a map of the biotin repressor-biotin operator interface: binding of a winged helix-turn-helix...
TRANSCRIPT

J. Mol. Biol. (1998) 278, 787±800
A Map of the Biotin Repressor-Biotin OperatorInterface: Binding of a Winged Helix-turn-helix ProteinDimer to a Forty Base-pair Site
Emily D. Streaker and Dorothy Beckett*
Department of Chemistry andBiochemistry, University ofMaryland Baltimore County1000 Hilltop Circle, BaltimoreMD 21250, USA
Abbreviations used: HTH, helix-twinged HTH; HSF, heat shock facto
0022±2836/98/190787±14 $25.00/0/mb9
The Escherichia coli biotin repressor is a member of the ``winged helix-turn-helix'' class of site-speci®c DNA binding proteins. The protein bindsas a dimer to the 40 bp biotin operator sequence. Although the structureof the aporepressor has been solved by X-ray crystallographic techniques,no structure of the holorepressor-DNA complex is yet available. In orderto characterize the structural features of the biotin repressor-biotin oper-ator interface we have applied a number of solution techniques includingDNase I, hydroxyl radical and dimethyl sulfate footprinting and the cir-cular permutation or ``bending'' assay. Results of these combined studiesindicate that each repressor monomer forms a bipartite interface witheach half-site of the biotin operator sequence. The results imply that, inaddition to the helix-turn-helix module of each monomer, a second struc-tural element participates in the protein-DNA interface. The two bipartiteprotein-DNA interfaces appear, moreover, to primarily involve the two12 bp termini of the operator site. Results of combined DNase I footprint-ing and circular permutation analysis indicate, furthermore, that the cen-tral 16 bp region that links the two termini becomes distortedconcomitant with binding of holoBirA.
# 1998 Academic Press Limited
Keywords: site-speci®c DNA binding; chemical footprinting; DNAbending
*Corresponding authorIntroduction
Structural studies of sequence-speci®c protein-DNA complexes have revealed general themes forinteraction of particular DNA binding motifs withtarget sites on DNA. For example, the helix-turn-helix motif is typically associated with binding inthe major groove of a DNA target site (Brennan,1992). Ribbon-helix-helix proteins that utilize twob-strands in site-speci®c DNA binding also contactthe DNA in the major groove (Raumann et al.,1994). In the context of these general models vari-ations are observed, for example, with respect toorientation of the recognition motif in the DNAgroove. In addition to the common recognitionmotifs, other structural elements in the DNA bind-ing domains of site-speci®c DNA binding proteinshave been shown to be signi®cant for speci®city.Among these are the N-terminal arms of the bac-teriophage lambda repressor which contact theminor groove on the backside of the DNA helix at
urn-helix; wHTH,r.
81733
the center of the operator site (Jordan & Pabo,1988) or the minor groove recognition helicesobserved in the PurR-purF complex (Schumacheret al., 1994). As the number of available high resol-ution structures of protein-DNA complexes con-tinues to grow, a broader range of variation inutilization of structural themes in DNA recognitionwill undoubtedly be revealed.
The Escherichia coli repressor of biotin biosyn-thesis, BirA, binds to a 40 bp operator site (bioO)to repress transcriptional initiation at the twodivergent promoters of the biotin biosyntheticoperon (Figure 1A; Otsuka & Abelson, 1978). Therepressor is an allosteric DNA binding protein andutilizes a derivative of biotin, biotinyl-50-AMP, asits corepressor (Prakash & Eisenberg, 1979). A highresolution structure of apoBirA has been obtainedusing x-ray crystallographic techniques and theDNA binding domain of the protein, as predictedfrom its amino acid sequence, has been shown tocontain a helix-turn-helix (HTH) motif (Wilsonet al., 1992). In addition to the HTH module, thedomain contains a third helix and a two-strandedantiparallel b-sheet (Figure 2). The overall fold of
# 1998 Academic Press Limited

Figure 1. A, Schematic representation of the Escherichia coli biotin operon with the transcriptional control regionexpanded. PA is the promoter for leftward transcription and PB is the promoter for rightward transcription. BioA-bioFcorrespond to the genes that encode the biotin biosynthetic enzymes. The boxed region represents the biotin operator.B, Detailed representation of the biotin operator sequence. The two half-sites have been arbitrarily designated 1 and2. The numbering scheme is taken from Otsuka & Abelson (1978). The underlining indicates the regions of inverteddyad symmetry in the sequence.
788 Map of Biotin Repressor-Biotin Operator Interface
the DNA binding domain of BirA, which has beenreferred to as the winged helix-turn-helix motif(wHTH), is similar to that found in several eukary-otic and prokaryotic site-speci®c DNA binding
Figure 2. A model of the three-dimensional structure ofapoBirA (Wilson et al., 1992). The helix-turn-helix andthe wing of the DNA binding domain are shown inblack. The Figure was generated using RASMOL (Sayle& Milner-White, 1995).
proteins. Among these are CAP (McKay & Steitz,1981) and LexA (Fogh et al., 1994) from E. coli, theyeast K. lactis heat shock factor (HSF; Harrisonet al., 1994) and hepatocyte nuclear factor (HNF-3g;Clark et al., 1993). No structure of the complex ofholoBirA bound to the biotin operator DNA is yetavailable.
Complexes of members of the wHTH class ofsite-speci®c DNA binding proteins bound to targetsites have been studied by a variety of techniques.Results of biochemical studies indicate that theseproteins can bind to target sites as monomers (Nyeet al., 1992), dimers (Huang & Igo, 1996), trimers(Sorger & Nelson, 1989), or even hexamers (Tianet al., 1992). Monomers of wHTH proteins havebeen reported to form interfaces with DNA targetsites characterized by a broad range of lengths. Forexample, while the heat shock factor (HSF) mono-mer binds to a 5 bp target (Perisic et al., 1989), amonomer of the fork head class of proteins bindsto sites ranging from 13 to 18 bp (Clark et al., 1993;Kaufmann et al., 1994), and each OmpR monomerbinds to a 9 bp site (Huang & Igo, 1996). Oligo-meric wHTH proteins have been shown to bind toeither inverted or tandem repeats of their respect-ive monomeric target sites (Xiao et al., 1991; Perisicet al., 1989; Wertman & Mount, 1985). High resol-ution structures of three complexes of wHTH pro-teins bound to DNA have been determined. Theseinclude E. coli CAP, HNF-3g and the PU.1 ETS-domain. In the CAP dimer-DNA cocrystal the pro-tein DNA interface formed by each monomer

Map of Biotin Repressor-Biotin Operator Interface 789
extends over 14 bp (Schultz et al., 1991; Parkinsonet al., 1996). The monomeric HNF-3g, which wascrystallized with a 13 bp target, appears to form aninterface along the entire length of that oligomer(Clark et al., 1993). The PU.1 ETS protein, which isalso monomeric, forms an interface with 10 bp ofDNA (Kodandapani et al., 1996). Although thethree proteins are by no means identical in struc-ture, there are similarities apparent in the macro-molecular interfaces of the complexes. First, asobserved in structures of the HTH class of site-speci®c DNA binding proteins, a number of thecontacts formed between the protein and DNAbases in these wHTH protein-DNA complexesinvolve insertion of the second or ``recognition''alpha helix of the HTH motif into a major grooveof the recognition sequence. Second, in all threestructures additional contacts are observedbetween one of the wings (W1) of the wHTH motifand phosphates or bases of the minor groove adja-cent to the core major groove of the target site.Finally, amino acids at the N termini a-helices 1and 2 of the DNA binding domain of all of theseproteins interact with backbone phosphates.
The 40 bp biotin operator sequence is character-ized by hyphenated inverted symmetry (Figure 1B).We have arbitrarily divided the sequence into two20 bp half-sites with the designations half-site 1and 2. Limited studies of the effects of alterationsin the bioO sequence on in vivo and/or in vitrofunction have been performed. For example, wheneither of the symmetrically placed G �C base-pairsat positions ÿ15 and �15 is changed to an A �Tbase-pair an operator constitutive phenotype in vivoand decreased af®nity for holoBirA in vitro areobserved (Otsuka & Abelson, 1978; Barker et al.,1981). In addition, alterations of the amino acidsequence in the recognition helix of the wHTHmotif of the DNA binding domain of BirA resultsin a loss of repression at the biotin control region(Barker & Campbell, 1981; Buoncristiani et al.,1986). Like CAP, holoBirA has been shown to bindto its target site as a dimer (Lin et al.,1991; Abbott& Beckett, 1993). As discussed above, recognitionsites of dimeric wHTH repressors are signi®cantlysmaller than bioO and range in size from the 18 bptarget of OmpR (Huang & Igo, 1996) to the 28 bpsite required for site-speci®c DNA binding of CAP(Liu-Johnson et al., 1986). The size of the BirA tar-get site suggests that the structural details of DNArecognition by this protein may differ from thoseobserved in other wHTH protein-DNA complexes.Results of DNase I footprinting studies of onestrand of the biotin operator indicate that as manyas 44 bases of DNA sequence are altered in theirsusceptibility to hydrolysis in the holoBirA-bioOcomplex. The pattern of digestion is, moreover,complex in that both protection from and enhance-ment of cleavage of the DNA are observed in thecomplex relative to free DNA. This enhancementof hydrolysis may indicate distortion of theDNA concomitant with repressor binding (Drew& Travers, 1984).
In order to develop a low resolution model ofthe structure of the holoBirA-bioO complex wehave applied a combination of solution techniquesincluding DNase I footprinting, methylation pro-tection and hydroxyl radical footprinting. Resultsof these studies indicate that the holoBirA dimercontacts the biotin operator DNA in four regions.Each repressor monomer thus forms a bipartiteinterface with the DNA, one involving only majorgroove contacts and the other involving minorgroove interactions. The majority of the contactsformed between the protein and DNA involve twoperfectly conserved 12 bp regions at the termini ofthe operator site. Results of gel mobility shiftassays indicate that binding of the protein to DNAis accompanied by a global distortion of the DNAfrom linearity. Results of DNase I footprinting indi-cate that one locus of structural change in theDNA is the 16 bp operator core that links the two12 bp termini. These combined results suggest astructural model for the complex in which the pro-tein serves to bridge the terminal segments of adistorted operator site. Although the results indi-cate that the details of the interaction of eachwHTH motif of the holoBirA dimer with DNAmay be similar to those observed in high resolutionstructures of other wHTH-DNA complexes, thepresence of a lengthy A � T-rich linker sequencethat undergoes a structural change concomitantwith repressor binding represents a novel depar-ture from observations in related systems.
Results
DNase I footprinting
The DNase I footprints of the holoBirA-bioOcomplex are shown in Figure 3. Footprints of boththe upper and lower strands of the operatorsequence were obtained. As indicated in Figure 3Athe footprint of the upper strand covers approxi-mately 39 bp. No change in the cleavage pattern ofthe operator is observed in the 4 bp at the 50 end ofthe operator sequence of this strand. The footprintof the lower strand is larger and extends over44 bp, four base-pairs into the 50 ¯anking region ofthe operator sequence. The DNase I footprints ofboth strands are complex as indicated by both pro-tection from and enhancement of hydrolysis of thephosphodiester backbone. The complexity is, how-ever, more extensive in the lower strand. Whileincreased reactivity of only one cleavage site, pos-ition ÿ11, in the upper strand of the the operatorsequence is observed, multiple sites of enhancedcleavage are observed in the lower strand of thebioO sequence (Figure 3B). These include theTAAC sequence immediately 30 to the central Atract as well as positions �11, �20 in the operatorsequence and �24 in the 50 ¯anking sequence ofthe operator site. DNase I, because of its size, is oflimited use in obtaining detailed information aboutthe structural features of a protein-DNA interface.In order to expand the footprinting studies to a
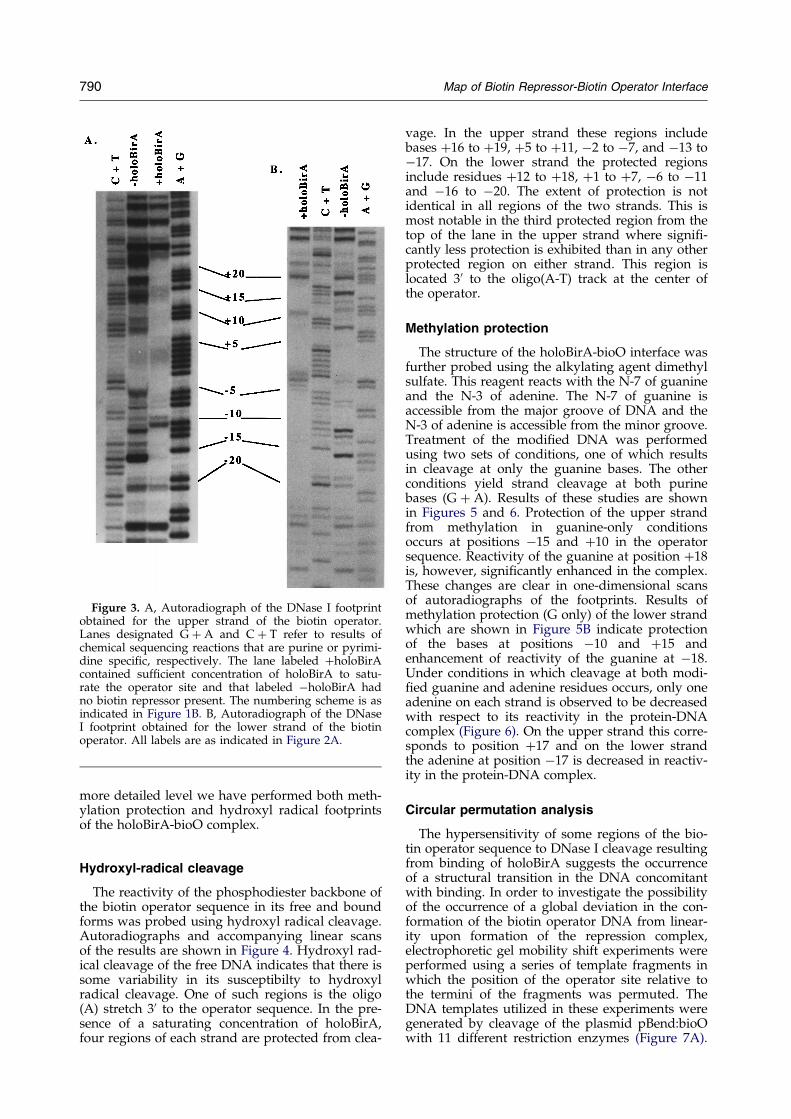
Figure 3. A, Autoradiograph of the DNase I footprintobtained for the upper strand of the biotin operator.Lanes designated G � A and C � T refer to results ofchemical sequencing reactions that are purine or pyrimi-dine speci®c, respectively. The lane labeled �holoBirAcontained suf®cient concentration of holoBirA to satu-rate the operator site and that labeled ÿholoBirA hadno biotin repressor present. The numbering scheme is asindicated in Figure 1B. B, Autoradiograph of the DNaseI footprint obtained for the lower strand of the biotinoperator. All labels are as indicated in Figure 2A.
790 Map of Biotin Repressor-Biotin Operator Interface
more detailed level we have performed both meth-ylation protection and hydroxyl radical footprintsof the holoBirA-bioO complex.
Hydroxyl-radical cleavage
The reactivity of the phosphodiester backbone ofthe biotin operator sequence in its free and boundforms was probed using hydroxyl radical cleavage.Autoradiographs and accompanying linear scansof the results are shown in Figure 4. Hydroxyl rad-ical cleavage of the free DNA indicates that there issome variability in its susceptibilty to hydroxylradical cleavage. One of such regions is the oligo(A) stretch 30 to the operator sequence. In the pre-sence of a saturating concentration of holoBirA,four regions of each strand are protected from clea-
vage. In the upper strand these regions includebases �16 to �19, �5 to �11, ÿ2 to ÿ7, and ÿ13 toÿ17. On the lower strand the protected regionsinclude residues �12 to �18, �1 to �7, ÿ6 to ÿ11and ÿ16 to ÿ20. The extent of protection is notidentical in all regions of the two strands. This ismost notable in the third protected region from thetop of the lane in the upper strand where signi®-cantly less protection is exhibited than in any otherprotected region on either strand. This region islocated 30 to the oligo(A-T) track at the center ofthe operator.
Methylation protection
The structure of the holoBirA-bioO interface wasfurther probed using the alkylating agent dimethylsulfate. This reagent reacts with the N-7 of guanineand the N-3 of adenine. The N-7 of guanine isaccessible from the major groove of DNA and theN-3 of adenine is accessible from the minor groove.Treatment of the modi®ed DNA was performedusing two sets of conditions, one of which resultsin cleavage at only the guanine bases. The otherconditions yield strand cleavage at both purinebases (G � A). Results of these studies are shownin Figures 5 and 6. Protection of the upper strandfrom methylation in guanine-only conditionsoccurs at positions ÿ15 and �10 in the operatorsequence. Reactivity of the guanine at position �18is, however, signi®cantly enhanced in the complex.These changes are clear in one-dimensional scansof autoradiographs of the footprints. Results ofmethylation protection (G only) of the lower strandwhich are shown in Figure 5B indicate protectionof the bases at positions ÿ10 and �15 andenhancement of reactivity of the guanine at ÿ18.Under conditions in which cleavage at both modi-®ed guanine and adenine residues occurs, only oneadenine on each strand is observed to be decreasedwith respect to its reactivity in the protein-DNAcomplex (Figure 6). On the upper strand this corre-sponds to position �17 and on the lower strandthe adenine at position ÿ17 is decreased in reactiv-ity in the protein-DNA complex.
Circular permutation analysis
The hypersensitivity of some regions of the bio-tin operator sequence to DNase I cleavage resultingfrom binding of holoBirA suggests the occurrenceof a structural transition in the DNA concomitantwith binding. In order to investigate the possibilityof the occurrence of a global deviation in the con-formation of the biotin operator DNA from linear-ity upon formation of the repression complex,electrophoretic gel mobility shift experiments wereperformed using a series of template fragments inwhich the position of the operator site relative tothe termini of the fragments was permuted. TheDNA templates utilized in these experiments weregenerated by cleavage of the plasmid pBend:bioOwith 11 different restriction enzymes (Figure 7A).

Figure 4. Hydroxyl radical footprints of (A) upper and (B) lower strand of the biotin operator. The line scans of auto-radiographs are shown to the right of each autoradiograph. The signi®cance of the lane labels is identical with thatindicated in Figure 2 and the numbering scheme is as indicated in Figure 1B.
Map of Biotin Repressor-Biotin Operator Interface 791
Results of electrophoresis of reaction mixtures con-taining holoBirA bound to each of the bioO tem-plate fragments are shown. The concentrations ofholoBirA and operator fragment present in eachmixture were designed to obtain approximately30% binding of the DNA. As indicated inFigure 7A, the mobility of the complex is inverselyproportional to the the distance of the operator sitefrom the termini of the template fragment. Com-plexes with the operator site positioned in the cen-ter of the fragments exhibited the lowest mobility.This is shown graphically in Figure 7B. The mobili-ties of the uncomplexed or free fragments show nosystematic dependence of mobility on the distanceof the site from the termini of the fragment(Figure 7C.) However, the characteristic pattern ofrelative mobilities of the unliganded DNA frag-ments indicated in the Figure were highly reprodu-cible. An estimate of the deviation of the DNAfrom linearity in its complex with holoBirA wasmade using the formalism of Thompson & Landy(1988). Results of this analysis performed asdescribed in Materials and Methods indicate thatin its complex with holoBirA the bioO DNA devi-ates from linearity by approximately 40 degrees.
Discussion
The winged helix-turn-helix DNA binding pro-tein, BirA, binds to a 40 bp target site, bioO. Gen-
etic studies of the biotin operator function arelimited and only point mutations at G �C base-pairs at positions �15 and ÿ15 have been shownto result in constitutive transcription initiation atthe biotin operon promoters and decreased af®nityfor holoBirA (Otsuka & Abelson, 1978; Barker et al.,1981). This work constitutes the ®rst detailed bio-chemical mapping of the BirA-bioO complex. Ingeneral dimeric wHTH proteins bind to target sitesthat range in size from 20 to 30 bp. Since the biotinoperator, at 40 bp, is signi®cantly longer, it is con-ceivable that formation of the repression complexinvolves binding of an oligomeric species of holo-BirA larger than a dimer. We have, however,previously shown, using direct stoichiometrictitrations, that two holoBirA monomers bind to theoperator site (Abbott & Beckett, 1993). An implicitassumption in interpretation of the titration data isthat the protein is 100% active in DNA binding.Although this is dif®cult to unequivoically estab-lish, we have demonstrated that BirA preparationsobtained in our laboratory are fully active in bothsynthesizing and binding to the allosteric effector,bio-50-AMP. Lin et al. (1991), using a completelyindependent technique to determine the holoBirA:bioO stoichiometry in which the amount of proteinand DNA present in an electrophoretic band corre-sponding to the complex was quanti®ed, alsoobtained a 2:1 stoichiometry for the repressioncomplex. These combined results thus rule out the
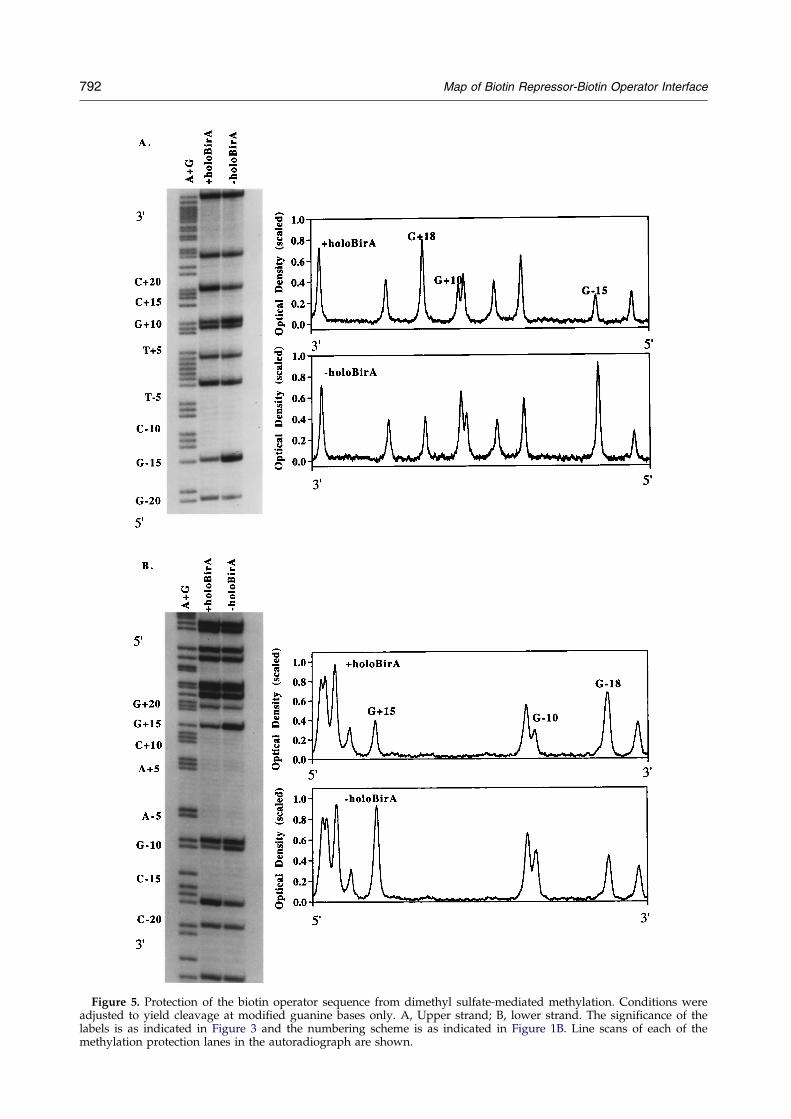
Figure 5. Protection of the biotin operator sequence from dimethyl sulfate-mediated methylation. Conditions wereadjusted to yield cleavage at modi®ed guanine bases only. A, Upper strand; B, lower strand. The signi®cance of thelabels is as indicated in Figure 3 and the numbering scheme is as indicated in Figure 1B. Line scans of each of themethylation protection lanes in the autoradiograph are shown.
792 Map of Biotin Repressor-Biotin Operator Interface
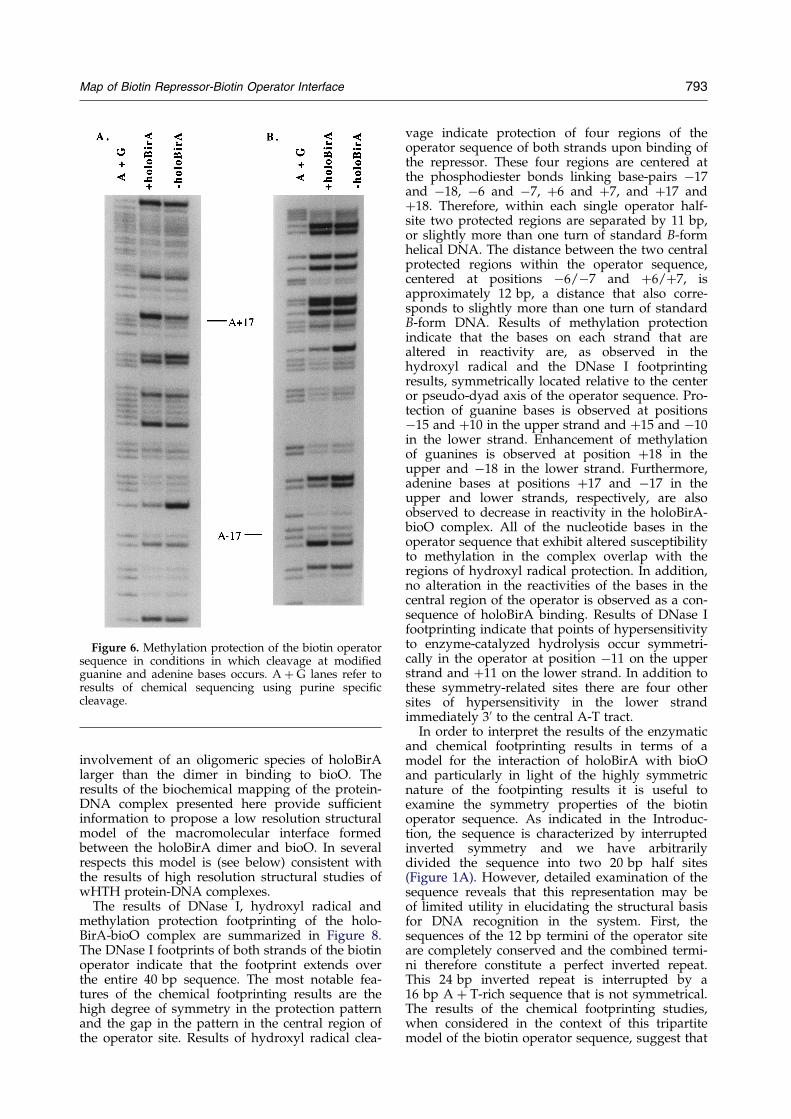
Figure 6. Methylation protection of the biotin operatorsequence in conditions in which cleavage at modi®edguanine and adenine bases occurs. A � G lanes refer toresults of chemical sequencing using purine speci®ccleavage.
Map of Biotin Repressor-Biotin Operator Interface 793
involvement of an oligomeric species of holoBirAlarger than the dimer in binding to bioO. Theresults of the biochemical mapping of the protein-DNA complex presented here provide suf®cientinformation to propose a low resolution structuralmodel of the macromolecular interface formedbetween the holoBirA dimer and bioO. In severalrespects this model is (see below) consistent withthe results of high resolution structural studies ofwHTH protein-DNA complexes.
The results of DNase I, hydroxyl radical andmethylation protection footprinting of the holo-BirA-bioO complex are summarized in Figure 8.The DNase I footprints of both strands of the biotinoperator indicate that the footprint extends overthe entire 40 bp sequence. The most notable fea-tures of the chemical footprinting results are thehigh degree of symmetry in the protection patternand the gap in the pattern in the central region ofthe operator site. Results of hydroxyl radical clea-
vage indicate protection of four regions of theoperator sequence of both strands upon binding ofthe repressor. These four regions are centered atthe phosphodiester bonds linking base-pairs ÿ17and ÿ18, ÿ6 and ÿ7, �6 and �7, and �17 and�18. Therefore, within each single operator half-site two protected regions are separated by 11 bp,or slightly more than one turn of standard B-formhelical DNA. The distance between the two centralprotected regions within the operator sequence,centered at positions ÿ6/ÿ7 and �6/�7, isapproximately 12 bp, a distance that also corre-sponds to slightly more than one turn of standardB-form DNA. Results of methylation protectionindicate that the bases on each strand that arealtered in reactivity are, as observed in thehydroxyl radical and the DNase I footprintingresults, symmetrically located relative to the centeror pseudo-dyad axis of the operator sequence. Pro-tection of guanine bases is observed at positionsÿ15 and �10 in the upper strand and �15 and ÿ10in the lower strand. Enhancement of methylationof guanines is observed at position �18 in theupper and ÿ18 in the lower strand. Furthermore,adenine bases at positions �17 and ÿ17 in theupper and lower strands, respectively, are alsoobserved to decrease in reactivity in the holoBirA-bioO complex. All of the nucleotide bases in theoperator sequence that exhibit altered susceptibilityto methylation in the complex overlap with theregions of hydroxyl radical protection. In addition,no alteration in the reactivities of the bases in thecentral region of the operator is observed as a con-sequence of holoBirA binding. Results of DNase Ifootprinting indicate that points of hypersensitivityto enzyme-catalyzed hydrolysis occur symmetri-cally in the operator at position ÿ11 on the upperstrand and �11 on the lower strand. In addition tothese symmetry-related sites there are four othersites of hypersensitivity in the lower strandimmediately 30 to the central A-T tract.
In order to interpret the results of the enzymaticand chemical footprinting results in terms of amodel for the interaction of holoBirA with bioOand particularly in light of the highly symmetricnature of the footpinting results it is useful toexamine the symmetry properties of the biotinoperator sequence. As indicated in the Introduc-tion, the sequence is characterized by interruptedinverted symmetry and we have arbitrarilydivided the sequence into two 20 bp half sites(Figure 1A). However, detailed examination of thesequence reveals that this representation may beof limited utility in elucidating the structural basisfor DNA recognition in the system. First, thesequences of the 12 bp termini of the operator siteare completely conserved and the combined termi-ni therefore constitute a perfect inverted repeat.This 24 bp inverted repeat is interrupted by a16 bp A � T-rich sequence that is not symmetrical.The results of the chemical footprinting studies,when considered in the context of this tripartitemodel of the biotin operator sequence, suggest that
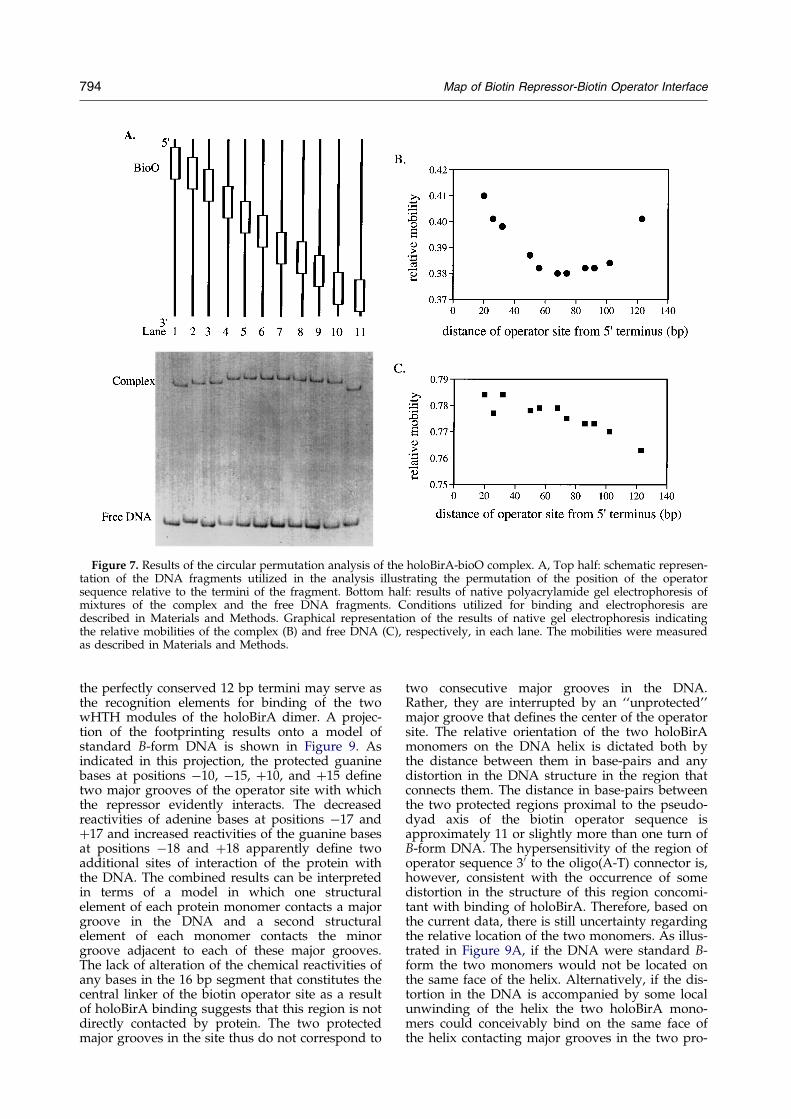
Figure 7. Results of the circular permutation analysis of the holoBirA-bioO complex. A, Top half: schematic represen-tation of the DNA fragments utilized in the analysis illustrating the permutation of the position of the operatorsequence relative to the termini of the fragment. Bottom half: results of native polyacrylamide gel electrophoresis ofmixtures of the complex and the free DNA fragments. Conditions utilized for binding and electrophoresis aredescribed in Materials and Methods. Graphical representation of the results of native gel electrophoresis indicatingthe relative mobilities of the complex (B) and free DNA (C), respectively, in each lane. The mobilities were measuredas described in Materials and Methods.
794 Map of Biotin Repressor-Biotin Operator Interface
the perfectly conserved 12 bp termini may serve asthe recognition elements for binding of the twowHTH modules of the holoBirA dimer. A projec-tion of the footprinting results onto a model ofstandard B-form DNA is shown in Figure 9. Asindicated in this projection, the protected guaninebases at positions ÿ10, ÿ15, �10, and �15 de®netwo major grooves of the operator site with whichthe repressor evidently interacts. The decreasedreactivities of adenine bases at positions ÿ17 and�17 and increased reactivities of the guanine basesat positions ÿ18 and �18 apparently de®ne twoadditional sites of interaction of the protein withthe DNA. The combined results can be interpretedin terms of a model in which one structuralelement of each protein monomer contacts a majorgroove in the DNA and a second structuralelement of each monomer contacts the minorgroove adjacent to each of these major grooves.The lack of alteration of the chemical reactivities ofany bases in the 16 bp segment that constitutes thecentral linker of the biotin operator site as a resultof holoBirA binding suggests that this region is notdirectly contacted by protein. The two protectedmajor grooves in the site thus do not correspond to
two consecutive major grooves in the DNA.Rather, they are interrupted by an ``unprotected''major groove that de®nes the center of the operatorsite. The relative orientation of the two holoBirAmonomers on the DNA helix is dictated both bythe distance between them in base-pairs and anydistortion in the DNA structure in the region thatconnects them. The distance in base-pairs betweenthe two protected regions proximal to the pseudo-dyad axis of the biotin operator sequence isapproximately 11 or slightly more than one turn ofB-form DNA. The hypersensitivity of the region ofoperator sequence 30 to the oligo(A-T) connector is,however, consistent with the occurrence of somedistortion in the structure of this region concomi-tant with binding of holoBirA. Therefore, based onthe current data, there is still uncertainty regardingthe relative location of the two monomers. As illus-trated in Figure 9A, if the DNA were standard B-form the two monomers would not be located onthe same face of the helix. Alternatively, if the dis-tortion in the DNA is accompanied by some localunwinding of the helix the two holoBirA mono-mers could conceivably bind on the same face ofthe helix contacting major grooves in the two pro-

Figure 8. Summary of the results of enzymatic and chemical footprinting of the holoBirA-bioO complex. The dia-monds represent sites of protection from hydroxyl radical-mediated cleavage. The ®lled circles represent sites of pro-tection from (small circles) or enhancement of (large circles) dimethyl sulfate modi®cation at guanine bases. The opensquares represent sites of protection from dimethyl sulfate modi®cation at adenine residues. The arrows representpoints of DNase I hypersensitivity in the complex relative to the free DNA.
Figure 9. Projection of the chemical protection results shown in Figure 8 onto a standard B-form helical DNA model(10 bp/helical turn). The Figure was generated using RASMOL (Sayle & Milner-White, 1995). Atoms in black indicateprotection from hydroxyl radical-mediated modi®cation (C40 and C50 positions are highlighted) and alteration indimethyl sulfate reactivity in the complex. The view in A, illustrates the methylation protection of the N-7 at G �10,G ÿ10, A �17, A ÿ17, G �18 and G ÿ18. The view in B, represents a 180� rotation around the helical axis of themodel in A and illustrates methylation protection at G �15 and G ÿ15.
Map of Biotin Repressor-Biotin Operator Interface 795

796 Map of Biotin Repressor-Biotin Operator Interface
tected regions proximal to the center of the oper-ator.
A three-dimensional structure of apoBirA isavailable the model of which is shown in Figure 2and it is tempting to utilize this structural infor-mation to construct a model of the protein-DNAcomplex. However, since binding of the allostericeffector, bio-50-AMP, results in structural changesin the protein at both tertiary and quaternarylevels, major limitations are placed on using theknown structure of apoBirA in proposing a modelfor the holoBirA-bioO complex. At the tertiarystructural level, binding of the effector induces asigni®cant conformational change involving, inpart, the formation of an interface between the N-terminal and central domains of the protein (Xuet al., 1995; Xu & Beckett, 1996). Furthermore,results of assembly studies of the protein indicatethat while the aporepressor crystallizes as a mono-mer, the holorepressor exhibits weak dimerization(KD � 30 micromolar; D. Beckett & E. Eisenstein,unpublished results). Two holoBirA monomershave been demonstrated to bind cooperatively tobioO and we hypothesize that the two monomersphysically interact in the protein-DNA complex(Abbott & Beckett, 1993). Although the crystalstructure provides no information about thelocation of the monomer-monomer interface on theprotein surface, genetic and biochemical studies oftwo BirA mutants, D197Y and �147A, provideinformation to speculate on its location. These twomutants are defective in repression of transcriptionfrom the biotin regulatory region. Results ofmeasurements of the af®nities of the puri®ed pro-teins for bioO indicate that they are indeed defec-tive in site-speci®c binding to the operator site(E.D.S. et al., unpublished observations). As indi-cated in Figure 2, the mutations are located on asurface of the central domain of the protein andare far removed from the catalytic site. Althoughmeasurements of the dimerization properties ofthese two mutants have yet to be completed, thesimplest interpretation of the biochemical results isthat the source of their decreased af®nity for bioOre¯ects defects in dimerization. Based on theassumption that these two mutants do indeedde®ne, in part, the location of the monomer-mono-mer interface in the DNA-bound protein, the orien-tations of the two wHTH modules with respect tothe pseudo-dyad axis of the biotin operator in theprotein-DNA complex can be proposed. In the pro-posed complex the HTH motifs of the two mono-mers would be proximal to the dyad axis. As suchthey are responsible for the protection of the sym-metry-related G ÿ10, G �15 and G �10 and G ÿ15,from methylation. These protected regions, there-fore, presumably re¯ect the interaction of the rec-ognition helices of two HTH motifs with majorgrooves of the DNA. The two additional regions ofaltered reactivity to dimethylsulfate de®ned bybases A ÿ17 and G ÿ18 and A �17 and G �18presumably re¯ect the two segments of the DNA
that contact the ``wings'' of the two wHTH mod-ules of the holoBirA.
The possibility of the occurence of bending ofthe biotin operator DNA upon formation of theholoBirA-bioO complex has been investigatedusing the circular permutation analysis technique(Wu & Crothers, 1984). Results of these measure-ments are consistent with the occurrence of limitedglobal distortion in the DNA upon protein binding.Estimates of the bending angle, however, indicatethat the deviation of the DNA in the complex fromlinearity is only on the order of 40 degrees. Thisestimate is, of course, based only on the results ofthe circular permutation assay and veri®cation ofbending as well as geometry of the bend can beobtained from application of phasing analysis(Zinkel & Crothers, 1987; Salvo & Grindley, 1987).The results of the circular permutation assay pro-vide information at a global level and do not indi-cate local points within the operator sequence atwhich deviations from standard B-form structuremay occur. Nevertheless, the existence of threeDNase I hypersensitive sites within the biotin oper-ator upon formation of the protein-DNA complexsuggests that deviations of the operator sequenceDNA from standard B-form are induced at thesethree loci. Within each of the two 12 bp segmentsthat serve as a recognition element for a wHTHmodule there is local distortion of the DNAde®ned by the observed hypersensitivities at pos-itions ÿ11 and �11 of the operator site. This distor-tion within the putative recognition site for eachwHTH motif is consistent with structural infor-mation obtained for the CAP-DNA complex(Schultz et al., 1991; Parkinson et al., 1996). Inaddition, the hypersensitivity observed in theregion adjacent to the central A-T tract suggeststhat there is some distortion of the DNA in the16 bp segment that links the recognition elementsfor the two wHTH motifs.
In the model proposed for the holoBirA-bioOcomplex we have suggested that two structuralelements of each holoBirA monomer are involvedin the interface of the protein-DNA complex. Asindicated in the Introduction, the DNA bindingdomain of BirA belongs to the class of site-speci®cDNA binding proteins referred to as winged helix-turn-helix proteins. The results of the footprintingstudies on the holoBirA-bioO complex presentedhere provide evidence that, analogous to otherwHTH proteins, each biotin repressor monomerforms a bipartite interface with the DNA in theoperator sequence. This interface extends overapproximately 12 bp, a length similar to thatformed by monomers of all wHTH that have thusfar been studied using high resolution structuraltechniques. As observed in these other structures,the recognition helix of the HTH motif of eachBirA monomer is most likely contacting the DNAvia the major groove that is de®ned by the methyl-ation protection observed at guanine bases ÿ10,ÿ15, �10 and �15. The altered methylationobserved at positions �17, �18, ÿ17 and ÿ18 pre-

Map of Biotin Repressor-Biotin Operator Interface 797
sumably results from the interaction of the wingwith the adjacent minor grooves of the DNA at thetwo termini of the operator site. The lack of appar-ent interaction between holoBirA and the largecentral linker region of the biotin operatorsequence represents a signi®cant departure of thebiotin repressor system from wHTH proteins thathave been previously characterized. As indicatedabove, the enhanced DNase I-mediated cleavagethat occurs in this region concomitant with holo-BirA binding is consistent with the occurrence ofdistortion of the DNA structure in this region. It islikely that this distortion is required for formationof the two interfaces between the two wHTH mod-ules and the two bioO termini. Thus, although theprotein does not appear to contact the centralregion of the operator site its sequence and/orlength undoubtedly contributes to the stability ofthe holoBirA-bioO interaction.
Materials and Methods
Enzymes and chemicals
Restriction enzymes were obtained from Promega orNew England Biolabs. The Klenow fragment, T4 DNAligase, and calf intestinal alkaline phosphatase were fromPromega. T4 polynucleotide kinase was from New Eng-land Biolabs. The [g-32P-]ATP and [a-32P]dATP anddGTP used in radiolabeling of DNA were purchasedfrom Amersham. Dimethyl sulfate, EDTA, ferrousammonium sulfate, and sodium ascorbate, all 99 � %,were from Aldrich. The d-biotin, 50-AMP, and ATP werefrom Sigma. All other chemicals used in the preparationof buffers were reagent or analytical grade. BirA waspuri®ed as previously described (Abbott & Beckett,1993) and biotinyl-50-AMP was synthesized in this lab-oratory and puri®ed as described by Abbott & Beckett(1993).
Plasmid construction
The plasmid pBioZ used in footprinting experimentswas constructed by insertion of a 210 bp EcoRI biotinoperator fragment from pAOC3 (Junuichi & Otsuka,personal communication) into the EcoRI site of theplasmid pZ150 (Abbott & Beckett, 1993). The plasmidpBend:bioO was constructed by inserting a double-stranded 50 bp bioO fragment with XbaI sticky ends intothe XbaI site of the plasmid pBend 2 (Kim et al., 1989).The biotin operator fragment, bioO, was assembled fromfour oligonucleotides. Two of the oligonucleotides werephosphorylated using T4 polynucleotide kinase and thenligated to the two remaining oligos using T4 DNA ligase.The bioO fragment was then inserted into XbaI-digestedpBend 2. The resulting plasmid, pBend:bioO, was usedto transform the strain JM109 made competent by theCaCl2 procedure. Finally, plasmids were prepared usingthe method of Birnboim & Doly (1979) and subjected to®nal puri®cation by CsCl density gradient centrifugation(Maniatis et al., 1982).
Preparation of DNA for footprinting
All DNA fragments for radiolabeling were obtainedby digesting the plasmid pBioZ with HindIII. End-
labeled fragments used for footprinting of the upperstrand (see Results) were produced by ®lling in therecessed 30 end with [a-32P]dATP and dGTP using Kle-now fragment (Brenowitz et al., 1986). DNA intended foruse in footprinting of the lower strand was depho-sphorylated using calf intestinal alkaline phosphataseand labeled at the 50 end using T4 polynucleotide kinaseand [g-32P]ATP as described by Titus (1991). All sub-sequent manipulations were identical for both strands.Labeled fragments were puri®ed over a NACS Prepaccolumn (Life Technologies) and then digested with PstI.Restriction fragments were separated by electrophoresison a 1% (w/v) agarose gel, visualized by autoradiog-raphy, and the desired fragment was recovered by elec-troelution as described by Maniatis et al. (1982). TheDNA was then puri®ed over a NACS Prepac columnand concentrated by precipitation. The DNA was resus-pended to a concentration of 420,000 cpm/ml in TE andstored at 4�C. Labeled DNA intended for Maxam-Gilbertsequencing, hydroxyl radical and methylation protectionfootprinting was exchanged into water immediatelyprior to use.
Preparation of DNA fragments for circularpermutation analysis
Single enzyme digests were made of pBend:bioOusing MluI, BglII, NheI, SpeI, XhoI, EcoRV, PvuII, StuI,NruI, SspI, and BamHI to cut 20 mg of plasmid each. Thereactions were subjected to phenol/chloroform extrac-tions to remove proteins and the DNA was concentratedby ethanol-precipitation. The DNA was ®nally resus-pended in TE at a concentration of 1 mg/ml. Since SspIdigestion generates three restriction fragments, one ofwhich is very close in size to the bioO-containing frag-ment, further puri®cation was necessary. Digestion pro-ducts were separated on a native 5% polyacrylamide gel,the bioO-containing fragment was excised from the gel,and electroeluted out of the gel piece (Maniatis et al.,1982). Recovered DNA was puri®ed over a NACS Pre-pac column (Life Technologies), concentrated by ethanol-precipitation and resuspended in 10 mM Tris-HCl(pH 8.2), 0.1 mM EDTA at a ®nal concentration ofapproximately 0.8 mg/ml.
DNase I footprinting
DNase I footprints were performed as previouslydescribed by Abbott & Beckett (1993). The reaction buf-fer used contained 10 mM Tris (pH 7.50 at 20�C), 50 mMKCl, 2.5 mM MgCl2, 1 mM CaCl2, 2 mg/ml sonicatedcalf thymus DNA, and 100 mg/ml BSA. Biotin and ATPwere also present at ®nal concentrations of 50 mM and250 mM, respectively. BirA is known to be saturated withbio-50-AMP at these concentrations of the two substrates(Xu & Beckett, 1994). Each 200 ml reaction mixture con-tained 12,000 cpm of labeled DNA and 4 � 10ÿ7 M holo-BirA. DNA and protein were allowed to equilibrate at20�C for one hour. DNase I digestion of the reaction wasinitiated by the addition of 5 ml of 0.2 mg/ml DNase Iprepared in binding buffer minus calf thymus DNA andBSA. The digestion was allowed to proceed for two min-utes at 20�C, quenched by addition of 33 ml of 50 mMEDTA and the DNA was precipitated by the addition of700 ml of a solution containing 0.4 M NH4OAc and0.1 mg/ml tRNA in ethanol. The pellets were washedtwice with cold 80% (v/v) ethanol in water, dried, resus-pended in 7 ml formamide gel-loading buffer, and the

798 Map of Biotin Repressor-Biotin Operator Interface
products were separated by electrophoresis on adenaturing 10% polyacrylamide gel.
Hydroxyl radical footprinting
Hydroxyl radical footprints were performed accord-ing to the method described by Strahs & Brenowitz(1994) with the following changes. The reaction bufferused was identical with that used for DNase I foot-printing. Each 35 ml reaction contained 12,000 cpm 32P-labeled DNA and 4 � 10ÿ7 M holoBirA. The reactionswere incubated for one hour at 20�C to allow them toreach equilibrium. Three solutions: 0.1 mM Fe(NH4)2-(SO4)2, 0.2 mM EDTA; 10 mM ascorbic acid; and 0.3%H2O2 were prepared and incubated at 20�C for 30 min-utes. Hydroxyl radical cleavage was initiated by add-ing 5 ml of each of the three radical-generating reagentsas one drop on the inside wall of the tube and mixingit with the reaction (Tullius & Dombroski, 1986). Reac-tions were incubated for two minutes at 20�C,quenched and precipitated with 193 ml of 0.63 MNH4OAc, 0.5 ml/ml tRNA in ethanol. Reprecipitatedand dried reaction products were resuspended in 6.5 mlof formamide gel-loading buffer and separated by elec-trophoresis on a denaturing 10% polyacrylamide gel.
Methylation protection footprinting
Methylation protection footprints were performed asdescribed by Wissman & Hillen (1991) with the follow-ing changes. The buffer used for these experiments con-tained 10 mM sodium cacodylate (pH 7.0 at 20�C),200 mM KCl, 2.5 mM MgCl2, 1 mM CaCl2, 2 mg/mlsonicated calf thymus DNA, and 100 mg/ml BSA. Bioti-nyl-50-AMP was added to a ®nal concentration of 50 mMjust prior to use. Each 200 ml reaction mixture, whetherstrong guanine or guanine � adenine, contained12,000 cpm 32P-labeled DNA and 4 � 10ÿ7 M holoBirA.DNA and protein were allowed to equilibrate at 20�C forone hour. Methylation attack was initiated by theaddition of 2.5 ml of 10% dimethyl sulfate in ethanol. Thereaction was incubated for two minutes at 20�C,quenched by addition of 50 ml of 1.5 M NaOAc, 1 Mb-mercaptoethanol, 100 mg/ml tRNA, and precipitatedwith 750 ml of ethanol. Two subsequent precipitationswere performed using 250 ml of 0.3 M NaOAc, 0.1 mMEDTA and 750 ml of ethanol. The guanine � adeninereaction required one further step. The lyophilizedDNA was resuspended in 20 ml of water and chilledon ice before the addition of 5 ml of 0.5 M HCl. Thereaction was gently mixed periodically during a onehour incubation on ice. Two precipitations identicalwith those described above prepared the DNA fortreatment with piperidine. DNA strand scission wasachieved by adding 100 ml of 1 M piperidine and heat-ing at 90�C for 30 minutes (Maxam and Gilbert,1980). The reactions were dried under vacuum andtwice resuspended in 10 ml of water and lyophilized.The dried reaction products were resuspended in6.5 ml of formamide gel-loading buffer and resolvedby electrophoresis on a denaturing 10% polyacryl-amide gel.
Circular permutation analysis
Bending experiments were performed following amodi®cation of the methods described by Kim et al.(1989). The buffer used in these reactions contained
10 mM Tris (pH 7.5 at 20�C), 50 mM KCl, 2.5 mMMgCl2, 1 mM CaCl2, 3% (v/v) glycerol, and 0.1 mg/mlBSA. The electrophoresis buffer used contained 6.7 mMTris (pH 7.5 at 20�C), 1 mM EDTA, and 3.3 mM NaOAc.Biotin and ATP were added to a ®nal concentrations of50 mM and 250 mM, respectively. Each 30 ml reaction wascomposed of 3 mg (5 � 10ÿ8 M) of the appropriateenzyme-digested pBend:bioO and BirA at a ®nal concen-tration of 3.35 � 10ÿ8 M. Incubation was carried out at20�C for 90 minutes to allow the reactions to reach equi-librium. During incubation, a 10% (19:1 acrylamide tobis-acrylamide, w/w) native polyacrylamide gel pre-pared in electrophoresis buffer was pre-electrophoresedat 100 V for one hour with recirculation of the electro-phoresis buffer provided by a peristaltic pump. The elec-trophoresis voltage was increased to 300 V, reactionswere loaded directly onto the gel, and the voltage wasdecreased to 100 V for the duration of the 11 hour run.The gel was stained with ethidium bromide, destained inelectrophoresis buffer, and photographed using PolaroidType 55 ®lm with a red ®lter. The negative was devel-oped with 18% sodium sul®te and wet with Photo¯o 200(Kodak) before drying.
Quantification and data analysis
Sequencing reactions, performed as described byMaxam & Gilbert (1980), were run alongside the foot-printing reaction products in order to provide orien-tation to the footprinting lanes. Gels were dried andexposed to pre-¯ashed X-ray ®lm (Kodak XAR-5) for36 to 38 hours. Densitometric analyses of autoradio-grams and quanti®cation of gel retardation assays wereperformed using a Molecular Dynamics personal laserdensitometer as described by Abbott & Beckett (1993).Retardation of gel mobility was quanti®ed by measure-ment of the y-coordinates corresponding to the protein-bound and free DNA fragments on the scanned ®lmnegatives. The DNA bending angle was estimatedusing the equation mM/mE� cos(a/2), where mM and mE
are the mobilities of DNA fragments in which the bio-tin operator sequence is positioned at the center or endof the molecule, respectively, and a is the magnitudeof the deviation of the DNA from linearity (Thompson& Landy, 1988).
Acknowledgments
This work was supported by NIH grant ROI-GM46511. The authors thank Dr S. Adhya for providingthe plasmid pBend 2.
References
Abbott, J. & Beckett, D. (1993). Cooperative binding ofthe Escherichia coli repressor of biotin biosynthesisto the biotin operator. Biochemistry, 32, 9649±9656.
Barker, D. F. & Campbell, A. M. (1981). Genetic and bio-chemical characterization of the birA gene and itsproduct: Evidence for a direct role of biotin holoen-zyme synthetase in repression of the biotin operonin Escherichia coli. J. Mol. Biol. 146, 469±492.
Barker, D. F., Kuhn, J. & Campbell, A. M. (1981).Sequence and properties of operator mutations inthe bio operon of Escherichia coli. Gene, 13, 89±102.

Map of Biotin Repressor-Biotin Operator Interface 799
Birnboim, H. C. & Doly, J. (1979). A rapid alkalineextraction procedure for screening recombinantplasmid DNA. Nucl. Acids Res. 7, 1513±1523.
Brennan, R. G. (1992). DNA recognition by the helix-turn-helix motif. Curr. Opin. Struct. Biol. 2, 100±108.
Brenowitz, M., Senear, D. F., Shea, M. A. & Ackers, G. K.(1986). Quantitative DNase footprinting titrations: amethod for studying DNA-protein interactions.Methods Enzymol. 130, 132±181.
Buoncristiani, M. R., Howard, P. K. & Otsuka, A. J.(1986). DNA-binding and enzymatic domains ofthe bifunctional biotin operon repressor (BirA) ofEscherichia coli. Gene, 44, 255±261.
Clark, K. L., Hurley, E. D., Lai, E. & Burley, S. K. (1993).Co-crystal structure of the HNF-3/fork head DNA-recognition motif resembles histone H5. Nature, 364,412±419.
Drew, H. R. & Travers, A. R. (1984). DNA structuralvariations in the E. coli tyrT promoter. Cell, 37, 491±502.
Fogh, R. H., Ottleben, G., Ruterjans, H., Schnarr, M.,Boelens, R. & Kaptein, R. (1994). Solution structureof the LexA repressor DNA binding domain deter-mined by 1H NMR spectroscopy. EMBO J. 13,3936±3944.
Harrison, C. J., Bohm, A. A. & Nelson, H. C. M. (1994).Crystal structure of the DNA binding domain ofthe heat shock transcription factor. Nature, 263,224±227.
Huang, K.-J. & Igo, M. M. (1996). Identi®cation of thebases in the ompF regulatory region, which interactwith the transcription factor OmpR. J. Mol. Biol.262, 615±628.
Jordan, S. R. & Pabo, C. O. (1988). Structure of thelambda complex at 2.5 AÊ resolution: details of therepressor-operator interactions. Science, 242, 893±899.
Kaufmann, E., Hoch, M. & Jackle, H. (1994). The Inter-action of DNA with the DNA-binding domainencoded by the Drosophila gene fork head. Eur. J. Bio-chem. 223, 329±337.
Kim, J., Zwieb, C., Wu, C. & Adhya, S. (1989). Bendingof DNA by gene-regulatory proteins: constructionand use of a DNA bending vector. Gene, 85, 15±23.
Kodandapani, R., Pio, F., Ni, C.-Z., Piccialli, G., Klemsz,M., McKercher, S., Maki, R. A. & Ely, K. R. (1996).A new pattern for helix-turn-helix recognitionrevealed by the PU. 1 ETS-domain-DNA complex.Nature, 380, 456±459.
Lin, K.-C., Campbell, A. & Shiuan, D. (1991). Bindingcharacteristics of Escherichia coli biotin repressor-operator complex. Biochim. Biophys. Acta, 1090, 317±325.
Liu-Johnson, H.-N., Gartenberg, M. R. & Crothers, D. M.(1986). The DNA binding domain and bendingangle of E. coli CAP protein. Cell, 47, 995±1005.
Maniatis, T., Fritsch, E. F. & Sambrook, J. (1982). Molecu-lar Cloning: A Laboratory Manual, Cold Spring Har-bor Laboratory Press, Cold Spring Harbor, NY.
Maxam, A. M. & Gilbert, W. (1980). Sequencing end-labeled DNA with base-speci®c chemical cleavages.Methods Enzymol. 65, 499±560.
McKay, D. B. & Steitz, T. A. (1981). Structure of catabo-lite gene activator protein at 2.9 AÊ resolutionsuggests binding to left-handed B-DNA. Nature,290, 744±749.
Nye, J. A., Petersen, J. M., Gunther, C. V., Jonsen,M. D. & Graves, B. J. (1992). Interaction of murineEts-1 with GGA-binding sites establishes the ETS
domain as a new DNA-binding motif. Genes Dev. 6,975±990.
Otsuka, A. J. & Abelson, J. (1978). The regulatory regionof the biotin operon in Escherichia coli. Nature, 276,689±693.
Parkinson, G., Wilson, C., Gunasekera, A., Ebright,Y. W., Ebright, R. E. & Berman, H. M. (1996).Structure of the CAP-DNA complex at 2 5 AÊ resol-ution: A complete picture of the protein-DNAinterface. J. Mol. Biol. 260, 395±408.
Perisic, O., Xiao, H. & Lis, J. T. (1989). Stable binding ofDrosophila heat shock factor to head-to-head andtail-to-tail repeats of a conserved 5 bp recognitionunit. Cell, 59, 797±806.
Prakash, O. & Eisenberg, M. A. (1979). Biotinyl-50-aden-ylate: corepressor role in the regulation of the biotingenes of Escherichia coli K-12. Proc. Natl Acad. Sci.USA, 76, 5592±5595.
Raumann, B. E., Brown, B. M. & Sauer, R. T. (1994).Major groove tecognition by b-sheets: the ribbon-helix-helix family of gene regulatory proteins. Curr.Opin. Struct. Biol. 4, 36±43.
Salvo, J. J. & Grindley, N. D. F. (1987). Helical phasingbetween DNA bends and the determination of benddirection. Nucl. Acids Res, 15, 9771±9779.
Sayle, R. A. & Milner-White, E. J. (1995). RASMOL: bio-molecular graphics for all. Trends Biochem. Sci. 20,pp. 374±376.
Schultz, S. C., Shields, G. C. & Steitz, T. A. (1991).Crystal structure of a CAP-DNA complex:The DNAis bent by 90�. Science, 253, 1001±1007.
Schumacher, M. A., Choi, K. Y., Zalkin, H. & Brennan,R. G. (1994). Crystal structure of LacI member,PurR, bound to DNA: minor groove binding by ahelices. Science, 266, 763±770.
Sorger, P. K. & Nelson, H. C. M. (1989). Trimerizationof a yeast transcriptional activator via a coiled-colimotif. Cell, 59, 855±864.
Strahs, D. & Brenowitz, M. (1994). DNA conformationalchanges associated with the cooperative binding ofcI repressor of bacteriophage l to OR. J. Mol. Biol.244, 494±510.
Thompson, J. F. & Landy, A. (1988). Empirical esti-mation of protein-induced DNA bending angles:applications to lambda site-speci®c recombinationcomplexes. Nucl. Acids Res. 16, 9687±9705.
Tian, G.-L., Lim, D.-B., Carey, J. & Maas, W. K. (1992).Binding of the arginine repressor of Escherichi coliK12 to its operator sites. J. Mol. Biol. 226, 387±397.
Titus, D. E., ed. (1991). Promega Protocols and ApplicationsGuide, 2nd edit., Promega Corporation, Madison,WI.
Tullius, T. & Dombroski, B. (1986). Hydroxyl radical``footprinting'': high resolution information aboutDNA-protein contacts and application to l repres-sor and Cro protein. Proc. Natl Acad. Sci. USA, 83,5469±5473.
Wertman, K. F. & Mount, D. W. (1985). Nucleotidebinding speci®city of the LexA repressor of Escheri-chia coli K-12. J. Bacteriol. 163, 376±384.
Wilson, K. S., Shewchuk, L. M., Brennan, R. G., Otsuka,A. J. & Matthews, B. W. (1992). Escherichia coli bio-tin holoenzyme synthetase/bio repressor crystalstructure delineates biotin- and DNA-bindingdomains. Proc. Natl Acad. Sci. 89, 9257±9261.
Wissman, A. & Hillen, W. (1991). DNA contacts probedby modi®cation protection and interference studies.Methods Enzymol. 208, 365±379.

800 Map of Biotin Repressor-Biotin Operator Interface
Wu, H.-M. & Crothers, D. M. (1984). The locus ofsequence-directed and protein-induced DNAbending. Nature, 308, 509±513.
Xiao, H., Perisic, O. & Lis, J. T. (1991). Cooperative bind-ing of Drosophila heat shock factor to arrays of aconserved 5 bp unit. Cell, 64, 585±593.
Xu, Y. & Beckett, D. (1994). Kinetics of biotinyl-50-aden-ylate synthesis catalyzed by the Escherichia colirepressor of biotin biosynthesis and the stability ofthe enzyme-product complex. Biochemistry, 33,7354±7360.
Xu, Y. & Beckett, D. (1996). Evidence for interdomaininteraction in the Escherichia coli repressor of biotinbiosynthesis from studies of an N-terminal domaindeletion mutant. Biochemistry, 35, 1783±1792.
Xu, Y., Nenortas, E. & Beckett, D. (1995). Evidence fordistinct ligand-bound conformational states of themultifunctional Escherichia coli repressor of biotinbiosynthesis. Biochemistry, 34, 16624±16631.
Zinkel, S. S. & Crothers, D. M. (1987). DNA bend direc-tion by phase sensitive detection. Nature, 328, 178±181.
Edited by R. Ebright
(Received 11 August 1997; received in revised form 13 February 1998; accepted 13 February 1998)