ocranol. Acra, 1 : 493-509.
MARGALEF, R. 1983. Lirnnología. Omega, Barcelona. MARGALEF, R., D. PLANAS, J. ARMENGOL, A.
VIDAL, N. PRAT, A. GUISET, J. TOJA, & M. ESTRADA 1976. Limriología de los embalses españoles. Dirección General de Obras Hidráulicas, Ministerio de Obras Públi- cas, Publ. n. 123. Madrid.
MARGALEF, R., M. MIR, & M. ESTRADA. 1982. Phytoplankton composition & distribution as an expression of properties of reservoirs. Can. Wat. Res. .J., 7:26-50.
MORGUI, J.A., J. ARMENGOL, & J.L. RIERA. 1990. Evaluación limnológica de los embalses españoles: compo-
sición iónica y nutrientes. Com~lnicacioi~s III Jornadas Españolas de Presas. Barcelona.
MOYA, G. & G. RAMON. 1981. Características limno- lógicas del embalse de Cúber Mallorca. Actas del Primer Congr.eso Español de Lirnnología, Barcelona, pp. 83-92.
O'NEILL, R.V., D.L. DEANGELIS, J.B. WAIDE, &
T.F.H. ALLEN. 1986. A hierarchic.al concept qf'ecosysrem. Princeton University Press, Princeton.
ORTIZ, J.L. & R. PEÑA. 1984. Applicability of the OECD eutrophication models to Spanish reservoirs. Verh. Internar. Verein. Lirnno1.,22:1521-1535.
OTSUKI, A. & R.G. WETZEL. 1972. Coprecipitation of phosphate with carbonates in a mar1 lake. Lirnnol. Ocea- nogt... 17:763-767.
PACKARD, T.T. 1985. Measurement of electron trans- port activity of microplankton. Adv. Ayuatic Mici.ohiology, 3:207-26 1.

PICAZO, M.T. & G. LEMEUNIER, eds. 1990. Aglra y
nzodo de prod~1cci611. Crítica, Barcelona.
RIERA, J.L., E. MARTI, & J.A. MORGUI. 1991. Changes
in the trophic state of the Spanish reservoirs during the last
sixteen years. Verh. 1ntel.nat. Vevein. Limnol., 24:1367-1370.
RIERA, J.L. & J. ARMENGOL. in [?rep. Relationships
between seston composition & water transparency in
Spanish reservoirs. SABATER, S. & J. NOLLA. 1991. Distributional pa-
tterns of phytoplankton in Spanish reservoirs: First results
& comparison after fifteen years. Verh. Internat. Vei'ein.
Limnol., 24: 137 1-1375.
SABATER, S. 1991. Size as a factor in centric diatoms
distribution:the Spanish reservoirs as an example. Oecolo-
gia uquutica, 10:45-60.
SIGG, L. 1985. Metal transfer mechanisms in lakes; the
role of settling particles. In: Chernical P~.ocesses in Lakes
W. Stumm, ed.. Wiley-Interscience. STUMM, W. & J.J. MORGAN. 198 1. Ayuutic r.hemisti.y.
An introduction emphasizirzg (.hemical eyuilihria in natlrral
waters. 2nd ed. J. Wiley & Sons, New York.
TOJA. J. 1984. Limnologíu (le los emha1.se.s de ahasteci-
miento de agua a Sevilla. Servicio de Publicaciones de
CEDEX, M.O.P.U., Madrid.
VIDAL, A. 1969. Evolution d'un lac de barrage dans le
NE de I'Espagne pendant les quatre premieres années de service. Verh. Internat. Verein. Limnol., 17: 19 1-200.
VIDAL, A. 1973. Development et evaluation du phyto-
plancton dans le reservoir de Sau. XI Congr.ess Int. Cornnz.
Large Dams, Madrid.

THE LOW MINERALIZED AND FAST TURNOVER WATERCOURSES OF GALICIA
E. Martínez-Ansemill and P. M e m b i e l a 2
1. Departamento de Bioloxía Animal e Bioloxía Vexetal, Facultade de Ciencias, Universidade da Coruña, Campus da Zapateira s/n, 1507 1 A Coruña. Spain. 2. Escola Universitaria de Formación do Profesorado de E.X.B., Universidade de Vigo, Vicente Risco s/n, 32001 Ourense. Spain.
Keywords: Running waters, Galicia, Physicochemistry, Soft waters, Communities.
ABSTRACT
The climate of Galicia, which is characterized by its oceanic influence, its geological composition, made up basically granite
and schist, and its relief, produce a wide hydrographic network with waters which are generally acid and very soft in this area
of the Iberian Peninsula. A relatively high specific richness with a high percentage of endemisms and a wide spectrum of sexual maturity periods and
species distribution ranges form the principal aspects to be taken into account in a characterization of its communities.
INTRODUCTION
Limnological studies in Galicia (NW Iberian Peninsula) are still scarce (see MEMBIELA et al., 1990). Yet, available information allows elucidation of the main patterns in the
physicochemical characteristics of these waters and the
composition and ecological behavior of their communities.
The limnological characterization of this area would help to describe the limnetic diversity of Spain, since most limno-
logical research in Spain has been focused on hard waters
over calcareous or clay substrates. This summarizes earlier research on the hydrochemical
characteristics of unpolluted watercourses in Galicia (MEM- BIELA er al., in press), and the ecology of the Plecoptera,
Trichoptera, Chironomidae and Oligochaeta that inhabit them.
THE ENVIRONMENT
General characteristics of the study area
The topography of Galicia (surface area 29.434 Km2; fig. 1 B) determines the existence of an extensive hydrographic
network (fig. 1 A). With the northern and easthern mountain ranges setting the boundary, there are a great number of
short watercourses draining into the Cantabrian-Atlantic Sea. The most important of them being the Eo (91 Km long
along its main course, catchment area 1032 Km*), the Tambre (139 Km long, catchment area 1770 Km2), and the Ulla (1 15 Km long, catchment area 2033 Km2). The eastern
and southeastern ranges, with maximal altitudes of around 2000 m, along with the above-mentioned ranges, delimit a
sparser and more hierarchical river network, where most of
the watercourses are joined into a single river, the Miño (around 350 Km long), draining an area of approximately
14,000 Km2 (nearly half of the surface area of Galicia). The
most important rivers show steep slopes, even in their lower sections. Some of the profiles are concave (the Miño, Anllóns); others are concave-convex (the Tambre, Eume, Xallas) os concave with a strong convexity (Umia).
The Galician terrain is included in what is known as the
Hesperian Massif and shows great lithological diversity (fig. IC). As a whole, there is an abundance of metamorphic
rock outcrops (schist, slate, quartzite, gneiss, migmatite,
etc.) and in the central western zone, the granitic rocks. This
area also contains complexes of basic and ultrabasic rocks. Most of the sedimentary material and the scarce limestone
and dolomite outcrops on Galicia are found in the eastern zone. As a whole, the lithology of Galicia is dominated by hard materials of low solubility.
Lirririciicü. 8 . 125-130 (1992) O Asociación Española de Limnologia, Madrid. Spain

Figure l . boiiit. \ciiiiig ol ilic I i \ci ~ , ~ i i . l i i i i t . ~ ~ i i i i ( ~ . i l i i i c i . . \ I \I,iiii i I i . i i i i , ~ ~ ~ ~ I ~ , i ~ i i ~ ~ . 13 i l < i l i ~ c i i \ I i i i i i ~ I i ~ i i i ~ I IL , IL . I \ . i l i i i \ i , w.i i c . \ i l i . ( I Lithology. modified froni MACIAS (1986): C I , \chit\ arid gneiase\ with low quasil conteni: C2, slatcs and schists with high quai .1~ conieni: C.3, limesto- nes and dolomites; C4, basic and ultrabasic rocks: CS. sediments; C6. granites. D) Annual average precipitations (in mm), modified from CARBALLEIRA et al. (1983).
The prevalling westerly winds extend the influence from tory reaches values of over 1000 mm (CARBALLEIRA er
the Atlantic to the entire region, and, although the average al., 1983). The mountainous strip that runs parallel to the
yearly rainfall varies (fig. ID), most of the Galician terri- coast has high annual precipitation, particularly in the west,

where they intercept the ciouds coming in frorn the sea, causing a significant decrease in the rainfall received in the inland areas of Galicia. Precipitation increases again in the
east-southeastern mountains owing to the higher altitudes.
In general, maximum rainfall occurs in December-January,
with a secondary peack sometime around April, and mini- mum rainfall is found in July.
There is a clear relationship between rainfall distributiori
and the flow regime of the watercourses, although in some
cases there is a snow component. On the whole, the 1ow water period extends from July to October, while high waters occur between January and March, and, exceptio-
nally, extends to May (fig. 2).
Physicochemistry o f the waters
Most of the hydrochemistry research on Galician water-
courses focusses on contaminated watercourses, which represent a small fraction of the fluvial network of Galicia. MEMBIELA et al. ( 1 99 1) describe the main physicochemi-
cal aspects of unpolluted waters. This study, made up over a
seasonal analysis in 25 sampling stations spread al1 over
Galicia provides a basis to examine the relationships between environmental characteristics and the biotic communities of the unpolluted watercourses in the Galician
territory. A multivariate analysis using the Factor Analysis of the BMDP4M (DIXON & BROWN, 1977) points out the
relative importance of the 14 chemical parameters analysed in the characterization of the waters as well as the correla-
tions between them, and shows,
- The extent of mineralization of the waters, as accoun- ting for differences among locations, which can be grou- ped, therefore, into river networks with calcareous mate-
rial on their catchments, and those on non-limestone geolo- gical basins (Table 1). The geological substrate of the
watershed is, therefore, the main factor constraining the physicochemical characteristics of unpolluted Galician
watercourses, as has been described for waters elsewhere
(e.g. GORHAM, 1961; GIBBS, 1971; CAPBLANCQ &
TOURENQ, 1978). - Maririe influences reflected in the concentrations of C1-
and Na+ ions, appear to be of secondary importance in diffe- rentiating the chemical characteristics of the watercourses.
This influence is best reflected by a strong negative corre- lation between the concentrations of those ions and distance from the sea (GARCIA et al., 1977) in agreement with
studies for coastal regions elsewhere (GORHAM, 1961; BALDWIN, 197 1; VITOUSEK, 1977).
_ TAMBRE IPORTOYOUPOI
M ~ / S E0 w3/s _ DUBRA IPORTOMOURO, 80 IN S TIRSO DE ABRES
80 1
40
_ MlNO 1 ~ ~ 0 0 1
MlNO I R I B A D E )
~ ' 1 5 CABE I N R l B A l ALTAS
I *=/S CABRERA
# N PTE OOl lNBO í L O R L 1
I
I - l ~ l l i ~ 2 . \ I i > l l i l l l > . i \ i~l , igi , \ i l i i i i i l i ~ i i i l i r 1 , \ . l. 1). .l.iilii.il> lil L)c- cember) of differeiii Galicinn aniercoui-sea (Tambre. Eo, Miiio, Cabe. Cabrera). Data from D.G.O.H. ( 1 9 7 9 ) .
- Temporal variability, although smaller than spatial variations, is also important. Physicochemical variability allows the identification of two distinct periods, July to
October and January to May, which represent the low and
high water periods respectively.
THE COMMUNITIES
Most biological studies in Galician waters focus on their lotic benthic rnacroinvertebrates, which have increased our knowledge on the major patterns of their ecological be-
havior.
Specific richness
The running waters of Galicia have a relatively high biotic richness, as can be deduced by the substantial number of species found in the groups that have received the most atten- tion (cf. MEMBIELA er al., 1990). The main circumstances that help account for this considerable specific richness are.
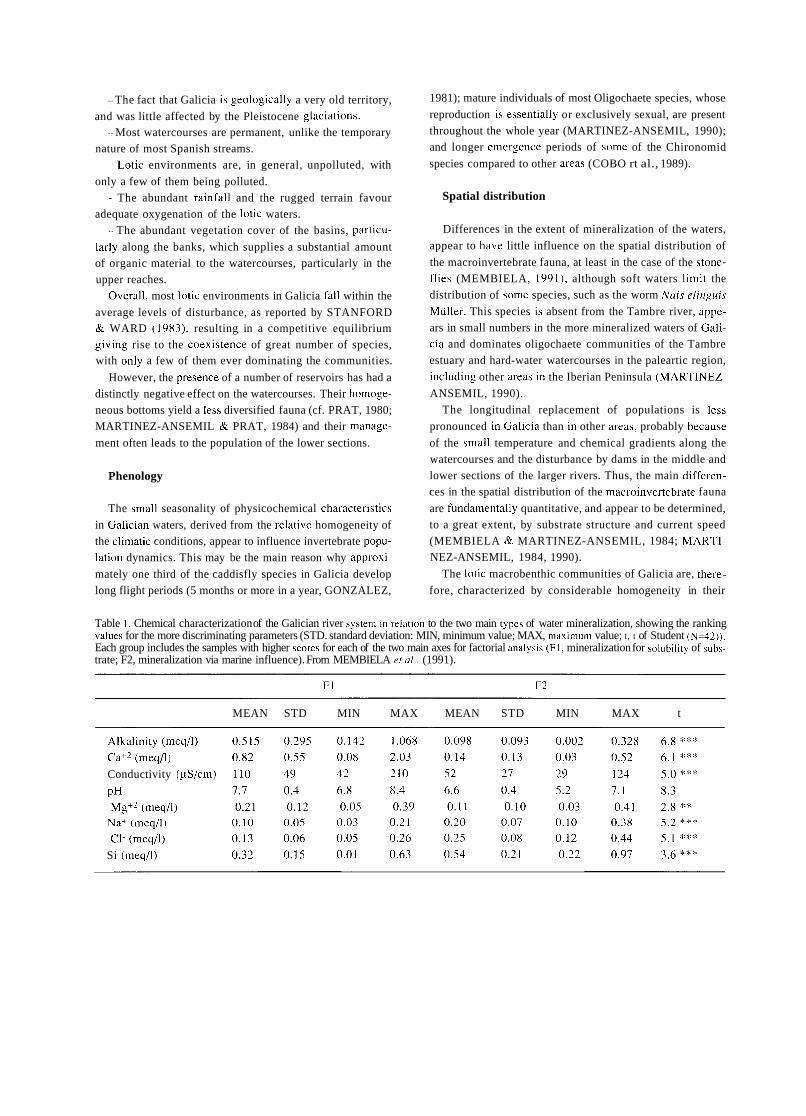
- The fact that Galicia is geologically a very old territory,
and was little affected by the Pleistocene glaciations. - Most watercourses are permanent, unlike the temporary
nature of most Spanish streams. - Lotic environments are, in general, unpolluted, with
only a few of them being polluted. - The abundant rainfall and the rugged terrain favour
adequate oxygenation of the lotic waters. - The abundant vegetation cover of the basins, particu-
larly along the banks, which supplies a substantial amount of organic material to the watercourses, particularly in the upper reaches.
Overall, most lotic environments in Galicia fall within the
average levels of disturbance, as reported by STANFORD & WARD (1983), resulting in a competitive equilibrium giving rise to the coexistence of great number of species,
1981); mature individuals of most Oligochaete species, whose
reproduction is essentially or exclusively sexual, are present throughout the whole year (MARTINEZ-ANSEMIL, 1990); and longer emergence periods of some of the Chironomid
species compared to other areas (COBO rt al., 1989).
Spatial distribution
Differences in the extent of mineralization of the waters,
appear to have little influence on the spatial distribution of
the macroinvertebrate fauna, at least in the case of the stone- flies (MEMBIELA, 1991), although soft waters limit the distribution of some species, such as the worm Nais el ingi~is
Müller. This species is absent from the Tambre river, appe- ars in small numbers in the more mineralized waters of Gali- cia and dominates oligochaete communities of the Tambre
with only a few of them ever dominating the communities. estuary and hard-water watercourses in the paleartic region,
However, the presence of a number of reservoirs has had a including other areas in the Iberian Peninsula (MARTINEZ-
distinctly negative effect on the watercourses. Their homoge- ANSEMIL, 1990).
neous bottoms yield a less diversified fauna (cf. PRAT, 1980; The longitudinal replacement of populations is less
MARTINEZ-ANSEMIL & PRAT, 1984) and their manage- pronounced in Galicia than in other areas, probably because
ment often leads to the population of the lower sections. of the small temperature and chemical gradients along the watercourses and the disturbance by dams in the middle and
Phenology lower sections of the larger rivers. Thus, the main differen- ces in the spatial distribution of the macroinvertebrate fauna
The small seasonality of physicochemical characteristics are fundamentally quantitative, and appear to be determined,
in Galician waters, derived from the relative homogeneity of to a great extent, by substrate structure and current speed
the climatic conditions, appear to influence invertebrate popu- (MEMBIELA & MARTINEZ-ANSEMIL, 1984; MARTI-
lation dynamics. This may be the main reason why approxi- NEZ-ANSEMIL, 1984, 1990).
mately one third of the caddisfly species in Galicia develop The lotic macrobenthic communities of Galicia are, there-
long flight periods (5 months or more in a year, GONZALEZ, fore, characterized by considerable homogeneity in their
Table 1 . Chemical characterization of the Galician river system in relation to the two main types of water mineralization, showing the ranking values for the more discriminating parameters (STD. standard deviation: MIN, minimum value; MAX, maxiinum value; t , t of Student (N=42)). Each group includes the samples with higher scores for each of the two main axes for factorial analysis (Fl, mineralization for solubility of subs- trate; F2, mineralization via marine influence). From MEMBIELA et u/.. (1991).
MEAN STD MIN MAX MEAN STD MIN MAX t
Alkalinity (meq/l) Ca+2 (meq/l)
Conductivity (pS/cm)
PH Mg+2 (rneqll)
Na+ (meq/l) Cl- (meqll)
Si (meqll)

distribution. However, the stoneflies communities (MEM-
BIELA, 1991) from large rivers, can be separated from those
of smaller watercourses, which constitute a heterogeneous group with clear discontinuities and multiple overlapping
distributions. In addition, communities of temporary water-
courses are few in Galicia, compared to other peninsular areas, where these watercourses are dominant.
Biogeographical aspects
Galicia contains a substantial fraction of the high number of endemisms present in the aquatic fauna of the
Iberian Peninsula. Thus, half of the stoneflies and appro-
ximately one third of the caddisflies species found in Gali- cia are peninsular endemisms, which are largely limited to the northwestern zone of the Iberian Peninsula (GONZA-
LEZ et al., 1987; MEMBIELA, 1990). AUBERT (1956)
and GONZALEZ er al. (1987) have reported that the
Hesperian Massif is the zone containing the highest number of endemisms for the stoneflies and caddisflies
respectively. This territory was isolated from the European
Pyrenees until the end of the Oligocene. Moreover, the fact
that the Iberian Peninsula served as a refuge during the glaciar periods of the Pleistocene (MARGALEF, 1955) has
also contributed to the development of a considerable
number of endemisms in the fauna of the northwestern area
of the peninsula.
REMARKS
We have outlined here the main environmental characteris- tics of unpolluted watercourses of Galicia, and provided
evidence of their influence on their benthic communities.
Altough the epicontinental waters of Galicia still have an acceptable level of conservation compared to many other areas
of the Peninsula, this situation is threatened by some activities,
(a) The increasing number of dams which are severely
influencing the major rivers. The D.G.O.H. (1979) counted 29 reservoirs with a capacity of between 4 and 645 Hm3. Others which are under construction or planned must be
added to this list. Also, a considerable number of small hydroelectric plants, some of which are already opperatio-
nal and others in various stages of construction, add to the environmental impact of dams.
(b) Increasing pollution from urban and industrial wastes. Galicia has traditionally been fundamentally a rural society with a population that is spread out in a number of small
nuclei, and whose main activities have been agriculture, lives-
tock and fishing. However since the early 1960's, a conside- rable part of the population has been shifting to the urban
centers which have undergone industrialization at a fast pace.
As a result of this substantial increase in population and indus- trialization, which has taken place mainly around the large
cities (Vigo, A Coruña, Ourense, Santiago, Lugo, etc.), some of tbe watercourses (the Sar, Sarela, Barbaña, Lagares, etc.)
have been notably polluted by both organic matter and chemi-
cals. Many watercourses have also been subject to chemical pollution by detergents, acid rain, purine from farms, herbi- cids, insecticids, etc., about which we have very little data. The low level of mineralization and acidic nature oP these waters,
which are poorly buffered, increases the fragility of Galician epicontinental waters. There is, therefore, an urgengy to act against the serious threats to these watercourses by the ever-
increasing trend towards increased pollution.
(c) Deforestation of watersheds by fires. In the last three decades one third of the total surface of Galicia has suffe- red the effects of fires. This is certainly causing great chan- ges in environmental conditions, although no evaluations
have been carried out as yet. The enormous loss of vegeta-
tion will considerably reduce the amount of organic mate- rial supplied to the watercourses and may even cause major changes in climate. Soil erosion related to fires, leads to
alterations in the aquatic environment with a substantial
increase in fine material in the watercourses.
REFERENCES
AUBERT, J., 1956. Contribution a I'étude des Plécoptk-
res d'Espagne. Mém. Soc. Vaucl. Sci. Nat., 11: 209-276. BALDWIN, S.D., 197 1 . Contribution of atmospheric
chloride in water from relected coastal stream of Central
California. Watc.r Resoul-ces Resear.ch, 7(4): 1007- 1012. CAPBLANCQ, J. & J.N. TOURENQ, 1978. Hydrochi-
mie de la rivikre Lot. Annls Lin~tzol., 14(1-2): 25-37.
CARBALLEIRA, A; C. DEVESA; R. RETUERTO; E. SANTILLAN & F. UCIEDA, 1983. Bioclimatología de Gulicia. Fundación Barrié de la Maza. A Coruña,
391 pp. COBO, F.; M.A. GONZALEZ & J . RAMIL, 1989. Quiro-
nómidos (Diptera: Chironomidae) del río Ulla, 1. Bol. Asoc. es/?. Enfom., 13: 299-308.
DIXON, W.J. & M.B. BROWN, 1977. BMDP-77 Biome- dical compirter.pl-ogl-ums P-seric>s. University of California Press. Berkeley, 880 pp.

DIRECCION GENERAL DE OBRAS HIDRAULICAS, 1979. Afo1.o~ l . Cuenca del Norte cle Espuria. Peifocio 1974- 75 a 1975-76. M.O.P.U. Madrid.
GARCIA, C.; F. MACIAS & F. DIAZ-FIERROS, 1977. Relación entre la composición química de las aguas super- ficiales y la mineralogía de los suelos de Galicia. Acta Cien-
rljricu Compostelana, 14: 337-363. GIBBS, R.J., 1971. Mechanisms controlling world water
chemistry. Science, 170: 1088- 1090. GONZALEZ, M.A., 1981. Tricópteros de Galicicl. Tesis
doctoral. Univ. Santiago de Compostela, 41 1 pp. GONZALEZ, M.A.; D. GARCIA DE JALON & L.S.W.
TERRA, 1987. Faunistic studies on Iberian Trichoptera: a historical survey and present state of knowledge. In: Proc.. 5th. Inr. Symp. T/.ichoptera. M. Bourneaud & H. Tachet (eds.). Junk. Dordrecht: 85-90.
GORHAM, E., 196 1. Factors influencing supply of major ions to island waters with special reference to the atmosp- here. Geol. Soc. Am. Bitll., 72: 795-840.
MACIAS, F., 1986. Materiais orixinais e solos de Gaiiza. In: O meio matur.al gczlego: 47-79. Edicións do Castro. Sada,
206 pp. MARGALEF, R., 1955. Comunidades bióticas de las
aguas dulces del noroeste de España. P. Inst. Biol. Apl., 21 : 5-85.
MARTINEZ-ANSEMIL, E., 1984. Oligoquetos dulcea- cuícolas de Galicia: catálogo y diversos aspectos ecológi- cos. Limnetica, 1 : 3 1 1-320.
MARTINEZ-ANSEMIL, E., 1990. Etude biologique et écologique des Oligoch2tes aquatiques de la rivikre Tambre et ses milieux associés (Galice, Espagne). Annls Limnol., 26(2-3): 131-151.
MARTINEZ-ANSEMIL, E. & N. PRAT, 1984. Oligo- chaeta from profunda1 zones of Spanish reservoirs. Hydro- hiologia, 1 15: 223-230.
MEMBIELA, P., 1984. Primera contribución al conoci- miento de los Plecópteros de Galicia: la cuenca del Tambre. Limnericu, 1 : 197-202.
MEMBIELA, P., 1990. Contribución al conocimiento faunístico de los Plecópteros de Galicia (Plecoptera). Bol.
Asoc. esp. Entom.. 14: 89-103. MEMBIELA, P., 1991. The stoneflies of Galicia (NW
Iberian Peninsula): an ecological study. In: J. Alba Tercedor & A. Sánchez Ortega (Eds.), Overiiew und strutegies of Ephe-
wieroptera ancl Plecoprera. Sanhill Crane Press: 413-424. MEMBIELA, P.; F. COBO; M.A. GONZALEZ & E.
MARTINEZ-ANSEMIL, 1990. A investigación limnolóxica en Galicia con especial referencia ós macroinvertebrados: precedentes, estado actual e perspectivas. Ingenium, 2: 8 1-94.
MEMBIELA, P. & E. MARTINEZ-ANSEMIL, 1984. Larvas de Plecópteros del río Tambre (Galicia): estudio ecológico. Bol. Asoc. esp. Enrom., 8: 101-109.
MEMBIELA, P.; C. MONTES & E. MARTINEZ-ANSE- MIL, 1991. Características hidroquímicas de los ríos de Galicia (NW Península Ibérica). Limnetica, 7: 163- 174.
PRAT, N,, 1980. Bentos de los embalses españoles. Oecolo~ ia uq~laticrr, 4: 3-43.
STANDFORD, J.A. & J.V. WARD, 1983. Insect species diversity as a function of environmental variability and distur- bance in stream systems. In: Streuw~ ecology. J.R. Barnes & G.W. Minshall (eds.). Plenum Press. New York, 265-278.
VITOUSEK, P.M., 1977. The regulation of element concentrations in mountain streams in the northeastern United States. Ecol. Monog., 47: 65-87.

THE ESLA RIVER BASIN: FROM THE CANTABRIAN MOUNTAIN TO T H E DUERO
M. Fernández-Aláez, C. Fernández-Aláez and E. de Luis Calabuig Asea de Ecología. Universidad de León. 24071 León. Spain.
Key words: River basin, Physico-chemical composition, Mineralization, Nutrients.
ABSTRACT
A general physico-chemical outline of the Esla basin (northwest Spain) is carried out, based on previous studies of the various sub-basins of which it is composed. With regard to the lithological composition of the drained lands, the River Tera and a large part of the Orbigo basin, with water low in mineral content, differ from the rest of the Esla basin, which generally contains
water of a higher mineral content. Bicarbonates and calcium are the predominant ionic forms. However, urban waste increases the amount of sulphates and chlorides as well as raising the leve1 of conductivity considerably. The regulation of the water by the reservoirs, agricultural activities and the presence of villages and towns are the determinating factors of the changes in the
concentration of nitrates and phosphates in the basin.
INTRODUCTION
The River Esla is the iongest Cantabrian tributary of the Duero basin and plays a very important role in the fluvial structure in the north-west of the Iberian Peninsula. It is
considered to be the most important tributary on the Penin- sula. Moreover, one of the most outstanding aspects of its basin is the presence of marked contrasts between the hydro- graphical networks which form it. These are a result of a
diversity of human actions, os in some cases a lack of them,
and which affect the fluvial geography. Broadly speaking, the Esla basin is a centre of intense
farming and cattle raising and with some industry, which is not particularly important and is restricted to only a few
areas, as well as the coa1 mines in large areas of the north of the basin which cause pollution that alters the quality of the water on the initial stretches of the rivers, where levels of higher quality would normally be expected. Only three
towns have a population of over 10,000, but none of these possess adequate systems for the purification of sewage. Moreover, more than 50% of the main rivers in the basin are regulated by reservoirs. Nevertheless, there are rivers in which none of the above-mentioned activities are of parti- cular relevante; their special importante within the basin is due to their abundance of fish.
The possibility of analysing such a heterogeneous river
system has led to a series of investigations over the last few years focused on the study of the different rivers making up
the basin, from both a biological (FERNANDEZ, 1986; FERNANDEZ et al., 1986a; FERNANDEZ et al . , 1987a; FERNANDEZ et u/., 1990; FERNANDEZ et al . , 1991 b;
PRESA et al., 1987; PRESA et al., 1988) and physical- chemical (FERNANDEZ et al., 1986b; FERNANDEZ et al.,
1987b; FERNANDEZ et al., 1988, FERNANDEZ et al.,
1989, FERNANDEZ et al., 199 1 a) point of view. This paper
is based on some of the results obtained.
DESCRIPTION OF THE AREA STUDIED
The geography of the river basin
The River Esla is the bigest water collector in the north- west sector of the Duero basin, and carries to it, on the right
hand side, the greatest discharge of al1 its tributaries: 182 m3/s (5,113 Hm3), which is even greater than its own flow at their confluente point. Its basin, with a surface area of 16,081 km2, becomes fan-shaped between the eastern slopes of the Montes de León and the western slopes of the Cantabrian Mountains, as far as the hills coming south of
Liiliiicii~a, 8: 131-140 (1992)
O Asociación Ebpañola de Limnologia, Madrid. Spain
Figure station
LOS BARRIOS DE LUNA
o -
11.4% 411 mm
PUEBLA DE S
1. The Esla catchment, together with ombrothermic diagrains from various meteorological stations and the situation of the s used in figure 4.
gauging
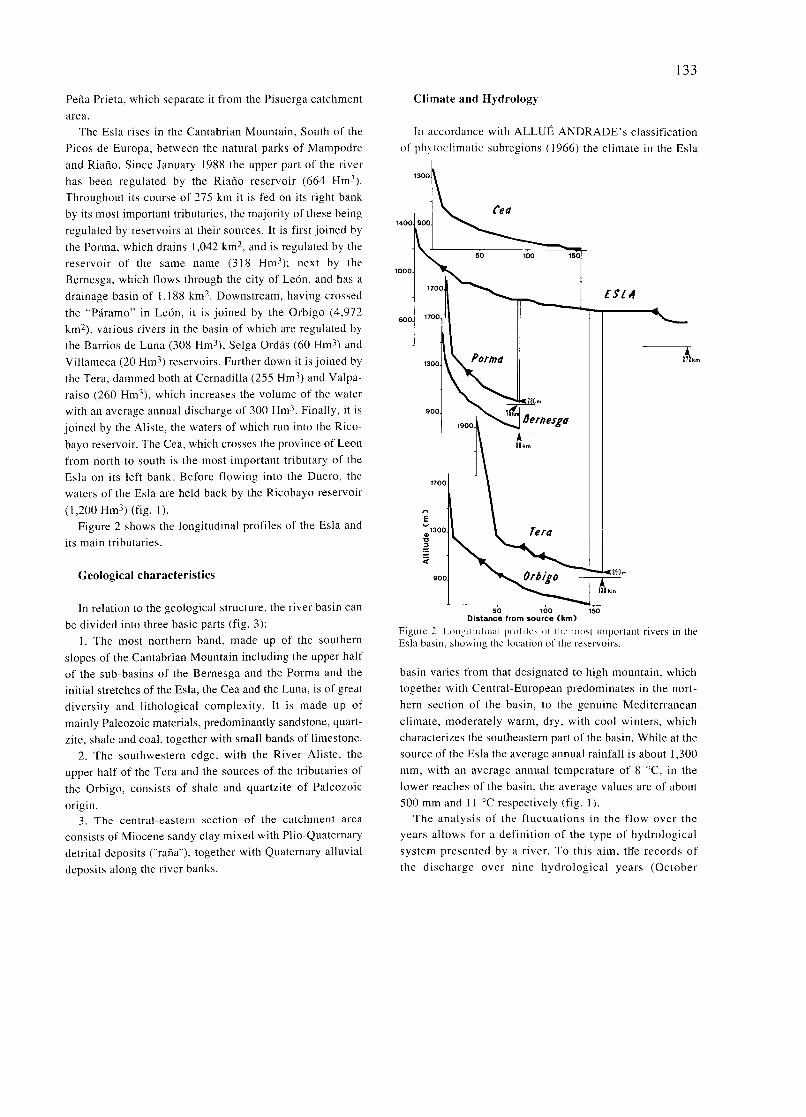

asandstones, shaies, mal and hestones shaies
shales, loams, dolomites, sandstones and lunestones sandy day
~haies and quartzites n
m,, U ~ ~ u v i u m and diiuvium
1980-September 1989) from four gauging stations situa- ted on the Esla were used. As can be seen from the figure 4, the annual variation in the discharge is basically simi- lar throughout the whole river, although a high increase in the flow is produced as the most important tributaries
join it. The maximum flows correspond to spring, in particular to
April(200.91 m'/s in E4) as a result of thawing at the headwa- ters in the mountains. However, since at the end of the autumn
the rainfall is generally heavy there is, of course, an increase in the discharge measured at the gauging stations in the lower
stretches, beyond the confluences of the Cea, the Orbigo and the Tera. During the summer period, in particular in August,
minimum flows are registered in the lower half of the river (E2:
10.54 mys, E3: 19.75 &/S, E4: 30.12 mvs) as opposed to the area around the source, where retention in the form of snow leads to a minimum flow in November (5.23 m3/s).
The marked variations in the flow throughout the year reflect the irregularity of the Mediterranean climate, and determine the hydrological system of the Esla as pluvial- nival. During the winter the leve1 of the water is low since it is accumulated in the form of ice and snow at the source. An important increase in the volume is produced in spring as a result of thawing.
The recent regulation of the Esla has led to a distortion in the system of the river, since it has given rise to a marked
increase in the volume of the water carried by the river
during the summer.
Population and land use
Large sections of the river basin which are situated on its
marginal areas show a density of not over 15 inhabitants per km2. In certain areas, generally those situated around the River Orbigo, there are 125 inhabitants per km2 and even up
to 500 in towns such as Benavente, Astorga and La Bañeza.
However, the greatest density corresponds to the city of
Leon with 3,409 inhabitants per km2. This is the most popu- lated city in the Esla basin, and is, therefore, the most
important centre of pollution, since the Bernesga and the
Torío receive its non-purified urban and industrial waste waters.
Land use in tlie catchment area allows for a clear diffe- rentiation of the mountainous areas which are situated in the
north and west and are largely covered with wide areas of
scrub land and grazing land. There are irrigated meadows around the river banks and there are wooded areas of coni-
ferous and deciduous trees, particularly around the source of the Esla. Tlie rest of the basin is generally given over to
intensive agriculture and there are orchards and irrigated farmland alorig the Orbigo and the Esla and on the "Páramo" in León.
The industrial development of the river basin is, howe- ver, of relatively little significance. It is mainly found in the city of León, where the iilost iiiiportant sectors are the phar- maceutical and the sugar factories. The latter is of impor-
Figure 4. Annual variatioii i i i ilic ki\cr;igc. cli\cli;irgc nt the four selec- ted gauging stations on the River Esla.

tance throughout the river basin, and its waste affects the Bernesga, the Orbigo and the Esla. Also of importance is the
energy sector, based on the production of coa1 and hydroe- lectricity. The most important coa1 deposits are in the nort- hern section, in the Ciñera-Matallana mining valley, situated
between the Bernesga and the Curueño, and the Sabero valley, between the Posma and the Esla.
METHODOLOGY
In order to characterize the Esla basin from a physical and chemical point of view, 126 sample stations were set up
along the different rivers. The collection of samples of water took place seasonally during the period from 1985 to 1990. The following physical and chemical parameters were deter- mined: temperature, total solids in suspension, pH, conduc-
tivity, dissolved oxygen, alkalinity, nitrates, nitrites, ammo-
nium, phosphates, sulphates, chloride, silicates, calcium.
magnesium, sodium and potassium. The analytical methods used are outlined in FERNANDEZ et al. (1987b).
After determining a series of intervals, figures 5 and 6 show the values of conductivity registered in the sainpling
stations of the basin. It has been decided to refer to the results of the spring and summer periods, since these are the
two extreme periods of volume with relation to the hydro- logical system in the river basin. The modifications produ- ced by the regulating effect of the reservoirs should, howe- ver, be taken into account.
Under high flow conditions, the values registered varied
between 24 pS/cm in the Tera and 412 uS/cm near the
mouth of the Bernesga. In summer the range of variations increased, although Ihe extreme values were still found in the same stretches of the river basin: the headwaters of the
Tera, with 19 uS/cm and the final stretch of the Bernesga with a conductivity slightly above 1,000 uS/cm. The least mineralised waters generally corresponded to the tributa- ries of the Orbigo, to the Tera and in particular to certain
sample 5tations at the head of the river basin. However, in
contrast to the low conductivity in the greater part of the tributaries of the Orbigo, the upper courses of this sub-basin
show a significant increase in ionic content, up to over 200 pS/cm. The particular lithological composition of the drai-
RESULTS AND DISCUSSION
Mineral composition of the river basin
The synthetic study of the physical and chemical charac- teristics of the main sub-basins that form the Esla basin
through the principal component analysis always resulted in
a basic factorial plane in which the sample points funda- mentally followed an order in relation to the mineralization of the water. This characteristic of the waters has been found to contain either a combination of the first two components,
or else with the two apart. In the second case, alkaline-type mineralization, more closely linked to the lithological composition of the river basin is related to component 1,
while component 11 represents the mineralization produced
by human activities. The global ionic content of water is known to be directly
related to conductivity (HAUBERT, 1975), and moreover its dependence both on the lithological composition of the
drained lands (OTTO, 1983) and on the dissolved waste of
human origin (MEYBECK, 1979) is evident. In order to offer a simple concept of the physical and chemical co~ripo- sition of the rivers in the Esla basin, the measurement of the electric conductivity of the water as a representative charac-
teristic of its mineralization has been chosen for this reason. Figure S. Location of the \ainpliiig \tatioiis. Distribution of the values of conductivity measured in the catchment during the spring.

SUMMER
Y CONDUCTIVITY
( pS cm-')
Figure 6. Distributi»n o1 tlic \ a l u c \ 0 1 cuii~liicri\ 1 1 ) i~irusurcd ¡ti the catchment during the summer.
ned lands is the cause of these differences, although silice- ous materials predominate there are also some small limes-
tone layers. During the spring, in the middle and lower stretches of
the Orbigo the conductivity was reduced with respect to the source up to 100-200 pS/cm due to the effect of dilution brought about by the incorporation of siliceous tributaries.
In summer, however, and after a slight decrease just after
the dam, levels of mineralization closer to those at the source were registered right along the Orbigo; even in the
lower river they were over 300 pS/cm. The reason for this is the contribution of ions, particularly chlorides and sodium,
which clearly influence the global mineralization of the
water during the period of minimum discharge, and which derive from the important influence of human activities on the riverbanks. It should be noted that in spite of the effect
of the reservoir, the flow of the Orbigo in the lower stretch is lower in summer (1 1 .O9 m3/s) than in spring (5 1.15 m3/s) due to the reduction in the water volume supplied by the
tributaries, the final sections of which also appear to be affected by pollution.
The rest of the Esla basin as a whole contains waters of higher mineral content, particularly the Bernesga sub-basin. Both in the Bernesga and the Torío there is a general
tendency in the course of the river towards a gradual incre- ase in conductivity from the headwaters to the mouth, a
result of the progressive accumulation of salts proceeding from the washing of the land and the waste of human origin (MARGALEF, 1983). In both rivers there is a sharp drop in flow in summer, as a result of which the mineralization
increases considerably and the effects of pollution are inten- sified. The conductivity at the upper Torío is situated at
about 200 uS/cm, whilst at its mouth it reaches 500 uS/cm. In the middle stretch of the Bernesga the impact of the coa1 inines together with the development of large centres of
population is important, provoking high contributions of
sulphates, chlorides and sodium. But the effect of contami- nation is even greater after the confluence with the Torío,
when the Bernesga receives the waste from the city of León
and its conductivity sises to 1,136 uS/cm.
The Curueño is similar to the Torío. The ionic content increases as the drained surface asea does, even more so in the summer with the decrease in volume. In contrast, and
due to the reservoir, the Porma shows little variation in
conductivity from its source to its mouth during the summer. Moreover, it does not generally reach over 200 uS/cm.
Although in spring no important variation was registered
along the river, these levels were surpassed after it was
joined by the Curueño. The mineralization also varied little along the Esla. The
conductivity is below 200 uS/cm until the entry of the rivers
of the Bernesga sub-basin; it then increases only slightly and does not rise above 300 uS/cm. The uniformity of the mine-
ralization values is explained on one hand by the presence of the Riaño reservoir, at the upper river, and on the other hand by its significant reduction in the water flow in
summer, even though the ionic content remains the same in the same season. When the Esla joins the Duero the conduc- tivity is below 300 uS/cm and even in summer it carries water that is relatively low in minerals to the Duero.
Typification of the river basin based on the ionic composition
Considering the samples of the basins of the Porma, the
Bernesga, the Orbigo and the Esla with its small tributaries at the source independently, the proportion of ions has been calculated and then represented in triangular diagrams.(figs 7 and 8) In this way a typification of the water in al1 the
samples is obtained, with a separate representation of the percentage of the majority anions (bicarbonates + carbona- tes, chlorides and sulphates) and cations (calcium, magne-

sium and sodium + potassium) for the two selected sample periods: summer and spring.
As can be observed in figure 7, the geo-chemical group into which al1 the samples from the River Esla and al1 the small tributaries at the source are included is the bicarbo-
nate-calcic one.
As regards the Porma basin, the dominant proportion of
mineralization is also made up of bicarbonates and cal- cium. The samples from the Curueño and the small tribu-
taries of the Porma contain more bicarbonates and are
slightly different from those from the Porma in the spring sampling. Although bicarbonates consistently represent
over 50% of the total anions, the proportion of sulphates increases at the source of the Porma because of the
influence of the lithological characteristics. Calcium conti- nues to be the main cation, there is a high proportion of
magnesium, and a minimum presence of sodium and potas- sium in the ionic content (fig. 7).
As shown in figure 8, the Orbigo basin is more heteroge-
neous as regards its proportions of ions. There is still a clear
predominance of bicarbonates in the relation of the anions; however certain samples in tributaries, such as the Tuerto and the Jamuz affected by the pollution from small villages, are more widely dispersed, due, basically, to the increase in
the percentage of chlorides. In the triangular diagram of the
cations, calcium is also dominant, but in a group of samples belonging to the Tuerto, the Jamuz, the Brañuelas and the Eria the other cations, in particular magnesium, are of grea-
ter importance. Among the ions in the Bernesga basin there is a high
predominance of bicarbonate and calcium. This is even more marked in the Torío, and it is maintained throughout
its course. In the Bernesga, however, there are some excep-
tions, due to the effects of pollution. On the one hand, the coa1 mines at the upper parts of the basin increase the amount of sulphates, and on the other. urban waste increases
Figure 7. TI-iari~ular diagraiiis 01' tlie iiiii~orit) ~ o i i \ . 1 i ic ~1ii i ; i irter- to the samples taken in spring and summer for the Esla river and the Porma catchment.
Figure 8. Triangular diagrams of thc iiiqlorii) i o i i \ . I lic diiia rcfer to the samples taken in spring and summer for the Orbigo and the Bernesga catchment.

the proportion of chlorides and sodium, especially in the summer (fig. 8).
Phosphates and nitrates
Figures 9 and 10 show the variation in space and time in the N-nitrate and P-phosphate content during spring and
summer in the rivers in the basin which have been the object of greater study.
With regard to soluble reactive phosphate, the relative singularity of each of these rivers determines the existence
of differences in the evolution followed by the said nutrient. In the first place, a differentiation can be made between the rivers which are regulated (the Porma. the Orbigo, the Esla) and those which are not (the Curueño, the Torío, the Bernesga). In the latter, the reduction in volume during the dry season leads to a general increase in the concentration of orthophosphate particularly in the stretches downstream from the larger towns. The content
of this nutrient is particularly high in the final stretch of
BERNESGA
Figure O . Vasiiiiioii ~ ) i ~ ~ i i l c ~ 111 I I I C 1'-l'lii~rl>li,iic ii i i i l N-Xitraie along the Esla, the Bernesga and the Torío.
the Bernesga; downstream from the city of León, with a maximum of 1.24 mgll (fig. 9).
In the rivers which are regulated, the use of water retai-
ned by the dam during the summer leads to a process of
dilution, and, therefore, to reduction in the concentration of orthophosphate compared with the spring. The greater incre- ases in this nutrient correspond to areas of greater density
of population in the catchment area, together with an incre- ase in agricultural activity and the washing of the land, which is particularly high in spring in the areas around the
source. The high pollution in the final stretch of the Bernesga
clearly affects the phosphate levels found in the Esla downs- tream from the confluence of the two rivers. However, the
decline in industrial activity during the summer in which the
Esla was sampled has led to a considerable decrease in the concentration of this nutrient in relation to the values obtai- ned in spring (fig. 9).
Nitrate is the most abundant inorganic form of nitrogen in the Esla basin, with a maximum concentration in the two
A LUNA-ORBIGO A
PORMA
Figure 10. \'aii,i~ioii ~ i io l i l '~ t i 1 I I I L 1' I ' l ~ i ~ ~ ~ ~ i i , i i c , i i i i I N-Nitrate along the Luna-Orbigo. thr Porrna arid [he Cui-ueíio.

seasons under consideration of 9.99 mg/l, registered in the Curueño, and a ininimum content of 0.09 1 mg/l, measured in the Torío.
The regulation of the Esla involves very little seasonal variation and the nitrate content is practically homogeneous throughout its course. However, the other regulated rivers do not offer the same findings, as the concentration of this
nutrient in the Orbigo and the Porma undergoes an increase in the summer and there are marked fluctuations. These are occasioned by contributions particularly of agricultural
origin and by an increase in the importance of ammonia in
the sections affected by urban waste, as well as by the reten-
tion generated, in come cases, by the reservoirs. In the Bernesga and the Torío there is no clear separation
between the nitrate contents measured in spring and
summer; nos do they reach the levels found in the other
rivers in the basin. The reduction in volume in the summer cuts down the quantity of nitrate in the lower reach of both
rivers, in particular that of the Bernesga, where the condi-
tions of almost total anoxia lead to the process of denitrifi-
cation, increasing the ammonia content. The Curueño is the only river in which a clear correlation
between the nitrate content and the drained surface asea can
be observed.(fig. 10). This characteristic is more evident during the very dry season, which is not accompanied by a drastic decrease in oxygen content, and produces a higher
concentration of this nutrient in the final stretch.
ACKNOWLEDGEMENTS
We should like to thank the Joint University of León-
León Provincial Council Commission for financing the rese- arch projects that have made this study possible.
REFERENCES
ALLUE, A.J.L. 1966. Subregiones fitoclimáticas de
España. IFIE. Ed. Ministerio de Agricultura, Madrid. FERNANDEZ ALAEZ, C. 1986. Gradiente estructziral
de la iyegetación macr4fita acuática y ribereña y i~aloración de factores ecológicos en el río Bernesga, León. Tesis
Doctoral. Universidad de León, 344 pp FERNANDEZ, C., E. LUIS & M. FERNANDEZ 1986a.
Gradiente longitudinal de distribución de la vegetación ribe- seña del río Bernesga (León) en su tramo de montaña. Anales de Biología, 8: 35-44.
FERNANDEZ, C., E. LUIS & M. FERNANDEZ. 1986b. Análisis de correlaciones canónicas aplicado al estudio de
la relación entre la composición físico-química del agua y suelo aluvial en el río Bernesga (León). Limnetic,~, 2: 85-93.
FERNANDEZ, C., E. LUIS & M. FERNANDEZ 1987a. Distribución de la vegetación macrófita en la cuenca del río Orbigo (León). Actas del IV Conjireso Español de Limnolo-
~ í a : 19 1-202. FERNANDEZ, M., Y. IBAÑEZ y C. FERNANDEZ
1987b. Estudio físico-químico de los ríos de la cuenca del Bernesga (León).Actas del IV Congr-eso Español de Limno-
logía: 179- 190.
FERNANDEZ C., M. FERNANDEZ & E. LUIS 1988. Variations in time and space of some physical and chemical variables in the Bernesga river (León, Spain). Anrzls. Limnol., 24(3): 285-29 1.
FERNANDEZ, C., E. LUIS & M. FERNANDEZ. 1989. Caracterización físico-química de la cuenca del río Orbigo
(León, España). Actas del Coloquio Luso-Espanhol sobre
Ecología das Bacias Hidrograficas e Recursos Z o o l o g i < ~ ~ s ,
103-111. FERNANDEZ, C., M. FERNANDEZ, & E. LUIS. 1990.
Estudio comparado de la vegetación macrófita acuática y ribereña de las cuencas altas de los ríos Sil, Porma y Orbigo (León). Monografías del Instituto Pirenaico de Ecología, 5: 387-396.
FERNANDEZ, C., E. LUIS & M. FERNANDEZ.
(1991a). Caracterización físico-química de los ríos de la
cuenca del Porma (León). Sc,ientia Gerundensis, 16
FERNANDEZ, M . , C. FERNANDEZ & A. DEL RIO. (1991b) Estudio comparativo de los ríos de la cuenca del
Orbigo (León) en base a las variaciones en la estructura y composición de las comunidades de macrófitos. Scienriu
Gerundensis, 16. HAUBERT, M. 1975. Bilan hydrochimique d 'un basin
vrrsant de moyenne montugne: la Dranse de Bellevaz<x (ou
Brevon), Hallte Saixoie. Thkse 3e cycle. Université Pierre et
Marie-Curie, París. MARGALEF, R. 1983. Limnología. Omega, Barcelona,
1.010 pp. MEYBECK, M. 1979. Concentrations des eaux fluviales
en éléments majeur et apports en solution aux oceans. Rei,. Geol. Dyn. Geog. Phys., 21(3): 215-246.
OTTO, A. 1983. Natural site-specific hasic values of the chemism qf' (suyface) waters (Base load) in different regions of the Federal Republic qf Germany. Proceedings of a MBA Project 5 Workshop. Budapest. Octubre 1983,
14 PP.

PRESA, Y., E. LUIS & J. DE SOTO. 1987. Análisis de 1988. Análisis comparativo de los macroinvertebra- las comunidades de macroinvertebrados de la cuenca del río dos en dos subcuencas de diferente sustrato litoló- Orbigo. León. Actris del 1V Congreso Español de Lin~nolo- gico. Actas do Coloqilio Luso-Espanhol sobre ecología
gia. Sevilla, 203-213. das hacías hidrográficas e recursos zoológicos. Porto,
PRESA, Y., E. LUIS, J . DE SOTO & M. POSTIGO. 327-334.

141
THE TER, A MEDITERRANEAN RIVER SYSTEM IN SPAIN
S. Sabater, H. Guasch, E. Marti, J. Armengol, M. Vila and F. Sabater Departament d'Ecologia, Universitat de Barcelona, Avgda Diagonal 645, 08028 Barcelona, Spain
Keywords: River, Mediterranean, Water characteristics, Biological communities, Primary production, Nutrient dynamics.
ABSTRACT
Irregular hydrology and human influences are common features in the Mediterranean river systems in Spain. The two affect both the water characteristics and the distributional patterns of the organisms. Current research on functional aspects of undis- turbed tributaries is ultimately directed to give light to the behaviour of Mediterranean rivers. Analysis of nutrient dynamics and epilithon metabolism is being performed in two low-order tributaries under different environmental conditions (essentially light
and substrata). Results are discussed with respect to the special character of the Mediterranean rivers.
CLIMATE AND MAN, INTERACTING FACTORS IN SHAPING THE TER.
The Mediterranean region is one of the most densely populated and industrially developed areas in Spain. Most
of the rivers in this region, including the Ter, reveal many of
the anthropogenic disturbances derived from the use of their waters (domestic, agricultural and industrial water supplies, power generation, waste disposal, recreation ...) which have
affected the river basins for centuries. Human influence on the Ter started in the Middle Ages
with iron industries (catalan "fargues"), which caused great
deforestation in large areas. During the Industrial Revolu-
tion, cotton mills and paper mills proliferated along its
banks, attracted by the quality of its water. An inheritance of this period is the presence of small dams and channels directing the water to the production of hydraulic power.
Regulation of the Ter was completed during the 1950's by the construction of two large reservoirs in the middle stretch of the river (fig. 1) . Since then, the high development of industrial facilities and, in general, the unplanned growth of
urban centers have caused the affluence of enormous sewage
inputs to the river. Human pressure on the river is diverse
and has increased in recent times. Intensive farming, urban
development and industry depend on its waters. Up to now the Ter remains undisturbed only in some of its first and
second order tributaries. In addition to this considerable anthropogenic impact,
fluctuations in water discharge due to the Mediterranean
climate creates variable conditions in the river. Summer is
usually very dry period while high discharge (due to heavy
rain) is common in autumn. The location of the Ter head- waters in the Pyrenees, where the discharge pattern is strongly regulated by snow accumulation and melting means
lower flow in winter and subsequent increase in spring. Although the annual average discharge is 554,6 f 36
~ m ' , this value varies considerably in successive years (fig. 2). This is a common feature in the Mediterranean region.
The water discharge in the Ter shows a very seasonal cycle,
but in practice climatic events introduce a source of varia-
Figure 1. Location of the htudird san~pI~ng-po~nr\ 111 rhc Icl K ~ l e r . The main geological features of the basin, and towns with more than 10,000 inhabitants are shown.
Limnetica. 8: 141-149 (1992)
0 Asociac16n Espafiola de Limnologia, Madrid. Spain

bility in runoffs (ARMENGOL et al.. 1991). Short fluctua-
tions of 3, 6 and 12 months determine the seasonality of the
annual cycle. Annual fluctuations have periods of 5.5, 8.6, 10.1 and 11.7 years. These longer term fluctuations can be
considered as an expression of cycles of 7 and 11 years which are characteristic of many climatic events in the
Mediterranean region. The state of the water and the biolo- gical communities in the river are obviously affected by this temporal varition.
PHYSIOGRAPHICAL CHARACTERISTICS OF THE TER.
The River Ter drains about 3,010 km2 of the north part of Catalonia (NE Spain). It rises at 2,400 m a.s.1. in the Pyre- nees and flows 208 km down to the Mediterranean sea. During its course i t receives a large number of tributaries.
The largest in the upper part is the river Freser (4th order). The upper Ter and Freser basins receive a high number of small streams feeding in the Pyrenees. In its middle stretch, the Ter receives small 2nd to 3rd order streams, coming from middle-mountain basins. Three reservoirs (Sau, Susqueda and El Pasteral; joint capacity of 0.402 km3; average residence time of the water of 11 7, 143 and 1.7 days respectively) are also located in this stretch (fig. 1). The
reservoirs exert a direct influence on the lower part of the
river (PUIG er al.. 1987). Before it reaches the sea the Ter receives the Onyar River, a 4th. order stream, which has a marked mediterranean regime, but does not highly alter the
Ter hydrography. When the Ter flows out into the Medite- rranean sea, it is a 5th order stream.
The middle and low parts of the main stretch of the river, including a large number of tributaries, drain calcareous areas.
However, the Ter headwaters and some of the tributaries in the
middle stretch flow over granites and slates, while a few tribu- taries drain areas which are rich in gypsum (fig. I ).
WATER CHARACTERISTICS AND SEDIMENT TRANSPORT
In the Ter basin the high diversity of land uses, vegeta-
tion cover and soil types cause differential influences on the physical and chemical features of the water.
The Ter waters are rich in Ca(HC03), (SABATER, 1988). Most of its basins have alkaline-earth bicarbonate waters: hydrocarbonate represents 63 % of the total anions
and calcium 60 % of the total cations. Other ions reach rele- vant co~icentrations in particular places along the river: sulp-
hate constitutes on the average 24 % and chloride only 13 % of the total anions. These last two anions are only abun- dant in particular parts of the river. Water conductivity rises
from less than 100 ks cm-' in the headwaters to more than
1000 ps cm-' in the lower reaches. Such a general increase
may be related both to the rock weathering and to human activity, this latter leading to particular increases in chlo-
ride, sodium and potassium.
Nutrient concentrations in the Ter waters are mainly rela- ted to land use. This defines important spatial variations throughout the basin. On the other hand, temporal variations
in nutrient concentrations are highly dependent on hydrolo- gical features. Both organic and inorganic forms of dissol-
ved phosphate decrease during high discharge, while the particulate form exhibits a slight increase in such periods
(fig. 3b). During high water levels there is an important
increase in nitrate whereas ammonium and nitrite decrease. Ammonium dynamics is only partially regulated by the
water discharge; increases are related to pollutant inflows. The organic forms of nitrogen predominate during high
discharge (fig. 3a). In the upper part of the river where the bedrock is silice-
ous and human activity is low, the content of total dissolved
solids (TDS) is very low, usually less than 20 mg I-'. TDS
values are high in sites upstream from the reservoirs (up to 80 mg I- ' ) . TDS reaches extremely high values (up to 600
mg 1.') in a polluted tributary which enters the river at the jonction with the reservoirs. Reservoirs act as decanting
lagoons; TDS at their outflow has been reduced to 10 mg I-'. Values of TDS in the tributaries vary according to their geochemical features (siliceous vs. calcareous) and the extent of forest cover.
The Ter River drains unforested areas which are inters-
persed among well forested areas. The basins of the Ter
F ig~ t t c 2 . A t ~ i ~ ~ t i t l ; t \~,t~t;c cIt~cl~iti:~,\ cltiriiig 111c [ I L Y I ( I ( / 1054 10 I O X X . The amplitude of the 95% confidence intervals cvidenceb the impor- tance of the seasonal variations.
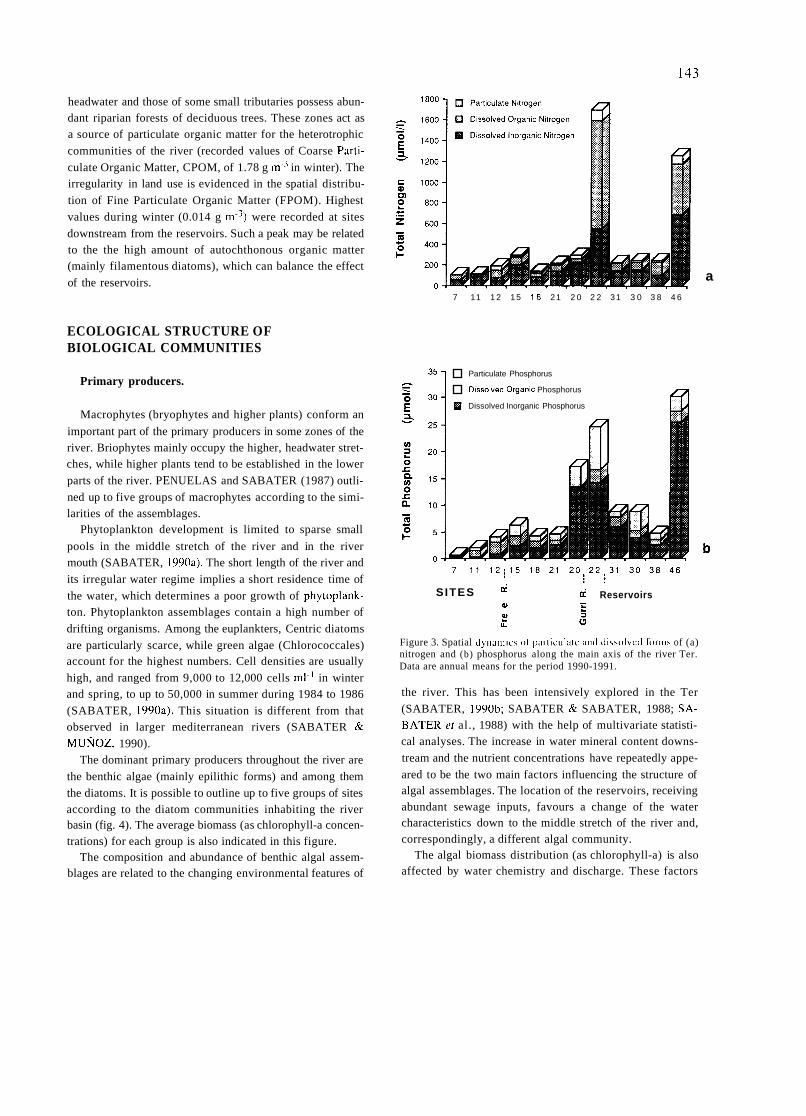
headwater and those of some small tributaries possess abun- dant riparian forests of deciduous trees. These zones act as a source of particulate organic matter for the heterotrophic
communities of the river (recorded values of Coarse Parti-
culate Organic Matter, CPOM, of 1.78 g m-' in winter). The irregularity in land use is evidenced in the spatial distribu-
tion of Fine Particulate Organic Matter (FPOM). Highest
values during winter (0.014 g m-') were recorded at sites
downstream from the reservoirs. Such a peak may be related to the the high amount of autochthonous organic matter (mainly filamentous diatoms), which can balance the effect
of the reservoirs. a 7 11 1 2 1 5 1 8 2 1 2 0 2 2 3 1 3 0 3 8 4 6
ECOLOGICAL STRUCTURE OF BIOLOGICAL COMMUNITIES
Primary producers.
Macrophytes (bryophytes and higher plants) conform an
important part of the primary producers in some zones of the river. Briophytes mainly occupy the higher, headwater stret- ches, while higher plants tend to be established in the lower
parts of the river. PENUELAS and SABATER (1987) outli-
ned up to five groups of macrophytes according to the simi- larities of the assemblages.
Phytoplankton development is limited to sparse small
pools in the middle stretch of the river and in the river mouth (SABATER, 1990a). The short length of the river and
its irregular water regime implies a short residence time of
the water, which determines a poor growth of phytoplank- ton. Phytoplankton assemblages contain a high number of
drifting organisms. Among the euplankters, Centric diatoms
are particularly scarce, while green algae (Chlorococcales) account for the highest numbers. Cell densities are usually
high, and ranged from 9,000 to 12,000 cells ml-' in winter and spring, to up to 50,000 in summer during 1984 to 1986
(SABATER, 1990a). This situation is different from that observed in larger mediterranean rivers (SABATER &
MUNOZ, 1990). The dominant primary producers throughout the river are
the benthic algae (mainly epilithic forms) and among them
the diatoms. It is possible to outline up to five groups of sites according to the diatom communities inhabiting the river basin (fig. 4). The average biomass (as chlorophyll-a concen- trations) for each group is also indicated in this figure.
The composition and abundance of benthic algal assem- blages are related to the changing environmental features of
35 7 Particulate Phosphorus
I D~ssolved Organ~c Phosphorus n 1 W Dissolved Inorganic Phosphorus
SITES rii 0)
2 LL
Reservoirs
Figure 3. Spatial dq~ la~ i i l c \ ot pal-t~culate a n d d~ssolved forms of (a) nitrogen and (b) phosphorus along the main axis of the river Ter. Data are annual means for the period 1990-1991.
the river. This has been intensively explored in the Ter
(SABATER, 1990b; SABATER & SABATER, 1988; SA- BATER et al., 1988) with the help of multivariate statisti- cal analyses. The increase in water mineral content downs-
tream and the nutrient concentrations have repeatedly appe-
ared to be the two main factors influencing the structure of algal assemblages. The location of the reservoirs, receiving abundant sewage inputs, favours a change of the water characteristics down to the middle stretch of the river and, correspondingly, a different algal community.
The algal biomass distribution (as chlorophyll-a) is also affected by water chemistry and discharge. These factors

interact with the natural dynamics of the photosynthesis of the benthic algae (SABATER, 1989). Spatial differences in chl-a along the river are higher in summer than in spring
(SABATER & SABATER, 1991), the two periods of highest production. Differences in discharge, associated with the Mediterranean climate (high in spring, reduced in summer), directly affect the algal communities (scouring in spring, accumulation in summer). Moreover, discharge can also contribute to the pollution-related patchy distribution of nutrients along the river, which can favour local increases in benthic algal biomass in summer.
Secondary producers.
Macroinvertebrate assemblages in the Ter are affected by similar environmental factors to those affecting the algal
communities. The longitudinal organization of the macroin- vertebrate guilds is according to the type of food available,
but is altered by the pollution of the waters. This ordination is
also affected by the reservoirs, which modify water conditions (PUIG et al., 1987). Gathering-collectors and shredders are dominant in the headwaters. The invertebrate predators are
more common in this part of the river. Filter-collectors are dominant in the headwater sites only when influenced by anthropogenic activities, with a lower proportion of grazers and scrapers. The macroinvertebrate which inhabit the middle
stretch of the river are mainly grazers and filter-feeders, while the collector-gatherers become reduced in comparison with the upper stream zones.
The community inhabiting below the reservoirs is made
up of large populations of benthic invertebrates feeding
upon nutrient rich seston (ARMITAGE, 1984). Algae, mosses and vascular plants, which are abundant in this stretch of the river, may be fed upon directly or through
breakdown and decomposition, while the release of detritus
particles becomes a food source for filter-feeders
Hydrurus foetidus Group 1 Ulothrix zonata
Ceratoneis arcus Group 2 Diatoma hiemale
Meridion circulare Group 3 /.-....- ..........-.....+...
- . , ,Chl-a: 20-100 mg m' /- ">%.---a N ,,..... -.-../
Group 5
~.\/---.-'
Gomphonema parvulum Nitzschia palea
/ Cyclotella meneahiniana n-
Navicula sp. pl. Nitzschia sp. pl. Gomphonema sp. pl. Cladophora glomerata
Chl-a: 1 12-1 33 (400) rng m-'
Cymbella sp
.. ~avicu la areaaria / W
Denticula tenuis var. crassula I Fragilaria sp. pl. Diploneis oblongella Amohora oediculus
Chl-a: 200-500 rng m-' ~it=schia dissipata Melosira varians Bangia atropurpurea
Figure 4. Representation of the five groups of sampling htatlon\ acco~.d~~ig lo ~l ic .~r \~nr~l:u-~t~c. \ in the diatom assemblages. The most characte- ristic species in every group are listed. Usual values of benthic algae biomasa (in mg of chl-a m-2) during the period 1982-1983 are also indi- cated.

(WALLACE & MERRITT, 1980), or may be deposited into the hyporheic zone where they will provide a nutrient-rich habitat for the hyporheos community (SABATER & VILA,
1990). The most abundant species within this group are
filter-feeding tricoptera which depend on epilimnial suspen- ded matter (WARD & STANFORD, 1979).
Hyporheos.
The hyporheic fauna in the Ter River is dominated by insects, (particularly Chironomidae), Crustacea and Oligocha-
eta. The different proportion among groups of taxa at each site suggests a relationship between hyporheos requirements and
physicochemical features in interstitial waters (SABA- TER,1987; SABATER & DE MANUEL, 1988). Crustacea,
mainly Harpacticoida and Cyclopoida species, are abundant in
the interstitial sites of the headwaters. The Isopoda PI-nasellus
phreaticus and Microcharon margalefi were first described from these sites (SABATER & DE MANUEL, 1988). Ostra-
coda and Chydoridae become more abundant in clean calca-
reous tributaries. In sites located further downstream, the occu- rrence and abundance of these different crustacean groups decrease in number of individuals and taxa. One reason for this is that inputs of fine particulate organic matter coming
from anthropogenic activities. cause clogging of the superfi-
cial layer of sediments, thus decreasing the oxygen content noticeably. Only some animal groups, such as Nematoda and
Oligochaeta, can resist these adverse conditions. In sites strongly affected by pollution there is a striking decrease in
interstitial fauna density which, in the worst case, is absent. In sites below the reservoirs, the interstitial environment is
not subjected to a such physical stress. In comparison to
upstream sites, the number of Crustacean species and other
groups of meio-fauna (e.g., Rotifera, Tardigrada, and hydrac-
nellid mites) become more abundant and persistent in time. However, the most abundant interstitial animal groups occu- rring below the reservoirs are cyclopoids, oligochaets and chyronomids. This fauna feeds on the fine particulate organic
matter accumulated in the sandy-gravel matrix.
CURRENT RESEARCH IN THE RIVER TER.
Production of epilithic communities in undisturbed tributaries.
of the river Ter. Recirculating light and dark metabolism chambers placed in situ (BOTT et al . , 1978) are being used to study the oxygen and carbon dioxide metabolism of two
streams of similar geomorphological and physical features
(2nd order, low nutrient content of the waters) but of diffe- rent geology. A phtosynthetic quotient of ca. 0.4 has been calculated for the epilithic community in the two streams,
and used to convert oxygen data tocarxbon units.
The first river, La Solana, is calcareous, colonized by
carbonate crusts of filamentous cyanobacteria (SABATER 1989). Net daily metabolism (NDM) during 1990 - 1991 was of 0.69-0.73 g C m-' day-' during summer and autumn
and 0.1 1 g C m-' day-' in spring, but negative in winter (- 0.16 g C m-' day-' ) (fig. 5) . The ratio between the Gross Primary Production (GPP) and respiration (R) was maxi- mum in autumn (2.69) and minimum in winter (0.57).
Although conclusions are still preliminary, differences in the
ratio suggest a very different behaviour of the epilithic community during the two periods of the year, possibly rela- ted to the lower activity of the algal component of the
community during winter. Results are different in the siliceous tributary, La Major.
This is a siliceous, well forested stream. Shaded reaches are colonized by conspicuous patches of the Rhodophycean
Hildenhr-andia I-ivular.is, which is the most abundant compo-
nent of the epilithic community during winter. In autumn
and early spring this alga is partially covered by a thin diatom layer. NDM increased from winter (0.24 g C m-'
NET DAILY METABOLISM
U
-2001 summer ' autumn winter spring 1
I 0 NDM Solana m NDM R.Major 1
Measurement of oxygen production and respiration of epilithic communities is in progress in two small tributaries
Figut-e 5 . l ' i ~ l u t . \ ot net d a ~ l ) t~letaholt\rn (111 2 C ni-2 clay-I) in La Major (siliceous) and La Solana (calcareous) streams during winter, spring and summer of the period 1990- 199 1.
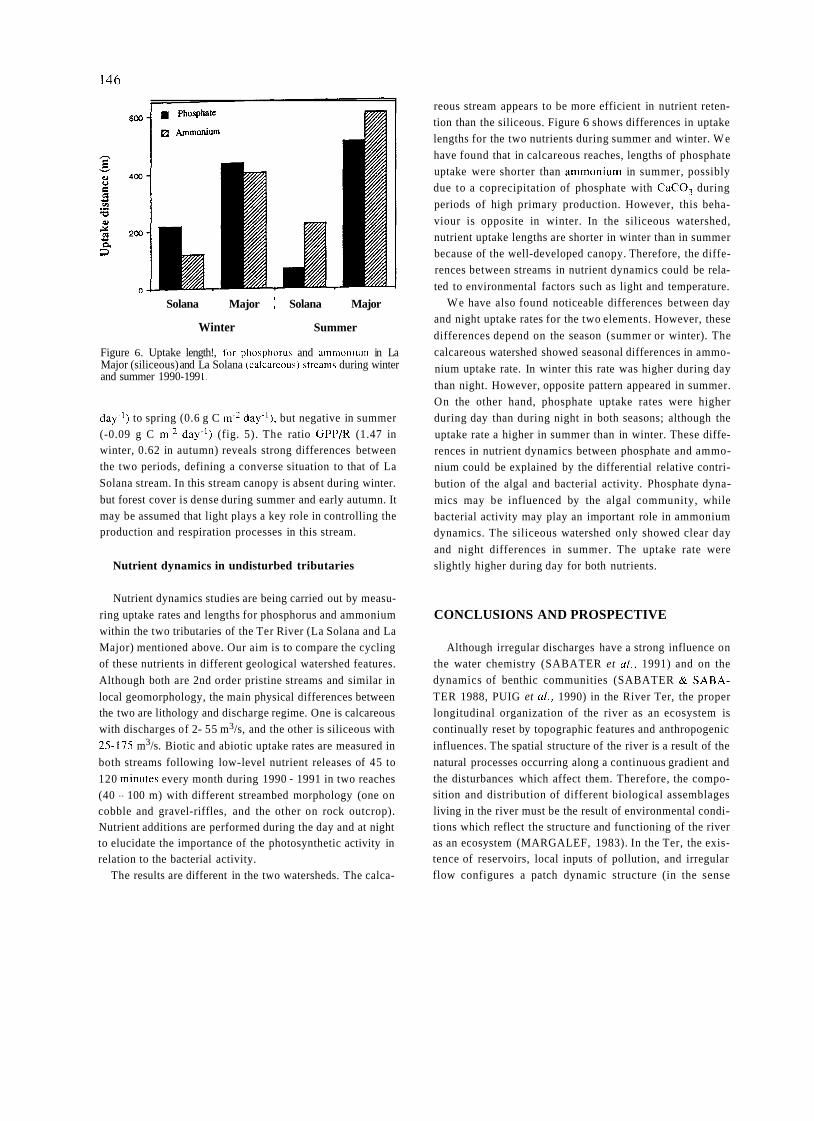
Solana Major : Solana Major
Winter Summer
Figure 6. Uptake length!, fol. pho5phorus and a~nmoniuin in La Major (siliceous) and La Solana (calcareous) streams during winter and summer 1990- 199 1 .
day-') to spring (0.6 g C m-2 day-'), but negative in summer (-0.09 g C m-2 day-l) (fig. 5). The ratio GPP/R (1.47 in winter, 0.62 in autumn) reveals strong differences between the two periods, defining a converse situation to that of La
Solana stream. In this stream canopy is absent during winter.
but forest cover is dense during summer and early autumn. It may be assumed that light plays a key role in controlling the production and respiration processes in this stream.
Nutrient dynamics in undisturbed tributaries
Nutrient dynamics studies are being carried out by measu-
ring uptake rates and lengths for phosphorus and ammonium within the two tributaries of the Ter River (La Solana and La Major) mentioned above. Our aim is to compare the cycling of these nutrients in different geological watershed features.
Although both are 2nd order pristine streams and similar in
local geomorphology, the main physical differences between the two are lithology and discharge regime. One is calcareous with discharges of 2- 55 m3/s, and the other is siliceous with
25-175 m3/s. Biotic and abiotic uptake rates are measured in
both streams following low-level nutrient releases of 45 to
120 minutes every month during 1990 - 1991 in two reaches (40 - 100 m) with different streambed morphology (one on cobble and gravel-riffles, and the other on rock outcrop). Nutrient additions are performed during the day and at night to elucidate the importance of the photosynthetic activity in relation to the bacterial activity.
The results are different in the two watersheds. The calca-
reous stream appears to be more efficient in nutrient reten- tion than the siliceous. Figure 6 shows differences in uptake lengths for the two nutrients during summer and winter. We
have found that in calcareous reaches, lengths of phosphate uptake were shorter than anlmonium in summer, possibly due to a coprecipitation of phosphate with C a C 0 3 during
periods of high primary production. However, this beha-
viour is opposite in winter. In the siliceous watershed, nutrient uptake lengths are shorter in winter than in summer because of the well-developed canopy. Therefore, the diffe- rences between streams in nutrient dynamics could be rela-
ted to environmental factors such as light and temperature. We have also found noticeable differences between day
and night uptake rates for the two elements. However, these
differences depend on the season (summer or winter). The
calcareous watershed showed seasonal differences in ammo-
nium uptake rate. In winter this rate was higher during day
than night. However, opposite pattern appeared in summer. On the other hand, phosphate uptake rates were higher during day than during night in both seasons; although the
uptake rate a higher in summer than in winter. These diffe- rences in nutrient dynamics between phosphate and ammo- nium could be explained by the differential relative contri-
bution of the algal and bacterial activity. Phosphate dyna-
mics may be influenced by the algal community, while bacterial activity may play an important role in ammonium dynamics. The siliceous watershed only showed clear day
and night differences in summer. The uptake rate were slightly higher during day for both nutrients.
CONCLUSIONS AND PROSPECTIVE
Although irregular discharges have a strong influence on the water chemistry (SABATER et ul., 1991) and on the dynamics of benthic communities (SABATER & SABA-
TER 1988, PUIG et ul., 1990) in the River Ter, the proper longitudinal organization of the river as an ecosystem is continually reset by topographic features and anthropogenic
influences. The spatial structure of the river is a result of the
natural processes occurring along a continuous gradient and the disturbances which affect them. Therefore, the compo- sition and distribution of different biological assemblages living in the river must be the result of environmental condi- tions which reflect the structure and functioning of the river as an ecosystem (MARGALEF, 1983). In the Ter, the exis- tence of reservoirs, local inputs of pollution, and irregular flow configures a patch dynamic structure (in the sense
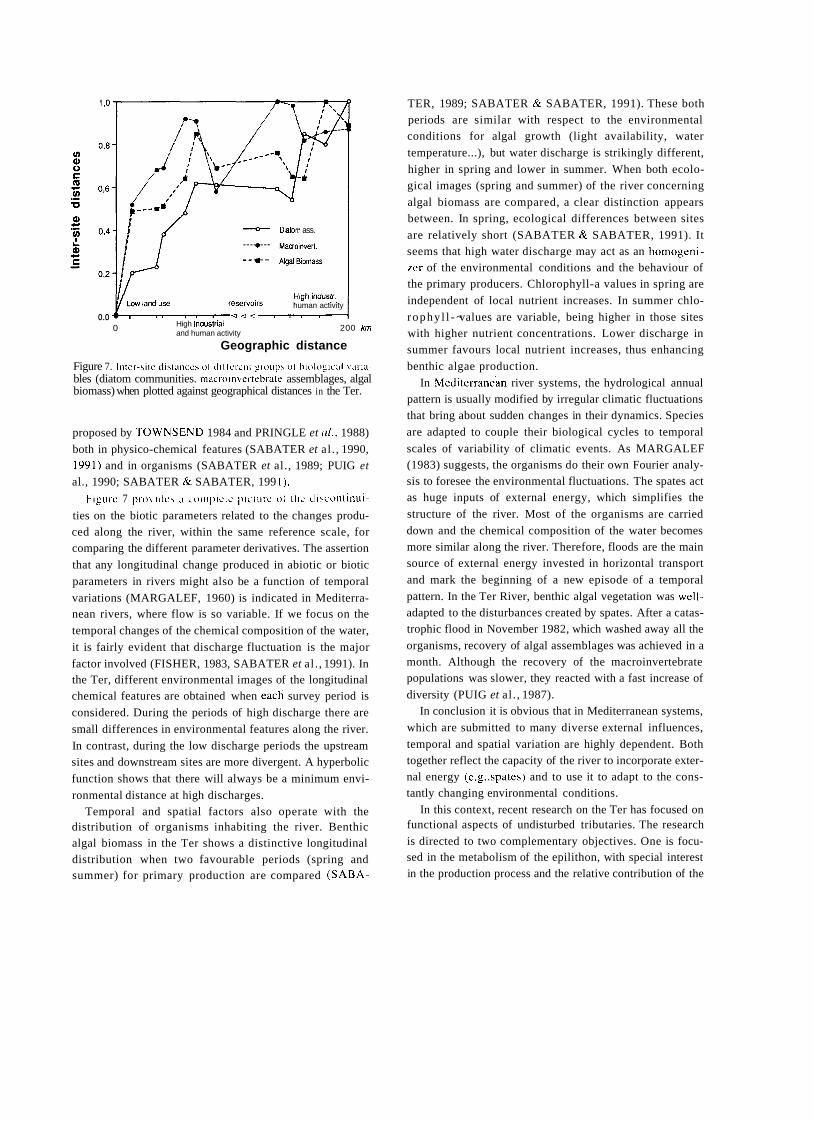
- Diatom ass.
reservoirs human activity
0.0 7- 4 -a - 0 High industrial
and human activity 200 km
Geographic distance
Figure 7. Inter-\ire d~stanceh ol tl11lc1c111 ~ I O L I ~ 01 I ~ I O ~ O ~ I C ; I ~ ~ a i . 13 -
bles (diatom communities. macroinvertebrate assemblages, algal biomass) when plotted against geographical distances in the Ter.
proposed by TOWNSEND 1984 and PRINGLE et al., 1988)
both in physico-chemical features (SABATER et al. , 1990, 1991) and in organisms (SABATER et al., 1989; PUIG et
al., 1990; SABATER & SABATER, 199 1).
t12~11.c 7 pro\ 1c1c\ ;I L ~ O I I I ~ ~ C ' I ~ I I I C I L I I . ~ 0 1 1 1 1 ~ ' ~ l ~ \ c o n ~ i n u i -
ties on the biotic parameters related to the changes produ- ced along the river, within the same reference scale, for comparing the different parameter derivatives. The assertion
that any longitudinal change produced in abiotic or biotic
parameters in rivers might also be a function of temporal
variations (MARGALEF, 1960) is indicated in Mediterra- nean rivers, where flow is so variable. If we focus on the
temporal changes of the chemical composition of the water,
it is fairly evident that discharge fluctuation is the major
factor involved (FISHER, 1983, SABATER et al. , 1991). In the Ter, different environmental images of the longitudinal chemical features are obtained when each survey period is
considered. During the periods of high discharge there are
small differences in environmental features along the river.
In contrast, during the low discharge periods the upstream sites and downstream sites are more divergent. A hyperbolic
function shows that there will always be a minimum envi-
ronmental distance at high discharges. Temporal and spatial factors also operate with the
distribution of organisms inhabiting the river. Benthic algal biomass in the Ter shows a distinctive longitudinal distribution when two favourable periods (spring and summer) for primary production are compared (SABA-
TER, 1989; SABATER & SABATER, 1991). These both periods are similar with respect to the environmental conditions for algal growth (light availability, water
temperature...), but water discharge is strikingly different,
higher in spring and lower in summer. When both ecolo- gical images (spring and summer) of the river concerning
algal biomass are compared, a clear distinction appears between. In spring, ecological differences between sites
are relatively short (SABATER & SABATER, 1991). It seems that high water discharge may act as an homogeni- zer of the environmental conditions and the behaviour of the primary producers. Chlorophyll-a values in spring are
independent of local nutrient increases. In summer chlo-
rophyl l -~ values are variable, being higher in those sites with higher nutrient concentrations. Lower discharge in summer favours local nutrient increases, thus enhancing
benthic algae production.
In ~edi terranean river systems, the hydrological annual pattern is usually modified by irregular climatic fluctuations that bring about sudden changes in their dynamics. Species
are adapted to couple their biological cycles to temporal
scales of variability of climatic events. As MARGALEF (1983) suggests, the organisms do their own Fourier analy- sis to foresee the environmental fluctuations. The spates act as huge inputs of external energy, which simplifies the
structure of the river. Most of the organisms are carried
down and the chemical composition of the water becomes more similar along the river. Therefore, floods are the main source of external energy invested in horizontal transport
and mark the beginning of a new episode of a temporal
pattern. In the Ter River, benthic algal vegetation was well- adapted to the disturbances created by spates. After a catas- trophic flood in November 1982, which washed away all the
organisms, recovery of algal assemblages was achieved in a month. Although the recovery of the macroinvertebrate populations was slower, they reacted with a fast increase of
diversity (PUIG et al. , 1987). In conclusion it is obvious that in Mediterranean systems,
which are submitted to many diverse external influences,
temporal and spatial variation are highly dependent. Both together reflect the capacity of the river to incorporate exter- nal energy (e.g.,spates) and to use it to adapt to the cons- tantly changing environmental conditions.
In this context, recent research on the Ter has focused on functional aspects of undisturbed tributaries. The research
is directed to two complementary objectives. One is focu- sed in the metabolism of the epilithon, with special interest in the production process and the relative contribution of the

algal and bacterial component. The second is the analysis of nutrient dynamics in the river under different environmental conditions (essentially light and substrata). Starting in undisturbed streams in order to evaluate the magnitude of the natural processes, our aim is to extend the approach to anthropogenically disturbed reaches of the river. Future research will also be directed to the specification of the role of the bacterial community in the processing both of allocht- honous and autochthonous material in the river.
ACKNOWLEDGEMENTS
This research has been supported by the funding of the CICYT grants nV78-81 and NAT 89-0886. H. Guasch and E. Marti benefit a FP1 grant to participate in the last project.
REFERENCES
ARMENGOL, J., SABATER,S., VIDAL, A. & SABA- TER, F., 1991. Using the rescaled range analysis for the study of hydrological records: the River Ter as an example. Oecologia Aquaticu,lO (in press).
ARMITAGE, R.D., 1984. Environmental changes indu- ced by stream regulation and their effect on lotic macroin- vertebrate communities. In: A.Lillehammer and S.J.Saltveit (Editors), Regulut. Rivers. Universitesforlaget, Oslo, pp. 139-165.
BOTT, T.L., BROCK, J.T., CUSHING,C.E., GREGORY S.V., KING, D. & PETERSEN, R.C., 1978. A comparison of methods for measuring primary productivity and commu- nity respiration in streams. Hydr-ohiologia, 60: 3-12.
BOTT, T.L., BROCK, J.T., DUNN, C.S., NAIMAN, R.J., OVINK. R.W. & PETERSEN, R.C., 1985. Benthic commu- nity metabolism in four temperate stream systems: An inter- biome comparison and evaluation of the river continuum concept. Hydrohiologia 123: 3-45.
CUMMINS, K.W., MINSHALL, G.W., SEDELL, J.R., CUSHING, C.E. & PETERSEN, R.C., 1984. Stream ecosys- tem theory. Verh. lnternar. Verein. Limnol., 22: 1818-1827.
CUSHING,C.E., 1988. Allochthonous detritus input to a small, cold desert spring-stream. Verh. Interxat. Ver-ein.
Theor. Limnol., 23: 1107-1 113. FISHER, S.G., 1983. Succession in streams. In:
J.R.Barnes & G.W.Minshal1 (Editors), Stream ecology : application and testing of general ecolo,gical theory. Plenum Press, New York, pp.7-27.
LIKENS, G.E., BORMANN, F.H., PIERCE, R.S., EATON, J.S. & JOHNSON, N.M., 1977. Biochemistry q f a
forested ecosystem. Springer, New York. 147pp. MARGALEF, R., 1983. Lin~nologiu. Omega, Barcelona.
101opp. MINSHALL, G.W., 1978. Autotrophy in stream ecosys-
tems. Bioscience, 28: 767-771. PENUELAS, J. & SABATER, F., 1987. Distribution of
macrophytes in relation to environmental factors in the Ter River, NE Spain. Internat. Rev. Gesamt. Hydrohiol., 72: 41-58.
PRAT, N., PUIG, M.A., GONZALEZ, G., TORT, M.I. &
ESTRADA, M,, 1984. The Llobregat : a Mediterranean river fed by the Pyrenees. In: B.A.Whitton (Editor), The ecology of E~lropean Ri1,ers. Studies in Ecology, Blackwell, London,
pp.527-552. PRINGLE, C.M., NAIMAN, R.J., BRETSCHKO, G.,
KARR, J.R., OSWOOD, M.W, WEBSTER, WELCOMME, R.L. & WINTERBOURN, M.J., 1988. Patch dynamics in lotic systems : the stream as a mosaic. .l. North Am. Bent-
hol. Soc., 7: 503-524. PUIG, M.A.,ARMENGOL, J., GONZALEZ, G., PENUE-
LAS, J, SABATER, S. & SABATER, F., 1987. Chemical and biological changes in the Ter River induced by a series of reser- voirs. In: J.F.Craig and J.B.Kemper (Editors), Advances in rugu- lated stream ecology. Plenum Press, New York, pp.373-382.
PUIG, M.A., GONZALEZ, G. & RECASEN S,L., 1987a. Las comunidades de macroinvertebrados del rio Ter, con especial referencia a Plecopteros, Efemeropteros, Tricopte- ros y Simulidos. Limnerica 3: 125-132.
PUIG, M.A., ABOAL, M. & SOSTOA, A., 1990. New approaches to Mediterranean fluvial communities. Oecolo-
~ i r r Aqr~crfica, 10 (in press). RESH, V.H., BROWN, A.V., COVICH, A.P., GURTZ,
M.E., LI,H.W., MINSHALL, G.W., REICE, S.R., SHEL- DON, A.L., WALLACE, J.B. & WISSMA R,R., 1988. The role of disturbence theory in stream ecology. .l. North Am.
Benrhol.Soc., 7: 433-445. SABATER, F. 1987. On the interstitial Cladocera of the
River Ter (Catalonia,NE Spain) with a description of the male of Alona phrerrticcr. Hydrohiologia, 144: 5 1-62.
SABATER, F., 1988. The geochemical character of the River Ter watershed. Oc~cologia Aquatics, 9: 77-87.
SABATER, F. & ARMENGOL,J., 1986. Chemical cha- racterization of the Ter River. Limnetica, 2: 75-84.
SABATER, F. & DE MANUEL,J., 1988. Interstitial isopoda of the Ter River (Catalonia,NE Spain) and descrip- tion of two new species, Mi~,rocharon margalefi n.sp. and PI-oa.sellus phreaticus n.sp.. Stygologia, 4: 267-278.

SABATER, F. & VILA, P., 1990. The hyporheic zone considered as an ecotone. Oecologia Ayuarica, 10 (in press).
SABATER, F., ARMENGOL, J. & SABATER, S., 1989. Measuring discontinuities in the Ter River. Regul. Ri~,ers , 3: 133-142.
SABATER, F., SABATER, S. Br ARMENGOL, J., 1990. Chemical characteristics of a Mediterranean river basin as influenced by land uses in the watershed. Wat. Res., 24: 143.155.
SABATER, F., ARMENGOL, J. & SABATER, S., 199 1. Physico-chemical disturbances associated with spatial and temporal variation in a Mediterranean river. J. North Am.
Benthol. Soi,., 10: 2- 13. SABATER, S., 1988. Composicion y ciclo de pigmentos
clorofilicos en las poblaciones del fitobentos del rio Ter durante un period0 de nueve meses. Oecologia Aq~(alica, 9: 61-75.
SABATER, S., 1989. Encrusting algal assemblages in a
Mediterranean river basin. Arch. Hydrohiol., 114: 55-573. SABATER, S., 1990a. Phytoplankton composition in a
medium-sized Mediterranean river : the Ter (Spain). Limne-
tics, 6:47-56. SABATER, S., 1990b. Composition and dynamics of a
highly diverse diatom assemblage in a limestone stream. Hydrohiologia, 190: 43-53.
SABATER, S. & SABATER, F., 1988. Diatom assem- blages in a Mediterranean river basin. Arch. Hydrohiol., 1 1 1 : 397-408.
SABATER, S. & MUNOZ, l . , 1990. Successional dyna- mics of the phytopiankton in the lower part of the River Ebro. J. Plankton Res., 3: 573-592.
SABATER, S. & SABATER, F., 1991. Longitudinal changes of benthic algal biomass in a Mediterranean river during two high production periods. Arch. Hydr-ohiol. (in press).
SABATER, S., SABATER, F. & ARMENGOL, J., 1988. Relationships between diatom assemblages and physico- chemical variables in the River Ter (NE, Spain). Internut.
Rev. Gesamt. Hydrohiol., 73: 171 - 179. TOWNSEND, C.R., 1984. The patch dynamics concept
of stream community ecology. .l. North Am. Benthol. Soc.,
8: 36-50. VANNOTE, R.L., G.W. MINSHALL, K.W. CU-
MMINS, J.R. SEDELL & C.E. CUSHING. 1980. The ri- ver continuum concept. Can. J O L I ~ . Fish. Aquaf . Sc. 37: 130-137.
WALLACE, J.B. & MERRITT, R.W., 1980. Filter feeding ecology of aquatic insects. Anttl. Rev. Etzromol., 25: 103.132.

ECOLOGY OF SPANISH SEMIARID STREAMS
M.R. Vidal-Abarca, M.L. Suárez and L. Ramírez-Díaz Animal Biology and Ecology Department. University of Murcia. Campus de Espinardo. 30100 Murcia, Spain
Keywords: Semiarid streams, Perturbations and disturbances, Floods, Aquatic invertebrates, Ecological unpredictability.
ABSTRACT
This chapter analyses the limnological peculiarities of Spanish semiarid streams. Their hydrological regime vary on spatial
and temporal scales depending on precipitation patterns. Their hydrological cycles are severely disturbed, in a fairly unpredic-
table way, by flash i'ioods.
Variations in physico-chemical parameters, especially on a temporal scale are highly variable due to floods, which make it difficult to stablish hydrochemical models in the short term. In response to this regime of disturbances, communities of aqua-
tic organisms show distinct ecological characteristics. The adaptative strategies of organisms vary considerably and seem to
respond to the unpredictability of the disturbances. Additionaly, the perspectives and future research lines of the group on the Ecology of Inland Waters of the University of
Murcia are commented.
INTRODUCTION
The term "semiarid" is applied in this chapter to regions
where, for different reasons, the balance of water is nega- tive, creating an environmental stress which, unlike those occurring in arid lands or deserts, is neither permanent nor
predictable. This condition is associated to particular clima-
tic characteristics, primarily the pattern of rains, which determines a very characteristic hydrological regime on a
pluriannual scale. The streams in these regions are subject to natural disturbances (droughts and floods), as a conse-
quence of their irregular regime.
In Spain, two regions exist with semiarid climatic charac- teristics, situated in the South-east and in the central region
of the Ebro depression (fig. 1). The latter, however, due to
different topographical, geological and hydrological factors
constitutes an endorrheic region disconnected from the fluvial network (MONTES & MARTINO, 1987). In the
Iberian South-east, the more abrupt and erosive topography
makes drainage exorrheic giving rise to semiarid streams that are the subject of this chapter.
Very few bibliographic references exist concerning the structure and functioning of Spanish semiarid streams. Only scattered information is available about the changes of the physical or chemical parameters after a flood (AVILA,
1984; ESCARRE et a l . , 1984; VIDAL-ABARCA, 1985; SUAREZ, 1986; PUIG et al . , 1987), or the washing and
sweeping away of algae (SABATER, 1981) and macroin-
vertebrates populations (PRAT et al., 1986, PUIG er al. ,
1990; SOLER, 1991). The group on Ecology of lnland Waters of Murcia
University has recently published some of the results of its
studies on semiarid streams, of the Iberian South-east (ORTEGA, 1988; ORTEGA et al., 1988; 1991a,b; VIDAL- ABARCA, 1990, etc).
ENVIRONMENTAL CHARACTERISTICS OF SPANISH SEMIARID REGIONS AND THEIR LIMNOLOGICAL IMPLICATIONS
Any discussion on semiarid streams has to deal with the
temporal and even the ephemeral character of the water- flow. However, these streams are not found exclusively in
these semiarid regions. In Spain, as in other parts of the
world, there is a great variety of terms that refer to the inte- rruption of the water-flow in a channel ("torrent", "torren- tera" and "riera" in the Catalan countries; "torrents" in the Balearic Islands; "yasa" in Aragon; "clamor" in the Aragon region of Monegros, and other of a more generalized use,
Liinnetica, 8: 151-160 (1992)
O Asociación Española de Limnología, Madrid. Spain

M,, hnual P) Interannual N" days Mean Annual precipitation [rn) Variability of P ( X ) with rain T* ( O C )
Semiarid 330 35 50 16
Arid 220 40 30 18
~ I C L I I C l . Ai-id aiid \riiiini-id Spaiii\li icgioiis niitl aoiiic. cliniatic para- meters (modified from GARCIA DE PEDRAZA, 1989).
such as "cárcava", "barranco", "rambla", etc; ARENILLAS & SAENZ, 1987). +Besides, in the semiarid Iberian South- east, streams with permanent water, are also found.
The singularity of these streams is due to their hydro-
logical regimes, which are very variable on both spatial
and temporal scales. On a spatial scale it is possible to distinguish channels with permanent water (with a conti- nuous flow during complete hydrological cycles), chan- nels with temporary water (that remain dry os have a
discontinuous flow in the summer season) and ephenieral channels (that only carry rapidly flowing water after strong storms). Moreover the same channel may Iiave sections of different typology. Figure 2 shows the drai-
nage network of the Segura river basin (Iberian South- east) according to the time during which water remains in its channels. As can be seen al1 possible combiriations and
situations are observed. Temporal variability in water-flow represents one of the
most significant stressing elements both during an annual hydrological cycle and at a pluriannual level. Figure 3A,
shows monthly flows and the daily maxima of the Rambla del
Moro, one of the semiarid tributaires of the Segura river in
the Iberian South-east (fig. 2). As can be observed the irre- gularity of the flows during the study period, was extreme (from O m3/month to 227.146 m3/month). The maximum values corsespond to floods, a natural hydrological phenome- non that disturbs the whole system, basically because its occurrence is quite unpredictable. Fig. 3B, shows the number of floods that occursed in the Segura river between the years
826 and 1990. The scarcity of data during certain periods makes it difficult to stablish any regularity, even on a plurian- nual scale, where sequences of these events are repeated.
These hydrological models are the consequence of the peculiar climate of semiarid regions. Of al1 the climatic parameters usually used to define the climate of an asea, the
most important is the pattern of precipitations. Figure 4
shows temperature and precipitation diagrams (A) and the value of the annual precipitation in relation to its median (B), during a period of 41 years, in various termopluviome- tric gauges situated in the Segura river basin. Al1 of these
explains the peculiarities of the precipitation patterns in the
Spanish South-east: Its seasonal variability on the one hand, and its scarce predictability, on the other, when pluriannual cycles are analysed.
As a consequence of the above, it is easy to deduce that
the biological communities which inhabit these streams, find
themselves subjected to considerable natural disturbances that determine their structure and the functioning of the whole system.
ASPECTS ABOUT STRUCTURE AND FUNCTIONING OF SPANISH SEMIARID STREAMS
Hydrochemical patterns
Floods are one of the most important natural hydrologi- cal disturbance phenomena of Spanish semiarid streams. Changes in the physico-chemical parameters induced by a
flash flood (October 7th, 1986) in the Rambla del Moro, a spatially intermittent stream located in the East of the Segura river basin (fig. 2), were studied analyzing the water during ten days after the flood. Suspended solids, alkalinity and nutrients (nitrate-nitrogen and phosphate-
phosphorus) increased markedly after the flood while
conductivity declined quickly in relation to the discharge peaks (fig. 5) .
However, this situation is sporadic and only occurs when
flooding takes place. Problems related to the general hydro- logical system are encountered when trying to stablish the hydrochemical models of these streams. An intesive study on the variations in the hydrochemistry of Mula river (one of the main tributaries of the Segura river: fig. 2) made it possible to establish that, on a spatial level, the physico- chemical characteristics of the river could be explained by two components: the mineralization of the water and the
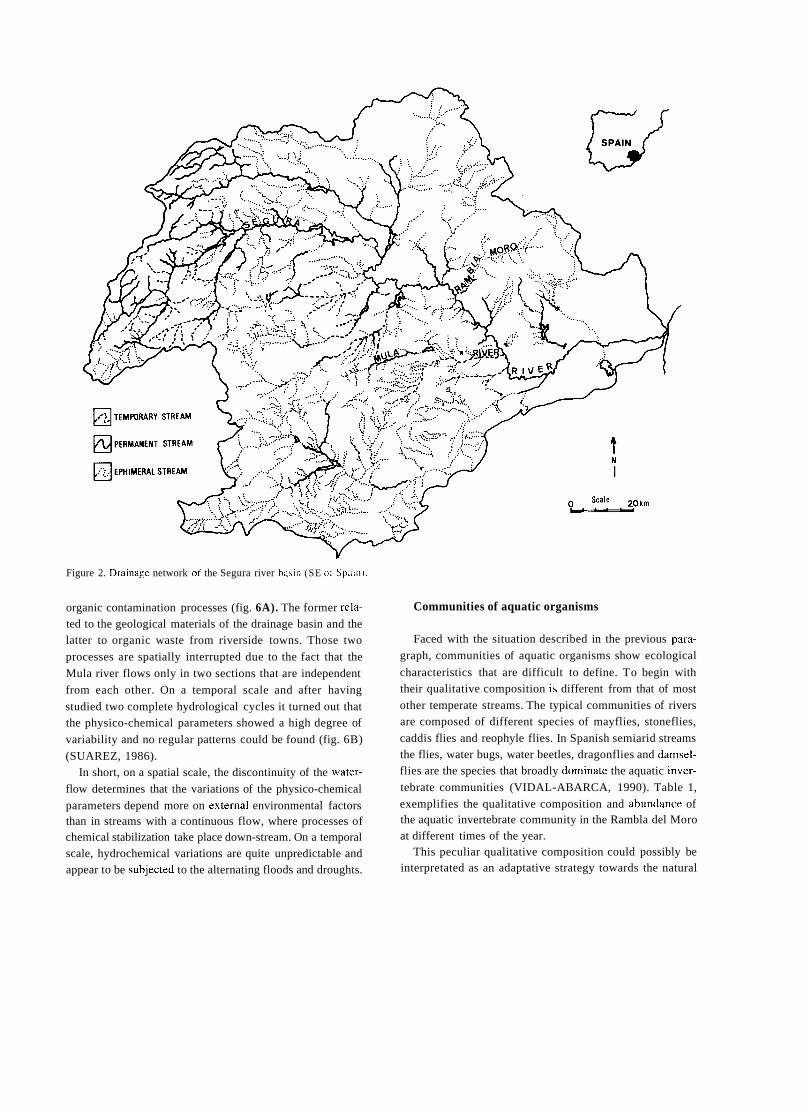
Figure 2. Drainage network of the Segura river basiti (SE ol Sp i i i i i ~ .
organic contamination processes (fig. 6A). The former rela- ted to the geological materials of the drainage basin and the latter to organic waste from riverside towns. Those two
processes are spatially interrupted due to the fact that the
Mula river flows only in two sections that are independent
from each other. On a temporal scale and after having
studied two complete hydrological cycles it turned out that the physico-chemical parameters showed a high degree of
variability and no regular patterns could be found (fig. 6B)
(SUAREZ, 1986). In short, on a spatial scale, the discontinuity of the water-
flow determines that the variations of the physico-chemical
parameters depend more on externa1 environmental factors than in streams with a continuous flow, where processes of chemical stabilization take place down-stream. On a temporal scale, hydrochemical variations are quite unpredictable and appear to be subjected to the alternating floods and droughts.
SPAIN i-i
Communities of aquatic organisms
Faced with the situation described in the previous para-
graph, communities of aquatic organisms show ecological
characteristics that are difficult to define. To begin with their qualitative composition is different from that of most
other temperate streams. The typical communities of rivers are composed of different species of mayflies, stoneflies,
caddis flies and reophyle flies. In Spanish semiarid streams the flies, water bugs, water beetles, dragonflies and damsel- flies are the species that broadly dominate the aquatic inver-
tebrate communities (VIDAL-ABARCA, 1990). Table 1,
exemplifies the qualitative composition and abundance of the aquatic invertebrate community in the Rambla del Moro at different times of the year.
This peculiar qualitative composition could possibly be interpretated as an adaptative strategy towards the natural
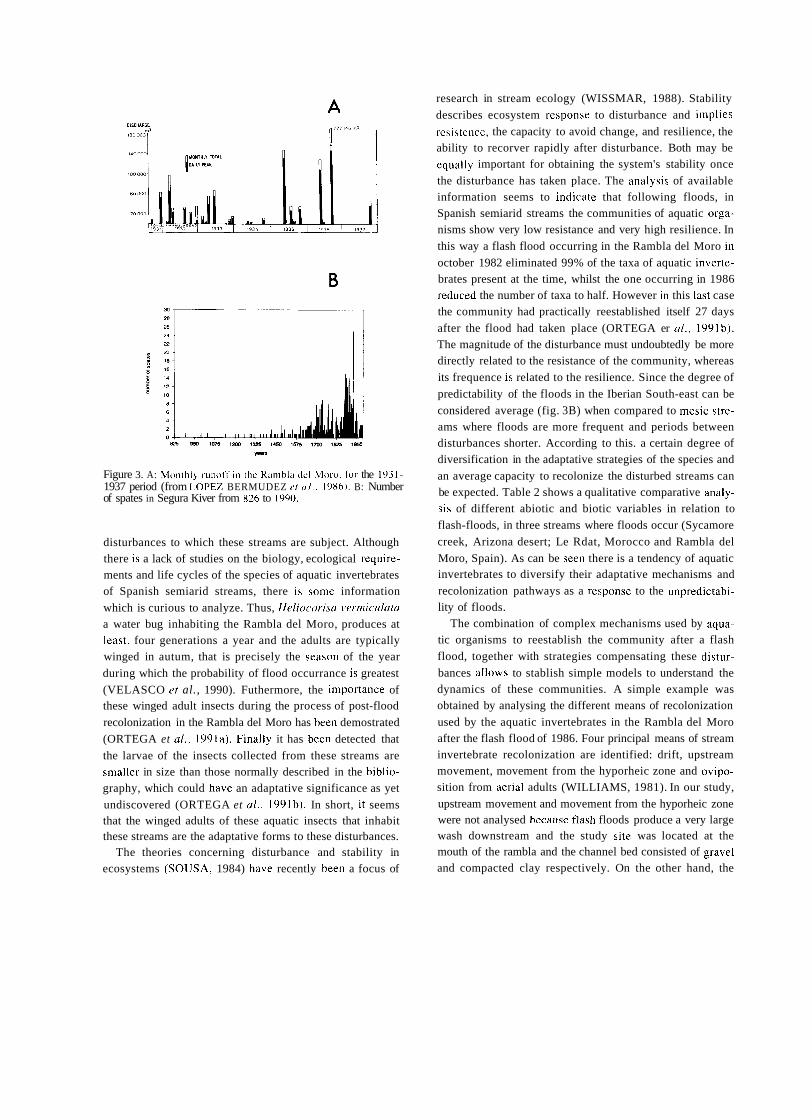
Figure 3. A: Moiithlq i-uiiol'l'iii tlic Kniiibla dcl Moro. for the 1931- 1937 period (from LOPEZ BERMUDEZ ct o / . . 1986). B: Number of spates in Segura Kiver from 826 to 1090.
disturbances to which these streams are subject. Although there is a lack of studies on the biology, ecological require- ments and life cycles of the species of aquatic invertebrates of Spanish semiarid streams, there is some information
which is curious to analyze. Thus, Hrlioc.or-isa i~er.rniculata a water bug inhabiting the Rambla del Moro, produces at least, four generations a year and the adults are typically winged in autum, that is precisely the season of the year
during which the probability of flood occurrance is greatest
(VELASCO e? al., 1990). Futhermore, the importance of these winged adult insects during the process of post-flood
recolonization in the Rambla del Moro has been demostrated
(ORTEGA et al.. 1991a). Finally it has been detected that
the larvae of the insects collected from these streams are smaller in size than those normally described in the biblio- graphy, which could have an adaptative significance as yet
undiscovered (ORTEGA et ul.. 1991b). In short, it seems that the winged adults of these aquatic insects that inhabit these streams are the adaptative forms to these disturbances.
The theories concerning disturbance and stability in ecosystems (SOUSA, 1984) have recently been a focus of
research in stream ecology (WISSMAR, 1988). Stability
describes ecosystem response to disturbance and implies
resistence, the capacity to avoid change, and resilience, the ability to recorver rapidly after disturbance. Both may be equally important for obtaining the system's stability once
the disturbance has taken place. The analysis of available information seems to indicate that following floods, in Spanish semiarid streams the communities of aquatic orga- nisms show very low resistance and very high resilience. In
this way a flash flood occurring in the Rambla del Moro in
october 1982 eliminated 99% of the taxa of aquatic inverte- brates present at the time, whilst the one occurring in 1986 reduced the number of taxa to half. However in this last case the community had practically reestablished itself 27 days
after the flood had taken place (ORTEGA er al., 1991b). The magnitude of the disturbance must undoubtedly be more directly related to the resistance of the community, whereas
its frequence is related to the resilience. Since the degree of
predictability of the floods in the Iberian South-east can be
considered average (fig. 3B) when compared to mesic stre- ams where floods are more frequent and periods between disturbances shorter. According to this. a certain degree of diversification in the adaptative strategies of the species and
an average capacity to recolonize the disturbed streams can be expected. Table 2 shows a qualitative comparative analy- sis of different abiotic and biotic variables in relation to
flash-floods, in three streams where floods occur (Sycamore
creek, Arizona desert; Le Rdat, Morocco and Rambla del
Moro, Spain). As can be seen there is a tendency of aquatic invertebrates to diversify their adaptative mechanisms and
recolonization pathways as a response to the unpredictabi- lity of floods.
The combination of complex mechanisms used by aqua- tic organisms to reestablish the community after a flash
flood, together with strategies compensating these distur-
bances allows to stablish simple models to understand the dynamics of these communities. A simple example was obtained by analysing the different means of recolonization
used by the aquatic invertebrates in the Rambla del Moro after the flash flood of 1986. Four principal means of stream invertebrate recolonization are identified: drift, upstream movement, movement from the hyporheic zone and ovipo- sition from aerial adults (WILLIAMS, 1981). In our study,
upstream movement and movement from the hyporheic zone were not analysed because flash floods produce a very large wash downstream and the study site was located at the mouth of the rambla and the channel bed consisted of grave1 and compacted clay respectively. On the other hand, the
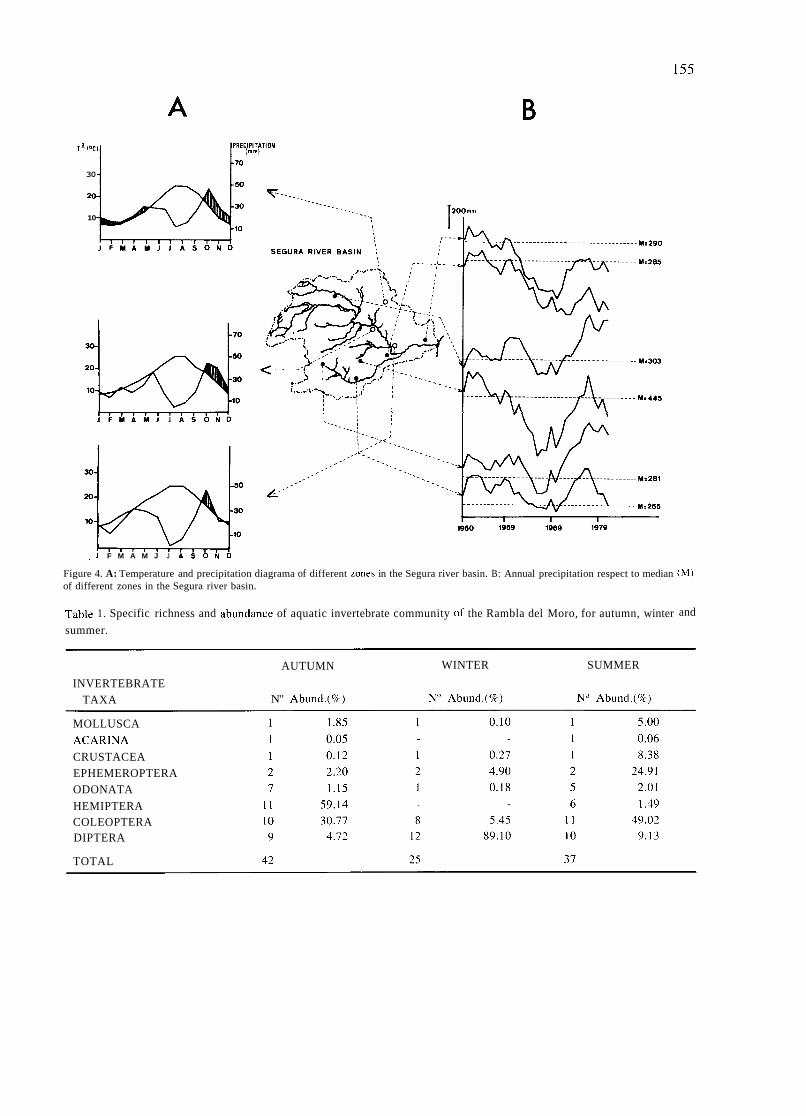
30
20 -. -. 10
. J F M A M J J A S O N D
Figure 4. A: Temperature and precipitation diagrama of different Lones in the Segura river basin. B: Annual precipitation respect to median of different zones in the Segura river basin.
Table 1. Specific richness and abundance of aquatic invertebrate community oí' the Rambla del Moro, for autumn, winter
summer.
(M)
and
INVERTEBRATE TAXA
AUTUMN
N" Abund.(%)
WINTER SUMMER
N"bund.(%) N V b u n d . ( % )
MOLLUSCA ACARINA CRUSTACEA EPHEMEROPTERA ODONATA HEMIPTERA COLEOPTERA DIPTERA
TOTAL
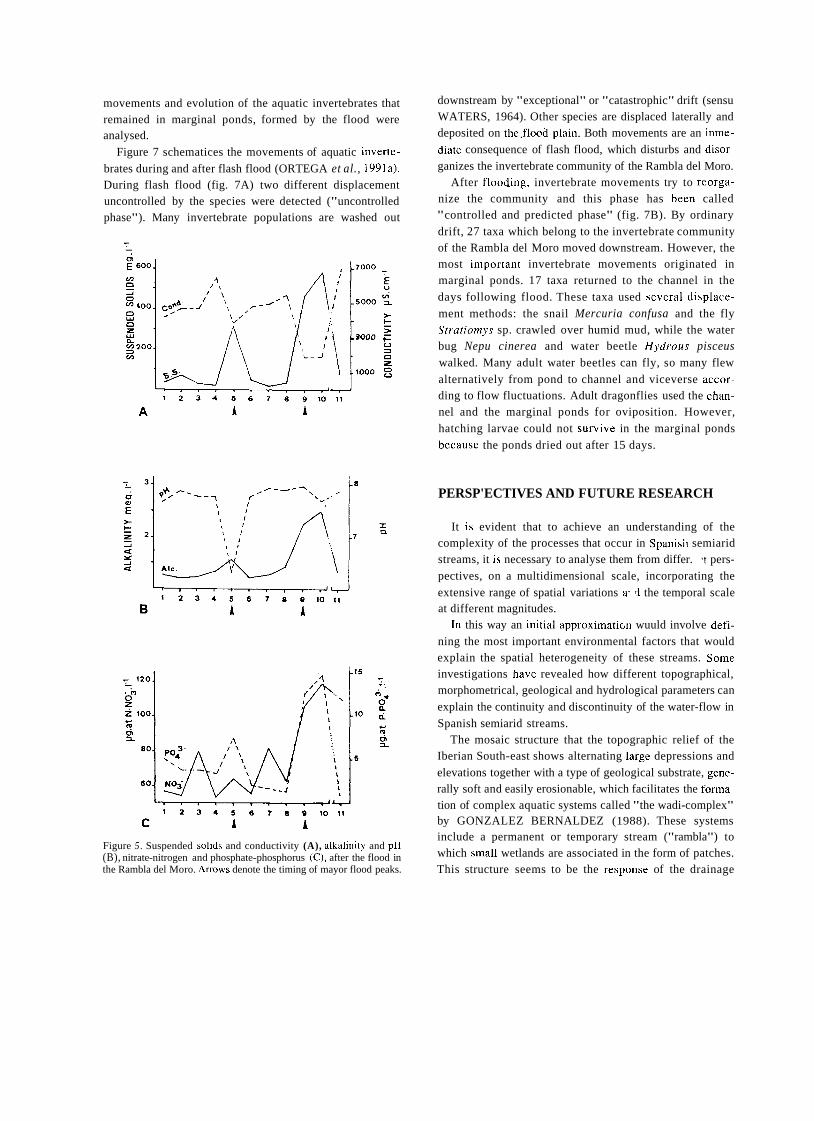
movements and evolution of the aquatic invertebrates that remained in marginal ponds, formed by the flood were analysed.
Figure 7 schematices the movements of aquatic inverte- brates during and after flash flood (ORTEGA et al., 1991a). During flash flood (fig. 7A) two different displacement uncontrolled by the species were detected ("uncontrolled phase"). Many invertebrate populations are washed out
Figure 5. Suspended solids and conductivity (A), alkalinity and pH (B), nitrate-nitrogen and phosphate-phosphorus (C), after the flood in the Rambla del Moro. Arrows denote the timing of mayor flood peaks.
downstream by "exceptional" or "catastrophic" drift (sensu WATERS, 1964). Other species are displaced laterally and deposited on the,flood-plain. Both movements are an inmc- diate consequence of flash flood, which disturbs and disor- ganizes the invertebrate community of the Rambla del Moro.
After flooding, invertebrate movements try to reorga- nize the community and this phase has been called "controlled and predicted phase" (fig. 7B). By ordinary drift, 27 taxa which belong to the invertebrate community of the Rambla del Moro moved downstream. However, the most important invertebrate movements originated in marginal ponds. 17 taxa returned to the channel in the days following flood. These taxa used severa1 displace- ment methods: the snail Mercuria confusa and the fly Stratiomys sp. crawled over humid mud, while the water bug Nepu cinerea and water beetle Hydrous pisceus walked. Many adult water beetles can fly, so many flew alternatively from pond to channel and viceverse accor- ding to flow fluctuations. Adult dragonflies used the chan- nel and the marginal ponds for oviposition. However, hatching larvae could not survive in the marginal ponds because the ponds dried out after 15 days.
PERSP'ECTIVES AND FUTURE RESEARCH
It is evident that to achieve an understanding of the complexity of the processes that occur in Spanisi~ semiarid streams, it is necessary to analyse them from differ. -t pers- pectives, on a multidimensional scale, incorporating the extensive range of spatial variations a. d the temporal scale at different magnitudes.
In this way an initial approximatio~i wuuld involve defi- ning the most important environmental factors that would explain the spatial heterogeneity of these streams. Some investigations have revealed how different topographical, morphometrical, geological and hydrological parameters can explain the continuity and discontinuity of the water-flow in Spanish semiarid streams.
The mosaic structure that the topographic relief of the Iberian South-east shows alternating large depressions and elevations together with a type of geological substrate, gene- rally soft and easily erosionable, which facilitates the forma- tion of complex aquatic systems called "the wadi-complex" by GONZALEZ BERNALDEZ (1988). These systems include a permanent or temporary stream ("rambla") to which small wetlands are associated in the form of patches. This structure seems to be the response of the drainage
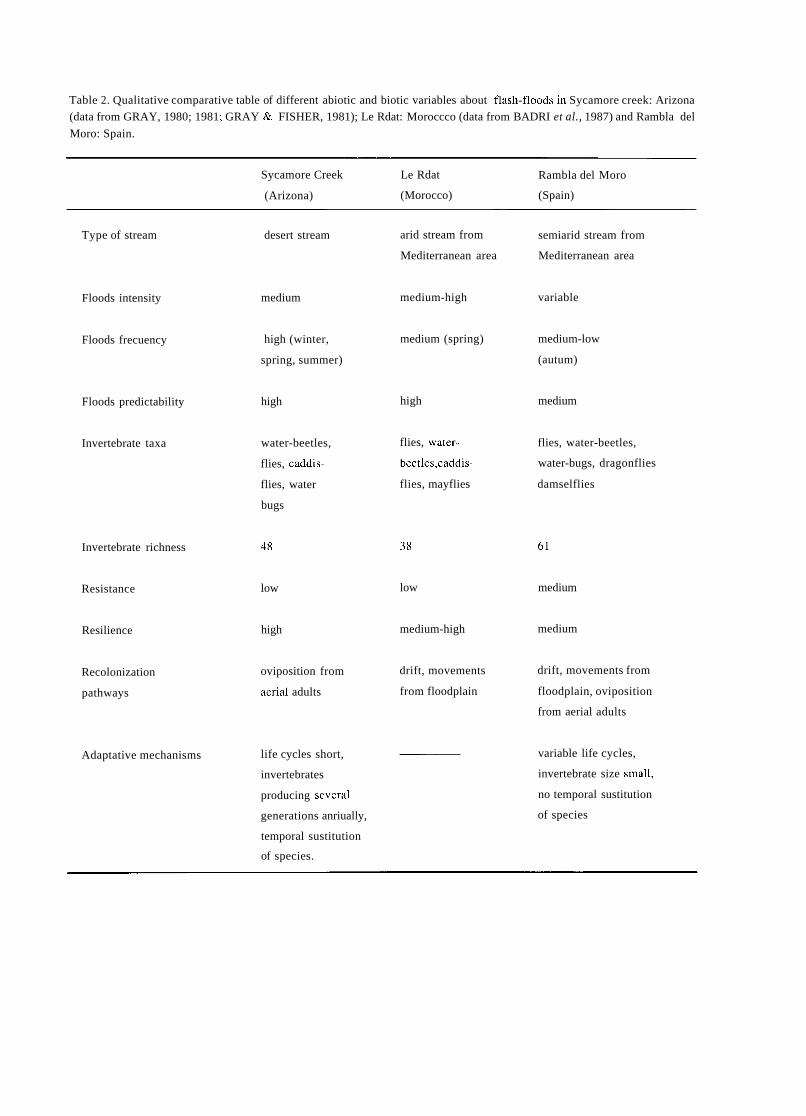
Table 2. Qualitative comparative table of different abiotic and biotic variables about flash-floods in Sycamore creek: Arizona
(data from GRAY, 1980; 198 1 ; GRAY & FISHER, 198 1); Le Rdat: Moroccco (data from BADRI et al., 1987) and Rambla del
Moro: Spain.
Sycamore Creek Le Rdat
(Arizona) (Morocco)
Rambla del Moro
(Spain)
desert stream arid stream from semiarid stream from
Mediterranean area Mediterranean area
Type of stream
Floods intensity medium medium-high variable
high (winter,
spring, summer)
medium (spring) medium-low
(autum)
Floods frecuency
high medium high Floods predictability
Invertebrate taxa water-beetles,
flies, caddis-
flies, water
bugs
flies, water-
beetles,caddis-
flies, mayflies
flies, water-beetles,
water-bugs, dragonflies
damselflies
Invertebrate richness
Resistance low low medium
Resilience high medium-high medium
oviposition from
aerial adults
drift, movements
from floodplain
drift, movements from
floodplain, oviposition
from aerial adults
Recolonization
pathways
variable life cycles,
invertebrate size small,
no temporal sustitution
of species
Adaptative mechanisms life cycles short,
invertebrates
producing several
generations anriually,
temporal sustitution
of species.
Figure 6. A: Relative position of physico-chemical parameters measured in 5 1 sampling stations located in the Mula river basin, during summer, in the planes defined by axes 1-11 and 1-111 of the Factor Analysis, and environmental interpretation of these axes. B: Projection of sarnples (months) in the plane defined by the first three axes of Factor Analysis, of two sampling stations located in the Mula river. Alk. = Alkalinity; Cond. = Conductivity; Sat. = O/o Saturation of dissolved oxygen; S.S. = Suspended solids.
system to the floods. The role of the floods in the genesis or
disappearance of these wetlands is another objetive of current investigation.
These "wadi complex" must have an essential role in the dynamics of the nutrients of the associated streams. The
studies that we are currently initiating will possibly explain the spatial and temporal hydrochemical models of Spanish semiarid streams.
On the other hand, the studies on the adaptative behaviour
of aquatic invertebrate species in these streams continue to
be the main research objetive. Undoubtedly, the accumula-
tion of information (intensity, frequency, magnitude, etc) of the disturbances of Spanish semiarid streams and the analy-
sis of the life cycles and ecological requirements of the invertebrates species, will enable to stablish reliable models of their dynamics.

t Flood-Plain
b
MOVEMENTS TO REORCANIZE THE BENTHIC COMMUNITV
¡CONTROLLEO ANO PREDICTED PHASEI
OVOPOS~T~ON FROM AERIAL AOULTS
1 OROINARY DRlFT I
1 Unhabitual Habtlual , dnfling drilling - \ \ i
* Marginal Pond
Figure 7. Summary of invertebrate taxa movements ocurring during (A) and after (B) flood, in the Rambla del Moro (modified from ORTEGA et al., 1991a).
Additionally, another objetive of the work has an applied character and pretends to design a complex methodology to
made possible elaborate maps with associated risk of floods. This will be most usefull for the studies on territorial plan- ning.
ACKNOWLEDGEMENTS
To Dr. F. Torrella for reviewing the english paper.
REFERENCES
ARENILLAS, M. & C.SAENZ. 1987. Guia Fí.siic de
España. 3.-Los ríos. Alianza Editorial. Madrid.
AVILA, A. 1984. Composición química de los arroyos del Montseny (Barcelona). Limnética 1( 1): 128- 135.
BADRI, A; J.GUIDICELL1 & G.PREVOT. 1987. Effects d'une crue sur la communaute d'invertébrés benthiques d'une riviere méditerranéenne, Le Rdat (Maroc). Acta Oeco-
logica. Oecol. Gener., 8(4): 48 1-500. ESCARRE, A.; C.GRACIA; F.RODA & J.TERRADAS.
1984. Ecología del bosque esclerófilo mediterráneo. Inves-
tigación y Ciencia, 95: 69-78. GARCIA DE PEDRAZA, L. 1989. Climas de transición
al árido. Climas áridos, dominios áridos y semiáridos en España. In: Seminario sobre Zonas Aridas en España. 45- 63. Real Academia de Ciencias Exactas, Físicas y Natura- les. Madrid.
GONZALEZ BERNALDEZ, F. 1988. Typology of wetlands and evaluation of the resources they represent. In: International Symposium on Hydrology of wetlands in Semiarid and arid regions. 7-36. Sevilla.
GRAY, L.J. 1980. Recolonization pathways and commu- nity development of desert stream macroinvertebrate. Diser- tation: Arizona State University.
GRAY, L.J. 1981. Species composition and life histories
of aquatic insects in a Lowland Sonoran Desert stream. Amer. Midland Natur., 106: 229-242.
GRAY, L.J. & S.G. FISHER. 1981. Postflood recoloni- zation pathways of macroinvertebrates in a Lowland Sono- ran Desert stream. Amer. Midland Natur. 106: 249-257.
LOPEZ BERMUDEZ, F.; F.CALVO & A.MORALES. 1986. Geografía de la Región de Murcia. Ketres Ed. Barce- lona.
MONTES, C. & P.MARTINO. 1987. Las lagunas salinas españolas. In: Bases cientí$icas para la protección de los
humedales españoles. 95-145. Real Academia de Ciencias Exactas, Físicas y Naturales. Madrid.
ORTEGA, M. 1988. La rambla del Moro (Cuenca del
Río Segura). Ambiente ,físico, biológico y alteraciones producidas por una riada. Tesis de Licenciatura. Univ. Murcia.
ORTEGA, M.; M.R.VIDAL-ABARCA; M.L.SUAREZ; J.L.GONZALEZ-BESERAN & L.RAMIREZ-DIAZ. 1988. Características fisico-químicas de las aguas superficiales de la Rambla del Moro despues de una riada (Cuenca del Río Segura, SE de España). Limnética, 4: 19-26.
ORTEGA, M.; M.L.SUAREZ; M.R.VIDAL-ABARCA; R.GOMEZ & L.RAMIREZ-DIAZ. 1991a. Aspects of post- flood recolonization of macroinvertebrates in a "Rambla" of south-east Spain ("Rambla del Moro": Segura River Basin). Verh. Internar. Verein. Limnol., 24: 1994-200 1.

ORTEGA, M.; M.L.SUAREZ; M.R.VIDAL-ABARCA & L.RAMIREZ-DIAZ. 1991b. Aspectos dinámicos de la composición y estructura de la comunidad de invertebrados acuáticos de la Rambla del Moro despues de una riada (Cuenca del Río Segura: SE de España). Linznética, 7: 1 1-24.
PRAT, N; G.GONZALEZ; I.MUNOZ & X.MILLET. 1986. Community structure and colonization process in a mediterranean drainage basin. Proc. 3rd Europen Cong.
Entomol.: 12 1 -124. PUIG, M.A.; J.ARMENGOL; G.GONZALEZ; J.PEÑUE-
LAS; S.SABATER & F.SABATER. 1987. Chemical and biological changes in the Ter River induced by series of reservoirs. In: CRAIG, J. (ed.) Regulated Str-eam: Advances
in Ecology. 363-373. Plenum Press. PUIG, M.A.; M.FERRERAS & A.GARCIA-ROJAS.
1990. Morphological variability of Tyrrhenoleuctra minuta (Klapalek, 1903) in south Spain. In: CAMPBELL, I.C.(ed.). Mayflies and Stoneflies. Kluwer Acad. Publ.
SABATER, S. 1981. ~istrtbución espacio-temporal de las poblaciones de algas del arroyo de L'Avencó (Barcelona). Actas 1 T o n g . Limnología: 159-166.
SOLER, G. 1991. Pautus de migración de las comunida- des hentónicas del Río Matarrana (Cuenca del Ebro) a lo largo de un ciclo anual y en función de la velocidad de la corriente y la heterogeneidad del sustrato colonizahle. Tésis de Licenciatura. Univ. Murcia.
SOUSA, W.P. 1984. The role of disturbance in natural communities. Aifiz. Rev. Ecol. Syst., 15: 353-391.
SUAREZ, M.L. 1986. Estructura y dinámica de la composición físico-química de las aguas superficiales de
una cuenca de características semiáridas del sureste espa- ñol (Río Mula: Cuenca del Segura). Tesis Doctoral. Univ. Murcia.
VELASCO, J.; A.MILLAN & N.NIESER. 1990. Obser- vaciones sobre la colonización y el ciclo de vida de Helio-
corisa vermiculata (Puton, 1874) (Heteroptera, Corixidae) en pequeños estanques del SE español. Limnética, 6: 101-108.
VIDAL-ABARCA, M.R. 1985. Las aguas superficiales
de la Cuenca del Río Segura (SE de España). Caracteriza-
ción físico-quimica en relación al medio físico y humano. Tesis Doctoral. Univ. Murcia.
VIDAL-ABARCA, M.R. 1990. Los ríos de las cuencas áridas y semiáridas: Una perspectiva ecológica y de sínte- sis. Scientia gerundensis, 16: 219-228.
WATERS, T.F. 1964. Recolonization of denuded stream bottom aseas by drift. Trans. Am. Fish. Soc., 93: 31 1-325.
WILLIAMS, D.D. 1981. Migrations and distributions of stream benthos. In: LOCK, M.A. & D.D. WILLIAMS (eds.). Perspectives in running ecology. 155-207. Plenum Press. New York.
WISSMAR, R.C. 1988. The role of disturbance in stream ecology. J. N. Am. Benthol. Soc.,7(4): 433-455.

161
ECOLOGY OF REGULATED STREAMS IN SPAIN: AN OVERVIEW
D. Garcia de Ja16n1, M. Gonzilez del Tinago1 and C. C a s a d o 2
1. E.T.S. Ingenieros de Montes,Universidad PolitCcnica de Madrid. Spain. 2. Lab. Limnologia. Dpto. Ecologia, Universidad Aut6noma de Madrid. 28049 Madrid. Spain.
Keywords: Stream regulation, Limnology, Spain.
ABSTRACT
A brief review of the main studies on flow regulation effects in Spain is presented, considering algae, fish and macroinver- tebrate communities.
The eel (Anguilla anguilla), salmon (Salmo salar-) and seatrout (Salmo frutta tr-utta) are perhaps the most affected species, having reduced their distribution area considerably. The response of macroinvertebrate communities to flow regulation chan- ges from one river to another, due to the multiple use of reservoirs. A decrease of species richness and diversity have been frequently reported, being flow fluctuation and lower water temperature the main factors responsible for that.
INTRODUCTION
Nowadays all Spanish large rivers have their flows regu- lated by reservoirs, and most of the median size streams are also regulated. This intensive flow regulation tends to ameliorate an irregular distribution of the water resources, both in terms of space and time.
The space irregularity is explained because of climatic conditions, following a gradient from the more rainy areas in the Northwest (Atlantic influence) to the drier zones in the Southeast (Mediterranean influence), having a mean annual effective rainfall of 215 mm. for the country.
The time irregularity is a consequence of the dominant mediterranean conditions, with intensive to very intensive precipitations during fall to early spring and a marked summer drought, having a large irregularity between years also.
Flow regulation has been one of the priorities in Spain during the 50's, and nowadays there are more than a thousand big dams constructed, taking advantage of the Iberian high relief. This amount of big reservoirs is completed with some water transfers, to reduce water differences between regions.
RESERVOIRS
The thousand of large reservoirs represents an storage capacity of near 45,000 million cubic'metres (DIRECCION
GENERAL OBRAS HIDRAULICAS, 1986), having most of them a multiple use. Hydropower generation is the most commun, using the 84 % of this capacity, followed by irri- gation that employes the 59 % and the industrial and urban supply that represents a 26 % of it.
Hydroelectric energy is mainly generated for absorbing the peaks of energy demand, which is basically attended in Spain by conventional thermic and nuclear plants, needing to import some energy from France as well. Sudden flow fluctuations take place below reservoirs used for energy production, with no consideration to the aquatic environ- ment. Since 198 1 there is an increase of the construction rate of small hydropower systems, subsidized by the Spanish governement and the EEC, under the erroneous idea of no environmental impact ("clean energy"). This last hydroe- lectic regulation is specially impacting recondite streams that otherwise would be in pristine conditions.
During the last years, an increase of irrigation land is taking place in Spain, needing to dam more water for this use despite the public opposition in some cases like "Riaiio reservoir". Horticulture and fruit crops are more demman- ded in the European Market and most of the traditional agri- culture is being abandoned and replaced by irrigation crops.
The installation and maintenance of reservoirs, even if designed primarly for irrigation pourposes, substantially reduce flood peaks in dryland rivers (ALDRIDGE & EYCHANER, 1984). High dams also cause a shift to the
Limnetica, 8: 161-166 (1992) O Asociaci6n Espaiola de Limnologia, Madrid. Spain
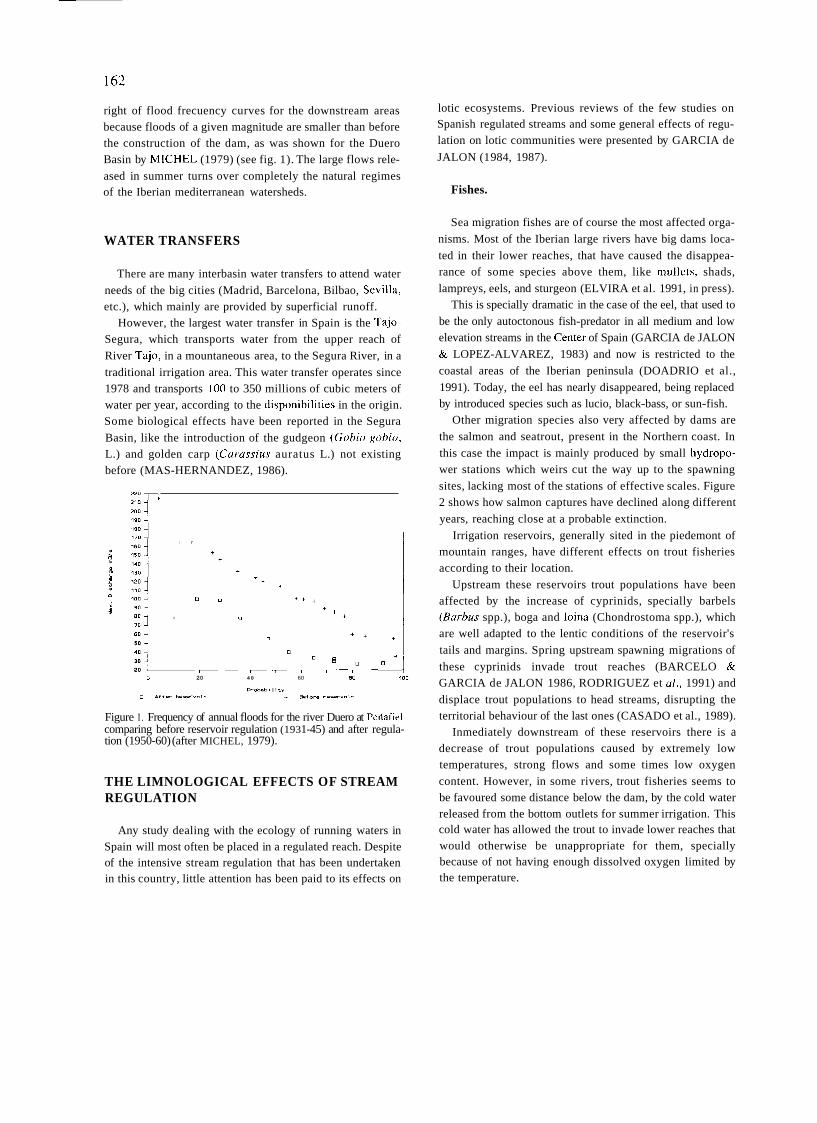
right of flood frecuency curves for the downstream areas because floods of a given magnitude are smaller than before the construction of the dam, as was shown for the Duero Basin by MICHEL (1979) (see fig. 1). The large flows rele-
ased in summer turns over completely the natural regimes of the Iberian mediterranean watersheds.
WATER TRANSFERS
There are many interbasin water transfers to attend water
needs of the big cities (Madrid, Barcelona, Bilbao, Sevilla, etc.), which mainly are provided by superficial runoff.
However, the largest water transfer in Spain is the Tajo- Segura, which transports water from the upper reach of
River Tajo, in a mountaneous area, to the Segura River, in a
traditional irrigation area. This water transfer operates since 1978 and transports 100 to 350 millions of cubic meters of water per year, according to the disponibilities in the origin. Some biological effects have been reported in the Segura
Basin, like the introduction of the gudgeon (Gobio gobio, L.) and golden carp (Curassius auratus L.) not existing before (MAS-HERNANDEZ, 1986).
: '!!l 70 , 1 O , D , o + , ; ; + + ; o + , 60
50
110
30 0 0 20
0 20 4 0 60 80 100
R ~ b * b 8 l i t y
0 &F*- D P ~ s r r o i r + r n f c r o re-voir
Figure I . Frequency of annual floods for the river Duero at Peftafiel comparing before reservoir regulation (193 1-45) and after regula- tion (1950-60) (after MICHEL, 1979).
THE LIMNOLOGICAL EFFECTS OF STREAM REGULATION
Any study dealing with the ecology of running waters in Spain will most often be placed in a regulated reach. Despite of the intensive stream regulation that has been undertaken in this country, little attention has been paid to its effects on
lotic ecosystems. Previous reviews of the few studies on Spanish regulated streams and some general effects of regu- lation on lotic communities were presented by GARCIA de
JALON (1 984, 1987).
Fishes.
Sea migration fishes are of course the most affected orga-
nisms. Most of the Iberian large rivers have big dams loca-
ted in their lower reaches, that have caused the disappea- rance of some species above them, like mullets, shads,
lampreys, eels, and sturgeon (ELVIRA et al. 1991, in press). This is specially dramatic in the case of the eel, that used to
be the only autoctonous fish-predator in all medium and low elevation streams in the Center of Spain (GARCIA de JALON
& LOPEZ-ALVAREZ, 1983) and now is restricted to the
coastal areas of the Iberian peninsula (DOADRIO et al.,
1991). Today, the eel has nearly disappeared, being replaced by introduced species such as lucio, black-bass, or sun-fish.
Other migration species also very affected by dams are
the salmon and seatrout, present in the Northern coast. In
this case the impact is mainly produced by small hydropo- wer stations which weirs cut the way up to the spawning
sites, lacking most of the stations of effective scales. Figure 2 shows how salmon captures have declined along different
years, reaching close at a probable extinction.
Irrigation reservoirs, generally sited in the piedemont of mountain ranges, have different effects on trout fisheries according to their location.
Upstream these reservoirs trout populations have been affected by the increase of cyprinids, specially barbels (Barbus spp.), boga and loina (Chondrostoma spp.), which are well adapted to the lentic conditions of the reservoir's
tails and margins. Spring upstream spawning migrations of
these cyprinids invade trout reaches (BARCELO & GARCIA de JALON 1986, RODRIGUEZ et ul., 1991) and
displace trout populations to head streams, disrupting the territorial behaviour of the last ones (CASADO et al., 1989).
Inmediately downstream of these reservoirs there is a decrease of trout populations caused by extremely low temperatures, strong flows and some times low oxygen
content. However, in some rivers, trout fisheries seems to be favoured some distance below the dam, by the cold water released from the bottom outlets for summer irrigation. This cold water has allowed the trout to invade lower reaches that would otherwise be unappropriate for them, specially because of not having enough dissolved oxygen limited by the temperature.
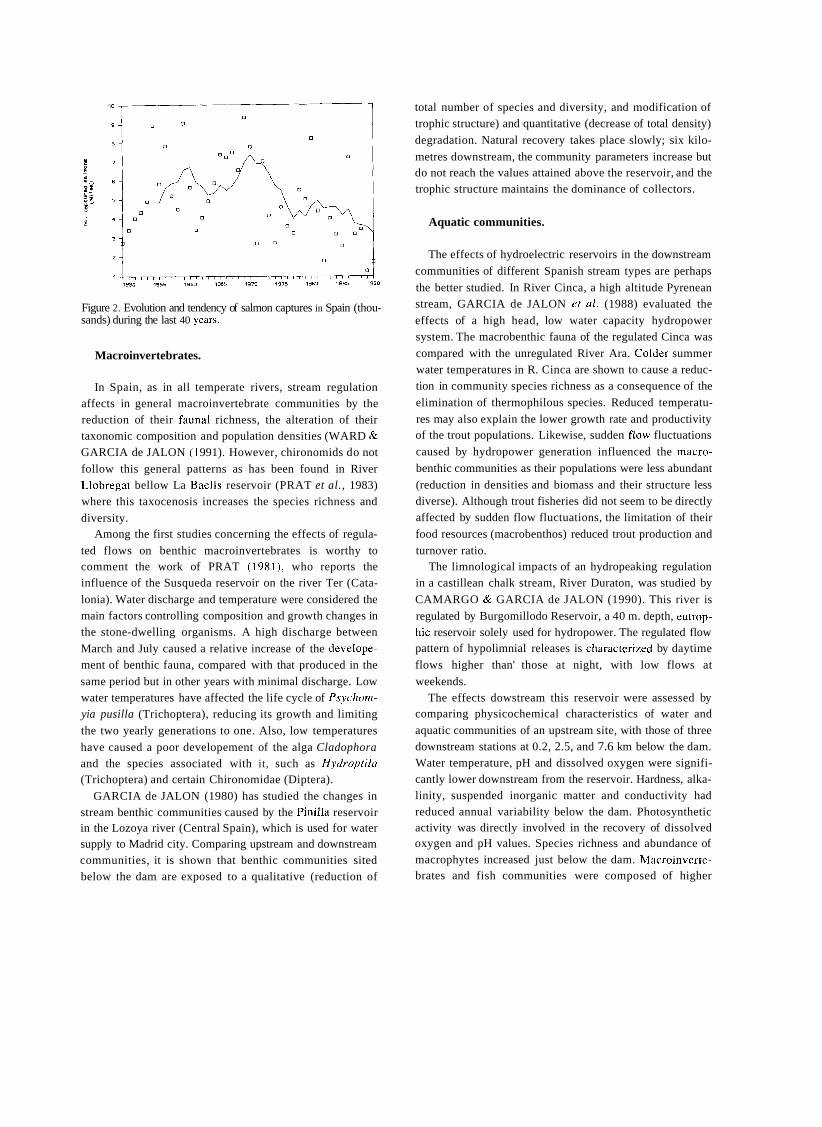
Figure 2. Evolution and tendency of salmon captures in Spain (thou- sands) during the last 40 years.
Macroinvertebrates.
In Spain, as in all temperate rivers, stream regulation affects in general macroinvertebrate communities by the reduction of their fauna1 richness, the alteration of their taxonomic composition and population densities (WARD &
GARCIA de JALON ( 1 991). However, chironomids do not
follow this general patterns as has been found in River Llobregat bellow La Baells reservoir (PRAT et al., 1983) where this taxocenosis increases the species richness and
diversity. Among the first studies concerning the effects of regula-
ted flows on benthic macroinvertebrates is worthy to comment the work of PRAT (1981), who reports the influence of the Susqueda reservoir on the river Ter (Cata-
lonia). Water discharge and temperature were considered the main factors controlling composition and growth changes in the stone-dwelling organisms. A high discharge between
March and July caused a relative increase of the develope- ment of benthic fauna, compared with that produced in the
same period but in other years with minimal discharge. Low water temperatures have affected the life cycle of Psychom- yia pusilla (Trichoptera), reducing its growth and limiting
the two yearly generations to one. Also, low temperatures
have caused a poor developement of the alga Cladophora and the species associated with it, such as Hydroptila (Trichoptera) and certain Chironomidae (Diptera).
GARCIA de JALON (1980) has studied the changes in stream benthic communities caused by the Pinilla reservoir in the Lozoya river (Central Spain), which is used for water supply to Madrid city. Comparing upstream and downstream communities, it is shown that benthic communities sited below the dam are exposed to a qualitative (reduction of
total number of species and diversity, and modification of trophic structure) and quantitative (decrease of total density)
degradation. Natural recovery takes place slowly; six kilo-
metres downstream, the community parameters increase but do not reach the values attained above the reservoir, and the trophic structure maintains the dominance of collectors.
Aquatic communities.
The effects of hydroelectric reservoirs in the downstream
communities of different Spanish stream types are perhaps
the better studied. In River Cinca, a high altitude Pyrenean stream, GARCIA de JALON rr al. (1988) evaluated the
effects of a high head, low water capacity hydropower system. The macrobenthic fauna of the regulated Cinca was
compared with the unregulated River Ara. Colder summer water temperatures in R. Cinca are shown to cause a reduc- tion in community species richness as a consequence of the
elimination of thermophilous species. Reduced temperatu-
res may also explain the lower growth rate and productivity of the trout populations. Likewise, sudden flow fluctuations
caused by hydropower generation influenced the macro-
benthic communities as their populations were less abundant
(reduction in densities and biomass and their structure less diverse). Although trout fisheries did not seem to be directly affected by sudden flow fluctuations, the limitation of their
food resources (macrobenthos) reduced trout production and
turnover ratio. The limnological impacts of an hydropeaking regulation
in a castillean chalk stream, River Duraton, was studied by
CAMARGO & GARCIA de JALON (1990). This river is
regulated by Burgomillodo Reservoir, a 40 m. depth, eutrop- hic reservoir solely used for hydropower. The regulated flow pattern of hypolimnial releases is characterized by daytime
flows higher than' those at night, with low flows at
weekends. The effects dowstream this reservoir were assessed by
comparing physicochemical characteristics of water and
aquatic communities of an upstream site, with those of three downstream stations at 0.2, 2.5, and 7.6 km below the dam. Water temperature, pH and dissolved oxygen were signifi- cantly lower downstream from the reservoir. Hardness, alka- linity, suspended inorganic matter and conductivity had reduced annual variability below the dam. Photosynthetic activity was directly involved in the recovery of dissolved oxygen and pH values. Species richness and abundance of macrophytes increased just below the dam. Macroinverte- brates and fish communities were composed of higher
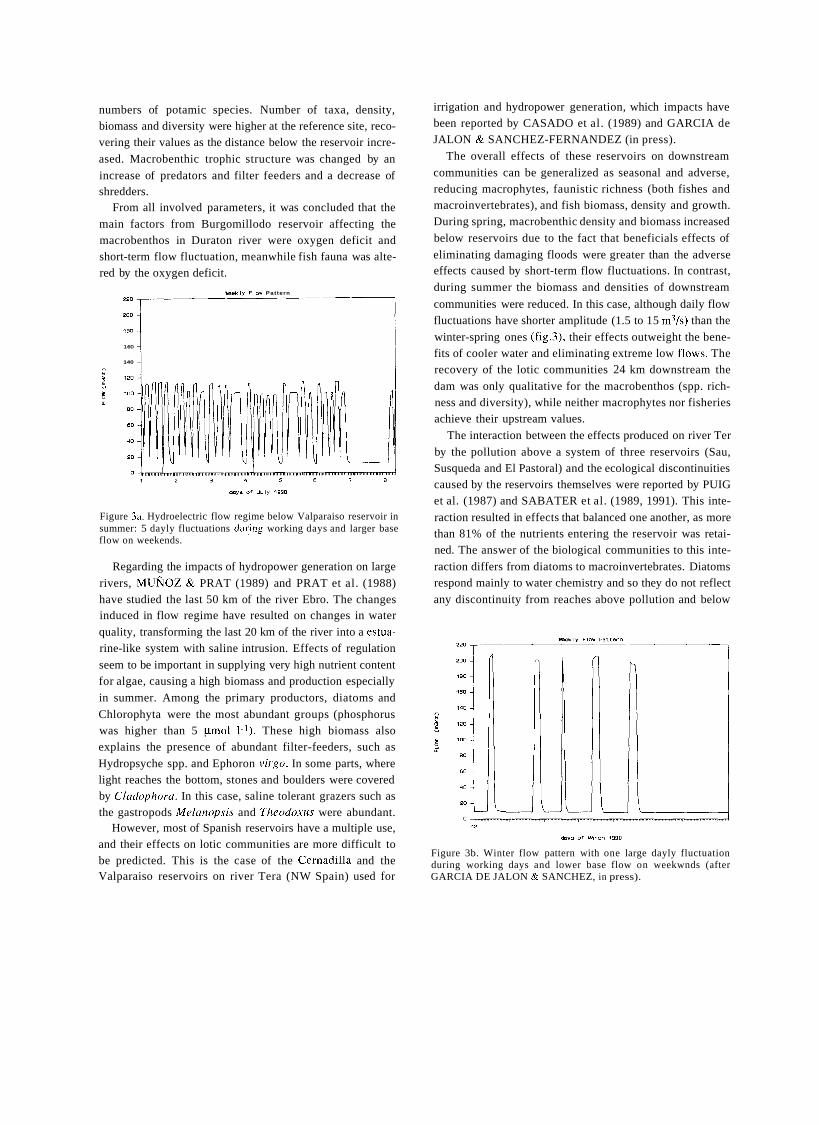
numbers of potamic species. Number of taxa, density, biomass and diversity were higher at the reference site, reco- vering their values as the distance below the reservoir incre-
ased. Macrobenthic trophic structure was changed by an
increase of predators and filter feeders and a decrease of shredders.
From all involved parameters, it was concluded that the
main factors from Burgomillodo reservoir affecting the macrobenthos in Duraton river were oxygen deficit and short-term flow fluctuation, meanwhile fish fauna was alte- red by the oxygen deficit.
week ly F o * Pattern
160
140
Figure 3a. Hydroelectric flow regime below Valparaiso reservoir in summer: 5 dayly fluctuations dur,ing working days and larger base flow on weekends.
Regarding the impacts of hydropower generation on large
rivers, MUNOZ & PRAT (1989) and PRAT et al. (1988) have studied the last 50 km of the river Ebro. The changes induced in flow regime have resulted on changes in water
quality, transforming the last 20 km of the river into a estua- rine-like system with saline intrusion. Effects of regulation
seem to be important in supplying very high nutrient content for algae, causing a high biomass and production especially
in summer. Among the primary productors, diatoms and
Chlorophyta were the most abundant groups (phosphorus was higher than 5 pm01 I-'). These high biomass also explains the presence of abundant filter-feeders, such as
Hydropsyche spp. and Ephoron virgo. In some parts, where light reaches the bottom, stones and boulders were covered by Cladophol-a. In this case, saline tolerant grazers such as the gastropods Melanopsis and Theodoxus were abundant.
However, most of Spanish reservoirs have a multiple use, and their effects on lotic communities are more difficult to be predicted. This is the case of the Cernadilla and the Valparaiso reservoirs on river Tera (NW Spain) used for
irrigation and hydropower generation, which impacts have been reported by CASADO et al. (1989) and GARCIA de JALON & SANCHEZ-FERNANDEZ (in press).
The overall effects of these reservoirs on downstream
communities can be generalized as seasonal and adverse, reducing macrophytes, faunistic richness (both fishes and macroinvertebrates), and fish biomass, density and growth. During spring, macrobenthic density and biomass increased
below reservoirs due to the fact that beneficials effects of
eliminating damaging floods were greater than the adverse effects caused by short-term flow fluctuations. In contrast,
during summer the biomass and densities of downstream
communities were reduced. In this case, although daily flow fluctuations have shorter amplitude (1.5 to 15 m3/s) than the
winter-spring ones (fig.3), their effects outweight the bene- fits of cooler water and eliminating extreme low flows. The recovery of the lotic communities 24 km downstream the
dam was only qualitative for the macrobenthos (spp. rich- ness and diversity), while neither macrophytes nor fisheries achieve their upstream values.
The interaction between the effects produced on river Ter
by the pollution above a system of three reservoirs (Sau, Susqueda and El Pastoral) and the ecological discontinuities caused by the reservoirs themselves were reported by PUIG et al. (1987) and SABATER et al. (1989, 1991). This inte-
raction resulted in effects that balanced one another, as more than 81% of the nutrients entering the reservoir was retai- ned. The answer of the biological communities to this inte-
raction differs from diatoms to macroinvertebrates. Diatoms respond mainly to water chemistry and so they do not reflect
any discontinuity from reaches above pollution and below
Figure 3b. Winter flow pattern with one large dayly fluctuation during working days and lower base flow on weekwnds (after GARCIA DE JALON & SANCHEZ, in press).

the reservoirs. While in the macroinvertebrate community. remarkable shifts in composition occur as a consequence of the disruption in the physical conditions of the river, although the community diversity did not changed.
CONCLUSIONS
Flow regulation in Spain has been one of the priorities to social developme~it during the last decades, attending to solve intensive irregularities of water resources distribution, both in terms of time and space. Today, there are more than a thousand big dams constructed, and more than 92 large reservoirs under constructiorl or projected to increase the actual storage water capacity of 45,000 Hm3 to 60,000 Hm3.
As a consequence, the ecology of rivers and streams has been deeply altered, although little attention is still been paid to study the regulation effects. Even less attention have been given to compensation or minimum flows released below reservoirs, that never have been evaluated from an ecological basis (GARCIA de JALON, 1990).
Migration fishes have been the most affected. Anguilla arzguilla has disappeared from central reaches where it used to be very abundant, and salmon (Salmo salar) and seatrout (Salmo trutta tr-utta) populations have declined to nearly extinction in many Cantabric streams.
Macroinvertebrate communities have been also affected by dams in their composition and structure, decreasing in most of the cases their species richness. Sudden flow fluc- tuations from hydropower reservoirs, as well as lower water temperature, have been reported as the main factors impac- ting these communities, although the multiple use of the majority of Spanish reservoirs makes rather complex the assessment of their effects on aquatic fauna.
REFERENCES
ALDRIGE, B.N. y J.H. EYCHANER. 1984. Floods of October 1977 in southern Arizona and March 1978 in New Mexico, and adjacent basins in Arizona. U.S. Geol. Surv. Wat. Supply Pap. 1850-C.
BARCELO, E. & D. GARCIA DE JALON. 1986. Edad y crecimiento de la boga de rio (Chondrostoma polylepis Steind., 1865) en la Cuenca del Duero. Limnetica 2: 235-240.
CAMARGO, J.A. y D. GARCIA DE JALON. 1991. The downstream impacts of the Burgomillodo reservoir, Spain. Regul. Riv1er.s: Res. & Mgt. 5, 305-17.
CASADO, C., D. GARCIA DE JALON, C. MONTES, E. BARCELO & F. MENES. 1989. The effect of an irrigation and hydroelectric reservoir on its downstream communities. Regulated Rivers. Research and Management 4: 275-284.
DGOH (Direction General de Obras Hidraulicas). 1986. Inventario de Presas Espatiolus 1986. Publ. MOPU, Madrid.
DOADRIO, I., B. ELVIRA y Y. BERNAT (eds.). 1991. Prc,es contitrentales espa6oles. Inl,enturiu y clasificac.i6n de zonasflux~iales. Col. TCcnica. Publ. ICONA, Madrid.
ELVIRA, B., A. ALMODOVAR y J. LOBON-CERVIA
1991. Recorded distribution of sturgeon (Acipenser sturio L.) in the Iberian Peninsula and actual status in spanish waters. Arch. Hydrohiol. 121(2), 253-258.
ELVIRA, B., I. DOADRIO, J. LOBON-CRVIA & A. SOSTOA (in press). Red list of the freshwater fishes of Spain. Procs. 6th Cong. Europ. Icthiol. Budapest.
GARCIA DE JALON, D. 1984. Stream Regulation in Spain. En: Regulated Rivers. A. Lillehammer & S.J. Saltveit (eds.), 48 1-494. Universitets forlaget, Univ. Oslo.
GARCIA DE JALON, D. 1987. River Regulation in Spain. Regulated Ri~sers. Reseurch and Mrrnagemrnt 4(1): 343-348.
GARCIA DE JALON, D. 1990. TCcnicas hidrobiologicas para la fijaci6n de caudales ecologicos minimos. En: Libro honzerzaje a1 Profesor D. M. Gurcfa de Viedmu. 183- 196. A. Ramos, A. Notario & R. Baragaiio (eds.). FUCOVASA. UPM. Madrid.
GARCIA DE JALON, D. & J.V. LOPEZ ALVAREZ. 1983. Contribucion a1 conocimiento de la distribuci6n geografica y mesol6gica de las principales especies piscico- las de la Cuenca del Duero en el verano de 198 1. Actas I
Congreso espatiol de Limnologia: 214-226, Barcelona. GARCIA DE JALON, D., C. MONTES, E. BARCELO,
C. CASADO & F. MENES. 1988. Effects of a Hydroelec- tric Regulation on Fluvial Ecosystems from the Spanish Pyrenees. Regulated Rivers. Research and Marzagement 4(2): 479-492.
GARCIA de JALON, D. & P. SANCHEZ-FERNANDEZ (in pr-ess). Evaluation of downstream effects caused by a new reservoir comparing the fluvial communities before and after its construction. Procs. Fith Int. Symp. Reg~il. Streams. Montana.
MAS HERNANDEZ, J. 1986. La ictiofauna continental de la Cuenca del rio Segura. Evolution historica y estado actual. Anls. Biol. Pub. Univ. Murcia (Biol. Amb. 2) 8, 3-17.
MICHEL, M. 1979. The effect of reservoir implementa- tion on the discharge pf the Rii7et- Dlrero in Central Spain. BS Diss, Dep. Geogr. King's College, London.

MUNOZ, I. & N. PRAT. 1989. Effects of river regulation on the Lower Ebro river (NE Spain). Reg. Rivers: Res. & Mngt. 3, 345-254.
PUIG, M.A., J. ARMENGOL, G. GONZALEZ, J. PE- NUELAS, S. SABATER & F. SABATER. 1987. Chemical and biological changes in the Ter river induced by a series of reservoirs. In: Regulated Streams: Advances in Ecology. J.F. Craig & J.B. Kemper (eds.). Plenum Press. New York.
PRAT, N., M.A. PUIG, G. GONZALEZ & X. MILLET 1983. Chironomid Longitudinal Distribution and Macroin- vertebrate Diversity along the Llobregat River (NE Spain). Mem. Amer. Ent. Soc. 34, 267-278.
PRAT, N., I. MUNOZ, J. CAMP, F.A. COMIN, J.R. LUCENA, J. ROMERO & M. VIDAL. 1988. Seasonal changes in particulate organic carbon and nitrogen in the
river and drainage channels of the Ebro Delta (NE Spain). Ve1.h. Internat. Verein. Limnol. 23, 1344-1 349.
SABATER, F., J. ARMENGOL & S. SABATER. 1989. Measuring discontinuites in the Ter river. Reg. Rivers: Res.
& Mngt. 3, 133-142. SABATER, S., J. ARMENGOL, E. MARTI, F. SABA-
TER & H. GUASCH. 1991. Benthic diatom communities as descriptors of discontinuities in the river Ter, Spain. In:
B.A. Whitton & R.E. Friedrich (eds.). Use of algae for monitoring of river-S. 157- 163. Univ. Insbrick.
WARD, J.V. & D. GARCIA de JALON. 199 1. Epheme- roptera of regulated mountain streams in Spain and Colo- rado. 567-578. In: J. Alba-Tercedor & A. Sanchez-Ortega (eds.). Overview and Strategies of Ephemeroptera and Plecoptera

THE SUBTERRANEAN ENVIRONMENT
A. 1. Camacho and A. G.-Valdecasas Museo Nacional de Ciencias Naturales. José Gutiérrez Abascal 2. 28006- Madrid. Spain.
Keywords: Groundwater, Interstitial fauna, Natural History, Phylogeny, Adaptation and Evolution.
ABSTRACT
This paper is a short essays on the fauna that inhabits the subterranean environment and its bearing to some critica1 and
present day problem in Evolutionary Biology, with a review of the work done on Spanish subterranean waters.
INTRODUCTION
Overview
The recent history of science should be a focus of atten-
tion for those areas of scientific research where a new disci- pline arises. Much sterile discussion and effort could be
avoided if we learnt how similar problems were posed and
solved in nearby fields. Or, at least, the kind of involvement
that should be avoided. Habitat classification in what we here cal1 the subterra-
nean environment is a good example of this. Caves have been known and used since human origins. The interstitial
layers, Epigean and Hypogean that are beneath and surround streams and lakes were known to have a fauna during the first half of this century. Both share the absence of light as
the main joint characteristic. Deep sea also lacks light, but
water composition makes a big difference. Al1 of them could
be properly called subterranean environments- under somet- hing, be it terra, freshwater or marine water-. We deal here
only with those subterranean that could properly be called
freshwater. In order to avoid discussion and to clarify the subject,
the first question to answer is what we understand by habi-
tat or "restricted environment". There are two ways of dealing with this problem. One is to make distinctions and
classifications of the physical space based on the disconti- nuities humans can discriminate visually or by any other way. After the environment is subdivided into homogene- ous parts it is assumed that, al1 else being constant, similar parts will have similar inhabitants. The alternative appro- ach is to look for similar inhabitants first and assume that
the physical space where they live is equivalent in some
way. Of course, many approaches are possible between these two lirnits and Nature is generally indifferent to our efforts to clarify issues.
In the area of Limnology, the trend on Regional Lim-
nology (NAUMANN, 1932) of the second quarter of this century, the efforts to classify lakes on a eutrophic-oligo-
trophic dimension or. more recently the discussion on conti-
nuum versus discrete in lotic environments (VANNOTE et al., 1980; BARNES & MANN, 1980), are good examples of the limited success of classificatory squemes in Nature.
Having said that, we would like to note that the subject of this chapter is the fauna that inhabits the interstitial envi-
ronment of streams, either superficial or inside caves, and al1 other aquatic organisms inside caves. Al1 of them share
the absence of light, and although we realize it is more or
less arbitrary to use this as a delimitatory criteria, we found
other criteria even more arbitrary. A more precise concept of this subterranean environment follows.
Cave and Interstitial fauna.
The subterranean aquatic environment is built up of two sets which have been long since clearly differentiated: subterranean water sensu strictu and interstitial water.
Subterranean water in its broadest sense is found in caves and pits, from fissures and cracks inaccessible to humans to big cavities where subterranean streams flow, to completely inundated galleries (siphons). The interstitial water flows through the sand grains and grave1 deposits of littoral beaches, alluvial layers at the edge of rivers and lakes and the upper part of the sediments below the river bed (hypo-
Liriiiietica, 8: 167-174 (1992)
O Asociación Española de Limnologia, Madrid. Spain

rreic, ORGHIDAM, 1953) as well as the hypotelminorreic layer (MESTROV, 1964).
In caves, the invertebrate fauna of the Iberian Peninsula lives in small ponds, gours, lakes, at the bottom of defiles, subterranean river shores and in any other source of water. Some of these animals, such as Crustacea Syncarida live strictly in this type of water; others have hypogean and epigean species like the copepods, and some, like the isopod Stenasellus live well in either environment. This has made it very difficult to delimit the subterranean fauna and a great variety of criteria ecological, etological os topographical has been used to produce classification. The most general classi- fication and perhaps the most accepted had its origin in Schi- ner-Racovitza which considers three categories of subterra- nean animals: troglobious, a strictly cave animals whose life cycle takes place completely in the cave; troglofilous, a frequent inhabitant of the caves, adapted ecologically but not morphologically, and trogloxenus, casual inhabitants, whose long permanence in the caves could cause them to die. More recently CHAPMAN (1986) takes the view that the only valid criteria for a classification is that based on a single common characteristic and with a deep biological signifi- cance: the absence of light. The scarce knowledge we have about the majority of the species that live in subterranean water projects no light on the degree of adaptation of this fauna to its environment. The interstitial os phreatic fauna that could be found in the other dominium of the subterra- nean waters, and that could only be reached through artifi- cial wells, the Karaman-Chappuis sampling method (CHAP- PUIS, 1942; MOTAS, 1962) os the Bou-Rouch method (BOU, 1974), is composed of permanent frequent, or occa- sional inhabitants as is the case of cave ariiinals.
The most complex classification of this environment could be found in HUSMANN (1966-1971), who creates categories for the fauna as a function of the stream stretch, in terms of the classical river classification, the size of the sediment, adaptation and permanence in the environment (habitual, frequent, os occasional). This kind of classifica- tion is usually avoided today, as it is very difficult to assign the fauna univocally to any of its categories. There are animals that spend only a part of their life cycle in this envi- ronment as do some insect larvae. At this stage, they should be properly considered subterranean inhabitants.
Brief historical review.
The studies on biospeleology have evolved in Spain in a similar fashion to those in other countries but at a lower intensity and with a certain delay at the beginning.
The first records of Spanish biospeleology, around 1861, come from foreign researchers, and the main emphasis is on
taxonomy of the terrestrial cavernicolous fauna. The first
aquatic animal, discovered by Racovitza in 1905 was the
isopod Typhlocirolana moraguesi in the Cuevas del Drach, Mallorca. The first half of this century witnessed an increase in knowledge in very different groups by researchers like
Chappuis, Viets, Coiffait os Delamare Deboutteville. In
Portugal, on the other hand, it was the local researchers who made the earlier contributions to their fauna: names like Braga, Frade os Mateus are the most prominent of this
period. By the fifties Margalef begins to publish his contri-
butions on cavernicolous crustacea. By 1960 there is an increased interest in sporting speleology, and inside the new
clubs, especially in the Basque and Catalan provinces, a growing group of naturalists creates the "criticum minimun
number" that makes biospeleology an established activity in
Spain. Foreign and local researchers have recently contri- buted to the knowledge of our fauna, and a visual summary of what is presently known in relation to the world subte-
rranean fauna is presented in figure 1 . It could be said that
subterranean fauna is still very poorly known in this area, although, as the figure 1 shows, the proportion of species in relation to what is known elsewhere is rather high, espe-
cially in groups like Isopoda, Copepoda and Amphipoda.
A REVIEW OF THE NATURAL HISTORY OF THE SUBTERRANEAN AQUATIC ENVIRONMENT.
On the adaptative value of form and the effects of the absence of light
Life in caves and the interstices have mistified the appro- aches of scientists to the understanding of adaptation. The
afore mentioned properties such as lack of light, constant
environmental variables like humidity os temperature and the scarcity of food, have led cave researchers to directly correlate some organism features with any of these proper-
ties. Interstitial space, for example, has been thought adequate for "verm" type organisms, while the lack of light is thought responsible for the absence of eyes and pigmen- tation (table 1).
As in any other area of field biology, close correlation between organism features and environment has led to two serious problems. First, the need to understand os to prove that the correlation is a real result of adaptation. Second, and

Table 1. Features of interstitial and cave animals.
INTERSTITIAL COMMON CAVES
Dwarfism
Positive Ticmotactism
Anoftalmia
Dispigmentation
Better touch and chemical
senses
Enlargement and compression of the body
Adhesive organs
Paedomorphism
Low fecundity Lesser number of eggs Increase in egg size
DwarfismJGigantism Appendages enlargement
Metabolic economy
Longevity Longer life cycle
mainly of historical interest- although there are still scien- tists that maintain unorthodox points of view- the mecha- nism which explain that adaptation. For some reason, caves have becn the subject of very involved debates between Darwiniari and Lamarckian theorists (e.g. the early century debate in Nort America), and even some contemporary rese- archers like .i8.,iNDEL (1964). DELAMARE DEBOUTTE- VILLE (1973) were dissatisfied with the Darwinian expla- nation.
Although the understanding of the mechanisms of adap- tation is a challenge, no less important is the previous step: establishing that some organ or set of morphology is the result of adaptation. Until now, descriptions of the features of organisms were offered as the only scientific result. The subject of the third part of this paper, is precisely how to prove that some feature is the result of a process of adapta- tion to certain environmental characteristics. But before dealing with this aspect, let us indicate how impressed other researchers have been by what seems to be very good "fitness" to the environment.
Our first example is the water mite Frontipodopsis reti-
culatifrons SZALAY, 1954. This species is usually found in the interstices of streams, as sampled by the Karaman-Chap- puis method, fig. 2a. It is a laterally compressed animal, with very strong fourth legs. There are only three basic body forms in water mites: globular, that are common in the still waters of ponds, lakes and streams, dorso-ventral compres-
poda, that live in still waters, and Frontipodopsis, show this
last body form. The two environments are so different in terms of the demands on locomotion, that it is difficult to accept that both cases are adaptations to the environment. We should not forget the critica1 advice of GOULD &
LEWONTIN (1979) to avoid the analysis of every aspect of an organism as an adaptation. But we are still confronted with the problem of recognizing those that really are.
The second example is a cave Syncarid of the genus lherobathynrlla (fig.2b). The most prominent features of this organism are its slender body form, the lack of eyes and
Mollusca Copepoda lsopoda Amphipoda Acari (Hidrachnellae)
sed forms, frequent i n streams and rivers, and laterally Figure 1. Relationship berween niimber of subterranean species in compressed forms. Only two kinds of water mites, Fronti- Spain versus the world.

the total dispigmentation of the body. As almost al1 synca- rids are cave inhabitants, we are not confronted here with contradictory evidence in the same group. One is, then,
tempted to assign these characteristics to the life in caves. Nevertheles, al1 these characteristics are found in other orga- nisms outside the cave environment (VANDEL, 1964), so
one is still uncertain of the origin of these characteristics in
Iber-ohathyizella. Finally, our third example is the isopod Canrahronisc~ls
prirnitii'us (fig. 2c) an amphibian inhabitant of caves that lacks eyes and is dispigmented. In this case, the temptation
to assign these characteristics to the life in caves is higher than with Iherohathynella, as the close relatives of Canta- broniscus, the wood louses, are heavily pigmented, have eyes and do not play in water.
What is needed in al1 these cases is an objective way of
establishing if a character is present in an organism as a result of its being inherited from an ancestor os if it has
arisen as a new character in the organism under study. Ways to as certain this follow.
UNDERSTANDING THE EVOLUTIONARY HISTORY.
A methodological preamble.
The understanding of the organic diversity (morphologi-
cal, developmental and ecological patterns) shown by the aquatic subterranean fauna can be treated with the tools of Systematics. Characters, classifications and cladograms are
the basic materials in the search for the evolutive process
that has led to those patterns. The first methodological step is to split the evolutive part
from the phylogenetic component (heredity). If a character is not inherited but acquired through the evolutive history
of the taxon we should then look for a casual explanation, be it adaptative (natural selection working as an answer to an environmental situation) or not (random changes, genetic
drift, etc.).
Only a few groups have detailed developed phylogenies,
but classifications are more common, even in subterranean taxa. We can move from a phylogenie to a classification using the convexity criterium (ESTABROOK, 1978; DUNCAN, 1980; MEACHAM, 1980; MEACHAM &
DUNCAN, 1988). With this criteria the results from cladis- tics studies and previus classifications could be compared, and estimations of the evolutive history revised.
Evolutive explanations can be derived from functional explanations in morphology ("completeness" sensu BOCK, 1988) using the tools of Systematics. Morphological studies
try to uncover functional explanations produced by experi-
mental methods and evolutive explanations by comparative methods, al1 embodied inside a phylogenetic framework.
The comparative method (RIDLEY, 1983; HUEY, 1978;
PAGEL & HARVEY, 1988, 1989, 1990; PAGEL, 1988) uses the fact that similar evolutive results come, very frequently, as a response to selective forces and not only randomly. Correlate changes support the evidence of a gene-
ral adaptative relation. With this method one can identify how phenotipic components tend to covariate among them- selves and with the type of life. Such patterns have been
used to suggest and test adaptative explanations of organic
diversity. Characters that are evolutively independent should
be examined together to detect adaptative convergence.
The main obstacle of the method is how to distinguish the similarities due to a common ancestor and the similarities due to parallel and convergent changes.
In order to study the evolution of ecological characters using the comparative method, the most adequate taxono-
mic leve1 is the species (TELENIUS et al., 1989) and their phylogenetic relationships. The phylogenetic trees should be
derived independently from the ecological data. Only those characters that phylogenies show to be new (autopomorp- hies) could be candidates to discover explanations that affect
actual environmental conditions and look for causal rela-
tionships between the characters and the environment (TELENIUS ef al., 1989). The ground for this hypothesis is the common connection between environment and charac-
ter.
Phylogenetic Systematics and Ecology answer qualitati- vely different questions: relations and history versus process and adaptation.
The renewed interest in comparative analysis in ecology
is due to the advent of new methods in Systematics and any field of Biology with a comparative component is favoured by the introduction of the phylogenetic view as one of its tools.
Using copepods as model organisms.
In 1968 ROUCH published a detailed study on the biology of subterranean copepods. His data has been used by other authors (e.&. CULVER, 1982), and is specially rele- vant to the matters we have been discussing up to now. Table 2 is a resumé of the data of Rouch.
Figure 2. Some examples of subterranean inhabitants. a) Fi.onril>odo[~sis i .cti<~~tlut~fi~otis. b) Ihr~i~ohuth.v~iella .Y[>. C ) Curituh,-onis<,us prirnitii~ris.
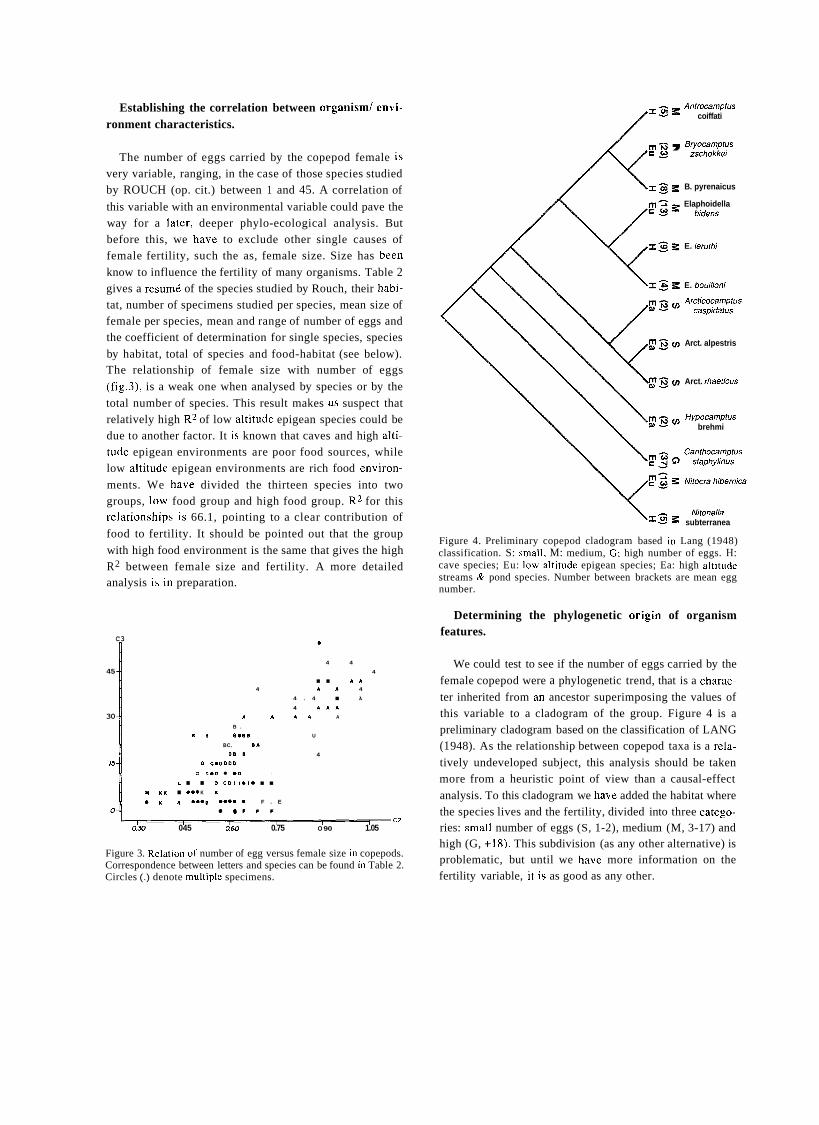
Establishing the correlation between organism/ envi- ronment characteristics.
ti Antrocamptus
/ Y coiffati
The number of eggs carried by the copepod female is very variable, ranging, in the case of those species studied by ROUCH (op. cit.) between 1 and 45. A correlation of
this variable with an environmental variable could pave the way for a later, deeper phylo-ecological analysis. But before this, we have to exclude other single causes of female fertility, such the as, female size. Size has been
know to influence the fertility of many organisms. Table 2 gives a resumé of the species studied by Rouch, their habi- tat, number of specimens studied per species, mean size of female per species, mean and range of number of eggs and the coefficient of determination for single species, species
by habitat, total of species and food-habitat (see below). The relationship of female size with number of eggs
(fig.3), is a weak one when analysed by species or by the
total number of species. This result makes us suspect that relatively high R2 of low altitude epigean species could be due to another factor. It is known that caves and high alti- tude epigean environments are poor food sources, while low altitude epigean environments are rich food environ-
ments. We have divided the thirteen species into two groups, low food group and high food group. R2 for this relationships is 66.1, pointing to a clear contribution of
food to fertility. It should be pointed out that the group
with high food environment is the same that gives the high R2 between female size and fertility. A more detailed analysis is in preparation.
C3
4 4
45 4 .. & A
4 A 4 4
4 . 4 . A
4 A l .
30 1 A 1 4 A
B .
B B B.BU U
BC. U A
DI) B 4
15 o C . O D D 0
D C.D * D I
L . . D C D , 1.1. . . " K K . 0.. K I
. I X . e . 0 o.... F . E
0 Q F F F
C 2 0.33 045 0.60 0.75 O 90 1.05
Figure 3. Relation of number of egg versus female size in copepods. Correspondence between letters and species can be found in Table 2. Circles (.) denote multiple specimens.
m l ~ j , B'yocamptus c zschokkei
B. pyrenaicus
Elaphoidella bidens
E. leruthi
E. bouilloni
Arcticocamptus caspidatus
Arct. alpestris
Arct. rhaeticus
Hypocamptus brehmi
= Nitonei subterranea
Figure 4. Preliminary copepod cladogram based in Lang (1948) classification. S: small, M: medium, G: high number of eggs. H: cave species; Eu: low altitude epigean species; Ea: high altitude streams & pond species. Number between brackets are mean egg number.
Determining the phylogenetic origin of organism features.
We could test to see if the number of eggs carried by the
female copepod were a phylogenetic trend, that is a charac-
ter inherited from an ancestor superimposing the values of this variable to a cladogram of the group. Figure 4 is a preliminary cladogram based on the classification of LANG (1 948). As the relationship between copepod taxa is a rela-
tively undeveloped subject, this analysis should be taken more from a heuristic point of view than a causal-effect
analysis. To this cladogram we have added the habitat where the species lives and the fertility, divided into three catego- ries: small number of eggs (S, 1-2), medium (M, 3-17) and high (G, +la). This subdivision (as any other alternative) is problematic, but until we have more information on the fertility variable, it is as good as any other.
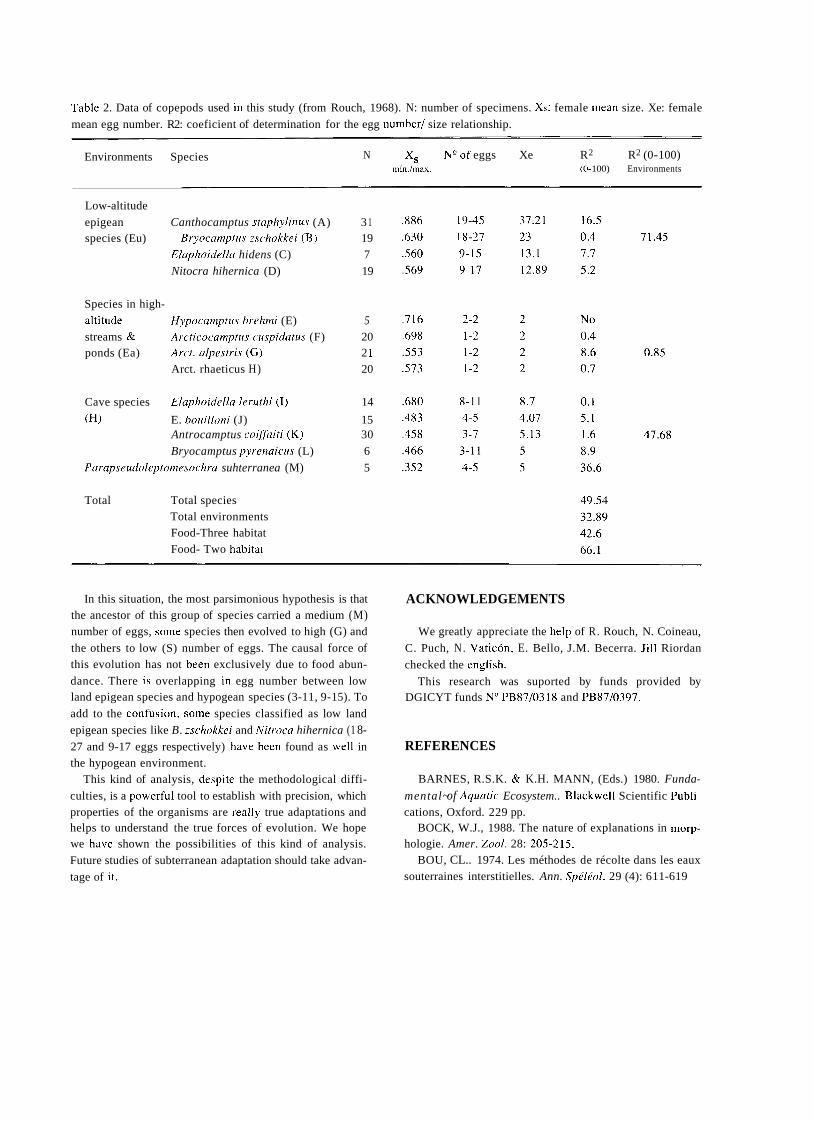
Table 2. Data of copepods used in this study (from Rouch, 1968). N: number of specimens. Xs: female rnean size. Xe: female
mean egg number. R2: coeficient of determination for the egg numberl size relationship.
Environments Species N X s N%f eggs Xe R2 R2 (0- 100) rnin.lrnax. (0- 100) Environments
--
Low-altitude
epigean Canthocamptus staphylinus (A) 3 1 species (Eu) Bryocamptus zschokkei (8) 19
Elal~hoidellu hidens (C) 7
Nitocra hihernica (D) 19
Species in high-
altitude Hypocumpt~ls hrehmi (E) 5 streams & Arcticocamptus cuspidatus (F) 20 ponds (Ea) Arct. alpestris (G) 2 1
Arct. rhaeticus H) 20
Cave species Eluphoidellu leruthi (1) 14
(H) E. houilloni (J) 15 Antrocamptus coiffaiti (K) 30
Bryocamptus pyrenaicus (L) 6
Parapseudoleptomesochra suhterranea (M) 5
Total Total species Total environments Food-Three habitat Food- Two habitat
In this situation, the most parsimonious hypothesis is that the ancestor of this group of species carried a medium (M) number of eggs, some species then evolved to high (G) and
the others to low (S) number of eggs. The causal force of this evolution has not been exclusively due to food abun-
dance. There is overlapping in egg number between low land epigean species and hypogean species (3-1 1, 9- 15). To
add to the confusion, some species classified as low land
epigean species like B. zschokkei and Nitroca hihernica (1 8- 27 and 9-17 eggs respectively) have been found as well in the hypogean environment.
This kind of analysis, despite the methodological diffi-
culties, is a powerful tool to establish with precision, which properties of the organisms are really true adaptations and helps to understand the true forces of evolution. We hope we have shown the possibilities of this kind of analysis.
Future studies of subterranean adaptation should take advan-
tage of it.
ACKNOWLEDGEMENTS
We greatly appreciate the help of R. Rouch, N. Coineau,
C. Puch, N. Vaticón, E. Bello, J.M. Becerra. Jill Riordan
checked the english.
This research was suported by funds provided by DGICYT funds NVB8710318 and PB8710397.
REFERENCES
BARNES, R.S.K. & K.H. MANN, (Eds.) 1980. Funda-
mental~ of Aquutic Ecosystem.. Blackwell Scientific Publi- cations, Oxford. 229 pp.
BOCK, W.J., 1988. The nature of explanations in morp-
hologie. Amer. Zool. 28: 205-215. BOU, CL.. 1974. Les méthodes de récolte dans les eaux
souterraines interstitielles. Ann. Spéléol. 29 (4): 61 1-619

CHAPMAN, P., 1986. Non-relictud cavernicolous inver- tebrates in tropical Asian and Australasian Caves. 9r. Corzg. Inr.Speleo1.. Barcelona. 2: 161 -1 63.
CHAPPUIS, P.A., 1942. Eine neue methode zur Untersu- chung der Grundwasser fauna. Acr. Sc,iential-unz math. nat.
6, Kolozsvar: 3-7. CULVER, D.C., 1982. Cave lifc~. E\~~l~ct ion rrrid Ecolo,qy.
Harvard University Press, Cambridge. 189 pp. DELAMARE DEBOUTTEVILLE, CI., 1973. Conclu-
sions Générales. Ann, Spélu'ol. 28: 343-345. DUNCAN,T., 1980. Cladistics for the practicing taxono-
mist- An eclectic view. Syst. Bor. 5: 149-1 72. 5: 149-172. ESTABROOK, G.F.,1978. Some concepts for the stima-
tion of evolutionary relationships in systematic botany. Syst.
Bot. 3: 146-150. GOULD, S.J. & R.C. LEWONTIN, 1979. The Spandrels
of San Marcos and the Panglossian paradigm: a critique of the adaptationist programme. Pl-oc,. Roy. Soc. Lorzdorz, s 205: 58 1-598.
HUEY, R.B., 1987. Phylogeny, history and the compara- tive method. In: M.E. Feder, A.F. Bennett, W. Burggren &
R.B.Huey (eds). N e n ~ Dil-ections in Ecnlogical Physiology, 76-98. University Press, Cambridge.
HUSMANN, S., 1966. Versuch einer okologischen Glie- derung des interstitiellen Grundwassers in Lebensbereiche eigener Pragung. Arch. Hydl-ohiol. 62: (2): 23 1-268.
HUSMANN, S., 1971. Ecological studies on Freshwater Meiobenthon in Layers of Sand and Gravel. Smithson. Contr. Zool. 76: 161-169.
LANG, K., 1948. Monogl-crphi<> der Harpac,ticides. 2 vol. Lund, 1682 pags.
MEACHAN, C.A., 1980. Phylogeny of the Berberidaceae with an evaluation of classifications. Sysr. Bot. 5: 149-172.
MEACHAN, C.A. & T. DUNCAN, 1987. The necessity of Convex Groups in Biological Classifications. Syst. Bot. 12 (1): 78-90.
MESTROV, M.,1962. Un nouveau milieu aquatique souterrain: le biotope hypotelminorheique. C.R. Acad. Se.
Paris, 254 (D) 14: 2677-2679.
MOTAS, C., 1962. Procédé des sondages phréatiques.
Division du domaine souterrain. - Classification écologique des animaux souterrains- Le psammon. Acta Mus. Maced.
sci. Nat. 8: 135-173.
NAUMANN, E., 1932. Grundzüge der regionalen limno-
logie. Die Binnengrwusser-, 1 1, 176 pp. ORGHIDAM, T., 1953. Un nou domein de viata subte-
rrana Biotopul hiporeic. Bull. sti. sect. Biologir si sfi-Agro-
tzon, si .sec.t. geologiu si geogl-. R.P.R. 7: 3. PAGEL, M.D. & P.H. HARVEY, 1988. Recent develop-
ments in the analysis of comparative data. Quart.Rei~.Biol.
63 (4): 4 13-440. PAGEL, M.D. & P.H. HARVEY, 1989. Comparative
methods for examining adaptation depend on evolutionary models. Folia Prinzutologica 53: 203-220.
PAGEL, M.D. & P.H. HARVEY, 1990. Taxonomic diffe-
rences in the scaling of brain on body size among mammals.
Science 244: 1589-1 593. RIDLEY. M., 1983. The e.xplanafion oforgrrnic. diversity:
the comparativu method und uduptations for mating. Oxford
University Press, Oxford.
ROUCH, R., 1968. Contribution h la connaissance des harpacticides hypogés (Crustacés, Copepods). Ann. Spr'léol.
23: 5-167. SZALAY, L., 1945. Eine neue Art der Gattung Fronti-
podopsis Walt. (Hydrachnellae, Acari) aus unterrirdis-
chen Gewassern der Karpatenbeckens. Fragm. ,faun.
hung. 8: 1-5.
TELENIUS, A,, A. ANGERBJORN & O. ERIKSSON,
1989. On phylogenetic ecology. Evolutiorzat-y Theor-; 8: 351-356.
VANDEL, A., 1964. Biospeologie: la hiologie des
animnu.~ crri~ernicoles. Gauthier-Villars Editeur, Paris. 619
pags. VANNOTE, R.C., G.W. MINSHALL, K.W. CUMMINS,
J.R. SEDELL & C.E. CUSHING, 1980. The river continuum
concept. Canadian Journal of Fisherirs and Aquatic Scien-
ces. 37: 130-137.

THE MICROBIAL COMPONENT IN SPANISH AQUATIC ECOSYSTEMS
R. Guerrerol and C. Pedr6s-Alib2
I Department of Microbiology, University of Barcelona. Spain. 2 Marine Sciences Institute, CSIC, Barcelona, Spain.
Keywords: Microbial ecology, Heterotrophic bacteria, Phototrophic bacteria, Flagellates, Ciliates, Plankton, Phagotrophy.
Regions ($.sorrow, dolef14l shades, wxhere peace A lake is the landscape's most beaut(f'u1 and e.vpressive
And rest can never dw7ell; hut torture without end featilre. Ir i s earth' eye; looking into which the beholder.
Still urges, and a fiery Deluge,,fed measures thedepth of his own nrrture. The f l~c~~iat i le trees
With ever-burning Sulphur unconsumed: next the shore are the slender eyelashes which ,fr-inge it , urzd
Such pluc,e Eterr7al Justic,e had prepar'd.. . the wooded hills and c,l(ffs around are its ol'erhanging brow's.
John Milton, Par-udisc~ Lost (1 667) Henry D. Thoreau, Walden (1854)
Ecology is physiology it1 the wlorst possible c,onditions.
Thomas D. Brock (1966) citing one of his distressed students.
ABSTRACT
Spain has a wide variety of aquatic ecosystems with peculiar chemical composition and hydrological regimes. Microorganisms are the sole inhabitants of many of these sytems and a very important component in most. Studies of microbial ecology in Spanish ecosystems, therefore, provide examples of both "conventional" and "peculiar" environments, where hypotheses about
the role of microorganisms can be tested. This paper provides an overview of the studies of microorganisms carried out in
Spanish aquatic ecosystems and summarizes their contributions to the current paradigm of the bacterioplankton.
INTRODUCTION
Microbial ecology is an extremely young discipline. The first book with such a title was published only 25 years ago
(BROCK, 1966). A good method to determine the number
of bacteria in natural waters, for example, was only esta- blished 15 years ago, and no satisfactory technique yet
exists to determine total production of bacterioplankton. Initially, microbial ecology was only a ragbag of disciplines of applied microbiology in which the Environment had to be
considered: pollution microbiology, environmental micro- biology, microbial leaching of minerals, nitrogen fixation by
symbiotic associations, biogeochemical cycling of nutrients, and so on. The introduction of techniques from medical microbiology, cell and molecular biology and bacterial physiology during the seventies and eighties has completely transformed the discipline. Nowadays microbial ecology is a
field in its own, with efforts centered in the understanding of the ecology of microorganisms. Moreover, the perspective provided by microbial ecology is considered to give the best
possible approach to the understanding of other widely
different fields such as biotechnology or medical microbio- logy. In order to fulfil1 such high hopes, microbial ecology needs to incorporate the ideas and concepts from general ecology and to improve the techniques for the study of
microorganisms in situ. Within the field of limnology, the past 15 years have
witnessed a change of paradigm. The classical food chain (diatom to copepod to fish), has been shown to be only a
part of the whole picture. This was already proposed by POMEROY in 1974, but only in the last ten years were techniques available to probe its intrincacies and to provide quantitative data. After a paradigm becomes established, the exceptions to the rules are probably more interesting than
L~mnet~ca , X: 175-184 (1992) O Asociacion Espaiiola de Llrnnologia, Madrid. Spain

most "orthodox" systems. Studies carried out in Spain offer examples of both orthodox and "heretic" microbial ecosys- tems. The latter, however, have received considerably more attention in our country, in particular the ecology of photo-
trophic bacteria has been intensively studied. This is perhaps, due to the abundance of "peculiar" aquatic envi-
ronments in Spain, which can be considered to be exceptions
Figure I. Map of the Iberian Peninsula with the location of study sites. Triangles pointing up indicate systems with planktonic communities of sulfur phototrophic bacteria. Triangles pointing down indicate systems with benthic communities of phototrophic bacteria. Empty circles indicate systems without communities of phototrophic bacteria. 1. Butron River (Biscaia); 2. Lake Estanya (Huesca); 3. Lake MontcortCs (Lleida); 4. La Massona Coastal lagoon (Girona); 5. Sant Miquel de Campmajor karstic region with Lakes Negre and Coromines (Girona); 6. Banyoles karstic region with Lakes Banyoles, Ciso, and Vilar (Girona); 7. Chiprana hyper- saline pond (Zaragoza); 8. Remolar coastal lagoon (Barcelona); 9. Alfacs Bay in the Ebro Delta (Tarragona); 10. Torcas karstic region, with Laguna de la Cruz and Lagunillo del Tejo (Cuenca); I I. Arcas- 2 and other karstic lakes (Cuenca); 12. Hypersaline lagoons in the endorheic region around Alciizar de San Juan (Ciudad Real); 13. Albufera de Valencia shallow coastal lagoon (Valencia); 14. Cullera coastal lagoon (Valencia); 15. PCtrola hyperasaline lagoon (Alba- cete); 16. Santa Pola saltworks (Alicante); 17. Mar Menor, shallow coastal lagoon (Murcia); 18. La Caldera, high mountain lake (Granada); 19. La Concepcidn reservoir (MBlaga); 20. SantoAa coas- tal lagoon (Santander); 21. Lake El Tobar (Cuenca); 22. Cibollar coastal lagoon (Mallorca).
to the rules (fig. 1). These rules, however, have been defined after the study of a biassed sample of world ecosystems (WILLIAMS, 1988). Therefore, the study of these "pecu- liar" systems should be rewarding and fascinating.
In this chapter we will first summarize the current para- digm of the plankton. Then, relevant studies carried out in
Spain will be reviewed. Finally, studies of aquatic ecosys- tems which do not fit this paradigm will be considered. Environmental and pollution microbiology will not be consi- dered. Likewise, phytoplankton studies will be dealt with in
other chapters.
THE CURRENT PARADIGM OF THE HETEROTROPHIC MICROBIAL PLANKTON
Numerous studies have recently established a new para- digm of the plankton. A few statements form the basis of
this model:
a) Primary production is considered to be carried out mostly by the pico- and nanoplankton.
b) Bacterioplankton biomass in widely different aquatic systems is close to 20 % of phytoplankton biornass
(BIRD & KALFF, 1984), although it can be higher than 100% or lower than 10% (DUCKLOW & CARL- SON, 1992).
C) Bacterioplankton heterotrophic production is thought to be around 50% of primary production (COLE et al., 1988, DUCKLOW & CARLSON, 1992). The former is not only supported by primary production, howe-
ver, but also by allocthonous inputs of dissolved and particulated organic matter.
d) Both the phototrophic and heterotrophic picoplankton are eaten by a large collection of microorganisms,
ranging in size from viruses up to rotifers and crusta-
ceans. Most of these predators select food particles based on their size, although some are able to taste food particles before either ingesting or rejecting them.
e) The slopy feeding activities of these predators, toget- her with their excretion, generate dissolved and parti-
culate organic matter, and nutrients which become available to phyto- and bacterioplankton.
f) The largest of these predators are adequate as food for large invertebrates and vertebrate predators and, thus, close the microbial loop: from phytoplankton to bacte- ria, to heterotrophic flagellates, to ciliates, to rotifers
and crustaceans, to fishes (AZAM et al., 1984).
THE HETEROTROPHIC BACTERIA IN SPANISH AQUATIC ECOSYSTEMS
a) Studies of phytoplankton and primary production will be dealt with in other chapters of the present book.
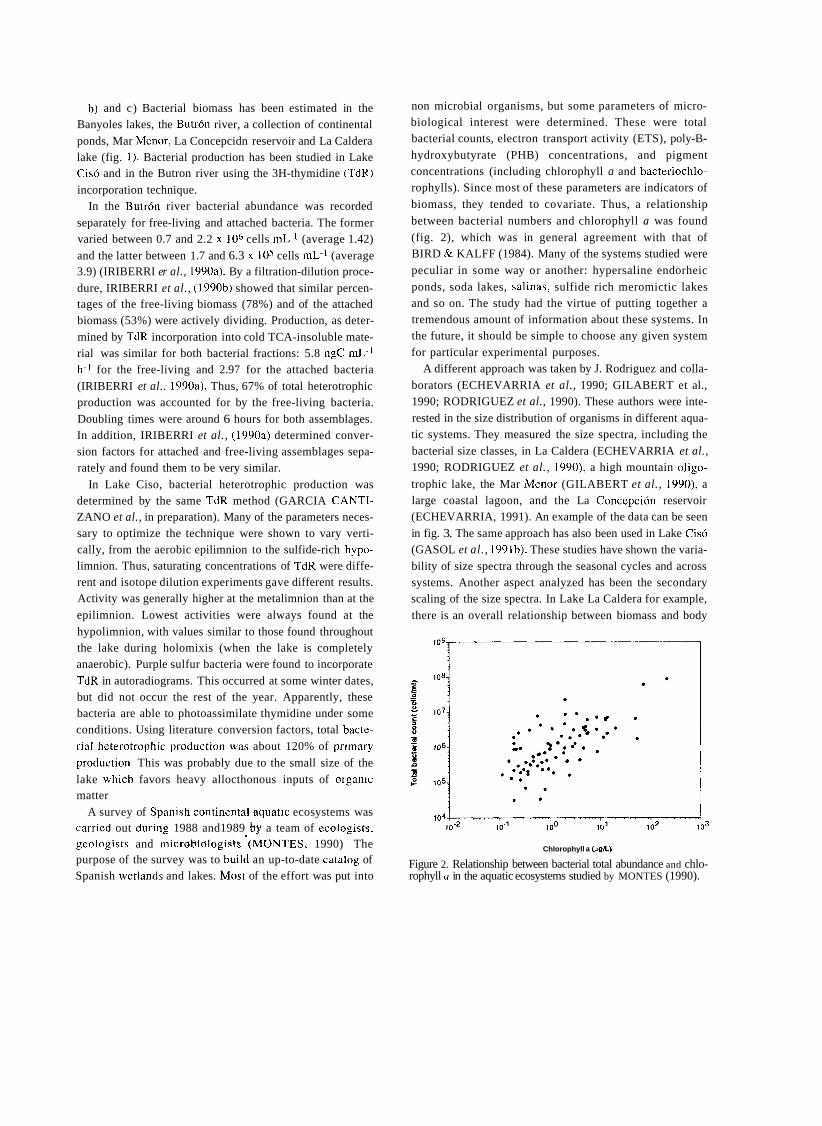
b) and c) Bacterial biomass has been estimated in the Banyoles lakes, the Butr6n river, a collection of continental ponds, Mar Menor, La Concepcidn reservoir and La Caldera lake (fig. l ) . Bacterial production has been studied in Lake Cis6 and in the Butron river using the 3H-thymidine (TdR) incorporation technique.
In the Butr6n river bacterial abundance was recorded separately for free-living and attached bacteria. The former varied between 0.7 and 2.2 X 106 cells mL-l (average 1.42) and the latter between 1.7 and 6.3 X 105 cells mL-l (average 3.9) (IRIBERRI er al., 1990a). By a filtration-dilution proce- dure, IRIBERRI et al., (1990b) showed that similar percen- tages of the free-living biomass (78%) and of the attached biomass (53%) were actively dividing. Production, as deter- mined by TdR incorporation into cold TCA-insoluble mate- rial was similar for both bacterial fractions: 5.8 ngC mL-' h-' for the free-living and 2.97 for the attached bacteria (IRIBERRI et al.. 1990a). Thus, 67% of total heterotrophic production was accounted for by the free-living bacteria. Doubling times were around 6 hours for both assemblages. In addition, IRIBERRI et al., (1990a) determined conver- sion factors for attached and free-living assemblages sepa- rately and found them to be very similar.
In Lake Ciso, bacterial heterotrophic production was determined by the same TdR method (GARCIA CANTI- ZANO et al., in preparation). Many of the parameters neces- sary to optimize the technique were shown to vary verti- cally, from the aerobic epilimnion to the sulfide-rich hypo- limnion. Thus, saturating concentrations of TdR were diffe- rent and isotope dilution experiments gave different results. Activity was generally higher at the metalimnion than at the epilimnion. Lowest activities were always found at the hypolimnion, with values similar to those found throughout the lake during holomixis (when the lake is completely anaerobic). Purple sulfur bacteria were found to incorporate TdR in autoradiograms. This occurred at some winter dates, but did not occur the rest of the year. Apparently, these bacteria are able to photoassimilate thymidine under some conditions. Using literature conversion factors, total bacte- rlal heterotroph~c product~on way about 120% of prlmary product~on This was probably due to the small size of the lake wh~ch favors heavy allocthonous inputs of organlc matter
A survey of Span~sh continental aquatlc ecosystems was carrled out durlng 1988 and1989 by a team of ecolog~sts, geologists and m~crobiolog~st$ '(MC)NTES, 1990) The purpose of the survey was to b u ~ l d an up-to-date catalog of Spanish wetlands and lakes. Most of the effort was put into
non microbial organisms, but some parameters of micro- biological interest were determined. These were total bacterial counts, electron transport activity (ETS), poly-B- hydroxybutyrate (PHB) concentrations, and pigment concentrations (including chlorophyll a and bacteriochlo- rophylls). Since most of these parameters are indicators of biomass, they tended to covariate. Thus, a relationship between bacterial numbers and chlorophyll a was found (fig. 2), which was in general agreement with that of BIRD & KALFF (1984). Many of the systems studied were peculiar in some way or another: hypersaline endorheic ponds, soda lakes, salinas, sulfide rich meromictic lakes and so on. The study had the virtue of putting together a tremendous amount of information about these systems. In the future, it should be simple to choose any given system for particular experimental purposes.
A different approach was taken by J. Rodriguez and colla- borators (ECHEVARRIA et al., 1990; GILABERT et al., 1990; RODRIGUEZ et al., 1990). These authors were inte- rested in the size distribution of organisms in different aqua- tic systems. They measured the size spectra, including the bacterial size classes, in La Caldera (ECHEVARRIA et al., 1990; RODRIGUEZ et al., 1990), a high mountain oligo-
trophic lake, the Mar Menor (GILABERT et al., 1990), a large coastal lagoon, and the La Concepcion reservoir (ECHEVARRIA, 1991). An example of the data can be seen in fig. 3. The same approach has also been used in Lake Cis6 (GASOL et al., 1991b). These studies have shown the varia- bility of size spectra through the seasonal cycles and across systems. Another aspect analyzed has been the secondary scaling of the size spectra. In Lake La Caldera for example, there is an overall relationship between biomass and body
Chlorophyll a @gK)
Figure 2. Relationship between bacterial total abundance and chlo- rophyll u in the aquatic ecosystems studied by MONTES (1990).
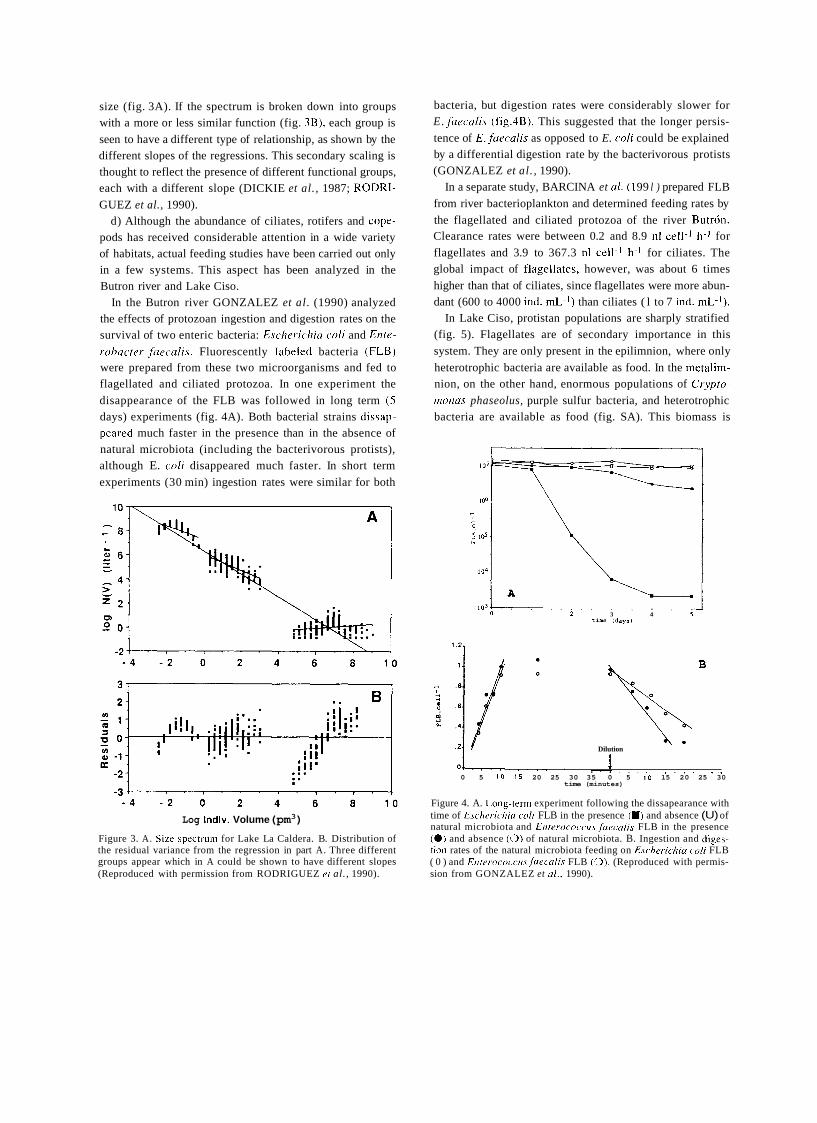
size (fig. 3A). If the spectrum is broken down into groups with a more or less similar function (fig. 3B), each group is
seen to have a different type of relationship, as shown by the different slopes of the regressions. This secondary scaling is thought to reflect the presence of different functional groups, each with a different slope (DICKIE et al., 1987; RODRI-
GUEZ et al., 1990). d) Although the abundance of ciliates, rotifers and cope-
pods has received considerable attention in a wide variety of habitats, actual feeding studies have been carried out only in a few systems. This aspect has been analyzed in the Butron river and Lake Ciso.
In the Butron river GONZALEZ et al. (1990) analyzed the effects of protozoan ingestion and digestion rates on the survival of two enteric bacteria: Escherichiu c,oli and Ente-
rohacter faecu1i.s. Fluorescently labeled bacteria (FLB) were prepared from these two microorganisms and fed to flagellated and ciliated protozoa. In one experiment the
disappearance of the FLB was followed in long term (S days) experiments (fig. 4A). Both bacterial strains dissap- peared much faster in the presence than in the absence of natural microbiota (including the bacterivorous protists), although E. coli disappeared much faster. In short term
experiments (30 min) ingestion rates were similar for both
Log Indlv. Volume (pm3)
Figure 3. A. S i ~ e spectrum for Lake La Caldera. B. Distribution of the residual variance from the regression in part A. Three different groups appear which in A could be shown to have different slopes (Reproduced with permission from RODRIGUEZ cjt al., 1990).
bacteria, but digestion rates were considerably slower for E. faec~ulis (fig.4B). This suggested that the longer persis-
tence of E.faecalis as opposed to E. col; could be explained by a differential digestion rate by the bacterivorous protists (GONZALEZ et al. , 1990).
In a separate study, BARCINA et u1. (1 99 l ) prepared FLB
from river bacterioplankton and determined feeding rates by
the flagellated and ciliated protozoa of the river Butson. Clearance rates were between 0.2 and 8.9 nl cell-) h-' for
flagellates and 3.9 to 367.3 nl cell-' h-l for ciliates. The global impact of i-lagellates, however, was about 6 times
higher than that of ciliates, since flagellates were more abun- dant (600 to 4000 ind. mL-l) than ciliates ( l to 7 ind. mL-l).
In Lake Ciso, protistan populations are sharply stratified (fig. 5). Flagellates are of secondary importance in this
system. They are only present in the epilimnion, where only
heterotrophic bacteria are available as food. In the metalim- nion, on the other hand, enormous populations of Crypto-
nlonas phaseolus, purple sulfur bacteria, and heterotrophic bacteria are available as food (fig. SA). This biomass is
io4
103. . . . . . . . . , . , . 3 4 5
time (daysi
kB Dilution
. , . , . , . , , , l , , , , , , , . , 0 5 10 15 20 25 30 35 0 5 10 l5 20 25 30
time (minutes)
Figure 4. A. Long-tenn experiment following the dissapearance with time of E.scher.ic,hirr c,oli FLB in the presence (M) and absence (U) of natural microbiota and Enter.ococ~c~~.sfaeculis FLB in the presence ( 0 ) and absence (0) of natural microbiota. B. Ingestion and diges- tiori rates of the natural microbiota feeding on E.sc./zer.ic~/zia col; FLB ( 0 ) and Errrrr.oc~oc~c~u.s fa~~cuIi.s FLB (3). (Reproduced with permis- sion from GONZALEZ et al., 1990).
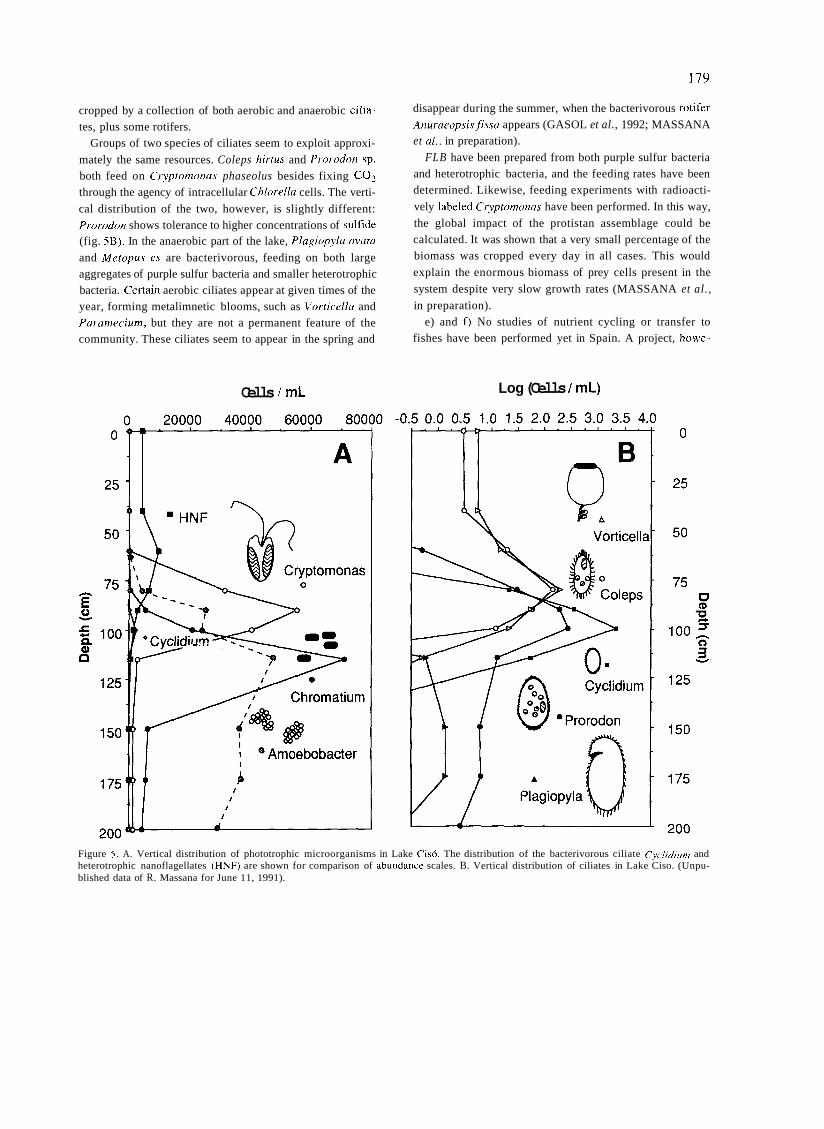
cropped by a collection of both aerobic and anaerobic cilia- tes, plus some rotifers.
Groups of two species of ciliates seem to exploit approxi-
mately the same resources. Coleps hirtus and Prot-odon sp. both feed on Cryptomonas phaseolus besides fixing CO2 through the agency of intracellular Chlorella cells. The verti-
cal distribution of the two, however, is slightly different: Prorodon shows tolerance to higher concentrations of sulfide (fig. 5B). In the anaerobic part of the lake, Plagiopyla ovuta
and Metopus es are bacterivorous, feeding on both large aggregates of purple sulfur bacteria and smaller heterotrophic bacteria. Certain aerobic ciliates appear at given times of the year, forming metalimnetic blooms, such as Vorticella and Paramecium, but they are not a permanent feature of the community. These ciliates seem to appear in the spring and
disappear during the summer, when the bacterivorous rotifer Anuraeopsisfi'ssa appears (GASOL et al., 1992; MASSANA et al.. in preparation).
FLB have been prepared from both purple sulfur bacteria and heterotrophic bacteria, and the feeding rates have been determined. Likewise, feeding experiments with radioacti-
vely labeled C1:yptomonas have been performed. In this way,
the global impact of the protistan assemblage could be calculated. It was shown that a very small percentage of the biomass was cropped every day in all cases. This would
explain the enormous biomass of prey cells present in the
system despite very slow growth rates (MASSANA et al.,
in preparation). e) and f) No studies of nutrient cycling or transfer to
fishes have been performed yet in Spain. A project, howe-
Cells 1 mL Log (Cells 1 mL)
Figure 5 . A. Vertical distribution of phototrophic microorganisms in Lake Cisd. The distribution of the bacterivorous ciliate Cyclidium and heterotrophic nanoflagellates (HNF) are shown for comparison of abundance scales. B. Vertical distribution of ciliates in Lake Ciso. (Unpu- blished data of R. Massana for June 11, 1991).

ver, has been initiated in Lake La Caldera (Granada) to determine the effects of zooplankton on phosphorous recy- cling in this oligotrophic high-mountain lake (L. CRUZ- PIZARRO, personal communication).
THE ECOLOGY OF AUTOTROPHIC PROKARYOTES
Distribution of phototrophic bacteria
Several authors have described the presence of autotrop-
hic bacteria in many aquatic ecosystems (fig. 1). GUERRE-
R 0 et al. (1987) reviewed information about many water bodies in the Mediterranean region of Spain. Further detai- led studies have been conducted in the karstic lakes of Cuenca (MIRACLE et al., 1992; VICENTE & MIRACLE, 1984, 1988). Their presence has also been noted in karstic lakes in the Pre-Pyreneean region (CAMPS et al., 1976; AVILA et al., 1984; GUERRERO et al., 1987) and in coas- tal lagoons such as La Massona (RIERA & ABELLA 1986, 1991), El Remolar (GUERRERO et al., 1987), Cullera
(MIRACLE & VICENTE 1985) and Cibollar (MOYA et al., 1987). Finally, the survey of Spanish continental systems (MONTES 1990) provided a wealth of information on the distribution of phototrophic bacteria throughout the country.
Only a brief summary is possible here. There are three types of aquatic environments where
phototrophic bacteria are usually present: inland hypersa-
line shallow lakes and salterns, coastal lagoons with fresh-
water above marine water, and karstic lakes. Spanish hyper- saline systems almost always show the presence of micro- bial mats (GUERRERO & DE WIT, 1992), while the other two types of systems usually have layers of planktonic
purple or green sulfur bacteria at the thermocline or chemo- cline (MIRACLE & VICENTE, 1985; MOYA et al., 1987; RIERA & ABELLA, 1986, 1991; GUERRERO & DE WIT, 1992; MIRACLE et al., 1992).
Due to the generally dry climate evaporation is strong and, as aconsequence, hypersaline systems are very abun- dant and extremely diverse in Spain (MONTES & MARTINO, 1987). In addition endorheic areas are quite
extensive. Therefore, comercially exploited and abandoned salterns are quite frequent both in coastal areas (such as Santa Pola in Alicante) and in the ~nterior (for example in the shallow lake of Quero, in Ciudad Real). Other shallow lakes are athalassohaline, with magnesium as the dominant anion, such as Chiprana (Zaragoza, GUERRERO et al.,
1991) or PCtrola (Albacete). Tide pools and marsh areas also become frequently hypersaline such as in the Alfacs Bay in the Ebro Delta (ESTEVE et al., 1992b). Most of these have been shown to have microbial mats with diffe- rent degrees of development. The variety of chemistries
and hydrological conditions suggest a diversity of species and adaptation mechanisms which should be extremely interesting to study.
Most of the planktonic communities of phototrophic bacteria are found in karstic lakes and coastal lagoons. Karstic lakes are very frequent in the Eastern portion of the Iberian Peninsula, which is mostly calcareous (fig. 1). A classification of such systems is given in GUERRERO et al. (1987). A more detailed scheme of karstic lakes can
be found in MIRACLE et al. (1992). These are usually deep, round shaped, dolines formed by dissolution and collapse of the terrain. If gypsum layers occur the lakes
have high sulfate concentration (for example in the Banyoles area, sulfate concentrations are about half of those in sea water). On the other hand, some of the torcas in Cuenca are very low in sulfate (for example Laguna de la Cruz, VICENTE & MIRACLE, 1984, 1988). The
presence of sulfate is important because it usually results in sulfate reduction by bacteria (Desulfovihrio-like), and consequently in the production of sulfide in the lower layers of lakes. This sulfide provides reducing power for the phototrophic sulfur bacteria and allows their develop- ment if enough light reaches the anaerobic waters (MIRA- CLE et al., 1992).
The coastal lagoons are usually old river mouths, with marine water under a layer of freshwater. In these cases, sulfate concentrations are the usual ones in sea water and sulfide is always present: The particular species of photo- trophic bacteria seem to be different from those found in
freshwaters (MIRACLE & VICENTE, 1985; MOYA et al., 1987; RIERA & ABELLA, 1986, 1991).
GUERRERO et al. (1 987) attempted a biogeography of sulfur phototrophic bacteria. They noted a significant trend
for species of purple bacteria with the carotenoid okenone to be dominant in planktonic systems, unlike in laboratory isolates. However, since most identifications of strains rely on isolation of pure cultures, a true species distribution has not been possible.
Planktonic phototrophic bacteria are always found in layers below the sulfide-oxygen interface. Usually, low light intensities in this region are sufficient to allow slow growth of the bacteria, while sulfide from below provides the neces- sary reducing power.

Light determines whether green or purple sulfur bacte- ria dominate planktonic communities. This was shown by MONTESINOS et al. (1983) and MONTESINOS & ESTEVE (1984). These authors showed how purple bacteria were, in general, more abundant when epilim- netic algae were few or absent. Green bacteria were dominant in the opposite case. Green bacteria also tended to be more abundant in lakes where the sulfide- oxygen interface was deep than in lakes where it was shallow. In fact, the presence of purple bacteria was shown to affect the quality of light in Lake Ciso in such a way that Chlorobium limicola was present, while Chlo- robium phaeobacteroides dominated in the absence of purple bacteria (MONTESINOS et al., 1983).
It is interesting to note that accumulations of phototrop- hic bacteria are usually accompanied by similar layers of other organisms. In particular Oscillatoria spp. and Crypto- monas spp. have been found slightly above or below the layers of phototrophic bacteria in several lakes (MIRACLE et al., 1992). These layers of primary producers coexist with heterotrophic organisms. In particular, ciliates such as Coleps hirtus and rotifers (see below) form extremely dense layers above those of phototrophs. Thus, starting from above, one usually encounters a rotifer, Chlorella bearing ciliates such as Coleps or Prorodon, Cryptomonas or a cyanobacterium, purple bacteria, and green bacteria. These layered structure is analogous to that found in the benthic microbial mats, but spans cm or m instead of mm. The whole community has been shown to migrate vertically through die1 cycles in Lake Cis6 (GASOL er al., 1991a; PEDROS-ALIO & SALA, 1990).
Growth
Production of autotrophic bacteria has been determined in Cullera, where green sulfur bacteria predominate (MIRACLE & VICENTE, 1985) and in Lake Cis6, where purple bacteria are generally more important (GARCIA CANTIZANO et al., 1987). In both cases the bacteria were observed growing very slowly. In Lake Cis6, doubling times ranged from 5 to 50 days (MAS et al., 1990; PEDROS-ALIO er al., in prep.). This extremely slow growth was caused by light limitation, as shown by the enhanced CO2 fixation when bacterial samples were incubated at light intensities higher than ambient (MIRA-
CLE & VICENTE, 1985; GUERRERO et al., 1985). Neit- her sulfide nor phosphate enhanced CO2 fixation in Lake Cis6.
Losses
The enormous biomasses accumulated by sulfur photo- trophic bacteria with such slow growth rates would argue for little losses. Sedimentation, decomposition, and washout were measured in Lake Cis6 through a year cycle (MAS et al., 1990). Estimates of these losses were in agreement with slow growth, but the impact of predation had not been deter- mined and, thus, it remained as a possibility. Potential predators such as the anaerobic ciliates Caenomorpha, Plagiopyla and Metopus, or metalinmetic Scuticociliates, Coleps or Prorodon have been found and identified in Arcas-2 (FINLAY er al., 1991). These ciliates were also found in Lake Ciso (DYER et al., 1986) but their quantita- tive importance had not been determined until recently (fig. 5 ; MASSANA et al., in prep.).
Another potential cause of predatory losses was the accu- mulation of rotifers in the metalimnion, which has been recorded in most meromictic lakes studied (GUERRERO et al., 1978; MIRACLE & VICENTE, 1983; GASOL et al., 1992). One of the most frequently found rotifers is Anurar- opsis fissu and its congenere A. miraclei. The former has been found in Lake Cis6 (PEDROS-ALIO et al., 1986), Laguna de la Cruz and Lagunillo del Tejo (MIRACLE &
VICENTE, 1983), forming abundant masses at the upper part of the metalimnion (up to 101 individuals mL~I in Lake Cis6). Several other rotifers have been found to accumulate at the metalimnion of meromictic lakes: Synchaeta grimpei,
S. tremula, Notholca salina and Others in Cullera, Filinia terminalis , F , h~ fmann i and A. ,fi.ssa in basin I11 of Lake Banyoles, and F. hnfmanni and Keratella quadrata in Laguna de la Cruz (GUERRERO et al., 1978; MIRACLE &
VICENTE, 1983; MIRACLE et al., 1992). Since dense layers of phototrophic bacteria are found
slightly below rotifer and ciliate accumulations in all cases, it is tempting to assume a trophic relationship. Morebver, purple sulfur bacteria have been observed in the interior of several of these ciliates (FINLAY et al., 1991), and seve- ral of the rotifers were known to be at least partially bacte- rivorou's. MASSANA and PEDROS-ALIO (in prep.) carried out experiments with fluorescently labeled hetero- trophic and purple sulfur bacteria in Lake Cis6 to quantify this potential relationship. It could be shown that both cilia- tes and rotifers were able to feed on the bacteria in experi- mental bottles. In the lake, however, the populations of predators and prey were slightly separated in space and, thus, the impact of predation was very small for the photo- trophic bacteria.

40000
t Dunallella
3 m
20000
l WOO
0
Figure 6. Abundance of the phototrophic eukaryote Durralic~llu sp. (e) along a salinity gradient in the Santa Pola salterns, and average counts of nonpigmented colonies (bacteria, 0) and pigmented colo- nies (archaea, U) in different media inoculated with samples from ponds of different salinities. (Redrawn, with permission, from RODRIGUEZ-VALERA et ul . , 198 1 , 1985).
The last possible source of predatory losses is the exis-
tence of predatory bacteria and viruses. Viruses have not
been studied in this context, but the existence of several
predatory bacteria has been shown in Lakes Estanya
(ESTEVE et al., 1983, 1992a; GUERRERO et al., 1986), Arcas-2 (VICENTE et al., 1991), Ciso (GUERRERO et al.,
1986; ESTEVE et al., 1992a; GAJU et al., 1992) and Vilar
(GAJU et al., 1992) always attacking purple sulfur bacteria. These predatory bacteria tend to be more abundant where
the purple bacteria are less active, that is, in the deeper parts
of the bacterial layer, suggesting that their r ~ l e is that of
opportunistic scavengers. Their real impact, however,
remains to be assessed. In summary, biomass and production of phototrophic
bacteria are controled by the light intensity reaching their
depths. Species composition, on the other hand, is contro-
lled by the light quality and the amount of sulfide present. These organisms generally form enormous accumulations of
biomass despite slow growth thanks to small losses. In parti-
cular predators are not efficient at cropping this biomass.
Alltogether, the studies of phototrophic bacteria in Spanish ecosystems constitute the most complete study of the
ecology of any particular group of bacteria available.
STUDIES OF HYPERSALINE ENVIRONMENTS
Another type of system which has received some attention
is that of hypersaline environments. Although not centered on the microbial ecology of the salterns, the studies provided
interesting ecological information on the distribution of halophilic microorganisms (RODRIGUEZ-VALERA, 1986).
The system studied was the Santa Pola salterns (fig. l ) in
Alicante. These salterns provided a series of ponds of incre-
asing salinity, from seawater up to salt saturation. Samples from the ponds were spread on agar plates with different
media for halophilic bacteria (RODRIGUEZ-VALERA et
al., 1981). Essentially all halotolerant bacteria disappeared
at salinities above 10%0 total salts (w/v). Only halophilic
bacteria remained (fig. 6). These could be divided into rnode- rate halophiles, found from 10 to 25%0 salinity, and extreme
halophiles, found from 25%0 salinity to saturation. The
former were a diverse assemblage of eubacteria, including
Pseudornonas, Alcu1igrne.r and other bacteria (RODRI-
GUEZ-VALERA et al., 1985). The latter, on the other hand,
were exclusively archaea of the genera Halohac~tei.iurn, Halo-
c o ~ ~ ~ ~ u . s and Haloarc~ la (RODRIGUEZ-VALERA et al.,
1985). Most of the eukaryotic microorganisms disappeared
above 15%0 salinities, except Dunaliella which had its maxi-
mal abundance at 25%0 salinity, with 105 cells m ~ - ' (fig. 6).
Finally, microbial mats were found in most ponds, with
higher diversity of microorganisms at about 10% salts. These ponds should provide an excellent laboratory for studies of
microbial ecology.
CONCLUSION
Due to its climate and topography, Spain provides a rich
variety of permanent and temporary wetlands. Many of
these systems are hypersaline, anaerobic or both. Thus, they
are extremely interesting from the microbial point of view.
The recently carried out survey of Spanish wetlands (MONTES, 1990) has been instrumental in providing an up-
to-date catalog of available systems. It should be possible to
use such a wide array of aquatic systems as a natural labo-
ratory for experimental microbial ecology.
REFERENCES
AVILA, A., J.L. BURRELL, A. DOMINGO, E. FER-
NANDEZ, J. GODALL, & J.M. LLOPART, 1984. Limno-
logia del Lago Grande de Estanya. Oec,ol. Ayuat. 7: 3-24. AZAM, F., T. FENCHEL, J.G. FIELD, J.S. GREY, L.A.
MEYER-REIL, & F. THINGSTAD, 1983. The ecological
role of water-column microbes in the sea. Mar. Ecol. Pt-ogr. Set.. 10: 257-263.

BARCINA, I., B. AYO, A. MUELA, L. EGEA, & J. IRIBERRI, 199 1. Predation rates of tlagellate and ciliated protozoa on bacterioplankton in a river. FEMS Mic.rohio1. Ecol. 85: 141-150.
BIRD, D.F., & J. KALFF, 1984. Empirical relationship between bacterial abundance and chlorophyll concentration in fresh and marine waters. Can. I . Fish. Aquat. Sci. 41: 1015-1023.
BROCK, T.D., 1966. Principles of Microbial Ecology. Prentice Hall, Englewood Cliffs, New York
CAMPS. J., 1. GONZALVO, J. G ~ ~ E L L , P. LOPEZ, A. TEJERO, X. TOLDRA, F. VALLESPINOS, & M. VICENS. 1976. El lago de MontcortCs, descripcionde un ciclo anual. Oecol. uyuur 2: 99- 1 10.
COLE, J.J., S. FSNDLAY, & M.L. PACE, 1988. Bacterial production in fresh and saltwater ecosystems: a cross-system overview. Mar. Ecol. Prog. Set-. 43: 1-10.
DICKIE, L.M., S.R. KERR, & P.R. BOUDREAU, 1987. Size dependent processes underlying regularities in ecosys- tem structure. Ecol. Monogr-. 57: 233-250.
DUCKLOW, H.W., & C.A. CARLSON, 1992. Oceanic bacterial production. Adv. Microbial Ecol. 12 (in press).
DYER, B.B., N. GAJU, C. PEDROS-ALIO, I. ESTEVE, & R. GUERRERO, 1986. Ciliates from a freshwater sulfu- return. Biosystems 19: 127- 135.
ECHEVARRIA, F., 1991. Variabilidad espacio-temporal de la estructura de tamaiios del plancton en dos ecosistemas acuiticos epicontinentales. Ph. D. Thesis, Urlit~er-sity of'
Mrilaga. ECHEVARRIA, F., P. CARRILLO, F. JIMENEZ, P.
SANCHEZ-CASTILLO, L. CRUZ-PIZARRO, & J. RODRI- GUEZ, 1990. The size-abundance distribution and taxono- mic composition of plankton in an oligotrophic, high moun- tain lake (La Caldera, Sierra Nevada, Spain). J. Plankton Res. 12: 41 5-422.
ESTEVE, I., R. GUERRERO, E. MONTESINOS, & C. ABELLA, 1983. Electron microscopy study of the interac- tion of epibiontic bacteria with Chromatium minus in natu- ral habitats.. Microh. Ecol. 9: 57-64.
ESTEVE, I., N. GAJU, J. MIR & R. GUERRERO, 1992a. Comparison of techniques to determine the abundance of predatory bacteria attacking Chromatiaceae. FEMS Mien- hiol. Ecol., 86: 205-2 l l .
ESTEVE, I., M. MARTINEZ ALONSO, J. MIR & R. GUERRERO, 1992b. Distribution, typology and structure of microbial mat communities in Spain. Preliminary studies. Limnetica 8.
FINLAY. B.J., K.J. CLARKE, E. VICENTE, & M.R. MIRACLE, 1991. Anaerobic ciliates from a sulphide-rich solution lake in Spain. Europ. J. Pmtistol. 27: 148-159.
GAJU, N., I. ESTEVE & R. GUERRERO, 1992. Distri- bution of predatory bacteria that attack Chromatiaceae in a sulfureous lake. Mic.rohio1. Er.ol., (in press).
GARCiA CANTIZANO, J., J.M. GASOL, & C. PEDROS-ALIO, 1987. Produccicin primaria por fototrofia y por quimiotrofia en la laguna de Cis6. A(,tus IV Cong.Esp.
Limnologia (Sevilla) pp. 75-84. GASOL, J.M., J. GARCiA-CANTIZANO, R. MASSANA,
F. PETERS, R. GUERRERO, & C. PEDROS-ALIO, 1991a. Diel changes in the microstratification of the metalimnetic community in Lake Ciso. Hydrohiologiu 2 11 : 227-240.
GASOL, J.M., R. GUERRERO & C. PEDROS-ALIO, 1991 b. Seasonal variations in size structure and procaryotic dominance in sulfurous Lake Ciso. Limnol. Oceunogr. 36: 860-872.
GASOL, J.M., F. PETERS, R. GUERRERO, & C. PEDROS-ALIO, 1992. Community structure in Lake Ciso: Biomass allocation to trophic groups and differing patterns of seasonal succession in the meta- and epilimnion. Arch. Hydrohiol. 123: 275-303.
GILABERT, J., J. RODRIGUEZ, & F. JIMENEZ- GOMEZ, 1990. The planktonic size-abundance spectrum in an oligotrophic, hypersaline coastal lagoon, the Mar Menor, Murcia, Spain. Proc. 24th Europ. Mar. Biol. Symp., M. Burnes and R N Gihson (eds.) , Trophic r-c.1ationship.s in the
marinc~ environment, Aberdeen U.P. : 18-27. GONZALEZ, J.M., J. IRIBERRI, L. EGEA, & I.
BARCINA, 1990. Differential rates of digestion of bacteria by freshwater and marine phagotrophic protozoa. Appl.
En~-ir.oti. Mic.rohio1. 56: 185 1 - 1857. GUERRERO, M.C. & R. DE WIT, 1992. Microbial mats
in the inland saline lakes of Spain. Limnetica 8. GUERRERO, M.c:, J , BALSA, M. PASCUAL, B.
MARTSNEZ & C. MONTES, 1991. Caracterizacion limno- logica de la laguna salada de Chiprana y sus comunidades de bacterias fototroficas. Limnetica 7: 83-96
GUERRERO, R., C.A. ABELLA, & M.R. MIRACLE, 1978. Spatial and temporal distribution of bacteria in a
meromictic karstic lake basin: relationship with physico- chemical parameters and zooplankton.. Verh. Internat.
Verein. Limnol. 20: 2264-2274. GUERRERO, R., E. MONTESINOS, C. PEDROS-ALIO,
I. ESTEVE, J. MAS, H. VAN GEMERDEN. P.A.G. HOFMAN, & J.F. BAKKER, 1985. Phototrophic sulfur bacteria in two Spanish lakes: Vertical distribution and limi- ting factors. Limnol. O c e a n o ~ r . 30: 9 19-93 1.
GUERRERO, R., C. PEDROS-ALIO, I. ESTEVE, J. MAS, D. CHASE, & L. MARGULIS, 1986. Predatory

prokaryotes: predation and primary consumption evolved in bacteria. Proc. Natl. Acad. Sci. USA 83: 2138-2142.
GUERRERO, R., C. PEDROS-ALIO, I. ESTEVE, & J. MAS, 1987. Communities of phototrophic sulfur bacteria in lakes of the Spanish mediterranean region. Acta Academ.
Ahoen. 47: 125-155. IRIBERRI, J., M. UNANUE, B. AYO, I. BARCINA, &
L. EGEA, 1990a. Bacterial production and growth rate esti- mation from [3H]thymidine incorporation for attached and free-living bacteria in aquatic systems. Appl. Environ.
Microhiol. 56: 483-487. IRIBERRI, J., M. UNANUE, B. AYO, I. BARCINA, & L.
EGEA, 1990b. Attached and free-living dividing bacteria in two aquatic systems. Letters Appl. Microhiol. l l : 87-89.
MAS, J., C. PEDROS-ALIO, & R. GUERRERO, 1990. In situ specific loss and growth rates of purple sulfur bacteria in Lake Cisb. FEMS Micr-ohiol. Ecol. 73: 271-281.
MIRACLE, M.R., & E. VICENTE, 1983. Vertical distri- bution and rotifer concentrations in the chemocline of mero- mictic lakes. Hydrohiologia 104: 259-267.
MIRACLE, M.R., & E. VICENTE, 1985. Phytoplankton and photosynthetic bacterial sulfur bacteria productionin the meromictic coastal lagoon of Cullera (Valencia, Spain). Verh. 1nter.nat. Vr~rein. Limnol. 22: 2214-2220.
MIRACLE, M.R., E. VICENTE & C. PEDROS-ALIO, 1992. Biological studies in Spanish meromictic and strati- fied karstic lakes. Limneticu 8.
MONTES, C & P. MARTINO, 1987. Las lagunas salinas espafiolas. In: Seminario sobre bases cientljeicas para la proteccidn de las h14madudes en Espafia: 95-145. Real Academia de Ciencias de Madrid.
MONTES, C. (Coord.), 1990. Estudio de I N S zonas hume- das de la Espafip peninsulur-. In~xentario y tip(ficacidn. INITEC, Ministerio de Obras Publicas y Urbanismo. Madrid.
MONTESINOS, E., & I. ESTEVE, 1984. Effect of algal shading on the net growth and production of phototrophic sulfur bacteria in lakes of the Banyoles karstic area.. Verh. Internat. Verein. Limnol. 22: 1 102-1 105.
MONTESINOS, E., R. GUERRERO, C. ABELLA, & I. ESTEVE, 1983. Ecology and physiology of the competition for light between Chlorobium limicolu and Chlorobium phueohacteroides in natural habitats. Appl. Environ. Micro- biol. 46: 1007-1016.
MOYA, G., G. RAMON, A. MARTiNEZ TABERNER, V. FORTEZA, C. PICO, C. PONSELL, R. ROSSELLO, &
M.A. SOBERATS, 1987. Limnology of a meromictic coas- tal lagoon, L'Estany Cibollar (Majorca, Balearic Islands). Limnetica 3(2): 255-262.
PEDROS-ALIO, C., J.M. GASOL, & R. GUERRERO, 1986. Microbial ecology of sulfurous Lake Cis6. In.: F.
Megusar & M. Gantar (Eds). Perspectives in Microbial Ecology. Slovenian Soc. Microhiol, Ljubjuna, Yugoslavia : 638-643.
PEDROS-ALIO, C., & M.M. SALA, 1990. Microdistri- bution and die1 vertical migration of flagellated vs. gas- vacuolate purple sulfur bacteria in a stratified water body. Limnol. Oceanogr. 35: 1637- 1644.
POMEROY, L.R.. 1974. The ocean's food web, a chan- ging paradigm. BioScience 24: 499-504.
RIERA, X G. & C.A. ABELLA, 1986. Dinamica anual de les poblacions de bacteris fototrbfics de la llacuna costanera de La Massona (Alt Emporda, Girona). Scientia gerund. 12: 35-48.
RIERA, X G. & C.A. ABELLA, 1991. Limnological cycle of the coastal lagoon La Massona (Girona, NE Spain). Origin, dynamics and influence of the sporadic sewater infalls on the meromixis. Vel-h. Internat. Verein. Limnol. 24: 1029-1031.
RODRIGUEZ, J., F. ECHEVARRIA, & F. JIMENEZ- GOMEZ, 1990. Physiological and ecological scalings of body size in an oligotrophic, high mountain lake (La Caldera, Sierra Nevada, Spain). J. Plank. Res. 12: 593-599.
RODRIGUEZ-VALERA, F., 1986. The ecology and taxo- nomy of aerobic chemoorganotrophic halophilic eubacteria. FEMS Microhiol. Reviews 39: 17-22.
RODRIGUEZ-VALERA, F., F. RUIZ-BERRAQUERO, & A. RAMOS-CORMENZANA, 198 1. Characteristics of the heterotrophic bacterial populations in hypersaline envi- ronments of different salt concentrations. Microb. Ecol. 7: 235-243.
RODRIGUEZ-VALERA, F., A. VENTOSA, G. JUEZ, & J.F. IMHOFF, 1985. Variation of environmental features and microbial populations with salt concentrations in a multi-pond saltern. Microh. Ecol. 11: 107-1 15.
VICENTE, E., & M.R. MIRACLE, 1984. Distribution of photosynthetic organisms in a temporal stratified karstic pond near Cuenca (Spain). Verh. Internat. Verein. Limnol.
22: 1504-1710. VICENTE, E., & M.R. MIRACLE, 1988. Physicochemi-
cal and microbial stratification in a meromictic karstic lake of Spain. Verh. Internat. Verein. Limnol. 23: 522-529.
VICENTE, E., M.A. RODRIGO, A. CAMACHO, & M.R. MIRACLE, 1991. Phototrophic prokaryotes in a karstic sulphate lake. Verh. Internal. Verein. Limnol. 24: 998-1004.
WILLIAMS, W.D., 1988. Limnological imbalances: an antipodan viewpoint. Freshwat. Biol. 20: 407-420.

DISTRIBUTION, TYPOLOGY AND STRUCTURE OF MICROBIAL MAT COMMUNITIES IN SPAIN: A PRELIMINARY STUDY
1. Esteve1, M. M a r t í n e z - A l o n s o l , J. M i r l and R. G u e r r e r o 2
1 Department of Genetic and Microbiology and Institute for Fundamental. Biology. Autonomous University of Barcelona, Bellaterra, Spain.
2 Department of Microbiology, University of Barcelona, Spain.
Keywords: Microbial mats, Cyanobacteria, Purple sulfur bacteria, Chlorophyll a , Bacteriochlorophyll a .
ABSTRACT
This work includes a preliminary study of some microbial mats located in the Mediterranean region of the Iberian Penin- sula. The structure of these laminated communities has been studied through the combined use of light microscopy, scanning electron microscopy and pigment analysis.
Al1 the mats studied were made up predominantly of cyanobacteria, with chlorophyll a as the main photosynthetic pigment. Coccoid cyanobacteria were the main organisms in mats with higher salinities. When salinity levels decreased, filamentous cyanobacteria became the principal mat-building organisms. In young mats, Spirulina was dominant, while in well developed mats, where Microcoleus was dominant, a purple layer with bacteriochlorophyll a was also present.
INTRODUCTION
It was more than a century and a half ago that stroma- tolites were first described (STEEL, 1825). These structu- res, defined as "organosedimentary structures produced by sediment trapping, binding, and/or precipitation as a result of the growth and metabolic activity of microorganisms, primarily by cyanobacteria" (MARGULIS et a l . , 1986), are fundamental for a better understanding of microbial mats.
Microbial mats have been described as stratified commu- nities of microorganisms that develop in the physico- chemical gradients established at the interfaces between water and solid substrates. These structures are composed prirnarily of phototrophic bacteria together with diverse microorganisms. They are frequently laminated due to variations of different parameters such as light, tempera- ture, salinity, etc. These laminations have different colors as a result of growth of phototrophs with different pigment composition, and thus, different patterns of spectral ut i l i -
zation of the available light. Nevertheless, non-laminated microbial mats can be found, depending on environmental fluctuations.
In the last few years, two books (COHEN et al . , 1984; COHEN & ROSENBERG, 1989), and many articles have been published which extensively revise the structure, physiology, genetics and evolution of microbial mats
(table 1). Today, microbial mats are confined to a restricted range
of habitats including hypersaline and coastal marine envi- ronments (BAULD, 1984; VAN GEMERDEN et al . , 1989), hot springs (CASTENHOLZ, 1984; JORGENSEN & NELSON, 1988) and alkaline lakes (BROCK, 1978). Micro- bial mats on deep-sea hydrothermal vents have also been studied. The emphasis on this particular community is of their chemosynthetic function (JANNASH, 1985; NELSON et al . 1989).
Marine microbial mats are inundated periodically because they are separated from the open sea by a shallow barrier: dunes (Le. Baja California), salinas (Santa Pola, San Rafael, etc.), even the vegetation of mangrove trees [Matanzas, Cuba (R. Guerrero, unpublished; MARGULIS ef al., 1986)l.
Some microbial mats can be placed in strange places, like those found in the slopes of the caldera in the Kilauea crater (Hawaii). They are small irregulary-shaped patches, not more
Limnetica, 8 : 185-195 (1992)
O Asociación Española de Limnología, Madrid. Spaiii

Table 1. Some important microbial mats of the world
Location Kind Reference
Yellowstone (Wyoming. USA) Hot spring CASTENHOLZ, 1984.
New Zealand
Iceland
Hot spring
Hot spring
CASTENHOLZ, 1976.
SCHWABE, 1960.
Guaymas Basin (East Pacific) Deep-sea hydrothermal vents. BELKIN & JANNASCH, 1989.
Galapagos Rift (Ecuador) Deep-sea hydrothermal vents. JANNASCH, 1985.
Solar Lake (Sinai) Marine KRUMBEIN et a l , 1977.
Gulf of Aqaba (Sinai) Marine KRUMBEIN & COHEN, 1974.
Shark Bay and
Spencer Gulf (Australia) Marine
Laguna Figueroa
(Baja California, México) Marine
Guerrero Negro
(Baja California, México) Marine
Sippewisset
(Woods Hole, Mass., USA) Marine
Hamei Mazor
(Dead Sea, Israel)
Meijendel (dune area)
(The Netherlands)
Mellum Islands
(North Sea, Germany)
BAULD, 1984.
HORODYSKI, ct al. 1977.
JAVOR & CASTENHOLZ, 1984
GIBSON et al., 1984.
Hypersaline sulfur spring OREN, 1989.
Marine WINDER et al., 1989.
Marine STAL et al., 1985.
Orkney Islands (UK) Sheltered beaches HERBERT, 1985.
Shackleford Banks (USA) Marine
Ebro Delta (Spain) " Marine
Canary Islands (Spain) Marine
PAERL et al., 1989.
MIR et u/., 1991.
MENDEZ & VINUESA
(personal communication).
Soda Lake (Nevada, USA) Hypersaline lake OREMLAND & DES MARAIS, 1983.
Hawaii (USA) Marine GUERRERO
(personal communication)
Cuba (Salina de Bido. Matanzas) Marine (protected by mangroves) MARGULIS et al., 1986.
Saltern of "Saline de Giraud"
(Camarge, France) Hypersaline pond CAUMETTE et al., 1988.

than 10 cin in diameter. They are scattered on the walls of
the crater, some times tilted more than 701> and presenting a different structure of those usually found in coastal zones.
They are covered by a layer of black or dark-brown cyano-
bacteria (M~r.sticqnc~la~l~rs Innzitlosus is dominani), clearly defi- ned purple and green layers (not more than 1 mm thick each
one), and a basal layer of a yellowish, translucent gelatin, prohably constituted by polysaccharides. Thus, the black
basal layer of anaerobic sulfate-reducers, which is almost universal in other microbial mats, is here absent. Al1 sulfide and water is coming from the vapors steaming interinittently from fissures in the volcano walls, and therefore the cycling of sulfur compounds is not complete, reminding us of the
earliest pre-ecosystems (~inpublished data). The development of microelectrodes to measure chemical
gradients (REVSBECH & JORGENSEN. 1985), micro-optic fibers for the study of spectral light distribution (PIERSON et al., 1987; JORGENSEN & DES MARAIS, 1988). and
electron microscopy techniques (STAL et ul . . 1985; CAUMETTE, 1984; D'AMELIO et al . 1987), to better
observe the structure and composition of these communities
and the intra- and interspecific relationship have increased in the last few years our knowledge ofmicrobial mats.
From an evolutionary point of view, microbial mats are of great interest because they are considered modern homo-
logs of some precambrian stromatolites. Also, microbial mats found in modern hot springs provide examples of precambrian communities. The high temperature of hot spring waters in combination with high H2S concentrations
or acidic conditions sometimes prevent the development of eukaryotes, facilitating the study of prokaryotic life.
Chemical fossils (organic compounds whose structures, in part, have survived in the fossil record) have been the starting point for many geochemical studies on modern
inicrobial mats. The aim of these studies has been to identify the types of microorganisms present in ancient sediment by examining the types of biochemical markers (PALMISANO
t>t al.. 1988). Nevertheless. the complexity of these commu-
nities makes it difficult to determine the microbial origins of specific compounds found in the mats.
The role that mat-forming bacteria play on prokaryotic evolution (DES MARAIS, 1990), their role in laminated
(STOLZ, 1990; KAZMERCZAK & KEMPE, 1990; Mc NAMARA. 1990) and non-laminated microbial comniuni- tics (CASTENHOLZ rt al . . 1990), the description of new microorganisms (WAHLUND er al . , 1991). and the elabo- ration of microcosms and models (GIANI et a l . , 1989) are
other topics published recently.
Nevertheless, one of the questions of great interest that is not well understood is Iiow microorganisms living in micro-
bial mats can survive under such extreme conditions as high light intensity, salinity, high temperatures and desiccation.
Hyperscums, inucilages and polymer inclusions seem to
play an important role here. Hyperscums have been descri-
bed as crusted. buoyant, cyanobacterial mats often decime- ters thick. in which the organisms are so densely packed that free water is not evident. ZOHARY (1985) postulated that
the role of these dry crusts is similar to that of resting spores or cysts of other organisms, providing the iririoculum for growth when conditions become favorable.
Also, adhesion by mucilage can serves as a survival
mechanism at least for cyanobacteria. The mechanisms for
attachment that have been found to operate in this group of microorganism~, involve a change of the cell surface proper-
ties (FALTON & SHILO, 1984). Less known is the role of biopolimers in the survival of
microorganisms in microbial mats. The coinpounds that can be regarded as reserve material are usually polymers such as glycogen, poly-B-hydroxyalkanoates. cyanophycins and
phycobiliproteins. Most of them are used by microorga-
nisms as energy, carbon and nitrogen sources (JENSEN &
SICKO, 1971 ; LAWRY & SIMON, 1982; GLAZER, 198 1). The total amount of these storage compounds accuin~ilated
in the cell can be considerable . depending on the environ- mental conditions (ALLEN & HUTCHISON, 1980; MAS-
CASTELLA, 199 1). During the last fifteen years, our group has been studying
freshwater stratified lakes, where phototrophic prokaryotes
often form multilayered planktonic microbial communities (MPM) (GUERRERO et crl., 1987; PEDRÓS-ALIÓ et u / . . 1983). Microbial mats and MPM have been compared to
ecosystems which depend on light as the primary energy
source (GUERRERO & MAS, 1989). Phototrophic orga- nisms place themselves at different positions in these struc-
tures according to the vertical distribution of the environ- mental factors.
Also, we have studied the accumulation of poly-B-hydrox- yalkanoates inclusions in Chromatiaceae in natural habitats
(ESTEVE al., 1990) and the characterization of predatory bacteria (GUERRERO eral. . 1986) and its dynamics contro- lling populations of Chromatiaceae in different karstic lakes (ESTEVE ' t u l . , 1992, and GAJU et al., i.n press).
Since 1987, we have been studying multilayered benihic microbial communities in the Ebro Delta and microbial mats froin different locations in Spain (GUERRERO & DE WIT,
1992).
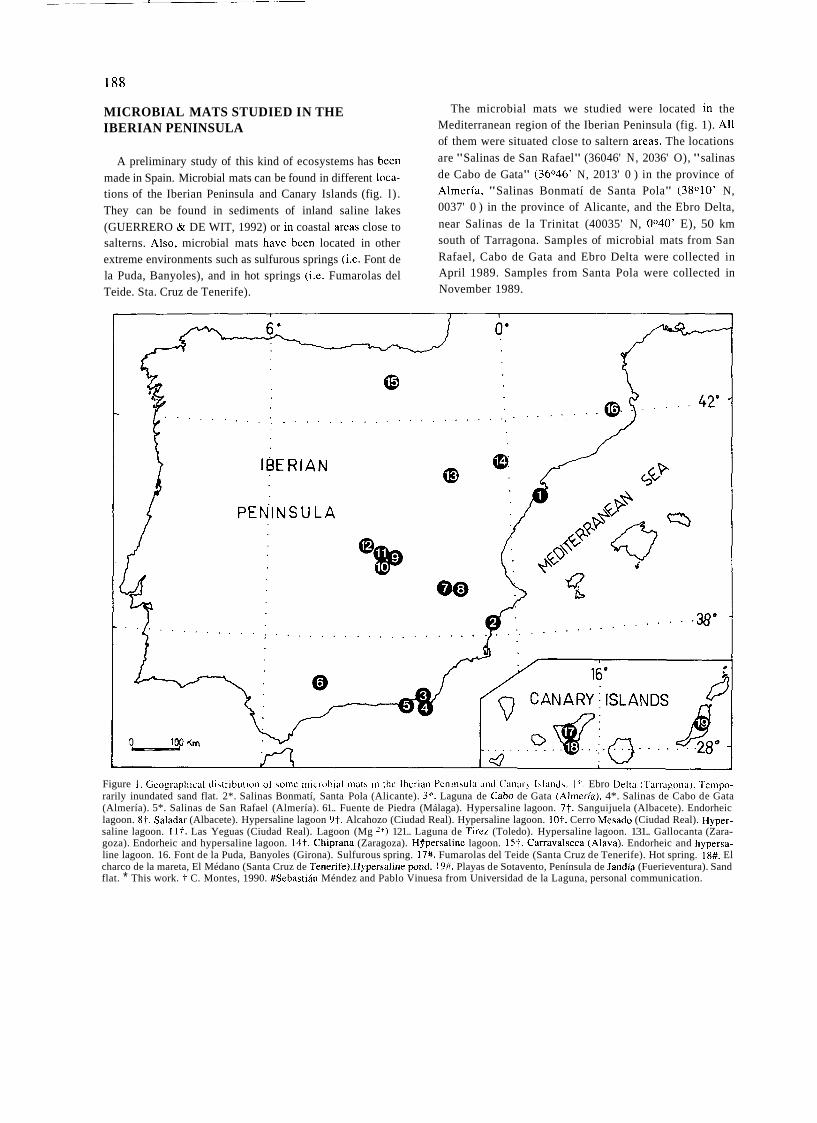
MICROBIAL MATS STUDIED IN THE IBERIAN PENINSULA
A preliminary study of this kind of ecosystems has been made in Spain. Microbial mats can be found in different loca- tions of the Iberian Peninsula and Canary Islands (fig. l) . They can be found in sediments of inland saline lakes (GUERRERO & DE WIT, 1992) or in coastal areas close to salterns. Also, microbial mats have been located in other extreme environments such as sulfurous springs (Le. Font de la Puda, Banyoles), and in hot springs (i.e. Fumarolas del Teide. Sta. Cruz de Tenerife).
The microbial mats we studied were located in the Mediterranean region of the Iberian Peninsula (fig. 1). All of them were situated close to saltern areas. The locations are "Salinas de San Rafael" (36046' N, 2036' O), "salinas de Cabo de Gata" (36O46' N, 2013' 0 ) in the province of Almería, "Salinas Bonmatí de Santa Pola" (38O10' N, 0037' 0 ) in the province of Alicante, and the Ebro Delta, near Salinas de la Trinitat (40035' N, 0°40' E), 50 km south of Tarragona. Samples of microbial mats from San Rafael, Cabo de Gata and Ebro Delta were collected in April 1989. Samples from Santa Pola were collected in November 1989.
Figure l . Geogi-aphical disiributioii of sonir iiiici.obinl iiials i i i tlie Iberiaii Peiiiiisula ;iiiil C'uiini.) Islniid\. 1 - . Ebro Delrii (l'iiri-agona). 'I'enipo- rarily inundated sand flat. 2*. Salinas Bonmatí, Santa Pola (Alicante). 3*. Laguna de Cabo de Gata (Almería). 4*. Salinas de Cabo de Gata (Almería). 5*. Salinas de San Rafael (Almería). 61.. Fuente de Piedra (Málaga). Hypersaline lagoon. 7 t . Sanguijuela (Albacete). Endorheic lagoon. 8 t . Saladar (Albacete). Hypersaline lagoon 9 t . Alcahozo (Ciudad Real). Hypersaline lagoon. lo f . Cerro Mesado (Ciudad Real). Hyper- saline lagoon. 117. Las Yeguas (Ciudad Real). Lagoon (Mg 2+) 121.. Laguna de Tirez (Toledo). Hypersaline lagoon. 131.. Gallocanta (Zara- goza). Endorheic and hypersaline lagoon. 14t . Chiprana (Zaragoza). Hgpersaline lagoon. 157. Carravalseca (Alava). Endorheic and hypersa- line lagoon. 16. Font de la Puda, Banyoles (Girona). Sulfurous spring. 17#. Fumarolas del Teide (Santa Cruz de Tenerife). Hot spring. 18#. El charco de la mareta, El Médano (Santa Cruz de Tenerife).Hypersaline pond. 19#. Playas de Sotavento, Península de Jandía (Fuerieventura). Sand flat. * This work. 1. C. Montes, 1990. #Sebastián Méndez and Pablo Vinuesa from Universidad de la Laguna, personal communication.

Figui-e 2. View of differeiit ,atiipling site\ \tudied (A-E) aiid croab-acctioiis ot niici-obial iiiats observed (E-J) . A ) and F) Salinas de San Rafael, Alrnería; B) and G) Salinas de Cabo de Gata, Almería: C ) aiid H) Laguna de Cabo de Gata, Almería; D) and 1) Salinas Bonrnatí, Santa Pola, Alicante; E) and J) Sand flats of Ebro Delta, Tarragona.
Temperature, conductivity, and salinity of the water cove-
ring the microbial mats were measured in siru using a Yellow Springs Instrument S-C-T meter model 33. The pH was measured with a micro-pH 2001 Crison pH-meter.
Vertical stratification of the mats was studied by exami-
ning 1 mm thick vertical slices from top to bottom by phase
contrast microscopy and scanning electron microscopy. Horizontal cross-sections of the mats were used to charac-
terize more accurately the different individual layers. For
pigment determination, the different colored layers were separated manually and extracted with methanol.
Sa1itzu.s de Suri Rufael (Almería) Two types of microbial mats were found in salinas San
Rafael, on the shore of the ponds (fig. 2A). The water of the
pond was brownish-red, presumably due to Halobacteria-
ceae. A~.temia salinu was also present. One of these micro- bial mats had developed on white sediment (fig. 2F). Three distinct colored layers could be distinguished. The upper-
most layer was brownish-white in color, with gypsum preci-
pitates on its surface. Diatoms were dominant in this layer. The second layer was green and coccoid cyanobacteria and some unidentified filamentous cyanobacteria about 3 y m in diameter were observed. The third layer was dimly pink, and
below these pigmented layers the sediment was white. The second type of microbial mat found in salinas de
San Rafael had two colored laminations. The upper layer was brown, and diatoms were the main phototrophic orga- nisms present. The second layer was green, and coccoid cyanobacteria and some filamentous cyanobacteria could

inain photosynthetic pigment in al1 colored layers, and little
amounts of bacteriochlorophyll u could be detccted (fig. 5).
Crrho c/c Gura (Almería)
T h e structure of rnicrobial mats from "Salinas del C a b o
nity, in conjunction with high levels of evaporation, has lead
Figure 3. I'lio~ofi:iphic pinte \lio\\iiif diltererii oi-giiii~\rii\ o1 ilic
microbial mats studied. A ) Gjro.ti,qmu \p.. a diatom bclongiiig to the family Naviculaceae: B ) Amphoru c~,qgrc',yiu., a diatom belonging to the family Cymbellaceae: C) L j r ~ ~ h j u oestucrr.ii filamentous cyano- bacteria with pigmented sheath; D) Mic~oc,olerts (~liror~oplusre.s. a sheathed filamentous cyanobacteria; E) 0.sc~illutor-iu sp; F) Spir.ulirrcr sp; G) Clrr.ooc,oí.c.u.s sp. a coccoid cyanobacteria belonging to the Gloco<.upsu group: H) Chromariirnl sp.. a purple sulfur bacteria.
be observed . Below this l ayer the sed iment was black in
color .
T h e absorption spectra of methanolic extracts of the diffe- I.-IELIIC 4. S C ; I I I I I I I I ~ C ~ C C I I . O I I i i i ~ ~ ~ o ~ i ~ ; i l ~ l l \lio\t 1112 , A ) .\11, I O , OI<>II\ rent colored laminations showed that chlorophyll a was the c ~ l i r o ~ i o / ~ l u . ~ ~ ~ ~ \ and B ) S/~ii.rrliiiir \p.
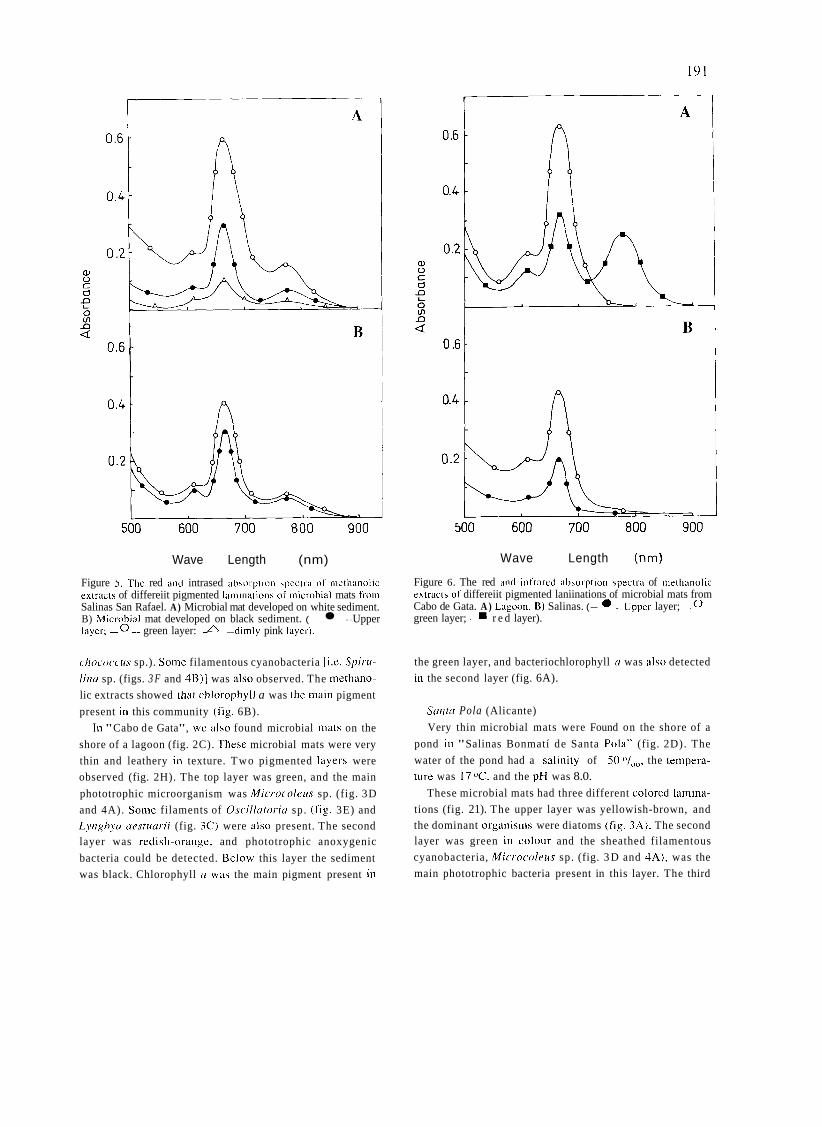
Wave Length (nm)
Figure 5 . Tlie red niid intrased absorptioii spectra 01' iiietlianolic extracts of differeiit pigmented laniinstions of inicsobial mats fro~n Salinas San Rafael. A) Microbial mat developed on white sediment. B) Microbial mat developed on black sediment. ( - - Upper layei: - 0 - green layer: -A d i m l y pink layes).
c,hococcrrs sp.). Some filamentous cyanobacteria 1i.e. Sl1i1.u-
l ino sp. (figs. 3 F and 4B)j was also observed. The methano-
lic extracts showed Ihat chlorophyll a was Ihe main pigment
present in this community (fig. 6B).
In "Cabo de Gata", wc also found microbial rnats on the
shore of a lagoon (fig. 2C). These microbial mats were very
thin and leathery in texture. Two pigmented Iayers were
observed (fig. 2H). The top layer was green, and the main
phototrophic microorganism was Mir,r.o<,olrus sp. (fig. 3D
and 4A). Some filaments of 0.scillatoriu sp. (fig. 3E) and L y t ~ g h y u C I C ' S ~ U U ~ ~ ~ (fig. 3C) were also present. The second layer was redish-orange, and phototrophic anoxygenic
bacteria could be detected. Below this layer the sediment
was black. Chlorophyll cr was the main pigment present ir1
Wave Length (nm)
Figure 6. The red aiid iiifrared ahaorption spectra of iiiethanolic extracts of differeiit pigmented laniinations of microbial mats from Cabo de Gata. A ) Lagoon. B) Salinas. (- a - -Upper layer; -0- green layer; -m r e d layer).
the green layer, and bacteriochlorophyll u was also detected
in the second layer (fig. 6A).
S n n f u Pola (Alicante)
Very thin microbial mats were Found on the shore of a
pond in "Salinas Bonmatí de Santa Pola" (fig. 2D). The
water of the pond had a salinity of 50 <)/,,, the tempera-
ture was 17OC, and the pH was 8.0. These microbial mats had three different colored lamina-
tions (fig. 21). The upper layer was yellowish-brown, and the dominant organisnis were diatoms (fig. 3A). The second layer was green in colour and the sheathed filamentous
cyanobacteria, Mic.roc.01eu.s sp. (fig. 3 D and 4A) , was the
main phototrophic bacteria present in this layer. The third
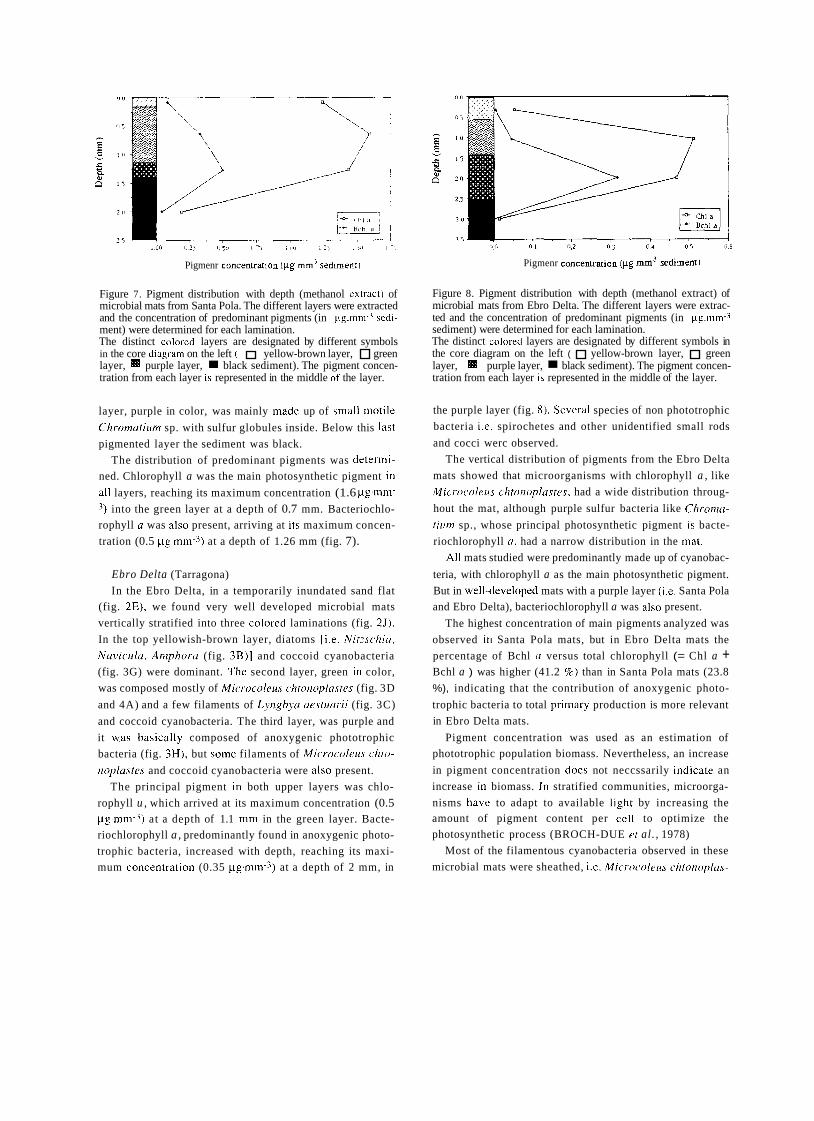
Pigmenr concenuarion !)lg mm3 sedimení)
Figure 7. Pigment distribution with depth (methanol extract) of microbial mats from Santa Pola. The different layers were extracted and the concentration of predominant pigments (in yg.mm-3 sedi- ment) were determined for each lamination. The distinct colored layers are designated by different symbols in the core diagram on the left ( yellow-brown layer, green layer, ES purple layer, black sediment). The pigment concen- tration from each layer is represented in the middle of the layer.
layer, purple in color, was mainly rnade up of small motile
Chrnrnatil~m sp. with sulfur globules inside. Below this last
pigmented layer the sediment was black.
The distribution of predominant pigments was determi- ned. Chlorophyll a was the main photosynthetic pigment in
al1 layers, reaching its maximum concentration (1.6 p g m m -
') into the green layer at a depth of 0.7 mm. Bacteriochlo- rophyll u was also present, arriving at its maximum concen-
tration (0.5 p g m m - 3 at a depth of 1.26 mm (fig. 7).
Ebro Delta (Tarragona)
In the Ebro Delta, in a temporarily inundated sand flat (fig. 2E), we found very well developed microbial mats
vertically stratified into three colored laminations (fig. 25).
In the top yellowish-brown layer, diatoms [¡.e. Nitrsckicc,
Navic~l lu . Amphor-u (fig. 3B)] and coccoid cyanobacteria (fig. 3G) were dominant. Tlie second layer, green in color,
was composed mostly of Microcoleus chtoriop1a.sre.s (fig. 3D
and 4A) and a few filaments of Lyngbya ur.stuar.ii (fig. 3C)
and coccoid cyanobacteria. The third layer, was purple and
it was basically composed of anoxygenic phototrophic
bacteria (fig. 3H), but some filaments of Micioco1eii.s c,hto-
noplastrs and coccoid cyanobacteria were also present.
The principal pigment in both upper layers was chlo-
rophyll u , which arrived at its maximum concentration (0.5 pg.mm-') at a depth of 1.1 mm in the green layer. Bacte- riochlorophyll a , predominantly found in anoxygenic photo-
trophic bacteria, increased with depth, reaching its maxi- mum concentration (0.35 pg.mm-') at a depth of 2 mm, in
Pigmenr concentrauon (pg mmhediment )
Figure 8. Pigment distribution with depth (methanol extract) of microbial mats from Ebro Delta. The different layers were extrac- ted and the concentration of predominant pigments (in yg.mm-' sediment) were determined for each lamination. The distinct colored layers are designated by different symbols in the core diagram on the left ( yellow-brown layer, green layer, purple layer, black sediment). The pigment concen- tration from each layer is represented in the middle of the layer.
the purple layer (fig. 8). Several species of non phototrophic
bacteria ¡.e. spirochetes and other unidentified small rods
and cocci werc observed.
The vertical distribution of pigments from the Ebro Delta
mats showed that microorganisms with chlorophyll a , like
Mic,i.ocoleus c,htonopla.stes, had a wide distribution throug-
hout the mat, although purple sulfur bacteria like Chroma-
tium sp., whose principal photosynthetic pigment is bacte-
riochlorophyll cr, had a narrow distribution in the mar.
Al1 mats studied were predominantly made up of cyanobac-
teria, with chlorophyll a as the main photosynthetic pigment.
But in well-developed mats with a purple layer (¡.e. Santa Pola and Ebro Delta), bacteriochlorophyll a was also present.
The highest concentration of main pigments analyzed was
observed in Santa Pola mats, but in Ebro Delta mats the
percentage of Bchl u versus total chlorophyll (= Chl a + Bchl a ) was higher (41.2 %) than in Santa Pola mats (23.8
%), indicating that the contribution of anoxygenic photo-
trophic bacteria to total primary production is more relevant
in Ebro Delta mats.
Pigment concentration was used as an estimation of phototrophic population biomass. Nevertheless, an increase
in pigment concentration does not neccssarily indicate an
increase in biomass. In stratified communities, microorga-
nisms have to adapt to available light by increasing the amount of pigment content per cell to optimize the photosynthetic process (BROCH-DUE et al . , 1978)
Most of the filamentous cyanobacteria observed in these
microbial mats were sheathed, ¡.e. Microcole~ls c,l~tonopla.s-

tes and Lyngbya aestuarii. The sheaths play protective roles against dangers in the environment. An example of this function is the shielding ability of scytonemin, a yellow- brown pigment located in the sheath of several cyanobacte- ria such as Lyngbya sp., that protects them from high levels of solar radiation (GARCIA-PICHEL & CASTENHOLZ, 1991). Lyngbya aestuarii located on the surface of the mats studied had more brownish coloration than the same orga- nisms situated below.
Cyanobacterial mats which grow undisturbed for long periods may develop a regular lamination as the result of periodic changes in response to seasonal varia- tions of the environment. Species composition, density of photosynthetic organisms, and levels of excretion of polysaccharides by bacteria change seasonally in response to changes in salinity, water cover, Iight inten- sity and temperature.
In hypersaline environments, the differences in composi- tion of microorganisms depends on salinity levels. Coccoid cyanobacteria are the main microorganisms in environments where salinity is high, for example in Cabo de Gata salinas. Moreover, the diversity increases when salinity levels decre- ase. In this case, filamentous cyanobacteria, mainly Micro- coleus sp., are the principal mat-building organisms, although coccoid cyanobacteria are also present.
This kind of microbial mats (Santa Pola and Ebro Delta mats) had three well distinguished colored laminations, and the dominant microorganisms were similar. Diatoms were present only in well established mats, building a new layer on the surface of these stratified communities.
In these photosynthetic communities, changes in struc- ture can be observed not only seasonally but also in terms of location. For example, in Ebro Delta mats different mat development states were found in only a few meters of distance. Filamentous cyanobacteria such as Spirulina sp. were dominant in young mats; however, Microcoleus sp. was the main phototrophic microorganism in the green layer of well-developed microbial mats (MIR et al., 1991).
Although extensive work has been done on the subject of these kinds of communities, there is still little known about productivity, the role of polymer inclusions in oxygenic and anoxygenic phototrophic bacteria, and the inter-and intras- pecific relationships among bacteria living on laminated structures,
The nekt studies in the Ebto Delta will provide fiore information about these intetesting ecosystems developed in extreme erivironments.
ACKNOWLEDGEMENTS
We thank the Ebro Delta Natural Park Service (Genera- litat de Catalunya) and Salines de la Trinitat for permis- sion to collect samples. We gratefully acknowledge Dr. J . Mas-Gordi for critically reading the manuscript, J. Checa for his technical assistance and J. Bloomgarden for helping us in the English version. This work was supported by grants CICYT PB87-0500 to 1. E. and BI089-0591 and MAR91-0874 to R. G. We also thank the Autonomous University of Barcelona for a research grant to M. M-A. J . M. research was supported by grant PP88-39158752 from the Spanish Ministry of Education and Science ascribed to grant PB87-083 to C. Pedrós-Alió.
REFERENCES
ALLEN, M. M., & F. HUTCHISON, 1980. Nitrogen limitation and recovery in the cyanobacterium Aphanocapsa 6308. Arch. Microbiol. 128: 1-7.
BAULD, J . , 1984. Microbial mats in marginal marine environments : Shark bay, Western Australia, and Spencer Gulf, South Australia. In: Cohen, Y., Castenholz, R. W Halvorson, H. O (Eds.), Microbial Mats Stromafolires: 39- 58. Alan R. Liss, New York.
BELKIN, S., & H. W. JANNASCH, 1989. Microbial mats at deep-sea hydrothermal vents: New observations In: Cohen, Y., & E. Rosenberg (Eds.), Microbial mats. Physiological ecology of benthic microbial communities: 16-21. American Society for Microbiology Washing- ton, D.C.
BROCH-DUE, M., J. ORMEROD & F. STRAND, 1978. Effect of light intensity on vesicles formation in Chloro- bium. Arch. Microbiol. 116: 269-274.
BROCK, T. D., 1978. The habitats. Thermophilic micro- organisms and life at high tempet-atures, Springer-Verlag. New York.
CASTENHOLZ, R. W., J. BAULD & B. B. JORGENSEN, 1990. Anoxygenic microbial mats of hot springs: thermophi- lic Chlorobium sp. FEMS Microbiol. Ecol. 74: 325-336.
CASTENHOLZ, R. W., 1976. The effect of sulfide on the blue green algae of hot springs. 1. New Zealand and Iceland. J. Phycol. 12: 57-68.
CASTENHOLZ, R. W.,1984. Composition of hot spring microbial mats: a summary. In: Cohen, Y., Castenholz, R. W and Halvorson, H. O (Eds.). Microbial Mats Stromatoli- tes: 101-1 19. Alan R. Liss, New York.

CAUMETTE, P., 1984. Distribution and characterization of phototrophic bacteria isolated from the water of Bietri
Bay (Ivory Coast). Cuw. .J. Mic.rohio1. 30: 273-284. CAUMETTE, P., R. BAULAIGUE & R. MATHERON,
1988. Characterization of Chromatii<wr .salrsigen.s sp. nov., a halophilic Chromatiaceae isolated from Mediterranean sali-
nas. Systern. Appl. Microhiol. 10: 284-292. COHEN, Y., R. W. CASTENHOLZ & H. O. HALVOR-
SON (Eds), 1984. Mic,rohial rnuts: Strornutolitrs. Alan R. Liss, Inc New York.
COHEN, Y. & E. ROSENBERG (Eds), 1989. Microhirrl
mcrts. Physiologic.c~l ec,ology of hrnthic nzic.rohin1 cornni~ini- ties. American Society for Microbiology, Washington, D.C.
D'AMELIO, E. D., Y. COHEN & D. J. DES MARAIS, 1987. Association of a new type of gliding, filamentous,
purple phototrophic bacterium inside bundles of Mic.1.oc.o- 1erl.s chthonopla.stes in hypersaline cyanobacterial mats. Ai.c.h. Mic,rohiol. 147: 2 13-220.
DES MARAIS, D.,1990. Microbial mats and the early evolution of life. TREE 5: 140-144.
ESTEVE, 1.. E. MONTESINOS, J . G. MITCHELL& R. GUERRERO, 1990. A quantitative ultrastructural study of Chvornutirlm rninus in the bacterial layer of lake Cisó (Spain). A1.c.h. Mic.r.ohio1. 153: 422-427.
ESTEVE, l., N. GAJU, J . MIR & R. GUERRERO, 1992. Comparison techniques to determine the abundance of predatory bacteria attacking Chromatiaceae. FEMS Micra- b i d . Ecol. 86: 205-2 1 1.
FALTON, A. & M. SHILO, 1984. Hydrophobicity as an adhesion mechanism of benthic cyanobacteria. Appl. Envi- ron. Microhiol. 47: 135- 143.
GAJU, N., 1. ESTEVE & R.GUERRERO. Distribution of predatory bacteria attacking Chromatiaceae in a sulfurous lake. Micvoh. Ecol. 24: (in press).
GARCIA-PICHEL, F.. R.W. CASTENHOLZ, 1991.
Characterization and biological implications of scytonemin, a cyanobacterial sheath pigment. .l. Phycol. 27: 395-409.
GIANI, D., J. SEELER, L. GIANI & W. E. KRUMBEIN. 1989. Microbial mats and physicochemistry in a saltern in
the Bretagne (France) and in a laboratory scale saltern model. FEMS Mici.ohio1. Ecol. 62: 151 -162.
GIBSON, J., E. R. LEADBETTER & H. B. JANNASCH, 1984. Great Sippewisset marsh: A summary of projects
carried out by students in the microbial ecology course of the marine biological laboratory, Woods Hole, during summers 1972-198 1. 111: Cohen, Y., Castenholz, R. W and Halvorson, H. O., (Eds.), Mici.ohia1 Muts Sfromnto1ite.s: 95- 100. Alan R. Liss, New York.
GLAZER, A. N, , 198 l . Photosynthetic accessory proteins with bilin prosthetic groups: 51-96. In: M.D. Hatch and
N.K. Boardman (Ed), Biochemistry of Plants, vol VI11 :
Photosynthesis. Academic Press, New York. GUERRERO, M. C. & DE WIT. R. 1092. Microbial mats
in the inland saline lakes of Spain. LinincJtic.rr 8. GUERRERO, R., C. PEDROS-ALIO. l . ESTEVE, J .
MAS, D. CHASE & L. MARGULIS, 1986. Predatory prokaryotes: Predation and primary consumption evolved in bacteria. Proc.. Nutl. Accrd. Sci. 83: 2 138-2 142.
GUERRERO, R., C. PEDROS-ALIO, 1. ESTEVE & J .
MAS. 1987. Cornmunities of phototrophic sulfur bacteria in lakes of the Spanish Mediterranean region. Acta Ac,ud. Aho. 47: 125-151.
GUERRERO, R. & J. MAS, 1989. Multilayered microbial
communities in aqu;itic ecosystems: growth and loss factors. In: Cohen, Y. & E. Rosenberg (Eds.), Mi(.rohiuI rnuts. Physiologic,crl ec.olog o$herithic. Mic.rohiu1 c.on7munitie.s: 37-
5 1. American Socicty for Microbiology Washington. HERBERT, R. A. 1985. Development of mass blooms of
photosynthetic bacteria on sheltered beaches in Scapa Flow, Orkney Islands, Proc.. Koy. Soc.. Edinhurglr 87: 15-25.
HORODYSKI, R. J., B. BLOESSER & S. J. VON DER
HAAR, 1977. Laminated algal mats from a coastal lagoon, Laguna Mormona, Baja California, Mexico. J. Sedimrnt. Prt., 47: 680-696.
JANNASH, H. W., 1985. The chemosynthetic support of life and the microbial diversity at deep sea hydrothermal verits. P~.oc.. R. Soc. Lond. Ser. B.: 225: 227-297.
JAVOR, B. J. & R. W. CASTENHOLZ, 1984. Producti- vity studies of microbial mats, Laguna Guerrero Negro, México, pp. 149-170. In: Y. Cohen, R.W. Castenholz &
H.O. Halvorson (Ed), Mic,r.ohiul Muts: Stron~utolites. Alan R. Liss, New York.
JENSEN, T. E. & L. M. SICKO, 197 1. Fine structure of poly-8-hydroxybutyrate acid granules in a blue-green alga,
Chloro,gloecr frit.sc.hii. J. Bact~riol . 106: 683-686. JORGENSEN, B. B. & D. J. DES MARAIS, 1988. Optical
properties of benthic photosynthetic communities: fiber-optic
studies of cyanobacterial mats. Lirnnol. Oceanogr. 33: 99-1 13. JORGENSEN, B. B. & D. C. NELSON, 1988. Bacteria1
zonation, photosynthesis, and spectral light distribution in hot spring microbial mats of Iceland. Microh. Ecol. 16: 133- 147.
KAZMERCZAK, J. & S. KEMPE, 1990. Modern cyano- bacterial analogs of Paleozoic stromatoporoids. Science 250: 1244- 1250.
KRUMBEIN. W. E. & Y. COHEN, 1974. Klastische und evaporitischc sedimentation in einem mesothermen mono-

iniktischeii ufernahen See (Golf von Aqaba, Sinai). Geol.
K~rndsc~li. 63: 1035- 1065. KRUMBEIN, W. E., Y. COHEN & M. SHILO, 1977.
Solar lake (Sinai). 4. Stroinatolitic cyanobacterial mats. Limviol. Oceariogr.. 22: 635-655.
LAWRY, N . H. & R. D. SIMON, 1982. The normal and induced occurrence of cyanophycin inclusion bodies in several blue-green algae. J. Phyc.01. 18: 391-399.
MARGULIS, L., L. LOPEZ-BALUJA, S. M. AWRA- MIK & D. SAGAN, 1986. Community living long before man: Fossil and living microbial mats and early life. 7'h(~ Scienc.~~ r?f rhe Total Erii1ironn7c'tif 56: 379-397.
MAS-CASTELLA, J . 199 1. Acitn~ulacicín de poli$-
hiclrnria1r~a)::iouto.s por bacterias. Distribucihn en /u riatiii.rr-
/era y hiote<.nolo~ía. Ph. D. Thesis. University of Barcelona. Mc NAMARA, K., 1990. Survivors from the primordial
soup. New Scientist 8: 50-52. MIR, J., M. MARTINEZ-ALONSO, 1. ESTEVE & R.
GUERRERO, 1991. Vertical stratification and microbial assemblage of a microbial mat in the Ebro delta (Spain). FEMS Mici.obio1. Ecol. 86: 59-68.
MONTES, C. (coord.) 1990. Estrtdio de las zonas Iiúwie-
das de la Espancr peni)::~.sula/.. Irzr~~tzturio y Tipologiu. Direc- ción General de Obras Hidráulicas, Ministerio de Obras Públicas y Urbanismo. Madrid.
NELSON, D. C., C. O. WIRSEN & H. W. JANNASCH, 1989. Characterization of large, autotrophic Beggiutou spp. abundant at hydrothermal vents of the Guayamas Basin. Appl. Etivirorr. Mic.r.ohiol. 55: 2909-29 17.
OREMLAND, R.S., & D.J. DES MARAIS, 1983. Distri- bution, abundance and carbon isotopic composition of gase- ous hydrocarbons in Big Soda Lake, Nevada: an alkaline, meromictic lake. Geoc,him. Cosrnochim. Acta 47: 2 107-2 1 14.
OREN, A., 1989. Photosynthetic and heterotrophic bent- hic bacterial communities of a hypersaline sulfur spring on the shore of the Dead Sea (Hamei Mazor). In: Cohen, Y., Castenholz, R. W & Halvorson, H. O. (Eds.), Mic.rohia1 Mats Str-omatolites: 64-76. Alan R. Liss, New York.
PAERL, H. W., B. M. BEBOUT & L. E. PRUFERT, 1989. Naturally ocurring patterns of oxygenic photosynthe- sis and N2 fixation in a marine microbial mat: physiological and ecological ramifications. In: Cohen, Y., and E. Rosen- berg (Eds.), Mic,robiul mats. Physiologicul ecology ofbeni- hic Mic.rohia1 commurrities: 326-341. American Society for Microbiology Washington, D.C
PALMISANO, A. C., S. E. CRONIN & D. J . DES MARAIS, 1988. Analysis of lipophilic pigments from a phototrophic microbial mat community by high perfor- mance liquid chromatography. .l. Mic.robio1 Methods 8 :
209-2 17. PEDROS-ALIO, C.. E. MONTESINOS & R. GUERRE-
RO. 1983. Factors determining annual changes in bacterial photosynthetic pigments in holomictic lake Cisó, Spain. AppI. Environ. Mic.robio1. 46: 999- 1006.
PIERSON, B., A. OESTERLE & G. L. MURPHY, 1987. Pigments, light penetration, and photosynthetic activity in the multi-layered microbial mats of Great Sippewissett Salt Marsh, Massachusetts. FEMS Mic.rohio1. Ecol. 45: 365-376.
REVSBECH, N. P., & B. B. JORGENSEN, 1985. Micro- clectrodes: their use in microbial ecology. Adv. Mic.roh.
Ecol. 9 : 293-352. SCHWABE, G. H., 1960. Uber den thermobioten
Kosmopoliten Mcrsti,yocluclus laniinosus Cohn. Blaugen und Lebenstraum. V. Schweeig. .l. Hydr.01. 22: 757-792.
STAL, L. J., H. VAN GEMERDEN & W. E. KRUM- BEIN, 1985. Structure and development of a benthic niarine microbial mat. FEMS Mic.rohio1. Ecol. 31: 1 1 1 - 125.
STEEL, J. H., 1825. A description of the oolitic forma-
tion lately discovered in the county of Saratoga, and state of N m , Y o I . ~ . Anl. .l. Sci. (ser. 1 j , 9: 16-19.
STOLZ, J . F., 1990. Distribution of phototrophic micro- bes in the flat lainiriated microbial mat at Laguna Figueroa, Baja California. Mexico. Bio.systrnis 23: 345-357.
VAN GEMERDEN, H., C. S. TUGHAN, R. DE WIT & R. A. HERBERT, 1989. Laminated microbial ecosystems on
sheltered beaches in Scapa Flow, Orkney Islands. FEMS Microbiol. Ecol. 62: 87- 102.
WAHLUND, T. M., C. R. WOESE, R. W. CASTEN- HOLZ & M. T. MADIGAN, 1991. A thermophilic green sulfur bacterium from New Zealand hot springs, Chlorobium tepidurn sp. nov. Arch. Microbiol. 156(2 j: 8 1-90.
WINDER, B., J. PLUIS, L. REUS & L. R. MUR, 1989. Characterization of a cyanobacterial algal crust in the coas- tal dunes oP the Netherlands. In: Cohen, Y., and E. Rosen- berg (Eds.), Microbial mata. Physiological ecology of bent- hic Microhicd con7mriniti~s: 77-83. American Society for Microbiology, Washington, D.C.
ZOHARY, T., 1985. Hyperscums of the cyanobacterium Myc.rocyxris aervginosa in a hypertrophic lake (Hartbeespo- ort Dam, South Africa ). .l. Plarikton Rcs. 7: 399-409.

MICROBIAL MATS IN T H E INLAND SALINE LAKES OF SPAIN
M. C. Guerrero' and R. de Wit2 1. Lab. Limnología. Depto. de Ecología. Universidad Autónoma de Madrid. 28049 Madrid. Spain. 2. Dept. of Environmental Chemistry. Centro de Investigación y Desarrollo (C.S.I.C.). Jordi Girona, 18-26; 08034 Barcelona.
Spain.
Keywords: Spain, Endorheic saline lakes, Alkaline lakes, Microbial mats, Annual cycles, Mic~rocoleus chthonopla.stes.
ABSTRACT
The Iberian Peninsula possesses a particular richness of inland saline lakes which mainly occur in three endorheic regions in Aragón, La Mancha and Andalucía. The distribution of benthic microbial mat communities in these lakes is described and rela-
ted to the ionic composition, salinity fluctuations, inundation period and sediment features. It was found that the cyanobacte- rium Microco1eu.s chthonoplastes is the most widespread species that occurs under a whole range of environmental conditions among which e.g., high Na+, high Mg2+ and high pH. The species composition and the annual cycle is described in more detail
for four different mat types. Finally, the scientific and naturalistic values of the microbial mats in inland lakes are discussed. We propose that the presence of interesting microbial communities should be used as an important element for wetland conserva-
tion policies.
INTRODUCTION
Microbial mats are benthic communities dominated by microorganisms which form cohesive layers at the sediment
surface. Commonly, these layers can be peeled off the sedi- ment and, apparently, the ecosystem owes its name to this feature. Phototrophic oxygen-producing microorganisms which include cyanobacteria and algae, are primary produ- cers in most microbial mats. In addition, a large chemoor- ganotrophic bacteria1 flora thrives on the excretion products and dead material from the phototrophs. Due to the high respiration activities, anoxic conditions are established only a few tenths of mm below the zone of oxygen photosynthe-
sis (REVSBECH et al., 1983). In the anoxic zone, sulphate- reducing bacteria produce sulphide. Hence, sulphide-oxidi- zing bacteria comprising phototrophic and chemotrophic
sulphur bacteria often form clearly visible laminas at the oxygen-sulphide interface just below the oxygenic photo-
trophs. Dense layers oí' benthic oxygenic phototrophs only deve-
lop if animal grazing is repressed and competition from macrophytes is virtually absent. Therefore, microbial mats only develop in extreme environments and are commonly found at high temperatures along the outflow of thermal
wells, temporarily flooded plains in marine environment and hypersaline environments. In this volume, microbial mats on coastal plains are described by ESTEVE et al. (1992) and those found in coastal salinas are described by DE WIT &
GRIMALT ( 1992). Continental saline and hypersaline lakes occur worldwide
in semiarid climates (WILLIAMS, 1986) and are also a charactefistic feature of the Iberian peninsula (MONTES & MARTINO, 1987). An extensive literature study on micro-
bial mats in inland saline lakes has been published by BAULD (1981). Different mats have been found in the lakes of the Great Rift Valley (Africa), the deserts of America and Australia. More information on mats in Australian lakes is
provided by BAULD (1 986). However, studies of microbial mats in inland salt lakes are restricted and mainly focused on descriptions of cyanobacterial and algal species. Thus, most current research on microbial mats that addresses more fundamental ecological and physiological questions, has been performed in thermal and coastal environments only. Nevertheless, Spanish inland salt lakes encompass a much wider variety of ionic composition types (MONTES &
MARTINO, 1987), seasonal fluctuations and sediment features than the thalassic water bodies. We will focus on the fact that these diverse inland environments represent an
Liiiiiictica, 8: 197-204 (1992) O Asoci;ición Espanola de Limnologia, Madrid. Spaiii

ideal research object to study which environmental factors allow the formation of microbial mats.
ABUNDANCE AND VARIETY OF INLAND SALT LAKES IN SPAIN
The presence of inland saline lakes is one of the most original limnological elements of the Iberian Peninsula. These waterbodies are associated with endorheic phenomena in semiarid ciimates and, in Western Europe, are thus confi- ned to Spain. These lakes occur in groups known as limno-
logical districts os endorheic centers. Their distribution is shown in figure l . In this figure we did not include the subsaline lakes (up to 10 g/L salinity) as for example found
in the Duero basins. As a rule, no microbial mats were found
in these lakes. However, an exception was made for the highly alkaline (pH 9.5 to 11) soda lakes found close to Coca (Segovia), since some interesting mats have been
found in thcm.
Figure 1. Map of the Iberiari Peninsula indicating the occurrence of microbial mats in inland saline or alkaline lakes (closed circles=saline lake with conspicuous microbial mats; open circles=saline lake in which no microbial mats have been found so far; closed square=subsaline soda lake ( p H 9 . 5 ) with microbial mats: open squares=subsaline soda lakes without conspicuous microbial mats).
The ionic composition of the iriland saline lakes has been studied previously by COMIN & ALONSO (1988) and MONTES & MARTINO (1 987). In summary, the Spanish inland lakes encompass a wide spectrum of ionic types: in the Andalucian lakes sodiurn predominates with either chlo- ride of sulphate as the dominant anion; the lakes in La
Mancha mainly contain magnesium sulphate; and the lakes in Aragón encompass a whole range from sodium chloride to magnesium sulphate. The unique permanent lake "La
Salada de Chiprana" belongs to the magnesium sulfate type (GUERRERO et al., 1991). Strikingly, the highly alkaline lakes close to Coca (Segovia) are of the sodium carbonate
type and, thus, are typical examples of soda lakes. The Spanish continental salt lakes encompass a broad
variety of different physico-chemical conditions, seasonal fluctuations and sediment features. Lake Chiprana (Sp. "Salada de Chiprana") (Chiprana, Zaragoza) is the only deep permanent hypersaline lake found in Spain. This lake is stra- tified during almost the entire year, has a maximum depth
of 5.6 m and a total surface of 31 ha; the general physico- chemical and geomorphological features have been descri-
bed by GUERRERO rt al. (1991). In contrast, al1 other
Spanish saline lakes are shallow and temporal water basins
which contain water during 3 to 10 months of the year. During the inundation periods, the water leve1 and conco-
mitantly the salinity of these shallow lakes is subjected to extreme temporal fluctuations.
Figure 2 shows the fluctuations of the salinity during the studied periods as found in the temporal shallow lake "La Dehesilla" (Cuenca, Manchego endorheic district) (period
1988- 1989) and the permanent saline lake Chiprana (1 989). In "La Dehesilla" the salinity fluctuations amounted to a
factor five (20 to 100 g/L). In contrast, in lake Chiprana, the salinity was remarkably constant in the anoxic bottom layer, while the salinity fluctuations of the oxic top layer were
aproximately two and half fold (30 to 73 g/L). During the year 1989, Lake Chiprana was only once overturned during
the stormy days of October: S o far, it is not known if the
overturn was an uncommon event os is repeated every year. Hence, this lake can not be classified yet as mono- of mero- mictic.
AN OVERVIEW OF THE MICROBIAL MATS FOUND IN THE SPANISH INLAND SALT LAKES
One of the most interesting examples of consistent micro- bial mats in Spanish inland lakes is found in the unique perma- nent hypersaline Lake Chiprana. The sediments of gentle slope from about 1.5 m water depth up to the shore, are covered with microbial mats alternated with prairies of the macrophyte R~cppia mur.irimu vas. mur.itimu. Both communities are clearly segregated in space. Very thick (4-9 mm) and cohesive mats
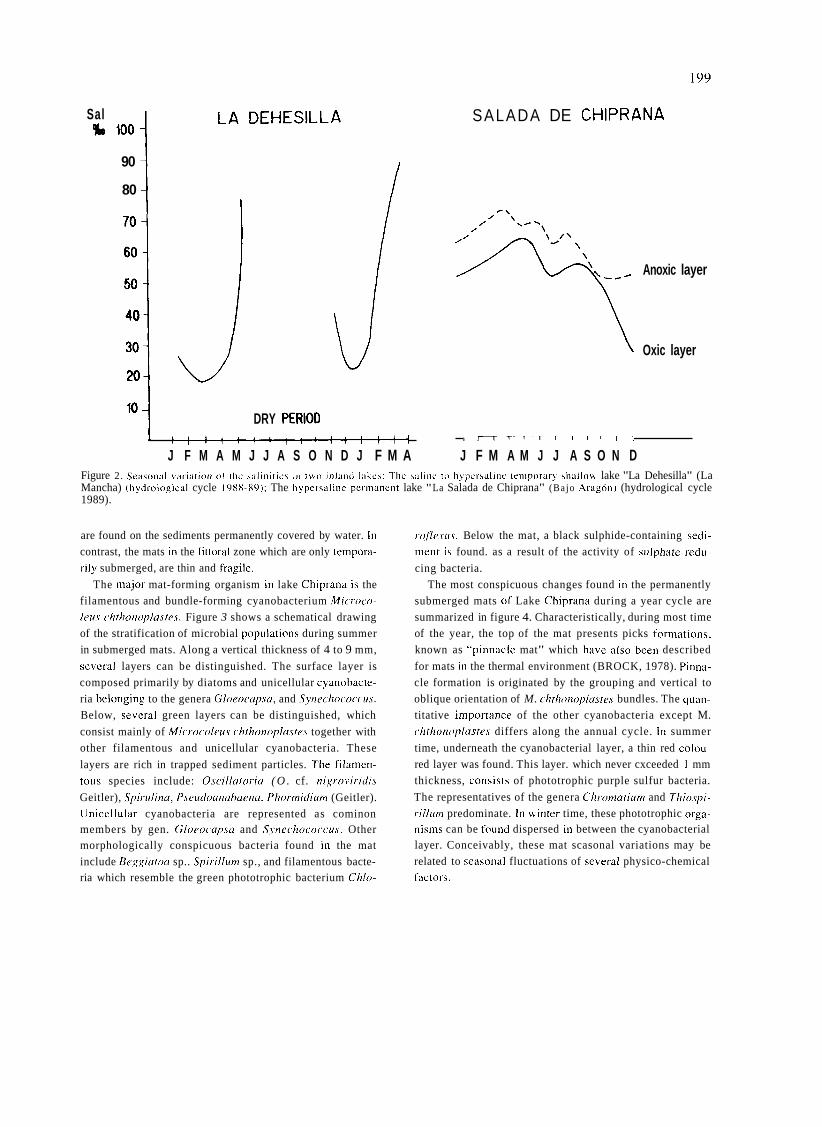
Sal
90
80
DRY PERIOD
SALADA DE CHIPRANA
Anoxic layer
Oxic layer
1 ; ; ; ; ; ; : ; ; ; ; I l ' l l l l l l 2 i l I I I ~ I I I I ~ I I
J F M A M J J A S O N D J F M A J F M A M J J A S O N D Figure 2. Seasotial varintioil ol ilie saliiiii~cs i i i tho i i i l i i i i< l lahe\: Tlic \aliiie to hqpersuliiie 1emporai.y shalloh lake "La Dehesilla" (La Mancha) (hydrological cycle 1988-89); The hypersaliiie pei-maiient lake "La Salada de Chiprana" (Bajo Aragón) (hydrological cycle 1989).
are found on the sediments permanently covered by water. In
contrast, the mats in the littoral zone which are only tempora-
rily submerged, are thin and fragile.
The majos mat-forming organism in lake Chiprana is the
filamentous and bundle-forming cyanobacterium Mic,l-o<,o-
1eu.s <~hthono/?la.stes. Figure 3 shows a schematical drawing
of the stratification of microbial popuiations during summer
in submerged mats. Along a vertical thickness of 4 to 9 mm,
several layers can be distinguished. The surface layer is
composed primarily by diatoms and unicellular cyanobacte-
ria belonging to the genera Glorocapsu, and .Tynechoc.occ.us.
Below, several green layers can be distinguished, which
consist mainly of Micr-ocoleus chthonoplastes together with
other filamentous and unicellular cyanobacteria. These
layers are rich in trapped sediment particles. Tlie filamen-
t o ~ i s species include: Osci1laroi.i~ ( O . cf. nigr-oi~i1.idi.s
Geitler), Spii.itlina, Pse~ldocrnahaena, Phor-midiurn (Geitler).
Unicellular cyanobacteria are represented as cominon members by gen. Gloc,oc,apsa and ~ne( .hococci t , s . Other morphologically conspicuous bacteria found in the mat
include Bc~~gia tou sp.. Spir.illum sp., and filamentous bacte- ria which resemble the green phototrophic bacterium Chlo-
i.qf1e.r~~. Below the mat, a black sulphide-containing sedi-
rrieirt is found. as a result of the activity of sulphate-redu-
cing bacteria.
The most conspicuous changes found in the permanently
submerged mats of Lake Chiprana during a year cycle are
summarized in figure 4. Characteristically, during most time
of the year, the top of the mat presents picks formations,
known as "pinnacle mat" which have also been described
for mats in the thermal environment (BROCK, 1978). Pinna-
cle formation is originated by the grouping and vertical to
oblique orientation of M. c,hrhonopkisres bundles. The quan-
titative irnportance of the other cyanobacteria except M. c~hthorzop1aste.s differs along the annual cycle. In summer
time, underneath the cyanobacterial layer, a thin red colou-
red layer was found. This layer. which never cxceeded 1 mm
thickness, consists of phototrophic purple sulfur bacteria.
The representatives of the genera Chr.ornuti~lm and Thiospi-
l-illi~nz predominate. In winter time, these phototrophic orga- nisms can be lound dispersed in between the cyanobacterial layer. Conceivably, these mat scasonal variations may be
related to seasonal fluctuations of several physico-chemical factors.

Among the temporal shallow lakes studied so far, cohe- sive mats with Microco1e~l.s chthonoplastes as the main mat-
LA SALADA DE CHIPRANA
LONGAR
Figure 3. Drawiiiph of the iiiaiii phototi-ophic nnd clieiiiotrophic organisms found in microbial mats in four inland saline lakes. a: diatoms; b: Gloeocapsa; c: Synechococcus; d: Microcoleus; e: Osci- Ilatoria; f: Pseudoanabaena; g: LPP; h: Spirulina; i: Chromatium; j: Thiospirillum; k: Anabaena; 1: Beggiatoa; m: eukaryotic green algae; A: green layer; B: red layer; D: white layer; C: sediment. (Drawing made by Ana Isabel López)
forming organism have been found in "La Muerte" and in "Guallar" in the Aragón endorheic center (Monegros, Zara- goza) and in "Alcahozo" in La Mancha (Ciudad Real). The total asea of the submerged sediments in these lakes is colo- nized by the rnat, and macrophytes are not present. During the summer, these lakes are dry and the benthic communities form typical drought crusts, in which the microorganisms survive in a physiologically inactive form. However, the organisms rapidly become active again upon inundation of the lake.
Microcoleus chthonoplastes is also the dominant mat- forming cyanobacterium in a the alkaline hyposaline soda lake "Caballo Alba" (Coca, Segovia). This lake has a pH of 10.5-1 1 and a concentration of dissolved salts of 3-4 g/L. Cohesive mats can be found in dispersed patches on submer- ged sediments.
A different kind of cohesive mat was found in lake "La Dehesilla" (La Mancha basins, Cuenca). A thin flexible green sheet covers the submerged sediments. The dominant mat-forming organisms in this mat are the large size unice- lular cyanobacteria, presumably G1oeocap.s~ sp., and diffe- rent very thin filamentous cyanobacteria belonging to the LPP-B group. This mat also forms a drought crust during the dry periods which persists in this lake for approximately nine months. The schematic drawing of the mat is shown in figure 3, and the yearly cycle, in figure 4.
Another type of mat was found in the hypersaline lake "Longar" (La Mancha). This lake is hypertrophic due to the input of waste water from the adjacent village Lillo. The surface of exposed wet sediments is covered by a cohesive mat composed of two morphologicaly distinct types of 0sc.i- Ilator-iu with minor contributions of Pseudoanabaena and LPP-forms. Under the green superficial layer (of 1-2 mm thickness), a thin red layer of photosynthetic purple sulphur bacteria developed. The highly eutrophic water column, that has a green colour due to a phytoplanktonic bloom, severely attenuates the irradiance at the sediment surface. As a result, the submerged microbial mats become detached and float on the water. The schematic drawing of microbial population and also the yearly cycle are shown in figures 3 and 4 respectively.
For the remainder of the saline temporal lakes studied so far, microbial mats occur only as very thin green layers. These occur in dispersed patches on exposed sediments which remain in a moisty state due to capillary rise of subsurface water. Characteristically, these mats loose cohe-

DRY P E r n
LlCROCWUS CHlHOrmPLASTES WRPLE PHOTOSYWTHETK: BACTERIA -
LONGAR L\ ~ Y W R P L E PHOTOSYNTHETIC BACTERIA
DRY PEm
MKROCOLEUS CHTHCNOPLASTES WlCROCOLEUS CHTHONOPLASTES PüRPlE PHOTOSYNTHETIC BACTEMA -
WRPLE PHOTOSYWTHTK: BACJERIA
Figure 4. Schematic representation of the seasonal variation of the benthic coniinuiiities iii lour iiilaiid \aliiic lahes ("La Salada de Chiprana" (Bajo Aragón); "Longar" (La Mancha); "La Albardiosa" (La Mancha); "La Dehesilla" (La Mancha).
LA SALADA DE CHIPRANA
siveness and become detached from the sediments upon However, presently, it is known that some lakes never
flooding at the onset of the inundation period. M a t - b u i 1- contain microbial mats. An intriging case is presented by
ding organisms in such thin microbial mats include Oscilla- lake "Piñol" (Aragones center, Monegros, Zaragoza) a toria subbrevis (as e.g. in "Carravalseca", Alava) and hypersaline lake which never contains microbial mats and
Anabaena sp. (as e.g. in "La Albardiosa", La Mancha, located only 500 m apart from lake "La Muerte" with very
Toledo). cohesive mats. Apparently, the climatological and physico- A very interesing succession has been found during a chemical properties of both lakes are very similar. Concei-
year-cycle (1991) in lake "La Albardiosa" (Toledo), which vably, the sediment features are another important prerequi- is depicted in figure 4. During this period, the lake changed site for microbial mat formation. Therefore, we analysed the from subsaline into hypersaline. In early spring, abundant granulometric composition of both lakes; the results are
growth occurred of aquatic macrophytes at low salinity and shown in figure 5 . It turned out that both compositions are
no mats were found. However, the macrophytes died massi- only slightly different. However, a clear difference was vely upon increasing salinity and, subsequently, thin micro- observed with respect to the fractions of small size (up to bial mats formed by Anabaena colonised the sediments. 0.1 mm diameter): these fraction were of negligable impor-
Below the cyanobacterial layer, a white layer was found tance in "La Muerte", but amounted up to 20 % in "Piñol".
which consisted of the chemotrophic sulphur bacterium Beggiatoa. A schematic drawing of mat microbial popula- tion is shown in figure 3. The succession started in shalow areas near the shore where hypersaline conditions were esta- blished earliest.
As can be apreciated from figure 1, microbial mats have not been found in al1 Spanish inland saline or alkaline lakes. Obviously, more microbial mats will be discovered in the future when more intensive studies will be performed.
DISCUSSION AND PERSPECTIVES FOR FUTURE RESEARCH
So far, the study on microbial mats of the inland saline lakes in Spain has been mainly descriptive as it has been in general for the microbial mats in inland saline lakes (BAULD, 198 1; BAULD, 1986). Presently, with respect to

202
the lberian Peninsula, we posses a fairly complete know- legde of the distribution and variability of microbial rnats in the inland saline lakes (see fig. 1). Especially, the Aragonés center, the basins in la Mancha, and the lakes in Coca have been visited very intensively. In contrast, Andalucian lakes received less attention so far, and consequently, in the near future, new mats may be discovered in this region. Nevert- heless, the data obtained, represent valuable information on
the distribution of rriicrobial mats and will serve as a basis to
address several questions with respect to microbial mat formation and dynamics.
The ubiquitous benthic cyanobacterium Mi(~r.oco1c~lr.s
c~kthorloplast~.~ turned out to be a very succesful organism in inland lakes. This organism formed cohesive mats in the permanent hypersaline Lake Chiprana, in several of the temporal shallow lakes of high salinity, and also in the unique subsaline soda lake "Caballo Alba". As a matter of
fact, Mic~rocolezrs chthoriop1uste.s was the mat-forming
cyanobacterium in most submerged cohesive mats in the inland lakes. Mic~roc~o1eu.s c~hthonoplastes is also common in the coastal environment; it occurs both in supratidal marine
mats (ESTEVE et al., 1992) as well as in coastal salinas at salinities from about 80 up to 140 g/L (THOMAS, 1984; DE WIT & GRIMALT, 1992). In addition, in coastal sali-
nas, very consistent mats of Aphunolher.~ spp. and Chroma- tiaceae are found at salinities ranging from 150 to 300 g/L.
These very high salinities, however, do not occur for prolon- ged periods in the inland saline lakes.
The tolerance of Miel-oc~o1eu.s (~htko/~opla.stc~.s to very alkaline conditions in "Caballo Alba", is not a surprise.
Microelectrode studies demostrated that highly alkaline conditions often occur during daytime in the cyanobacterial layer of the mats and must be attributed to a very high rate
of C0,-fixation (REVSBECH et al., 1983).
Bivalent cations at high concentrations, especially M&*+, may be toxic and can inhibit growth. It has been reported, that the sensitivity towards these cations is actually increased at higher salinities, and that cyanobac-
teria are only able to grow at 4 mM NaCl when bivalent cations are absent (BAAS BECKING. 1931 cited in BAULD, 198 1 ). Nevertheless, abundant Mic~r.o(~olcos mats
were found in Lake Chiprana at 70 g/L salinity with Mg2+
as the predominant cation. Obviously, the actual Mg2+ tolerance has to be assessed in more detail in growth expe- riments. It may turn out that this tolerance is actually strüin specific, when studying strains from Mg*+ poor and Mg2+
rich environments. Such studies are very relevant for the understanding of evolution in bacteria1 species. The inland
salt lakes constitute an ideal study object since they provide an enormous variation within a geographically
relatively small area. In several lakes, microbial mats only occurred as very
thin layers on exposed sediments. These mats with 0.sc'illa-
rol-in spp. os Anc~haencr sp. as the dominant species, became detached from the sediment upon temporal flooding in subsaline water. Apparently, in these mats the ability to
maintain cohesiveness in subsaline water is indeed a limi-
ting factor for its occurrence. In contrast, the Microc,oIeu.s
arid Gloeocapsa sp. mats in more saline waters maintained cohesiveness and remained adhered to the sediment throug-
hout the inundation period. Microbial mats in inland saline lakes are exposed to
strong variations of the salinity (2.5 t 5-fold see fig. 2), such in clear contrast to the mats found in the coastal salinas. Apparently, Mic,l-ocole~ls c.hthonop1a.ste.s is adapted to fluc-
tuating environmental conditions, but it is also competitive
under very constant conditions. In the inland lakes, two stra- tegies have been observed which allow the cyanobacteria to
cope with the fluctuating conditions. Firstly, cohesive
submerged mats of Microc~o1eu.s c,hthonoplastes and Gloeo-
czpsn formed drought crust which survive the dry periods
and which rapidly become active again upon wetting. Secondly, the cyanobacteriu'm Anuhoerla in "La Albardiosa"
did not survive the adverse subsaline periods, but demons-
trated a very rapid colonisation rate upon the establishment of the favourable conditions.
Mats with heterocystous cyanobacteria as the mat- forming organisms are very rase, even in inland lakes (BAULD, 1981). STAL & KRUMBEIN (1985) reported
that non-heterocystous filamentous cyanobacteria in micro- bial mats are actively fixing nitrogen during limited periods
of the dial cycle, thus achieving a temporal separation
between incompatible oxygen production and nitrogen fixa- tion. S o far, it remains to be investigated why in lake "La Albardiosa" the heterocystous Anahaerla sp. predominates over such non-heterocystous species.
This study confirmed that at low salinities up to 40 g/L the macrophytes rnainly Rzrppia maritimc~ are more compe- titive than the microbial mats, but at higher salinities the reverse occurs. Similar observations have been reported for
coastal salinas (THOMAS, 1984; DE WIT & GRIMALT,
1992). In the lake "La Albardiosa" a very interesting cycli- cal succession was observcd which was directly related to salinity changes. Surprisingly, in lake Chiprana, the micro- bial mats occurred together with prairies of Rzrppia mai.itimu
at salinities fluctuating from 30 to 75 g/L (see fig. 2). The

PINOL (1 Ocm) LA MUERTE (10 cm)
Figure 5. Si/e distrihutioii ot tlie ~eiliiiiciit pnrticlc\ (top 10 cm) in two hypersaline temporal lakes in "Loa Monegros" Aragón. B«th lakes are separated by 500 m and have similar climatic and salinity conditions. The sediments of lake "La Muerte" are completely cove- red by microbial mats: in contrast, microbial mats are always absent in lake "Piñol".
actual competition between these entities has to be studied
in more detail. As shown above, the study of microbial mats in the
diverse inland lakes can significantly contribute to study
several ecological, physiological and genetic questions,
which are not easily addressed in the coastal environments.
Nevertheless, more experimental studies wili be required
to entangle these questions. The information actually avai-
lable represents a valuable basis to choose the relevant rese-
arch objects and sites. It is, however, highly important
to maintain the enormous variability found among diffe-
rent inland saline lakes. Therefore, we discuss its actual
state of conservation and future perspectives in the follo-
wing section.
THE URGENT NEED FOR A CONSERVATION POLICY OF THE SPANISH INLAND SALINE LAKES
The inland saline lakes possess very important naturalis-
tic, scientific, esthetic, economic and cultural values as has been mentioned by various authors (HAMMER, 1986;
WILLIAMS, 1986; GONZALEZ BERNALDEZ, 1987). With respect to its scientific values, these lakes are espe- cially important study objects for microbial ecology, because of their interesting microbial ecosystems. Espe- cially, as aforementioned, the study of microbial mats in
inland saline lakes may contribute to a better understanding of the general features of these ecosystems. Due to the high
population densities and their conspicuous features come of
which can even be appreciated with the naked eye, the
microbial mats in saline environments represent ideal study
cites which can be succesfully studied with current techni-
ques available in microbial ecology. As such, these mats can
be studied as model systems, thus allowing to enlarge gene-
ral knowledge on microbial ecology. Comparably. in the
United States, the study of microbial ecosystems in extre-
mely thermal environments greatly expanded general know-
ledge on general microbiology and microbial ecology
(BROCK, 1978). Unfortunately, the conservation of the inland saline lakes
is highly threatened. The general conservation problematics
are described by MONTES & MARTINO (1987), and have
been discussed at local scientific meetings (GUERRERO et al.. 1992). The saline lakes have suffered form contamina-
tion and eutrophication by discharge of waste water.
Presently, large scale extraction of brines causes great
damage in some hypersaline lakes. In contrast, traditional
small scale salt harvesting in salinas connected with saline
lakes can be compatible with the conservation of these lakes
and may even give sise to interesting habitats for microbes
such as ponds for extremely halophilic archabacteria (DE WIT & GRIMALT, 1992). The recent large scale extension
of agricultural irrigation has a very negative impact on the
water regime of the saline lakes. Depending on the local
conditions, irrigation can lead either to the increase of the
water mass and concurrent desalination due to runoff, os to
a drop in the ground water table resulting in increasing sali-
nities and shorter inundation periods.
The interesting microbial mat ecosystems, which have
been described in this Chapter, constitute a valuable element
of the inland saline lakes. Therefore, we believe it is esssen-
tial to conserve these lakes for future generations and coun-
teract the various abovementioned negative developments.
Unfortunately, microorganisms and microbial ecosystems
often have not been taken into account when establishing
priorities for wetland preservation. We believe that the
presence of original microbial ecosystems highly justifies a
conservation of these wetlands in their natural state.
ACKNOWLEDGEMENTS
We are indebted to Dr. Carlos Montes for helpfull discus- sions. Jesús Balsa, M- Luisa Pascual, Ana Belén Tadeo

contributed during sampling and analyses. This project was partly subsidized by the Spanish Ministry of Education (CAYCIT proy. No. NAT 90-0721-C02-01). Rutger de Wit has been supported by fellowships from the Spanish Ministry of Education and Science and European Environ- mental Research Organization (E.E.R.O.).
REFERENCES
COMIN, F. & M. ALONSO. 1988. Spanish salt lakes. Their chemistry and biota. Hydrobiologia 158:237-245
BAULD, J. 1981. Occurrence of benthic microbial mats in saline lakes. Hydrobiologia 8 137-1 11
BAULD, J. 1986. Benthic microbial communities of Austra- lian saline lakes. In: P. de Deckker & W.D. Williams (ed.) Limnology in Australia, pp. 95-1 11. Dr.W. Junk Pub., Boston.
BROCK, Th.D. 1978. Thermophilic microorganisms and lifi~ ( r i Iri:.ll tcr~r/~o.otr~i.c, .r . Sprinpci-. Ve\\ York.
DE WITT, R. & J. GRIMALT. 1992. Microbial ecosis- tems in Spanish coastal salinas: an ecological and geoche- mical study of biomarkers. Limnetica, 8.
ESTEVE, 1.; M. MARTINEZ, J. MIR & R. GUE- RRERO. 1992. Distribution, typology and structure of microbial mat communities in Spain: Preliminary studies. Limnetica, 8.
GONZALEZ BERNALDEZ, F. 1987. Las zonas enchar- cables españolas. El marco conceptual. In: Bases CientijFi- cas para la protección de los humedules en España, pp. 9-30. Real Academia de Ciencias Exactas, Físicas y Naturales de Madrid.
GUERRERO, M.C., J. BALSA, M.L. PASCUAL, B. MARTINEZ, B. & C. MONTES. 1991. Caracterización limnológica de la laguna Salada de Chiprana (Zaragoza, España) y sus comunidades de bacterias fototróficas. Limné- tira, 7: 83-96.
GUERRERO, M.C., B. VIDONDO, J. BALSA & C. MONTES. 1992. La Salada de Chiprana: Características y singularidades. Proceedings of: Jornadas sobre el futuro de las Saladas de Monegros y el Bajo Aragón. Grupo Cultural Caspolino (ed.). Zaragoza (in press).
HAMMER, V.I. 1986. Saline lakes ecosystems of the world. Dr.W. Junk Pub., Boston.
MONTES, C. & P. MARTINO. 1987. Las lagunas sali- nas españolas. In: Bases Cientficas para la protección de los humedales en España, pp. 95-145. Real Academia de Ciencias Exactas, Físicas y Naturales de Madrid.
REVSBECH, M.P., B.B. JORGENSEN, T.H. BLACK- BURN, T.H. & Y. COHEN. 1983. Microelectrode studies of the photosynthesis and 02, H2S and pH profiles in a microbial mat. Limnol. Oceanogr. 28: 1062-1 074.
STAL, L., H. VAN GEMERDEN & W.E. KRUMBEIN. 1985. Structure and development of a benthic marine micro- bial mat. FEMS Microbiol. Ecol. 31: 11-125.
THOMAS, J.C. 1984. Formations benthiques: Cyanobac- téries des salins de Santa Pola (Espagne): Composition spécifique, morphologie et caractéristiques biologiques des principaux peuplements. Rev. Inv. Geol. 38/39:139-158.
WILLIAMS, W.D. 1986. Limnology, the study of inland waters: a comment on perception of studies of salt lakes, past and present. In: P. de Deckker & W.D. Williams (ed.) Limno- logy in Australia, pp. 471-496. Dr.W. Junk Pub., Boston.

MICROBIAL ECOSYSTEMS IN SPANISH COASTAL SALINAS; AN ECOLOGICAL AND GEOCHEMICAL STUDY OF BIOMARKERS.
R. de Wit and J. 0. Grimalt Dept. of Environmental Chemistry. C.1.D.-C.S.I.C. Jordi Girona 18-26. 08034 Barcelona. Spain.
Keywords:Biomarker, Salina, Hypersaline, Microbial mat, Cyanobacteria, Lipids, Highly branched C20 alkene.
ABSTRACT
Coastal salinas (solar salterns) are human-made environments that comprise an array of evaporation ponds of increasing sali- nities. Therefore, salinas are appropriate systems to study the biotic communities and the different biogeochemical processes in relation to the water column salinity. At higher salinities the microorganisms are the main organisms; above 300 g/l salinity the
extremely halophilic archaebacteria predominate. Microbial mats are the conspicuous benthic ecosystems found from 50 to 300
g/l salinity. Lipid biomarkers have been used to trace microorganism and their remains at various depth layers in the sediments covered
by the microbial mats. The microscale vertical distribution of some compounds as e.g., a highly branched C20-alkene, is evalua- ted in relation to the stratification of the bacterial populations. The use of biomarkers in ecology and geology is discussed.
INTRODUCTION
Coastal salinas have been studied frequently by micro- biologists and geologists. These human-made ecosystems are extreme environments predominated by microorganisms that represent a source of novel extremophiles. Furthermore,
the salinas allow the study of how the salinity determines the composition of the communities. For the geologists the salinas represent a model system to study salt precipitation
and related biogeochemical processes in shallow evaporitic environments. The formation of evaporite deposits along the earth history is well documented; about 25 % of the conti- nental areas are underlain by rocks which originated in such environments (BLATT et al., 1980).
We have chosen the salinas within the framework of our multidisciplinary studies on organic geochemistry and microbial ecology of hypersaline microbial ecosystems. Our
interest is to trace microorganism by characteristic lipid
components. It will be shown that lipid components can be
used as biomarkers to analyze the living communities. Furt- hermore, biomarkers are good tools in palaeolimnological and geological studies, because several organisms leave lipid traces in the sedimentary record. First, the features of the coastal salinas and their biota will be treated in detail. Subsequently, the use of biomarkers in microbial ecology
and organic geochemistry will be discussed using some
examples from our work as an illustration.
THE COASTAL SALINAS AND THEIR BIOTA
Salinas are human-made environments that are exploited to obtain halite (NaCI) for human consumption and indus- trial purposes. The process is based on the evaporation of brines using the sun and the wind as the energy sources.
Thus, salinas are restricted to areas with climates characte- rized by periods during which evaporation exceeds precipi- tation such as the Mediterranean region. In Spain salinas are found on the coast and were common inland. Coastal Atlan-
tic salinas are found as far north as the French Bretagne
(POISBEAU-HEMERY, 1980) and even on the river Mersey (Manchester, U.K.).
Figure 1 gives a distribution map of the coastal salinas that are actually exploited in the Spanish part of the Iberian
peninsula and the Balearic islands. Most salinas are located along the Mediterranean coast, but some are found along the Atlantic coast in the provinces of Huelva and Cadiz. A comparison of figure 1 with the maps provided by LEFOND (1969) and DRONKERT (1985) demonstrate that the number of coastal salinas under exploitation has declined
Limnetica, 8: 205-2 12 ( 1992) O AsociaciCIn Espaiiola de Limnologia, Madrid. Spain

Figure l . Map ot'ilie consial \alirlas tliat are act~ia l ly exploited in Spain (Peninsula aiid Balearic 15l;inds).
dramatically during the last two decades. This is the result of low salt prices on the international market due to competi- tion from fossil salt mines, the need for mechanization of
the exploitation and the high prices paid for the urbaniza-
tion of coastal areas. Most coastal salinas use marine water (thalassic waters),
which has a very constant ionic composition world round. In Spain, the salina of Torrevieja (Nc 7 in fig. 1) is an excep- tion because it is fed with a mixture of marine water and brine obtained from Triassic salt rocks. In the Atlantic sali-
nas, the sea water intake normally proceeds at high tide through a system of channels; at decreasing tide, sluices are
closed to keep the sea water in these systems. In the Medi-
terranean region, this system is less eHicient due to the small tidal difference. Formerly, seepage through dune barriers used to feed the ponds in some salinas. Presently, however, pumping is used instead of or in addition to direct inflow
arid seepage. A very characteristic continuous process is used to obtain
enriched halite in the salinas. Sea water is initially stored in
large lagunar shaped ponds, tlie so-called del~ositos. Subse- quently, it flows through a system of consecutive ponds (see
fig. 2), while continuously evaporating and concomitantly increasing its salinity. This process results in sequeirtial
precipitation of different salts (USIGLIO, 1848) which is shown in figure 2. First, sea water is reduced to approxima- tely 115 th while calcium carbonate precipitates and, subse- quently. to approximately 1/10 th of the original volume while gypsum precipitates. The brine thus obtained is used to feed the cristalizers where halite is obtained. Finally, a brine highly enriched in potassium salts remains, which only
comprises 1 %, of its original volume. These brines are called bitterns, and, gerierally, are either discarded into the sea or used to wash the halite. As a result from this exploitation, a salina encornpasses a salinity gradient ranging from sea
water (37 g/¡) up to halite saturation (350-370 g/]), corres-
ponding to domains characterized by the precipitation of calcite, gypsum and halite (fig. 2).
The most effective exploitation o f t h e salinas depends on
[he constancy of the salinities of the different ponds in the
array. Therefore, the salinity in the ponds is checked repea- tedly and the water flow readjusted in order to prevent sali- nity tluctuations. The salt concentrations are monitored by
density measurements which are expressed in degrees Baumé
(Specific Density= 145/(145-'Bé) kg/l). Not al1 salinas are exploited on a year round basis. Especially in the northern salinas, as e.&., in "Les Salines de la Trinitat", the salinity of
the ccrlentadorc~s, <,uhec.r.i.us and r~ristc11izadore.s decreases
during winter. In contrast, in the southeastei-n salinas (N" 5-
Depósitos Calentadores Cri<,alirrdura
Cabeceras
Volume of the brine m t of the salts Dreci~itated
Concentrating
Caco3 ponds
1 Cristalizadores 1 1 Cristalizers 1
F i f ~ i i e 1. (irapliic rcpi-e\eiit:~tioii ol tlic Iiriiic IIo\\ ,iiitl tlir e\apora- tion and precipitation processeh in Spiiriish conatal salinas. The names oF the different evaporation ponds are given in Spaniah and in English.

10 in fig. 1). the salinity gradient is maintained throughout the year, hence, gypsum and halite precipitation are even observed in winter, albeit at very reduced rates. These salinas
represent an ideal site for scientific research, because they
can be studied as a model system in which the biotic commu- nities and the different biogeochemical processes can be rela- ted directly to the salinity of the water column.
The biota found in hypersaline environments have been extensiveiy reviewed by JAVOR (1989). The orange-red
and the pink to purple-red colours of the most concentrated c,uhec,er.us arid the c~~.i.strrlizudo~.es ponds, represent perhaps the most well-known feature of the salinas. These colours
are due to carotenoids in planktonic microorganisms;
orange-red is related to the presence of the halophilic alga D~~nal ie l lu ; pink to purple-red indicates the presence of extreme halophilic bacteria that predominate at salinities exceeding 300 g/l. These extreme halophilic bacteria combine with methanogens and thermoacidophilic bacteria, forming the kingdom of the archaebacteria, that possess features which distinguish them sharply from both the
eukaryotes and the rest of the bacteria (presently named
eubacteria). The halophilic archaebacteria have been exten- sively studied by RODR~GUEZ-VALERA and coworkers (University of Alicante, Spain). Remarkably, out of 14 halophilic archaebacterial species described in Bergeys
Manual of Systematic Bacteriology (GRANT & LARSEN, 1989), 3 have been originally isolated from Spanish coastal salinas by this group (see table 1).
Salinas are typical exampies of "extreme environments"
in the sense that only a limited number oftaxonomic groups
is present. Fishes, macrophytes, i.e. Ruppia muritimu, and macroalgae, e.g., Cludophora spp., are found only in low salinity ponds (up to approximately 60 g/l). The invertebrate fauna has been studied extensively in the French Mediterra-
nean salina the Salin de Giraud (BRITTON & JOHNSON,
1987). It was observed that as the salinity increases the diversity of invertebrate species decreases. The depositas
have an invertebrate fauna poorer than the sea. The drop of
invertebrate species numbers is most pronounced from 40 to
70 g/l salinity. At 300 g/l salinity only the brine shrimp Artcrnicr .salir?a remains. RODR~GUEZ-VALERA et al.
(198 1, 1985) studied the planktonic microorganisms along the salinity gradient. Up to 250 g/l salinity, the planktonic
communities consist of halotolerant and halophilic microal- gae and eubacteria, whereas the halophilic archaebacteria predominate above 300 g/l.
The reduced activities of macrophytes and invertebrate animals, allow the proliferation of benthic cyanobacteria
resulting in the formation of microbial mats. Figure 3 gives a schematic representation of the different microbial mat communities found along the salinity gradients in the sali-
nas. This figure has been compiled from personal observa-
tions and previous reports (THOMAS, 1984; NOEL, 1984; CORNÉE, 1984). Microbial mats do occur from 50 up to 300 g/l saliriity. In the lowest salinity range, mats consist of
a top layer of the cyanobacterium Phor-midium i'alderiani/m
underlaid by B r g ~ i u t o u sp. and filamentous heterotrophic bacteria that can be grown chemoorganotrophically on pepton and yeast extract under anoxic conditions. Subse-
quently, mats can be found that consist of the ubiquitous
cyanobacterium Micr-ocoleus chthonoplastes accompanied by purple sulphur bacteria and the green filamentous bacte- rium Chlor(fl1e.i-us that has only recently been cultured by
LARSEN et al. (1991). Above 150 g/l salinity, the micro- bial mats have a typical undulated structure with gypsum crystals incrustations. The dominant cyanobacterium is Aphanothccc, sp. accompanied by layers of the purple sulp- hur bacteria Clironzutium su1c.i-i,qerr.s 2nd Thioc.upsa lzulo-
Table 1. Halophilic archaebacteria (fam. Halobacteriaceae) originally isolated from Spanish Salinas by F. Rodríguez Valera
and coworkers.
Organism: Morphology: Special features:
flattened disk or cup
flattened disk or cup
uses starch, found at salinities > 100 g/l(l) does not use starch
Haloar.c~ula hi.spanic,í< small pleomorphic rods uses starch
(1) In the natural environment, the other members of the Halobacteriaceae do not occur in high densities at salinities below 250
g/i.

Diatoms m
compact mats of Phonnidium valderianum Be~yiafoa sp.: heterotrophic
filamentous bacteria m
laminated mats of Microcoleus chthonoplostes Chloroflexaceae; niiocapsa hahphiia
mucous mat of Myxosarcina
Undulated mats with gypsum cristals Aphanothece , Phormidium Thir,cu~>.<a hulophila,Chromarium salexzyem
Concentration of dissolved salts in water column above the sediment (g/l)
Figure 3. Microbial inat cotiiiiluiiiiir.\ foutid i i i Spaiii\h con\tiil sali- nas at various dissolved salt concentrations.
phila that have been isolated recently by CAUMETTE et al. (1988, 1991) from French salinas.
THE BIOMARKER CONCEPT.
The use of "biomarkers" in microbial ecology and in orga-
nic geochemistry is based on the observation that some compounds are exclusively synthesized by certain taxa of organisms. The compounds most frequently used comprise
photosynthetic pigments and fatty compounds. A extensive referente on lipids in microorganisms of different groups is given in RATLEDGE & WILKINSON (1988). The phosp- holipid bilayers of the cellular membranes are the main
source of fatty acids, steroids and hopanoids. The rapid
development of gas chromatography (GC) and GC coupled to mass spectrometry (GC-MS) has enabled the identifica- tion and quantification of lipid compounds in environmental samples of small size. Commonly, mixtures of (2: 1 ) methy- lene chloride-methanol are used to extract the lipids from the samples. The extracts are saponified to recover fatty acids from phospholipids (so-called phospholipid-linked
fatty acids). Subsequently, fractionation techniques are used to aeparate the lipid mixtures in hydrocarbon, ketone, aidehyde, alcohol and fatty acid fractions. These procedures
have been described in detail by ALBAIGÉS et al. (1984), BARBÉ et al. (1991) and others.
In microbial ecology a "biomarker" has been defined as "chemical components of microorganisms which can be
analyzed directly from the environment and be interpreted
both quantitatively and qualitatively in terms of in situ microbial biomass" (PARKES, 1987). Presently, the quali- tative and quantitative determination of the bacterial species present in ecosystems is still unreliable due to technical
iimitations. It has been claimed that the classical counting techniques, which involve culturing on agar plates (colony forming units) os in liquid media (most probable number),
underestimate the bacterial population by a factor between 10 and 106 (PARKES, 1987). Obviously, new techniques are
required to get better estimations of microbial biomass and diversity, hence, the use of lipid biomarkers seems a promi-
sing tool (PARKES, 1987; VESTAL & WHITE, 1989). So
far, lipid biomarkers have proven to be especially useful for tracing various groups of sulphate-reducing bacteria in anoxic sediments (TAYLOR & PARKES, 1985).
In geological chemistry a "biomarker" has been defined as
"an organic compound of widespread occurrence in geologic samples that can be related to the original carbon-containing constituents of organisms" (ENGLINTON & CALVIN,
1967). For example, geological biomarkers indicative of hypersalinity have been recognized by comparison of fossil
facies (TEN HAVE et al., 1988), os can be recognized by studying the fate of different compounds in actual ecosystems
(see below). In microbial ecology, a biomarker is an actual constituent of a microorganism that ideally disappears upon
death. In contrast, in geology, a biomarker has to be conser- ved after death in its original form os as a post-depositional transformation product. Hence, the ecological and geological biomarker concepts are similar, but can be conflicting in their application. In practice, the living cell comprises a whole range of compounds of different biodegradabilities. For example, adenosine triphosphate (ATP) is a compound found
in al1 living organisms, that rapidly disappears upon cellular
death. Hence, ATP is commonly used to estimate the total biomass in different ecosystems. Likewise, phospholipids are believed to have a rapid turnover and, therefore, have also been used to estirnate total living biomass (KARL, 1986). In contrast, neutral lipids, steroids and other terpenoid lipids can be preserved for extended periods. However, the conservation and transformation processes strongly depend on the envi-
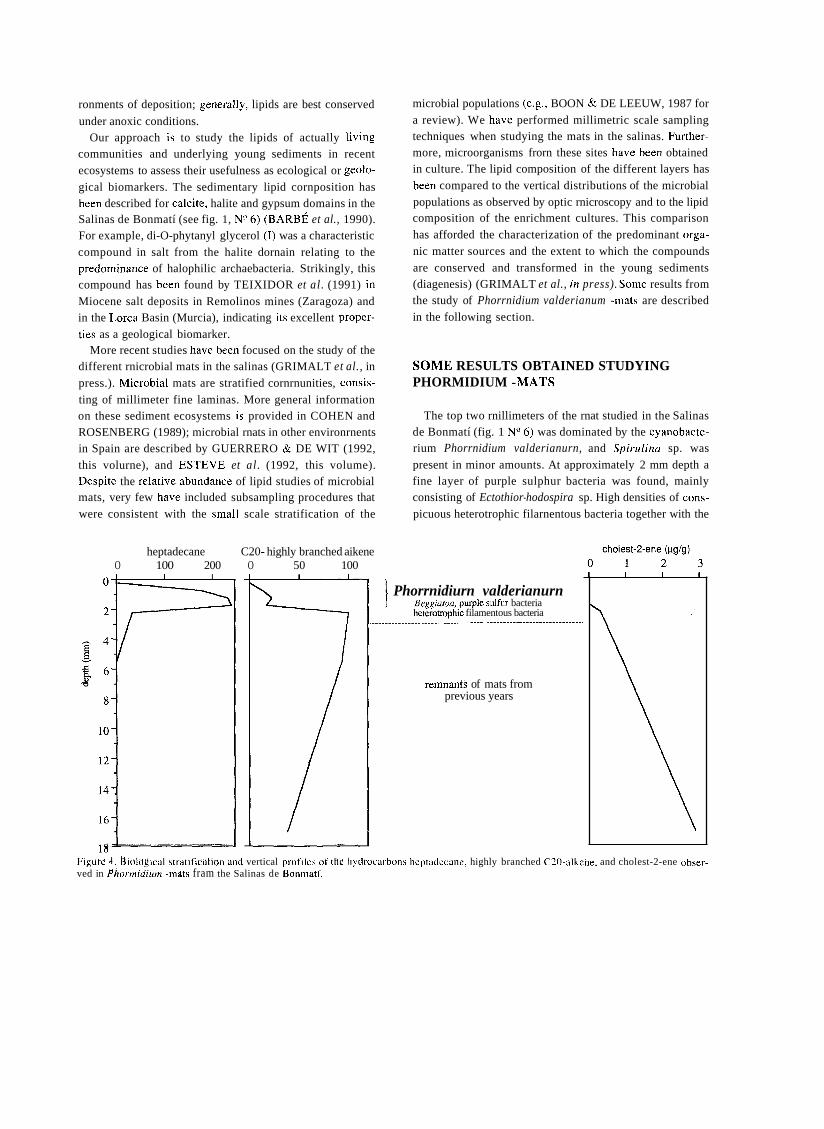
ronments of deposition; generally, lipids are best conserved
under anoxic conditions. Our approach is to study the lipids of actually living
communities and underlying young sediments in recent ecosystems to assess their usefulness as ecological or geolo- gical biomarkers. The sedimentary lipid cornposition has been described for calcite, halite and gypsum domains in the Salinas de Bonmatí (see fig. 1, N%) (BARBÉ et al., 1990). For example, di-O-phytanyl glycerol (1) was a characteristic compound in salt from the halite dornain relating to the predominance of halophilic archaebacteria. Strikingly, this compound has been found by TEIXIDOR et al. (1991) in Miocene salt deposits in Remolinos mines (Zaragoza) and in the Lorca Basin (Murcia), indicating its excellent proper- ties as a geological biomarker.
More recent studies have been focused on the study of the different rnicrobial mats in the salinas (GRIMALT et al., in press.). Microbial mats are stratified cornrnunities, consis- ting of millimeter fine laminas. More general information on these sediment ecosystems is provided in COHEN and ROSENBERG (1989); microbial rnats in other environrnents in Spain are described by GUERRERO & DE WIT (1992, this volurne), and ESTEVE et al. (1992, this volume). Despite the relative abundante of lipid studies of microbial mats, very few have included subsampling procedures that were consistent with the small scale stratification of the
heptadecane C20- highly branched aikene O 100 200 O 50 100
microbial populations (e.g., BOON & DE LEEUW, 1987 for a review). We have performed millimetric scale sampling techniques when studying the mats in the salinas. Further- more, microorganisms frorn these sites have been obtained in culture. The lipid composition of the different layers has been compared to the vertical distributions of the microbial populations as observed by optic rnicroscopy and to the lipid composition of the enrichment cultures. This comparison has afforded the characterization of the predominant orga- nic matter sources and the extent to which the compounds are conserved and transformed in the young sediments (diagenesis) (GRIMALT et al., in press). Some results from the study of Phorrnidium valderianum -rnats are described in the following section.
SOME RESULTS OBTAINED STUDYING PHORMIDIUM -MATS
The top two rnillimeters of the rnat studied in the Salinas de Bonmatí (fig. 1 N") was dominated by the cyanobacte- rium Phorrnidium valderianurn, and Spirulina sp. was present in minor amounts. At approximately 2 mm depth a fine layer of purple sulphur bacteria was found, mainly consisting of Ectothior-hodospira sp. High densities of cons- picuous heterotrophic filarnentous bacteria together with the
1 Phorrnidiurn valderianurn Beggiaroa, purple sulfur bacteria heteroaophic filamentous bacteria
remnants of mats from previous years
Figu1.P 4. Bioldgicai str~tificiltion aiid vertical profi le of t l lP hydrocarbons Iieptadecane, highly branched C20-alkene, and cholest-2-ene obser- ved in Phoiwtidium -mats fram the Salinas de Bonmati.

chemotrophic sulphide oxidizer Beggicitoc~ were found from
1 .S to 3 mm depth. Below 2.5 mm depth the sediment was
black-coloured indicating continuously anoxic conditions
and sulphide production. Bioturbation of these mats was not
observed. The laminations below the actually living mat
correspond to buried mats of former years. The vertical
distribution of some of the hydrocarbons in these sediments
is given in figure 4.
The high amounts of heptadecane in the 0.5-2 mm sedi-
ment sections is related to the presence of the cyanobacteria;
this compound has been found in several cyanobacterial and
alga1 species (e.g., GELPI et al., 1970) and is the major
hydrocarbon in cultures of Phoi.niidium i~alder.iavrum strain
P4. However, this hydrocarbon is virtually absent below 2
mm depth indicating a rapid degradation. In general, it has
been found that the cyanobacteria leave only minor lipid
features in the sedimentary record (GRIMALT er al., in
pr.ess). Thus, heptadecane is a good tracer for living cyano-
bacteria in these ecosystems.
A most interesting feature is the occurrence of the highly
branched C20-alkene (11) in the sediments of the Phornzi-
dium -mat; this compound is not found in Mi<.t.or.ole~r.s and
Aphunothec,e -mats. Corresponding C20-alkanes of the saine
structure have been found in older strata (TEN HAVE et al . ,
1988). Strikingly, the highly branched CZo-alkene has not
been found in c~iltiire\ oí' Phoi.tiritlir~ti~ i~iililrr.iiiiir~rti strniii
Figure 5 . ('o~i\ersioil o f cliolestari-313-01 ¡rito cliolrst-2-ciic (dehy- dration) that occura iii aalina sediinents.
P4, and its concentration was very low in the top 2 millime-
ters of the mat considered in this study. Most likely, the
compound is synthesized by anaerobic heterotrophic bacte-
ria that grow on the cyanobacterial remains. Nevertheless,
the biosynthesis of this compound has not been demonstra-
ted unambiguously by laboratory culture experiments.
The depth profile distribution of another hydrocarbon,
cholest-2-ene, is not related to a direct biogenic input, but
rather to a diagenetic dehydration of cholestan-3B-o1 as
depicted in figure 5. In addition, various transformation
processes are known; concomitantly with dehydration, hydrogenation takes place in layers of strongly reducing
conditions (GRIMALT et al., in press).
Figure 6. Sonie compourids cited ir1 the text: 1 = di-O-phytanyl glycerol: 11 = highly branched CZO-alkene.
In summary, these examples show something of the
complex biogeochemistry of the lipid compounds in micro-
bial mat communities. The combination of millimetric scale
sampling in combination with culturing experiments provides
a useful tool for its understanding. For more detailed infor-
mation the reader is referred to GRIMALT et al. (in pr-ess).
CONSERVATION OF THE SALINA ENVIRONMENTS
The coastal salinas are very useful for integrated ecologi-
cal, microbiological and geological research. The biotic
communities have a low species diversity, but are highly
original because they are composed of extremophiles. Thus,
they are a source of novel organisms (see e.g., table 1). The
occurrence of these interesting communities is dependent on
salt-harvesting activities by human beings. For al1 these
reasons, it is regrettable that many salinas have been aban-
doned during the las1 decades.
We strongly recommend al1 efforts to maintain the inte-
resting salinas in cooperation with the exploiting companies.
I t is to be hoped that salinas will remain along the Spanish
coast for future generations.
ACKNOWLEDGEMENTS
We are rnost thankful to Dr. Mariona Hernández (Dept. of
Natural Products, Faculty of Pharmacy, University of Barce- lona) for help in determination of the cyanobacterium Phor- mi~lillnz i~cildri-iarutm. We are indebted to Dr. Francisco Rodríguez-Valera for valuable discussions. Mr. Agustín

Codina, Mr. Antonio Albalata (UNION SALINERA DE ESPANA, S.A.) and Mr. Miguel Cuervo-Arango (Salinas Bras del Port) informed us on the actual exploitation of the Spanish salinas (data used in Figure 1 ) and granted permis- sion for studying the salinas in Santa Pola.
Rutger de Wit has been supported by grants from the Commission of the European Community (STEP Programme) and the European Environmental Research Organization (E.E.R.O.).
REFERENCES
ALBAIGÉS, J., ALGABA, J. & GRIMALT, J.O. 1984. Extractable and bound neutral lipids in some lacustrine sedi- ments. Org. Geoc,hc.m. 6: 223-236
BARBÉ, A,, GRIMALT, J.o., PUEYO, J.J. & ALBAI-
GÉS, J. (1990). Characterization of model evaporitic envi- ronments through the study of lipid components. OI'S.
Geochem. 16: 815-828 BLATT, M., MIDDLETON, G. & MURRAY, R. 1980.
Origin of'sedimentary roc,ks. 2nd Ed. Pentice-Hall, Engle- wood Cliffs, N.S.
BOON, J.J. & DE LEEUW, J.W. 1987. Organic geoche- mical aspects of cyanobacterial mats. In: P. Fay & C. Van Balen (Eds.) The Cyanohac,teriu: 471-492. Elsevier,
Amsterdam BRITTON, R.H. & JOHNSON, A.R. (1987). An ecologi-
cal account of a Mediterranean Salina: The Salin de Giraud, Camargue (S. France). Biological Conseriation 42: 185-230
CAUMETTE, P.. BAULAIGUE, R. & MATERON, R. 1988. Characterization of Chromutium .salesigens sp. nov., a halophilic Chromatiaceae isolated from Mediterranean Sali- nas. System Appl. Microhiol. 10: 284-292
CAUMETTE, P., BAULAIGUE, R. & MATERON, R. 1991. Thio(,upila hulophilu sp. nov., a new halophilic photo- trophic purple sulfur bacterium. Arch. Microhiol. 155: 170-176
COHEN, Y. & ROSENBERG, E. Microbial Mats; Physio- logical Ecology of Benthic Microbial Communities. A.S.M., Washington D.C.
CORNÉE A. 1984. Étude préliminaire des bacté- ries des saumures et des sédiments des salins de Santa Pola (Espagne). Comparison avec les marais salants de Salin-de Giraud (Sud de la France). Kev. Ini,est. Geol. 38- 39: 109-122
DRONKERT, H. 1985. Eixapor-ite models and .sedimento- logy oj' Mes.viun and i.ec,c2nt e~~upor i t e s . Ph.D. Thesis. University of Amsterdam.
ENGLINTON. G. & CALVIN. M. 1967. Chemical fo\\il\. Sr.; \ I I / 216: 7 2 - J
ESTEVE. l . , M. MARTINEZ, J. MIR & R. GUERRERO, 1992. Distribution, typology and structure of inicrobial mat communities in Spain. Preliminary studies. Lim17etic.a 8.
CiELi'I. t.. SC'1iNLII)EK. t i . . M A N h , J . bC ORO. J . 1970.
Hydrocarbons of geocheinical significace in microscopic algae. Phytoc,henii.str.y 9: 603-6 12
GRANT, W.D. & LARSEN, H. 1989. Extremely Iialophi- lic archaeobacteria. Order Halobacteriales Ord. Nov. Itz: J.C. Holt (Ed.). Bei.,qey's Man~ccll (?f'Systcmutic Bac.ter.iology. Vol
3: 2216-2236 GRIMALT, J.O., DE WIT, R., TEIXIDOR, P. & AL-
BAIGÉS, J. (in press). Lipid biogeochemistry of Phnrmi-
dium and Mic~roc~o1eu.s mats. Adv. of Org. Geochem. Proc..
of'XV Irrt. M e e t i n ~ o11 O r ~ a n i c G~ochcnrisri-y, Manches- ter. 100 l
GUERRERO, M. C. & DE WIT, R. 1992. Microbial mats in the inland saline lakes of Spain. Limneticu 8.
JAVOK. B . 1989. t / \ . /~c i . \~~ / / r~ ( , I.i/i.i~-o~itrr<~r~\. rMlc.i.ohio- lo,yy i.l.117i.l.l Biogeoc~hcn/i.stt:),. Springcr, Berlin.
KARL, D.M. 1986. Dctermination of in situ microbial biomass, viability, metabolism and growth. In: J.S. Poin- dexter & E.R. Leadbetter (Eds.) Ba(.teria itz Nciture. Vol. 2. Methods atid Spe(,iul App1ic.ation.s in Ba<~teria/ Ecology: 85- 176. Plenum Press. New York.
LARSEN, M., MACK, E.E. & PIERSON, B.K. 1991. Mesophilic C/zlot~oflexu,v -1ike organisms from marine and hypersaline environments. Ahstr. VI / . Int. Symp. Photosynr- hetic Pi~okulyotes: 169.
LEFOND, S.J. 1963. Hundhook of w~orld sult resoui.c~cJs. Plenum Press, New York.
NOEL, D. 1984. Les diatornées des saumures et des sédi- ments de surface du Salin de Bras del Port (Santa Pola province d'Alicante, Espagne). Rev. Inivst. Geol. 38-39: 79-1 07
PARKES, R.J. 1987. Analysis of microbial communities within sediments using biomarkers. Iti: M. Fletcher, T.R.G. Gray & J.G. Jones (Eds.) Ecology :~'?f'mic~rohiul c,omm~lnitie.s.
Society for General Microbiology, London. POISBEAU-HÉMERY, J. (Ed.), 1980. Marais .sulant.s.
Connui.ssanc,e des Richesses tiatur~el1e.s de la Loire-Atlantiq~~c. Société des Sciences naturelles de I'Ouest de la France, Nantes.
RATLEDGE, C. & WILKINSON, S.G. (Eds.), 1988. Microbial lipids. Academic Press, London
RODR~GUEZ-VALERA, F., R U I Z BERRAQUERO, F. & CORMENZANA, R. 198 1. Characteristics of the hetero- trophic bacteria1 populations in hypersaline environments of different sal1 concentrations. Mirr-ohiol. Ecol. 7: 235-243

2 12
RODRÍGUEZ-VALERA, F., VENTOSA, A,, JUEZ, G. & IMHOFF, J.F. 1985. Variation of environmental features and microbial populations with salt concentration in a multi- pond saltern. Microbiol. Ecol. 11: 107-1 15
TAYLOR, J. & PARKES, R.J. 1985. Identifying different populations of sulphate-reducing bacteria within marine sediment systems, using fatty acid biomarkers. J. Gen.
Microbiol. 131: 63 1-642 TEIXIDOR, P., PUEYO, J.J., RODRÍGUEZ-VALERA, F.
& GRIMALT, J.O. 1991. Alkylglycerol diethers in recent and ancient evaporites. In: D.A.C. Manning (Ed.). Organic Geochemistry. Advances and applications in the natural envi- ronment: 563-565. Manchester University Press, Manchester
TEN HAVE, H.L., DE LEEUW, J.W., SINNINGHE DAMSTE, J.S., SCHENK, P.A., PALMER, S.E. & ZUM-
BERGE, J.E. 1988. Application of biological markers in the recognition of palaeohypersaline environments. In: A.J. Fleets, K . Kelts, & M.R. Talbot (Eds.). Lacustrine Petro- leum Source Rocks: 123-130. Geological Society Special Publication NVO.
THOMAS, J.C. 1984. Formations benthiques 2 Cyano- bacteries des salins de Santa Pola (Espagne): composition spécifique, morphologie et caractéristiques biologiques des principaux peuplements. Rev. Invest. Geol. 38-39: 139-158
USIGLIO, J. 1848. Etudes sur la composition de I'eau de la Méditerranée et sur I'exploitation des sels qu'elle contient. Annales des Chem. Phys. 3d. Ser., Vol. XXVZZ: 172-191.
VESTAL, J.R. & WHITE, D.C. (1989). Lipid analysis in Microbial Ecology. Bioscience 39: 535-541.

FILAMENTOUS GREEN ALGAE OF SPAIN: DISTRIBUTION AND ECOLOGY
J. C a m b r a l and M. Aboa12 1. Departamento de Biología Vegetal. Facultad de Biología. Universidad de Barcelona. Avgda. Diagonal, 645. Barcelona
08028. Spain. 2. Departamento de Biología Vegetal (Botánica). Facultad de Biología. Universidad de Murcia. Campus de Espinardo. Murcia
30 100. Spain.
Keywords: Filament, Green Algae, Flora, Distribution, Ecology, Spain.
ABSTRACT
A survey on the studies of filarnentous green algae in Spain is presented. The main part of the information was produced by Margalef in the 50's. Recently, there is an increasing interest on filarnentous green algae. Data on distribution and ecology are compiled.
INTRODUCTION
The relative importance of macrophytes in benthic communities is much more higher in freshwater than in marine habitats dominated by macroalgae. In epicontinental waters benthic communities are composed mainly of micro- algae in close relationship with phanerogamic macrophytes.
Most of the microscopic benthos of freshwater habitats have such a passive role like plankton but they have redu- ced their losses by sedimentation to zones 8Utside the photic zone. In lakes and especially small ponds, primary produc- tion of littoral and benthic algae is twice that of plankton one and the production of macrophytes is higher than both together (MARGALEF, 1983);
Filamentous green algae constitute an important compo- nent of algal communities in freshwater habitats. Neverthe- less, their enormous morphological plasticity and the diffi- culty of observing some features have complicated their taxonomy. These and other facts explain the scarcity of data on their ecology.
On a worldwide scale there is a lack of information as regards their geographical distribution and it seems neces- sary to undertake a taxon~mical review of some of these algal groups.
The filament is an excellent adaptative form: the plant grows rapidly in length and can use new volumes of water maintaining the area/volume ratio constant. Branches repre-
sent a variation which permit the algae a better use of water in confined spaces and a better control of absorption (MARGALEF, 1983).
A sucession from simple filaments with fast growth to bran- ched filaments with a slower growth can be observed very frequetltly in freshwater ecosystems (MARGALEF, 1983).
FLORA. DISTRIBUTION.
The most important contribution to our knotvledge of Spanish algal flora was made 84. ALLORGE & ALLORGE (1930), ALLORGE & MANGUIN (1941), BESCANSA (1907, 1908), CABALLERO (1942, 1945, 1947, 1950),
GONZALEZ GUERRERO (1927, 193 1 , 1940a,b, 1942, 1946, 1953, 1960), LEWIN (1888) and MARGALEF (1944,1946a y b, 1947, 1948, 1949a y b, 1950, 1951, 1952a y b, 1953, 1955, 1956, 1957).
A total of 431 filamentous green algae taxa were recor-
ded (ALVAREZ COBELAS, 1983; ALVAREZ COBELAS & GALLARDO, 1986; CAMBRA, 1985).
More recently new reports were presented (CAMBRA, 1987, 1988, 1989 a y b; SABATER, 1989; ABOAL, 1988 a y b, 1991, 1992).
The most frequently cited genus was Oedogoniurn (fig. 1) followed by Spirogyra and Mougeotia and the most impor- tant order is Zygnernatules (fig. 2).
Lininetica, 8: 2 13-220 (1992) 8 Asociación Espenola de Limnología, Madrid. Spain
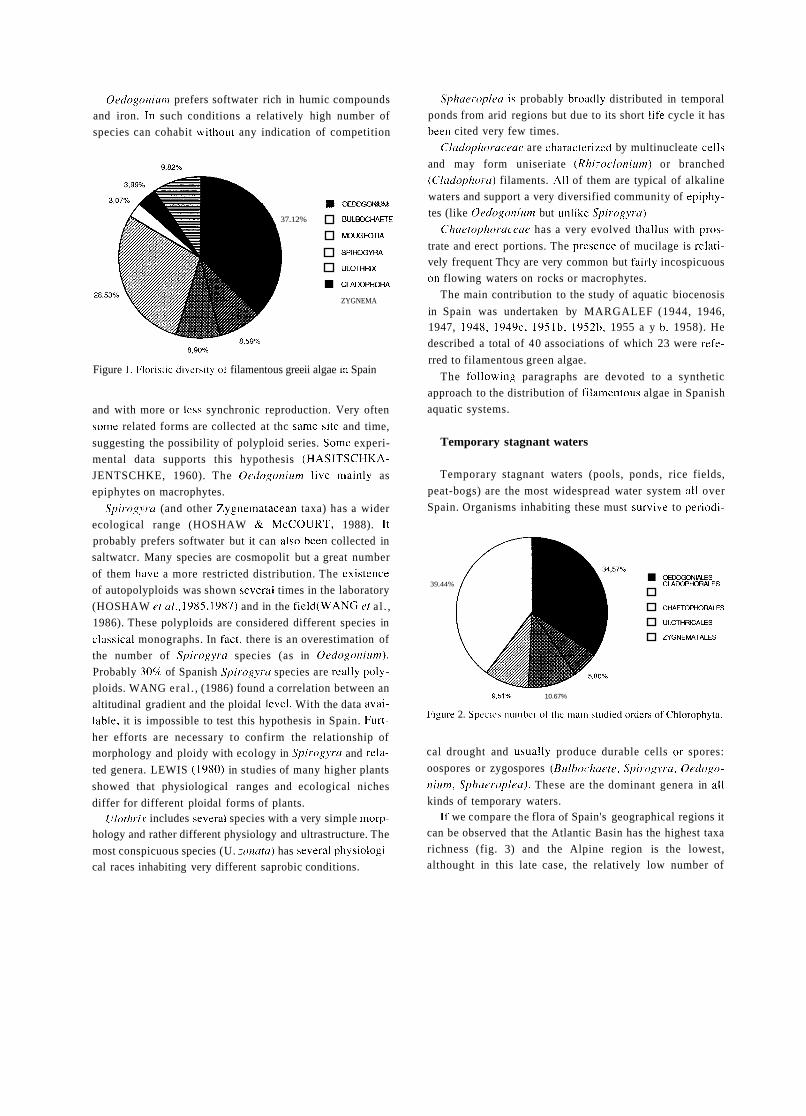
Oedogoniirnr prefers softwater rich in humic compounds
and iron. In such conditions a relatively high number of
species can cohabit without any indication of competition
m- 37.12% BULBOCHAETE
MOUGEOTIA
SPIROGYRA
ULOTHRIX
ClrnPHOFlA
ZYGNEMA
8,59% 8,9070
Figure 1 . Floristic tlivei-sity of filamentous greeii algae in Spain
and with more or less synchronic reproduction. Very often
some related forms are collected at thc same site and time,
suggesting the possibility of polyploid series. Some experi-
mental data supports this hypothesis (HASITSCHKA-
JENTSCHKE, 1960). The Oedogot~ilrnl live mainly as
epiphytes on macrophytes.
Spiroggrrr (and other Zygneinatacean taxa) has a wider
ecological range (HOSHAW & McCOURT, 1988). It
probably prefers softwater but it can also been collected in
saltwatcr. Many species are cosmopolit but a great number
of them have a more restricted distribution. The existence
of autopolyploids was shown several times in the laboratory
(HOSHAW et a1.,1985,1987) and in the field(WANG er a l . ,
1986). These polyploids are considered different species in
classical monographs. In fact, there is an overestimation of
the number of Spirogyra species (as in Oedogoriiuvri).
Probably 30% of Spanish Spirogyru species are really poly-
ploids. WANG eral . , (1986) found a correlation between an
altitudinal gradient and the ploidal level. With the data avai-
lable, it is impossible to test this hypothesis in Spain. Furt-
her efforts are necessary to confirm the relationship of morphology and ploidy with ecology in Spirogyra and rela-
ted genera. LEWIS (1980) in studies of many higher plants
showed that physiological ranges and ecological niches
differ for different ploidal forms of plants. Ulorkrir includes several species with a very simple Inorp-
hology and rather different physiology and ultrastructure. The
most conspicuous species (U. zonatu) has several physiologi- cal races inhabiting very different saprobic conditions.
Sphcre/.oplcu is probably broadly distributed in temporal
ponds from arid regions but due to its short life cycle it has been cited very few times.
C ludop l io r~c~~uc . are characteri~ed by multinucleate cells
and may form uniseriate (Khi:oc.lonium) or branched
(Clatlo/?liorcr) filaments. All of them are typical of alkaline
waters and support a very diversified community of epiphy-
tes (like Ocn'o,goniilnz but unlike Spirogyru)
Choetophorcrc,eue has a very evolved thallus with pros-
trate and erect portions. The presence of mucilage is relati- vely frequent Thcy are very common but fairly incospicuous
on flowing waters on rocks or macrophytes.
The main contribution to the study of aquatic biocenosis
in Spain was undertaken by MARGALEF (1944, 1946, 1947, 1948, 1949c, 1951b, 1952b, 1955 a y b, 1958). He
described a total of 40 associations of which 23 were refe-
rred to filamentous green algae.
The following paragraphs are devoted to a synthetic
approach to the distribution of filainentous algae in Spanish
aquatic systems.
Temporary stagnant waters
Temporary stagnant waters (pools, ponds, rice fields,
peat-bogs) are the most widespread water system al1 over
Spain. Organisms inhabiting these must survive to periodi-
cxzcCa4IALEs 39.44% ClADoPHoRnLES
CHAETOPHORALES
ULOTHRICALES
ZYGNEMATALES
9,51% 10.67%
Figiire 2. Specic.5 i i~ i i i ibc . i ol ~lic i i i ~ i i i i \tiidied orders of Chlorophyta.
cal drought and usually produce durable cells os spores:
oospores or zygospores (Bulhoc.Ciuete, Spirogyra, Oedogo-
nilrwi, Sphaeroplea). These are the dominant genera in al1
kinds of temporary waters. I i we compare the flora of Spain's geographical regions it
can be observed that the Atlantic Basin has the highest taxa
richness (fig. 3) and the Alpine region is the lowest, althought in this late case, the relatively low number of

species recorded is related with the scarcity of data and the
difficulty in finding fructified inaterial(indispensable for
species identification).
In the Atlantic systems (fig. 4), Oerlogoniuc,c~rrc are the
doininant group: Oeclogor~iun~ and Btrlhoc~liuc~te together
represent 63% of the total number of taxa. In these systems,
Oedogoniuc~eue are accompanied by many Trihor~eniu and
Desmids species aild constitute a community which is very
frequent in Northern Europe (MARGALEF, 1955).
The Mediterranean freshwater systems (fig. 4) are domi-
nated by Zy~rienicrtrrc.erre (73%).
The algal corrirnunities of endorrheic lagoons are similar
to the Mediterranean communities, with a great number of
Zygric~rnutuc~eae. However, the relative proportion of
C1cruc.hc~r.icr species is higher.
In alpine teniporary waters the percentage of Zj,qtrc~nlutcr-
c,ecrt. (¡.e. figo,yoniitwi er.ic.etorurn) is slightly higher than
that of Oedo,goniuc,enr but they are usually sterile.
Ulorhi.ic.hu1e.s are well represented in peat-bogs and acid
waters (¡.e. Birruclerrr.ia tcJc.ror.um, Chlorhor.rnidirrnr c.r.cnrrlo-
tirrn. C.wlucositn7. Mic~1.o.spor.a pachid¿,r.rncr. M.an~oer~u) but
much more floristic information is required.
Rice fields are located on the Eastern coast of Spain.
Important alga1 growths of S[~huerop/eu unnrr/inu, alinost
completely covering the water surface can be observed at
the beginning of the cuitivation. Hydrodir,tyori i~etic~irlrrritrn
rnay develop a thallus of several meters. Later on, befnre
harvest, Clado/~horrr masses became dominant.
In more permanent waters, which do not dry up in
summer, the filamentous masses of Oedogoniitn~. Spirogyrcr
and others are replaced by Cludophora,fi-ac,tcr and C.,qlorne-
r.attr i*,c~r~as.sior. mats, in mesotrophic and eutrophic waters
(MARGALEF, 1983). In this more stable situation, the
macroalgal filaments are covered by many Chaetophoralean
epiphytes (Aphunochaete repens, Colt~oc.licrere s(.utcrtcr.
Gloeoplcrv ~ ' e h c ~ r i , Protoderr~ru i<iride).
In brackish or saline waters (endorrheic os coastal lago-
ons), algal mats of Rliiroc~loriii<m Iziel.ogIjphic,un~, Clicreto-
nioipha sp.p/.. Cludophorcr ,ylonzer.crta iar~.cru,s.sioi~. C..sc.i,i-
(,ea, C.i*a,qah~tnclu are dominant (fig.4). ~gnc~nicr/crlr.s and
Oedocqonicrle.s are not frequent (i.e.Oedo~orlilrn7 c,ripilli-
for.nie, S/~ii.ogyr.u rnorrserratii. S..sirh.sul.su. S.n~ic~r~o~~irr~ctcrtu).
Flowing waters
In flowing waters the most widespread species is Cluilop- 11or.u ,qloniel.urcr, especially in mesotrophic alkaline waters
(MARGALEF, 1983), although it can be collected in a very
ATLANTIC
ALPINE
51,49% ~NDORREIC
MEDITERRANEAN
Fig~11.e 3. Species tilirnbct- of filainentou\ algae in several biogeo graphic areas of Spain.
wide ecological range from oligotrophic to heavily organic
pollutcd waters (ABOAL, 1989a; SABATER, 1987). Seve-
sal filamentous green microalgae can live on C1adophor.u
cells (Apharroc~licrc~r~ [)olyc.liuctc~. Colcoc~hac~e o/~hic~uluris.
Mi(,r.othanini1rn7 .str.i<.ti.s.siniirm, iJr.otr~rnu c~or~feri~ic,ir l~tn~ or.
Pr.orocler.nicr i.iride). Other groups such as Clicretoplioi.tz1e.s and Ulotkr~ic~hule.~
can develop important growths depending on the alkalinity
values and water nutrient content (ABOAL. 1987: SABA-
TER, 1982, 1987).
Drcrpm~riuldiu ,qlornc~r.ura and D.nrirtirhi1i.s frequently
cover the aquatic phanerogams or rocks in the littoral zone
of slowly flowing streams with relatively low conductivity
and low level of nutrients (ABOAL, 1987, 1989).
Attached to the aquatic phanerogarn roots several species
of Sti,q~oc~lor~irrm can he found in alkaline flowing walers,
from oligo-mesotrophic (S.lori,qi/~ilirn~, S.nuti~tn1. S.suhsr~-
c~~rriel~rnr, S. i~crr.iuhilt~) to heavily polluted (S.teri~rc).
The hemispherical lobuiated thalli of Chuc~tophorcr
species cover rocks or rnacrophytes in several flow condi-
tions but always in pure alkaline water: Ck.,qlornc~ratu is
rnainly epiphytic and Ch.irrc~ru,s.sutu is epilithic. Both can
develop a very hard encr~istation. Sorrie of these filamentous algae strongly adhered to
rocks and are usually intensely encrusted. They constitute a
very important and very inconspicuous portion of benthic
cornmunities (Gorigr.osir-cr, Chloroc.loniun7). Species of
Gorr~r~o.sir~a can inhabit both saltwater and freshwater stre-
ams (G.deho~yciria, G.iric~r.ustans) and can even grow on macroalgae such as Clodoplzor.c~ (G.sc~otri;fi'eldiiJ. The only
known species of Chlor.otyli~rn~ (Cl~.c~ar«r~ac~rui~irniJ is characteristic of pure fast flowing waters al1 around the
world (ABOAL, 1989c).
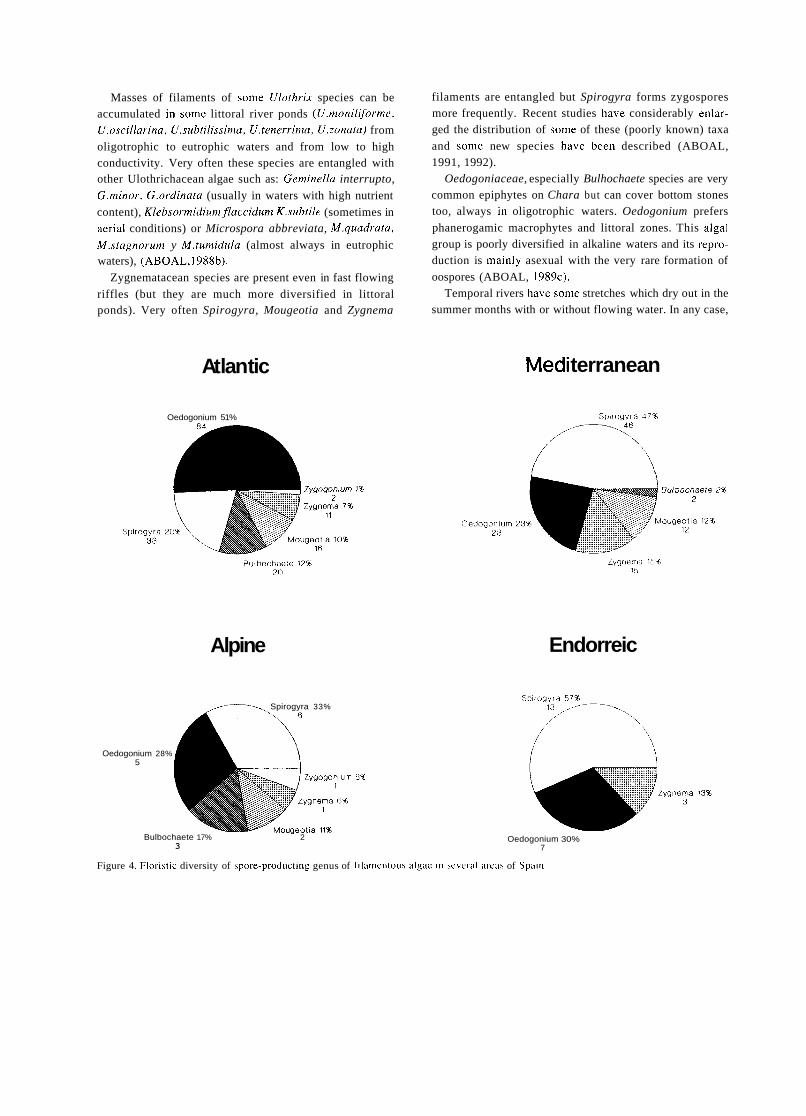
Masses of filaments of some Ulothrix species can be accumulated in some littoral river ponds (U.monillforme, U.oscillarina, U.subtilissima, U.tenerrima, U.zonata) from
oligotrophic to eutrophic waters and from low to high conductivity. Very often these species are entangled with other Ulothrichacean algae such as: Geminella interrupto,
G.minor, G.ordinata (usually in waters with high nutrient
content), Klebsormidium flaccidum K.subtile (sometimes in aerial conditions) or Microspora abbreviata, M.quadrata,
M.stagnorum y M.tumidula (almost always in eutrophic waters), (ABOAL,1988b).
Zygnematacean species are present even in fast flowing
riffles (but they are much more diversified in littoral ponds). Very often Spirogyra, Mougeotia and Zygnema
At lan tic
Oedogonium 51%
Alpine
Spirogyra 33%
Oedogonium 28% 5
zygogonium 8%
Bulbochaete 17% 2 3
filaments are entangled but Spirogyra forms zygospores more frequently. Recent studies have considerably enlar- ged the distribution of some of these (poorly known) taxa
and some new species have been described (ABOAL, 1991, 1992).
Oedogoniaceae, especially Bulhochaete species are very common epiphytes on Chara but can cover bottom stones too, always in oligotrophic waters. Oedogonium prefers
phanerogamic macrophytes and littoral zones. This alga1 group is poorly diversified in alkaline waters and its repro- duction is mainly asexual with the very rare formation of
oospores (ABOAL, 1989~) . Temporal rivers have some stretches which dry out in the
summer months with or without flowing water. In any case,
Medi terranean
Endorreic
Oedogonium 30% 7
Figure 4. Floristic diversity of spore-producting genus of filarnentous algae in ~everül at-ens of Spaiti
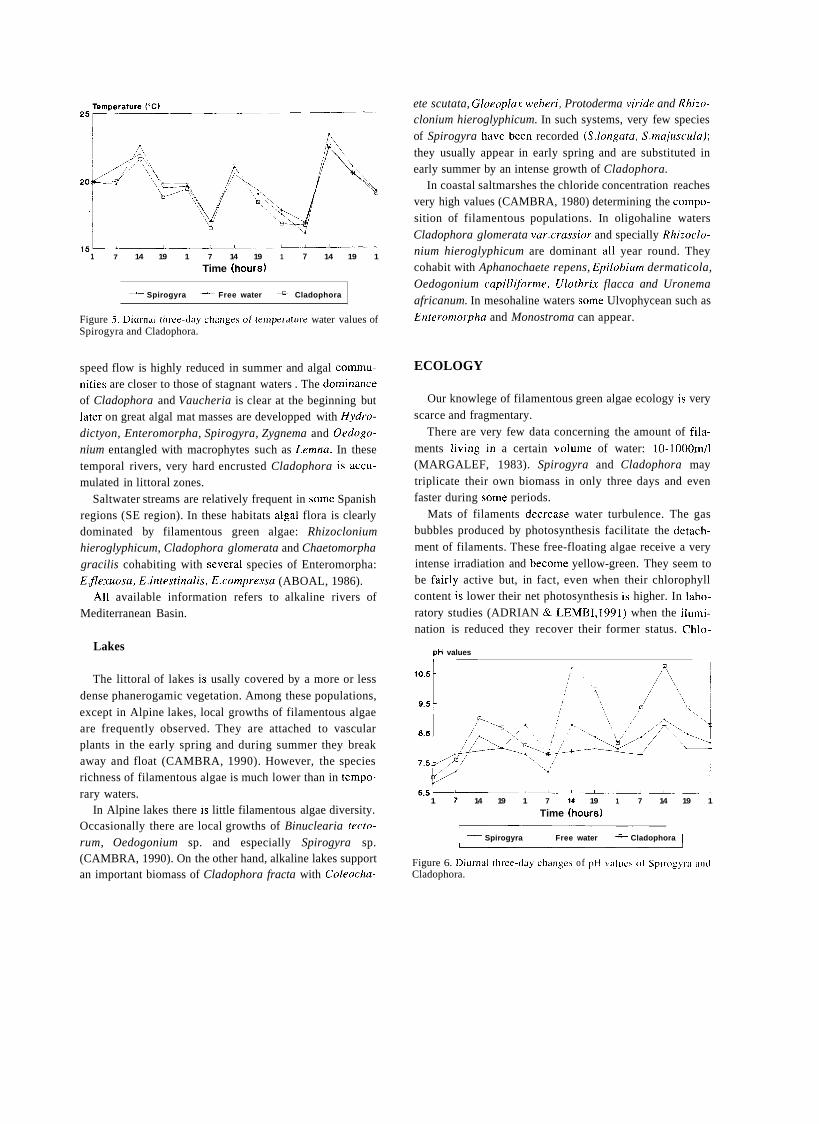
1 5 ' ' ' ' 1 ' 1 7 14 19 1 7 14 19 1 7 14 19 1
Time (hours)
l - Spirogyra - Free water -Et Cladophora
Figure 5. Diurnal ihi-ee-day ct~aiigeh of teinpei-atui-e water values of Spirogyra and Cladophora.
speed flow is highly reduced in summer and algal commu- nities are closer to those of stagnant waters . The dominance of Cladophora and Vaucheria is clear at the beginning but later on great algal mat masses are developped with Hydro- dictyon, Enteromorpha, Spirogyra, Zygnema and Oedogo- nium entangled with macrophytes such as Lemna. In these temporal rivers, very hard encrusted Cladophora is accu- mulated in littoral zones.
Saltwater streams are relatively frequent in some Spanish regions (SE region). In these habitats algal flora is clearly dominated by filamentous green algae: Rhizoclonium hieroglyphicum, Cladophora glomerata and Chaetomorpha gracilis cohabiting with severa1 species of Enteromorpha: E.flexuosa, E.intestinalis, E.compressa (ABOAL, 1986).
Al1 available information refers to alkaline rivers of Mediterranean Basin.
Lakes
The littoral of lakes is usally covered by a more or less dense phanerogamic vegetation. Among these populations, except in Alpine lakes, local growths of filamentous algae are frequently observed. They are attached to vascular plants in the early spring and during summer they break away and float (CAMBRA, 1990). However, the species richness of filamentous algae is much lower than in tempo- rary waters.
In Alpine lakes there is little filamentous algae diversity. Occasionally there are local growths of Binuclearia tecto- rum, Oedogonium sp. and especially Spirogyra sp. (CAMBRA, 1990). On the other hand, alkaline lakes support an important biomass of Cladophora fracta with Coleocha-
ete scutata, Gloeoplax weheri, Protoderma vir-ide and Rhizo- clonium hieroglyphicum. In such systems, very few species of Spirogyra have been recorded (S.longata, S.majuscu/a); they usually appear in early spring and are substituted in early summer by an intense growth of Cladophora.
In coastal saltmarshes the chloride concentration reaches very high values (CAMBRA, 1980) determining the compo- sition of filamentous populations. In oligohaline waters Cladophora glomerata var.crassior and specially Rhizoclo- nium hieroglyphicum are dominant al1 year round. They cohabit with Aphanochaete repens, Epilohium dermaticola, Oedogonium capilliforme, Ulothris flacca and Uronema africanum. In mesohaline waters some Ulvophycean such as Enteromorpha and Monostroma can appear.
ECOLOGY
Our knowlege of filamentous green algae ecology is very scarce and fragmentary.
There are very few data concerning the amount of fila- ments living in a certain volume of water: 10-1000m/l (MARGALEF, 1983). Spirogyra and Cladophora may triplicate their own biomass in only three days and even faster during some periods.
Mats of filaments decrease water turbulence. The gas bubbles produced by photosynthesis facilitate the detach- ment of filaments. These free-floating algae receive a very intense irradiation and become yellow-green. They seem to be fairly active but, in fact, even when their chlorophyll content is lower their net photosynthesis is higher. In labo- ratory studies (ADRIAN & LEMBI,1991) when the ilumi- nation is reduced they recover their former status. Chlo-
PH values 7
6 . 5 1 " 1 ' 1 1 1 4 1 1 1 1 1 7 14 19 1 7 14 19 1 7 14 19 1
Time (hours)
- Spirogyra + Free water + Cladophora (
Figure 6. Diurtial three-day changea of pH vnlue\ of Spirogy1-;I ;iii<1
Cladophora.
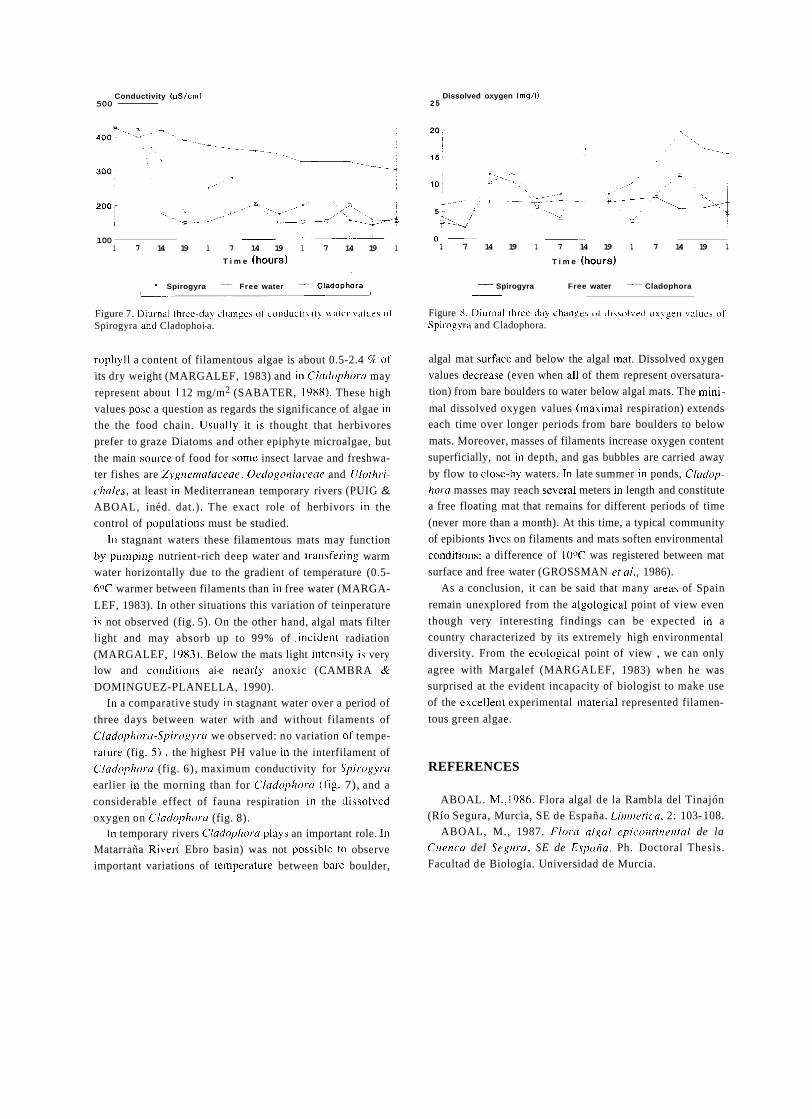
Conductivity ( ~ S l c r n ) 500 - - Dissolved oxygen (mg/l)
2 5 -- -
100 - - A -
l 7 14 19 l 7 14 19 l 7 14 19 l
T i m e (hours)
- Spirogyra - Free water - Cladophora
Figure 7. Di~iriinl iliree-tlay cliaiigr\ o i coii(luctivii) unte¡- vnlue\ ot Spirogyra and Cladophoi-a.
rophyll a content of filamentous algae is about 0.5-2.4 %, of
its dry weight (MARGALEF, 1983) and in C1crdol1hor.a may
represent about 1 12 mg/m2 (SABATER, 1988). These high
values pose a question as regards the significance of algae in
the the food chain. Usually it is thought that herbivores
prefer to graze Diatoms and other epiphyte microalgae, but
the main source of food for some insect larvae and freshwa-
ter fishes are Zy,qnematcrceae, 0edogoniac.eae and Ulothr-i-
c.lzulc~s, at least in Mediterranean temporary rivers (PUIG & ABOAL, inéd. dat.). The exact role of herbivors in the
control of populations must be studied.
In stagnant waters these filamentous mats may function
by pumping nutrient-rich deep water and transfering warm
water horizontally due to the gradient of temperature (0.5-
60C warmer between filaments than in free water (MARGA-
LEF, 1983). In other situations this variation of teinperature
is not observed (fig. 5). On the other hand, algal mats filter
light and may absorb up to 99% of ,incident radiation
(MARGALEF, 1983). Below the mats light intensity is very low and coridilioiis ai-e riearly anoxic (CAMBRA &
DOMINGUEZ-PLANELLA, 1990). In a comparative study in stagnant water over a period of
three days between water with and without filaments of
C1aciopho1.a-Sl,ir-otqy~~u we observed: no variation of tempe-
sature (fig. S ) , the highest PH value in the interfilament of
Cladophor-a (fig. 6), maximum conductivity for Spir-ogyr.cr
earlier in the morning than for Cludo/7ho1.u (fig. 7), and a
considerable effect of fauna respiration in the ciissolved
oxygen on C1adophor.u (fig. 8). In temporary rivers Clndophor-o plays an important role. In
Matarraña River( Ebro basin) was not possible to observe
important variations of temperature between bare boulder,
0 -- 1 7 14 19 1 7 14 19 1 7 14 19 1
T i m e (hours)
- Spirogyra Free water -+ Cladophora -- J
Figure 8. Diurii:il ilircc-day cliaiiges ot tli\\ol\rd ox)geii valueh of Spirogyra and Cladophora.
algal mat surface and below the algal mar. Dissolved oxygen
values decrease (even when al1 of them represent oversatura-
tion) from bare boulders to water below algal mats. The mini-
mal dissolved oxygen values (maximal respiration) extends each time over longer periods from bare boulders to below
mats. Moreover, masses of filaments increase oxygen content
superficially, not in depth, and gas bubbles are carried away
by flow to close-by waters. In late summer in ponds, Cladop-
hoi.a masses may reach severa1 meters in length and constitute a free floating mat that remains for different periods of time
(never more than a month). At this time, a typical community
of epibionts lives on filaments and mats soften environmental
condjtions: a difference of 1O0C was registered between mat
surface and free water (GROSSMAN er u/., 1986).
As a conclusion, it can be said that many areas of Spain
remain unexplored from the algological point of view even
though very interesting findings can be expected in a
country characterized by its extremely high environmental diversity. From the ecological point of view , we can only
agree with Margalef (MARGALEF, 1983) when he was surprised at the evident incapacity of biologist to make use
of the excellent experimental material represented filamen-
tous green algae.
REFERENCES
ABOAL. M.,1986. Flora algal de la Rambla del Tinajón (Río Segura, Murcia, SE de España. Limneticu, 2: 103- 108.
ABOAL, M., 1987. F1ol.a ulgul epicontirlenral de la Cuenc.u del Seg~o.a, SE de España. Ph. Doctoral Thesis.
Facultad de Biología. Universidad de Murcia.

ABOAL, M., 1988a. Zygnen~ataceae (Co?zj~rgales, Chlo- rophycecre) of the River Segura Basin, Southeastern Spain. Noi'a Hedil,igia, 47(3-4): 389-402.
ABOAL. M., 1988b. Aportación al conocimiento de las algas del SE de España.VI1. Clorofíceas (Chloi.ophyc,eae Wille in Warming 1884). Carzdollea, 43: 521-548.
ABOAL, M., 1989a. Los poblamientos algales bentóni- cos del Río Segura y su relación con la calidad del agua.
Carzdollea, 44: 199-208. ABOAL, M., 1989b. Flora algal del Río Benamor
(Cuenca del Segura, SE de España). Limnetica, 5: 1 - 1 1. ABOAL, M., 1989c. Epilithic algal communities from the
River Segura Basin, Southeastern Spain. Arch. Hydrohiol.,
106 (1): 113-124. ABOAL, M., 1991. New records of Zygnemataceae
(Zygnernales, Chloroplzyceae) from Southeastern Spain. Noila Hedwigia, 52 (3-4): 365-373.
ABOAL, M., 1992. A new species of Zygizenza (Zygne- matales,) from Southeastern Spain. Algological Studies, 65. (in press).
ADRIAN, H. L. & C. A. LEMBI., 1991. Effects of light and temperature on growth, photosynthesis and chlorophyll content of Spir.o,gy.a. 4th Inrei.narionci1 Phycologic,al ~ l l / l ~ l ~ f ' \ \ . l ~ ~ l l ~ l l ~ l l l l . \or1I1 r:ll~olill:l. ( ' S . ' .
ALVAREZ COBELAS, M.,1984. Catálogo de las algas continentales españolas. 111. Zygophyceae Widder, 1960. Collect Bot., 15: 17-37.
ALLOKCil:. V . b: 1'. .ALLOKGE.. 1930. Hiiéroco~ites, Euchlorophycées et Conjuguées de Galice. Re,. Algol., 5: 327-382.
ALLORGE, P. & E. MANGUIN.,1941. Algues d'eau douce (IL>\ P!,~CIIC!C\ I ~ : I \ ~ L I L ~ \ . Ij1111 SO¡ I ~ I I I / F I . ( / / I ~ 1 ~ . SS: l $0- 101
ALVAREZ COBELAS, M. & T. GALLARDO, 1986. Catálogo de las algas continentales españolas. IV. Chlo- rophyceae Wille in Warming 1884, Prasinophyceae T. Christensen ex Silva 1980. Acta. Bot. Malacitana, 11: 17-38.
UL:SC'XhS,A. t... 1'107. ,-\lg~iii;i\ coiij~igacla\ tlC I;i pi-o\ 1 1 1 -
cia de Orense. Bol. R. Soc. Esp. Hist. Nar., 7 : 65-68. BESCANSA, F., 1908. Conjugadas para la flora de Gali-
cia. Bol. R. Soc. Esp. Hist. Nat., 8: 234-238. CABALLERO, F., 1942. Contribución al conocimiento de
la flora algológica de España. An. J. Bot. Madrid, 3: 299-320. CABALLERO, F., 1945. Algas de las aguas de Santi-
llana. An. .T. Bor. Madrid, 5 (2): 345-364. CABALLERO, F. ,1947. Una Mougeotia nueva de
Guadalupe. An. J. Bot. Madrid, 8: 285-295. CABALLERO, F., 1950. Algas de Gredos. An. Irzsf. Bot.
Cai~anilles, 10 (1): 23 1-260.
CAMBRA, J.,1985. Catalegs de les especies de cianofi- cies i d'algues continentals y del plancton mari dels Paisos Catalans. Annex 1: 501-522. In: Historia Nar~rrcrl dels PaiSos Catulans. Plantes Inferiors. X . LLIMONA (ED.). ENCICLOPEDIA CATALANA S.A.
CAMBRA, J., 1987. Flore et végétation algologiques des eaux épicontinentales de la Réserve Naturelle des Vallées d'Ordesa (Pyrénées Aragonaises). Cundollea, 42: 475-490.
CAMBRA, J., 1989a. Fresh water algae from the Cape of Creus Peninsula, Catalonia (NE Spain). Nova Hedwigia, 49 (3-4): 281-321.
CAMBRA, J., 1989b. Contribución al conocimiento del género Oedogonium en España.1. Acta Bot. Mala<,ituna, 14: 20 1 -206.
CAMBRA, J.,1990. Estudi sobre les algues epifítiques en sisfemes larustres. Tesis Doctoral. Univ. Barcelona.
CAMBRA, J., 1991. Contribució a I'estudi de les algues epifítiques dels estanys de Banyoles, Basturs i Estanya. Orsis. 6: 27-44.
CAMBRA, J. & A. DOMINGUEZ-PLANELLA., 1990. Datos para el estudio de las algas filamentosas en arrozales de 1'Alt Empordi (Girona, NE de España). Scienfia G e ~ u n -
densis, 16(2): 43-53. GONZALEZ GUERRERO, P. ,1927. Contribución al
estudio de las algas y esquizófitas de España. Trah. Mus.
Ciencias, 22: 1-52. GONZALEZ GUERRERO, P., 193 1. Algunos datos algo-
lógicos de la Península Ibérica, Baleares y Marruecos. Bol. R. Soc. Esp. Hist. Nat., 31: 633-639.
GONZALEZ GUERRERO, P., 1940a. Observaciones ficobiológicas en aguas de Valencia. An. J. Bot. Madrid, 1 : 107-140.
GONZALEZ GUERRERO, P. , 1940b. Novedades bioló- gicas en algas de Cuenca. An. J. Bor. Madrid, 1 : 107-140.
GONZALEZ GUERRERO, P., 1942. Algas del norte y centro de España. An. .T. Bol. Madrid, 3: 269-278.
GONZALEZ GUERRERO, P., 1946. Algas invernales gaditanas. An. J. Bot. Madrid, 7: 433- 456.
GONZALEZ GUERRERO, P., 1953. Datos estivales de las algas pacenses. An. Inst. Bot. Cavanilles, 12: 5-76.
GONZALEZ GUERRERO, P., 1960. Algas de Aragón. An. Ins. Bot. Ca~xanilles, 18: 149.197.
GROSSMAN, G.; LOBON-CERVIA, J; PUIG, M. A. & A. de SOSTOA., 1986. Relaciones ecológico-amhien- tales de alg~cnas comunidades de peces del río Mara- rraña. Proyecto del Comité conjunto Hispano-Nortea- mericano de Cooperación científica y técnica. Annual Report 1986.

220
HASITSCHKA-JENSCHKE, G., 1960. Vergleichende Untersuchungen an haploiden und durch Colchicineinwir- kung dipioid gewordenen Stiimmen von Oedo~otii~lrii c~ardia(~unz. 0s terr . Bor. Zeirsch., 107: 194-21 1.
HOSHAW, R. W. & R. M. McCOURT., 1988. The Zy,qtiemuta(.eae (Chlorophyrcri: a twenty-year update of research. Phycologia, 27(4): 5 1 1-548.
HOSHAW, R. W.; WANG, J.C.; McCOURT, R. M. & H. M. HULL., 1985. Ploidal changes in clonal cultures of
Spirogyrcr conimunis and implications for species definition. Ame/.. .l. Bot., 72 (1): 1005-101 1.
HOSHAW, R. W., WELLS, C.V. & R. M. McCOURT.,
1987. A polyploid species complex in Spii.ogyra ma.vimcí
(Chloropkya, Zygneniataceae): a species with large chro- mosomes. .l. Phyc'ol., 25: 267-273.
LEWIN, M., 1888. Uber Spanische Süsswasseralgen. Bih.
Si~ensk. Vedrnsk. Akad. Harzdl. Aisdl., 14 ( 1 ): 1-24.
LEWIS. W. H., 1980. Polyploidy: hiolo~ic,al r.eIei.aticu. Plenum Press, New York.
MARGALEF, R. ,1944. Datos para la flora algológica de
nuestras aguas dulces. P14hl. Itist. Bot. Brir.celona, 4 ( l ) , 130 pp. MARGALEF, R., 1946a. Materiales para el estudio del
Lago de Bañolas (Gerona). P. Inst. Biol. ApI. 1 : 27-78. MARGALEF. R., 1946b. Contribución al conocimiento
hidrobiológico del País Vasco-Navarro. Aportación al estu-
dio de la fauna y flora vasco-navarra (Sierra de Aralar). Inst. Est. Pirenaicos, 44 pp.
MARGALEF, R., 1947. Materiales para una flora de las algas del NE de España. 1. Zy,qnematale,s. 1 vol.1, fasc. 11, n" 9: 1-21.
MARGALEF, R., 1948. Flora, fuutlu y c~omitriidacles hicíticas de las agilas del Pirineo de la Cer.claña. Inst. Est. Pirenaicos. 226 pp.
MARGALEF, R.,1949a. Datos para la hidrobiología de
la Sierra de Guadarrama. P. Inst. Biol. Apl., 6: 5-21. MARGALEF, R. , i949b. Materiales para una flora de las
algas del NE de España. 111. Euchlorophyceae. Coll. Bot., 2 (2): 234-293.
MARGALEF, R., 1949c. Las asociaciones de algas en las
aguas dulces de pequeño volumen del nordeste de España. Vegetatio, 1 : 258-284.
MARGALEF, R., 1950. Datos para la hidrobiología de la
Cordillera Cantábrica, especialmente del Macizo de los Picos de Europa. P. Inst. Biol. Apl., 7: 35-76.
MARGALEF, R., 195 la. Materiales para la hidrobilogía de la isla de Ibiza. P. Insr. Biol. Apl., 8: 9-70.
MARGALEF, R., 195 1 b. Regiones limnológicas de Cata- luña y ensayo de sistematización de las asociaciones de
algas. Collect. Bot., vol. 111, fasc. 1, n" 2: 43-67. MARGALEF, R., 1952a. Materiales para la hidrobiolo-
gía de la isla de Menorca. P. Inst. Biol. Apl.. 1 1 : 5-1 12. MARGALEF, R., 1952b. La vida en las aguas dirlces de
Atidor.ru. Mot~ogt.uf: Inst. Est. Pirenaicos, 69, 107 pp.
MARGALEF, R., 1953. Materiales para la hidrobiología de las isla de Mallorca. P. Inst. Biol. Apl., 1 S: 5-1 1 1.
MARGALEF, R., 1955. Comunidades bióticas de las
aguas dulces del NO de España. P. Inst. Biol. Apl., 21: 5-85. MARGALEF, R., 1955. Los organismos indicadores de
la Lin~tiologíu. Ministerio de Agricultura Ed. 300pp. MARGALEF, R., 1956. Materiales para una flora de las
algas del NE de España. VII. Suplemento. Collect. Bnt., vol.
V, fasc. 11, n": 87-107. MARGALEF, R., 1957. Materiales para una flora de las
algas del NE de España. VIII. Suplemento. Collect. Bot., vol. V, fasc. 11, no 15: 353-365.
MARGALEF, R., 1958. Trophic typology verus biotic
typology, as exemplified in the regional limnology of Nort- hern Spain. Verh. intrrn. Verein Limnol., 13: 339-349.
MARGALEF, R., 1983. Limnologíu. Omega. 1 O1 O pp.
MARGALEF, R.; PLANAS, M. D.; ARMENGOL, J.;
VIDAL, A.; PRAT, N,; GUISSET, A,; TOJA, J. & M. ESTRADA., 1976. Liwinologíu de los embolses españoles.
MOPU. Madrid. 422 pp. SABATER, S., 1982. Estsrdio de lela sucesicín ecol6gica
cle 11ria pohlucicín alga1 en I'Ai,en(,ó (Montseny). Graduate St. Dissert. Barcelona Univ.
SABATER, S., 1987. Estudi de les poblacions d'algues del r.i~c Ter. Ph. Doctoral Thesis. Barcelona Univ.
SABATER, S., 1988. Composición y ciclos de pigmentos clorofílicos en las poblaciones del fitobentos del río Ter durante un periodo de nueve meses. Oe(.ologia aquatica., 9: 61-75.
SABATER, S., 1989. Encrusting alga1 assemblages in a
Meditterranean River Basin. Arch. Hydrohiol. 114 (4): 555.573.
WANG, J. CH.; HOSHAW, R. W. & R. M. McCOURT., 1986. A poliploid species complex of Spirogyra commu-
nis (Chlorophj~tu) occurring in nature. J. Phycol., 22:
102- 107.

BENTHOS OF SPANISH LAKES AND RESERVOIRS
N. Prat, M. Real and M. Rieradevall Dept. Ecologia. Universitat Barcelona, Avda. Diagonal, 645; 08028 Barcelona. Spain
Keywords: Benthos, Meiofaunal loop, Benthic typology. Oxygen depletion.
ABSTRACT
A summary of the results from the study of benthos of lakes and reservoirs in Spain is provided, with a list of the species
found to date. Spanish natural lakes are smaller than European lakes; the largest is Lake Sanabria, of glacial origin, which is 3 Km long and half a kilorneter wide. Many are very small and situated in the mountains; more than 200 hundred have been recorded in Spain, but only in Lake Sanabria and Lake Banyoles have the benthos been studied. Lake Sanabria is a cold oligo- trophic, monomictic lake with oxygen always present in the deepest zones. Its fauna is similar to that of other central European
lakes, with Mici.opsectra c.orztractu (a chironomid) as the dominan1 species. Lake Banyoles is a karstic, monomictic and multi- basin lake; despite the low primary productivity, due to the abundante of sulphate in the water, the allochthonous inflow of organic matter and the inflow of water from bottom springs, the profunda1 environinent is very stressing for benthic fauna. Very low oxygen concentrations and high sulphide content in the water and sediments dueto meromixis mean that only the larva
of the dipteran Chaohoi.lcs flai7icans was present in one of the 5 basins of the lake. In other basins, when oxygen is available (no meromixis), the fauna is similar to that of tlie inineralized lakes of the Aegean region and some lakes in central Italia. On the other hand, preliminary data from the Pyrenean lakes and from Sierra Nevada ponds reveal no differences with northern cold
lakes. Tlie largest lakes in Spain are the reservoirs. There are nearly 1000 and data are available on 100 of them, including
the kargest. In addition to oxygen and sulphide content in the bottom waters, water level fluctuation and high sedimentation rates are disturbance factors that prevented the organization of the community. Allochthonous inputs of organic matter are also an important factor both in the reservoirs and also in the small, oligotrophic lakes like Banyoles and Sanabria. As a result the meiofaunal loop is very important in many of the Spanish water bodies . For this reason the natural lakes and reservoirs of
Spain are dominated by Oligochaeta, small crustaceans and the microcarnivore chironomids (such as Procladi~ls, Cladopelma
and Mi(,rnc.hil-onnmus) that feed on these meoifaunal elements. The phytophagous chironomids, like Chironomus, are only abundan1 in the shallow areas of mesotrophic and eutrophic reservoirs. This situation makes it difficult to apply the typologi-
cal system of SAETHER which predicts with some confidence only the benthic communities of Spanish natural lakes above
1500 m in the Pyrenees or the ponds above 2000 m in Sierra Nevada mountains. Higher temperatures (which originate a longer stratification period), the presence of sulphate in the waters of the eastern part of Spain and high inputs of sediments and allocht- honous organic matter seem to be the factors that originated the differences between the benthic profunda1 faunas of Spanish
lakes and reservoirs and those of the temperate lakes of north and central Europe.
INTRODUCTION AND SITES SAMPLED.
In the last 20 years there has been a considerabl advance in research on benthos in Spanish lakes and reservoirs. Most of the work has been done in connection with different rese- arch programmes, especially those devoted to the extensive studies on Spanish reservoirs (MARGALEF et al., 1976). To date. there are data from 131 reservoirs scattered al1
around Spain (PRAT, 1976, 1978, 1980b, 1 9 8 0 ~ ; REAL &
PRAT, 1991; LIMNOS S.A., 1989; REAL & PRAT, in
press) as well as from lakes Banyoles and Sanabria (RIERADEVALL & PRAT, 1989, 199 1 ; PRAT & RIERA- DEVALL, in pi.ess; RIERADEVALL & REAL, in press, RIERADEVALL et al., in press). These are the two largest natural lakes in Spain. We also have data from the commu- nities of lakes from the Pyrenees (not published) and we
L ~ ~ i i ~ i c t ~ c a , 8 : 221-22') ( 1 0 0 2 ) O Asociación E s p ~ ~ i o l ü de 1.iniriologia. >Madrid. Spain

know the presence in Sierra Nevada lakes and ponds of some species through the adults or pupal exuviae of Chiro- nomidae collected (LAVILLE & VILCHEZ, 1986). On the
other hand the communities of some Pyrenean lakes are well known from the studies in the French Pyrenees (LAVILLE, 1972) and the communities of Spanish Pyrenees should be
very similar. The geographical distribution of the sites
sampled in Spain is providcd in figure 1.
Figure l . Site5 \oiiiplctl Ioi- hciitlio\ I I I Spaiti. Opeti circles are reser- voirs, the open triangle Lake Baiiyoles, the S is Lake Saiiübria and the squares denotes the position of the areas of mountain lakes from which some information exist including those of the lakes investi- gated in France by LAVILLE (1972).
MACROBENTHIC COMMUNITIES
We have studied the macrobenthos after sieving the mud
taken with Ekman or Van Veen grabs with nets of 150
microns (Banyoles) or 250 microns (other studies). The most complete list for a single lake is that of Banyoles, with a total of 66 species collected in the sublittoral (5-1 2 m) and the profundal (20 m) zones in 8 different sampling stations. This list includes some meiobenthic forms (Nematoda, Ostracoda, Copepoda and Cladocera). The list is provided as an appendix to this paper.
Some species frequently found in Spanish lakes and reser-
voirs are unusual in the species list from central and northern Europe lakes, such as Microchironornus tener and Clado- plerna virescens, which have been more frequently and abun- dantly found in the southern part of Europe (Italy) (BAZZANTI, 1983; SEMINARA & BAZZANTI, 1988) and in the Aegean lakes (STANKOVIC, 1959) as well as in two lakes in central Europe which have hard waters, ¡.e. the
Wolilensee aiid the Balaton. It is also interesting to note that, to date, only 3 species of Chironorwus (Ch. plurnosu.~, Ch. hernensis and Ch. nuditarsis ) have been found in the lake of Banyoles (RIERADEVALL & PRAT, 1989) and the Spanish
reservoirs (unpublished data) using the chromosomal patterns of collected larvae. Moreover, in lake Banyoles (RIERADEVALL & REAL, in pre.c.s) and in some reservoirs
(PRAT et al., in press) the worm Potarnothri-r heuscheri is more frequent and abundant than in other European lakes. The similarity to the Italian lakes is also evident in this case.
THE KEY FACTORS.
The study of lakes Banyoles and Sanabria and Spanish reservoirs has given us some insights into the importance of
several factors in defining the sublittoral and profundal communities
Temperature
Temperatures of Spanish lakes and reservoirs are higher than those of other European lakes, which influences the species composition. Thus many of the species from artic and alpine lakes are not found in our lakes or reservoris. Only the species composition of the mountain lakes and ponds in Spain over 1500 m altitude are similar to those of
lakes from northern and central Europe. The characteristic
chironomid larvae of these lakes such as Heterotrissocla- dius have been found only in the Pyrenean lakes (unpublis- hed data) and other typical cold-stenotermic species such as Mic.rop.~ec.tra hiderztata, only in Sierra Nevada mountains
(Granada, SE Spain) (LAVILLE & VILCHEZ, 1986). Indirect effects of temperature are caused by the duration
of thermocline, which can last, in the reservoirs of southern Spain and in Lake Banyoles from the beginning of April
until the middle November (RIERADEVALL & PRAT, 1989). This leads to a depletion of oxygen in the hypolim- nion of lakes with lower primary productivity than in central and northern Europe.
Oxygen and sulphide.
The lack of oxygen in the profundal water has been recognized as the key factor in determining the species composition in the bottom of many lakes. Recent laboratory experiments have shown that physiological adaptations allow some species to live in depleted oxygen environments
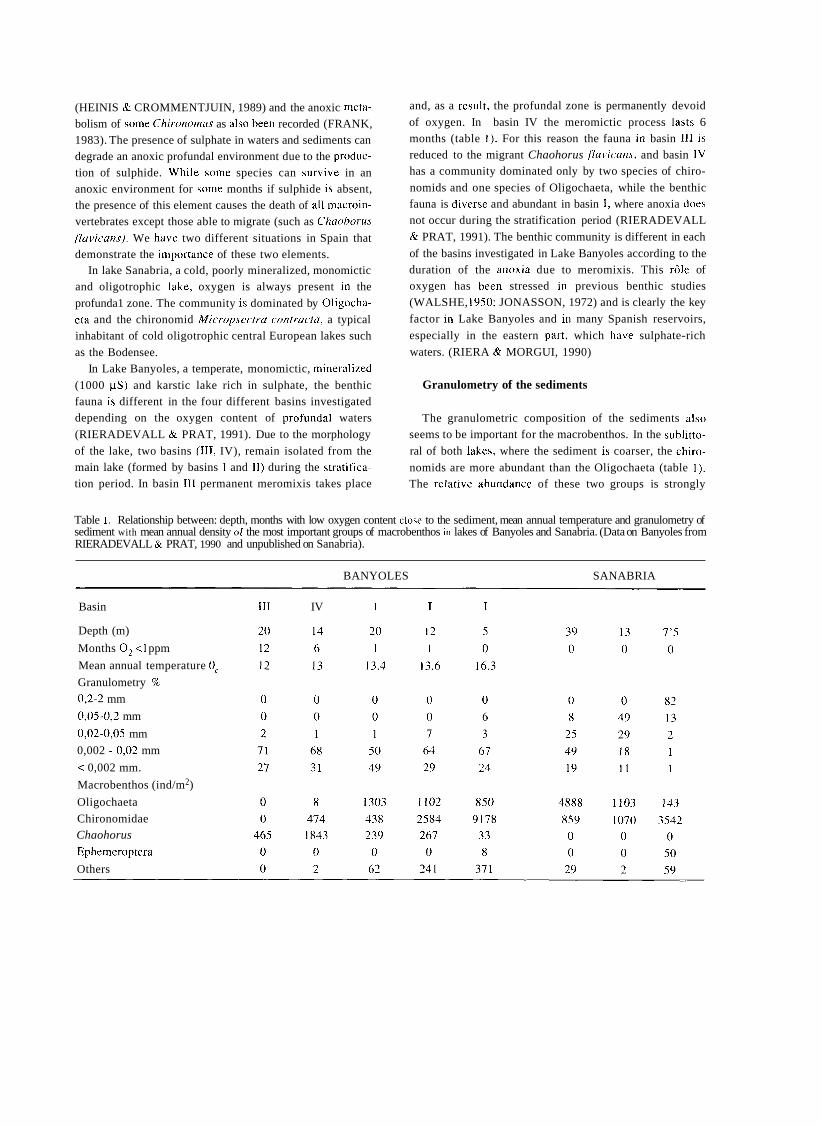
(HEINIS & CROMMENTJUIN, 1989) and the anoxic meta- bolism of some Chironornrts as also been recorded (FRANK, 1983). The presence of sulphate in waters and sediments can degrade an anoxic profundal environment due to the produc- tion of sulphide. While some species can survive in an anoxic environment for come months if sulphide is absent, the presence of this element causes the death of al1 macroin- vertebrates except those able to migrate (such as Chaohorus
flui~icuns). We have two different situations in Spain that
and, as a result, the profundal zone is permanently devoid
of oxygen. In basin IV the meromictic process lasts 6 months (table 1 ) . For this reason the fauna in basin 111 is reduced to the migrant Chaohorus ,flui~ic~ans, and basin 1V has a community dominated only by two species of chiro-
nomids and one species of Oligochaeta, while the benthic fauna is diverse and abundant in basin 1, where anoxia does not occur during the stratification period (RIERADEVALL
& PRAT, 1991). The benthic community is different in each
demonstrate the importance of these two elements. of the basins investigated in Lake Banyoles according to the
In lake Sanabria, a cold, poorly mineralized, monomictic duration of the anoxia due to meromixis. This r6le of
and oligotrophic lake, oxygen is always present in the oxygen has been stressed in previous benthic studies
profunda1 zone. The community is dominated by Oligocha- (WALSHE, 1950; JONASSON, 1972) and is clearly the key
eta and the chironomid Micropsecrru contracta, a typical factor in Lake Banyoles and in many Spanish reservoirs,
inhabitant of cold oligotrophic central European lakes such especially in the eastern part, which have sulphate-rich
as the Bodensee. waters. (RIERA & MORGUI, 1990)
In Lake Banyoles, a temperate, monomictic, mineralized
(1000 &S) and karstic lake rich in sulphate, the benthic Granulometry of the sediments fauna is different in the four different basins investigated depending on the oxygen content of profunda] waters The granulometric composition of the sediments also (RIERADEVALL & PRAT, 1991). Due to the morphology seems to be important for the macrobenthos. In the sublitto-
of the lake, two basins (111, IV), remain isolated from the ral of both lakes, where the sediment is coarser, the chiro- main lake (formed by basins 1 and 11) during the stratifica- nomids are more abundant than the Oligochaeta (table 1). tion period. In basin 111 permanent meromixis takes place The relative abundance of these two groups is strongly
Table l . Relationship between: depth, months with low oxygen content cloie to the sediment, mean annual temperature and granulometry of sediment with mean annual density of the most important groups of macrobenthos in lakes of Banyoles and Sanabria. (Data on Banyoles from RIERADEVALL & PRAT, 1990 and unpublished on Sanabria).
BANYOLES SANABRIA
Basin 111 IV I 1 1
Depth (m)
Months O2 < I ppm
Mean annual temperature Oc Granulometry %
0,2-2 mm
0,05-0,2 mm
0,02-0,05 mm
0,002 - 0,02 mm
< 0,002 mm.
Macrobenthos (ind/m2)
Oligochaeta
Chironomidae Chaohorus
Ephemeroptera
Others
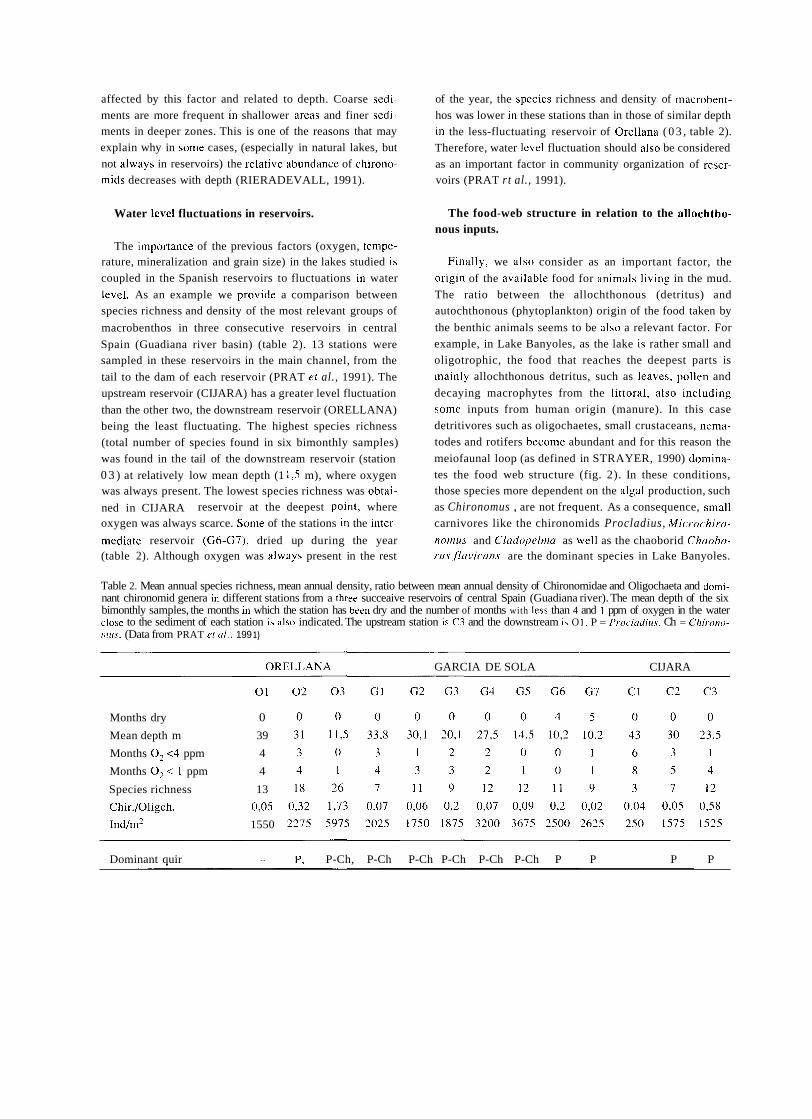
affected by this factor and related to depth. Coarse sedi- of the year, the species richness and density of macrobent-
ments are more frequent in shallower areas and finer sedi- hos was lower in these stations than in those of similar depth ments in deeper zones. This is one of the reasons that may in the less-fluctuating reservoir of Orellana ( 0 3 , table 2). explain why in some cases, (especially in natural lakes, but Therefore, water leve1 fluctuation should also be considered not always in reservoirs) the relative abundance of chirono- as an important factor in community organization of reser- inids decreases with depth (RIERADEVALL, 199 1). voirs (PRAT rt al., 1991).
Water level fluctuations in reservoirs. The food-web structure in relation to the allochtho- nous inputs.
The importance of the previous factors (oxygen, tempe- rature, mineralization and grain size) in the lakes studied is Finally, we also consider as an important factor, the
coupled in the Spanish reservoirs to fluctuations in water origin of the available food for animals living in the mud.
level. As an example we provide a comparison between The ratio between the allochthonous (detritus) and species richness and density of the most relevant groups of autochthonous (phytoplankton) origin of the food taken by
macrobenthos in three consecutive reservoirs in central
Spain (Guadiana river basin) (table 2). 13 stations were sampled in these reservoirs in the main channel, from the
tail to the dam of each reservoir (PRAT et al., 1991). The
upstream reservoir (CIJARA) has a greater level fluctuation
than the other two, the downstream reservoir (ORELLANA)
being the least fluctuating. The highest species richness
(total number of species found in six bimonthly samples)
was found in the tail of the downstream reservoir (station
0 3 ) at relatively low mean depth (1 1,5 m), where oxygen
the benthic animals seems to be also a relevant factor. For
example, in Lake Banyoles, as the lake is rather small and
oligotrophic, the food that reaches the deepest parts is
mainly allochthonous detritus, such as leaves, pollen and
decaying macrophytes from the littoral, also including
some inputs from human origin (manure). In this case
detritivores such as oligochaetes, small crustaceans, nema-
todes and rotifers become abundant and for this reason the
meiofaunal loop (as defined in STRAYER, 1990) domina-
tes the food web structure (fig. 2). In these conditions,
was always present. The lowest species richness was obtai- those species more dependent on the alga1 production, such
ned in CIJARA reservoir at the deepest point, where as Chironomus , are not frequent. As a consequence, small
oxygen was always scarce. Some of the stations in the inter- carnivores like the chironomids Procladius, Microchiro-
mediate reservoir (G6-G7), dried up during the year nomus and Cladopelma as well as the chaoborid Chaoho- (table 2). Although oxygen was always present in the rest i x s flai,ic.an.s are the dominant species in Lake Banyoles.
Table 2. Mean annual species richness, mean annual density, ratio between mean annual density of Chironomidae and Oligochaeta and domi- nant chironomid genera in different stations from a three succeaive reservoirs of central Spain (Guadiana river). The mean depth of the six bimonthly samples, the months in which the station has been dry and the number of months with less than 4 and 1 ppm of oxygen in the water close to the sediment of each station is also indicated. The upstream station is C3 and the downstream is O l. P = Procludius. Ch = Chirono- 1 1 1 1 ( ~ . (Data from PRAT e! al., 199 1 )
ORELLANA GARCIA DE SOLA CIJARA
Months dry 0
Mean depth m 39
Months O2 <4 ppm 4
Months O2 < 1 ppm 4
Species richness 13
Chir./Oligch. 0,OS
1nd/m2 1550
Dominant quir - P, P-Ch, P-Ch P-Ch P-Ch P-Ch P-Ch P P P P
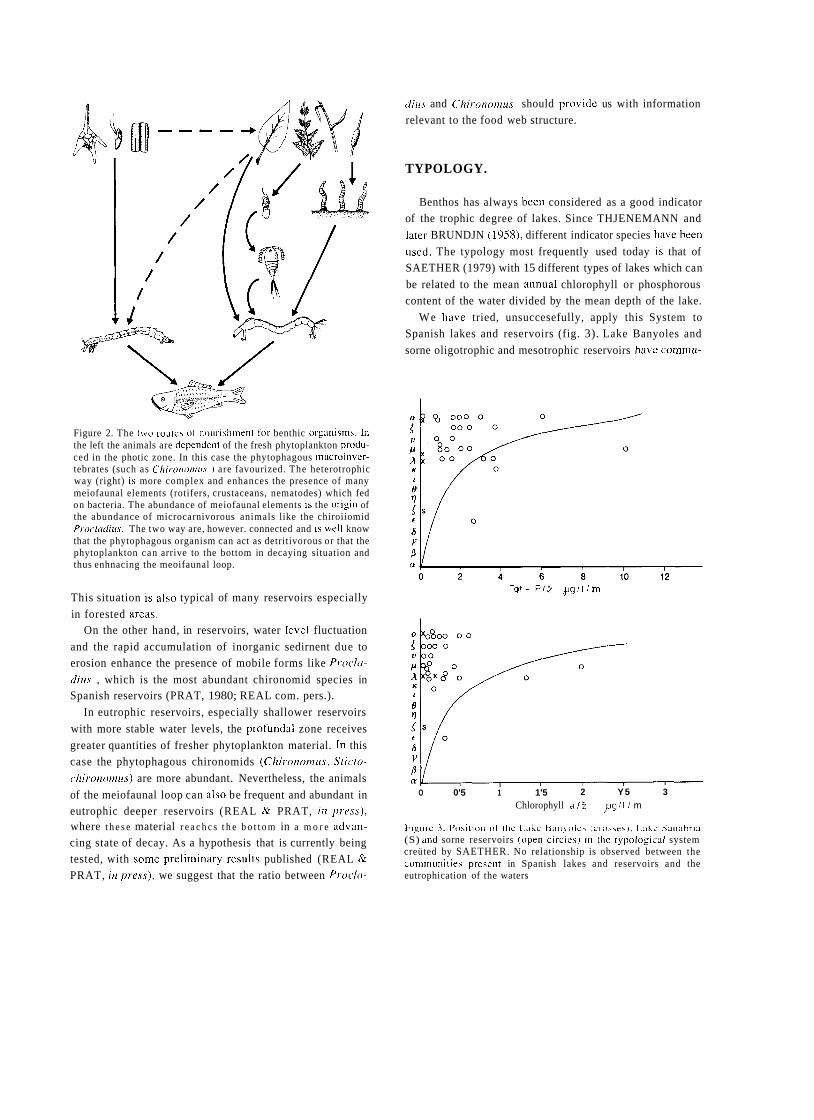
Figure 2. The t w o route\ ol iio~irishiiient f o r benthic organisins. In the left the animals are dependen1 of the fresh phytoplankton produ- ced in the photic zone. In this case the phytophagous macroinver- tebrates (such as Chirot~on~lrs ) are favourized. The heterotrophic way (right) is more complex and enhances the presence of many meiofaunal elements (rotifers, crustaceans, nematodes) which fed on bacteria. The abundance of meiofaunal elements is the origin of the abundance of microcarnivorous animals like the chiroiiomid Proclarli~rs. The two way are, however. connected and is well know that the phytophagous organism can act as detritivorous or that the phytoplankton can arrive to the bottom in decaying situation and thus enhnacing the meoifaunal loop.
This situation is also typical of many reservoirs especially
in forested areas. On the other hand, in reservoirs, water leve1 fluctuation
and the rapid accumulation of inorganic sedirnent due to erosion enhance the presence of mobile forms like Procla-
dizrs , which is the most abundant chironomid species in Spanish reservoirs (PRAT, 1980; REAL com. pers.).
In eutrophic reservoirs, especially shallower reservoirs
with more stable water levels, the profunda1 zone receives
greater quantities of fresher phytoplankton material. Jn this case the phytophagous chironomids (Chironom~ls, Sticto-
c.hironornus) are more abundant. Nevertheless, the animals
of the meiofaunal loop can also be frequent and abundant in eutrophic deeper reservoirs (REAL & PRAT, i t ~ piess) . where t h e s e material r e a c h c s t h e b o t t o m in a m o r e advan- cing state of decay. As a hypothesis that is currently being tested, with some preliminary results published (REAL & PRAT, in press), we suggest that the ratio between Pl.o<,la-
dius and Chir.o/ionllrs should provide us with information relevant to the food web structure.
TYPOLOGY.
Benthos has always been considered as a good indicator of the trophic degree of lakes. Since THJENEMANN and
later BRUNDJN (1958), different indicator species have been
used. The typology most frequently used today is that of SAETHER (1979) with 15 different types of lakes which can
be related to the mean annual chlorophyll or phosphorous content of the water divided by the mean depth of the lake.
We have tried, unsuccesefully, apply this System to Spanish lakes and reservoirs (fig. 3). Lake Banyoles and
sorne oligotrophic and mesotrophic reservoirs have commu-
O 0'5 1 1'5 2 Y5 3 Chlorophyll a l ? , ~ ~ g / l 1 m
Figure 3. Positioil ol ih t . 1.iilc. K,iii!oIc\ c i i o \ \ r \ j . Liihc Saiiabi-ia (S) and sorne reservoirs (open circles) ir1 tlie typological system creüted by SAETHER. No relationship is observed between the coriiinunities preseiit in Spanish lakes and reservoirs and the eutrophication of the waters

nities normally considered as typical of eutrophic lakes. The different basins of Banyoles are classed in different parts of the system and nearly al1 of the reservoirs considered (20, al1 in the Duero basin) are classified in the last 5 classes of
the SAETHER's system (fig. 3). Only the lakes of the Pyre- nees and the Sierra Morena ponds fit well into the first. ultraoligotrophic classes. We can consider Lake Sanabria
and only one reservoir in the to the mountains in the 5 inter-
mediate levels As a conclusion, the typology used for northern European
and American lakes is not very useful for the southern part
of Europe; it can be applied only to natural lakes over 1500 m in the Pyrenees and ponds over 2000 m in Sierra Nevada. As many of our reservoirs are below these altitudes. the thermal characteristics prevent the presence of many indi-
cator species. The benthic community of the profundal areas of our reservoirs are largely determined by the oxygen
content, the water level fluctuations and the allochthonous inputs of organic matter. As the oxygen content close to the bottom in Mediterranean and subtropical climates is not
related to eutrophy to such an extent as in northern lakes (Me CULLOGH & JACKSON, 1985), our communities do not fit the SAETHER's typology, which has been developed for cold temperate lakes. We have also to consider the origin
of the food consumed by the benthic animals, especially in the oligotrophic lakes and reservoirs, and the importance of watershed uses and erosion as well as water level fluctua- tions in the reservoirs The communities developed under such conditions cannot be related to eutrophication and for
this reason the typological system of SAETHER is not appropriate.
CONCLUSIONS.
The main conclusion of our work is that in Spain, and in general in the southern European lakes, at altitudes lower than 1500 m, owing to the thermal conditions (long thermo- cline period) coupled in many cases to high sulphate content in the waters, oxygen depletion close to the bottom is found
at lower eutrophication levels than in northern and central
European lakes. The comparison between Banyoles and Sanabria lakes (both oligotrophic, at similar latitudes, but at different altitudes and with -very different thermal regime and water mineralization) is clear in this respect. In addi-
tion, the abundance of allochthonous inputs eriliarices the meiofaunal loop, which could explain the abundance of small microcarnivores in our lakes and reservoirs (like the
chironomid Pl.oc,ladius). The presence of this chironomid in reservoirs is also enhanced by the high sedimentation rate. The phytophagous chironomids, like Clii/.onomlts, are only abundant in shallower areas of eutrophic reservoirs espe- cially in winter or spring, being absent from the profundal areas during summer especially in sulphate-rich waters. This reduces thc community to the most resistent species and
makes it difficult to use the indicator system adopted for
other lakes.
ACKNOWLEDGMENTS.
This work has been rnade possible by different CAYCIT contracts and due to the collaboration of many people who assisted us in sampling and sorting. Special thanks are due
to Enrique Martínez-Ansemil, Josefina Sanz, Isabel Muñoz
and Carles Ibáñez. LIMNOS S.A. have provided us with an unpublished report used in our work.
REFERENCES.
BAZZANTI, M., 1983. Composition and diversity of the
profundal rnacrozoobenthic community in the polluted lake Nemi (Central Italy). 1979-80. Acta Oecol. oecol. appl.,
4(3): 2 1 1-220. BRUNDIN, L., 1958. The bottom faunistical lake type
system and its application to the southern hemisphere. More- oever a theory of glacial as a factor of productivity in lakes and oceans. Verli. internat. ver.c.iri. Limnol., 13: 288-297.
FRANK , C.. 1983. Ecology, production and anaerobic metabolism of Chironomus plzrmoslrs L. in a shallow lake TI.
Anaerobic metabolism. Ai.ch. Hyu'r.ohio1.. 96: 354-362. HEINIS, F. & T. CROMMENTUIJN, . 1989. The natural
habitat of the deposit feeding chironomid larvae Stictochi-
ronomus histrio (Fab) and C h i r o n o m ~ ~ s anrhrac,iri~rs Zett. in relation to their responses to changing oxygen concentra- tions. Acta Biol. Dehrec,. Oec,ol. Hung., 3: 141-146.
JONASSON, P.M.. 1972. Ecology and production of the profundal benthos. Oikos, suppl., 14: 1-148.
LAVILLE, H., 1972. Recherches sur les chironomides
(Dipt) des lacs du massif de Néouvielle (Hautes-Pyrénées). Premiere partie: Systématique. Ecologie, Phénologie. Ann. Lirnnol. 7: 173-332.
LAVILLE. H. & A. VILCHEZ. 1986. Les Chironomidés (Diptera) de quelques "lagunas" de haute altitude de la Sierra Nevada (Granada. Espagne). Annls. Lirrinol. 22(1): 53-63.

LIMNOS S.A., 1989. Esrirtlio diu,gncí.stic.o dc la c~ilidatl
tle 1cz.s crglrus erriha1.sutlu.s de la t.oni~rriidac1 de Cn.stilln y
Lróir: Determinación y previsión de impactos, propuesta de corrección y red de vigilancia. 3 tomos.
MARTINEZ-ANSEMIL, E. & N. PRAT, 1984. Oligo- chaeta frorn profundal zones of Spanish reservoirs. Hyi1l.o- h i o l o ~ i u , 1 15: 223-230.
MARGALEF et. al. 1976. Lirrinologic~ da los eviihalses
pspot?oles. Public. Minist. Obras Públicas. n". 123. Mc CULLOUGH. J.D. & D. W. JACKSON,1985.
Composition and productivity of the benthic macroinverte- brate community of a subtropical reservoir. lnt . Revire ,gcs. Hydi.ohiol., 70: 22 1-233.
PRAT, N,. 1976. Fauna bentónica de los embalses espa- ñoles. Rcsiíniencs 11 A.sumhlc~ci Nnc.ionc11 tle Geodesirr y Geqfl.sico. págs. 204 1-2054.
1978. Benthic typology of Spanish reservoirs. C7er.h. inter-
nut. veiein. Limnol., 20: 1647- 165 1. 19793. Quironómidos de los embalses españoles (1).
Gi.uellsirc. XXXIII: 37-96. 1979b. Fauna marginal de los embalses españoles. Misc..
Zoologicu, 5: 149- 160. 1980a. Quironómidos de los embalses españoles (11).
Gi.uellsicl, XXXIV: 59- 1 19. 1980b. Bentos de los embalses españoles. Oec.01. ciclrrut.,
4: 3-43. 1980c. Benthic population dynamics in artificial samplers
in an Spanish reservoir. IN: Chivonomidar: Ecology. Syste- marics und Physiolo,yy (D.A. Murray ed.). págs, 239-246. Pergamon Press. Oxford and New York.
PRAT, N. & V. DAROCA, 1983. Eutrophication proces- ses of Spanish reservoirs as revealed by biological records in profundal sediments. Hydvohiologia, 103: 153-158.
PRAT, N., J. SANZ, & E. MARTINEZ-ANSEMIL, 1991. El bentos litoral y profundo de una cadena de tres embalses españoles del rio Guadiana. Limnetica. , 7 : 133-152
PRAT, N. & M. RIERADEVALL, in press. Life cycle and production of Cladopelmc~ iivescens (Mg.) (Dipt. Chiro- nomidae) in lake of Banyoles (NE Spain). Hydvobiol. Bii11.
227
REAL, M. & N. PRAT, 1991. Changes in the benthos of five Spanish reservoirs ir1 the last 15 years. C7eih. intc~inat.
i~rrc~irr. Lin~nol. , 24: 1377- 138 1. in press. Factors influencing the profundal macrozoo-
benthos of Spanish reservoirs. Hydr.ohio1. Bull. REAL, M.; F. SABATER, & J.A. MORGUI,. in press.
Significant physiographic disturbances in the Ebro basin (NE Spain) reflected by Oligochaeta size spectra. Hydvo-
hiologiu. RIERA, J.L. & J.A. MORGUI, 1990. Limnología regional
de los embalses españoles. Mundo Cicvziíji'co, 104: 720-726. RIERADEVALL, M. 199 1. Ec.ologici i produc'cicí del
hcntos (le1 Ilat de Bunjoles. Tesi Doctoral. Universitat de
Barcelona. 223 págs. RIERADEVALL, M. & N. PRAT, 1989. Chironomidae
from profuridal samples of Banyoles lake. Ac,tu Biol. Dehr.
Oecol. Hurig., 3: 267-274. 1991. Benthic fauna of Banyoles Iake. Vevh. inter.riut.
veruin. Lin~nol . , 24: 1020- 1023. RIERADEVALL, M. & M. REAL, in press. Sublittoral
and profundal fauna from Lake Banyoles (Catalonia, NE Spain). Hydi-ohiologia.
RIERADEVALL, M.; C. IBAÑEZ; 1. MUNOZ; M. REAL & N. PRAT, in press. Grupos de macroinvertebrados del bentos del lago de Sanabria y su relación con la granu- lometria del sedimento. Actus VI Congreso Español de
Limnologia.
SAETHER, O.A. 1979. Chironomid communities as water quality indicators. Holai.c,t. Eco[., 2: 65-74.
STRAYER, D.L. 199 1 . Perspectives on the size structure of lacustrine zoobenthos, its causes and its consequences. Journcrl N. Am. Bc.nrol. Soc., 10 (2 ) : 2 10-22 1 .
SEMINARA, M. & M. BAZZANTI,. 1988. Trophic leve1 assessment of profundal sediments of the artificial lake Campotosto (Central Italy), using midge larval community (Dipt. Chir.). Hydr.ohio1. B~rll., 22(2): 183-193.
WALSHE, B. M. 1950. The function of haemoglobin in ~ h i v o n o m u s p1urrio.su.s under natural conditions. J. exp.
Biol., 27: 73-95.

APENDIX.
List of species found in the profunda1 and sublittoral areas from Spanish lakes and reservoirs. B= Banyoles, S= Sanabria, R= Reservoirs.
NEMATODA
Daptotzemcr duhium, B Doryluinzus stagnalis, B , R Liniiiorner.mis hostrychorles. R
Tohrilus sp. (T . gracilis), B , R
OLIGOCHAETA Der.o n'igitata, B , R , Aulodr.ilus plitriseta, R A14lodi.il~rs pigueti, B , R Branchi~o-a sowerhyi. B , R . Limnodri1u.s c~lupuredriunus, B , R
L. hcd'nzeistei.i, B , R , Lin7nodi.ilir.s pi.ofirtidicola, R L. udekernianus, R Porumothri.~- bavarici4s, B , R
P. hammoniensis. B , R , P. heltsc'heri, B . R Tithifefes tuhife.~, R Psunirnoryctides hui.hc~til~, B , R
R. h l ~ h h o l z i , R, B. appetidic~irlatc~
CRUSTACEA MACROTRICIDAE
1l.yoc.1.ypt~t.s ~ o r d i d ~ l s . B , R ,
CHIDORIDAE Alona affiinis, B
Alona q~radrangltIai.is, B
Scccpho1ehei.i~ sp., B ,
COPEPODA Cyc,lops .SI.>., B , R, Diac,yc,lops hicuspidarlrs, B Eucyclops srt.r-ularus, B
OSTRACODA Candona neglecta, B -'c,loc,ypris oi3Ltnz. B
Porc~nais ,fr.ic.i, R Nais pardalis, R Nais comrnltnis, R Nais hai-bata, R. Nais i~ai.iahilis, R. Nais pselrdohri4sa, R. Ophionais serpentinc~, R Pr. rnenorii, R , Uncinuis unc,inata, R
Spirosperma feros, R , S Haber p y r e n a i c . ~ ~ ~ , S
Emholocepahlus velufinus, S Bothrioner~trn vqjdowskyanunr, R Ft.idet.icia sp., R.
Leydigia acanthoceiv,oides, B
Leydigia leydigii, B
Mac rocyc 1ops albidirs. B
Paracyc'lops fimhriatus, B
Cyprideis toi.osa, B
1Iyocypr.i~ hradyi, B

Cypria Iírcit.sti.is, B Darn3itzula stei3erisoni
GAMMARIDAE Echinogammarus pitngeris. B
HYDRACARINA Ar.renurus sirz~taror., B Nel<munia deltoides , B
INSECTA EPHEMEROPTERA Ephemera glaucops. B DIPTERA CHAOBORIDAE Chuohor~~s,flavic.rrns, B. R. CHIRONOMIDAE Procludius <f. chore~*s, B , R Tanypus punctiperrrris, B , R, Ahlahesmyiu sp., B , R Cricotopus ( I . ) sylvestris, B , R, Cricotopus gr. hicinctus, B , R, Cricotopus gr. ,festivell~ts, B Chironomu.~ plumosus, B . R , Ch. nuditarsis, B , R, Ch. herrze/zsis, B, R, Cludopelma virescens, B , R , Cryptochironomus sp., B , R, Cryptotendipes sp., B , , Harnischia sp., B. R, Microchironomus tener, B , R Microtendipes sp., B , R, Purucludopelma sp. B, Paratendipes sp., B Polypedilum spp., B , R , Stictochironomus maculipennis, B , R, Cladotanytarsus atridorsum, B , R , Micropsectra sp., R , S Stempellina sp., B , R, Tanyrarsus gr. lestagei, B.
I1yoc.ypi.i.s gihha, B
I.rocypcrs heuut harnpi
Neunzrrrzia imitara, B Uriionic.ola crassipc.~ , B
A. longistyla, R Prodiamesa o l i~ucea , R Parakiejferiella hutophila, R
Ch. halophilus, R.
S . hisrrio, R C . mctncus, R M . c.ontractu, R S . ha~tsei, R T . hatophilus, R

PRESENT LEVEL OF KNOWLEDGE REGARDING FLUVIAL MACROINVERTEBRATE COMMUNITIES IN SPAIN
J. Alba-Tercedorl, G. G o n z a l e z 2 and M.A. Puig3 1. Departamento de Biología Animal y Ecología. Facultad de Ciencias, Universidad de Granada. Spain. 2. LIMNOS S.A..Bruc, 168. Barcelona 08037. Spain. 3. Centro de Investigaciones del Agua. CSIC. La Poveda, 28500 Arganda del Rey, Madrid. Spain.
Keywords: River systems, Macroinvertebrates, History, Knowledge level. Spain
ABSTRACT
A synthetic review examines present knowledge of fluvial macroinvertebrates in Spain. Many works have been strictly dcscriptive, and most have aimed at the application of biotic indices in water quality studies. Nevertheless, new awareness in the 1980s, fostered by the Spanish Association of Limnology, has begun to stir works that carry more profound implications regarding community ecology and lotic ecosystems.
INTRODUCTION
A knowledge of benthic macroinvertebrate communities is widely considered to be vital in understanding the ecology of waterways (MARGALEF, 1947, 1960). The principal necessity in developing the study of macrobenthic ecology is a good taxonomic knowledge of the different faunistic groups involved. In Spain two major problems exist. Firstly, a great diversity of species inhabit fluvial environments. and the role of the Iberian Peninsula as a refuge and/or disper- sion zone at different geological moments has made the area faunistically very rich, with numerous endemic species (SANCHEZ-ORTEGA & ALBA-TERCEDOR, 1987). Secondly, if in fact the number of taxonomists who work on these groups has increased, the number is still small, and in any case inadequate (VALDECASAS et al., 1990). One reason for this is the comparatively low value generally placed on taxonomy. For example, studies on the water quality of rivers, using more or less simple biotic indices, abound because of the great acceptability and social interest of this work. Consequently, we are currently reaching a paradox in which we have studied water quality in a good part of our waterways without knowing the composition of species inhabiting those environments.
In Spain a good index of the true situation surrounding faunistic knowledge is the publication of the faunistic lists and bibliographies of the different fluvial macrobenthic
groups. Apart from works in preparation, to date there have been publications on Ephemeroptera (ALBA-TERCEDOR, 1981), Plecoptera (SANCHEZ-ORTEGA & ALBA- TERCEDOR, 1987), Heteroptera (NIESER & MONTES, 1984), Coleoptera (MONTES & SOLER, 1986; RICO et al. . 1990; VALLADARES & MONTES, 199 1 ), Mollusca bival- via (VIDAL-ABARCA & SUAREZ, 1985), Hydrachnellae (VALDECASAS, 1988); and a preliminary list of chirono- mid Diptera (COBO et al., 1987).
In this work we present a general synthesis of current knowledge regarding sensu lato ecology of fluvial macroin- vertebrates in Spain. For this we have included only those studies which involve macroinvertebrates as a whole, and therefore we have not considered those works that include only one or two faunistic groups, or those that contribute no information other than a list of taxa.
HISTORY
The beginning of the study of fluvial benthic communi- ties can be traced to the works of MARGALEF (1947, 1948, 1949, 1952, 1953, 1955, 1960), which offer descriptions of communities (1946, 1948, 1949, 1952, 1953, 1955) as well as theoretical proposals for the stages in community deve- lopment and for comparisons between fluvial and lacustrine systems (1947). Professor Margalef's role as founder and
Limnetica, 8: 23 1-24 1 (1992) O Asociación Española de Lirnnología, Madrid. Spain
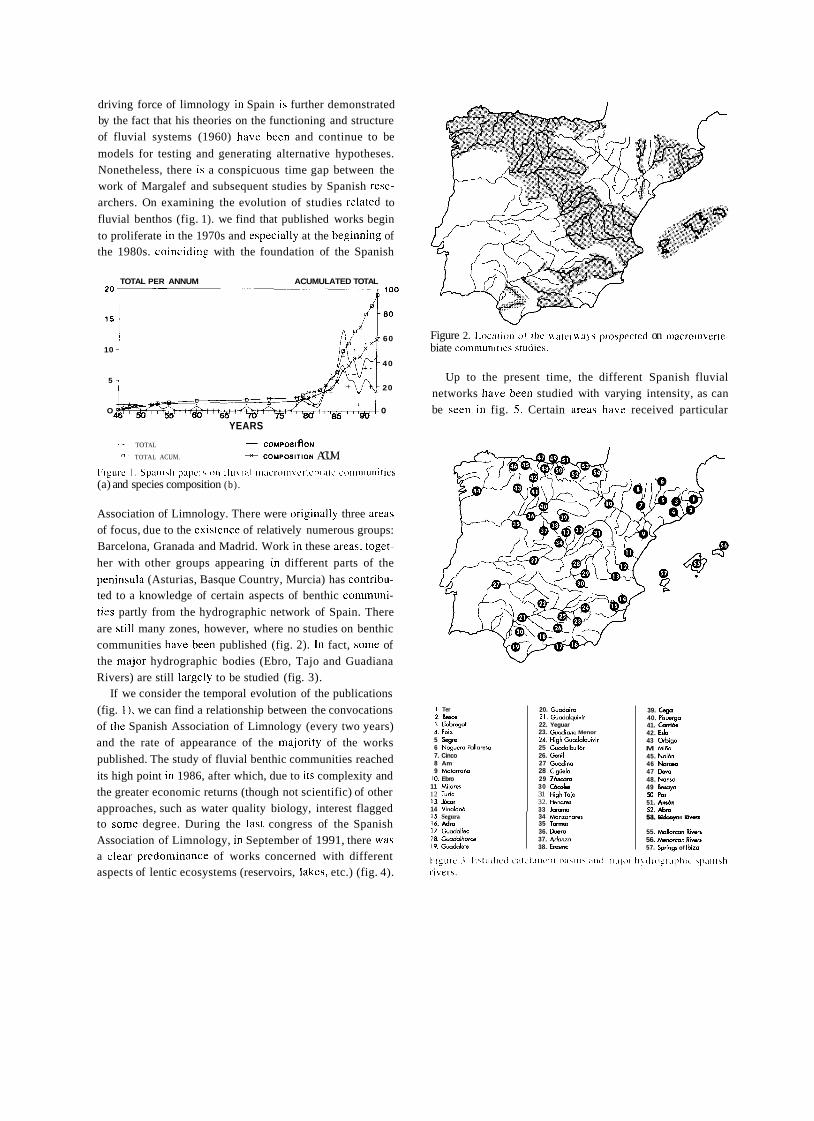
driving force of limnology in Spain is further demonstrated by the fact that his theories on the functioning and structure of fluvial systems (1960) have been and continue to be
models for testing and generating alternative hypotheses. Nonetheless, there is a conspicuous time gap between the work of Margalef and subsequent studies by Spanish rese- archers. On examining the evolution of studies related to
fluvial benthos (fig. 1). we find that published works begin
to proliferate in the 1970s and especially at the beginning of the 1980s. coinciding with the foundation of the Spanish
TOTAL PER ANNUM ACUMULATED TOTAL
15
60 10
40
5 20
O o
YEARS - TOTAL + C O M P O S I ~ ~ O N
+ TOTAL ACUM. - COMPOSITION ACUM. Figu~t l . S[ I~II I I \ I I ~ : I I ) C I \ o11 I I L I I I ; I I I I I : I L I O I I I \ C ~ I C ~ > I ~ I I C ~ o ~ n ~ ~ i u ~ ~ i t i e s (a) and species composition (b).
Association of Limnology. There were originally three aseas of focus, due to the existente of relatively numerous groups: Barcelona, Granada and Madrid. Work in these aseas, toget-
her with other groups appearing in different parts of the
peninsula (Asturias, Basque Country, Murcia) has contribu- ted to a knowledge of certain aspects of benthic communi-
ties partly from the hydrographic network of Spain. There
are still many zones, however, where no studies on benthic communities have been published (fig. 2). In fact, some of the major hydrographic bodies (Ebro, Tajo and Guadiana Rivers) are still largelfi to be studied (fig. 3).
If we consider the temporal evolution of the publications
(fig. l ) , we can find a relationship between the convocations of tlie Spanish Association of Limnology (every two years) and the rate of appearance of the majority of the works
published. The study of fluvial benthic communities reached
its high point in 1986, after which, due to its complexity and the greater economic returns (though not scientific) of other approaches, such as water quality biology, interest flagged to some degree. During the last congress of the Spanish Association of Limnology, in September of 1991, there was a clear predominance of works concerned with different aspects of lentic ecosystems (reservoirs, lakes, etc.) (fig. 4).
Figure 2. LOL~IIIOII 01 1 1 1 ~ U rltel I\ d) 5 pro\pe~ted on mdcroinverte- biate cornmunitie\ \tudie\.
Up to the present time, the different Spanish fluvial networks have been studied with varying intensity, as can be seen in fig. 5 . Certain areas have received particular
1 Ter 2. Besos 3. Liobregot 4. Foix 5 segre 6 Noguero Polloreia 7. Cinco 8 Am 9 MabrroM
10. Ebro 11 Mijarer 12 Turio 13. Júcar 14 Vinalwó 15. Segura 16. Adra 17. Guodalb 18. Gvadalhwce 19. Guodalete
20. Gucdaira 21. Gucdalquirir 22. Yeguar 23. Gucdiana Menor 24. High Gucdalquirir 25 Gucdalbullbn 26. Ganil 27 Gucdino 28 Cigiiela 29 Z6ocora 30 C k o h 31 HighTajo 32. Henotes 33 Joro- 34 Manzanares 35 Torme. 36. Duem 37. hbnra 38. Ereimo
I I I C I I I l1.1\111\ .11111
39. Cego 40. Piruergo 41. Carribn 42. Esb 43 Ohigo M Mi60 45. Nol6n 46 Narcea 47 Da0 48. Nonro 49 Beriyo 50 Pos 51. Andn 52. Abr. 53. 54. Vircopn B i&m Rivan
55. Mallorcon Riwo 56. M o r c o n Rivso 57. Spn'ngs of Ibiza
I I I C I I O I I I ~ L I I ~ I ~ I ~ I I ~ I I I L \ I ~ , I I I I \ ~
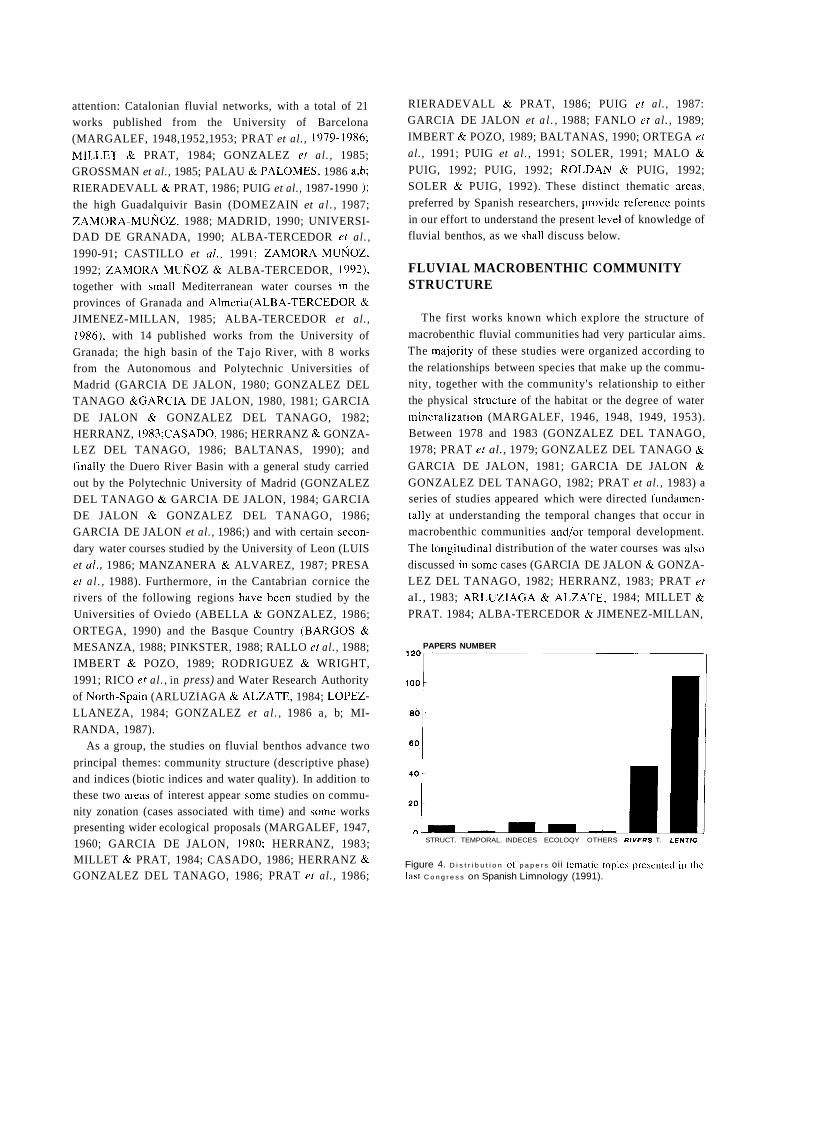
attention: Catalonian fluvial networks, with a total of 21 works published from the University of Barcelona (MARGALEF, 1948,1952,1953; PRAT et al., 1979-1986; MlLLET & PRAT, 1984; GONZALEZ et al . , 1985; GROSSMAN et al., 1985; PALAU & PALOMES, 1986 a,b; RIERADEVALL & PRAT, 1986; PUIG et al., 1987-1990 ); the high Guadalquivir Basin (DOMEZAIN et a l . , 1987; ZAMORA-MUÑOZ, 1988; MADRID, 1990; UNIVERSI- DAD DE GRANADA, 1990; ALBA-TERCEDOR et al . ,
1990-9 1 ; CASTILLO et al . , 199 1 ; ZAMORA-MUÑOZ, 1992; ZAMORA-MUÑOZ & ALBA-TERCEDOR, 1992), together with small Mediterranean water courses in the provinces of Granada and Almeria(ALBA-TERCEDOR &
JIMENEZ-MILLAN, 1985; ALBA-TERCEDOR et al.,
1986), with 14 published works from the University of Granada; the high basin of the Tajo River, with 8 works from the Autonomous and Polytechnic Universities of Madrid (GARCIA DE JALON, 1980; GONZALEZ DEL TANAGO &GARCIA DE JALON, 1980, 198 1; GARCIA DE JALON & GONZALEZ DEL TANAGO, 1982; HERRANZ, 1983;CASADO, 1986; HERRANZ & GONZA- LEZ DEL TANAGO, 1986; BALTANAS, 1990); and finally the Duero River Basin with a general study carried out by the Polytechnic University of Madrid (GONZALEZ DEL TANAGO & GARCIA DE JALON, 1984; GARCIA DE JALON & GONZALEZ DEL TANAGO, 1986; GARCIA DE JALON et al., 1986;) and with certain secon- dary water courses studied by the University of Leon (LUIS et al., 1986; MANZANERA & ALVAREZ, 1987; PRESA er a l . , 1988). Furthermore, in the Cantabrian cornice the rivers of the following regions have been studied by the Universities of Oviedo (ABELLA & GONZALEZ, 1986; ORTEGA, 1990) and the Basque Country (BARCOS & MESANZA, 1988; PINKSTER, 1988; RALLO et al., 1988; IMBERT & POZO, 1989; RODRIGUEZ & WRIGHT, 1991; RICO et al . , in press) and Water Research Authority of North-Spain (ARLUZIAGA & ALZATE, 1984; LOPEZ- LLANEZA, 1984; GONZALEZ et a l . , 1986 a, b; MI- RANDA, 1987).
As a group, the studies on fluvial benthos advance two
principal themes: community structure (descriptive phase) and indices (biotic indices and water quality). In addition to these two areas of interest appear some studies on commu- nity zonation (cases associated with time) and some works presenting wider ecological proposals (MARGALEF, 1947, 1960; GARCIA DE JALON, 1980; HERRANZ, 1983; MILLET & PRAT, 1984; CASADO, 1986; HERRANZ & GONZALEZ DEL TANAGO, 1986; PRAT et al., 1986;
RIERADEVALL & PRAT, 1986; PUIG et al., 1987: GARCIA DE JALON et a l . , 1988; FANLO et al . , 1989; IMBERT & POZO, 1989; BALTANAS, 1990; ORTEGA et
al., 1991; PUIG et al . , 1991; SOLER, 1991; MALO &
PUIG, 1992; PUIG, 1992; ROLDAN & PUIG, 1992; SOLER & PUIG, 1992). These distinct thematic areas, preferred by Spanish researchers, provide reference points in our effort to understand the present leve1 of knowledge of fluvial benthos, as we shall discuss below.
FLUVIAL MACROBENTHIC COMMUNITY STRUCTURE
The first works known which explore the structure of macrobenthic fluvial communities had very particular aims. The majority of these studies were organized according to the relationships between species that make up the commu- nity, together with the community's relationship to either the physical ftructure of the habitat or the degree of water mineralization (MARGALEF, 1946, 1948, 1949, 1953). Between 1978 and 1983 (GONZALEZ DEL TANAGO, 1978; PRAT et al., 1979; GONZALEZ DEL TANAGO &
GARCIA DE JALON, 1981; GARCIA DE JALON &
GONZALEZ DEL TANAGO, 1982; PRAT et al., 1983) a series of studies appeared which were directed fundamen- tally at understanding the temporal changes that occur in macrobenthic communities andlor temporal development. The longitudinal distribution of the water courses was also discussed in some cases (GARCIA DE JALON & GONZA- LEZ DEL TANAGO, 1982; HERRANZ, 1983; PRAT er
al., 1983; ARLUZIAGA & ALZATE, 1984; MILLET &
PRAT. 1984; ALBA-TERCEDOR & JIMENEZ-MILLAN,
PAPERS NUMBER
I z o o
STRUCT. TEMPORAL. INDECES ECOLOQY OTHERS RIVERS T. LENTlC
Figure 4. D i s t r i b u t i o n o f p a p e r s oii t e n i a t i c t o p i c s p r c s c n t e d iii t l i e
l a s t C o n g r e s s on Spanish Limnology (1991).

1985; GONZALEZ ut al.. 1985; ALBA-TERCEDOR et al . , 1986; PRESA ut al. , 1988; ZAMORA-MUÑOZ, 1988).
Of the overall number of works we have compiled, 95, only 58 (61 .OS%) offer information on the composition of these communities. This situation is primarily because.
between 1984 and 1986, works were continued on known
courses for which communities had already been described (Catalonia). Beginning in 1986, the year of the majos work describing benthic communities (fig. l ) , there was an appre-
ciable decrease in the number of new works concerning the composition and structure of the Spanish fluvial benthic
communities. On carefully analysing the state of knowledge of this type
of community composition we find that most studies focu-
sed on only a part of the problem and not always on the numerically predominant groups. In addition, studies of macroinvertebrate communities that include an identifica- tion of Diptera (especially the chironomids), at a level lower
than the family, are an exception (PRAT e t crl., 1983, 1985;
PUIG et al., 1984, 1987; RALLO et al., 1988; ROLDAN &
PUIG, 1992). For this reason, to provide a general characte- rization of the composition by taxa according to waterways, we have considered only the six groups that have been most
thoroughly studied in most cases: Ephemeroptera, Trichop- tera, Plecoptera, Coleoptera, Odonata and Heteroptera. On the basis of specific composition we have looked for tenden-
cies, similasities and differences among fluvial courses
which present different structural characteristics. Various types of faunistic composition have been detec-
ted (fig. 6): 1 . In the waterways of northeastern Spain (Cinca,
Noguera Pallaresa, Ter. Llobregat. Besos, Foix and Mata-
Figut-e 5 . l i i i r r l i ~ ~ i ~ 111 I ~ , I ~ C I \ ~)uhl~\lied ior firstly river basins.
rraña Rivers). that is, the sub-basin »f ihe Ebro River and ihc \111:1ll C : I I : I ~ O I ~ I : L I ~ \ \ : I I L , ~ C O L I I - \ C \ I I O I . I I ~ 0 1 . I I I C Fhro.
3 b@
0 Ephemeroptera m Heteroptera m Ple<optern Odonnh
Figure 6. Types of fauni\iic composition detected in the \panish basins.
dominance is shared by Trichoptera and Ephemeroptera, with Plecoptera as an accompanying species. For the al1 of the orders combined, moreover, the reophile species cons- titute the largest part of the community (PRAT et al . , 1983,
1985; GROSSMAN e t al . , 1985; PUIG e t al . , 1987; GARCIA DE JALON et al . , 1988; PUIG e t a l . , 1990; PUIG, 1992).
2. In waterways of the northwestern and central part of
Spain (small Asturian courses, Nalón, Duero and Jarama Rivers), Trichoptera remains dominant, with Ephemeroptera
as an accompanying group. But two subgroups are distin- guishable on the basis of the second accompanying group:
small courses with Plecoptera as the second accompanying order (Asturian rivers and the Jarama River) (GONZALEZ DEL TANAGO & GARClA DE JALON, 198 1 ; GARCIA DE JALON & GONZALEZ DEL TANAGO, 1982; LOPEZ- LLANEZA, 1984; ABELLA & GONZALEZ, 1986); and

large courses of generally little slope with Coleoptera as the second accompanying order (Duero River) (GARCIA DE JALON & GONZALEZ DEL TANAGO, 1986).
3. In al1 of the permanent waterways in the southern half of Spain Ephemeroptera is the order with the greatest number of species. Two subgroups can be distinguished: courses in which only Ephemeroptera dominates (the medium-high stretch of the Guadalquivir River and the Guadalhorce River) (GARCIA DE JALON & GONZALEZ DEL TANAGO, 1986; ALBA-TERCEDOR et al., 1990); courses with moun- tain systems, with Ephemeroptera and Coleoptera sharing dominance, and Trichoptera the basic accompanying group (sources of the Genil, Guadalete, Guadalfeo and Tajo Rivers) (ALBA-TERCEDOR & JIMENEZ-MILLAN, 1985; ZA- MORA-MUÑOZ, 1988; BALTANAS, 1990; GALLAR-DO, 199 1 ; ZAMORA-MUÑOZ, 1992).
4. In seasonal waterways with permanent reophile stret- ches (Yeguas, Adra and Vinalopó River) no order appears to dominate having a very balanced representation of species belonging to Ephemeroptera, Trichoptera, Coleop- tera, Odonata and Heteroptera. As a whole, lentic species dominate (GARCIA-ROJAS, 1985; ALBA-TERCEDOR et
al., 1986; ROLDAN & PUIG, 1992). 5. In completely seasonal waterways (Guadaira River and
Moro Guich), systems resembling ponds more than rivers, lentic Coleoptera are clearly dominant (GALLARDO & TOJA, 1988; ORTEGA, 1988; GALLARDO, 1991)
From the groupings of waterways according to commu- nity composition, a fairly dintinct typology can be identi- fied, which might be completed in the future when the remaining Spanish fluvial systems have been studied. New data would be especially interesting Spanish fluvial systems have been studied. New data would be especially interesting from general studies of the major river such as the Ebro, Tajo, Guadiana and Guadalquivir, though stretches of some of these have been partialty investigated (Guadalquivir, Guadiana Menor).
BIOTIC INDICES
Of the works published on the benthos of Spain 47.37% concern water quality, and use macroinvertebrate as indices of pollution. These studies, which are still on the rise, have as their precedent the works of MARGALEF (1952, 1969). Not until the 1970s, however, were any Spanish ideas effec- tively put into practice (GONZALEZ DEL TANAGO et al., 1979; PRAT et al., 1979), and in any case it should be
mentioned that the first practica] experiment dealing with Spanish territory was carried out in 1973 by MEYNELL. The major development began in 1984 (fig. 7) and continued its crescendo until the present. Regarding this group of works we should point out that only few authors have used the saprobios system (SLADECEK, 1966; MARGALEF,
,o i""' PER ANNUM ACUMULATED ToTAi
~ i k k ' ' '$oU '$5' ' ' 'do' O'
YEARS
?r WATER QUALITY " WATER QUALITY ACUM.
Figure 7. Spanish papers on biological pollution indicatoi-s (commu- nities).
1969; GARCIA DE JALON & GONZALEZ DEL TA- NAGO, 1986; VALDECASAS & BALTANAS, 1990). In the rest of the works authors have tried to test and adapt different European indices (Vernaux and Tuffery, Chandler, BMWP system) (TUFFERY & VERNEAUX, 1967; CHAN- DLER, 1970; ARMITAGE et al., 1983). There has been an obvious tendency toward obtaining maximum information with minimum effort. Leaving behind the inital use of biotic indices, which require the use of species identification (SLADECEK, 1966; TUFFERY & VERNEAUX, 1967), researchers are at present in more or less general agreement on the use of the BMWP' (ALBA-TERCEDOR & SAN- CHEZ-ORTEGA, 1988), an adaptation of the english BMWP (ARMITAGE et al., 1983) for the Iberian Peninsula, with quality guidelines which require the recognition of taxa
at the family leve1 only. From the studies considered in this article, concerning
both quality and any other perspective of fluvial macro- benthos for which the data allow the application of the BMWP' (GONZALEZ DEL TANAGO et al., 1979; GONZALEZ DEL TANAGO & GARCIA DE JALON, 1980; GARCIA DE JALON & GONZALEZ DEL TANAGO, 1982; PRAT et al., 1983; LOPEZ-LLANEZA, 1984; SOLER, 1984; ALBA-TERCEDOR & JIMENEZ- MILLAN, 1985; GARCIA-ROJAS, 1985; GROSSMAN et
al. , 1985; PRAT et al., 1979; ALBA-TERCEDOR et ul.,
1986; CASADO, 1986; GARCIA DE JALON & GONZA-

LEZ DEL TANAGO, 1986; GONZALEZ et a l . , 1986A; PALAU & PALOMES, 1986B; PRAT et al. , 1986; PUIG et al., 1987,1990; GARCIA DE JALON et al., 1988; RALLO et a1.,1988; ZAMORA-MUÑOZ, 1988; ALBA-TERCEDOR et al. , 1990; MADRID, 1990; ORTEGA, 1990; ALBA- TERCEDOR & PICAZO, 1990, 1991; CASTILLO et al. , 1991; GALLARDO, 1991; PUIG, 1992; ROLDAN & PUIG, 1992; ZAMORA-MUÑOZ & ALBA-TERCEDOR, 1992; MUÑOZ & PRAT, in press), two maps have been drawn to synthesize the water quality of Spanish rivers. One map represents water quality during the summer (fig. 8) while the other indicates the average annual values from those works spanning more than one season (fig. 9). In fact, due to extent of the territory and the cartographic scale, it has not been possible to include in these maps al1 the sampling points included in the original works. For this reason the result must be intespreted as an approximation of the general situa- tion (escecially when data was used from widely ranging years) for which partial works should be consulted to gain a more detailed interpretation of specific zones.
In most cases good biological conditions are found in unaltered communities, found only in stretches at the sour-
Figure 8. Estiniatioii 01' the unter quality dui-ing surnrner.
ces of waterways and in waters above the effluents of popu- lation centers. Water quality deteriorates abruptly below effluent points, creating altered zones, with degrees of alte- ration in many stretches which can be classified as "critical" (deplorable) and "very critical" (intolerable). It is also necessary to point out that studies reated in successive years report a definite worsening of the situation (PRAT et al . ,
1983; ALBA-TERCEDOR & PICAZO, 1990, 199 1 ; MUÑOZ & PRAT, in press).
ZONATION OF FLUVIAL COMMUNITIES
For the few works that have focused on the division os Spanish river commuriities into zones, we find two majos tendencies. The first is the exhaustive cataloguing of different communities based on the zonation proposed by ILLIES &
BOTOSANEANU (1963). The best exponents of this appro- ach in Spain have been GARCIA DE JALON & GONZALEZ DEL TANAGO (1986), working in the Duero Basin. In addi- tion, there have been other studies directed at a more restric- ted locations (GARCIA DE JALON & GONZALEZ DEL TANAGO, 1982; ALBA-TERCEDOR et al., 1986). The second tendency is the consideration of the community struc- ture of fluvial macroinvertebrates as being a longitudinal gradient of group and species substitution, in which the subs- titution hampers a clear division of the completely isolated macroinvertebrate communities characterizing precise stret- ches (VERNEAUX, 1973; VANNOTE, 1981). This approach uses different multivariant analyses (PCA and CCA), and has been the most used system since 1984 (GONZALEZ DEL TANAGO & GARCIA DE JALON, 198 1 ; GARCIA-ROJAS, 1985; GARCIA-AVILES, 1990).
Overall, the various studies published on zonation seem to indicate that the known Spanish fluvial systems allow us to differentiate their sources clearly, in contrast to the rest of the basin, where, strongly affected by the pollution in many cases, a gradient is found and therefore clear zonation is impeded (MILLET & PRAT, 1984; PUIG et al., 1987, 1991), except perhaps the Duero Basin (GARCIA DE JALON & GONZALEZ DEL TANAGO, 1986).
SPECIAL SYSTEMS (CASES RELATED TO SEASONALITY)
In spite of their singularity and their relative importance in Spanish geography, the seasonal rivers have not been considered irnportant until the middle of the 1980s (GARCIA-ROJAS, 1985; GROSSMAN et al., 1985; PRAT et al., 1985, 1986; PUIG et al., 1986, 1991; GARCIA- AVILES, 1990; ORTEGA et al. , 1991; SOLER, 1991; ROLDAN & PUIG, 1992; SOLER & PUIG, 1992 ). Even since then work has been insufficient, producing only 10 finished studies. The results obtained up to now, however, have refuted two ideas: the concept that diversity in these systems is less than that of permanent waterways (GARCIA-ROJAS, 1985; PUIG et al., 1991); and the idea that the structures of these communities are homogeneus.

V
Figure 9. Average annual valuea of water quality (BMWP' used).
This latter concept can be rejected simply with the obser- vation that seasonal river communities have been studied within three different typological groups defined in the section on community composition. Futhermore this subject is treated in depth in another chapter of this book.
FUTURE PERSPECTIVES
The tendency observed in the communications presented at the VI Spanish Congress on Limnology, showed the dominance of the works on biotic indices, although we can see a greater stant towards carrying our more complex studies on the community ecology of fluvial macrobenthos (fig. 4). Taken altogether, however, the number of works presented has been low (10.19%), and therefore at present Spanish limnology is dominated by studies related to lentic systems, as opposed to those focused on lotic systems.
The needs involved in the study of the macroinvertebrate communities is Spain include the exploration of large water- ways which are still completely unknown. This requires taxonomic work as a base. It must in fact approach a study of community ecology, scarcely begun and concentrated at the moment primarly on rates of colonization and displacement.
REFERENCES
ABELLA, M.A. & M.J. GONZALEZ, 1986. Variación estaciona1 de la fauna dulceacuícola del Alto Nalón, Astu- rias. Lirnnética 2: 173-179.
ALBA-TERCEDOR, J., 198 1. Recopilación de citas de Efemerópteros en la Península Ibérica e Islas Baleares. Trah. Monogr. Dep. Zool. Univ. Granada 4 (2): 41-81.
ALBA-TERCEDOR, J., L.F. CAPITAN, M. ESPIGA- RES, C. COCA, 1. GUISASOLA & J.M. MARTIN, 1990. Estudio de las condiciones ecológicas, sanitarias, qui- micas y de calidad de las aguas de la cuenca media-alta del río Guadalquivir. Universidad de Granada, Granada. 428 pp.
ALBA-TERCEDOR, J. & F. JIMÉNEZ-MILLAN, 198s. Evaluación de las variaciones estacionales de la calidad de las aguas del río Guadalfeo basada en el estudio de las comunidades de macroinvertebrados acuáticos y de los factores físico-químicos. ICONA Monograflas 48: 11-91
ALBA-TERCEDOR, J. & J. PICAZO, 1990. Calidad Biológica de las aguas de cauces de la margen izquierda de la Cuenca Alta del rio Guadalquivir, Proi'incias de Granada y Jaén. I.A.R.A., Granada. 178 pp
ALBA-TERCEDOR, J. PICAZO, 1991. Calidad Bioló- gica de las aguas de cauces de la margen izquierda de la Cuenca Alta del rio Guadalquivir: Provincias de Granada y Jaén, 11. I.A.R.A., Granada. 143 pp.
ALBA-TERCEDOR, J. & N. PRAT, (in press). Spanish experience in the use of macroinvertebrate as biological pollution indicators. Actas lnternational Conference on river water quality-ecological assessment and control.
ALBA-TERCEDOR, J . & A. SANCHEZ-ORTEGA, 1988. Un método rápido y simple para evaluar la calidad biológica de las aguas corrientes basado en el de Hellawell (1978). Limnética 4: 5 1-56.
ALBA-TERCEDOR, J., A. SANCHEZ-ORTEGA & 1. GUISASOLA, 1986. Caracterización de los cursos perma- nentes de agua de la cuenca del río Adra: Factores físico- quimicos, macroinvertehrados acuuticos y calidad de las uguas. Universidad de Granada, Granada. 54 pp.
ANTELO, J.M., F. FERNANDEZ, M.R. SOLORZANO & D. PRADO, 1990. Calidad de las aguas del río Anllons. Tecnologia del Agua 69.
ARLUZIAGA, 1. & J. ALZATE, 1984. Introducción a la ecología de los ríos guipuzcoanos. Lirnnética 1 : 2 14-22 1.
ARMITAGE, P.D., D. MOSS, J.F. WRIGHT & M.T. FURSE, 1983. The perfomance of a new biological water quality score system based on macroinvertebrates over a wide range of unpolluted running-water. Water Res. 17(3): 333-347.
ARMITAGE, P.D., 1. PARDO, M.T. FURSE & J.F. WRIGHT, 1990. Assessment and prediction of biological quality. A demonstration of a british macroinvertebrates- based method in two spanish rivers. Lirnnética 6: 147-156.

BALTANAS, A,, 1990. Estructura y organizac.ión de las comunidades de macroinvertebrados bentónicos del Alto Tajo: E.scula, patrones aleatorios y perturbación. Tésis Doctoral, Universidad Autonoma de Madrid. 331 pp.
BARGOS, D. & J.M. MESANZA, 1988. Variaciones estacionales de los grandes grupos taxonómicos de macroin- vertebrados bénticos en la red hidrográfica de Bizkaia. Actas 11 Congreso Mundial Vasco 11: 97-1 10.
CASADO,C., 1986. Composición y estructura de las comunidades de macroinvertebrados de un rio intensamente regulado del Sistema Central: río Lozoya (Cuenca del Tajo). Tesis Doctoral, Universidad Autónoma de Madrid. 580 pp.
CASTILLO, A., J. ALBA-TERCEDOR, L.F. CAPITAN- VALLVEY, L. CRUZ-PIZARRO & A. RAMOS, 1991. Ejemplo de un estudio interdisciplinar para la caracteriza- ción integral de la calidad y contaminación de las aguas de una cuenca de superficie. Actas 111 Simp. Agua Andalucia 11: 277-288.
COBO, F, M.JIMÉNEZ, J. CASAS, A. VILCHEZ, O. SORIANO, M. SEVILLANO, M. RIERADEVALL, 1. MUNOZ & N. PRAT, 1987. Inventario faunístico de los quironórnidos conocidos de la Península Ibérica. Alquihlu 12: 56-65.
CHANDLER, J.R., 1970. A biological approach to water quality rnanagement. Wat. Poll. Contr. 69: 415-422.
DOMEZAIN, A., 1. GUISASOLA & J. ALBA-TERCE- DOR, 1987. Estudio de la incidencia de una piscifactoría en las comunidades de macroinvertebrados acuáticos. Limné- tica 3: 151-157.
FANLO, E., S. MITJANS, R. UBEDA, P. CASALS & F . SOLEY, 1989. Influencia del sustrato en las comunidades de macroinvertebrados de los ríos del Moncayo. Turiaso 9: 597-616.
GALLARDO, I., 199 1. Respuesta de macroinvertebrudos fluviales a la salinidad. Tésis de Doctorado, Universidad de Sevilla. 125 pp.
GALLARDO, A. & J. TOJA, 1988. Efecto de la conta- minación orgánica en los macroinvertebrados acuáticos en la cuenca del río Guadaira (Sevilla, SW España). Actas Col. Luso-Esp. Ecol. Bac. Hidrogr. e Rec. Zoo1ogica.s: 163-170.
GARCIA-AVILÉS, J., 1990. Insectos acuáticos de Bale- ares (Odonata. Ephemeroptera, Heteroptera, Plecoptera y Coleoptera). Tesis Doctoral, Universidad Cornplutense de Madrid. 690 pp.
GARCIA DE JALON, D., 1980. Efectos del Embalse de Pinilla (Madrid) sobre las comunidades de rnacroinverte- brados benticos del río Lozoya. Boletin Esfación Central de Ecologia 9: 47-52.
GARCIA DE JALON, D. & M. GONZALEZ DEL TANAGO, 1982. Introducción a una zoosociología del rnacrobentos en los ríos de la Sierra de Guadarrama. Boletín de la Estucirín Central de Ecología 1 l(2 1): 63-7 1.
GARCIA DE JALON, D. & M. GONZALEZ DEL TANAGO, 1986. Métodos biológicos para el estudio de la calidad de las aguas. Aplicación a la cuenca del Duero. ICONA, Madrid. 241 pp.
GARCIA DE JALON, D. & M. GONZALEZ DEL TANAGO, 1986. Efemeroptera, Plecoptera y Trichoptera de los principales ríos de Malaga. Actas Simposium el Agua en Andaluc,ia: 33 1-346.
GARCIA DE JALON, D., M. GONZALEZ DEL TANAGO, E. BARCELO, C. MONTES, F. MENES & C. CASADO, 1986. Contribución al estudio faunístico de algu- nas taxocenosis de insectos acuáticos (Ephemeroptera, Plecoptera, Trichoptera, Coleoptera y Heteroptera) del Parque Natural del Lago de Sanabria y Río Tera (Zamora). Actas de las VIII .Tornadas As. esp. Limn. 1.145-1 156.
GARCIA DE JALON, D., C. MONTES, E. BARCELO, C. CASADO & F. MENES, 1988. Impacto ecológico de la regulación hidroeléctrica en ríos del Pirineo aragones. Actas del Congreso 11 Congreso Mundial Vasco 2: 140- 162.
GARCIA DE JALON, D., C. MONTES, C. CASADO & F. MENES, 1988. Effects of hydroelectric scherne on fluvial ecosystems within the spanish pyrenees. Reguluted Rivers 2: 479-49 1 .
GARCIA-ROJAS, A.M., 1985. Estudio de los insectos acuáticos del río Yeguas. Tesina de Licenciatura, Universi- dad de Córdoba. 23 1 pp.
GONZALEZ, F., A. ALVAREZ-BRYLLA, A.
MIRANDA, J.R. ALONSO & M.A. ABELLA, 1986 a. Estudio comparativo de la calidad del agua en dos ríos astu- rianos. Limnética 2: 2 17-223.
GONZALEZ, F., A. ALVAREZ-BRYLLA, A. MIRANDA, J.R. ALONSO & M.A. ABELLA, 1986 b. La calidad del agua en la cuenca del rio Piloña (Asturias). Limnética 2: 225-234.
GONZALEZ, G., X. MILLET, M.A. PUIG & N. PRAT, 1985. Patterns of the macroinvertebrate distribution in the Llobregat river Basin (NE-Spain). Verh. inter. Verein. Limnol. 22(4): 208 1-2086.
GONZALEZ, M.C., M.P. LANZA, M.P. ALVAREZ & Y. PRESA, 1986. Análisis de los macroinvertebrados bentó- nicos y su helinintofauna en el río Porrna (León, NO España). Limnética 2: 18 1-1 87.
GONZALEZ DEL TANAGO, M., 1978. Ephemeroptera, Odonata y Plecóptera del Coto Nacional de las Sierras de

Cazorla y Segura. Tésis Doctoral, Universidad Politécnica de Madrid. 172 pp.
GONZALEZ DEL TANAGO, M. & D. GARCIA DE JALON, 1980. Estimación de la contaminación de las aguas
nzediatzte indicadores biológicos: Aplicación al río Jarama. Icona, Madrid. 156 pp.
GONZALEZ DEL TANAGO, M. & D. GARCIA DE JALON, 1981. Estudio de la estructura biotipológica del río Jarama. Boletín de la Estación Central de Ecología 19: 33-51.
GONZALEZ DEL TANAGO, M. & D. GARCIA DE JALON, 1984. Desarrollo de un índice biológico para esti- mar la calidad de las aguas de la Cuenca del Duero. Limné-
tica 1: 263-272. GONZALEZ DEL TANAGO. M. & D. GARCIA DE
JALON, 1989. Elaboración y aplicación de índices biológi- cos. In: W. Lockhart (ed.), Proceedings of the fir-sf Symp. on the ~ t u d y , conservation and rational use of Salmonidar Fisheries, pp. 65-81. Madrid.
GONZALEZ DEL TANAGO, M., D. GARCIA DE JALON & I.M. ELCORO, 1979. Estudio sobre la fauna de macroinvertebrados de los ríos Cigüela, Zancara y Corco- les: Aplicación de indices biológicos para el estudio de la calidad de sus aguas. Boletín de la Estación Central de Ecología 8(15): 45-59.
GROSSMAN, G.D., A. DE SOSTOA, J. LOBON- CERVIA & M.A. PUIG, 1985. Ecological-eni~ironmentul relationships in some Río Matarraña fish a.ssemblages.
Comité Hispano-Norteamericano de Cooperación Científica y Técnica, Madrid. Interna1 Repport, 46 pp.
HERRANZ, J.M., 1983. Estudio de la fauna macroinver- tehrada de la cuenca del Alto Tajo. Tésis Doctoral, Univer- sidad Politécnica de Madrid. 464 pp.
HERRANZ, J.M. & M. GONZALEZ DEL TANAGO, 1986. La colonización de sustratos artificiales por macroin- vertebrados benticos en las aguas del Alto Tajo. Compara- ción de métodos de muestreo. Limnética 2: 163-17 1.
ILLIES, J. & L. BOTOSANEANU, 1963. Problemes et méthodes de la classification et de la zonation écologique des eaux courantes, considerées surtout du point de vue faunistique. Mitt. inr. Verein. theor. angew. Limnol. 12: 1-57.
IMBERT, J.B. & J. POZO, 1989. Breakdown of four leaf litter species and associated fauna in a Basque Country forested stream. Hydrobiologia 182: 1-14.
LATORRE, E., 1987. La fauna de artrópodos acuáticos de la cuenca del río Razón (Soria). Tésis Doctoral, Univer- sidad Complutense de Madrid. 160 pp.
LOPEZ-LLANEZA, J., 1984. Estudio de la calidad del agua en el río Nalón y su cuenca. Consejería de Ordena-
239
ción del Territorio, Vivienda y Medio Ambiente, Oviedo. 127 pp.
LUIS, E., V. ABAD, E. BAYON. F. FERNANDEZ, A. GUTIERREZ, A. HERNANDEZ, C. POLANCO áC M.M. POSTIGO, 1986. Calidad estival de las aguas en los ríos influenciados por la ciudad de León. Importancia de bioin- dicadores artrópodos. Actas de las VI11 Jornadas Aso. esp.
Limn.: 1167-1 178. MADRID, F., 1990. Factores ,físico-químicos y comuni-
dudes de macr-oinvertebrados de la cabecera del Río Genil (Sierra Nevada), aguas arriba de Granada. Estudio de la
calidad biológica de sus aguas. Tesina de Licenciatura,
Universidad de Granada. 198 pp. MALO, J. & M.A. PUIG, 1992. Efecto de las fluctua-
ciones sobre la estabilidad de la comunidad bentónica en un tramo permanente de un cauce mediterráneo (Río Mata- rraña, Cuenca del R. Ebro). Actas VI Congreso As. esp.
Limnología. MANZANERA, M.A. & M.P. ALVAREZ, 1987. Estudio
de las comunidades de macroinvertebrados bentónicos y su
helmintofauna en un tramo del río Omañas (León, NO. de España). Limnética 3: 133-139.
MARGALEF, R., 1946. Contribución al conocimiento hidrobiológico del país vasco-navarro. In: Margalef, R., M.R. de San Miguel & J. Rodríguez-Roda (eds.) Aportacio-
nes al estudio de la fauna y jlora vasco-navarr-as (Siera de Aralar). CSIC, Zaragoza.
MARGALEF, R., 1947. Limnosocio1ogía.Monografías de Ciencias Modernas, Madrid. 93 pp.
MARGALEF, R., 1948. Flora, fauna y comunidades bióticas de las aguas dulces del Pirineo de la Cerdaña. Monogr. Est. Pirenáicos 11.
MARGALEF, R., 1949. Datos para la hidrobiología de la Sierra de Guadarrama. P.lns. Biol. Apl. 6 : 5-21.
MARGALEF, R., 1952. La vida en las aguas dulces de Andorra. Actas del Primer Congreso Internacional del Piri-
neo : 5-107. MARGALEF, R., 1953. Algunos organismos interesan-
tes de las aguas dulces de los Pirineos. P. Ins. Biol. Apl. 28: 407-420.
MARGALEF, R., 1955. Comunidades bióticas de las aguas dulces del noroeste de España. P. Inst. Biol. Apl. 21: 5-85.
MARGALEF, R., 1960. Ideas for a synthetic approach to the ecology of running waters. Int. Revue ges. Hydrohiol. 45: 133-153.
MARGALEF, R., 1969. El concepto de polución en limnología y sus indicadores biológicos. Documentos Inv. Hidr-o[. 7 : 103-1 33.

MEYNELL, P.J., 1973. A hydrobiological survey of a small Spanish river grossly polluted by oil refinery and
petrochemical wastes. Freshwat. Biol. 3: 503-520. MILLET, X. & N. PRAT, 1984. Las comunidades de
macroinvertebrados a lo largo del río Llobregat. Limnéticu
1: 222-233. MIRANDA, A., 1987. Utilización de macroinvertebrados
bénticos como indicadores biológicos de la calidad del agua
en el río Viao-Piloña (Asturias). Limnética 3: 141- 150.
MONTES, C. & A.C. SOLER, 1986. Lista faunística y bibliográfica de los coleópteros acuáticos Dryopoidea (Dryopidae & Elmidae) de la Peninsula Ibérica e 1sla.s Baleares. Asociación española de Limnología. 38 pp.
MUÑOZ, 1. & N. PRAT (in press). Cambios en la cali-
dad del agua de los ríos Llobregat y Cardener en los últimos diez años. Tecnología del Agua.
NIESER, N. & C. MONTES, 1984. Lista faunística y
Bibliográfica de los Heterópteros acuuticos (Nepomorpha
& Gerromorpha) de España y Portugal. Asociación espa- ñola de Limnología. 66 pp.
ORTEGA, C., 1990. La calidad del agua en la red
fluvial de Cantabria. Tésis Doctoral, Universidad de Oviedo. 496 pp.
ORTEGA, M., 1988. La Rumbla del Moro (Cuenca del Río Segura). Ambiente físico, biológico y alteraciones
producidas por una riada. Tesina de Licenciatura, Univer-
sidad de Murcia. 224 pp. ORTEGA, M., M.L. SUAREZ, M.R. VIDAL-ABARCA,
R. GOMEZ & L. RAMIREZ-DIAZ, 1991. Aspects of post-
flood recolonization of macroinvetebrates in a "Rambla" of South-East Spain ("Rambla del Moro": Segura River Basin).
Verh. Internat. Verein. Limnol. 24: 1994-2001. PALAU, A. & A. PALOMES, 1986 a. Los macroinverte-
brados bentónicos como elementos de juicio para la evalua- ción de la calidad biológica del río Segre (Lerida, España).
Limnética 2: 205-2 15. PALAU, A. & A. PALOMES, 1986 b. Diagnrísticofísico-
quimico y biológico del rio Segre. Escuela Técnica Superior de Ingenieros Agrónomos, Lerida. 500 pp.
PINKSTER, S., 1988. Changes in the condition of basque rivers during the last 15 years. Actas 11 Congreso Mundial
Vasco 11: 133-143. PRAT, N., M.I. BAUTISTA, G. GONZALEZ & M.A.
PUIG, 1979. Els cursos d'aigua. In: R. Folch (ed.) Elpatrimoni natural d'Andorra, pp. 261-309. Ketres, Barcelona. 446 pp.
PRAT, N,, G. GONZALEZ & X. MILLET, 1986. Comparación critica de dos índices de calidad del agua: ISQA y BILL. Tecnología del Agua 31: 33-49.
PRAT, N., G. GONZALEZ, X. MILLET & M.A. PUIG, 1983. Chironomid longitudinal distribution and macroin-
vertebrate diversity along the river Llobregat (NE Spain). Mem. Amer. Ent. Soc. 34: 267-278.
PRAT, N., G. GONZALEZ, X. MILLET & M.A. PUIG, 1985. El Foix entre I'eiuutesu i la confaminació. Servei del
Medi Ambient, Diputació de Barcelona. 92 pp. PRAT, N., G. GONZALEZ, 1. MUÑOZ & X. MILLET,
1986. Community structure and colonization process in a mediterranean drainage basin. Proceedings of the 3rd Euro- pean Congress oj' Entomology: 12 1 - 124.
PRAT, N,, M.A. PUIG & G. GONZALEZ, 1983. Predic- ció i control de la qualitat de les aigües dels r i ~ s Besós i
Llobregat, 11: El poblament faunístic i la seva relació amb la qualitat de les aigües. Servei del Medi Ambient, Diputa- ció de Barcelona. 164 pp.
PRAT, N,, M.A. PUIG, G. GONZALEZ, M.J. TORT &
M. ESTRADA, 1983. The Llobregat: A mediterranean river fed by the Pyrenees. In: B.A. Whitton (ed), Ecology of euro- pean rivers. pp. 527-552. Blackwell, London. 644 pp.
PRESA, Y., E. LUIS & J. SOTO, 1988. Consideración de los macroinvertebrados en la zonación de la cuenca del río Orbigo (León). Actas del 11 Congreso Mundial Vasco 2: 207-215.
PUIG, M.A., 1992. Relaciones tróficas de la comunidad de macroinvertebrados en el río Matarraña (Cuenca del
Ebro). Actas VI Congreso As. esp. Limnología. PUIG, M.A., M. ABOAL & A. DE SOSTOA, 1991. New
approaches to mediterranean fluvial communities. Oecolo- gía aquática 10.
PUIG, M.A., J. ARMENGOL, G. GONZALEZ, J. PEÑUELAS, S. SABATER & F. SABATER, 1987. Chemi- cal and biological changes in the Ter River induced by a series of reservoirs. In: J.F. Craig & J.B. Kemper (eds)
Regulated Streums: Advances in Ecology. pp. 373-382. Plenum Press, New York. 421 pp.
PUIG, M.A., M. FERRERAS-ROMERO & A.M. GARCIA-ROJAS, 1986. Ecosistemas de ríos temporales:
ecología de las poblaciones de efemerópteros de la cuenca del río Bembezar (Sierra Morena). Anales de Biología 8: 65-69.
PUIG, M.A., G. GONZALEZ & L. RECASENS, 1987. Módelos de distribución de Plecópteros, Efemerópteros, Tricópteros y Simúlidos en el río Ter. Limnética 3:
125-132. PUIG, M.A., G. GONZALEZ & O. SORIANO, 1984.
Introducción al estudio de las comunidades de los ríos astu- rianos: Efemerópteros, Plecópteros, Tricópteros, Simúliidos y Quironómidos. Limnética l(1): 187-196.

PUIG, M.A., J. MALO, F. CASALS, M. ABOAL, D. VINYOLAS & A. DE SOSTOA, 1990. Comunidades acuá- ticas de la Nogueia Palluresa: estado actual y cambios asociados a la gestión hidroeléctrica de los cauces. (PIE 121.43). Barcelona. 158 pp.
RALLO, A., M.A. SEVILLANO, M. OJEA, E. RICO, L. DOCAMPO & J.C. ITURRONDOBEITIA, 1988. Niveles d calidad del agua en las distintas cuencas fluviales vizcainas: Clasificación obtenida por estudio faunístico de diversos taxones animales y en dos épocas del año. Actas II Congreso Mundial Vasco 11: 217-222.
RICO, E., L.C. PEREZ & C. MONTES, 1990. Lista faunística y bibliográfica de los Hydradephaga (Coleop- tera: Haliplidae, Hygrobiidae, Gyrinidae, Noteridae, Dytis- cidae) de la Península Ibérica e Islas Baleares. Asociación española de Limnología. 2 16 pp.
RICO, E., A. RALLO, M.A. SEVILLANO & M.L. ARRETXE, (in press). La calidad de las aguas de las redes fluviales de Alava y Guipuzcoa (N. España). Estudio comparado de diversos índices biológicos. Annales de Limnologie.
RIERADEVALL, M. & N. PRAT, 1986. Deriva nicte- mera1 de macroinvertebrados en el río Llobregat (Barce- lona). Limnética 2: 147-156.
RODRIGUEZ, P. & J.F. WRIGHT, 1991. Description and evaluation of a sampling strategy for macroinvertebrate communities in Basque rivers (Spain). Hydrobiologia 213: 113-124.
ROLDAN, A. & M.A. PUIG, 1992. Influence of aquatic invertebrate on the aquatic hyphomycetes occurring on leaves in a Mediterranean stream. Annales de Limnologie.
SANCHEZ-ORTEGA, A. & J. ALBA-TERCEDOR, 1987. Lista faunística y bibliográfica de los Plecópteros (Plecoptera) de la Perzínsula Ibérica. Asociación española de Limnología. 133 pp.
SLADECEK, V., 1966. Water quality system. Verh. Intern. Verein. Limnol. 16: 809-816.
SOLER, A. (ED.), 1984. Estudios y directrices para el Saneamiento del Río Segura. Comunidad Autónoma de la Región de Murcia, Murcia. 473 pp.
SOLER, G., 1991. Pautas de migración de las comu- nidades bentónicas del río Matarraña (Cuenca del Ebro) a lo largo de un ciclo anual y en función de la velocidad de la corriente y la heterogeneidad del sustrato coloni- zable. Tesina de Licenciatura, Universidad de Murcia.
91 PP. SOLER, G. & M.A. PUIG, 1992. Similitud y diferencias
de las pautas de colonización en tramos permanentes y
temporales de un río mediterráneo (R. Matarraña, Cuenca del Ebro). Actas VI Congreso As. esp. Limnología.
TUFFERY, G. & J.Q. VERNEAUX, 1967. Une metode zoologique practique de determination de la qualité biologi- que des eaux courantes. Indices biotiques. Ann. Scient. Univ. Besacon zoologie 3: 79-90.
UNIVERSIDAD DE GRANADA, 1990. Caracterización físico-químico-biológica de las aguas del Alto Genil. Estu- dio Integral de la calidad y contaminación de las aguas. Universidad de Granada, Granada. 278 pp.
VALDECASAS, A., 1988. Lista sinonímica y bibliográ- fica de las Hidracnelas (Acari, Hydrachnellae) de la Penín- sula Ibérica, Islas Baleares e Islas Canarias. Asociación española de Limnología. 8 1 pp.
VALDECASAS, A. & A. BALTANAS, 1990. Jacknife and bootstrap estimation of biological index of water quality. Wat. Res. 24(10): 1279-1283.
VALDECASAS, A., E. BELLO, E. REYES & J. BECE- RRA, 1990. DIRTAX: A taxonomist database. Science Soft- ware 5(4): 303-308.
VALLADARES, L.F. & C. MONTES, 1991. Lista faunísticu y bibliográfica de los Hydraenidae (Coleoptera) de la Península Ibérica e Islas Baleares. Asociación espa- ñola de Limnología. 93 pp.
VANNOTE, R.L., G.M. MINSHALL, K.W. CUMMINS, J.R. SEDELL & C.E. CUSHING, 1981. The river conti- nuum concept. Can. J. Fish. Aqlaaí. Sci. 37: 130-1 37.
VERNEAUX, J., 1973. Recherches écologiques sur le réseau Hydrographique du Doubs. Essai de biotypologie. Ann. Sc. Univ. Besancon 9: 1-260.
VIDAL-ABARCA, C. & M.L. SUAREZ, 1985. Lista faunística y bibliográfica de los wroluscos (Gastropoda & Bivalviuj de las aguas continentales de la Penínsz~la Ibérica e Islas Balearrs. Asociación española de Limnología. 190 pp.
ZAMORA-MUNOZ, C., 1988. Caracterización y calidad de las aguas del río Monachil (Sierra Nevada, Granada). Factores físico-químicos y comunidades de macroinverte- brados acuáticos. Tesina de Licenciatura, Universidad de Granada. 173 pp.
ZAMORA-MUÑOZ, C., 1992. Macroinvertehrados acuáticos, caracterización y calidad de las aguas de los cauces de lu cuenca alta del Río Genil. Tésis Doctoral, Universidad de Granada. 255 + 109 pp.
ZAMORA-MUNOZ, C. & J. ALBA-TERCEDOR, 1992. Caracterizucin'tz y calidad de las Aguas del rio Monachil (Sierra Nevada, Granada). Factores físico-químicos y com~tnidades de macroinvertebrados acuáticos. Junta de Andalucia, Agencia del Medio Ambiente, Granada.

BIOGEOGRAPHY OF THE FRESHWATER FISH OF THE IBERIAN PENINSULA
J. A. Hernando and M. C.-Soriguer Faculty of Marine Science, University of Cádiz. Biology. Ap. de Correos no. 40. 11510 Puerto Real. Cádiz, Spain.
Keywords: Iberian fish, Distribution, Biogeography, Allochthonous species, Sectorisation, Iberian peninsula, .
ABSTRACT
This paper reviews and presents new data on the composition and distribution of fish species in the continental waters of the Iberian Peninsula, and proposes a division of the Peninsula into three subregions: the Ebro-Cantabrian, the Atlantic and the
Betico-Mediterranean, based on the distribution of the 45 species (native and endemic) in the 22 river basins with surface aseas of 990 square kilometres os more.
INTRODUCTION
The lberian Peninsula is enormously interesting from an ichthyological point of view. Located at the Southwestern tip
of Europe, it is a point of contact between North Atlantic and tropical coastal fish fauna. Much of its coastline is on the
Mediterranean, the Straits of Gibraltar, which separate the
Mediterranean from the Atlantic, have always been more of a bridge than a barrier for North African and Iberian fauna.
The epicontinental fish fauna has also been affected by
the geological history of the Peninsula. Possibilities for
dispersion have been, and still are, limited, due to the isola- tion caused by the Pyrenees and the Straits of Gibraltar. The number of species relative to the surface asea is lower than in the rest of Europe, leading to a comparatively less diverse
ichthyofauna. Isolation, along with the orographic and
climatic peculiarities of the Peninsula, has led to the deve- lopment of endemisms. Most of the large rivers flow East- West, to the Atlantic, and the variable rainfall means that many of the rivers are torrential, while others are confined to
small survival areas in the long dry season (June - October). Al1 of this means that extreme life conditions are imposed on the animals which live in the rivers, favouring the deve-
lopment of adaptation mechanisms.
From the point of view of the ecological characteristics of the rivers, the ecosystems forming a frontier between sea and freshwater should not be forgotten: estuaries, wetlands, coastal lagoons, marshlands and salt marshes, which for different periods, experience the alternating influences of
sea and freshwater. The ecosystems of variable salinity lead us to consider marine species as forming part of the Iberian continental fauna.
THE ICTHYOFAUNA OF THE IBERIAN PENINSULA
MYERS (1960) classified freshwater fish into three cate- gories: primary, secondary and peripheral (vicariant, diadro- mous, sporadic and complementary). Under this classifica-
tion, the continental fauna of the Iberian Peninsula consists of 64 species in 24 families (table 1). Of these, 45 are autochthonous (native os endemic) and 19 allochthonous, with differing degrees of acclimatization (SOSTOA et al., 1984; ICONA, 1986). To these we should add 18 sporadic
os sedentary marine species which thrive in the estuaries, marshlands, coastal lagoons, marshes and salt flat ecosys- tems, and belong to 16 marine families (table 2), raising the number of species to 98.
The taxonomy of the species has been studied by ALMACA (1964, 1967, 1972), COLLARES-PEREIRA ( 1 983), DOADRIO ( 1 984, 1987) and ELVIRA ( 1 987), and their distribution in some river basins by HERNANDO
(1975a, 1975b), DEMESTRE et al., (1977), GARCIA DE JALON & GONZALEZ DE TANAGO (1983), GARCIA DE JALON & LOPEZ (1983), FERNANDEZ-DELGADO et al., (1986), LOBON-CERVIA rt al., (1989) and SOSTOA
& LOBON-CERVIA (1989).
Linirietica, 8: 243-253 (1992) O Asociación Española de Limnologia, Madrid. Spain
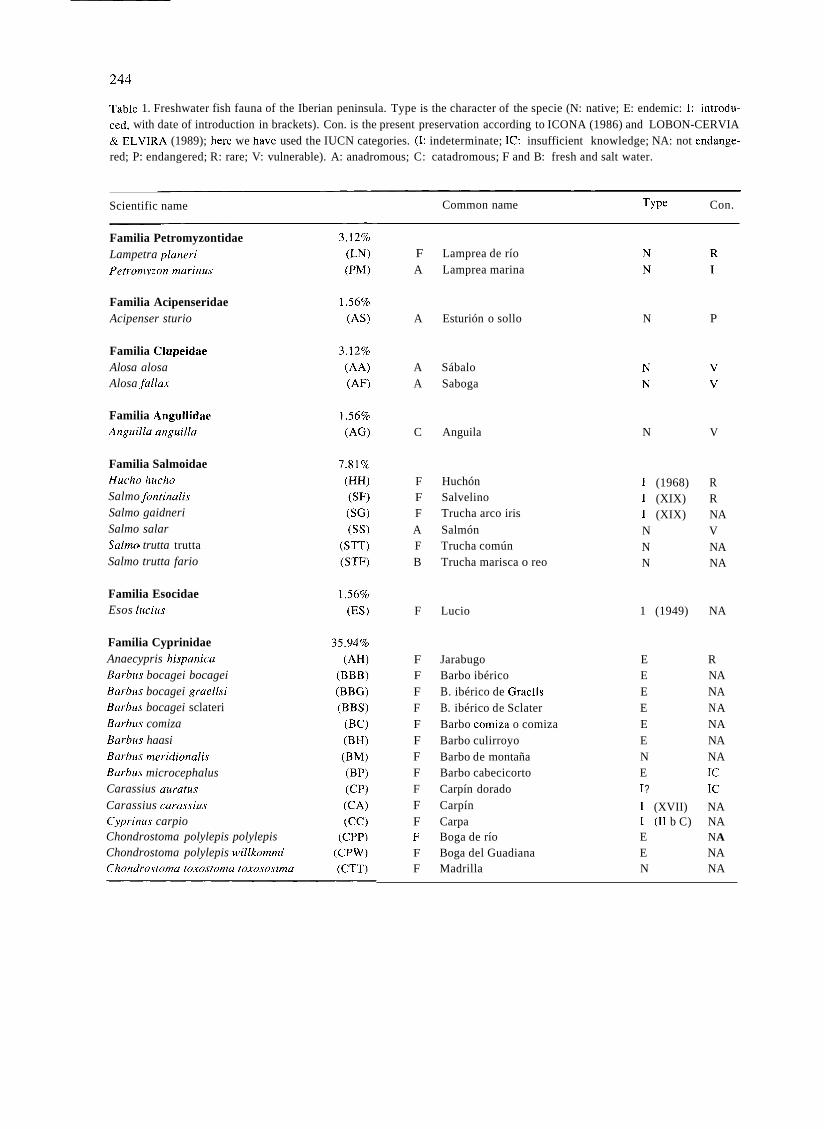
Table 1. Freshwater fish fauna of the Iberian peninsula. Type is the character of the specie (N: native; E: endemic: 1: introdu- ced, with date of introduction in brackets). Con. is the present preservation according to ICONA (1986) and LOBON-CERVIA & ELVIRA (1989); here we have used the IUCN categories. (1: indeterminate; IC: insufficient knowledge; NA: not endange- red; P: endangered; R: rare; V: vulnerable). A: anadromous; C: catadromous; F and B: fresh and salt water.
-
Scientific name
Familia Petromyzontidae Lampetra planeri Petromyzon marinus
Familia Acipenseridae Acipenser sturio
Familia Clupeidae Alosa alosa Alosa fallax
Familia Angullidae Ang~~il la anguilla
Familia Salmoidae Hucho hucho Salmo fontiriulis Salmo gaidneri Salmo salar Salmo trutta trutta Salmo trutta fario
Familia Esocidae Esos lucius
Familia Cyprinidae Anaecypris hispanica Barbus bocagei bocagei Barbus bocagei graellsi Barbus bocagei sclateri Barbus comiza Barbus haasi Barbus meridionalis Barbus microcephalus Carassius auratus Carassius carassius Cyprinus carpio Chondrostoma polylepis polylepis Chondrostoma polylepis willkommi Chorzdrostoma to,uostoma toxosostma
Common name T Y P ~ Con.
F Lamprea de río A Lamprea marina
A Esturión o sollo N P
A Sábalo A Saboga
C Anguila N V
F Huchón F Salvelino F Trucha arco iris A Salmón F Trucha común B Trucha marisca o reo
1 (1968) R 1 (XIX) R 1 (XIX) NA N V N N A N NA
F Lucio 1 (1949) NA
F Jarabugo E R F Barbo ibérico E N A F B. ibérico de Graells E N A F B. ibérico de Sclater E N A F Barbo comiza o comiza E N A F Barbo culirroyo E N A F Barbo de montaña N N A F Barbo cabecicorto E IC F Carpín dorado 1 ? IC F Carpín 1 (XVII) NA F Carpa 1 (11 b C) NA F Boga de río E N A F Boga del Guadiana E N A F Madrilla N N A
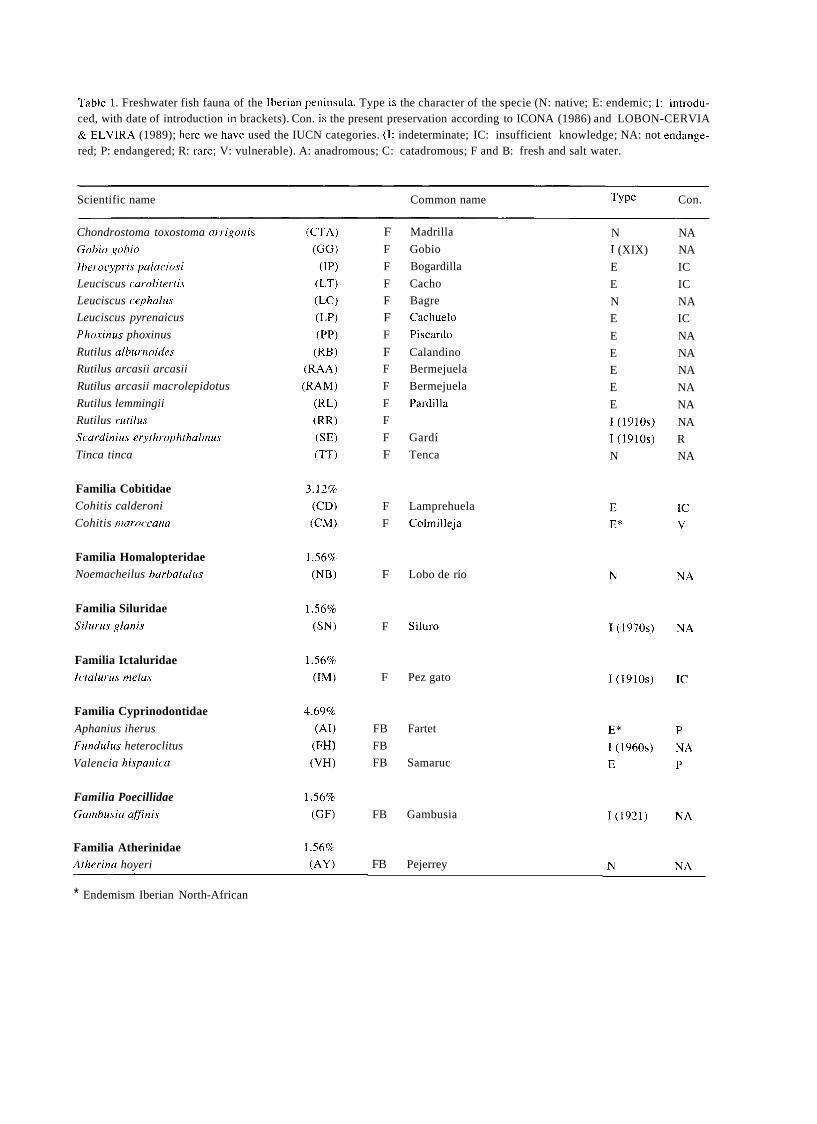
Table 1. Freshwater fish fauna of the lberian peninsula. Type is the character of the specie (N: native; E: endemic; 1: introdu- ced, with date of introduction in brackets). Con. is the present preservation according to ICONA (1986) and LOBON-CERVIA & ELVIRA (1989); here we have used the IUCN categories. (1: indeterminate; IC: insufficient knowledge; NA: not endange- red; P: endangered; R: rase; V: vulnerable). A: anadromous; C: catadromous; F and B: fresh and salt water.
Scientific name Common name T Y P ~ Con.
Chondrostoma toxostoma arrigonis
Gobio gobio
Iherocypris palaciosi
Leuciscus carolitertis
Leuciscus cephalus
Leuciscus pyrenaicus
Phoxinus phoxinus
Rutilus alburnoides
Rutilus arcasii arcasii
Rutilus arcasii macrolepidotus
Rutilus lemmingii
Rutilus rutilus
Scardinius erythrophthalmus
Tinca tinca
F Madrilla
F Gobio
F Bogardilla
F Cacho
F Bagre
F Cachuelo
F Piscardo
F Calandino F Bermejuela
F Bermejuela
F Pardilla
F
F Gardí
F Tenca
N NA
1 (XIX) NA
E IC
E IC
N N A
E IC
E N A
E N A
E N A
E N A
E NA
I(1910s) NA
I(1910s) R
N NA
Familia Cobitidae Cohitis calderoni
Cohitis rnuroccana
F Lamprehuela
F Colmilleja
Familia Homalopteridae Noemacheilus barbatulus F Lobo de río
Familia Siluridae Silurcls glanis F Siluro
Familia Ictaluridae Ictalurus melas F Pez gato
Familia Cyprinodontidae Aphanius iherus
Fundulus heteroclitus
Valencia hispanica
FB Fartet
FB
FB Samaruc
Familia Poecillidae Gumhusiu affinis FB Gambusia
Familia Atherinidae Atherinu hoyeri FB Pejerrey
* Endemism Iberian North-African
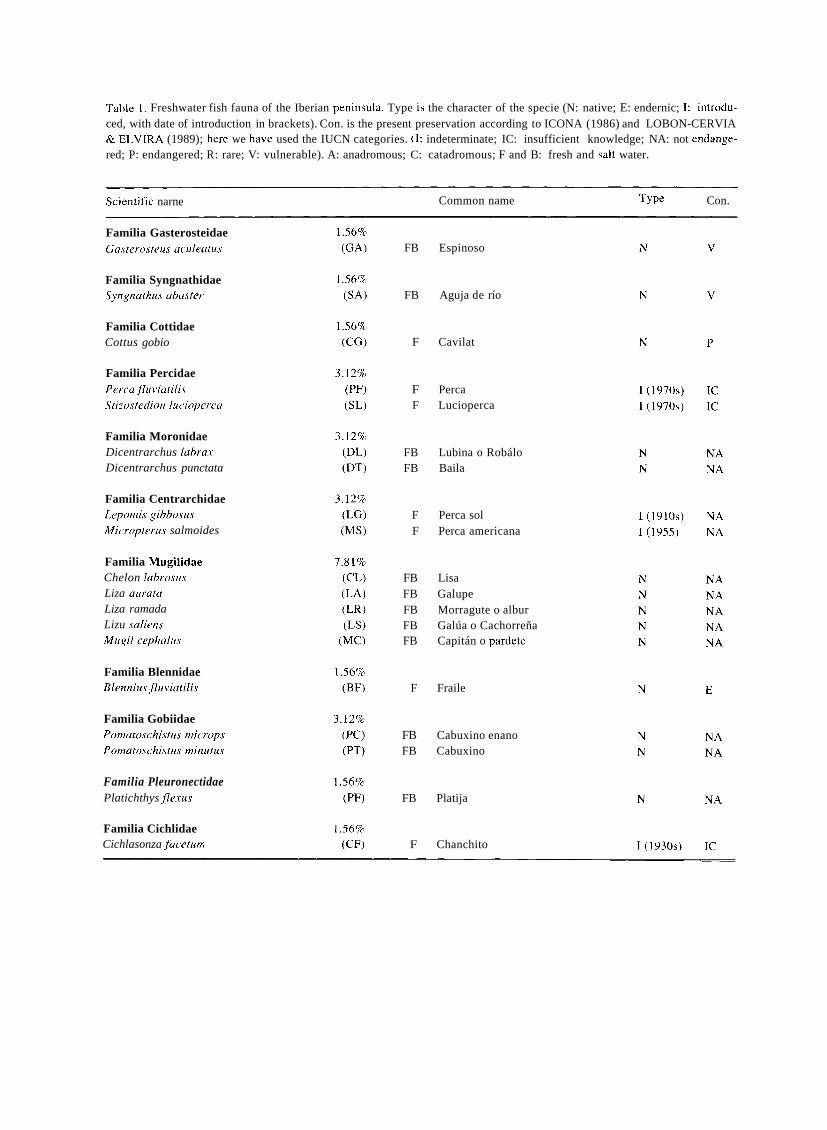
Table l . Freshwater fish fauna of the Iberian peninsula. Type is the character of the specie (N: native; E: endernic; 1: introdu- ced, with date of introduction in brackets). Con. is the present preservation according to ICONA (1986) and LOBON-CERVIA & ELVIRA (1989); here we have used the IUCN categories. (1: indeterminate; IC: insufficient knowledge; NA: not endange- red; P: endangered; R: rare; V: vulnerable). A: anadromous; C: catadromous; F and B: fresh and salt water.
- - - -
Scientific narne Common name T Y P ~ Con.
Familia Gasterosteidae Casterosteus acilleatus
Familia Syngnathidae Syrrgnatlrus ahuster
Familia Cottidae Cottus gobio
Familia Percidae Perca flui~iatilis Stizostediori luc.iopercu
Familia Moronidae Dicentrarchus lahrax Dicentrarchus punctata
Familia Centrarchidae Leponzis gihhosus Micropter~ls salmoides
Familia Mugilidae Chelon lahrosus Liza auratu Liza ramada Lizu salierzs Mugil cephal~rs
Familia Blennidae Blennius flur~iatilis
Familia Gobiidae Pomatoschistus rnicrops Pomatoschistiis minurus
Familia Pleuronectidae Platichthys ,flexus
Familia Cichlidae Cichlasonza fucetum
FB Espinoso
FB Aguja de río
F Cavilat
F Perca F Lucioperca
FB Lubina o Robálo FB Baila
F Perca sol F Perca americana
FB Lisa FB Galupe FB Morragute o albur FB Galúa o Cachorreña FB Capitán o pardete
F Fraile
FB Cabuxino enano FB Cabuxino
FB Platija
F Chanchito

The Cyprinidae genera which inhabit the southern provin- ces originated in Siberia, and arrived in Europe through the
drying out of the Sea of Obi during the Holocene period, the time of Alpine orogenesis, when the Pyrenees were formed. During the same period, the Oligocenic transgression divi-
ded Europe allowing communication of the North Sea and the Tethys Sea via Southern Russia (TERMIER, 1960).
Both of these created a great barrier to the arrival of freshwater fish in the Iberian Peninsula (MYERS, 1960; BANARESCU, 1972), and therefore the colonization of the
Peninsula must have happened before the end of the Oligo-
cene. The Siberian immigrants (BANARESCU, 1973a, 1973b)
are of the genera Pseudopholtinus (Phoxinell~*.s), Tropidop- ho.~inellus, Tinca, Chondrostoma, Rut i l i~~s and Leucisus; and the Barhus from East Asia. The differentiation of species such as Barhus meridionalis appears to arise in to this
period, while the Iberian endemisms of post-Oligocenic
formation, possibly during the Pontiense, form two groups. The first, with geographical and ecological incompatibilities
with the North African Cyprinidae, comprises the species Barhus hocugei graellsi, Barhus hocagei hocagei, Chron- drostoma polyepis polypepis and Chrosdrostoma tosostoma arrigonis and the second group, possibly formed well into the Pontiense, and which is geographically and ecologically compatible with the North African Cyprinidae, includes
Barhus harhus sclarteri. Barbus comiza, Barhus microcep-
halus, Chondrostoma polyepis willkommi, Leuc,iscus pyre- naicus, Rutilus arcasi, Rutilus lemmingii and Rutilus alhur- noides.
According to ALMACA (1976) and ELVIRA (1986), the Rutilus genus, still of uncertain taxonomy due to the inter-
mediate features of the population at both species and genus levels, may represent a speciation os sub-speciation.
Cobitidae and Homalopteridae are of Euro-Mediterranean
origin. In the case of Cobitidae a speciation may have occu-
rred in the Peninsula, in such a way that one species is rela- ted to the Euro-Mediterranean and the other endemism to the North African fauna (Cohitis maroccana), underlining the fact that both are considered endemisms, the formes
Iberian and the latter Iberian-North African.
THE CYPRINIDAE GENERA
The Barbus genera has been considered to include Barbus harbus, Barhus comiza and Barhus meridionalis, conside- ring the first and last having two subspecies: B. h. hocagei,
B . h . sclateri, B. m. meridionalis and B. rrr. graellsi
(LOZANO, 1935). Later, ALMACA (1967), revising the genera, considered that it was necessary to raise some subs-
pecies such as B. sclateri, B. hocagei, B. graellsi. B. cornizu was maintained, and two new species were defined: R. microcephalus and B. steindachner-i. Finally, DOADRIO
(1984) changed the taxonomy, classifying five species, B. hocugei, B. comiza, B. haasi, B. microcephal~~s and B.meri- dionalis, al1 without subspecies except for the first, the Iberian barbel, which he divided into 3 subspecies: B. h.
hocagei, B. h. graellsi und B. h. sclateri.
As for zoogeographical origin, ALMACA (1984) divides Iberian barbels into two groups: 1 (hocagei, graellsi and sclateri), together with North African and Asian barbels, and 11 (comiza, wiicrocephalu.s and sreindachneri) related to Western Asian and North African barbels.
There are two species of the Chondro.stoma gerius in the Iberian Peninsula, Ch. polylepis and Ch. toxostoma. The
first is endemic, and is distributed throughout tlie Atlantic
river basins, while the second is native, with a Cantabrian-
Mediterranean distribution. Each is divided into two subspecies: parental (Ch. p.
polylepis and Ch. t . tosostoma) and Ch. p. w~illkommi and Ch. t . arr-igonis. They have very different distributions: the parental subspecies (Ch. p. polylepis and Ch. t . tosostowiu) are to be found in the north, and Ch. p. ~lillikommi and Ch. t . ar-rigonis in the south (ELVIRA, 1987).
The Rutilus genus has so far had a more uncertain classi-
fication (COLLARES-PEREIRA, 1984; ELVIRA, 1987) since some of the species included have shifted between
L ~ I L ~ ~ S C L I S , C h o n d r ~ ~ t o m a , Pararutilus, Rutilus and Tropi-
dophoxinellus genera. At present it is considered to be formed by two species, R. arc,a.sii and R. lemmigii.
The first has two subspecies, the parental. distributed throughout the northern river basins, and R. a. macrolepi-
dotzis, found in North West Portugal, between the Limia and the Tajo, and in the Duero basin. Both are endemic to the Peninsula. (LOBON-CERVIA et al., 1989) include the subs-
pecies R. a. mucrolepidotus as a species in the text and in Table 1 of a paper, but do not include it in Table 2, referring
to its relict distribution in the most westerly streanis. Here we must refer to another endemism, Tropidophoxi-
nellus alburnoides, which was included in the same genera as
R . alh~~rnoides. Due to its peculiar biological characteristics, the presence of two different genotypical groups (2n = 50; 3n = 75) and triploid associated gynogenesis and the production of cloliic females, the "Rutilus alhurrzoides Complex" (COLLARES-PEREIRA, 1984, 1985) was created.

Table 2. Marine dependent species found in the estuaries, wetlands, marshes, salt flats, and brackish or salty lagoons of the Iberian Peninsula.
Familia Ammody tidae Familia Pomatomidae Ammodytes tobianus Pomatomus saltator Gymmnamodytes cicerellus
Familia Sciaenidae
Familia Batrachoididae Argirosomus regius Halobatrachus didactylus Umbrina canariensis
Umhrina cin-osa
Familia Blennidae Blennius pavo Familia Soleidae
Dicologoglosa cuneta Familia Bramidae Solea senegalensis Brama brama Solea vulgaris
Familia Clupeidae Sardina pilchardus
Familia Engraulidae Engraulis eucrasicholus
Familia Gobiidae Aphia minuta Gobius niger Gohius paganellus Gobius cruentatus
Familia Mugilidae Oeadalechilus labeo
Familia Mullidae Mullus barbatus Mullus surmuletus
Familia Sparidae Diplodus sargus Litognathus mormyrus Sparus nurata
Familia Stromateidae Stromateus fa'a fola
Familia Syngnathidae Hippucampus hippocampus
Familia Trachinidae Echiichthys vipera
It was finally decided (ELVIRA, 1987) to include it in the Tiopidophoxinellus genus, fully acknowledging the need for athorough review. Furthermore, LOBON-CERVIA et al., (1989) consider it as belonging to the Rutilus genus, and LOBON-CERVIA & ELVIRA (1989) as Tropidophoxine- Ilus.
In addition, the Iberian endemism Anaecypris hispanica has a complex taxonomy, and has been included in the Phoxinellus and Pseudophoxinellus genera. Its distribution, limited to the Guadiana basin, has been enlarged since its capture in the River Bembezar (Azuaga, Bajadoz), an affluent of the River Guadalquivir (DOADRIO, 1986).
As for the Leuciscus genus, similar problems have arisen, although traditionally (LOZANO, 1935; BUEN, 1935) it has been considered as comprising the species Leuciscus cepha- lus with two subspecies: L. c. cabeda in the north and L.c. pyranaicus in the south. However, DOADRIO (1987) has revised the genus, concluding that there are three species: L. cephalus, L. carolitertis and L. pyrenaicus. For the time being, and until the distribution of L. carolitertis is better known, it is restricted to the Duero basin. This suggests that this is a case of speciation similar to that of Rutilus, possibly due to the effect of glaciations.
Our own opinion, however, is in line with that of ALMACA (1976) and ELVIRA (1986), in that it is neces- sary to review thoroughly the Rutilus and Leuciscus genera, and determine the distribution of each species.
The group of strictly freshwater species is formed by the Cyprinidae, Cobitidae and Homalopterdae, and two vica- riant species: Blennius fluviatilis and Cottus gobio, both with a very concise distribution (table 3). Cyprinidae, Cobi- tidae and Homalopteridae have the largest number of ende- mic or native species. Of a total of 26 species (40.62% of fish fauna), 15 are endemic (23.42%), 5 are native (7.8%) and 6 (9.4%) introduced.
ALLOCHTHONOUS SPECIES
The introduction of allochthonous species in the Iberian peninsula began with the Romans, who acclimatised the carp, Cyprinus carpio, which today, with 19 species, makes up 29.7% of the total Iberian continental fish fauna. Their distribution among the river basins is unequal, with the lowest number of introduced species being found in the South of Spain (4) and the highest in the rivers of the Eastern Pyrenees, where 12 species in al1 have been found. These two areas include the five large river basins: the Ebro, Guadalquivir, Guadiana and Tajo, with 10 introduced species, and the Duero, with 8 (table 3).
The origins of these species, and the reasons for their introduction, are several. Firstly, there are "accidental" intro- ductions, mainly escapes from fish farms, where these fish were raised for sale or as live food for other species (brown trout, rainbow trout, grudgeon). These accidental introduc- tions also include Scardinius erythophtalmus, lctalurus rnelas and Silurus glanis. The government has caused other introductions: Esox lucius, Microterus salmoides, Lepomis gibbosus etc., for angling, and Gambussia affinis for health reasons (malaria control). Esox lucius, on the other hand, is
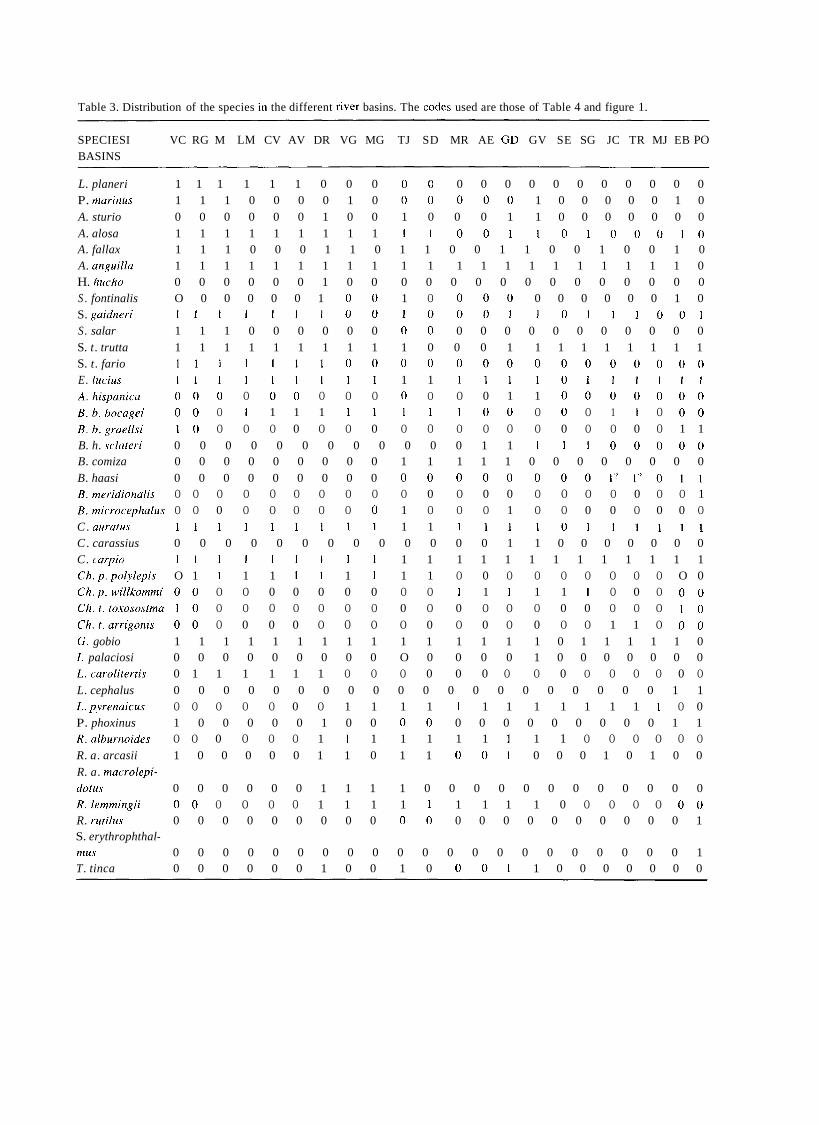
Table 3. Distribution of the species in the different river basins. The codes used are those of Table 4 and figure 1.
SPECIESI VC RG M LM CV AV DR VG MG TJ SD MR AE GD GV SE SG JC TR MJ EB PO BASINS
L. planeri 1 1 1 1 1 1 0 0 0 O 0 0 0 0 0 0 0 0 0 0 0 0 P. marinus 1 1 1 0 0 0 0 1 0 O 0 O 0 0 1 0 0 0 0 0 1 0 A. sturio 0 0 0 0 0 0 1 0 0 1 0 0 0 1 1 0 0 0 0 0 0 0 A. alosa 1 1 1 1 1 1 1 1 1 1 1 0 0 1 I O 1 0 0 0 1 0 A. fallax 1 1 1 0 0 0 1 1 0 1 1 0 0 1 1 0 0 1 0 0 1 0 A. anguilla 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 H. hucho 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 S . fontinalis O 0 0 0 0 0 1 O 0 1 O O 0 0 0 0 0 0 0 0 1 0 S. gaidneri 1 1 I I 1 1 1 0 0 1 0 0 0 1 1 0 1 1 1 0 0 1 S. salar 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 S. t . trutta 1 1 1 1 1 1 1 1 1 1 0 0 0 1 1 1 1 1 1 1 1 1 S. t . fario I 1 1 1 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 E. lucius 1 1 1 1 1 1 1 I 1 1 1 1 1 1 1 0 1 1 I 1 1 1 A.hispanica 0 0 0 O O 0 O O O 0 O O 0 1 1 0 0 0 0 0 0 0 B.h.bocagei O 0 O 1 1 1 1 1 1 1 1 1 0 0 O 0 O 1 1 O O 0 B.h.graellsi 1 0 O O O 0 O O 0 O O O O O O 0 O O 0 O 1 1 B. h. sclateri 0 0 0 0 0 0 0 0 0 0 0 0 1 1 I 1 1 0 0 0 0 0 B. comiza 0 0 0 0 0 0 0 0 0 1 1 1 1 1 0 0 0 0 0 0 0 0 B. haasi 0 0 0 0 0 0 0 0 0 0 0 O 0 0 0 0 0 1 ? 1 ~ 0 1 1 B.meridionalis O O O O O O O O O O 0 O 0 0 O O O O 0 O O 1 B.microcephalus O O 0 O O O O O O 1 O O O 1 O O 0 0 O 0 0 O C. auratus 1 1 I 1 1 1 1 1 1 1 1 I 1 1 1 0 1 1 1 1 1 1 C. carassius 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 C. calpio 1 1 1 I 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 Ch.p.polylepis O 1 1 1 1 1 1 1 1 1 1 O 0 O O O O O O O O 0 C h . p . w i l l k o m m i O 0 O O 0 0 0 0 0 O O 1 1 1 1 1 I O 0 O 0 0 C h . t . t o , ~ o s o s t m a I O O O O O O O O 0 O 0 O O O 0 0 O O O 1 0 Ch.t.arrigonis O 0 O 0 0 0 O O O O 0 O 0 O 0 O O 1 1 O O 0 C. gobio 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 0 1. palaciosi 0 0 0 0 0 0 0 0 0 O 0 0 0 0 1 0 0 0 0 0 0 0 L.carolitertis 0 1 1 1 1 1 1 O O O 0 0 0 O O 0 O O O O 0 O L. cephalus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 L.pyrenaicus O O O O O 0 O 1 1 1 1 1 1 1 1 1 1 1 1 1 O 0 P. phoxinus 1 0 0 0 0 0 1 0 0 O 0 0 0 0 0 0 0 0 0 0 1 1 R.alburnoides O O 0 O O O 1 1 1 1 1 1 1 1 1 1 O O O O O O R. a. arcasii 1 0 0 0 0 0 1 1 0 1 1 O 0 1 0 0 0 1 0 1 0 0 R. a. macrolepi- dotus 0 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 R.lemrningii 0 0 O O O O 1 1 1 1 1 1 1 1 1 0 O O O O O 0 R. rutilus 0 0 0 0 0 0 0 0 0 O 0 0 0 0 0 0 0 0 0 0 0 1 S. erythrophthal- mus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 T. tinca 0 0 0 0 0 0 1 0 0 1 0 O 0 1 1 0 0 0 0 0 0 0

Table 3. Distribution of the species in the different river basins. The codes used are those of Table 4 and figure 1.
SPECIES/ VC RG M LM CV AV DR VG MG TJ SD MR AE GD GV SE SG JC TR MJ EB PO
BASINS
C. calderoni
C . muroccana
N. harhutulus
S. glanis
I. rnelus A. iherus F. heteroc,litus V . hispanica
G. affinis A. hoyeri G. aculeatus
S . ahaster
C . gobio P. flu~~iutilis
S . Iiicioperca D. 1ahru.i.
D . punctata L. gibbosus
M. salmoides
Ch. Iahrosus L. aurata L. ramada
L. saliens M. cephalus
P . fl~rviatilis P . miel-ops
P. minutus
P . fle-vus
C . facetum
more of are introduction, as it has been found in the fossil
layer of the early Pleistocene in Arganda, Madrid (MORA-
LES, 1980). When more data is available, it may well be considered a native species of the Iberian Peninsula.
At present many changes are occurring, as species are being moved between different river basins. Live bait has
been used to catch predator species, which are then released in other river basins, as has been the case with Lepomis gibhosus, introduced into the Guadalquivir in the provirice of Cordoba from the Guadiana. Indiscriminate release by aquarium owners is another cause, and may explain the presence of Fundulus heteroclitus, Cichlasoma fac.etum. Carassilis auratus and Curassius carassius.
The Government, to satisfy ever increasing demand from anglers, has made massive introductions of fish from
Germany, Italy. France and the USA. The fish involved are brown trout, rainbow trout, pike, largemouth bass and pumkinseed sunfish.
Some species such as Hucho hucho, Sali,elinus fontina-
lis. Ictalurus melus and S ~ I U ~ L I S glanis show a limited distri- bution despite having been introduced in several areas. On the other hand, species such as Esox 1uciu.r. Micropterus salmoides, Lepornis gihbosits and Gamhusia aflinis are very widely distributed. F~lndulus heteroclitus and Cichlasoma ,facetum have been found in the river basins of the South of Portugal, the Guadiana and the Guadalquivir, but their

C Y P R I N I O A E , C O B I T I O A E 8 HOMALOPTERIDAE -0.40 o.,w a.0 a .m 1.20
distribution area is spreading. As for Cichlu.sowiu ,fuc.etum,
the first known samples are from the River Mira (HELLING, 1943). spreading later towards the South of
Portugal (ALMACA, 1964; COLLARES-PEREIRA, 1985), ~ i n d subsequently to the Guadiana basin (PEIRO, 1987).
SECTORISATION OF THE IBERIAN PENINSULA
There have been several attempts to divide the Iberian Periirisula into sectors of freshwater fauna. AREVALO
( 1929) proposed a model based on the presence or absence
of salmon and ciprinodontiforms, with three "provinces": Cantabria, the Atlantic and the Betico-Mediterranean. LOZANO (1952), based his model on the presence or
1 absence of Cyprinidae, Cobitidae and Cyprinodantidae, and
Figure , , Secrorlzation os the Peninsula, The ini t ia ls os feduced the number of "regions" to two: Northern and Sout-
river basins are those oí' Table 4. hern. ALMACA (1978) and HERNANDO et al., (1982)
Table 4. Number of species catalogued in the Iberian Peninsula river basins, using those whose surface is equal to or greater than
990 square kilometres. (NAT: native; END: endemic: INT: introduced).
RIVER BASINS SURFACE AREA (Km2)
-
SPECIES NAT END INT TOTAL
CANTABRIAN BASINS RIVERS OF GALICIA MIÑO
LIMI A
CAVADO AVE DUERO VOUG A
MONDIGO TAJO
SADO MIRA
ALGARVE GUADIANA GUADALQUIVIR BASINS O F SOUTHERN SPAIN
SEGURA JUCAR TURIA MIJARES EBRO WESTERN PYRENEES

propose two different models, both based on the distribution of Cyprinidae and Cobitidae. In the first, there are three "subdistricts" or "subsectors": Ebro-Cantabria, Central and Southern, and in the second there are two subregions: the North Atlantic and the Betico-Meditteranean.
Although HERNANDO (1990) considers this latter sub- regionalization to be valid, we believe that the new data on Cyprinidae, Cobitidae and Homalopteridae (strictly freshwater species), the description of a new species of the Lezisicus genus, the variation in the distribution of Anaecypris hispanica, etc., justify a new attempt to clarify the sectorisation or subregionalisation of the Iberian Peninsula.
This will be based on the distribution matrix (presence- absence) of the species and subspecies of these families in the 22 river basins with a surface area of more than 990 km2
(table 2). The river basins are grouped using the PHI simi- larity index (ROHLF, 1988) and then a UPGM-type cluster analysis (ROHLF, op. ct.), considering species and subspe- cies as identical.
Three different models were made to determine which most closely represents the distribution of species in river basins. The first includes species and subspecies of Cypri- nidae, Cobitidae and Homalopteridae (fig. 1). The second adds Blenniusjlu~~iatilis, Cottus gobio and Lurnpetra planeri to these families, and the third includes al1 35 species and subspecies under consideration (table 3).
The results show three groups of river basins (fig 1). The first group includes the Ebro, Cantabria and the eastern Pyrenees. The second has two subgroups: the Gallegos, Miño, Limia, Cavado and Ave, and the Duero, Vouga, Mondego, Tajo and Sado. The third group includes the South of Spain and the Segura, and the Jucar, Turia and Mijares on the Mediterranean.
This pattern is repeated for the other models, and demonstrates the strong influence of the Cyprinidae, Cobi- tidae and Homalopteridae families with respect to the other species. It is thus proposed to consider the Iberian Penin- sula as being divided into three subregions or subsectors: the Ebro-Cantabrian, the Atlantic and the Betico-Medi- terranean.
REFERENCES
ALMACA, C., 1964. Contribucao para o conhecimiento da fauna ictiologica das aguas interiores portuguesas. Bol. Soc. Port. Cien. Natur. 2% Serie, Vol X: 228-237.
ALMACA, C., 1967. Estudo das populacoes portuguesas do Géri. Barbus Cuvier, 1817 (Pisces: Cyprinidae). Rev. Fac. Cien. Lisboa, 14 (2): 15 1-400.
ALMACA, C., 1972. Sur la systématique des Barbeaux (genre et sousgenre Barbus) de la Péninsule Iberique et de I'Afrique du Nord. Arq. Mus. Boc. 3, nQ 10: 319-346.
ALMACA, C., 1976. La speciation chez les cyprinidae de la Peninsule Iberique. Rev. Trav. Inst. Peches Mart. 40 (3-4): 399-41 1.
ALMACA, C., 1978. Repartition geographique des Cyprinidaes iberiques et secteurs ichthyogeographiques de la Péninsule Iberique. Ves. Cesk. Spole Zool. XLIl 4: 241- 248.
ALMACA, C., 1984. Form relationships among western paleartic species of Barhus (Cyprinidae, Pisces). Arq. Mus. Boc. Serie A, Vol 11, 12: 207-248.
AREVALO, C., 1929. La vida en las aguus dulces. Ed. Labor. Barcelona. 198 pp.
BANARESCU, P., 1972. The zoogeographical position of the East Asian fresh-water fish fauna. Rev. Roum. Biol.
Zool. 17 (5): 315-323. BANARESCU, P., 1973 a. Origin and affinities of the
freshwater fish fauna of Europe. Ichthyologia. 5: 1-8. BANARESCU, P., 1973 b. Some reconsiderations on the
zoogeography of the euro-mediterranean fresh-water fish fauna. Rev. Roum. Biol. Zool. 18 (4): 257-264.
BUEN, F. DE ., 1935. Fauna ictiológica. Catálogo de los peces ibéricos de la planicie continental, aguas dulces, pelá- gicos y de los abismos próximos. Inst. Esp. Ocean. Nat. y Res. Sr. 11 88.
COLLARES-PEREIRA, M. J., 1983. Estudio ~itematico e citogenetico dos pequenos cyprinideos ibericos pertene- cientes aos generos Chondrostoma Agassiz, 1835, Rutilus Rafinesque, 1820 e Anaecjpris. Ph. D. Thesis Mus. Boc., University of Lisboa, 51 1 pp.
COLLARES-PEREIRA, M. J., 1984. The "Rutilus albur- noides (Steindachner, 1866) complex" (Pisces: Cyprinidae). 1.- Biometrical analisys of zone portuguese populations. Arq. MLLS. Boc. 11 (8) 2: 11-143.
COLLARES-PEREIRA, M. J., 1985 a. The "Rutilus alburnoides (Steindachner, 1866) complex" (Pisces: Cypri- nidae). 11.- First data on the caryology of a well established diploid-triploid group. Arq. Mus. Boc. 111 (S): 69-90.
COLLARES-PEREIRA, M. J., 1985 b. Ciprinideos do Alentejo. De (FROM) Congresso sohre o Alentejo, Evora. Vol 11: 537-545.
DEMESTRE, M., A. ROIG, A. DE SOSTOA & F. M. DE SOSTOA., 1977. Contriboció a l'estudi de la ictiofauna

continental del delta de I'Ebre. Trah. Insr. Cat. Hist. Nat. 8: 145- 226.
DOADRIO, l., 1984. Relaciones filogenéticas y hiogeo-
gr.áficas de los ha~.hos (Barhus, Cyprinidae) de la Península
Ibérica y aportes c~orológicos y hiogeográficos a su icrio- fauna continental. Ph. D. Thesis. Complutense University of Madrid. 1 11 9 pp.
DOADRIO, I., 1986. Ictiofauna de la Comunidad Autó-
noma de Extremadura. Informe preliminar. Direccicín Gene- ral de Medio Ambiente. Junta de Extremadura.
DOADRIO, I., 1987. Leuciscus curolitertis n. sp. from the Iberian Peninsula (Pisces: Cyprinidae). Senken. Biol.,
68: 4-6. ELVIRA, B., 1986. Re~~isicín taxonón~ica y distrihucirín
geográfica del género Clzoizdrostonza Agassiz, 1835 (Pisc.es: Cyprinidae). Ph. D. Thesis. INIA. Ministry of Agricultura. Madrid. 62: 1- 530.
ELVIRA, B., 1987. Taxonomic revisión of the genus Chondrostoma Agassiz, 1835 (Pisces: Cyprinidae). Cyhium 11 (2): 11-140.
FERNANDEZ-DELGADO, C., J. A. HERNANDO, M. HERRERA & M. BELLIDO, 1986. Sobre el status taxonó- mico del género Vulencia (Myers, 1928) en el suroeste de Iberia. Doñana Acta Vertebratu 13: 161 -163.
GARCIA DE JALON, D & M. GONZALEZ DEL TANAGO, 1983. Estudio biotopológico de las comunidades piscícolas de de la cuenca del Duero. Bol. Est. Cent. Ecol. 12: 57-66.
GARCIA DE JALON, D & J. V. LOPEZ ALVAREZ, 1983. Distribución geográfica y mesológica de las especies piscícolas de la cuenca del Duero en el verano de 1981. Actas I C o n g r . Esp. Limnol. 227-235.
HELLING, H., 1943. Novo catálogo dos peixes de Portu- gal em colecqao no Museu de Zoologia de Universidade de Coimbra. Men. Est. Mus. 2001. Univ. Coimbra, 149. 110 pp.
HERNANDO, J. A., 1975 a. Notas sobre la distribución de los peces fluviales en el suroeste de España. Doñuna Acta
Vertehrata 2 (2): 263-264. HERNANDO, J. A,, 1975 b. Nuevas localidades de
Valencia hispanica (Pisces: Cyprinodontidae) en el suroeste de España. Doñanu Acta Vertehrata. 13: 265 267.
HERNANDO, J. A., 1990. Ictiofauna del río Guadalqui- vir: Características y perspectivas. In: Avances en el cono-
253
cimiento y gestion del medio ambiente de Córdoba. A. López & J. M. Recio (Eds) . 35-62.
HERNANDO, J. A. 1. DOADRIO & C. FERNANDEZ- DELGADO, 1982. Distribución geográfica y diversidad de la ictiofauna continental ibérica. Modelo inicial. Res. 11 .lar.
Icriol. Iherica. Barcelona. Mayo 1983. ICONA, 1986. Lista roja de los Vertehrados de España.
Publ. Minist. Agric., Pesca y Aliment. Madrid. 400 pp. LOBON- CERVIA, J., B. ELVIRA & P. RINCON, 1989.
Historical changes in the fish fauna of the river Duero basin. In Historicul change of la,-ge alluvial rivers: Western Europe. (G.E. Pett, H. Moller & A. L. Roux, eds.): 221 -
232. John Wiley & Sons. Chichester. 355 pp. LOBON-CERVIA, J & B. ELVIRA, 1989. Estado de conser-
vación de los peces fluviales ibéricos. Queicus 44: 24-27. LOZANO REY, L., 1935. Los pec,es,fluviales de España.
Mem. Real Acad. Cien. Ex. Fis. Nat. Tomo V. Madrid. LOZANO REY, L., 1952. Los peces,fl~4iliales de España.
Min. Agricultura. Madrid. MORALES, A,, 1980. Los peces fósiles del yacimiento
Achelense de Aridos- 1 (Arganda, Madrid). Ocupaciones
Achelenses en el valle del Jarama. Publ. Exma. Diputación Prov. Madrid. 93-1 04.
MYERS, G. S., 1960. The endemic fish fauna of the lake Lanao, and the evolution of higher taxonomic categories. Ei~olution, 14 (3): 323-333.
PEIRO, S., 1987. Una excursión por la cuenca del Guadiana. Aquamar, 25: 32-33.
ROHLF, F. J., 1988. NTSYS-pc: Numerical tasonomic
and Multivnr.iarite Anulysis System. Exeter Publishing Ltd. New York. 129 pp.
SOSTOA, A., F. J. SOSTOA, J. LOBON- CERVIA, B. ELVIRA, J. A. HERNANDO & M. AVILA., 1984. Atlas y distribución de los peces de agua dulce de España: el proyecto, métodos y resultados preliminares. Bol. Est. Cent. Ecol. 13 (25): 75-81.
SOSTOA, A. & J. LOBON-CERVIA., 1989. Fish and fisheries of the river Ebro: actual state and recent history. In: Historical change of large alluvial rivers: Western E ~ ~ r o p e . (G.E. Pett, H. Moller & A. L. Roux, eds.): 233 - 247. John Wiley & Sons. Chichester. 355 pp.
TERMIER, H. et G.. 1960. Atlas de Paléogéogruphie. Masson. Paris.

FISH SPECIES ECOLOGY IN SPANISH FRESHWATER ECOSYSTEMS
C. Granado-Lorencio Departamento de Biología Vegetal y Ecología. Facultad de Biología, Apdo. 1095, 41080 Sevilla. Spain.
Keywords: Fish assemblage, Life-history, Fluctuating stream, Spain.
ABSTRACT
The life strategies of the Iberian ichthyofauna, poor in species and with a high number of endemisms, are the result of the
functioning of the aquatic systems (streams and reservoirs). The majorety of the species wager for iteroparity, high fertility, predominance of males over females and omnivorous feeding habits (short- chain trophic relationships). The fish assemblage
is not very structured and depends on the autoecology of the species.
A HISTORICAL APPROACH
Two factors determine, or have determined, life strate- gies and structure of the fish assemblages in Spanish fresh-
water ecosystems: biogeography of the species and func- tioning of the aquatic systems. Relatively recently (from the
beginning of the twentieth century), reservoir construction policy has resulted in adaptive divergences from the gene- ral patterns.
The Iberian Peninsula is in the extreme south of the conti-
nent, as a bridge towards Africa. The Pyrenean mountain system and the Straits of Gibraltar give this enclave singular biogeographical characteristics. In contrast to the diversity of the Palaearctic region and the European continent, the
Iberian fish fauna comprises just two freshwater families (Cyprinidae and Cohitidae), and ten with a certain tolerante to saltwater, of marine origin or with a cycle shared between river and sea (Cyprinodontidae, Acipenseridae, Anguillidae, Clupeidae, Salmonidae, Sygnatidae, Gasterosteidae, Bleni- dae, Gobiidae and Cottidae). Of the 61 Iberian species (44 native and 17 exotic), the Cyprinidae are the majority (22 species). The isolation of the area has determined the post-
oligocenic differentiation of the present-day Iberian ende-
misms (16 taxa). The exotic species belong to the families Esocidae, Ictaluridae, Poecilidae and Centrarchidae.
At the same time, the region is poor in lentic ecosystems, and the river systems are not very large compared with others in the world. Most of the Iberian river systems are characterised by their intermittence, with periods during the
year of torrential rain and drought, typical of semiarid and
arid geographical areas. Moreover. the seasonal irregularity
of the rainfall causes marked differences in the interannual flow. These hydrological peculiarities have determined the water policy of the present century, with the construction of
more than 1200 large dams (48 1 between 1938 and 1973).
Something over 45,000 ~ m ~ / y e a r of the flow is controlled (40 % of the total flow). To sum up, it can be said that the Spanish aquatic environment comprises few lakes and lago-
ons, with a group of sh ort hydrographic networks of irre-
gular, seasonal flow, controlled along practically their whole length by reservoirs.
In this "hydrological schizophrenia", the Iberian ichthyo- fauna - poor in species, with a high number of endemisms
(as a result of isolation), and evolving in the functional framework of natural environments (rivers), some of which resemble those of the past while others are totally discon- nected from current reality (by reservoirs and the reaches
below them) - uses its life strategies to survive. Paleoautoe-
cology and "adaptive reconversion" is the evolutionary wager for the Iberian ichthyofauna.
RIVER ICHTHYOFAUNA
The Iberian river ichthyofauna comprises three species types: sedentary and potamodromous natives, sedentary exotics, and migratory species (anadromous and catadro- mous).
Limnetica, 8: 255-261 (1992) O Asociación Española de Limnología, Madrid. Spain

The recent history of the Spanish migratory species is a phenomenon with scarce data. Their abundant populations in historical times were due to the location of the Iberian Peninsula in the migratory path of temperate diadromous fish species to the north Atlantic Ocean and the Mediterra- nean. The most important migratory fish species of the
Iberian Peninsula were sea lamprey (Petr-omyzon mar-inus ), Atlantic salmon (Sulrno salar-), sturgeon (Aripensel- sf~l-io),
European eel (Anguilla unguilla), shads (Alosa spp) and mullet (Chelon spp, Liza spp, Mugil spp). Since the early twentieth century, the blocking of upstream migration by reservoir construction has led to a progressive decline in the populations of the Iberian migratory species. Some of them are very scarce os have disappeared (sea lamprey, sturgeon. Atlantic salmon and shads); European eel and mullets are still abundant in the lower reaches.
As stated above, the family Cypr-inidae is the most inte-
resting group of the Spanish ichthyofauna (for its number of species and endemisms, fish assemblages, feeding habits, etc.). The Iberian Cyprinidae are characterised by short age pyramids (few age classes); short lifespan; early sexual maturity (between the second and fourth year of life); high
levels of fertility, in one os more reproductive periods during the year (3000 to 8000 ovocytes per mature female) varying according to age; the sex ratio usually favours the ma les (with interannual variations); the reproductive season is associated to the spring and early summer; high levels of mortality both at population recruitment level and throug- hout the age classes; cyclic growth rhythm, synchronised with the most favourable period and of 3 to 5 months in
duration (slowing down os becoming zero in summer arid winter).
There are rheophile species (mainly Iberian barbel and nase) which migrate upstream to reproduce, in search of
zones with current, grave1 and sand bottom, and well oxyge-
nated (RODRIGUEZ, 1992). The beginning of migration appears to be related to water temperature (between 12 and 14 O C). It does not usually last more t h a ~ one month (deter-
mined by the migratory distance). Occasionally it takes place globally (continuous flow to the reproduction aseas), and in some species (barbel) in a series of spurts. In both
cases, retur n to the normal aseas of occupation is en masse. These reproductive rnovements, in the Iberian rivers, should
not be considered as di splacement of the whole population to a single zone of the river (as with the salmon), but more a displacement of population groups localised in different sectors of the river that move upstream and reproduce in different zones ("meet und spawn hypothesis").
Feeding is at four trophic levels: detritus, phytoberithos,
benthonic macroinvertebrates, and emergent forms. Exploi- tation is sometimes at a single level (Chondr-ostoma-detri- tus) and sometimes at several levels (Bar-hz1.s-niacroinverte- brates, phytobenthos, drift). There can even be an ontogenic variation of food within the same species (Leuciscus-fry (algae and zooplankton), adults (microcarni vorous)). Ichth- yophagy is rase in Iberian fish, although fish remains can be
found sporadically in analysed stomachs.
Thus the taxa in the Iberian river communities show a low trophic diversity, due to their similar adaptations to an
environment whose fluctuation prevents more sophisticated evolutionary wagers. This incurs the need to develop time- separated patterns of resource exploitation (segregation),
when the feeding resource, if unequal, is spatially in the same microhabitat. Generally (DIANA, 1979), it may be said that with more ichthyophagy, the daily feeding rhythm is more acyclic. Herbivorous, benthophagic, and other
behaviours produce cyclic rhythms, which can be diurna1
or nocturnal, depending on the type of prey, competitive phenomena, etc. The latter feed continually os during a long
period o i the day and digest while they eat (they obtain
little energy from the food, forcing them to eat a lot - high energy cost in feeding). The others eat rapidly (over a short period) os severa1 times a day, and digest the food before eating again (they obtain high energy from the food and do
not need to eat frequently - they spend little energy in feeding). The detritovores obtain little energy from the food, forcing them to eat continually, while the herbivores obtain somewhat greater energy yield from the food, so that
they do not need to be continually feeding. The results obtained by ENCINA & GRANADO LORENCIO (1991) in a study of diet and diel feeding chronology in three ende- mic species demonstrate this phenomenon. Iberian nase
(detritovores) have a mean energy from a full stomach of
1450 callg dry, ash-free weight, Iberian barbel 3600 cal/g, and chub 5000 cal/g.
The conflict comes when the feeding habitat coincides, even though the food does not. On these occasions, the
species temporarily displace their daily feeding cycle so as not to interfere with each other. The latter authors found that the nase has maximum feeding between solar hours 19 and 20, while for the barbel it is between 13 and 15, in a sector
of the river in which the benthonic habitat is not very diverse and the zones of sediment accumulation are scarce. When there is great diversity of trophic habitats for the species, there is no need for displacemeiit - the barbel main-
tains the same timetable and the nase takes the hours 14 to

16, much closer to the other species (which may be its
potential feeding rhythm in non-competitive conditions). Fish inhabit a patchy environment (TOWNSEND, 1989),
reconnoitring the habitat to exploit the available resources necessary Sor their growth, base metabolism, and reproduc- tion. They move in function of the spatial concentration of
the trophic resources. If the resource is abundant and uniformly distributed, there is no need Sor a large home range. In contrast, if the resource is scarce or very concen-
trated, they need to range over a very large area to obtain the energy necessary (WIENS. 1976). Moreover, as the Iberian river s fluctuate throughout the year, sometimes we
find large home ranges and other times small ones (neces- sarily so at low water). Better quality habitats produce more
restricted living domains (MATTHEWS, 1990). How do the Iberian Cyprinidae behave?. The response is a littie
speculative: there are not enough data. If we consider the results obtained by PRENDA & GRANADO LORENCIO
(in press), using phytobenthonic production as potential indicator of the available resources and hydrology of the different reaches of the stream, we find some interesting behaviour. In the high reach of the stream during winter, when there is still a great amount of food in the riffles and raceways, these are the least occupied (mechanical incapa- city). The pools, although with less food, are more visited
(there is a negative gradient of current speed nearer the
bottom of the pool). That is, the pool is a shelter against the periodic winter disturbance. However, when the river is wider and less deep, with a greater capacity to reduce the disturbing effect of spate (mi dreach), the ichthyofauna is
spatially structured in a different way. Below a certain speed, the fish is able to keep swimming and overcome the drag effect, occupying the zones of the river with greater resources. If we could see what happens in other parts of th
e year, with less flow, the tendency would probably be towards the riffles.
If a greater home range is needed with greater concentra-
tion of resources, the response of the Iberian Cyprinidae in
the results obtained by the latter authors does not disagree with the conclusion of WIENS (op. cit.): species have a smaller home range when the re sources are uniformly
distributed. Estimates of biomass and production can b l used to quan-
tify "degree of welfare" of the popuIations. The existing data for Iberian rivers are few and to some degree incom- plete (no data for temperature regime, nos primary and secondary production, os any other measu rement permitting integration of fish production value into the aquatic ecosys-
tem). The estimates per species vary between 54 and 59 O Kg/Ha in biomass and between 21 and 353 Kg/Ha/year in
production (values of P/B between 0.49 and 0.70) - higher values than those obtained in most European rivers. There are distinct causes: predominance of Cyprinidae in the
assemblages; scarce, low diversity; and high thermal regime in the rivers. Nevertheless, partial analysis of these results
may lead to erroneous interpretations. Values of this type may result from the confluence of positive environmental
factors in a particular year, due to the unpredictability of the
mediterranean environments and the autoecology of the species. However, lengthy following of biomass and produc- tion would undoubt edly show the strong interannual fluc-
tuations which are typical of this type of ecosystem subjec- ted to physical disturbances (STRONG , 1983).
RESERVOIR ICHTHYOFAUNA
Three life-cycle-types of species can be found in Spanish reservoirs. There are those that live their whole cycle in the
reservoir (common carp, goldfish, tench, pumpkinseed, black bullhead, largemouth bass, mosquitofish, and pike); those that migrate to the high reaches of the rivers to repro- duce (barbel and nase); and those that have remained trap-
ped in the reservoir in their trophic or reproductive migra-
tion from the sea (mainly sandsmelt and eel). In coldwater
reservoirs, it is possible to find certain native or introduced salmonids (Salmo tvutta, Onchorrynchus mykiss and Salve- linus j'ontinalis).
There have been few studies at ichthyofauna leve1 in Spanish reservoirs. Nevertheless. the results obtained throw light on the effec t of the reservoir on the fish popu- lations. It has been possible to demonstrate the interaction
between limnology and plasticity of the species in a
comparative study of populations of Iberian nase (Chon- drosrornu polylepis) from three in-chain reservoirs of the Tagus River : Valdecañas, Torrejon and Arrocampo. The
factors having greatest effect on growth are primary productivity of the water mass (expressed as units of chlo rophyll "a") and mean depth of the reservoir (GRANADO LORENCIO et al . , 1985).
The spatial, temporal, and bathymetric distribution of the populations in the reservoir depend on endogenic-type factors (reproductive cycle) - genetically fixed - and exogenic ones (environmental) - determined by the physico-chemical variables. The river origin o f certain species (mainly barbel and nase), which move upstream for

25 8
reproduction, determines the maintenance of this type of cycle once they occupy the water mass of the reservoir. For a relatively long period of the year (generally from February to May or June, depending on the species), the
populations go up the tributaries in search of zones of well oxygenated, clear water with sand or grave1 bottom, where they spawn. In this way, the species assure their maximum
reproductive viability, and temporarily exploit the produc-
tion of the river, functioning as "a mature ecosystem which exploits other less mature ones" (c t : MARGALEF, 1960). This process results in swings of population to the rivers at spawning, on one hand, and recruitment of young indi-
v i d u a l ~ to the reservoir population, on the other. Other
species, belonging to the genus C.vprinils, Carassius. Micropterus, Esos, Lepomis, etc., reproduce in shallow
zones of the reservoir with vegetation, migrating within the reservoir from the areas of normal localisation to those of
reproduction. The exogenic factor is determined by the annual evolu-
tion of certain limnological parameters of the water mass. The reservoirs in these latitudes have annual holomictic-
monomictic cycles. Stratifications are established during the
summer period and a potent hypolimnion develops which increases with depth and eutrophycation level of the reser- voir. The cyclic behaviour of the phenomenon precludes a homogenous distribution of the ichthyofauna, which is loca- lised in surface layers and shallow creeks.
The period of stratification does not affect only the gene- ral pattern of population distribution, varying the home
range of the species. It also has a very important effect on the feeding regime of those species having a benthonic
andlor detritophagic diet (the niajority of the natives), since during this period the potentially exploitable resource is reduced to those areas where the hypolimnion does not
develop. If the food of the species in the river is based on detritus,
periphyton, and drifting and benthonic macroinvertebrates, associated with the large number of habitats in the river
courses, colonisation of the reservoir alters this feeding regime. The trophic chain of the reservoirs is not very diverse (unless there are exotic species) due to the drastic reduction of habitats and scarce structuring of a truly bent-
hic littoral, as a result of the continuous variations in level. Because of this, and the anatomic-structural incapacity of the species to exploit the new resources (mainly phyto- plankton and zooplankton), the reservoir leaves few possi- bilities o f a wide diet for the species (GRANADO LOREN- CIO & GARCIA NOVO, 1986).
The production of phytoplankton and zooplankton is not efficiently exploited by the fish, and sediments on the bottom (a process tha t may increase in cases of high eutrophy). A great part of the available energy in the system
is channelled detritically and benthon ically. The only trop- hic strategy of the native species is the exploitation of both subsystems. This implies diets based on detritus, insect larvae (mainly chironomids, being one of the few groups
represented), and bankside phytobenthos. However, it is not unusual to find planktonic elements in the alimentary tracts. The latter can be interpreted as "passive" feeding, forming
part of the diet without localisation and capture on the part of the fish. It enters the stomach only as a result of the abun-
dance of prey in the medium, being ingested with this other,
actively hunted prey. If the absence of species able to exploit the plankton and
ichthyophage levels (excepting the introduced species) is
added to the sc arce structuring of Spanish reservoir assemblages, the development of evaluative fishery models such as those applied since the 50s in other countries is
difficult (SCHNEIDER & HAEDRICH, 1989). In view of
this, a study was carried out in seven reservoirs in the
south of the Iberian Peninsula (GRANADO LORENCIO & SANCHO, 1987). The aim was to study the behaviour of different climatic. edaphic, soil-use, limnological, and fish
production variables, to detect which were most important, using multiple regressions. The two variables best related with reservoir biomass were average depth and the shore- line development factor.
There are works which demonstrate the relationship
between environmental variability and community structure. The assemblages do not depend only on the phenomena of competitive interaction. If the populations have no problems
in exploiting the resources, their swi ngs are caused by other
factors (EBELING et al., 1985). This is the case of the Torrejon reservoir (GRANADO LORENCIO, 1991). There was a large variation in population number throughout the
period 1980-1988. Spanish reservoirs have characteristics which make the system relatively unstable: swings of level, hypolimnetic water outflow, etc. The effect on the ichthyo- fauna depends on the autoecology of the species. Population stability is the result of interactions between the effect of
environmental changes (disturbances) and the adaptive capacity of the species. The disturbing factors can vary fish association in different ways, depending on the impact and effect on each taxa. The species of the lberian reservoirs are of an elastic and high reproductive rate type, and thus even- tually tend to return to the pre-disturbance state.

THE SPANISH FISH MODEL
The life strategies of the extant species can be understood only within the framework of biogeography and the func-
tioning of the ecosystems which they inhabit. From a pool of species which colonised the Iberian rivers in the past, and
from the evolution of their hydrological regime, it is possi- ble to understand the biological patterns developed by these species. Up to a point, some of them have developed
recently, and change (although minimally) in each annual cycle (occupation of space, variations in the qualitative composition of diet, changes in the population through catastrophic losses in recruitment). Others are possibly the
inheritance of a past much more adverse than the present, as
in the upstream migratory behaviour for reproduction (typi-
cal of colder eras). The ichthyofauna of most lberian rivers has life strategies
that show the best possible adaption to an irregularly fluc-
SPECIES
SELECTION
ASSEMBLAGE
DOMINANCE / NlCHE
tuating envir onment. It is possible to observe different forms of organisation of the fish communities or assemblages (fig.
1). When the time variation is small, the species can develop strategies to exploit resources and occupation of space which
occasionally lead to competitive relationships. In those rivers, the "community" may be controlled by the niche or
by phenomena of dominante by one species (YODZIS, 1986). These communities typically comprise native or intro- duced salmonids and other species with scarce populations.
In contrast. in stream with greater time variation (typical of the Iberian Peninsula), assemblage is not very structured
and depends on the autoecology of the species. To lessen the effect of disturbances, the majority of the species wager for iteroparity, high fertility, predorninance of one sex over the
other, and possibly even monosex populations (although
there are no reliable data on those processes). These are explosive population phenomena which counteract losses due to environmental factors, omnivorous feeding habits, and
COMPETlTlVE LOTTERY
TEMPORAL VARIATION - ENVIRONMENT L O W HlGH UNPREDlCTED
S T R E A M reservoir Figure 1. Schematic relationships betweeri teinporal variation and life sti-ategies and control of fish asseriiblageb in spanish aquatic ecoays- tems.

short-chain tropliic relationships, etc. Higher specialisation might be a cul-de-sac in a strongly íluctuating environment.
Assemblages respond more to an individual strategy of
survival than to the result of competitive interactions. Envi- ronmental fluctuations impede stable structuring of the "community", producing continua1 processes of reorganisa-
tion following the destructive phenomena caused by floods
and very low waters. Thus it is understandable to find asso- ciations in neighbouring rivers, formed by three species or more than eight, depending exclusively on the biogeograp- hical component. This type of association in the Iberian rivers is a good example of the "competitive lottery model" (WARNER & CHESSON, 1985).
From a synecological point of view, the relationships between functioning of the river and autoecology of the
species are obvious. In the rivers of the mediterranean region, there is a general destruction of biological associa- tions during the winter period. The high torrential flow removes everything that existed previously. As the environ-
mental conditions become more favourable in the spring, aquatic organisms begin to recolonise. The invasion invol- ves an acceleration in the life cycles, and thus high biologi- cal production that is transferred to the higher levels of the
trophic chains of the river. In this annual pattern, what is the best adaptive role for
the ichthyofauna?. The synchronisation of its life cycle to exploit resources as efficiently as possible and optimise the viability of its descendants. The species reduce the feeding
leve1 during winter, with zero growth (unfavourable period). In spring, trophic resources are used with more diverse feeding strategies; the beginning of the somatic growth period; and part of the energy consumed is transferred to the
production of reproductive tissue. Most species reproduce in this period or at the beginning of summer. Synchronisa- tion is perfect when it is seen that eclosion of the larval forms takes place in summer, the period when there is grea-
test abundance of small-sized prey (diatoms, bankside
plankton, and others) - the food base of the individuals of class O+ in most species.
The reservoirs are at the other extreme of time variation. Here, the importance of this factor is not so much its size as
its unpredictability. The seasonal disturbance of the stream
is internalised by the species (synchronisation), but that due to reservoir management is not. Consequently, only those species having a high degree of opportunist life cycle or
selection, with reproductive migrations out of the reservoir or with omnivorous feeding habits, will be capable of remai- ning in the reservoir.
Undoubtedly, the environmental severity of stream in the mediterranean region is a good "learning ground" for the Iberian species which colonise reservoirs. Those able to
make the evolutionary wager of all-or-nothing are the best prepared to survive in man-made environments which currently make up the greater part of the aquatic environ- ments of this region.
REFERENCES
DIANA, J.S. 1979. The feeding pattern & daily ration of a top carnivore, the northern pike (Esox lucius).Can. J. Zool.
57: 2121-2 127. EBELING, A.W.; LAUR, D.R. & ROWLEY, R.J. 1985.
Severe storm disturbances & reversal of community struc-
ture in a southern california Kelp forest. Marine Biology 84:
287-294. ENCINA, L. & GRANADO LORENCIO, C. 1991. Diet
& diel feeding chronology of three iberian fish species.
Ecology International Bulletin 19: 43-64. GRANADO LORENCIO, C.; GUILLÉN, E. &
CUADRADO, M. 1985. The influence of some environ-
mental factors on growth of iberian nase, Chondrostoma
polylepis, in three reservoirs of western Spain. Cybium 9 (3): 225-232.
GRANADO LORENCIO, C. & GARCIA NOVO, F. 1986. Feeding habits of the fish community in a eutrophic
reservoir in Spain. Ekologiu Polska 34 (1): 95-1 10. GRANADO LORENCIO, C. & SANCHO, F. 1987.
Producción piscícola en siete embalses españoles: Memoria de un Proyecto y resultados preliminares. Actas IV Congreso
Espatiol de Limnologi'a: 359-367. GRANADO LORENCIO, C. 1991. Fish communities of
spanish reservoir systems : a non-deterministic approach. I'crh. /r~t<~i.iic~t. I'<,r.ciii. Lin~riol 74: 2478-743 1 .
MATTHEWS, K.R. 1990. An experimental study of the habitat preferences & movement patterns of copper, qull- back & brown rockfishes (Sebastes spp.). Env. Biol. Fish.
29: 161-178. MARGALEF, R. 1960. Ideas for synthetic approach to
the ecology of running waters. Int. Rev. Gesamten. Hydro- biol. 45: 133.153.
RODRIGUEZ KUIZ, A. 1992. Comunidad icrica y estructura del háhitat en un vio de régimen mediterráneo. Doctoral Dissertation, Sevilla University, Spain, p.p. 430.
SCHNEIDER, D.C. & HAEDRICH, R.L. 1989. Predic- tion limits of allometric equations: a reanalysis of Ryder's

morphoedaphic index. Can. J . Fish. Aquat. Sci. 46: 503- 508.
STRONG, D.R. 1983. Natural variability & the manifold mechanisms of ecological communities. Amer. Natur. 122:
636-660. TOWNSEND, C.R. 1989. The patch dynamics concept of
stream community ecology. J . N . Am. Benrhol. Soc. 8 (1): 36-50.
WARNER, R.R. & CHESSON, P.L. 1985. Coexistence mediated by recruitment fluctuations: a field guide to the storage effect. Amer. Natur. 125: 769-787.
WIENS, J.A. 1976. Population responses to patchy envi- ronments. Ann. Rev. Ecol. Syst. 7: 81-120.
YODZIS, P. 1986. Competition, mortality & community structure, p.p. 480-491. Irz J.M. Diamond & T.J. Case (eds.). Community ecology. Harper & Row, New York.

EUTROPHICATION IN SPANISH FRESHWATER ECOSYSTEMS
M. Alvarez Cobelas, A. Rubio Olmo and P. Muñoz Ruiz Centro de Investigaciones del Agua (CSIC), La Poveda. Arganda del Rey, E-28500 Madrid. Spain.
Keywords: Eutrophication, Phosphorus, Ecosystem typology, Major Watersheds, Spain.
ABSTRACT
An account on the eutrophication of Spanish freshwaters is given here, based upon both published and unpublished data. Phosphorus was chosen as eutrophication index. Major watersheds and ecoaystem types were ranked by using yearly P-avera-
ges. On the whole, 713 sites are known for P-data. Júcar, Tajo and Eastern Pyrenees are the most heavily eutrophicated waters-
heds, whereas Duero, Segura and Southern watersheds show the least P contents. However, within and between variabilities
are very high. On the whole, streams are more eutrophic than stagnant waters but again variability in P contents within and between ecosystem types is high. Lakes show very strikingly trophic degrees, depending upon lake tipology and their envi- ronment. On an average basis, 80% Spanish lakes, 70% reservoirs and 60% river sites are eutrophic. In water courses, hyper- trophy is increasing downstream.
INTRODUCTION
In Spain, water quality standards have been largely neglected in the past despite the fact that water itself is an endangered, very scarce natural resource in our country.
Average rainfall is very low in most aseas, as compared with other temperate countries (OLIVER & FAIRBRIDGE, 1987), and experiences a remarkable seasonality. This results both in high water leve1 (in stagnant waterbodies)
and water discharge (in streams) fluctuations throughout the
year. These facts, ¡.e. neglecting water quality and fluctua- ting rainfall, are the main features operating on Spanish water resources and rendering them highly polluted in most
cases.
Unfortunately, there have been but a few attempts to fully survey water quality of Spanish freshwater ecosystems up to date (i.e., MINGO, 1983). Of course, severa1 independent
reports have been produced (MARGALEF et al., 1977;
ORTIZ CASAS & PENA, 1984; PRAT et al., 1985) but they are either local os not updated, which is a problem because of the high variability of water quality in Spanish waterbodies (RIERA et al., 1991).
Therefore, it is our aim here to provide a brief, up-dated account of water quality of Spanish freshwater ecosys- tems. Eutrophication is perhaps the best known of al1 processes threatening water quality. Nowadays, the key
role of phosphorus in eutrophication events is widely
recognized (OECD, 1982), and, accordingly, we shall use
phosphorus concentrations as an index of eutrophication in Spanish freshwater ecosystems, and hence as a water
quality index.
MATERIALS AND METHODS
This study has been carried out with already existing data
sets (published or not). All members of the AEL (Spanish Limnological Association) were requested to contribute data on phosphorus contents in freshwater ecosystems, but only
those data sets covering a whole year were used. Also published literature was searched on the topic. When two
distinct data sets on any given site were found, the most recent one was chosen. Phosphorus data have been yearly averaged for any site throughout the study to provide a
proper basis for comparisons since the sharp seasonality in Spanish freshwaters prevents meaningful comparisons if only single samples were used.
We classified the study sites according to ecosystem type and their affiliation to the majos watershed. The formes were included: irrigation channels, lakes, reservoirs, rivers, salt marshes, salt works, springs and temporary streams. The latter included: Northern watershed, Duero, Tajo, Guadiana,
Limnetica, 8: 263-266 (1992) O Asociación Española de Lirnnologin, Madrid. Spain
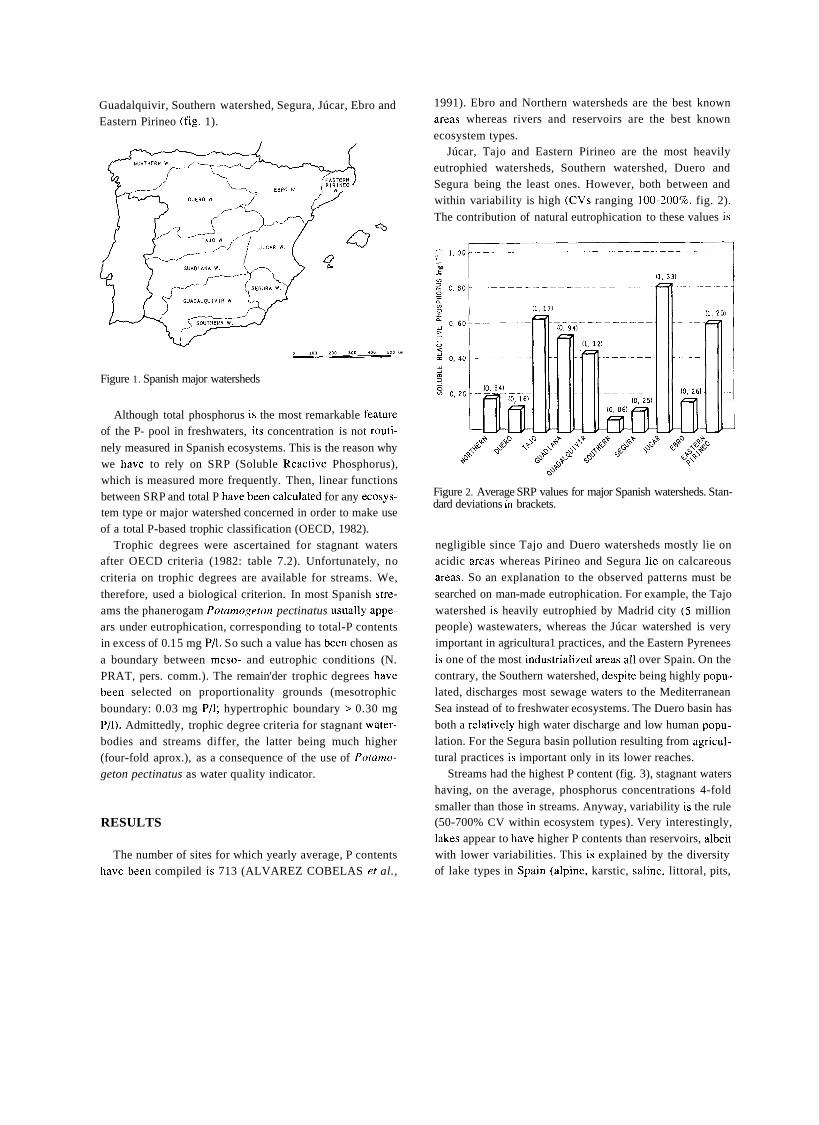
Guadalquivir, Southern watershed, Segura, Júcar, Ebro and Eastern Pirineo (fig. 1).
Figure 1. Spanish major watersheds
Although total phosphorus is the most remarkable feature of the P- pool in freshwaters, its concentration is not routi- nely measured in Spanish ecosystems. This is the reason why we have to rely on SRP (Soluble Reactive Phosphorus), which is measured more frequently. Then, linear functions between SRP and total P have been calculated for any ecosys- tem type or major watershed concerned in order to make use of a total P-based trophic classification (OECD, 1982).
Trophic degrees were ascertained for stagnant waters after OECD criteria (1982: table 7.2). Unfortunately, no criteria on trophic degrees are available for streams. We, therefore, used a biological criterion. In most Spanish stre- ams the phanerogam Potamogeton pectinatus usually appe- ars under eutrophication, corresponding to total-P contents in excess of 0.1 5 mg P/1. So such a value has been chosen as a boundary between meso- and eutrophic conditions (N. PRAT, pers. comm.). The remain'der trophic degrees have been selected on proportionality grounds (mesotrophic boundary: 0.03 mg P/1; hypertrophic boundary > 0.30 mg P/1). Admittedly, trophic degree criteria for stagnant water- bodies and streams differ, the latter being much higher (four-fold aprox.), as a consequence of the use of Potamo- geton pectinatus as water quality indicator.
RESULTS
The number of sites for which yearly average, P contents have been compiled is 713 (ALVAREZ COBELAS et al.,
1991). Ebro and Northern watersheds are the best known areas whereas rivers and reservoirs are the best known ecosystem types.
Júcar, Tajo and Eastern Pirineo are the most heavily eutrophied watersheds, Southern watershed, Duero and Segura being the least ones. However, both between and within variability is high (CVs ranging 100-200%, fig. 2).
The contribution of natural eutrophication to these values is
Figure 2. Average SRP values for major Spanish watersheds. Stan- dard deviations in brackets.
negligible since Tajo and Duero watersheds mostly lie on acidic areas whereas Pirineo and Segura lie on calcareous areas. So an explanation to the observed patterns must be searched on man-made eutrophication. For example, the Tajo watershed is heavily eutrophied by Madrid city (5 million people) wastewaters, whereas the Júcar watershed is very important in agricultura1 practices, and the Eastern Pyrenees is one of the most industrialized areas al1 over Spain. On the contrary, the Southern watershed, despite being highly popu- lated, discharges most sewage waters to the Mediterranean Sea instead of to freshwater ecosystems. The Duero basin has both a relatively high water discharge and low human popu- lation. For the Segura basin pollution resulting from agricul- tural practices is important only in its lower reaches.
Streams had the highest P content (fig. 3), stagnant waters having, on the average, phosphorus concentrations 4-fold smaller than those in streams. Anyway, variability is the rule (50-700% CV within ecosystem types). Very interestingly, lakes appear to have higher P contents than reservoirs, albeit with lower variabilities. This is explained by the diversity of lake types in Spain (alpine, karstic, saline, littoral, pits,
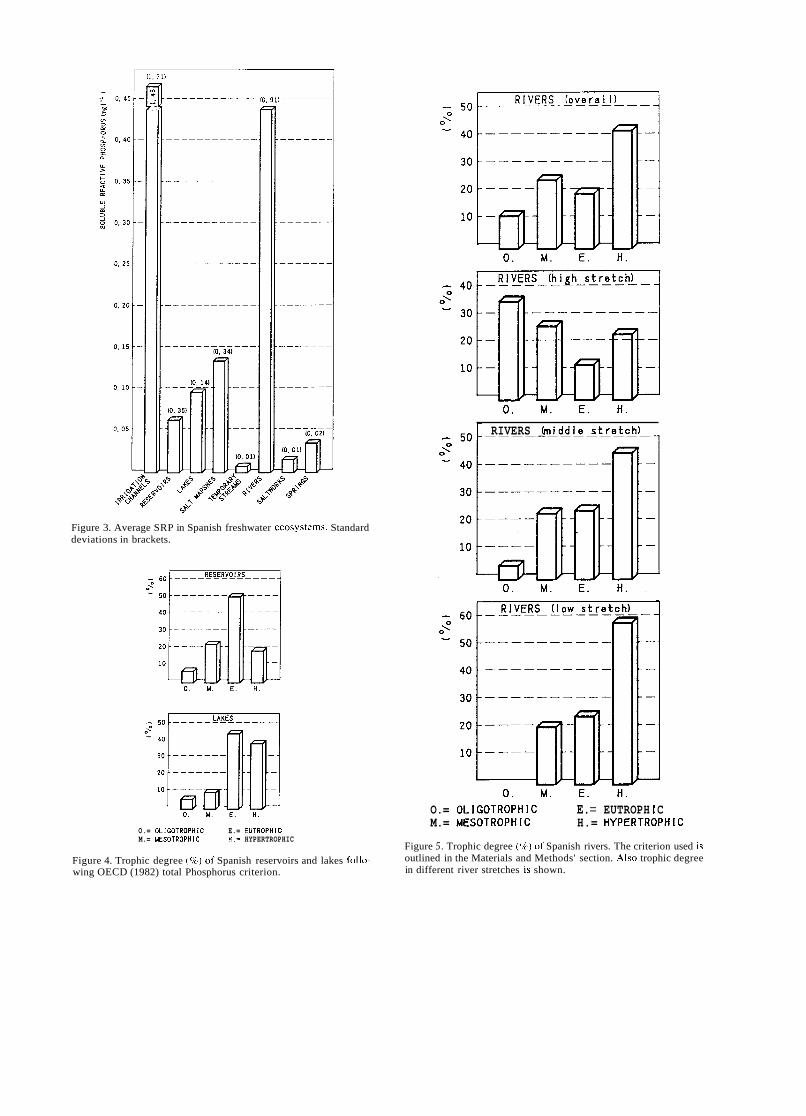
Figure 3. Average SRP in Spanish freshwater ecosysterns. Standard deviations in brackets.
O.= OLlGOTROPHlC E.= EUTROPHIC M.= MESOTROPHlC H.= HYPERTROPHIC
Figure 4. Trophic degree (%) of Spanish reservoirs and lakes follo- wing OECD (1982) total Phosphorus criterion.
RIVERS h i d d l e stretch) ----A--------
O.= OLIGOTROPHIC E. = EUTROPH 1 C M.= MESOTROPHlC H.= HYPERTROPHIC
Figure 5. Trophic degree (54) of Spanish rivers. The criterion used is outlined in the Materials and Methods' section. Also trophic degree in different river stretches is shown.

etc.). So karstic and alpine lakes show 0.030 - 0.034 mg P/1 (measured as SRP) whereas the remainder lakes are much more eutrophic (0.106 - 0.144 mg P/I).
As shown by OECD criteria using total P, 80% Spanish lakes are eutrophic, with hypertrophy accounting for half such a figure. Around 70% reservoirs are eutrophic but hypertrophy is still much lower (20%, fig. 4). Concerning
rivers, 60% out of the total sites are eutrophic (fig. 5 ) , the least eutrophic stretches being at the high course (fig. 5), but in middle and low stretches most sites are eutrophic (68% and 82%, respectively), and hypertrophy increases dramati-
cally downstream.
CONCLUSIONS
As a concluding remark, it is obvious that surhce water quality in Spain is far from good standards. But we hope that some remedial actions could be implemented in the near future.
ACKNOWLEDGEMENTS
This study could have not been undertaken without the financia1 support by Henkel Ibérica. Mr. Antonio Rosas (Environmental Chairman of Henkel Staff) was extremely helpful in coordination and suggestions. Prof Narcis Prat has provided a lot of useful comments and ideas before the study and on earlier drafts of manuscript. AEL (Spanish Limnological Association) promoted this study and many
of its Members supplied unpublished data. They are too
numerous to mention them al1 but we are very grateful to
them. Our deep gratitude is also due to an anonymous refe- ree whose suggestions have greatly improved this study. Finally, Fernando Delgado has kindly drawn the figures and Julián Michel checked the English language of the manus-
cript.
REFERENCES
ALVAREZ COBELAS, M.; P. MUÑOZ & A. RUBIO. 1991. La Eutrofizacicín de las Aguas continentales espuño- las. HENKEL IBERICA, S.A. Barcelona. 338 pp.
MARGALEF. R.; D. PLANAS; J. ARMENGOL; A. VIDAL; N. PRAT; A. GUISET; J. TOJA & M. ESTRADA. 1977. Limnología de los Embalses españoles. MOPU. Madrid. 422 pp. + appendixes.
MINGO, J. 1983. La Vigilancia de la Contaminación fluvial. MOPU. Madrid. 75 pp. + appendixes.
O.E.C.D. 1982. Eurrophication of Waters. Monitoving.
A.ssvssment and Control. O.E.C.D. Paris. 154 pp. OLIVER, J.E. & R.W. FAIRBRIDGE (eds). 1987. The
Encyclopedia o f Climatology. Encyclopedia of Earth Sciences, XI. Van Nostrand Reinhold Co. New York. xvi + 986 pp.
ORTIZ-CASAS, J.L. & R. PENA. 1984. Applicability of tlie OECD eutrophication models to Spanish reservoirs. Verh. Internat. Verein. Limnol. 22: 1521-1535.
PRAT, N,; G. GONZALEZ, X. MILLET & M.A. PUIG. 1985. El Fois, enti-e 1'ei.uutesa i la contaminacicí. Servei
Medi Ambient. Dipt. Barcelona. Est. Mongr., 1 1 . 92 pp. RIERA, J.L., E. MARTI & J.A. MORGUI. 1991. Changes
in the trophic state of the Spanish reservoirs during the last
sixteen years. Verh. Internat. Verein. Limnol. 24: 1367.1370.

FACTORS CONTROLLING ALGAL BIOMASS IN THE COMPLEX OF WATER SUPPLY RESERVOIRS OF SEVILLE (SPAIN)
J. ~ o j a ' , A. S as anta^ and R. Fernández ~ l e s l
l . Dept. Biología Vegetal y Ecología. Fac. Biología. Apartado 1095. 41080 Sevilla. Spain. 2. Empresa Municipal de Abastecimiento y Saneamiento de Aguas de Sevilla (EMASESA). Escuelas Pias 1 . 410 Sevilla. Spain.
Keywords: Reservoir, Alga1 biomass, Eutrophy. Water renewal rate. Nutrients
ABSTRACT
This paper summarizes more than a decade of research on Aracena. La Minilla and El Gergal reservoirs, which belong to the
water supply reservoir complex of Sevilla city (Spain). Aracena and La Minilla reservoirs were studied since July 1973 and El Gergal reservoir since October 1979. The results allowed the examination of the different factors which control thc interannual
variability of the trophic state of the reservoirs. Because of the high variability of average yearly rainfall, that is characteristic of the Mediterranean climate asea, the water
renewal rate of the reservoirs varies greatly from year to year. This water renewal rate plays a more important role than nutrient
(nitrogen and phosphorus) availability in controlling algal biomass and consequently, primary production. Nevertheless, its relative importance depends on the phosphorus loading of each reservoir. In the case of La Minilla reservoir, water renewal
rate is the only factor significantly correlated with phytoplankton biomass. In Aracena and El Gergal reservoirs both renewal rate
and nutrient concentrations can account for the differences in biomass. In Aracena reservoir the highest phytoplankton biomass
peaks occured in certain summers at the beginning of its history (when N/P ratio is less than 10) due to blooms of nitrogen fixing Cyanophyceae, but water renewal rate turned to be a more important controlling factor than nutrient concentration as years passed by. Although El Gergal reservoir presents the highest phosphorus input, summer blooms of Cyanophyceae do not
take place, due to its natural high water renewal rate (which has even been increased by human management). The largest algal
crops tend to occur during spring in El Gergal reservoir. where they reach the highest values of the 3 rcservoirs. Then, the phytoplankton biomass is maintained to similar standards during the stratification period (aproximately, 7-8 months). When phosphorus loading is extremely high, it becomes the only factor with significant importance on the control of the trophic state
of a reservoir, such as in the Sau reservoir (ARMENGOL, 1988).
INTRODUCTION
Water supply to Seville city (1 million inhabitants) is supported by a complex of 5 reservoirs located in the Rivera de Huelva basin, a tributary of the Guadalquivir river. Four of them (Aracena, Zufre, La Minilla and El Gergal reser-
voirs) are located on the main river while the fifth (Cala reservoir) is on one of its tributaries: the Rivera de Cala (fig. 1). The Aracena reservoir was first filled in 1969, Zufre in 1988, La Minilla in 1946, El Gergal in 1979 and Cala in
1936. The main use of Aracena. Zufre, La Minilla and El Gergal reservoirs is the water supply to Seville, whereas Cala reservoir is devoted to hydroelectric production, being
its outlet waters collected downstream in El Gergal reser-
voir. Since 1987 La Minilla reservoir has produced electri-
city too. La Minilla and El Gergal reservoirs allow extrac- tion of water at 4 different depths. In the other three it is only possible to extract water from the top and from the
botton of the dam. The basin of the reservoirs consists of Silurian acid rocks
and is hardly affected by human activity. The river water contains less than 250 mg.L-' of total dissolved solids (MARGALEF er c r l . . 1976: TOJA, 1976; ARMENGOL er
u / . , 1991) but has a relatively high alcalinity (1.2-1 .8 meq. L-') due to some tributaries from the calcareous zone of the Aracena Mountains (TOJA, 1976). The classification of
Limncticü. 8: 267-277 (1992)
O Amciación Española de Liiniiología. Madrid. Spain
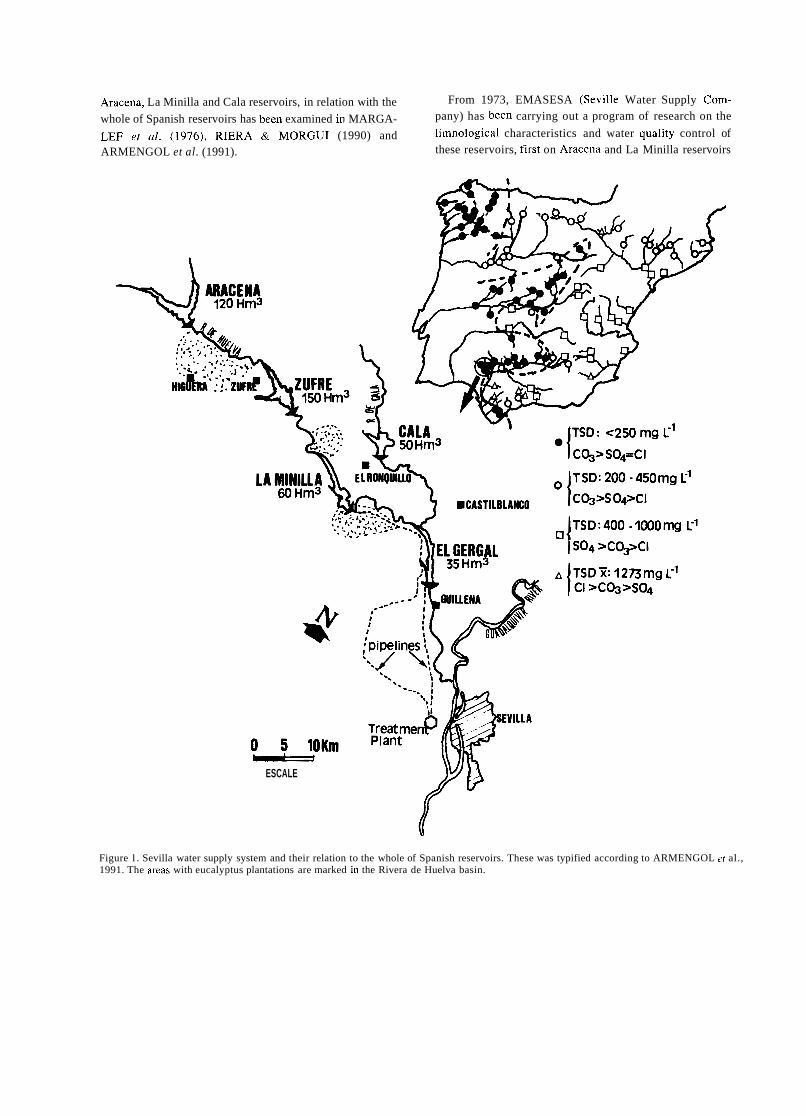
Aracena, La Minilla and Cala reservoirs, in relation with the From 1973, EMASESA (Seville Water Supply Com-
whole of Spanish reservoirs has been examined in MARGA- pany) has been carrying out a program of research on the
LEF et al. (1976), RIERA & MORGUI (1990) and limnological characteristics and water quality control of
ARMENGOL et al. (1991). these reservoirs, first on Aracena and La Minilla reservoirs
iCASTILBLAWE0
ESCALE
Figure l . Sevilla water supply system and their relation to the whole of Spanish reservoirs. These was typified according to ARMENGOL er al., 1991. The areas with eucalyptus plantations are marked in the Rivera de Huelva basin.

and later on El Gergal and Zufre reservoirs since their first inflow. Cala reservoir has only been studied sporadically.
The results of this research have been published in papers
describing aspects of the physicochemical characteristics of water (CASCO, 1990; TOJA, 1976, 1980a, 1984, 1990; TOJA er al., 1981) and sediments (GABELLONE & GUISANDE, 1989), phytoplankton (CASCO, 1990; TOJA, 1976, 1980a, 1984, 1990; TOJA et al., 198 1, 1983; TOJA &
CASCO, 1990), periphyton (CASCO, 1990; CASCO &
TOJA, 1991; TOJA & CASCO, 1990), zooplankton (TOJA, 1980b; 1983), fishes (SANCHO ROYO & GRANADO
LORENCIO 1987) and management (TOJA, 1982). In this work we will summarize the information obtained
from 1973 to 1989 in Aracena, La Minilla and El Gergal reservoirs, with emphasis on the observation of the factors that control their algal crop.
MATERIAL AND METHODS
Samples were taken monthly at one sampling station in
each reservoir located at S00 m from the dam. Aracena and La Minilla reservoirs were sampled from 1973 to 1989 and El Gergal reservoir from 1979 to 1989. In each reservoir samples were taken at different depths (0, 2, S, 10 m then at
every S m to the bottom). Nitrite was analyzed by the Shinn method (APHA 1985), nitrate by reduction to nitrite (APHA 1985), amnonia by Nessler reactive after precipitation with ZnS04 (APHA 1985) and soluble phosphorus according to
Musphy and Riley 1963 (in APHA 1985). Volumes of 1-2
L of water were filtered through fiber glass (WATMAN GF/C) filters, and the photosynthetic pigments were extrac-
ted in darkness in cold methanol. The Talling and Driver formula (VOLLENWEIDER, 1969) was used to calculate
chlorophyl1.a concentrations. Samples of 125 ml of water were fixed with solutions of iodine in potassium iodide for
later counts of phytoplankton in the inverted microscope according to the Utherm Ih technique.
The water renewal rate (RR) for each month was calcu-
lated according to the formula (TOJA, 1982):
Volume inflow + Volume stored in reservoir
RR (month-') =
total capacity of reservoir
This model is particularly appropiate for the Mediterra- nean reservoirs that have high level fluctuations both within a year and from year to year (PEREZ-MARTINEZ er ul.,
1991). It includes both the changes of the water flow and the dictance of the epilimnion to the bottom that determines the
posibility of the nutrient return to the euphotic zone. Relationships between algal biomass and limnetic
conditions were described using Multiple Regression Analysis (Statgraphics Statistical Computes Program 1985). Phytoplanktonic biomass was represented by the average monthly epilimnetic (O to 10 m of depth) chlo-
rophy1l.a concentration ( p g . ~ . ' ) . The logarithmic trasfor-
mation of the concentration of ch1orophyll.a was used since primary producers respond normally to environmen-
tal factors in an exponential way. The following factors
related to biomass were considered: the monthly water
renewal rate (RR in month-'), the average monthly con- centrations of total inorganic nitrogen (NO3 + NO2 + NH,) (N in pg-at N.L.') and of soluble reactive phosphorus (P in pg-at P.L.') in the water column, and the ratio between
both nutrients (N/P).
RESULTS
The main characteristics of the three reservoirs are shown in table 1. Inter-annual variability in al1 parameters is very high
and is not reflected by the average values. The climatic irre- gularity of the Mediterranean area is reflected in the volume of water stored in the reservoirs over the years (fig. 2), espe-
cially in Aracena reservoir, which is used as a regulation dam. Although the interannual variation in La Minilla and El Gergal
reservoirs is lower than in Aracena reservoir, the fluctuation
within the annual cycle is much greater. Therefore, the yearly water renewal rate in the three reservoirs has been very varia-
Figure 2. Evoluiion in time of the water volume ( ~ r n j ) stored in each reservoir

ble, even higher than reflected in the annual average (Table 1 ) (Aracena between 0. 18 and 2.20 yr-'; La Minilla: 0.68-9.44 yr-'; El Gergal: 1 .lo-14.85 yr-').
The level of eutrophy in Aracena and La Minilla is not very
high at present, as they are located in a basin that is relatively
unaffected by man. Eutrophy is higher in El Gergal due to its more recent construction and to the following circumstances. During the period 1981-83, water from the river Guadalquivir was let in, with a concentration of nitrogen and phosphorus
about ten times greater than that of the river Rivera de Huelva. Moreover, this reservoir receives wastewaters, directly from the village of Castilblanco (approximately 8.5 Tm P.yr'l) and indirectly from the Cala reservoir which receives the waste-
waters from El Ronquillo village. This has caused greater phosphorus inputs than in other reservoirs. The estimated phosphorus loadings are 0.36 g.m-2.yr-1 in Aracena , 1.22 g.m- 2.yr-1 in La Minilla and 5.1 1 g.m-2.yr-' in El Gergal.
When multiple regresions with the monthly data are carried out, taking the logarithm of the chlorophyll concen- tration as the dependent variable and considering the monthly water renewal rate, the average concentration of
total inorganic nitrogen, the average concentration of solu-
ble reactive phosphorus, and the N/P ratio, as independent variables, the following equations are found:
For Aracena: LogChl. = 1.135 - 0.268 RR - 0.0003 N (1) (n=161, r2=0.152, MS=0.519, F=7.035, p 50.001)
For La Minilla: LogChl. = 0.47 - 0.21 8 RR (2) (n=193, r2=0. 14, MS=0.666, F=8.787, p 50.001)
For El Gergal: Log CHI. = 1.1 16 - 0.243 RR - 0.162 P (3) (n=118, r2=0.23, MS=1.163, F=8.456, p áO.OO1)
Only the variables with a significant correlation coeffi- cient were considered.
When we consider the yearly averages, only the water renewal rate seems to be related with algal biomass in
Aracena and La Minilla reservoirs, although this may be due to the small number of cases, as the significant level of the coefficient of N in the equation 1 is very low (p 10.05). The equations found are the following:
For Aracena: LogChl. = 1.305 - 0.384 RR (4) (n=12, r2=0.79, MS=0.054, F=7.789, p 50.01)
For La Minilla: LogChl. = 0.671 - 0.194 RR (5) (n=15, r2=0.49, MS=0.029, F=2.72, p 10.05)
There is no significant relationship in El Gergal reservoir, possibly due to the few years of operation of this reservoir.
Table 2 shows the correlations obtained from these para- meters. In al1 reservoirs the higher correlation coefficients are those between the log chlorophyll and the water renewal rate as the independent variable. Relationships with the main
nutrients vary among reservoirs. In La Minilla reservoir, although there is no significant
correlation between chlorophyll and nutrients, a progressive increase of the winter phosphorus input has been recorded
through the years, bringing about a shift in the phytoplank- ton species composition (fig. 3). Thus, on diminishing the
N/P ratio, the relative importance of nitrogen fixing
Cyanophyceae has increased. Figure 4 represents the simple regresion of log chlo-
rophyll with water renewal rate obtained in each reservoir both from monthly data (fig. 4A) and annual average data
(Fig 4B). It can be observed in fig 4A that most data form a compact cluster, except for a few data corresponding to very rainy months. To test whether there was a functional corre- lation among these parameters and no stadistic artefact exis-
ted, a simple correlation analysis was done only with data
from this data cluster (RR < 2 month'l). As a result, the correlation coefficient was lower but still significant in La Minilla reservoir (r=-0.19, p 10.01) and it was slightly lowe- red in El Gergal reservoir (r=-0.38, p 20.001).
In both reservoirs, most of the monthly data which presented a water renewal rate of less than 1 month-' and a log chlorophyll value below 0.5, corresponded to either dry
winters or to stratification periods when water was outflo-
wed from the epilimnion. When the average annual values are considered, a signi-
ficant negative correlation between log chlorophyll and water renewal rate is maintained in Aracena and La Minilla
(r=-0.56, p 10.05 for Aracena and r=-0.61, p 10.05 for La Minilla) reservoirs (fig.4B). In El Gergal the relationship is negative but not significant, probably due to the scarce data.
DISCUSSION
Although such factors as nutrient concentration in water
and incoming radiation are fundamentally responsible for the algal crop, additional factors such as dilution (or water renewal), sinking and grazing can be also important. The latter two factors do not exclude recycling of nutrients back from the sediments. Hence, increasing the water renewal rate is potentially the most effective mechanism
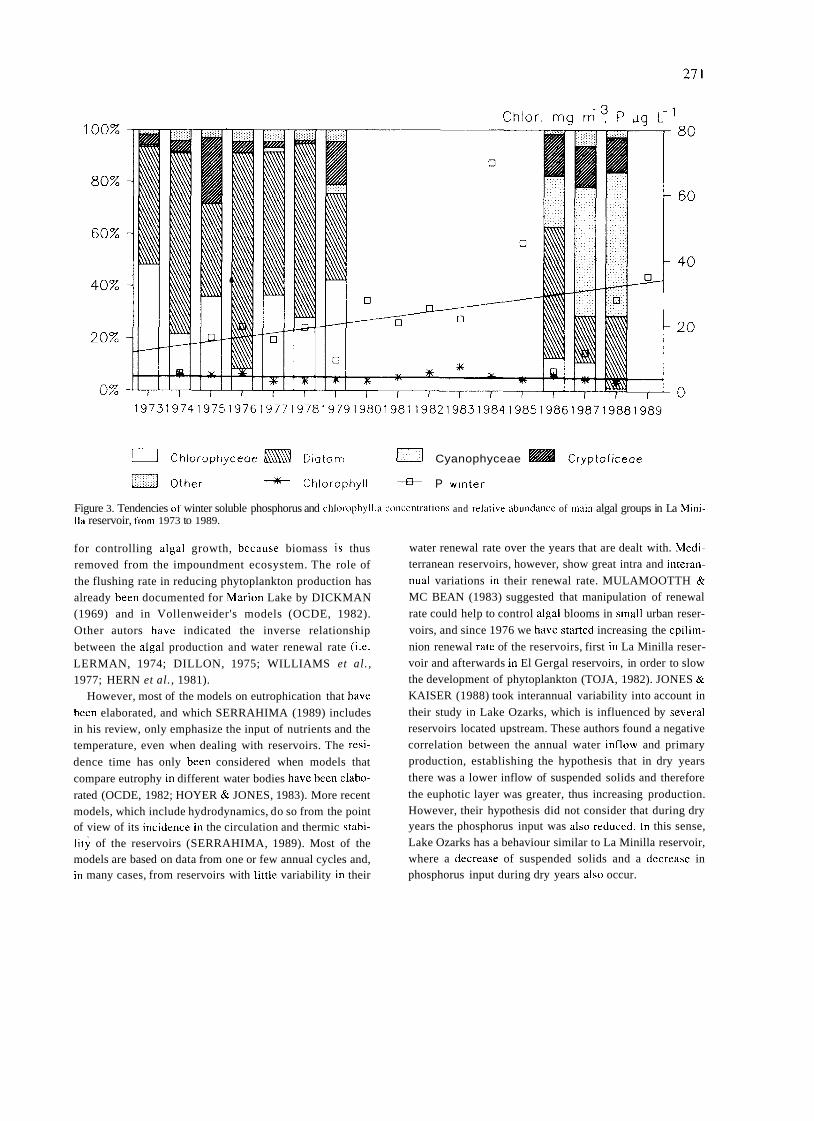
Chlorophyceae m Diatorri 0 Cyanophyceae m Cryptoficeoe
0 0ther iIc Chlorophyll P winter
Figure 3. Tendencies of winter soluble phosphorus and chlorophyll.a coriceiitrations and relative abundante of main algal groups in La Mini- Ila reservoir, from 1973 to 1989.
for controlling alga1 growth, because biomass is thus removed from the impoundment ecosystem. The role of
the flushing rate in reducing phytoplankton production has already been documented for Marion Lake by DICKMAN (1969) and in Vollenweider's models (OCDE, 1982).
Other autors have indicated the inverse relationship
between the algal production and water renewal rate (¡.e.
LERMAN, 1974; DILLON, 1975; WILLIAMS et al. , 1977; HERN et al., 1981).
However, most of the models on eutrophication that have
been elaborated, and which SERRAHIMA (1989) includes in his review, only emphasize the input of nutrients and the temperature, even when dealing with reservoirs. The resi-
dence time has only been considered when models that
compare eutrophy in different water bodies have been elabo-
rated (OCDE, 1982; HOYER & JONES, 1983). More recent models, which include hydrodynamics, do so from the point of view of its incidence in the circulation and thermic stabi- lity of the reservoirs (SERRAHIMA, 1989). Most of the models are based on data from one or few annual cycles and, in many cases, from reservoirs with little variability in their
water renewal rate over the years that are dealt with. Medi- terranean reservoirs, however, show great intra and interan-
nual variations in their renewal rate. MULAMOOTTH & MC BEAN (1983) suggested that manipulation of renewal
rate could help to control algal blooms in small urban reser-
voirs, and since 1976 we have started increasing the epilim-
nion renewal rate of the reservoirs, first in La Minilla reser- voir and afterwards in El Gergal reservoirs, in order to slow the development of phytoplankton (TOJA, 1982). JONES &
KAISER (1988) took interannual variability into account in
their study in Lake Ozarks, which is influenced by several reservoirs located upstream. These authors found a negative correlation between the annual water intlow and primary production, establishing the hypothesis that in dry years
there was a lower inflow of suspended solids and therefore
the euphotic layer was greater, thus increasing production. However, their hypothesis did not consider that during dry years the phosphorus input was also reduced. In this sense, Lake Ozarks has a behaviour similar to La Minilla reservoir, where a decrease of suspended solids and a decrease in phosphorus input during dry years also occur.
m -Q3 . m. m 1
O 45 1 1.5 2 2.5 3
Renewal rate month-'
A ARACENA
Renewal rate . month-'
1.6- m 1.28 . .y=i.004 -0.21~ 1.16:
k
Renewal rate month-'
. y=1.09-0.25~ r =- 0.56,pt0.05
Renewal rate . month-' Figure 4. Linear regressions between the water i-enewal riite niicl the logaritliiii ot'chloi-opliyll.n coiicentratioii in eacli re\er\'ois. A: using monthly data, B: using the annual average.
: z c a o Q96: leo? 7 O 0.3 0.6 0.9 1.2 1.5 O 0.2 0:4 0:6 Ci8 i 1:2
Renewal rate. month-' Renewal rate . mont h-'
m
. m
L L
O - O - 5 Q8G, L: O 0.4- d . O 4
0 -
. - m 5 m=' . ¤ • ' 8
b . % 0.761 4
Q66
Table 1. Main Characteristics of the 3 reservoirs (Anual average and standard desviation). For chlorophyll only epilimnetic values was used. For inorganic nitrogen (N03+N02+NH4) and soluble phosphorus values from al1 water column were used.
HYDROLOGIC YEAR
RENEWAL RATE CHLOROPHYLL INORGANIC N SOLUBLE P month-' mg. m-' pg-at. L-' pg-at. L.'
X u X (3 X u X u
ARACENA 1973-74 1974-75 1975-76 1976-77 1977-78 1978-79 1979-80 1980-8 1 1981-82 1982-83 1983-84 1984-85 1985-86 1986-87 1987-88 1988-89
LA MINILLA 1973-74 1974-75 1975-76 1976-77 1977-78 1978-79 1979-80 1980-8 1 198 1-82 1982-83 1983-84 1984-85 1985-86 1986-87 1987-88 1988-89
EL GERGAL 1979-80 1980-8 1 1981-82 1982-83 1983-84 1984-85 1985-86 1986-87 1987-88 1988-89
16.20 8.90 3.58 15.27 5.93 7.66 9.90 4.40 34.62 7.75 5.1 1 10.70 9.00 3.83 4.66 7.70 3.37 5.30 10.61 3.85 2.24 empty most of the time empty most of the time empty most of the time 5.69 5.29 39.89 10.38 8.04 9.80 7.90 1.63 16.31 7.39 1.78 28.4 1 6.61 4.64 28.82 4.69 2.61 16.75

The negative relationship between algal biomass and
water renewal rate is expected since an increaae in the latter
implies an extraction of biomass from the reservoir, espe-
cially if epilimnion water is extracted preferently. as it
happened in La Minilla reservoir (since 1976) and in El
Gergal reservoir (since it came into service). the two reser-
voirs that show the highest correlation coeficient between
these two factors. An increase in the renewal rate can also
lead to a losa of nutrients (LERMAN, 1974), which may
determine a drop in biomass in the following months. In
years with high rainfall during the mixing period there will be a lower production during the stratification period, and
average annual biomass will be lower than in drier years.
In Aracena reservoir, an increase in water renewal rate
leads to a reduction in phosphorus concentration (negative
correlation between RR and P in table 2). Despite this nega- tive relationship, there is no relationship between soluble
phosphorus and log chlorophyll. In this reservoir we find a
negative relation between nitrogen and log chorophyll, since
the largest algal crops can be related to Cyanophyceae
blooms, when NIP ratio is low (below 10). Although, equa- tion 1 does not include the N/P ratio, a negative correlation
exists between log chlorophyll and N/P ratio (table 2). In the
first years of the study there was a relatively high phospho- rus concentration, higher than in La Minilla reservoir, due
to the nutrients released from the flooded terrestrial vegeta-
tion when the reservoir was first filled. On the springs of
these first years the nitrogen was rapidly removed from the
epilimnion (due to phytoplankton uptake) and from the hypolimnion (due to denitrification), thus favouring the
proliferation of nitrogen fixing Cyanophyceae in summer
(TOJA, 1976; 1984). The initial nutrient stock did not incre-
ase, as the reservoir's catchment area is small and in a good state of conservation. Consequently, water inflow during
wet years contributed to the dilution of these nutrients and
their removal from the reservoir, which, except on rase
occassions, takes place through the outlet at the bottom. We
can conclude that in Aracena reservoir nutrients seem to
have been controlling algal biomass, although lately they
have less importance than the water renewal rate (equations
1 and 4). In El Gergal reservoir, although the water renewal rate is
still the most important factor, phophorus concentration also
seems to detemine algal biomass and, probably because this reservoir has a continuous input ofthis element (17.9 Tm.yr-'), the soluble phosphorus concentration appears with a signifi- cant coeficient in equation 3. Nevertheless, relationsiphs between algal biomass and phosphorus concentration are nega-
tive. This can be partly due to a statistical interference. In El Gergal reservoir, an increase in water inflow leads to an
increase in phosphorus concentration as it is shown by the
positive correlation between RR and P (table 2 ) since it receives waters with high phosphorus concentration (see
above). The strong negative correlation between the water
renewal rate and the algal biomass, together with the nega-
tive correlation between water renewal rate and phosphorus
may consequently produce a negative correlation between
phosphorus and chlorophyll.
El Gergal reservoir bears a higher P loading, which favours
phytoplankton growth in such a way that algal biomass reachs the highest levels of the 3 reservoirs during spring prolifera-
tions. However, its high water renewal rate makes algal
biomass decrease os remain the same during the rest of the
stratification period (approximately 7 months). Also, the
nitrogen fixing Cyanophyceae blooms did not take place
during summer as it happened in Aracena reservoir during its
first years of operation, although a nitrogen deficit can also
be detected in El Gergal reservoir during the summer. This is
due to the management of this reservoir, with water extrac-
tions taking place from the epilimnion during the stratifica-
tion period. This artificial increase of the water renewal rate
from the productive layer resulted in a continuous decrease
of the algal biomass, thus preventing algal blooms. Therefore, the highest algal crop is not registered when the N/P ratio is
lower and thus, there is no negative correlation between algal
biomass and nitrogen , as it happened in Aracena reservoir.
Then, the average chlorophyll concentration in El Gergal
reservoir is similar os everlower than in Aracena reservoir, in
spite of the highern phosphorus input in El Gergal reservoir.
Nevertheless, this P loading not being excessively high can
be almost totally consummed very rapidly. Then, the concen- tration of soluble phosphate is rather low during the stratifi-
cation period, accounting Sor the negative correlation between
P and algal biomass (table 2).
There is not significant correlation between water rene- wal rate and nutrients in La Minilla reservoir (table 2). It has
a larger catchment area and the nutrients input can vary a
great deal as the inflow also varies, that is, it depends on
how rainfall is distributed. If, after several dry years, a large
water inflow occurs, the nutrients input (especially phosp-
horus) increases very much. This happened, for example in
1976-77 (phosphorus loading was 2.12 g.m-'.yr-') and in 1983-84 (4.33 g.m-2.yr-'). There was also an increase in the
proportion of suspended solids (as in Ozarks lake), but they soon settled and during the stratification period (7 -8 months long, aproximately) the amount of inorganic seston was

Table 2. Significant correlations between the different parameters analyzed in each reservoir using monthly data. RR= water renewal rate month-', N= inorganic nitrogen (N03+N02+NH4) pg-at. L.', P= soluble phosphorus kg-at. L.', N/P ratio between inorganic nitrogen and soluble phosphorus
ARACENA (N = 162)
logchl. N P NIP
logchl. N
EL GERGAL (N = 1 18)
logchl N
independent variables N P
-0.23*** 0.31****
0.31**** 0.33**** .0.26****
similar to the drier years (CASCO, 1990). In drier years, with lit tle water inflow, there was a small phosphorus loading, i.e. in 1982-83 (0.22 g.m-2.yr-') (fig. 3).
In La Minilla reservoir, only the water renewal rate determines algal biomass, nutrients not seeming to have any importance (equations 2 and 5 ) . However, from 1973 to 1989, the phosphorus input into the reservoir tended to increase (fig. 4); firstly because of the removal of natural vegetation upstream in order to plant Eucalyptus (fig. 1) in 1976 (TOJA, 1984; TOJA et al., 1983), secondly, because of the effect of the cons~uc t ion of the Zufre reservoir, and after, because of the release of the nutrients stocked in this reservoir that were sent with water downstrean to La Mini- lla reservoir (CASCO, 1990). However, the phytoplankton
biomass tended to remain at the same level and even, to fall slightly, due to the high natural water renewal rate as well as to an increase in the epilimnion renewal rate, caused by the opening of the reservoir's upper outlets (TOJA, 1982).
However, the composition of the phytoplankton has chan- ged appreciably (fig. 3). Cyanophyceae, especially nitrogen
fixative types, used to be very scarce in this reservoir (TOJA, 1984). As far back as 1977-79, they began to appear in higher proportions than during the previous years (TOJA et al., 1983). In 1986-89 (CASCO, 1990) with the building and coming into service of Zufre reservoir, the population has changed noticeably, with a predominance of Cyanophy- ceae during the stratification periods.
In Mediterranean reservoirs with a continuous phospho- rus input much greater than the one El Gergal reservoir receives, the importance of the mentioned factors can be the opposite. The Sau reservoir (on the river Ter in Catalonia) is the reservoir where the longest series of data have been recorded in Spain, as it has been studied continuously since 1963 (VIDAL, 1976; ARMENGOL, 1988; ARMENGOL et
al., 1986; ARMENGOL & VIDAL, 1988). It has a similar water renewal rate than the Seville's reservoirs (2.12-7.50 yr-1) and it has been receiving increasing amounts of phosp- horus every year since it carne into service (from 30 Tm.yr-
in 1965 to 215 in 1985), with an average loading of 26 g m-2yr-1 while the water renewal rate fluctuates with time. Using data obtained from ARMENGOL (1988), the equa-

tion of multiple regression bet-ween the log chlorophyll, the annual water renewal rate (yr-') and the soluble phosphorus has been calculated as follows:
In this case phosphorus is the factor more correlated with algal biomass. Even when considering the water renewal rate it seems to be different to the Seville's reservoirs. For exam- ple, with water renewal rates that were high and similar in 1971 and 1982, concentrations of P in the reservoir were 123 and 240 pg L-' respectively, which meant a greater algal biomass in 1982. When comparing the evolution of the water renewal rate and the chlorophyll concentration, we can see an inverse relationship, though not significant (r=-0.12, p: n.s.). Yet there is a significant inverse relationship as far as phosp- horus is concerned (r=-0.46, p 10.05).
That is to say that an increase in the water renewal rate always tends to diminish algal crop, even though the beha- viour of each reservoir throughout its life is different accor- ding to the nutrients input from its catchment area. In reser- voirs where this contribution is low, the water renewal rate is the main controlling factor as far as the eutrophic state is concerned (and may be the only one, as in the case of La Minilla reservoir), while if the amount of phosphorus incre- ases, its relative importance in algal biomass increases as well until it completely overlaps the effect of the water rene- wal rate (as in the case of Sau reservoir). As REICHLE et
al. (1980) have pointed out: biomass in ecosystems depends on two factors: environmental favourability (in this case, nutrient availability) and disturbance. When disturbance that destructs biomass is very frequent (as a high water renewal rate), it may be the main factor controling biomass, counte- racting the positive effect of the habitat favourability.
It is dificult to determine the phosphorus threshold input above which the phosphorus loading becomes the main controlling factor. DILLON (1975) found that Cameron Lake (Ontario), with a phosphorus loading of 1.7 - 2.21 g.m- 2.yr-', was not eutrophic owing to its high flushing rate. The results obtained from Sevilla's reservoirs suggest that when the phosphorus loading remains at low levels (less than 2 g.m-2.yr-1) the water renewal rate is the controlling factor of algal biomass, even if the nutrient concentration and N/P ratio affect the qualitative composition of phytoplankton. However, when the phosphorus loading is higher, it beco- mes the dominant factor controlling algal crop, although the renewal rate is still significant.
This rapid control effect of water renewal rate on algal biomass has been and is still used to manage the water supply reservoirs of Sevilla (La Minilla and El Gergal reser- voirs). During the stratification period water is preferently extracted from the epilimnion thus, increasing proportio- nally the water renewal rate of the photic zone (TOJA, 1982) and keeping algal biomass at a lower level than the one that could be expected from the nutrient concentration in the water. Also, during the mixing periods, when outflow operations are needed in El Gergal reservoir (in wet winters), the water is extracted from the outlet at the botton, increasing the hypolimnion renewal rate and removing nutrients from the reservoir. This water management has improved the water quality and therefore has implied a great economical advantage for the drinking water supply mana- gement to Seville city (TOJA, 1982, 1984).
ACKNOWLEDGEMENTS
This work was supported by Sevilla Water Supply Company (EMASESA).
REFERENCES
APHA, 1985.- Standard methods for the examination of
water and wastewater. 16th ed. American Public Health Assoc. Washington DC.
ARMENGOL, J., 1988.- Contaminació de conques hidro- gráfiques y malmenament d'aquifers. Degradació de rius i eutrofizació d'embassaments. Natura ús i abus?. En R. Folch (ed) Llibre Blanc de la gestió de la Natura (2 Ed). Ed Barcino: 141 -150.
ARMENGOL, J. & VIDAL, A., 1988.- The use of diffe- rent phosphorus fractions for the estimation of the trophic evolution of the Sau reservoir. Arch. Hydrobiol. Beih. Ergebn. Limnol. 30: 61-70.
ARMENGOL, J., CRESPO, M., MORGUI, J.A. & VI- DAL, f.., 1986.- Phosphorus budgets and forms of phosp- horus in the Sau reservoir sediment: An interpretation of the limnological record. Hydrobiologia, 143: 33 1-336.
ARMENGOL, J., RIERA, J.L. & MORGUI , J.A. 1991. Major ionic composition in the Spanish reservoirs. Verh. Internat. Verein. Limnol., 24: 1363-1366.
CASCO, M.A., 1990.- El perifiton del embalse de La Minilla. Relaciones con el fitoplancton y contribución a la producción total. Tesis Doctoral. Univ. de Sevilla.

CASCO, M.A. & TOJA, J. 1991. Benthic microalgae of La Minilla reservoir (SW, Spain). Verh. Internat. Verein. Limnol., 24: 1386.1389.
DICKMAN, M., 1969.- Some effects of lake renewal and phytoplankton productivity and species composition. Limnul. Oceanogr.. 14; 660- 666
DILLON, P.J., 1975.- The phosphorus bugget of Came- ron Lake Ontario. The importance of flushing rate to the degree of eutrophy of lakes. Limnol. Oceanogr., 20: 28-39
GABELLONE, N.A. & GUISANDE, C. 1989. Relations- hip between texture and fractions of inorganic phosphorus in the surface sediment of a reservoir. Aquatic Sciences,
5 / (4 ) : 306-316 HERS, S.C., LAMBOU, V.W., WILLIANS, L.R. &
TAYLOR, W.D., 198 1.- Modifications of models predicting trophic state of lakes. Adjustamen t of models to account for the biological manifestation of nutrients. U.S. Environmen- tal Protrcction Agency EPA600lS3-81-001. Las Vegas.
HOYER, M.V. & JONES, J.R., 1983.- Factors affecting the relation between phophorus and chlorophyl1.a in Midwes- tern reservoirs. Can. J . Fish. Aqiat. Sci. 40: 192-199.
LERMAN, A. 1974.- Eutrophication and water quality of lakes: control by water residence time and transport to sedi- ments. Nyclr.olo~ic~cr1 S r i . Rltll.. 19: 25-45
MARGALEF, R., PLANAS, D., ARMENGOL, J., VIDAL, A., PRAT, N., GUISET, A., TOJA, J. & ESTRADA, M. 1976. Limnologia de los embal.ses españoles. D.G. Obras Hidraúli- cas, Serv. Publ. M.O.P. Madrid: 422+85 pp.
MULAMOOTTH, G. & MCBEAN, E.. 1083.- Detention time - A key decision factor in controling alga1 blooms in man-made lakes. Can. J. of Civil Enginering, lO(3): 450-455
OCDE (VOLLENWEIDER, R.A. ED.) 1982. Eutrophi- sation des eaulc. Methodes de surveillance, d'evaluation et de lutte. OCDE. Paris: 164 pp.
PEREZ-MARTINEZ, C., MORALES-BAQUERO, R. & SANCHEZ-CASTILLO, P., 1991: The effect of the volume
decreasing on the trophic status in four reservoirs from Sout- hern Spain. Verh. Internat. verein. Limnol., 24: 1382-1385
REICHLE, D.E., O'NEILL, R.V. & HARRIS, W.F., 1980.- Principios de intercambio de energía y de materia en los ecosistemas. En W.H. van Dobben & R.H. Lowe- McConnell (Eds.) Conceptos unificadores en Ecología. Ed Blume. Barcelona: 36-57.
RIERA J.L. & MORGUI , J.A., 1990.- Limnología regio- nal de los embalses españoles. Mundo cientljcico, 10: 720-727
SANCHO ROYO, F. & GRANADO LORENCIO, C. 1988. La pesca en los embalses andaluces. Cuadernos del Inst. Desarrollo Regional de Sevilla n 28: 225 pp.
277
SERRAHIMA, F., 1989.- Modelo matemático de simula- ción y control de la calidad del agua para embalses. Tesis Doctoral. Univ. Politécnica. Barcelona.
STATGRAPHICS, 1985.- STSC User's Guide. Software
Publ. Group. Rockville: 635 pp. TOJA, J., 1976.- Estudio ecológico comparado de dos
embalses con distinto grado de eutrofia: Aracena y La Mini- Ila. Tesis Doctoral. Univ. Barcelona.
TOJA, J. 1980a. Limnología del embalse de La Minilla durante 1976. 1. Ciclo del fitoplancton en relación con los factores del medio. Oecol. aquat., 4: 71 -88.
TOJA, J. 1980b. Limnología del embalse de La Minilla durante 1976. 11. Distribución del zooplancton. Oecol.
aquat., 4: 89-1 10. TOJA, J. 1982. Control de la eutrofia de embalses por
utilización selectiva de agua a distintas profundidades. Rev. de Obras Públicas. Abril-Mayo: 223-231.
TOJA, J. 1983. Zooplancton de los embalses de Aracena y La Minilla durante 1977. In: N. Prat (ed) Actas del I Congreso Español de Limnologia: 105-1 14. Barcelona.
TOJA, J. 1984. Limnologia de los embalses de abasteci- miento de agua a Sevilla. Publ. del CEDEX. MOPU. Madrid: 159+30pp.
TOJA, J. 1990. Longitudinal differentiation according to environmental factors and phytoplankton in Aracena and La Minilla reservoirs. Arch. Hydrobiol. Beih. Ergebn. Limnol. 33: 733-747.
TOJA, J. GONZALEZ RULL, J.A. & RAMOS, D., 1981.- Evolución del embalse de El Gergal en sus dos primeros años de vida. Actas del 1 Simposio del Agua en Andalucía: 1: 167- 180.
TOJA, J. GONZALEZ RULL, J.A. & RAMOS, D. 1983. Phytoplankton succession in Aracena, La Minilla and El Gergal reservoirs. Water supply, l ( 1 ) : 103-1 13.
TOJA, J. & CASCO, M.A. 1990. Contribution of phytoplankton and periphyton to the production in a reser- voir of SW Spain. Oecol. aquat ., 10 (in press).
VIDAL, A,, 1976.- Eutrofización del embalse de Sau en el transcurso de sus primeros datos (1 963-1 972). Conf. Hidrog. Pirineo Oriental. M.O.P. Cardedeu, Barcelona: 43 pp.
VOLLENWEIDER, R.A. 1969.- Primary production in aquatic environments. 1.B.P.Handbook n 12, Blackwell Sci. Publ., Oxford.
WILLIANS, J.R., LAMBOU, V.W., HERN, S.C. & THOMAS, R.W., 1977.- Relationships of productivity and
problem conditions to ambient nutrients: National eutrophi- cation findings for 418 eastern lakes. Nat. Eutrophication Survey. U.S.IEPA. Las Vegas.