helix lutescens rossmässler, 1837 (gastropoda - folia
TRANSCRIPT

HELIX LUTESCENS ROSSMÄSSLER, 1837(GASTROPODA: PULMONATA: HELICIDAE)– ITS STRUCTURE, BIOLOGY AND ECOLOGY
EL¯BIETA KORALEWSKA-BATURA
Department of General Zoology, Institute of Environmental Sciences,Adam Mickiewicz University, Fredry 10, 61-701 Poznañ, Poland
ABSTRACT: Helix lutescens Rossmässler, 1837 is a xerothermophilous species. It reaches its NW distribution bor-der in SE Poland. The studies, carried out in 1990–1997, involved the distribution of H. lutescens in Poland, itsbiology and ecology. Besides, shell structure and internal organs were studied, with special reference to differ-ences between H. lutescens and the related H. pomatia L. In H. lutescens the shell is roundish-conical, of a yellow-ish-white colour and much smaller than that of H. pomatia. The body is greyish and covered with numerouswrinkles and grooves; fine, whitish granules are located in the grooves, especially in those that form two deli-cate light streaks along the darker back of the animal; these streaks are characteristic of the species. The repro-ductive system of H. lutescens is of a structure similar to that in H. pomatia, but the duct of the gametolythicgland never bears a diverticle while flagellum, epiphallus and penis in adults are pigmented. The diurnal activ-ity of H. lutescens varies seasonally, depending on environmental factors (air temperature and relative humid-ity in ground layer, and substratum humidity – dew point). The reproductive activity reaches its peak in Mayand June. Courtship and copulation are in accordance with the typical helicid pattern. The copulation lasts ca.15 mins, and the entire mating process takes over 3 hrs. Eggs are laid in nests dug in the soil, the mean numberof eggs per nest being 35. In two weeks young hatch and remain in the nest for ca. 16 days. The abundanceand density of three age classes in a selected population of H. lutescens have been estimated on a permanentsampling plot, using marking-release-recapture method, with JOLLY-SEBER‘S model for an open population.The results made it possible to trace seasonal changes in the abundance within the whole population. In Po-land H. lutescens, because of its insular occurrence and the threat resulting from confusion with the edible H.pomatia, is a protected species.
KEY WORDS: Helix lutescens, Helicidae, land snails, distribution, reproduction, biology, ecology, population dy-namics

CONTENTS
Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 199Study area . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 200Material and methods . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 201Results . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 204
A. Morphology and anatomy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2041. Shell . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2042. External appearance . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2063. Musculature and body covers . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2074. Alimentary tract . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2075. Pallial complex . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2116. Excretory system . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2127. Circulatory system . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2138. Respiratory system . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2149. Nervous system and sense organs . . . . . . . . . . . . . . . . . . . . . . . . . 21410. Reproductive system. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 215
B. Distribution of H. lutescens in Poland . . . . . . . . . . . . . . . . . . . . . . . 227C. Biology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 228
1. Diurnal and seasonal activity of adults . . . . . . . . . . . . . . . . . . . . . 2282. Courtship and copulation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2313. Egg-laying and hatching. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 233
D. Ecology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2361. Abundance and density . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2362. Population abundance changes . . . . . . . . . . . . . . . . . . . . . . . . . . 236
Concluding remarks. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 237References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 238
198 Koralewska-Batura E.

INTRODUCTION
Contrary to the common Helix pomatia Linnaeus,1758, H. lutescens Rossmässler, 1837 was rarely an ob-ject of malacologists’ interest. Till now it is among theleast studied species in the fauna of Poland. Informa-tion on its occurrence in the country is contained infairly numerous publications (e.g. POLIÑSKI 1917,1919, 1927, GROCHMALICKI 1932, CZUBIÑSKI &URBAÑSKI 1933, URBAÑSKI 1937, 1947, 1948, 1957,1964, 1973, 1977, RIEDEL 1954, 1988, BARGA-WIÊC£AWSKA 1989, 1997, PIECHOCKI 1990, 1991,KORALEWSKA-BATURA 1993c), but its morphologyand anatomy have been only poorly studied. Basic in-formation on the species can be found in generalpublications dealing with European malacofauna,e.g. EHRMANN (1933), LIKHAREV & RAMMELMEIER
(1952), GROSSU (1955, 1983), LOZEK (1956, 1964),KERNEY et al. (1983), LISICKÝ (1991), and in some pa-pers dealing with the gastropod fauna of Poland(B¥KOWSKI & £OMNICKI 1892, URBAÑSKI 1957).ROSSMÄSSLER (1837), based on few specimens, de-scribed the shell, radula and reproductive system, thelatter in very general terms. POLIÑSKI (1924) dis-cussed, in a cursory way, the structure of its circulatorysystem, and SHILEYKO (1978) gave a shell descriptionand photograph. Recently, KORALEWSKA-BATURA
(1993a, b, 1994a, 1997a) has studied shell biometricsin H. lutescens, its love dart, postembryonic develop-ment of the reproductive system and lung structure;the author has analysed the structure of radula, onthe background of other helicid species (KORA-
Helix lutescens 199
Fig. 1. Distribution of H. lutescens in Europe

LEWSKA-BATURA 1994b). A sinistral specimen of thisspecies has also been described (KORALEWSKA-BATURA 1997b).
H. lutescens is a xero-thermophilous species. It in-habits localities of steppe character, dry shrubs, grass-lands, sunny slopes, especially on calcareous substra-tum. It is very often found also in gardens and ceme-teries where it lives among ruderal vegetation ofsunny places, close to fences or walls of old buildings.
Zoogeographically, H. lutescens is a Dak-Podolianspecies (Fig. 1). POLIÑSKI (1919) reports that it“reaches from its Podolian motherland to the north-ern fringes of the Lublin and Ma³opolska uplands”.According to URBAÑSKI (1948), the dry and ratherwarm Subboreal period affected favourably the dis-persal of south-eastern, often xero-thermophilous,species. The presence of H. lutescens in south-easternPoland would testify to its being a postglacialpseudorelict. Its distribution range is divided in twoparts by the Carpathian range: a south-western part,including Spisz and Slovakian Karst, northern andwestern Hungary and the plains of innerTransylvania, and a north-eastern part comprisingMoldavia, Besarabia, Podolia and southern uplands ofPoland (RIEDEL 1988). According to the latter authorH. lutescens is probably a receding species in Poland.
The very scanty knowledge of H. lutescens has in-duced me to attempt a more detailed and more exten-sive study, combined with summarizing the existingliterature data with a view of producing a monographof this species. My own studies included faunistic fieldsurvey of areas where the snail had been observed orits occurrence seemed likely, observations on the ac-tivity of adult individuals, their reproduction andpopulation studies, as well as anatomical-histologicalstudies in which special attention was paid to the re-productive and alimentary systems. The structure ofthe remaining systems, which mostly corresponds tothat found in other members of the genus Helix (seebelow), even if not analysed in detail and based partlyon literature data, was discussed for the sake ofcompletness of this monograph. Attention was fo-cused on the shell morphology and structure; mor-phology and anatomy of soft parts, includinghistological structure of the reproductive system;up-dating information on the distribution in Poland;diurnal and seasonal activity; reproductive biology; es-timation of abundance and density of particular ageclasses on a constant study plot; seasonal changes ofpopulation density.
STUDY AREA
The faunistic survey included the whole distribu-tion area of H. lutescens in Poland – south eastern partof the country i.e. uplands Wy¿yna Ma³opolska andWy¿yna Lubelska, lowland Nizina Sandomierska andthe foothills of the Beskid Wschodni mountains.
Behavioural and population studies were carriedout on a study plot located in the north-eastern part ofthe town of Kielce next to Maria Konopnicka street, inthe district of Szyd³ówek (Fig. 2). The habitat was aphytocoenosis of grassland character. The dominantgrasses were: Arrhenatherum elatius (L.) P. B., Poaangustifolia L., Agropyron repens (L.) P. B. and Dactylisglomerata L. The plant community resembled atall-herb community because of the abundant pres-ence of Tanacetum vulgare L., on the other hand, it alsoresembled thermophilous xerothermic grassland ofthe class Festuca-Brometea, because of the presence ofsuch species as Poa compressa L., Euphorbia cyparissias L.,Plantago media L. and Salvia verticillata L. The vegeta-tion was probably formed on an abandoned field, as in-dicated by the presence of weed the class Stellarietea me-diae, representing a relict of agrophytocoenosis. Like-wise, plants of the class Artemisietea, with Artemisiavulgaris L., Cirsium arvense (L.) Scop. and Solidagoserotina Ait. indicated a synantropic nature of thesyntaxon. The grassland character of the vegetation re-
sulted from a dry, warm, calcium-rich substratum ofpH 7.58. Chemical analysis of the soil revealed highquantities of calcium (4.64 mg/g), which favourscalciphilous plants. As early as May they form a highand dense cover. The absence of trees results in a lackof shadow, which causes large fluctuations of tempera-ture and air humidity (Figs 3, 4). The soils in Kielce areclassified as rendzina formed of slowly eroding Devo-nian crystalline limestones and dolomites. They areskeletal soils, of a high content of CaCO3 or CaCO3 ×MgCO3. The study plot is covered by anthropogenicsoil, deeply dug and mixed, with an unformed profile.
In the regional division of climatic areas of Poland(OKO£OWICZ & MARTYN 1979) the area is located inthe Œwiêtokrzyski climatic district, which is a part ofthe Silesian-Ma³opolska climatic region. Data on se-lected climatic elements of Kielce during the studiesin 1993–1996 were obtained from the IMiGW weatherstation in Su³ków near Kielce. The mean annual tem-perature is 9.1°C. In the summer, mild effects of con-tinental climate prevail (mean temperature of July18.7°C), and mild winters (mean temperature of Jan-uary -2.3°C) testify to the effect of western oceanic cli-mate. The mean annual precipitation in Kielce is 625mm, with the maximum in August (91.6 mm) and theminimum in February (23.6 mm). Snowfall takes
200 Koralewska-Batura E.

place from November till April, the mean being 68days per year. Westerly, south-westerly and southerlywinds prevail, and to a lesser extent south-easterly.The location of the city, in a wide valley opened to
south-east and north-west, results in relatively fewcalm periods.
MATERIAL AND METHODS
Field and laboratory studies were carried out in1990–1997. During the first three years I surveyed
the current distribution of H. lutescens in Poland, spe-
cial attention being paid to its typical habitats, such
Helix lutescens 201
Fig. 2. A fragment of map of the town of Kielce; with study plot indicated

as xerothermic grasslands, shrubs, copses, cemeter-ies and gardens. The aim was both finding new local-ities of the species and confirming its earlier records.The field studies involved the vegetation season,from the second half of April till the end of September.
In 1994, the field work involved estimation ofabundance of a selected population of H. lutescens,and in 1995 – observations of diurnal and seasonal ac-tivity of adult individuals in the site in Kielce, de-scribed above. Earlier, two separate study plots hadbeen established, which were subject to detailed ob-servations. The first plot was 500 m2 in area. For prac-tical reasons, I divided it in 20 squares, 5×5 m (25 m2)each. The squares made it possible to locate all theobserved snails more precisely. The other study plot,of 25 m2, was divided into 25 numbered squares, 1×1m each. The plots and their component squares weremarked with wooden poles or plastic plates stuck inthe corners of squares.
The abundance of snails was studied from half ofMay till half of September 1994, at monthly intervals,on the larger of the two plots (500 m2). The studiesconsisted in catching, marking, release and recaptureof H. lutescens. During catches, I searched the squaresthoroughly, inspecting the places below plants andpartly uncovering the surface layer of soil.
During consecutive catches all the snails caughtwere marked and left where they had been found.The snails were marked on the suture between thebody and penultimate whorls, with a waterproof,quick-drying marker, each time of different colour.The marker had no negative effect on the snails.Since during subsequent observations the paint wasobserved to wear off, I corrected the marks duringsubsequent marking. In each consecutive catch Ifound snails marked during the previous catches andunmarked individuals, and both categories weremarked with a new colour. Based on the colour com-bination of individual snails, I could tell which hadbeen caught earlier, when and how many times. Outof the total of 1,815 H. lutescens caught during thestudies, 280 (15.4%) were caught and marked at leasttwice.
Based on measurements of shell diameter and thedegree of development of genitalia, three age classeswere distinguished:
D – adults, in which the shell growth had been ter-minated and lip was present, with fully developedgenitalia;
M – young individuals (2 years and older), of shelldiameter 10.1–21.0 mm, with incompletely developedgenitalia;
J – yearlings and younger (including those justleaving their nests), of shell diameter up to 10.0 mmand only slightly developed genitalia.
The abundance was estimated independently foreach age class, with JOLLY-SEBER‘S model for an open
population (JOLLY 1965, SEBER 1982, SUTHERLAND1996). Calculations were made with the programmeJOLLY–1989, considering the correction of theJOLLY-SEBER model (the so called JOLLY-SEBER Bmodel) with the variable individual viability and vari-able probability of catch within consecutive timeunits.
The population abundance (N) was calculatedwith the formula:
Ni = Mi (ni + 1)/(mi + 1)where:Mi – number of marked individuals present in
the population in the ith catch= mi + (Ri + 1)zi/(ri + 1);
mi – number of individuals marked during thepreceding catch and present in the sample;
Ri – number of individuals left in the placewhere they had been found;
zi – number of individuals caught before the ith
catch, not caught in the ith catch butcaught in any of the next catches;
ri – number of individuals caught and left inthe ith catch, which were caught in any ofthe next catches;
ni – total number of individuals caught in theith catch.
In 1995, at 2 week intervals, from May till half ofSeptember, I observed the activity of H. lutescens onthe 25 m2 plot. Only adult individuals were included.They were marked individually (waterproof marker),with numbers written on the body whorl. The mark-ing made it possible to observe activities of particularindividuals, and the division of the plot into squaresenabled me to locate them precisely. In May 1995, Icollected and marked 63 adult snails. The study plotwas not fenced in any way, so that the marked snailscould escape and return, while unmarked individualscould enter the plot at will. For this reason, duringconsecutive controls I numbered adult individualsthat were found for the first time, and included themin further observations.
The diurnal activity of H. lutescens was observedregularly, irrespective from weather conditions, dur-ing eight observation sessions: May 11/12th and25/26th, June 7/8th and 21/22nd, July 9/10th and25/26th, August 10/11th and 24/25th 1995. Each ob-servation session of 24 hrs started at 7.00 a.m., the ob-servations being repeated every two hours, the last ob-servation was at 5.00 next morning. During the obser-vations I determined precisely the position of eachmarked snail within the study plots (squares) and theplace where each individual stayed (e.g. on theground, on vegetation etc.). At the same time I notedthe state of activity of each snail (activity versus rest).The snails, which during the observations were crawl-ing, feeding, displaying activity associated with court-
202 Koralewska-Batura E.

Helix lutescens 203
Fig. 4. A fragment of the study plot in the summer
Fig. 3. A fragment of the study plot in the spring

ship, copulation or egg-laying, were regarded as ac-tive. During the eight observation sessions, I observeda total of 84 marked individuals. The marked snailswere the most numerous in the sample of May 11th1995 (63 individuals), the fewest (12 individuals) inthat of August 10th 1995. In ninth observation, onSeptember 13th 1995, I found no adult specimen(marked and unmarked) within the whole study area.
During the observations, I measured the air tem-perature in the ground layer and the relative humid-ity (hair hygrometer, WSZ Kraków T¯-7/48249) andnoted the weather.
The snail activity was also observed outside theconstant study plot, especially in the spring. These ob-servations were aimed mainly at tracing the breedingbiology of H. lutescens.
The analysis of diurnal activity of H. lutescens wasbased on mean values of observations at particularhours, repeated several times, depending on changesin the relative humidity and temperature of theground layer, in the spring and summer.
Live specimens of H. lutescens for morphological,anatomical and histological studies were caught dur-ing faunistic trips. The specimens destined for mor-phological and anatomical examination weredrowned in boiled water, cleaned of mucus and pre-served in ethyl alcohol, of a concentration graduallyincreasing from 30 to 75%. For histological studies, I
removed from the snails reproductive and alimentarysystems under stereomicroscope (magn. 40×) andfixed them in BOUIN liquid. Serial histological sec-tions of particular organs were stained with hema-toxylin and eosin. Some sections were stained withMAYER‘S method (detection of mucous substances),and VAN KOSS‘S method (calcium detection). Thestatining procedures applied are described in detailin ZAWISTOWSKI (1975). Photographs were taken withAGFA 15 DIN films under light microscope.
The structure of shell walls of H. lutescens, and of itsjaw, radula and love dart were examined mainly inscanning electron microscope Philips 515. Drawingswere made based on photos, supplied with details ob-served during dissection. When estimating the size ofshell, I considered such parameters as height, breadthand diameter. During measurements, especially ofthe diameter, I followed STÊPCZAK (1976) who first in-troduced this measurement.
All the basic statistics were calculated as describedin SOKAL & ROHLF (1995); the nomenclature and sig-nificance level of the characteristics and tests aregiven in the respective parts of the text. Calculationswere made with the programme Statistica for Win-dows. The programme JOLLY–1989 was made avail-able by CEPE/CNRS, Montpellier Cedex, France.
The illustrations in the text are original, if not indi-cated otherwise.
RESULTS
A. MORPHOLOGY AND ANATOMY
1. Shell
The shell of H. lutescens (Fig. 5) is sphaerical-coni-cal, of 4–4.5 poorly convex whorls separated by a shal-low suture. It is more conical than in H. pomatia. Theprotoconch (Fig. 8) has a wide apical part and quickincrement. The border between the protoconch andthe definitive whorls is fairly distinct. The surface ofthe protoconch at a low magnification is smooth;viewed at a higher magnification in SEM (Fig. 9) it iscoarse-grained, with numerous, irregularly arrangedgrooves. Definitive whorls increase quickly. The aper-ture is large, roundish, in most cases slightly higherthan wide. Its outer margin is usually developed as apoorly marked, whitish lip. The shell is, as a rule,unicolour, yellowish-white. The shell measurementsin adult specimens are within 28.5–34.1 mm height,26.9–32.1 mm breadth and 22.5–26.9 mm diameter.The shells are only slightly elongated, and the meanvalue of their height/breadth ratio is 1.06 whichmeans that the mean shell height is not much largerthan its mean breadth (KORALEWSKA-BATURA 1993a).
All the examined adult H. lutescens had their shellsmuch smaller than those of adult H. pomatia.
The internal structure of the shell of H. lutescens isshown in Figure 6. In juvenile individuals the umbili-cus is partly visible; in adults it is completely coveredwhile in H. pomatia it is usually partly open.
The macrosculpture of definitive whorls, ana-lysed in SEM, is rather poorly distinct (Fig. 10). Itconsists of radial growth stripes of various width, re-sulting from uneven growth of the shell, and of spi-ral lines. The distinct growth stripes combined withvery delicate spiral lines create an impression ofgrating. The macrosculpture is more delicate thanin H. pomatia.
On shell fractures, perpendicular to its growthstripes, analysed in SEM, four layers can be distin-guished, as reported by KILIAS (1960) for H. pomatia:periostracum, ectostracum, mesostracum andendostracum (Fig. 11). However, their relative thick-ness in these two species is different. The perio-stracum (Fig. 11a) is the thinnest layer, built ofconchiolin, which in adult snails wears off against thesubstratum and vegetation. The remaining, calcare-ous layers, also contain conchiolin, but inorganic
204 Koralewska-Batura E.
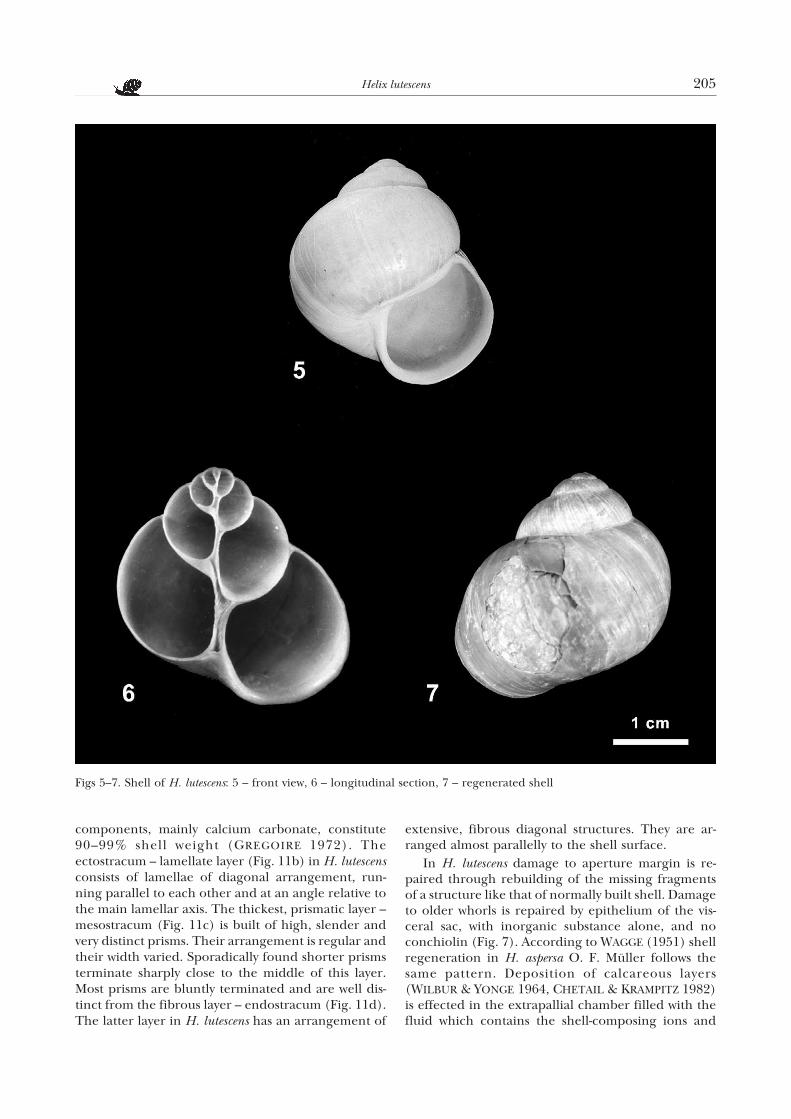
components, mainly calcium carbonate, constitute90–99% shell weight (GREGOIRE 1972). Theectostracum – lamellate layer (Fig. 11b) in H. lutescensconsists of lamellae of diagonal arrangement, run-ning parallel to each other and at an angle relative tothe main lamellar axis. The thickest, prismatic layer –mesostracum (Fig. 11c) is built of high, slender andvery distinct prisms. Their arrangement is regular andtheir width varied. Sporadically found shorter prismsterminate sharply close to the middle of this layer.Most prisms are bluntly terminated and are well dis-tinct from the fibrous layer – endostracum (Fig. 11d).The latter layer in H. lutescens has an arrangement of
extensive, fibrous diagonal structures. They are ar-ranged almost parallelly to the shell surface.
In H. lutescens damage to aperture margin is re-paired through rebuilding of the missing fragmentsof a structure like that of normally built shell. Damageto older whorls is repaired by epithelium of the vis-ceral sac, with inorganic substance alone, and noconchiolin (Fig. 7). According to WAGGE (1951) shellregeneration in H. aspersa O. F. Müller follows thesame pattern. Deposition of calcareous layers(WILBUR & YONGE 1964, CHETAIL & KRAMPITZ 1982)is effected in the extrapallial chamber filled with thefluid which contains the shell-composing ions and
Helix lutescens 205
Figs 5–7. Shell of H. lutescens: 5 – front view, 6 – longitudinal section, 7 – regenerated shell

compounds. They are provided to the extrapallialfluid through diffusion and actively transported byamebocytes. Calcium is taken up with food and accu-mulated in calcium cells of the hepatopancreas andconnective tissue. According to HELLER & MAGARITZ
(1983), oxygen, carbon and probably also calciumoriginate from water absorbed through the skin fromthe surface layer of the soil. Calcium transport is en-hanced by alkaline phosphatase, detected in mostmolluscs. The carbonate residue is derived from met-abolic carbonic acid, while deposition of crystalline
carbonates is enhanced by carbonic anhydrase(WILBUR & YONGE 1964). The ultimate deposition ofcalcium carbonate is extracellular.
2. External appearance
The external appearance of the body of H.
lutescens does not depart from that found in othermembers of the genus Helix. The body is greyish-coloured and covered by numerous wrinkles andgrooves. In the grooves there are fine, whitish granu-
206 Koralewska-Batura E.
Figs 8–11. Shell structure in H. lutescens: 8 – protoconch, 43×; 9 – sculpture of protoconch, 4,650×; 10 – sculpture of bodywhorl, 46.5×; 11 – shell fracture perpendicular to growth lines, a – periostracum, b – lamellate layer, c – prismatic layer, d– fibrous layer, 1,725×

lations. They are the most numerous in the groovesthat form two light streaks along the darker back ofthe snail (Fig. 12), which are characteristic of thespecies. The sole of the large, strongly muscled foot,is yellowish-grey. A large pneumostome is sur-rounded by two large lobes and a flat disc whosewhole free margin is rounded, while in H. pomatiathe margin is straight (JACKIEWICZ & KORA-LEWSKA-BATURA 1999).
3. Musculature and body covers
The structure of the main retractor muscles of H.lutescens is shown in Figure 13; it does not differ fromthat of H. pomatia. In the foot there are numerousstrands of longitudinally, transversely and dorso-ventrally arranged muscles.
The body wall of the snail is built of a monolayer epi-thelium, whose cells are provided with numerousciliae; the latter are the most numerous on the sole.Among them there are mucous glands. Around themouth, on sole edges and on the tentacles there arealso groups of sensory cells. The basal membrane ofthe epithelium is adjoined by a layer of connective tis-sue and strongly developed musculature of the bodywall.
4. Alimentary tract
The alimentary tract of H. lutescens (Fig. 14) corre-sponds to such tract in H. pomatia with respect to itsmorphological, anatomical and histological structure.
The mouth (Figs 15a, 16), in the shape of a trans-verse slit, is located on the underside of the anteriorpart of head. It is bordered by the upper and the
Helix lutescens 207
Fig. 12. A crawling H. lutescens
Fig. 13. Musculature of a snail: a – columella and columellarmuscle, b – foot retractors, c – pharynx retractors, d –ommatophore retractors, e – lower tentacle retractors, f– penial retractor, g – visceral sac, h – pedal gland, i –pharynx (after TRAPPMANN, from KILIAS 1960)
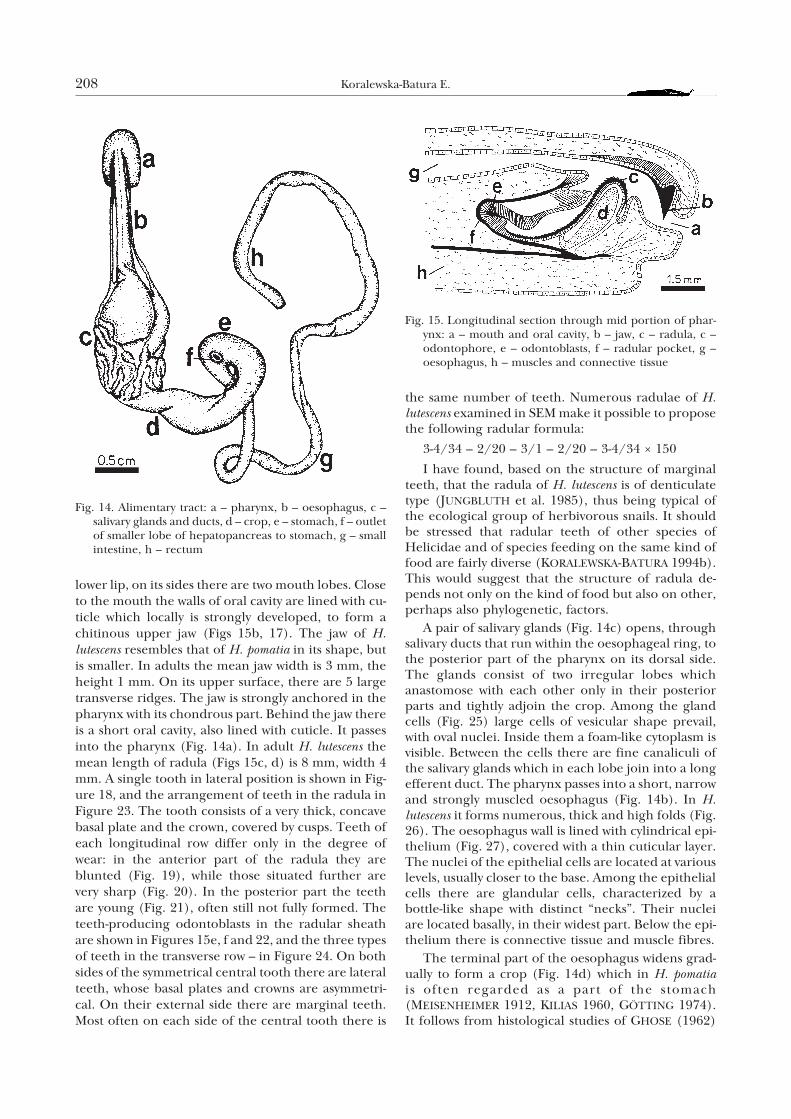
lower lip, on its sides there are two mouth lobes. Closeto the mouth the walls of oral cavity are lined with cu-ticle which locally is strongly developed, to form achitinous upper jaw (Figs 15b, 17). The jaw of H.lutescens resembles that of H. pomatia in its shape, butis smaller. In adults the mean jaw width is 3 mm, theheight 1 mm. On its upper surface, there are 5 largetransverse ridges. The jaw is strongly anchored in thepharynx with its chondrous part. Behind the jaw thereis a short oral cavity, also lined with cuticle. It passesinto the pharynx (Fig. 14a). In adult H. lutescens themean length of radula (Figs 15c, d) is 8 mm, width 4mm. A single tooth in lateral position is shown in Fig-ure 18, and the arrangement of teeth in the radula inFigure 23. The tooth consists of a very thick, concavebasal plate and the crown, covered by cusps. Teeth ofeach longitudinal row differ only in the degree ofwear: in the anterior part of the radula they areblunted (Fig. 19), while those situated further arevery sharp (Fig. 20). In the posterior part the teethare young (Fig. 21), often still not fully formed. Theteeth-producing odontoblasts in the radular sheathare shown in Figures 15e, f and 22, and the three typesof teeth in the transverse row – in Figure 24. On bothsides of the symmetrical central tooth there are lateralteeth, whose basal plates and crowns are asymmetri-cal. On their external side there are marginal teeth.Most often on each side of the central tooth there is
the same number of teeth. Numerous radulae of H.lutescens examined in SEM make it possible to proposethe following radular formula:
3-4/34 – 2/20 – 3/1 – 2/20 – 3-4/34 × 150
I have found, based on the structure of marginalteeth, that the radula of H. lutescens is of denticulatetype (JUNGBLUTH et al. 1985), thus being typical ofthe ecological group of herbivorous snails. It shouldbe stressed that radular teeth of other species ofHelicidae and of species feeding on the same kind offood are fairly diverse (KORALEWSKA-BATURA 1994b).This would suggest that the structure of radula de-pends not only on the kind of food but also on other,perhaps also phylogenetic, factors.
A pair of salivary glands (Fig. 14c) opens, throughsalivary ducts that run within the oesophageal ring, tothe posterior part of the pharynx on its dorsal side.The glands consist of two irregular lobes whichanastomose with each other only in their posteriorparts and tightly adjoin the crop. Among the glandcells (Fig. 25) large cells of vesicular shape prevail,with oval nuclei. Inside them a foam-like cytoplasm isvisible. Between the cells there are fine canaliculi ofthe salivary glands which in each lobe join into a longefferent duct. The pharynx passes into a short, narrowand strongly muscled oesophagus (Fig. 14b). In H.lutescens it forms numerous, thick and high folds (Fig.26). The oesophagus wall is lined with cylindrical epi-thelium (Fig. 27), covered with a thin cuticular layer.The nuclei of the epithelial cells are located at variouslevels, usually closer to the base. Among the epithelialcells there are glandular cells, characterized by abottle-like shape with distinct “necks”. Their nucleiare located basally, in their widest part. Below the epi-thelium there is connective tissue and muscle fibres.
The terminal part of the oesophagus widens grad-ually to form a crop (Fig. 14d) which in H. pomatiais often regarded as a part of the stomach(MEISENHEIMER 1912, KILIAS 1960, GÖTTING 1974).It follows from histological studies of GHOSE (1962)
208 Koralewska-Batura E.
Fig. 14. Alimentary tract: a – pharynx, b – oesophagus, c –salivary glands and ducts, d – crop, e – stomach, f – outletof smaller lobe of hepatopancreas to stomach, g – smallintestine, h – rectum
Fig. 15. Longitudinal section through mid portion of phar-ynx: a – mouth and oral cavity, b – jaw, c – radula, c –odontophore, e – odontoblasts, f – radular pocket, g –oesophagus, h – muscles and connective tissue
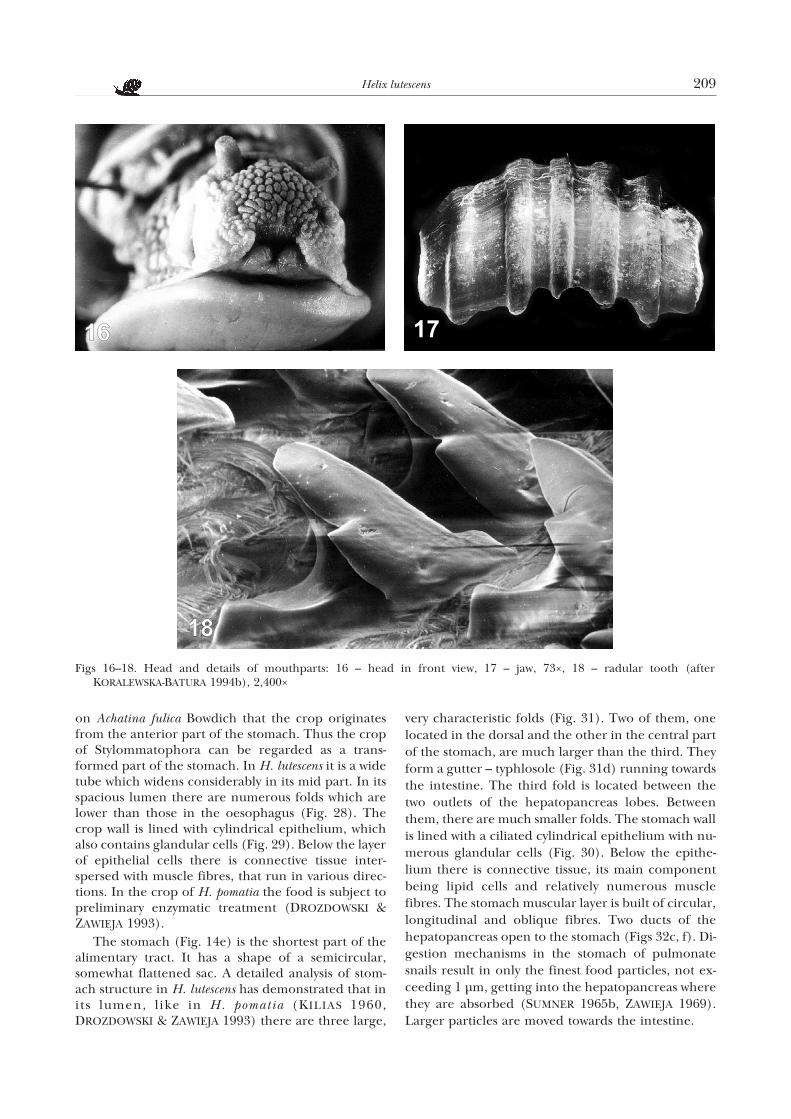
on Achatina fulica Bowdich that the crop originatesfrom the anterior part of the stomach. Thus the cropof Stylommatophora can be regarded as a trans-formed part of the stomach. In H. lutescens it is a widetube which widens considerably in its mid part. In itsspacious lumen there are numerous folds which arelower than those in the oesophagus (Fig. 28). Thecrop wall is lined with cylindrical epithelium, whichalso contains glandular cells (Fig. 29). Below the layerof epithelial cells there is connective tissue inter-spersed with muscle fibres, that run in various direc-tions. In the crop of H. pomatia the food is subject topreliminary enzymatic treatment (DROZDOWSKI &ZAWIEJA 1993).
The stomach (Fig. 14e) is the shortest part of thealimentary tract. It has a shape of a semicircular,somewhat flattened sac. A detailed analysis of stom-ach structure in H. lutescens has demonstrated that inits lumen, like in H. pomatia (KILIAS 1960,DROZDOWSKI & ZAWIEJA 1993) there are three large,
very characteristic folds (Fig. 31). Two of them, onelocated in the dorsal and the other in the central partof the stomach, are much larger than the third. Theyform a gutter – typhlosole (Fig. 31d) running towardsthe intestine. The third fold is located between thetwo outlets of the hepatopancreas lobes. Betweenthem, there are much smaller folds. The stomach wallis lined with a ciliated cylindrical epithelium with nu-merous glandular cells (Fig. 30). Below the epithe-lium there is connective tissue, its main componentbeing lipid cells and relatively numerous musclefibres. The stomach muscular layer is built of circular,longitudinal and oblique fibres. Two ducts of thehepatopancreas open to the stomach (Figs 32c, f). Di-gestion mechanisms in the stomach of pulmonatesnails result in only the finest food particles, not ex-ceeding 1 µm, getting into the hepatopancreas wherethey are absorbed (SUMNER 1965b, ZAWIEJA 1969).Larger particles are moved towards the intestine.
Helix lutescens 209
Figs 16–18. Head and details of mouthparts: 16 – head in front view, 17 – jaw, 73×, 18 – radular tooth (afterKORALEWSKA-BATURA 1994b), 2,400×
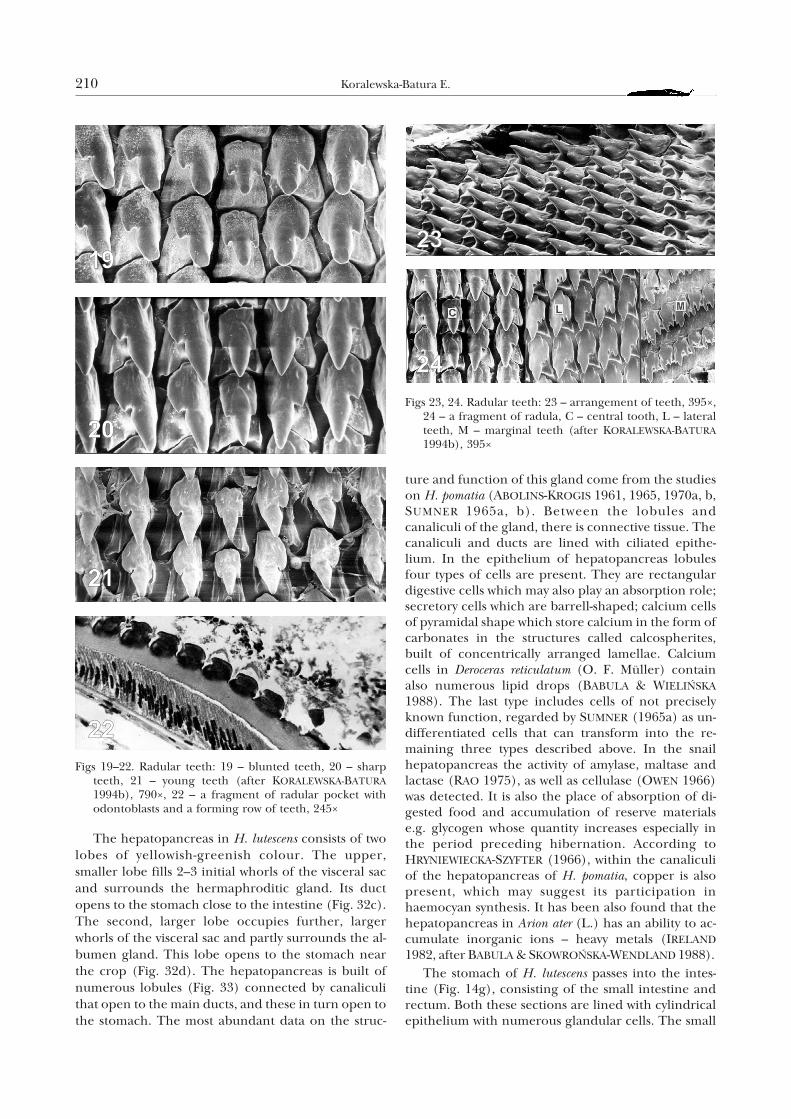
The hepatopancreas in H. lutescens consists of twolobes of yellowish-greenish colour. The upper,smaller lobe fills 2–3 initial whorls of the visceral sacand surrounds the hermaphroditic gland. Its ductopens to the stomach close to the intestine (Fig. 32c).The second, larger lobe occupies further, largerwhorls of the visceral sac and partly surrounds the al-bumen gland. This lobe opens to the stomach nearthe crop (Fig. 32d). The hepatopancreas is built ofnumerous lobules (Fig. 33) connected by canaliculithat open to the main ducts, and these in turn open tothe stomach. The most abundant data on the struc-
ture and function of this gland come from the studieson H. pomatia (ABOLINS-KROGIS 1961, 1965, 1970a, b,SUMNER 1965a, b). Between the lobules andcanaliculi of the gland, there is connective tissue. Thecanaliculi and ducts are lined with ciliated epithe-lium. In the epithelium of hepatopancreas lobulesfour types of cells are present. They are rectangulardigestive cells which may also play an absorption role;secretory cells which are barrell-shaped; calcium cellsof pyramidal shape which store calcium in the form ofcarbonates in the structures called calcospherites,built of concentrically arranged lamellae. Calciumcells in Deroceras reticulatum (O. F. Müller) containalso numerous lipid drops (BABULA & WIELIÑSKA
1988). The last type includes cells of not preciselyknown function, regarded by SUMNER (1965a) as un-differentiated cells that can transform into the re-maining three types described above. In the snailhepatopancreas the activity of amylase, maltase andlactase (RAO 1975), as well as cellulase (OWEN 1966)was detected. It is also the place of absorption of di-gested food and accumulation of reserve materialse.g. glycogen whose quantity increases especially inthe period preceding hibernation. According toHRYNIEWIECKA-SZYFTER (1966), within the canaliculiof the hepatopancreas of H. pomatia, copper is alsopresent, which may suggest its participation inhaemocyan synthesis. It has been also found that thehepatopancreas in Arion ater (L.) has an ability to ac-cumulate inorganic ions – heavy metals (IRELAND
1982, after BABULA & SKOWROÑSKA-WENDLAND 1988).The stomach of H. lutescens passes into the intes-
tine (Fig. 14g), consisting of the small intestine andrectum. Both these sections are lined with cylindricalepithelium with numerous glandular cells. The small
210 Koralewska-Batura E.
Figs 19–22. Radular teeth: 19 – blunted teeth, 20 – sharpteeth, 21 – young teeth (after KORALEWSKA-BATURA1994b), 790×, 22 – a fragment of radular pocket withodontoblasts and a forming row of teeth, 245×
Figs 23, 24. Radular teeth: 23 – arrangement of teeth, 395×,24 – a fragment of radula, C – central tooth, L – lateralteeth, M – marginal teeth (after KORALEWSKA-BATURA1994b), 395×
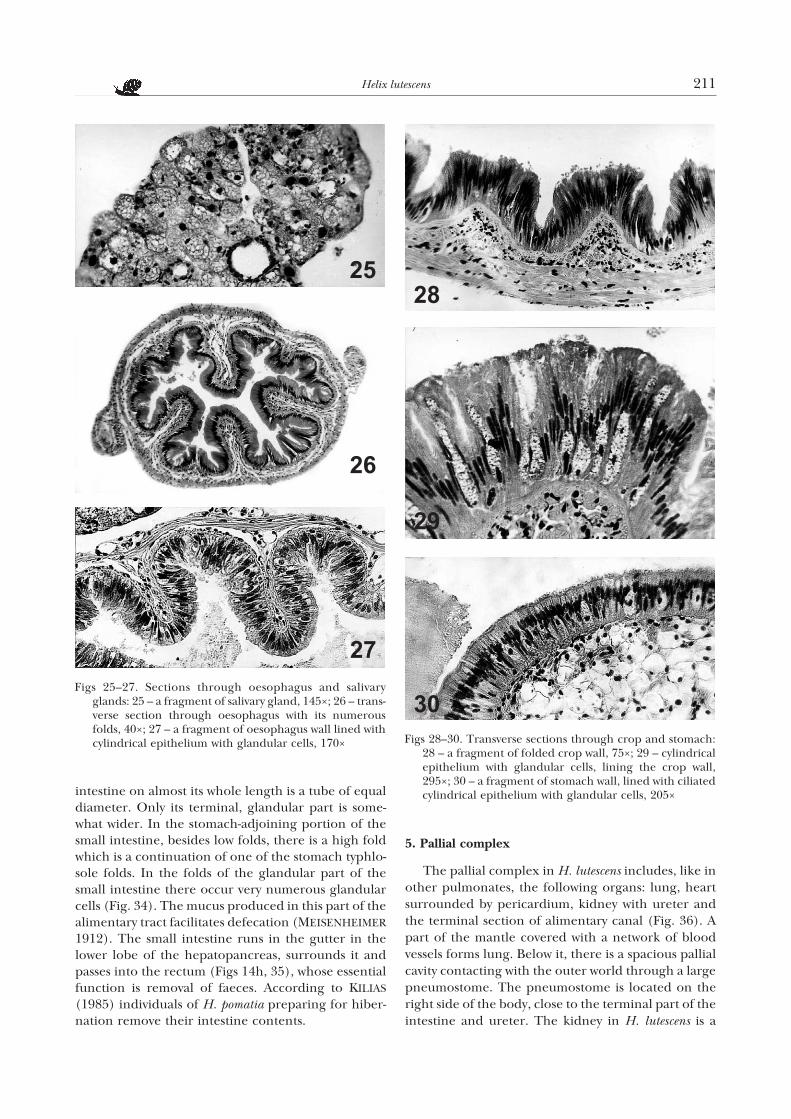
intestine on almost its whole length is a tube of equaldiameter. Only its terminal, glandular part is some-what wider. In the stomach-adjoining portion of thesmall intestine, besides low folds, there is a high foldwhich is a continuation of one of the stomach typhlo-sole folds. In the folds of the glandular part of thesmall intestine there occur very numerous glandularcells (Fig. 34). The mucus produced in this part of thealimentary tract facilitates defecation (MEISENHEIMER
1912). The small intestine runs in the gutter in thelower lobe of the hepatopancreas, surrounds it andpasses into the rectum (Figs 14h, 35), whose essentialfunction is removal of faeces. According to KILIAS
(1985) individuals of H. pomatia preparing for hiber-nation remove their intestine contents.
5. Pallial complex
The pallial complex in H. lutescens includes, like inother pulmonates, the following organs: lung, heartsurrounded by pericardium, kidney with ureter andthe terminal section of alimentary canal (Fig. 36). Apart of the mantle covered with a network of bloodvessels forms lung. Below it, there is a spacious pallialcavity contacting with the outer world through a largepneumostome. The pneumostome is located on theright side of the body, close to the terminal part of theintestine and ureter. The kidney in H. lutescens is a
Helix lutescens 211
Figs 25–27. Sections through oesophagus and salivaryglands: 25 – a fragment of salivary gland, 145×; 26 – trans-verse section through oesophagus with its numerousfolds, 40×; 27 – a fragment of oesophagus wall lined withcylindrical epithelium with glandular cells, 170× Figs 28–30. Transverse sections through crop and stomach:
28 – a fragment of folded crop wall, 75×; 29 – cylindricalepithelium with glandular cells, lining the crop wall,295×; 30 – a fragment of stomach wall, lined with ciliatedcylindrical epithelium with glandular cells, 205×
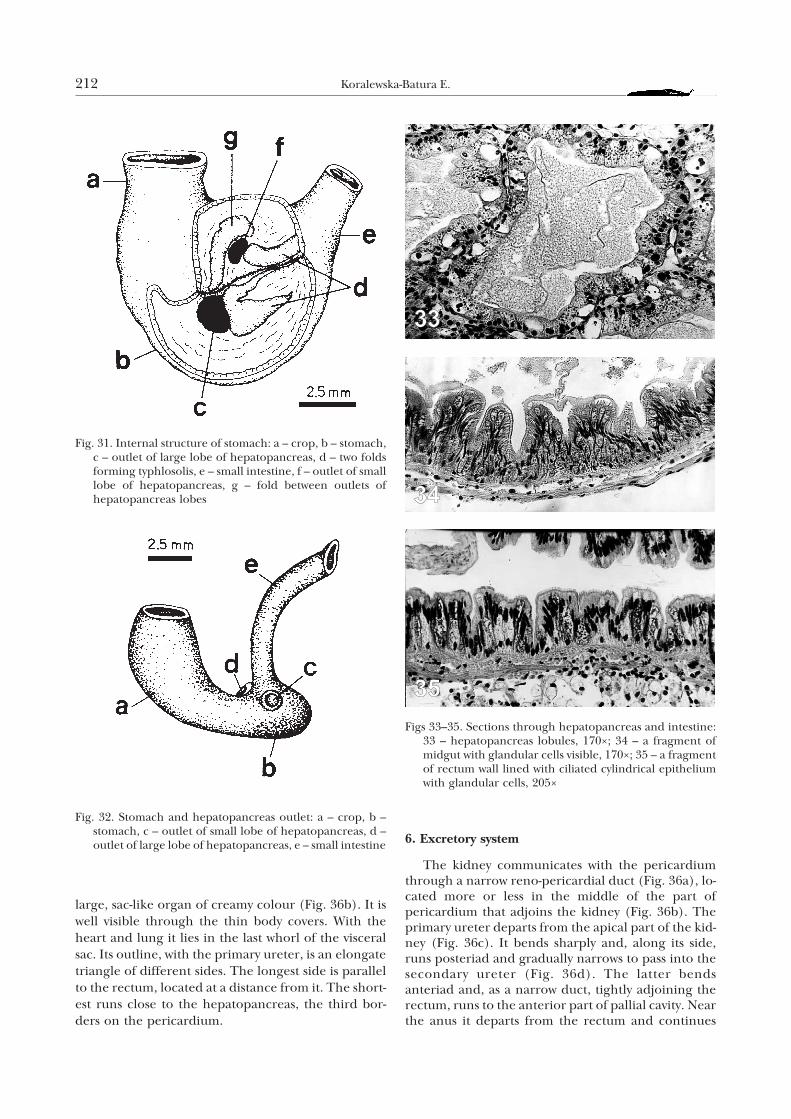
large, sac-like organ of creamy colour (Fig. 36b). It iswell visible through the thin body covers. With theheart and lung it lies in the last whorl of the visceralsac. Its outline, with the primary ureter, is an elongatetriangle of different sides. The longest side is parallelto the rectum, located at a distance from it. The short-est runs close to the hepatopancreas, the third bor-ders on the pericardium.
6. Excretory system
The kidney communicates with the pericardiumthrough a narrow reno-pericardial duct (Fig. 36a), lo-cated more or less in the middle of the part ofpericardium that adjoins the kidney (Fig. 36b). Theprimary ureter departs from the apical part of the kid-ney (Fig. 36c). It bends sharply and, along its side,runs posteriad and gradually narrows to pass into thesecondary ureter (Fig. 36d). The latter bendsanteriad and, as a narrow duct, tightly adjoining therectum, runs to the anterior part of pallial cavity. Nearthe anus it departs from the rectum and continues
212 Koralewska-Batura E.
Fig. 31. Internal structure of stomach: a – crop, b – stomach,c – outlet of large lobe of hepatopancreas, d – two foldsforming typhlosolis, e – small intestine, f – outlet of smalllobe of hepatopancreas, g – fold between outlets ofhepatopancreas lobes
Fig. 32. Stomach and hepatopancreas outlet: a – crop, b –stomach, c – outlet of small lobe of hepatopancreas, d –outlet of large lobe of hepatopancreas, e – small intestine
Figs 33–35. Sections through hepatopancreas and intestine:33 – hepatopancreas lobules, 170×; 34 – a fragment ofmidgut with glandular cells visible, 170×; 35 – a fragmentof rectum wall lined with ciliated cylindrical epitheliumwith glandular cells, 205×
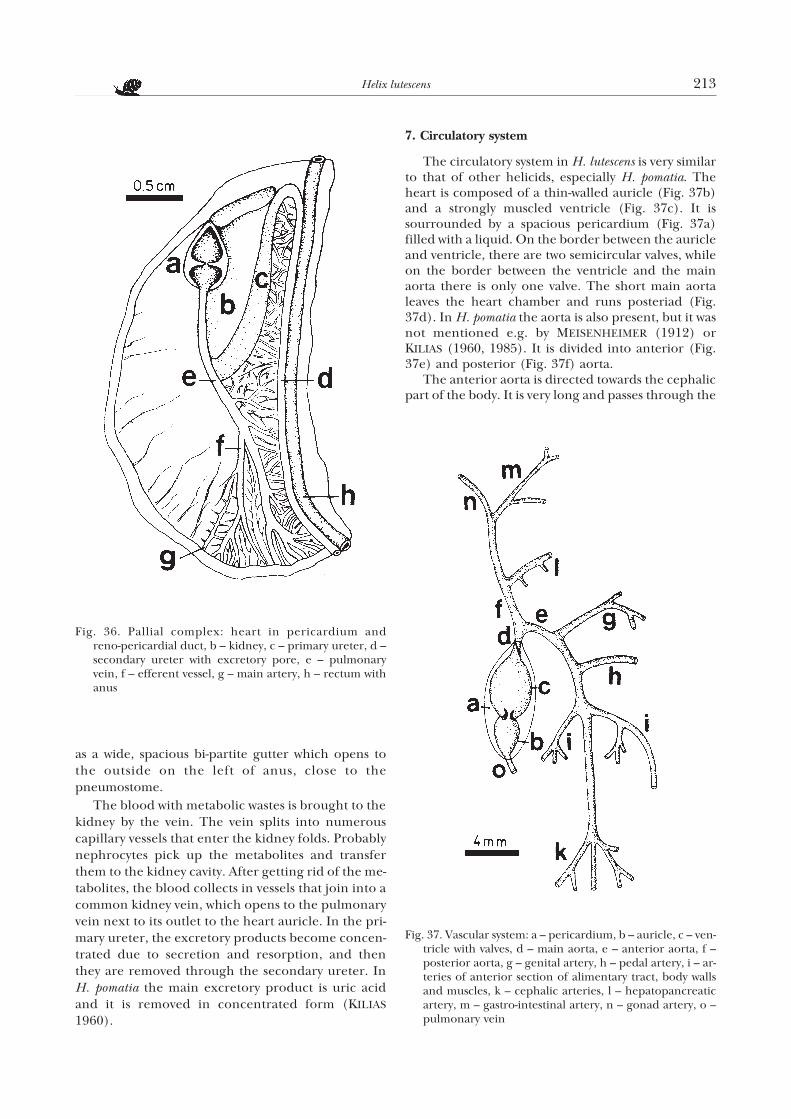
as a wide, spacious bi-partite gutter which opens tothe outside on the left of anus, close to thepneumostome.
The blood with metabolic wastes is brought to thekidney by the vein. The vein splits into numerouscapillary vessels that enter the kidney folds. Probablynephrocytes pick up the metabolites and transferthem to the kidney cavity. After getting rid of the me-tabolites, the blood collects in vessels that join into acommon kidney vein, which opens to the pulmonaryvein next to its outlet to the heart auricle. In the pri-mary ureter, the excretory products become concen-trated due to secretion and resorption, and thenthey are removed through the secondary ureter. InH. pomatia the main excretory product is uric acidand it is removed in concentrated form (KILIAS
1960).
7. Circulatory system
The circulatory system in H. lutescens is very similarto that of other helicids, especially H. pomatia. Theheart is composed of a thin-walled auricle (Fig. 37b)and a strongly muscled ventricle (Fig. 37c). It issourrounded by a spacious pericardium (Fig. 37a)filled with a liquid. On the border between the auricleand ventricle, there are two semicircular valves, whileon the border between the ventricle and the mainaorta there is only one valve. The short main aortaleaves the heart chamber and runs posteriad (Fig.37d). In H. pomatia the aorta is also present, but it wasnot mentioned e.g. by MEISENHEIMER (1912) orKILIAS (1960, 1985). It is divided into anterior (Fig.37e) and posterior (Fig. 37f) aorta.
The anterior aorta is directed towards the cephalicpart of the body. It is very long and passes through the
Helix lutescens 213
Fig. 36. Pallial complex: heart in pericardium andreno-pericardial duct, b – kidney, c – primary ureter, d –secondary ureter with excretory pore, e – pulmonaryvein, f – efferent vessel, g – main artery, h – rectum withanus
Fig. 37. Vascular system: a – pericardium, b – auricle, c – ven-tricle with valves, d – main aorta, e – anterior aorta, f –posterior aorta, g – genital artery, h – pedal artery, i – ar-teries of anterior section of alimentary tract, body wallsand muscles, k – cephalic arteries, l – hepatopancreaticartery, m – gastro-intestinal artery, n – gonad artery, o –pulmonary vein
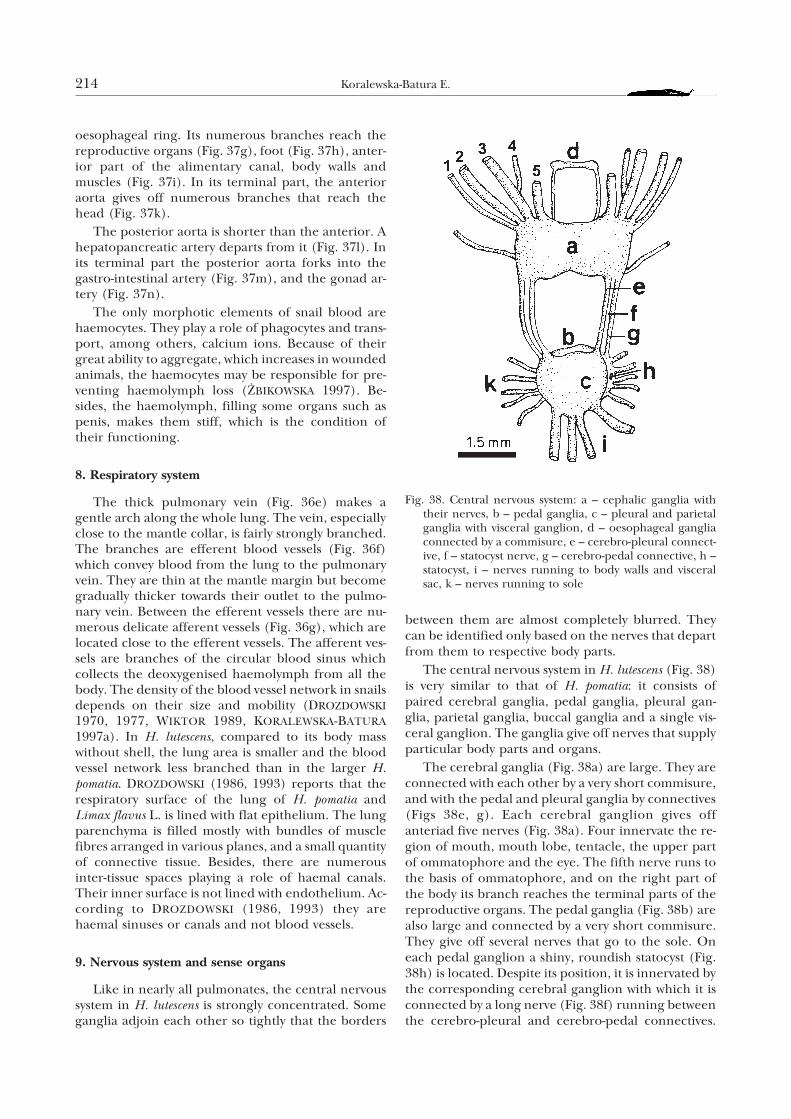
oesophageal ring. Its numerous branches reach thereproductive organs (Fig. 37g), foot (Fig. 37h), anter-ior part of the alimentary canal, body walls andmuscles (Fig. 37i). In its terminal part, the anterioraorta gives off numerous branches that reach thehead (Fig. 37k).
The posterior aorta is shorter than the anterior. Ahepatopancreatic artery departs from it (Fig. 37l). Inits terminal part the posterior aorta forks into thegastro-intestinal artery (Fig. 37m), and the gonad ar-tery (Fig. 37n).
The only morphotic elements of snail blood arehaemocytes. They play a role of phagocytes and trans-port, among others, calcium ions. Because of theirgreat ability to aggregate, which increases in woundedanimals, the haemocytes may be responsible for pre-venting haemolymph loss (¯BIKOWSKA 1997). Be-sides, the haemolymph, filling some organs such aspenis, makes them stiff, which is the condition oftheir functioning.
8. Respiratory system
The thick pulmonary vein (Fig. 36e) makes agentle arch along the whole lung. The vein, especiallyclose to the mantle collar, is fairly strongly branched.The branches are efferent blood vessels (Fig. 36f)which convey blood from the lung to the pulmonaryvein. They are thin at the mantle margin but becomegradually thicker towards their outlet to the pulmo-nary vein. Between the efferent vessels there are nu-merous delicate afferent vessels (Fig. 36g), which arelocated close to the efferent vessels. The afferent ves-sels are branches of the circular blood sinus whichcollects the deoxygenised haemolymph from all thebody. The density of the blood vessel network in snailsdepends on their size and mobility (DROZDOWSKI1970, 1977, WIKTOR 1989, KORALEWSKA-BATURA1997a). In H. lutescens, compared to its body masswithout shell, the lung area is smaller and the bloodvessel network less branched than in the larger H.pomatia. DROZDOWSKI (1986, 1993) reports that therespiratory surface of the lung of H. pomatia andLimax flavus L. is lined with flat epithelium. The lungparenchyma is filled mostly with bundles of musclefibres arranged in various planes, and a small quantityof connective tissue. Besides, there are numerousinter-tissue spaces playing a role of haemal canals.Their inner surface is not lined with endothelium. Ac-cording to DROZDOWSKI (1986, 1993) they arehaemal sinuses or canals and not blood vessels.
9. Nervous system and sense organs
Like in nearly all pulmonates, the central nervoussystem in H. lutescens is strongly concentrated. Someganglia adjoin each other so tightly that the borders
between them are almost completely blurred. Theycan be identified only based on the nerves that departfrom them to respective body parts.
The central nervous system in H. lutescens (Fig. 38)is very similar to that of H. pomatia: it consists ofpaired cerebral ganglia, pedal ganglia, pleural gan-glia, parietal ganglia, buccal ganglia and a single vis-ceral ganglion. The ganglia give off nerves that supplyparticular body parts and organs.
The cerebral ganglia (Fig. 38a) are large. They areconnected with each other by a very short commisure,and with the pedal and pleural ganglia by connectives(Figs 38e, g). Each cerebral ganglion gives offanteriad five nerves (Fig. 38a). Four innervate the re-gion of mouth, mouth lobe, tentacle, the upper partof ommatophore and the eye. The fifth nerve runs tothe basis of ommatophore, and on the right part ofthe body its branch reaches the terminal parts of thereproductive organs. The pedal ganglia (Fig. 38b) arealso large and connected by a very short commisure.They give off several nerves that go to the sole. Oneach pedal ganglion a shiny, roundish statocyst (Fig.38h) is located. Despite its position, it is innervated bythe corresponding cerebral ganglion with which it isconnected by a long nerve (Fig. 38f) running betweenthe cerebro-pleural and cerebro-pedal connectives.
214 Koralewska-Batura E.
Fig. 38. Central nervous system: a – cephalic ganglia withtheir nerves, b – pedal ganglia, c – pleural and parietalganglia with visceral ganglion, d – oesophageal gangliaconnected by a commisure, e – cerebro-pleural connect-ive, f – statocyst nerve, g – cerebro-pedal connective, h –statocyst, i – nerves running to body walls and visceralsac, k – nerves running to sole
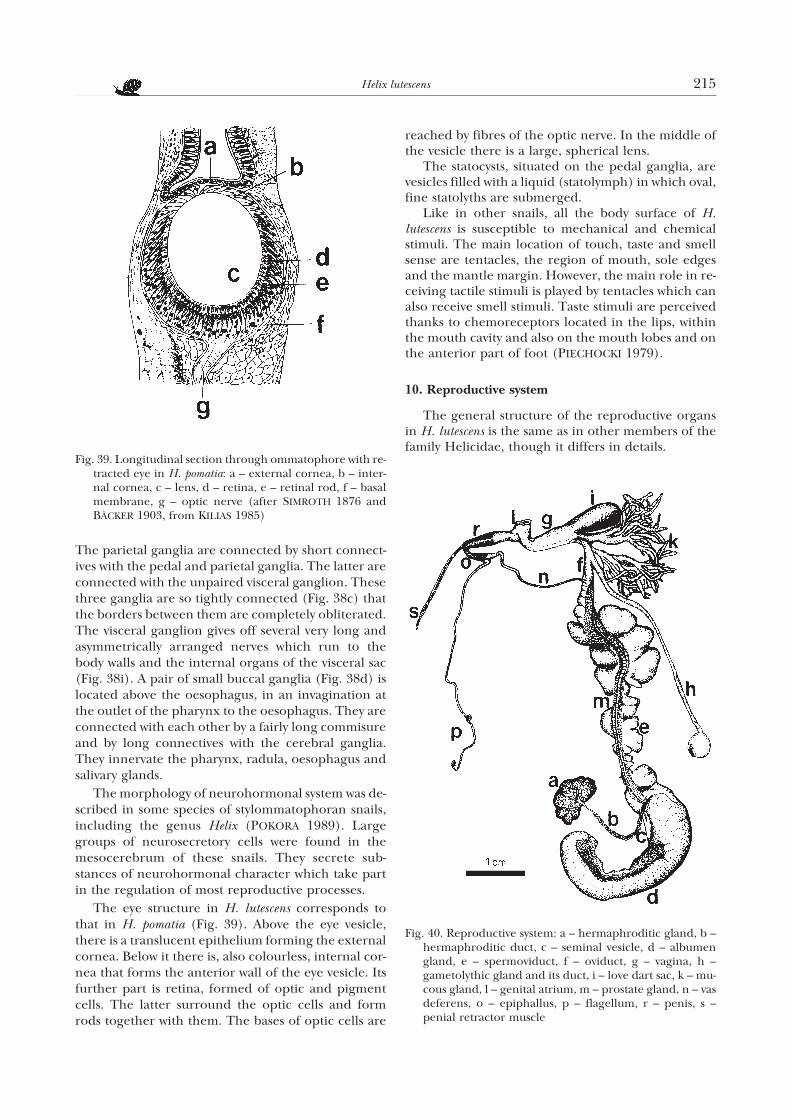
The parietal ganglia are connected by short connect-ives with the pedal and parietal ganglia. The latter areconnected with the unpaired visceral ganglion. Thesethree ganglia are so tightly connected (Fig. 38c) thatthe borders between them are completely obliterated.The visceral ganglion gives off several very long andasymmetrically arranged nerves which run to thebody walls and the internal organs of the visceral sac(Fig. 38i). A pair of small buccal ganglia (Fig. 38d) islocated above the oesophagus, in an invagination atthe outlet of the pharynx to the oesophagus. They areconnected with each other by a fairly long commisureand by long connectives with the cerebral ganglia.They innervate the pharynx, radula, oesophagus andsalivary glands.
The morphology of neurohormonal system was de-scribed in some species of stylommatophoran snails,including the genus Helix (POKORA 1989). Largegroups of neurosecretory cells were found in themesocerebrum of these snails. They secrete sub-stances of neurohormonal character which take partin the regulation of most reproductive processes.
The eye structure in H. lutescens corresponds tothat in H. pomatia (Fig. 39). Above the eye vesicle,there is a translucent epithelium forming the externalcornea. Below it there is, also colourless, internal cor-nea that forms the anterior wall of the eye vesicle. Itsfurther part is retina, formed of optic and pigmentcells. The latter surround the optic cells and formrods together with them. The bases of optic cells are
reached by fibres of the optic nerve. In the middle ofthe vesicle there is a large, spherical lens.
The statocysts, situated on the pedal ganglia, arevesicles filled with a liquid (statolymph) in which oval,fine statolyths are submerged.
Like in other snails, all the body surface of H.lutescens is susceptible to mechanical and chemicalstimuli. The main location of touch, taste and smellsense are tentacles, the region of mouth, sole edgesand the mantle margin. However, the main role in re-ceiving tactile stimuli is played by tentacles which canalso receive smell stimuli. Taste stimuli are perceivedthanks to chemoreceptors located in the lips, withinthe mouth cavity and also on the mouth lobes and onthe anterior part of foot (PIECHOCKI 1979).
10. Reproductive system
The general structure of the reproductive organsin H. lutescens is the same as in other members of thefamily Helicidae, though it differs in details.
Helix lutescens 215
Fig. 40. Reproductive system: a – hermaphroditic gland, b –hermaphroditic duct, c – seminal vesicle, d – albumengland, e – spermoviduct, f – oviduct, g – vagina, h –gametolythic gland and its duct, i – love dart sac, k – mu-cous gland, l – genital atrium, m – prostate gland, n – vasdeferens, o – epiphallus, p – flagellum, r – penis, s –penial retractor muscle
Fig. 39. Longitudinal section through ommatophore with re-tracted eye in H. pomatia: a – external cornea, b – inter-nal cornea, c – lens, d – retina, e – retinal rod, f – basalmembrane, g – optic nerve (after SIMROTH 1876 andBÄCKER 1903, from KILIAS 1985)
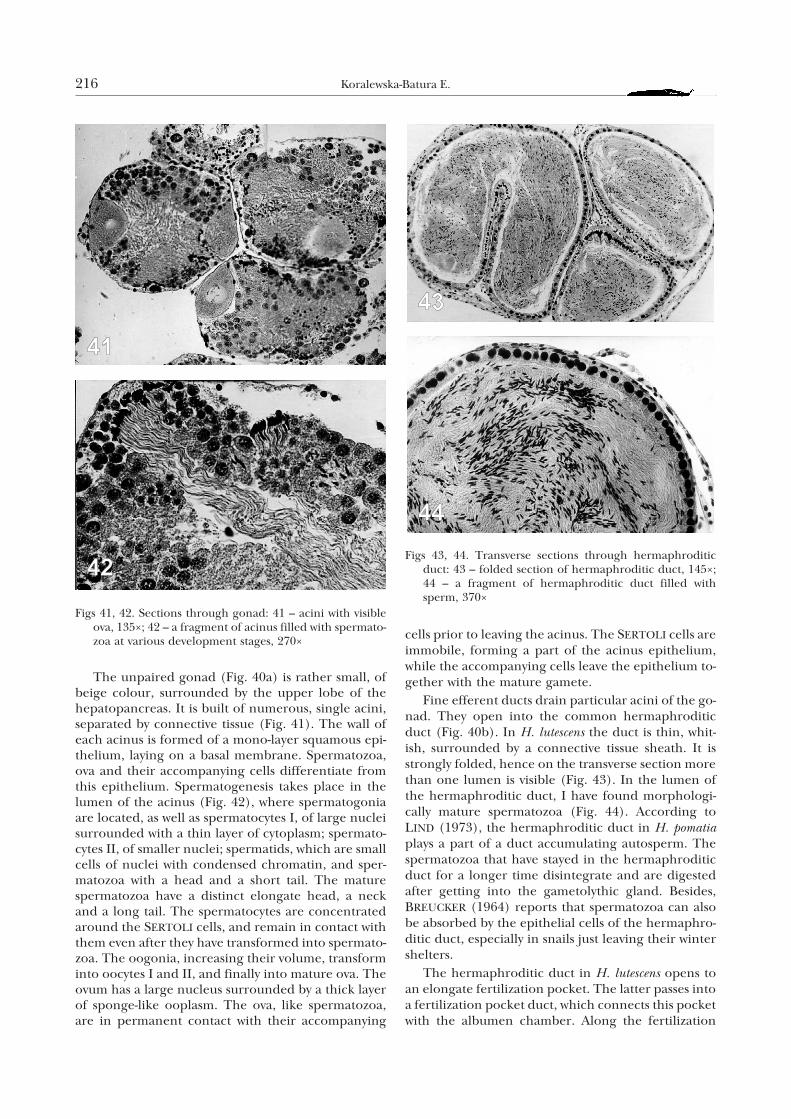
The unpaired gonad (Fig. 40a) is rather small, ofbeige colour, surrounded by the upper lobe of thehepatopancreas. It is built of numerous, single acini,separated by connective tissue (Fig. 41). The wall ofeach acinus is formed of a mono-layer squamous epi-thelium, laying on a basal membrane. Spermatozoa,ova and their accompanying cells differentiate fromthis epithelium. Spermatogenesis takes place in thelumen of the acinus (Fig. 42), where spermatogoniaare located, as well as spermatocytes I, of large nucleisurrounded with a thin layer of cytoplasm; spermato-cytes II, of smaller nuclei; spermatids, which are smallcells of nuclei with condensed chromatin, and sper-matozoa with a head and a short tail. The maturespermatozoa have a distinct elongate head, a neckand a long tail. The spermatocytes are concentratedaround the SERTOLI cells, and remain in contact withthem even after they have transformed into spermato-zoa. The oogonia, increasing their volume, transforminto oocytes I and II, and finally into mature ova. Theovum has a large nucleus surrounded by a thick layerof sponge-like ooplasm. The ova, like spermatozoa,are in permanent contact with their accompanying
cells prior to leaving the acinus. The SERTOLI cells areimmobile, forming a part of the acinus epithelium,while the accompanying cells leave the epithelium to-gether with the mature gamete.
Fine efferent ducts drain particular acini of the go-nad. They open into the common hermaphroditicduct (Fig. 40b). In H. lutescens the duct is thin, whit-ish, surrounded by a connective tissue sheath. It isstrongly folded, hence on the transverse section morethan one lumen is visible (Fig. 43). In the lumen ofthe hermaphroditic duct, I have found morphologi-cally mature spermatozoa (Fig. 44). According toLIND (1973), the hermaphroditic duct in H. pomatiaplays a part of a duct accumulating autosperm. Thespermatozoa that have stayed in the hermaphroditicduct for a longer time disintegrate and are digestedafter getting into the gametolythic gland. Besides,BREUCKER (1964) reports that spermatozoa can alsobe absorbed by the epithelial cells of the hermaphro-ditic duct, especially in snails just leaving their wintershelters.
The hermaphroditic duct in H. lutescens opens toan elongate fertilization pocket. The latter passes intoa fertilization pocket duct, which connects this pocketwith the albumen chamber. Along the fertilization
216 Koralewska-Batura E.
Figs 41, 42. Sections through gonad: 41 – acini with visibleova, 135×; 42 – a fragment of acinus filled with spermato-zoa at various development stages, 270×
Figs 43, 44. Transverse sections through hermaphroditicduct: 43 – folded section of hermaphroditic duct, 145×;44 – a fragment of hermaphroditic duct filled withsperm, 370×
Helix lutescens 217
Figs 46–48. Transverse sections through fertilisation pocket and seminal vesicles: 46 – fertilisation pocket and five seminalvesicles, 175×; 47 – fertilisation pocket and seminal vesicle with ciliated cylindrical epithelium, 150×; 48 – seminal vesiclefilled with spermatozoa arranged in packets, 230×
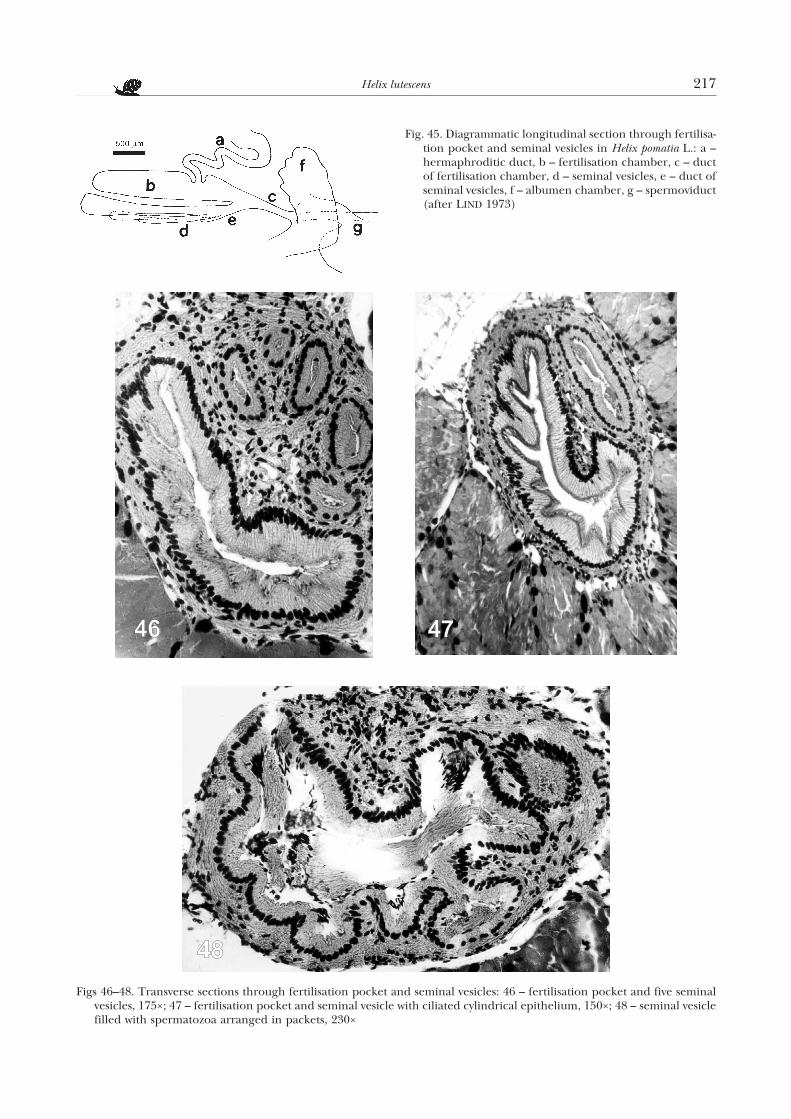
Fig. 45. Diagrammatic longitudinal section through fertilisa-tion pocket and seminal vesicles in Helix pomatia L.: a –hermaphroditic duct, b – fertilisation chamber, c – ductof fertilisation chamber, d – seminal vesicles, e – duct ofseminal vesicles, f – albumen chamber, g – spermoviduct(after LIND 1973)
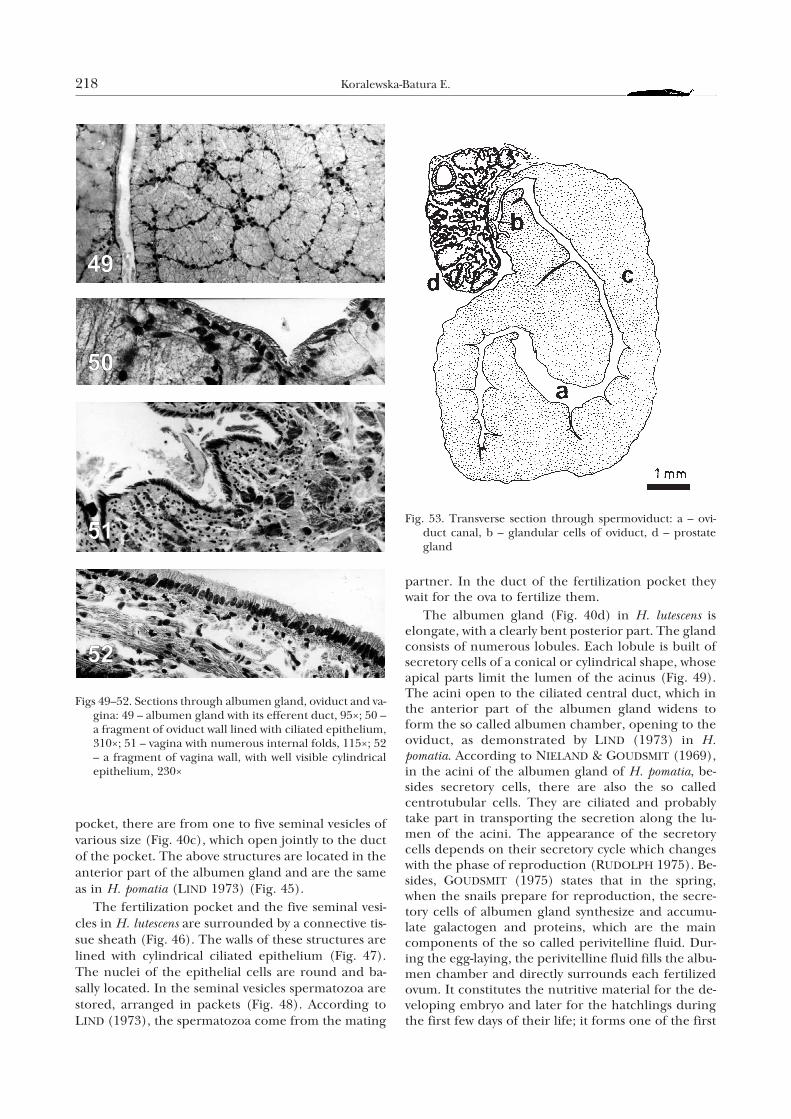
pocket, there are from one to five seminal vesicles ofvarious size (Fig. 40c), which open jointly to the ductof the pocket. The above structures are located in theanterior part of the albumen gland and are the sameas in H. pomatia (LIND 1973) (Fig. 45).
The fertilization pocket and the five seminal vesi-cles in H. lutescens are surrounded by a connective tis-sue sheath (Fig. 46). The walls of these structures arelined with cylindrical ciliated epithelium (Fig. 47).The nuclei of the epithelial cells are round and ba-sally located. In the seminal vesicles spermatozoa arestored, arranged in packets (Fig. 48). According toLIND (1973), the spermatozoa come from the mating
partner. In the duct of the fertilization pocket theywait for the ova to fertilize them.
The albumen gland (Fig. 40d) in H. lutescens iselongate, with a clearly bent posterior part. The glandconsists of numerous lobules. Each lobule is built ofsecretory cells of a conical or cylindrical shape, whoseapical parts limit the lumen of the acinus (Fig. 49).The acini open to the ciliated central duct, which inthe anterior part of the albumen gland widens toform the so called albumen chamber, opening to theoviduct, as demonstrated by LIND (1973) in H.pomatia. According to NIELAND & GOUDSMIT (1969),in the acini of the albumen gland of H. pomatia, be-sides secretory cells, there are also the so calledcentrotubular cells. They are ciliated and probablytake part in transporting the secretion along the lu-men of the acini. The appearance of the secretorycells depends on their secretory cycle which changeswith the phase of reproduction (RUDOLPH 1975). Be-sides, GOUDSMIT (1975) states that in the spring,when the snails prepare for reproduction, the secre-tory cells of albumen gland synthesize and accumu-late galactogen and proteins, which are the maincomponents of the so called perivitelline fluid. Dur-ing the egg-laying, the perivitelline fluid fills the albu-men chamber and directly surrounds each fertilizedovum. It constitutes the nutritive material for the de-veloping embryo and later for the hatchlings duringthe first few days of their life; it forms one of the first
218 Koralewska-Batura E.
Figs 49–52. Sections through albumen gland, oviduct and va-gina: 49 – albumen gland with its efferent duct, 95×; 50 –a fragment of oviduct wall lined with ciliated epithelium,310×; 51 – vagina with numerous internal folds, 115×; 52– a fragment of vagina wall, with well visible cylindricalepithelium, 230×
Fig. 53. Transverse section through spermoviduct: a – ovi-duct canal, b – glandular cells of oviduct, d – prostategland
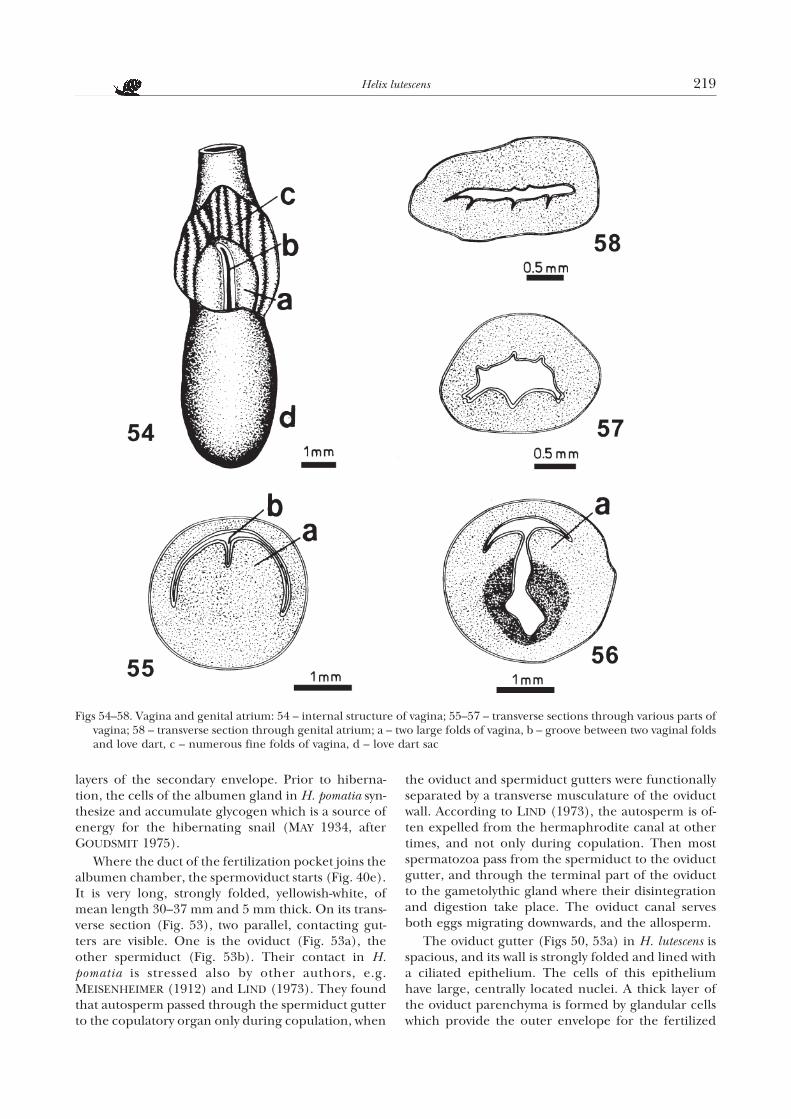
layers of the secondary envelope. Prior to hiberna-tion, the cells of the albumen gland in H. pomatia syn-thesize and accumulate glycogen which is a source ofenergy for the hibernating snail (MAY 1934, afterGOUDSMIT 1975).
Where the duct of the fertilization pocket joins thealbumen chamber, the spermoviduct starts (Fig. 40e).It is very long, strongly folded, yellowish-white, ofmean length 30–37 mm and 5 mm thick. On its trans-verse section (Fig. 53), two parallel, contacting gut-ters are visible. One is the oviduct (Fig. 53a), theother spermiduct (Fig. 53b). Their contact in H.pomatia is stressed also by other authors, e.g.MEISENHEIMER (1912) and LIND (1973). They foundthat autosperm passed through the spermiduct gutterto the copulatory organ only during copulation, when
the oviduct and spermiduct gutters were functionallyseparated by a transverse musculature of the oviductwall. According to LIND (1973), the autosperm is of-ten expelled from the hermaphrodite canal at othertimes, and not only during copulation. Then mostspermatozoa pass from the spermiduct to the oviductgutter, and through the terminal part of the oviductto the gametolythic gland where their disintegrationand digestion take place. The oviduct canal servesboth eggs migrating downwards, and the allosperm.
The oviduct gutter (Figs 50, 53a) in H. lutescens isspacious, and its wall is strongly folded and lined witha ciliated epithelium. The cells of this epitheliumhave large, centrally located nuclei. A thick layer ofthe oviduct parenchyma is formed by glandular cellswhich provide the outer envelope for the fertilized
Helix lutescens 219
Figs 54–58. Vagina and genital atrium: 54 – internal structure of vagina; 55–57 – transverse sections through various parts ofvagina; 58 – transverse section through genital atrium; a – two large folds of vagina, b – groove between two vaginal foldsand love dart, c – numerous fine folds of vagina, d – love dart sac
eggs. The external envelope is the main source of cal-cium for the developing embryo. Where thespermoviduct ends, the oviduct canal passes into thefree oviduct and then into the rather long vagina (Fig.40f, g).
The internal structure of the vagina in H. lutescensis much varied. In its lumen, there are two large folds(Figs 54a – 56a). Between them, there is a fissure inwhich the terminal part of the love dart is located(Figs 54b – 55b). Towards the dart sac, the fissure-likelumen between the large vagina folds deepens into acommon lumen of both these organs (Fig. 56). Closerto the genital atrium, on the vagina wall, there are nu-
merous folds (Figs 51, 54c, 57). The vagina (Fig. 52) islined with cylindrical epithelium whose cells haveelongate, basally located nuclei. Below the epitheliumthere is a connective tissue layer, with single musclefibres and glandular (mucous) cells embedded in it.The connective tissue is adjoined by a thick layer ofbundles of variously arranged muscle fibres.
The duct of the gametolythic gland opens to thevagina (Fig. 40h). The mean length of the duct is 30mm. I have found, as was reported by ROSSMÄSSLER
(1837), that in H. lutescens it is always devoid ofdiverticle, which is present e.g. in the related H.pomatia. Like the vagina, the spermatheca duct on its
220 Koralewska-Batura E.
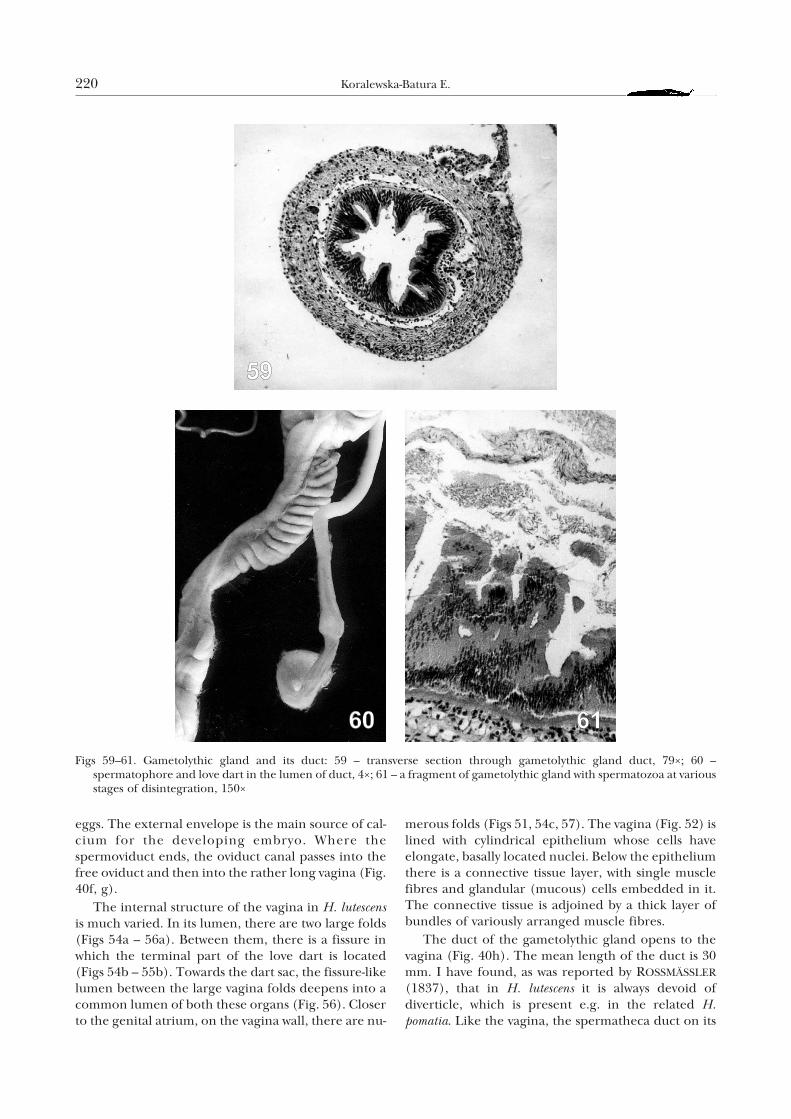
Figs 59–61. Gametolythic gland and its duct: 59 – transverse section through gametolythic gland duct, 79×; 60 –spermatophore and love dart in the lumen of duct, 4×; 61 – a fragment of gametolythic gland with spermatozoa at variousstages of disintegration, 150×

whole length is built of cylindrical epithelium andmuscular layer. Based on this, LIND (1973) postulatedthat in H. pomatia it was a continuation of the vagina.The lumen of the gametolythic gland duct is narrowand irregular (Fig. 59). Some cells of its epitheliumare higher, others lower, surrounded by a thin layer ofcircular muscle fibres. Below, there is a thick layer oflongitudinal muscle fibres, interspersed with connect-ive tissue. Following copulation, in the gametolythicgland duct of H. lutescens, the spermatophore andlove dart are present (Fig. 60). According to LIND
(1973), in H. pomatia strong unidirectional contrac-tions of the muscular layer of the duct move thespermatophore to the gametolythic gland.
The gametolythic gland (Fig. 40h) in H. lutescens isspherical or oval, 3–4 mm in diameter and ofred-brown colour. It is located just next to the kidney.In the lumen of the gland I found mucus and fewspermatozoa at various stages of disintegration (Fig.61). Following copulation, the whole spermatophoreenters the gametolythic gland, and the unshot lovedart may also enter it (Fig. 60). According to LIND
(1973) the gland in H. pomatia is the place of disinte-gration of allosperm, excess of autosperm and ofother products of the reproductive system. This isconfirmed by the studies of HRYNIEWIECKA-SZYFTER &RÊDZINIAK (1976). LIND (1973) suggests that a part ofthe products decomposed in the gametolythic glandremains as a red mass composed of substances whichare not absorbed by the epithelium and will never beremoved from the gland. REEDER & ROGERS (1979)found that the gametolythic gland in the genusSonorella Pilsbry was a storage, digestive and absorp-tion organ.
The dart sac (Fig. 40i) opens to the vagina. In H.lutescens the sac is large, strongly muscled, of whitishcolour, and its shape is sac-like. Its mean length is5.0–5.5 mm, the diameter ca. 3 mm. On its longitudi-nal section (Fig. 62), a canal is visible, with a small pa-pilla, built of a thick layer of muscles. It is separated bya very narrow lumen from the bottom sac wall. The in-ner surface of the sac and the papilla are lined with cy-lindrical epithelium (Figs 63, 64). The cells of this epi-thelium have elongate, basally located nuclei. Underthe epithelium, there is a fairly thick layer of looseconnective tissue. Then, there is a thick muscle layer,with circularly and longitudinally arranged fibres.Calcium concretions were found in the sac tissues.They are the most numerous in the papilla (Figs 63,65), and much less so in the walls of the sac.DILLAMAN (1981) reports that in H. aspersa the lovedart is formed as a result of the secretory function ofthe papilla epithelium and the inner surface of thesac. It is built of aragonite. The anterior part of thelove dart in H. lutescens is set on the papilla. A more orless open calyx can be distinguished in it, which con-stitutes ca. 1/6 dart length (Fig. 66). The calyx con-
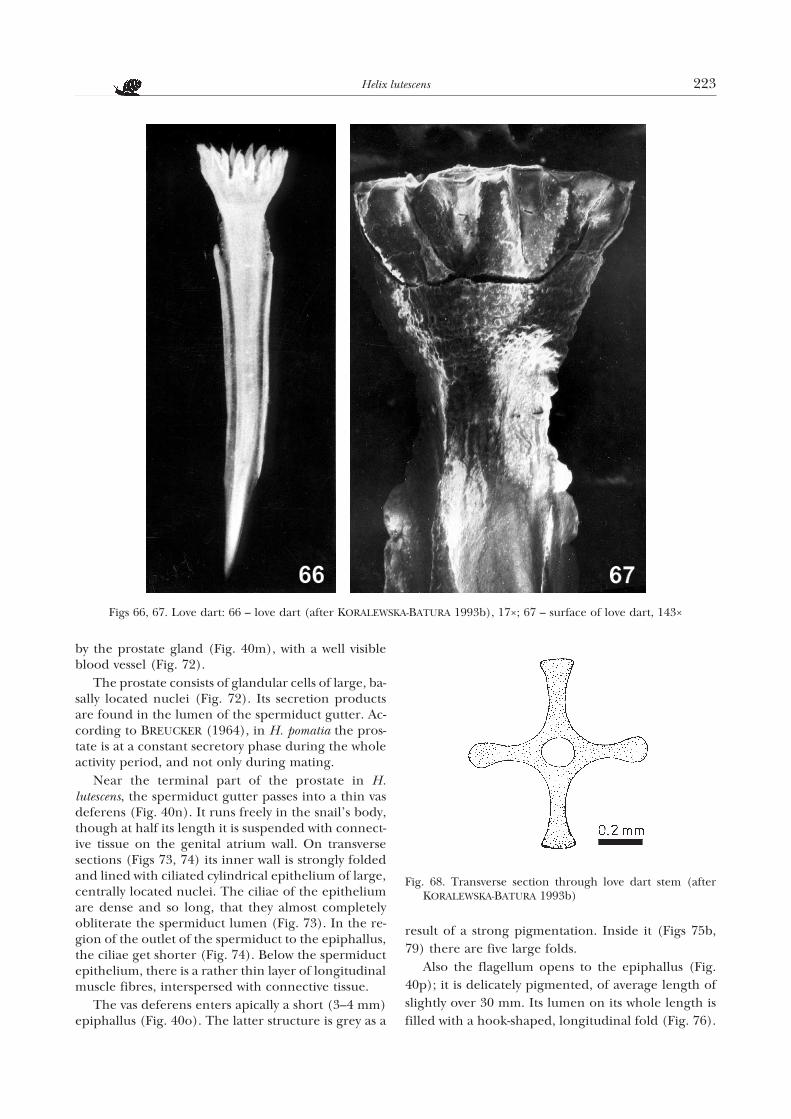
sists of a base on which there are “petals” with deli-cately serrated margins. They are rather wide andsometimes overlap. The calyx passes gently into aslightly bent stem which, gradually narrowing, isstrongly tapered at its end. On the stem there are fourribs. They start at the base of the calyx and continuealmost to the end of the stem. Perhaps they play a roleof reinforcing structures, as was observed by HUNT
(1979). The terminations of the love dart ribs in H.lutescens are of different shape (Fig. 68). Two oppositeribs are club-shaped, and the remaining two are onlydelicately rounded at their ends. Inside the stem ofthe love dart there is a canal. It is spacious, roundedin outline. The surface of the love dart (Fig. 67) isrough, covered with numerous tubercles. The latterare the most numerous on the surface of the calyxbase and undoubtedly also play a reinforcing role.FEDOSEEVA (1994) reports that in members of theHelicoidea the whole structure of the love dart in-creases its mechanical resistance. The mean length ofthe dart in H. lutescens is 6–7 mm, the width being 0.8mm. It is almost by 1/3 longer than the sac lumen, sothat its terminal part is located in the groove betweenthe two large vaginal folds (Fig. 54a).
Characteristic mucous glands (Fig. 40k) also opento the vagina. They are finger-shaped or tubular, ofwhite colour. They are arranged in two bundles, leftand right. The right bundle consists of an average of61 finger-like branches and opens to the vaginathrough a thick trunk. The left bundle, of an average
Helix lutescens 221
Fig. 62. Diagrammatic longitudinal section through lovedart sac: a – papilla, b – lumen between the papilla andthe bottom of the sac, c – sac canal, d – love dart, e – sacmuscles, f – vaginal folds
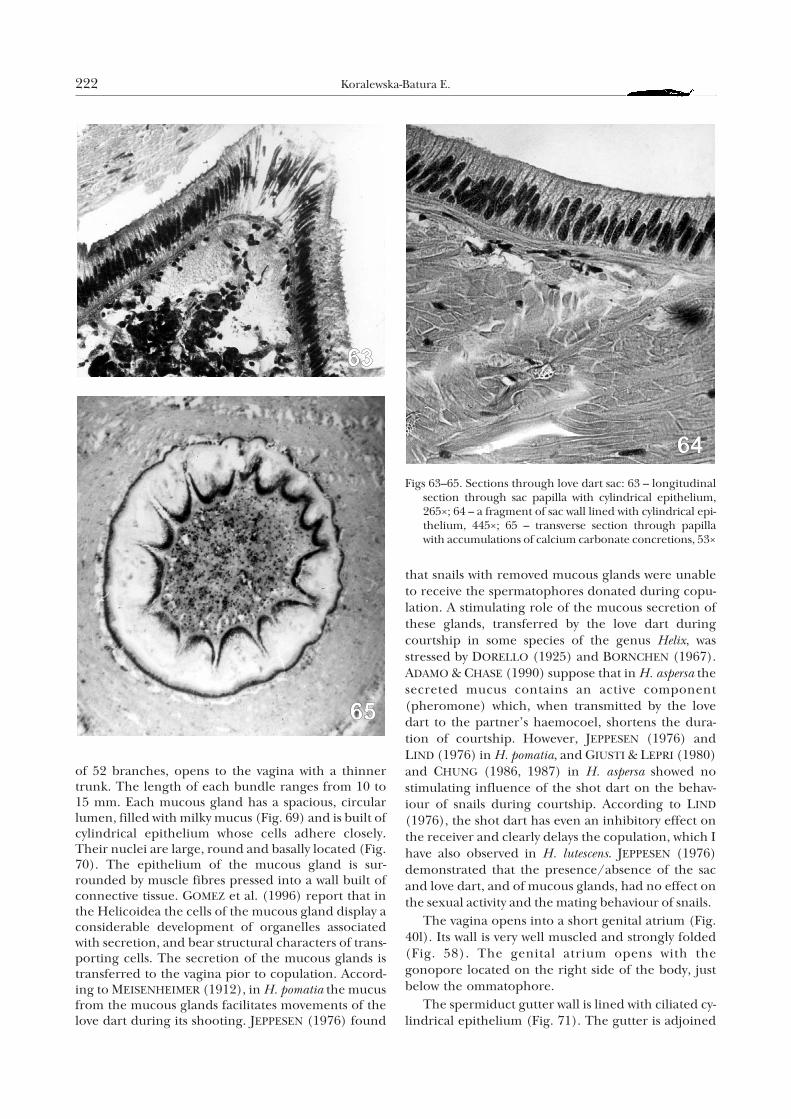
of 52 branches, opens to the vagina with a thinnertrunk. The length of each bundle ranges from 10 to15 mm. Each mucous gland has a spacious, circularlumen, filled with milky mucus (Fig. 69) and is built ofcylindrical epithelium whose cells adhere closely.Their nuclei are large, round and basally located (Fig.70). The epithelium of the mucous gland is sur-rounded by muscle fibres pressed into a wall built ofconnective tissue. GOMEZ et al. (1996) report that inthe Helicoidea the cells of the mucous gland display aconsiderable development of organelles associatedwith secretion, and bear structural characters of trans-porting cells. The secretion of the mucous glands istransferred to the vagina pior to copulation. Accord-ing to MEISENHEIMER (1912), in H. pomatia the mucusfrom the mucous glands facilitates movements of thelove dart during its shooting. JEPPESEN (1976) found
that snails with removed mucous glands were unableto receive the spermatophores donated during copu-lation. A stimulating role of the mucous secretion ofthese glands, transferred by the love dart duringcourtship in some species of the genus Helix, wasstressed by DORELLO (1925) and BORNCHEN (1967).ADAMO & CHASE (1990) suppose that in H. aspersa thesecreted mucus contains an active component(pheromone) which, when transmitted by the lovedart to the partner’s haemocoel, shortens the dura-tion of courtship. However, JEPPESEN (1976) andLIND (1976) in H. pomatia, and GIUSTI & LEPRI (1980)and CHUNG (1986, 1987) in H. aspersa showed nostimulating influence of the shot dart on the behav-iour of snails during courtship. According to LIND
(1976), the shot dart has even an inhibitory effect onthe receiver and clearly delays the copulation, which Ihave also observed in H. lutescens. JEPPESEN (1976)demonstrated that the presence/absence of the sacand love dart, and of mucous glands, had no effect onthe sexual activity and the mating behaviour of snails.
The vagina opens into a short genital atrium (Fig.40l). Its wall is very well muscled and strongly folded(Fig. 58). The genital atrium opens with thegonopore located on the right side of the body, justbelow the ommatophore.
The spermiduct gutter wall is lined with ciliated cy-lindrical epithelium (Fig. 71). The gutter is adjoined
222 Koralewska-Batura E.
Figs 63–65. Sections through love dart sac: 63 – longitudinalsection through sac papilla with cylindrical epithelium,265×; 64 – a fragment of sac wall lined with cylindrical epi-thelium, 445×; 65 – transverse section through papillawith accumulations of calcium carbonate concretions, 53×
by the prostate gland (Fig. 40m), with a well visibleblood vessel (Fig. 72).
The prostate consists of glandular cells of large, ba-sally located nuclei (Fig. 72). Its secretion productsare found in the lumen of the spermiduct gutter. Ac-cording to BREUCKER (1964), in H. pomatia the pros-tate is at a constant secretory phase during the wholeactivity period, and not only during mating.
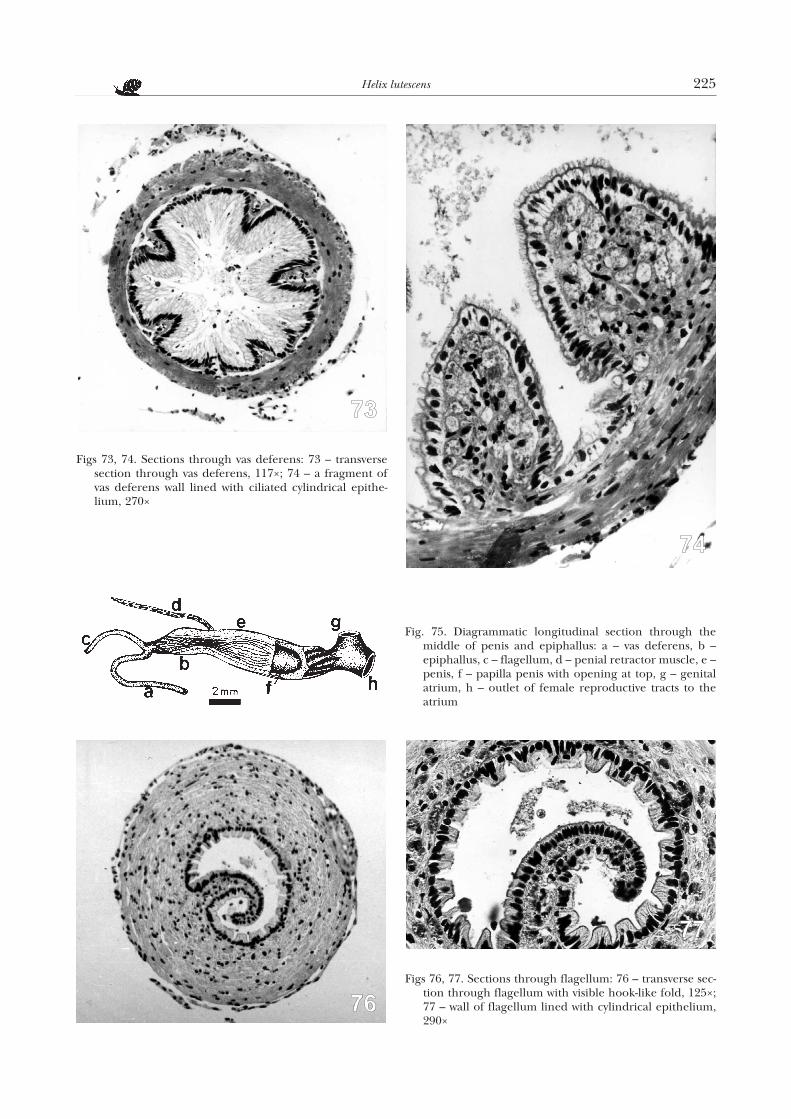
Near the terminal part of the prostate in H.lutescens, the spermiduct gutter passes into a thin vasdeferens (Fig. 40n). It runs freely in the snail’s body,though at half its length it is suspended with connect-ive tissue on the genital atrium wall. On transversesections (Figs 73, 74) its inner wall is strongly foldedand lined with ciliated cylindrical epithelium of large,centrally located nuclei. The ciliae of the epitheliumare dense and so long, that they almost completelyobliterate the spermiduct lumen (Fig. 73). In the re-gion of the outlet of the spermiduct to the epiphallus,the ciliae get shorter (Fig. 74). Below the spermiductepithelium, there is a rather thin layer of longitudinalmuscle fibres, interspersed with connective tissue.
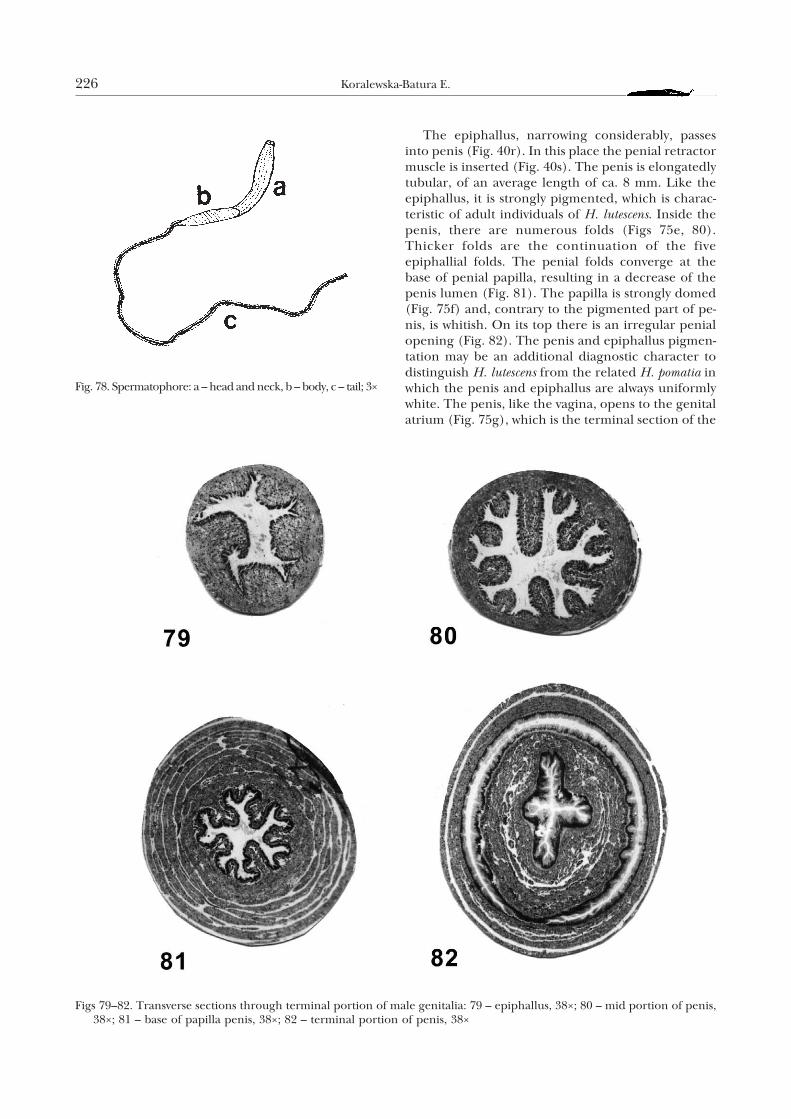
The vas deferens enters apically a short (3–4 mm)epiphallus (Fig. 40o). The latter structure is grey as a
result of a strong pigmentation. Inside it (Figs 75b,79) there are five large folds.
Also the flagellum opens to the epiphallus (Fig.40p); it is delicately pigmented, of average length ofslightly over 30 mm. Its lumen on its whole length isfilled with a hook-shaped, longitudinal fold (Fig. 76).
Helix lutescens 223
Fig. 68. Transverse section through love dart stem (afterKORALEWSKA-BATURA 1993b)
Figs 66, 67. Love dart: 66 – love dart (after KORALEWSKA-BATURA 1993b), 17×; 67 – surface of love dart, 143×
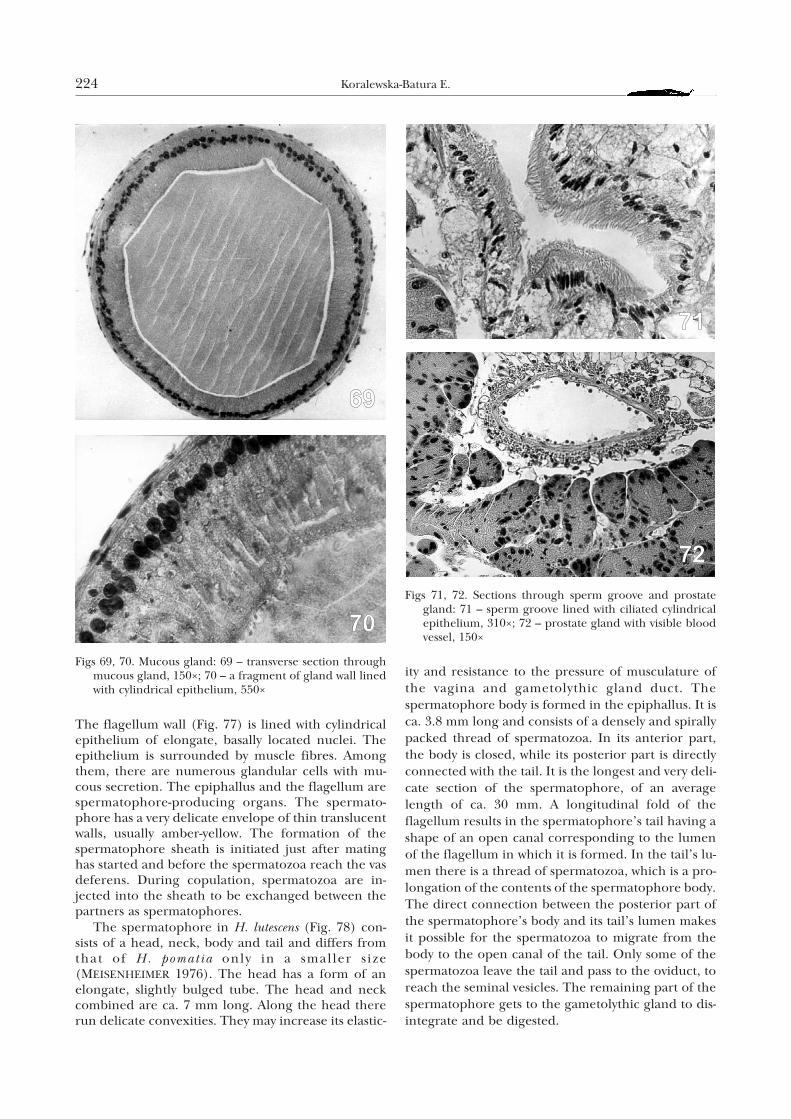
The flagellum wall (Fig. 77) is lined with cylindricalepithelium of elongate, basally located nuclei. Theepithelium is surrounded by muscle fibres. Amongthem, there are numerous glandular cells with mu-cous secretion. The epiphallus and the flagellum arespermatophore-producing organs. The spermato-phore has a very delicate envelope of thin translucentwalls, usually amber-yellow. The formation of thespermatophore sheath is initiated just after matinghas started and before the spermatozoa reach the vasdeferens. During copulation, spermatozoa are in-jected into the sheath to be exchanged between thepartners as spermatophores.
The spermatophore in H. lutescens (Fig. 78) con-sists of a head, neck, body and tail and differs fromthat of H. pomat ia only in a smal ler s ize(MEISENHEIMER 1976). The head has a form of anelongate, slightly bulged tube. The head and neckcombined are ca. 7 mm long. Along the head thererun delicate convexities. They may increase its elastic-
ity and resistance to the pressure of musculature ofthe vagina and gametolythic gland duct. Thespermatophore body is formed in the epiphallus. It isca. 3.8 mm long and consists of a densely and spirallypacked thread of spermatozoa. In its anterior part,the body is closed, while its posterior part is directlyconnected with the tail. It is the longest and very deli-cate section of the spermatophore, of an averagelength of ca. 30 mm. A longitudinal fold of theflagellum results in the spermatophore’s tail having ashape of an open canal corresponding to the lumenof the flagellum in which it is formed. In the tail’s lu-men there is a thread of spermatozoa, which is a pro-longation of the contents of the spermatophore body.The direct connection between the posterior part ofthe spermatophore’s body and its tail’s lumen makesit possible for the spermatozoa to migrate from thebody to the open canal of the tail. Only some of thespermatozoa leave the tail and pass to the oviduct, toreach the seminal vesicles. The remaining part of thespermatophore gets to the gametolythic gland to dis-integrate and be digested.
224 Koralewska-Batura E.
Figs 69, 70. Mucous gland: 69 – transverse section throughmucous gland, 150×; 70 – a fragment of gland wall linedwith cylindrical epithelium, 550×
Figs 71, 72. Sections through sperm groove and prostategland: 71 – sperm groove lined with ciliated cylindricalepithelium, 310×; 72 – prostate gland with visible bloodvessel, 150×
Helix lutescens 225
Figs 73, 74. Sections through vas deferens: 73 – transversesection through vas deferens, 117×; 74 – a fragment ofvas deferens wall lined with ciliated cylindrical epithe-lium, 270×
Fig. 75. Diagrammatic longitudinal section through themiddle of penis and epiphallus: a – vas deferens, b –epiphallus, c – flagellum, d – penial retractor muscle, e –penis, f – papilla penis with opening at top, g – genitalatrium, h – outlet of female reproductive tracts to theatrium
Figs 76, 77. Sections through flagellum: 76 – transverse sec-tion through flagellum with visible hook-like fold, 125×;77 – wall of flagellum lined with cylindrical epithelium,290×
The epiphallus, narrowing considerably, passesinto penis (Fig. 40r). In this place the penial retractormuscle is inserted (Fig. 40s). The penis is elongatedlytubular, of an average length of ca. 8 mm. Like theepiphallus, it is strongly pigmented, which is charac-teristic of adult individuals of H. lutescens. Inside thepenis, there are numerous folds (Figs 75e, 80).Thicker folds are the continuation of the fiveepiphallial folds. The penial folds converge at thebase of penial papilla, resulting in a decrease of thepenis lumen (Fig. 81). The papilla is strongly domed(Fig. 75f) and, contrary to the pigmented part of pe-nis, is whitish. On its top there is an irregular penialopening (Fig. 82). The penis and epiphallus pigmen-tation may be an additional diagnostic character todistinguish H. lutescens from the related H. pomatia inwhich the penis and epiphallus are always uniformlywhite. The penis, like the vagina, opens to the genitalatrium (Fig. 75g), which is the terminal section of the
226 Koralewska-Batura E.
Fig. 78. Spermatophore: a – head and neck, b – body, c – tail; 3×
Figs 79–82. Transverse sections through terminal portion of male genitalia: 79 – epiphallus, 38×; 80 – mid portion of penis,38×; 81 – base of papilla penis, 38×; 82 – terminal portion of penis, 38×

reproductive system. During copulation the penis andvagina of each partner are everted, and the penes areinserted into the vaginae simultaneously. This enablestransfer of sperm in spermatophores which leave thesnail’s body through the opening at the top of thepenial papilla.
B. DISTRIBUTION OF H. LUTESCENS IN POLAND
In Poland H. lutescens occurs only in thesouth-eastern part of the country: in Roztocze, the up-lands Wy¿yna Lubelska, Wy¿yna Malopolska, the low-land Nizina Sandomierska and the foothills of theBeskid Wschodni mts (up to ca. 300 m a.s.l.). In the firsthalf of this century, its distribution extended westwardto the line Kraków-Busko-Kielce-Skar¿ysko Kamienna-Kazimierz Dolny, and northward to the northern egde
of the upland Wy¿yna Lubelska (RIEDEL 1988). In Po-land H. lutescens reaches its north-western distributionborder. Below I list its up-dated localities in Poland (Fig.83), based on UTM grid system.
The following localities, listed in older literatureand then regarded as doubtful, have been recentlyconfirmed by me: Lubelskie voivodeship: FB 17Abramowice near Lublin, EB 68 Janowiec –xerothermic hills, EB 68 Kazimierz Dolny – cemetery,FB 30 Zwierzyniec – parks, gardens; Podkarpackievoivodeship: FA 32 vic ini ty of Przemyœl ;Œwiêtokrzyskie voivodeship: DB 73 Kielce – cemetery,gardens, shrubs at the foot of Wietrznia, DB 61Korytnica near Jêdrzejów, DB 60 xerothermic hills be-tween Piñczów and Skowronno.
Recently, BARGA-WIÊC£AWSKA (1989) has foundlocalities of H. lutescens in the Œwiêtokrzyski National
Helix lutescens 227
Fig. 83. Distribution of H. lutescens in Poland

Park and its protecting belt, i.e.: DB 93 £ysa Góra, EB03 Baszowice, EB 03 Góra Che³mowa, EB 03 Rudki,EB 03 Serwis D¹browa and DB 73 Kielce – cemeteryand gardens. Besides, PIECHOCKI (1990, 1991) liststhe following localities in the Roztoczañski NationalPark: FB 30 Zwierzyniec, FB 30 Tartaczna Góra nearZwierzyniec, FA 58 Rebizanty on the Tanew river, FA58 Rybnica, FA 69 Góra Wapielnia, FA 78 Machniównear Lubycz Królewska.
New records of the snail, found by me, are the fol-lowing: Lubelskie voivodeship: GB 03 Gródek –bushes at a field road to the river Huczwa, FB 45Suchodo³y – bushes in the park, FB 91 Tyszowce –cemetery; Ma³opolskie voivodeship: DA 36 Maszkównear S³omniki; Mazowieckie voivodeship: EB 45Czekarzewice – shrubs, EB 55 Józefów – bushes on theVistula river; Podkarpackie voivodeship: FA 02 Babice– San river escarpment, FA 32 Buszkowice – bushes onthe San river, FA 13 Bystrowice – roadside bushes, EA98 Bystre – bushes, FA 11 – bushes between Krzywczaand Ruszelczyce, EA 67 Nowa Wieœ Czudacka – busheson a small river, EA 52 Ubieszyn – bushes on the Sanriver, EA 64 Zwiêczyca – bushes; Œwiêtokrzyskievoivodeship: DB 73 Bia³ogon near Kielce –xerothermic grassland, DA 89 Busko Zdrój – gardens,nature reserve Zimne Wody of limestone-gypsum sub-stratum and steppe vegetation, DB 73 Kielce – areacovered with grass vegetation, DB 70 Kije – cemetery,EA 18 £ubnice – bushes, DB 60 Piñczów – sunnyslope, DA 89 Sies³awice near Busko Zdrój – steppevegetation on gypsum-limestone susbtratum, DB 96Skar¿ysko Kamienna – cemetery, bushes on a drain-age plot, DB 61 Sobków near Korytnica – cemetery,DB 60 Skowronno – roadside ditches, DA 99 Stopnica– garden, DA 89 Szczawory¿ – Garb Piñczowski, DB 80Œladków Ma³y – hills with rock outcrops, DA 79Winiary near Piñczów.
I have never found H. lutescens to co-occur with H.pomatia.
The new records of H. lutescens from Poland do notshift its distribution border given by RIEDEL (1988).The latter author cites URBAÑSKI (1977) who reportsthat the snail has gone or is going extinct near Kraków(DA 14); likewise, my own studies have not confirmedthe occurrence of H. lutescens in the region of Kraków.Contrary to what is reported by RIEDEL (1988), H.lutescens is not going extinct in the Œwiêtokrzyskie Mts;in anthropogenic sites it forms there abundant insu-lar populations (KORALEWSKA-BATURA 1993c,BARGA-WIÊC£AWSKA 1997). Likewise, the populationsof H. lutescens from Roztocze are not endangered.They are numerous and of high density (PIECHOCKI1990).
Because of its distribution (north-western distribu-tion border), insular occurrence and possibility of acci-dental collection for culinary purposes, H. lutescens islegally protected in Poland (WIKTOR & RIEDEL 1992).
C. BIOLOGY
1. Diurnal and seasonal activity of adult individuals
In the wild, H. lutescens is active only during thevegetation season. The snails spend the remainingpart of the year buried in the soil ca. 6 cm deep. Theirshells are then positioned with their apertures up-wards, the aperture being closed with the calcifiedepiphragm. At the end of April and in the first days ofMay the snails reject their epiphragms and leave theirwinter shelters. The activity of adult H. lutescens in-cludes seasonal behaviour: feeding, courtship andcopulation, egg-laying and preparations to hibernate.
During rest, at a considerable air humidity, thesnail’s head is retracted into the shell, and the foot ad-heres to the substratum. With increasing humidity thesnails may immediately get active. Low or high tem-peratures cause the snails to retract completely intothe shell and lie on the ground. Fairly often, duringdrought, the snails form a membrane of solidifiedmucus, which attaches them to the substratum. Atsuch times they do not react to tactile stimuli and areunable to regain activity immediately.
The diurnal activity of the snails in the spring ispresented in Figure 84. The vertical section denotesstandard deviation, for three observation sessions: onMay 11/12th (Fig. 84a), 25/26th (Fig. 84b) and June7/8th (Fig. 84c) 1995.
The curve of the diurnal activity indicates that inthe spring the activity of the snails starts with the in-crease in relative humidity. The further increase inthe proportion of active individuals is correlated withthe increase in relative humidity to 100%, which ismost often associated with a decrease in air tempera-ture in the ground layer to the so called dew point.According to my observations, this usually fell on thehours from 15.00 to 11.00 next day. The percentageof active snails was the highest between 5.00 and 9.00.At noon and in the afternoon, from 11.00 till 15.00, atan increase of temperature till 26°C and relative hu-midity of 74%, the snail activity was close to zero.
The considerable standard deviation from themean activity, between 17.00 and 3.00, is associatedwith rather widely varying weather conditions duringthe consecutive spring observations. Gusty winds andlow temperatures in the night considerably decreasethe snail activity (Fig. 84a). On a sunny day, a rapid in-crease in activity was associated with a passing rain ofonly several minutes duration (Fig. 84b). A gradualincrease in relative humidity results in a slow increasein activity which gets more intense at the dew point,already at a temperature of 15°C (Fig. 84c).
H. lutescens has no permanent shelter to which itwould return to rest; the snail ceases to be active andstays where it was during the final phase of its activity.However, the preferable resting places change sea-
228 Koralewska-Batura E.
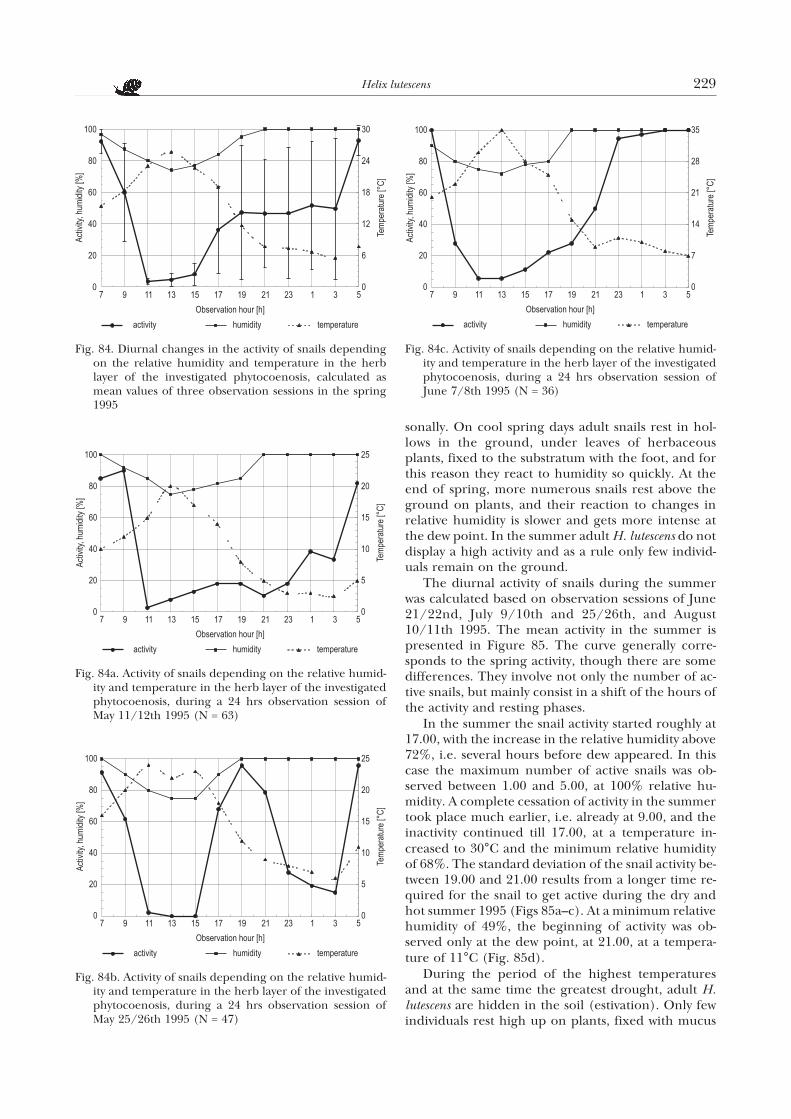
sonally. On cool spring days adult snails rest in hol-lows in the ground, under leaves of herbaceousplants, fixed to the substratum with the foot, and forthis reason they react to humidity so quickly. At theend of spring, more numerous snails rest above theground on plants, and their reaction to changes inrelative humidity is slower and gets more intense atthe dew point. In the summer adult H. lutescens do notdisplay a high activity and as a rule only few individ-uals remain on the ground.
The diurnal activity of snails during the summerwas calculated based on observation sessions of June21/22nd, July 9/10th and 25/26th, and August10/11th 1995. The mean activity in the summer ispresented in Figure 85. The curve generally corre-sponds to the spring activity, though there are somedifferences. They involve not only the number of ac-tive snails, but mainly consist in a shift of the hours ofthe activity and resting phases.
In the summer the snail activity started roughly at17.00, with the increase in the relative humidity above72%, i.e. several hours before dew appeared. In thiscase the maximum number of active snails was ob-served between 1.00 and 5.00, at 100% relative hu-midity. A complete cessation of activity in the summertook place much earlier, i.e. already at 9.00, and theinactivity continued till 17.00, at a temperature in-creased to 30°C and the minimum relative humidityof 68%. The standard deviation of the snail activity be-tween 19.00 and 21.00 results from a longer time re-quired for the snail to get active during the dry andhot summer 1995 (Figs 85a–c). At a minimum relativehumidity of 49%, the beginning of activity was ob-served only at the dew point, at 21.00, at a tempera-ture of 11°C (Fig. 85d).
During the period of the highest temperaturesand at the same time the greatest drought, adult H.lutescens are hidden in the soil (estivation). Only fewindividuals rest high up on plants, fixed with mucus
Helix lutescens 229
Fig. 84. Diurnal changes in the activity of snails dependingon the relative humidity and temperature in the herblayer of the investigated phytocoenosis, calculated asmean values of three observation sessions in the spring1995
Fig. 84a. Activity of snails depending on the relative humid-ity and temperature in the herb layer of the investigatedphytocoenosis, during a 24 hrs observation session ofMay 11/12th 1995 (N = 63)
Fig. 84b. Activity of snails depending on the relative humid-ity and temperature in the herb layer of the investigatedphytocoenosis, during a 24 hrs observation session ofMay 25/26th 1995 (N = 47)
Fig. 84c. Activity of snails depending on the relative humid-ity and temperature in the herb layer of the investigatedphytocoenosis, during a 24 hrs observation session ofJune 7/8th 1995 (N = 36)
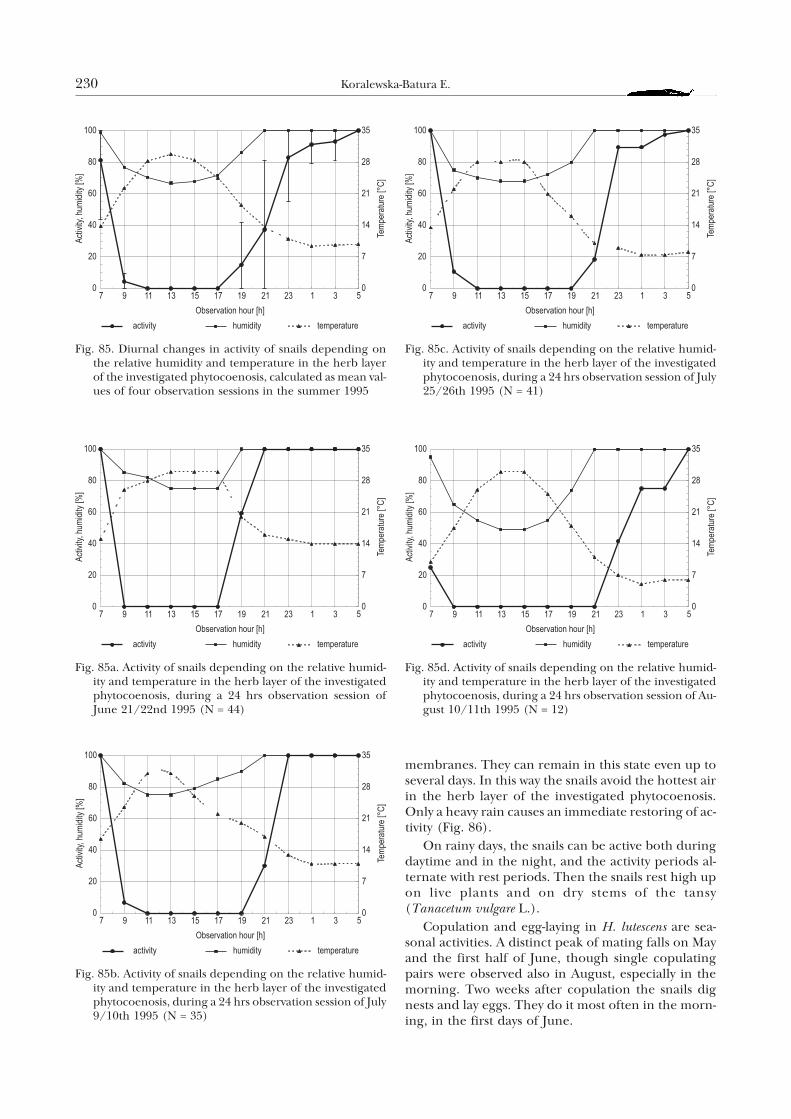
membranes. They can remain in this state even up toseveral days. In this way the snails avoid the hottest airin the herb layer of the investigated phytocoenosis.Only a heavy rain causes an immediate restoring of ac-tivity (Fig. 86).
On rainy days, the snails can be active both duringdaytime and in the night, and the activity periods al-ternate with rest periods. Then the snails rest high upon live plants and on dry stems of the tansy(Tanacetum vulgare L.).
Copulation and egg-laying in H. lutescens are sea-sonal activities. A distinct peak of mating falls on Mayand the first half of June, though single copulatingpairs were observed also in August, especially in themorning. Two weeks after copulation the snails dignests and lay eggs. They do it most often in the morn-ing, in the first days of June.
230 Koralewska-Batura E.
Fig. 85. Diurnal changes in activity of snails depending onthe relative humidity and temperature in the herb layerof the investigated phytocoenosis, calculated as mean val-ues of four observation sessions in the summer 1995
Fig. 85a. Activity of snails depending on the relative humid-ity and temperature in the herb layer of the investigatedphytocoenosis, during a 24 hrs observation session ofJune 21/22nd 1995 (N = 44)
Fig. 85b. Activity of snails depending on the relative humid-ity and temperature in the herb layer of the investigatedphytocoenosis, during a 24 hrs observation session of July9/10th 1995 (N = 35)
Fig. 85c. Activity of snails depending on the relative humid-ity and temperature in the herb layer of the investigatedphytocoenosis, during a 24 hrs observation session of July25/26th 1995 (N = 41)
Fig. 85d. Activity of snails depending on the relative humid-ity and temperature in the herb layer of the investigatedphytocoenosis, during a 24 hrs observation session of Au-gust 10/11th 1995 (N = 12)
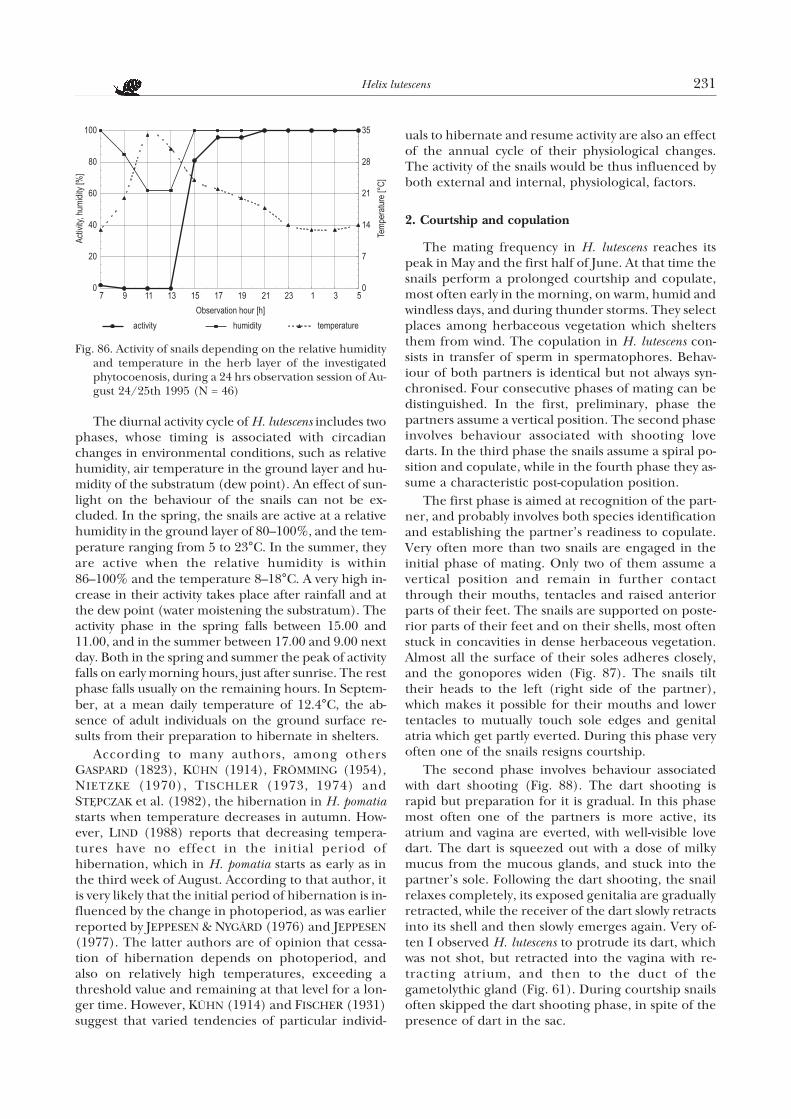
The diurnal activity cycle of H. lutescens includes twophases, whose timing is associated with circadianchanges in environmental conditions, such as relativehumidity, air temperature in the ground layer and hu-midity of the substratum (dew point). An effect of sun-light on the behaviour of the snails can not be ex-cluded. In the spring, the snails are active at a relativehumidity in the ground layer of 80–100%, and the tem-perature ranging from 5 to 23°C. In the summer, theyare active when the relative humidity is within86–100% and the temperature 8–18°C. A very high in-crease in their activity takes place after rainfall and atthe dew point (water moistening the substratum). Theactivity phase in the spring falls between 15.00 and11.00, and in the summer between 17.00 and 9.00 nextday. Both in the spring and summer the peak of activityfalls on early morning hours, just after sunrise. The restphase falls usually on the remaining hours. In Septem-ber, at a mean daily temperature of 12.4°C, the ab-sence of adult individuals on the ground surface re-sults from their preparation to hibernate in shelters.
According to many authors, among othersGASPARD (1823), KÜHN (1914), FRÖMMING (1954),NIETZKE (1970), TISCHLER (1973, 1974) andSTÊPCZAK et al. (1982), the hibernation in H. pomatiastarts when temperature decreases in autumn. How-ever, LIND (1988) reports that decreasing tempera-tures have no effect in the initial period ofhibernation, which in H. pomatia starts as early as inthe third week of August. According to that author, itis very likely that the initial period of hibernation is in-fluenced by the change in photoperiod, as was earlierreported by JEPPESEN & NYGÄRD (1976) and JEPPESEN(1977). The latter authors are of opinion that cessa-tion of hibernation depends on photoperiod, andalso on relatively high temperatures, exceeding athreshold value and remaining at that level for a lon-ger time. However, KÜHN (1914) and FISCHER (1931)suggest that varied tendencies of particular individ-
uals to hibernate and resume activity are also an effectof the annual cycle of their physiological changes.The activity of the snails would be thus influenced byboth external and internal, physiological, factors.
2. Courtship and copulation
The mating frequency in H. lutescens reaches itspeak in May and the first half of June. At that time thesnails perform a prolonged courtship and copulate,most often early in the morning, on warm, humid andwindless days, and during thunder storms. They selectplaces among herbaceous vegetation which sheltersthem from wind. The copulation in H. lutescens con-sists in transfer of sperm in spermatophores. Behav-iour of both partners is identical but not always syn-chronised. Four consecutive phases of mating can bedistinguished. In the first, preliminary, phase thepartners assume a vertical position. The second phaseinvolves behaviour associated with shooting lovedarts. In the third phase the snails assume a spiral po-sition and copulate, while in the fourth phase they as-sume a characteristic post-copulation position.
The first phase is aimed at recognition of the part-ner, and probably involves both species identificationand establishing the partner’s readiness to copulate.Very often more than two snails are engaged in theinitial phase of mating. Only two of them assume avertical position and remain in further contactthrough their mouths, tentacles and raised anteriorparts of their feet. The snails are supported on poste-rior parts of their feet and on their shells, most oftenstuck in concavities in dense herbaceous vegetation.Almost all the surface of their soles adheres closely,and the gonopores widen (Fig. 87). The snails tilttheir heads to the left (right side of the partner),which makes it possible for their mouths and lowertentacles to mutually touch sole edges and genitalatria which get partly everted. During this phase veryoften one of the snails resigns courtship.
The second phase involves behaviour associatedwith dart shooting (Fig. 88). The dart shooting israpid but preparation for it is gradual. In this phasemost often one of the partners is more active, itsatrium and vagina are everted, with well-visible lovedart. The dart is squeezed out with a dose of milkymucus from the mucous glands, and stuck into thepartner’s sole. Following the dart shooting, the snailrelaxes completely, its exposed genitalia are graduallyretracted, while the receiver of the dart slowly retractsinto its shell and then slowly emerges again. Very of-ten I observed H. lutescens to protrude its dart, whichwas not shot, but retracted into the vagina with re-tracting atrium, and then to the duct of thegametolythic gland (Fig. 61). During courtship snailsoften skipped the dart shooting phase, in spite of thepresence of dart in the sac.
Helix lutescens 231
Fig. 86. Activity of snails depending on the relative humidityand temperature in the herb layer of the investigatedphytocoenosis, during a 24 hrs observation session of Au-gust 24/25th 1995 (N = 46)

232 Koralewska-Batura E.
Figs 87–92. Courtship and copulation: 87 – phase I, vertical position; 88 – phase II, behaviour associated with love dart shoot-ing; 89, 90 – phase III, spiral position and copulation; 91, 92 – phase IV, post-copulation position

In the third phase, the snails assume a spiral posi-tion (Fig. 89). That phase is preceded by resuming ofvertical position by both partners, and then repeatedtilting of the heads to the left and twisting anteriorbody parts clockwise, by 90°, the posterior sole parttightly adjoining the partner’s sole. The head tiltingand body twisting initially enables touching of thepartner’s exposed atrium with the lip and short righttentacle, till strongly everted genitalia touch eachother. In the spiral phase the snails make attempts atcopulation. However, for the copulation to be accom-plished it is necessary for penes and vaginae to geteverted; then the penes are mutually inserted into thevaginae (Fig. 90) which makes it possible to transferspermatophores. In H. lutescens the copulation (trans-fer of spermatophores) lasts ca. 15 min, then theeverted genitalia are retracted; in H. pomatia thecopulation lasts only 5 minutes (LIND 1976). Very of-ten attempts at copulation fail. The copulation is in-terrupted when both partners fail to insert theirpenes simultaneously and when the everting penes ofthe partners are misdirected. Following such failed at-tempts, the everted genitalia are quickly retracted,and in a short time another attempt is made.
Following copulation, the fourth phase starts dur-ing which the snails remain immobile and assume apost-copulation position. In this position the wholesnail bodies contact except the heads which are nowmoved away from the partner. The soles become verywide and motionless, directed upwards and togetherforming a cup (Figs 91–92). Perhaps in this phase apart of sperm is released from the wholly transferredspermatophores, the sperm migrating then to the ovi-ducts and to seminal vesicles. The remnants of sper-matophores get to the gametolythic gland where they
are disintegrated and digested. In H. lutescens thefourth phase lasts ca. 2 hrs, and the whole mating pro-cess ca. 3 hrs, depending on thermal conditions. Theduration of the first two phases is very much variable.
3. Egg laying and hatching
Two weeks after copulation H. lutescens starts nestdigging and egg laying. The intensity of these activities isthe highest in June, the are sporadic in the first days ofJuly. A high air humidity, preceded by a short period ofintense rain, creates very good conditions for nest dig-ging. The nests are located in places with loose soil andsparse vegetation, most often under the plants of tansy(Tanacetum vulgare L.) and in sheltered places. In suchplaces the nest digging snails tend to gather in groups ofseveral (Fig. 93). This results from a particularly goodstructure of the soil. The nest depth from the bottom tothe soil surface is 4–6 cm and depends on the kind ofsubstratum. During the egg laying, the snail buries itsbody in the nest, so that its shell is completely coveredby the excavated soil (Fig. 94).
The eggs of H. lutescens (Figs 95–96) are spherical,with a pearl-like sheen, 4.0–4.5 mm in diameter.Their outer envelope is very delicate and calcified.The eggs are laid together with mucus, in groups of afew or about a dozen. The high viscosity of the mucusresults in the eggs in the nest being arranged reg-ularly, forming a cluster which tightly adjoins one ofthe nest walls. H. lutescens lays from 16 to 67 eggs(KARSZNIA 1997), and on an average 35 eggs per nest.Following the egg laying, the snail leaves the nest,closes its with mucus-glued soil and evens the soil sur-face thus masking the nest. The process lasts from twoto three days.
Helix lutescens 233
Figs 93, 94. Egg laying: 93 – distribution of clutches in the study plot; 94 – nest digging

234 Koralewska-Batura E.
Figs 95, 96. Egg laying: 95 – arrangement of eggs in the nest; 96 – eggs taken out of the nest, ca. 3×
Fig. 97. Hatching, ca. 3×

Helix lutescens 235
Fig. 98. Hatchlings three days old, ca. 3×
Fig. 99. Juveniles upon leaving their nest, ca. 3×

In developing eggs of H. lutescens, already on the14th day, shells of juvenile snails can be seen shiningthrough, and the eggs become light yellow (Fig. 97).Young snails leave the egg shell thanks to its gradualresorption. The newly hatched snails stay in the nestand feed on the remnants of egg shells (Fig. 98).Earlier hatched juveniles may consume eggs andsmaller juveniles; they eat also dead eggs. The eggcannibalism is common in some helicids, e.g. H.pomatia and Arianta arbustorum (L.) (BAUR 1988,1990). From 14 to 16 days after hatching the snailsemerge from the nest to the soil surface (Fig. 99). Infavourable humidity conditions I found them on thesoil as early as in the first days of July. The develop-ment of earlier laid eggs (first days of June) is fa-voured by the high humidity in the nest, remainingafter winter. The mean mortality of eggs is 48.5%(KARSZNIA 1997). A drought preceding reproductiondecreases chances for hatching, increasing the meanmortality of eggs up to over 70% (KARSZNIA 1997).The effect of humidity on egg development was ob-served also by POLLARD (1975) in H. pomatia. I havefound that one of the reasons for destruction of wholeclutches in H. lutescens is an excess of water in the sub-stratum; then the egg shells break.
Natural enemies of H. lutescens and its eggs are antsand earthworms. The latter dig the eggs out which re-sults in their drying. Ants attack and destroy eggs thatwere laid near ant hills.
D. ECOLOGY
1. Abundance and density
The mean abundance, its extreme values, as well asmean densities of the three age classes of H. lutescenson the study plot of 500 m2, are presented in Table 1.The density of individuals aged one year and younger(just after leaving their nests), of shell diameter up to10 mm and little advanced development of genitalia,is the highest. As a result of mortality of consecutiveage classes, their number gradually decreases.
Young individuals display a much lower density, onan average 0.82 individual per m2. It seems likely thatthe snails become sexually mature in the fourth yearof their life. Since adult individuals live a few years,this age class includes adults of various generationsi.e. of varied age. Most probably this accounts for thehigh density of adults (1.15 indiv./m2).
2. Changes in population abundance
Seasonal changes in abundance of the three ageclasses of H. lutescens are presented in Figure 100.
In the control plot I took the first sample (in themodel – sample i: 0) on May 15th 1994. I found andmarked 236 adult, 142 young (two years and older)individuals and 126 yearlings. The analysis of the datawith JOLLY-SEBER‘S model indicates that the peak ofpopulation abundance fell on June and persisted tillthe end of July, with a marked decrease at half of Sep-tember. Adult individuals were the most numerous inJune. This resulted no doubt from their intense activ-ity, which consisted in searching for a mating partner,copulation, selecting places for nest digging and egglaying. The decrease in abundance of adults andyoung in July was associated with drought. It can besupposed that in the study period high temperaturesaccompanied by drought, which persisted till half ofAugust, caused adult individuals to hide in the soil.The abundance of yearlings, which was high alreadyin June, reached its peak at half of July, thanks toemergence of juveniles (less than one year) from thenests. Already in August, quickly growing yearlings in-creased the abundance of the class of young snails. Atthe same time, the long draught resulted in a highmortality of snails of all the age classes. Yearlings andindividuals of less than one year were active till half ofSeptember, when adult snails had already retired totheir winter shelters.
236 Koralewska-Batura E.
Table 1. Mean abudance and density of the three ageclasses in H. lutescens on the study plot
Age classMean
abudance+SE
Range(min–max)
Meandensityper m2
adult 573.3+124.4 329–817 1.15
young(2 years
and older)408.2+133.5 147–670 0.82
1 year andyounger
1,988.8+946.3 134–3,844 3.98
Fig. 100. Abundance of the three age classes of H. lutescenson the study plot during consecutive controls in 1994(control: i = 0 of May 1994, whose values are given in thetext, has been omitted from the graph)

CONCLUDING REMARKS
1. Helix lutescens Rossm. is a xero-thermophilous spe-cies. It inhabits steppe habitats, dry shrubs, grass-lands, and sunny slopes, especially on calcareoussubstratum; it is often found in gardens and ceme-teries. I have not found it together with H. pomatiain any of the studied localities. In Poland H. lute-scens is found only in the south-eastern part of thecountry, where it reaches its north-western dis-tribution border.
2. The shell of H. lutescens is spherical-conical, yellow-ish-white, of 4–4.5 poorly convex whorls separatedby a shallow suture. It is more conical than in H.pomatia. The shell measurements in adults are28.5–34.1 height, 26.9–32.1 width and 22.5–26.9diameter, thus being considerably smaller than inH. pomatia. In adult H. lutescens the umbilicus iscompletely covered, while in H. pomatia it is partlyopen. The embryonic shell sculpture differs fromthat of definitive whorls, which consists of growthlines of various width and delicate spiral lines, re-sulting in a grating-like pattern.
3. The body of H. lutescens is greyish and coveredwith numerous grooves and wrinkles; in thegrooves there are fine, whitish grains, which arethe most numerous in the grooves forming twocharacteristic light streaks along the darker back.The character is species-specific.
4. In the reproductive system of H. pomatia it is note-worthy that the spermatheca duct never bears adiverticle, which is present in H. pomatia. Be-sides, the delicate pigmentation of the flagellumand intense pigmentation of the epiphallus andpenis in adult H. lutescens is an additional di-agnostic character to distinguish it from H. po-matia whose corresponding organs are alwayspure white.
5. The activity of adult H. lutescens is strictly associ-ated with seasonal behaviour: crawling, feeding,mating and egg laying, as well as preparing for hi-bernation. The diurnal activity of H. lutescens in-cludes two phases, their timing depending onseasonal fluctuations of environmental factors. Inthe spring, the snails are active at a relative humi-dity from 80 to 100% and a temperature of5–23°C. In the summer they are active when therelative humidity is within the range of 86–100%and the temperature – 8–18°C. Their activity in-creases very much especially after a rain or at thedew point. The active phase in the spring falls be-tween 15.00 and 11.00, and in the summer be-tween 17.00 and 9.00 next day. Both in the springand summer the peak of activity falls on earlymorning hours, just after sunrise. The restingphase usually includes the remaining hours. InSeptember, at a mean diurnal temperature of12.4oC, on the soil surface there are no adult in-
dividuals which results from their preparation tohibernate in winter shelters.
6. The highest frequency of mating in H. lutescens wasobserved in May and the first half of June. Fourconsecutive mating phases can be distinguished.The first, initial, phase involves probably recogni-tion of the partner’s species identity and its readi-ness to mate. During that phase the partners assu-me a vertical position and remain in contact. Thesecond phase includes behaviour associated withlove dart shooting, but this is often not accom-plished, in spite of the presence of the dart in thesac. In the third phase, the snails assume a spiralposition and copulate. The sperm is transferred inspermatophores and the process lasts ca. 15 min.Following sperm transfer, the fourth phase beginsduring which the snails remain motionless. Thelast phase lasts ca. 2 hrs, and the whole mating ta-kes over 3 hrs and depends on temperature.
7. Two weeks from mating H. lutescens starts diggingits nests and laying eggs. The eggs are spherical, ofpearl-like lustre and 4.0–4.5 mm in diameter. Thesnail lays from 16 to 67 eggs, on an average 35 eggsper nest. In developing eggs, already 14 days fromegg laying, shells of juvenile snails shine through.The juveniles leave the egg envelopes resorbingthem gradually. The hatchlings remain in the nestand leave them most often in 14–16 days.
8. The mean densities of the three age classes of H.lutescens found in the constant study plot were thefollowing: 1.15 individuals/m2 (adults), 0.82 in-div./m2 (two years and older) and 3.98 indiv./m2
(one year and younger). The population abun-dance reached its peak in June and persisted tillhalf of July, and a distinct decrease was observed athalf of September. The adults were the most nu-merous in June. The abundance of the youngestage class, which was high already in June, reachedits peak at half of July, due to the emergence of ha-tchlings from the nests. The individuals aged oneyear and younger, as well as few young individuals,were active till half of September when the adultshad already sheltered for winter.
9. The new localities of H. lutescens in Poland do notchange the distribution limits of the species repor-ted by RIEDEL (1988). Contrary to RIEDEL‘S (1988)data, the species is not going extinct in the Œwiêto-krzyskie Mts: in anthropogenic sites it forms abun-dant insular populations (KORALEWSKA-BATURA1993c, BARGA-WIÊC£AWSKA 1997). Likewise, theRoztocze populations of H. lutescens are not endan-gered (PIECHOCKI 1990). In Poland, because of thenorth-western distribution border and the insularoccurrence, as well as threat of unintentional collec-ting for culinary purposes, H. lutescens is a protectedspecies (WIKTOR & RIEDEL 1992).
Helix lutescens 237

REFERENCES
ABOLINS-KROGIS A. 1961. The histochemistry of the hepato-pancreas of Helix pomatia (L.) in relation to the regener-ation of the shell. Ark. Zool. 13: 159–201.
ABOLINS-KROGIS A. 1965. Electron microscope observationson cells in the hepatopancreas of the snail Helix pomatia(L.). Ark. Zool. 18: 85–92.
ABOLINS-KROGIS A. 1970a. Electron microscope studies ofthe intracellular origin and formation of calcifying gran-ules and calcium spherites in the hepatopancreas of thesnail Helix pomatia (L.). Z. Zellforsch. 108: 501–515.
ABOLINS-KROGIS A. 1970b. Alterations in the fine structureof cytoplasmic organelles in the hepatopancreatic cellsof shell-regenerating snail, Helix pomatia (L.). Z. Zell-forsch. 108: 516–529.
ADAMO SH. A., CHASE R. 1990. The “love dart” of the snailHelix aspersa injects a pheromone that decreases court-ship duration. J. Exp. Zool. 225: 80–87.
BABULA A., SKOWROÑSKA-WENDLAND D. 1988. Histologicaland histochemical studies of the digestive system of theslug Deroceras reticulatum (Müller) (Pulmonata). Bull.Soc. Amis Sci. Lett. Poznañ Ser. D biol. 26: 65–71.
BABULA A., WIELIÑSKA L. 1988. Ultrastructual studies of thedigestive system of the slug Deroceras reticulatum (Müller)(Pulmonata). Bull. Soc. Amis Sci. Lett. Poznañ Ser. Dbiol. 26: 73–78.
BARGA-WIÊC£AWSKA J. 1989. Helix lutescens Rossmässler, 1837(Gastropoda: Helicidae) in the Holy Cross MountainsNational Park and its vicinity. Folia Malacologica 3:23–33.
BARGA-WIÊC£AWSKA J. 1997. Sukcesja œlimaków na ha³dachregionu Œwiêtokrzyskiego. Wydawnictwo WSP, Kielce.
BAUR B. 1988. Egg-species recognition in cannibalistichatchlings of the land snails Arianta arbustorum and Helixpomatia. Experientia 44: 276–277.
BAUR B. 1990. Possible benefits of egg cannibalism in theland snail Arianta arbustorum (L.). Functional Ecology 4:679–684.
B¥KOWSKI J., £OMNICKI A. M. 1892. Miêczaki (Mollusca).Muz. im. Dzieduszyckich we Lwowie. III. Lwów.
BORNCHEN M. 1967. Untersuchungen zur Sekretion der fin-gerformigen Drusen von Helix pomatia. Z. Zellforsch. Mi-krosk. Anat. 78: 402–426.
BREUCKER H. 1964. Cytologische Untersuchungen des Zwit-terganes und des Spermatoviduktes von Helix pomatia.Protoplasma 58: 1–41.
CHETAIL M., KRAMPITZ G. 1982. Calcium and skeletal struc-tures in molluscs: concluding remarks. Malacologia 22:337–339.
CHUNG D. J. D. 1986. Stimulation of genital eversion in theland snail Helix aspersa by extracts of the glands of thedart apparatus. J. Exp. Zool. 238: 129–139.
CHUNG D. J. D. 1987. Courtship and dart shooting behaviorof the land snail, Helix aspersa. Veliger 30: 24–39.
CZUBIÑSKI Z., URBAÑSKI J. 1933. Szcz¹tki zespo³ów pontyj-skich na Wietrzni ko³o Kielc. Ochrona Przyrody 13:186–188.
DILLAMAN R. M. 1981. Dart formation in Helix aspersa (Mol-lusca, Gastropoda). J. Zoomorphol. 97: 247–261.
DORELLO P. 1925. Sulla funzione delle glandole digitate nelgen. Helix. Atti della Reale Academia Nazionale dei Lin-cei. Ser. 6. Rendiconti 1: 47–51.
DROZDOWSKI A. 1970. Vergleichende Studien über die Lun-genmorphologie bestimmter Schneckenarten (Gastro-poda, Pulmonata). Zool. Pol. 20: 217–265.
DROZDOWSKI A. 1977. Versuch einer Analyse der entwick-lungsgeschichtlichen Veränderungen der Schnecken-lungen (Gastropoda, Pulmonata). Malak. Abh. 5:203–212.
DROZDOWSKI A. 1986. Histologische Untersuchungen derLungenhemalsystem von Helix pomatia L. (Gastropoda,Pulmonata, Helicidae). Malak. Abh. 11: 167–178.
DROZDOWSKI A. 1993. Badania nad struktur¹ histologiczn¹centralnego uk³adu hemalnego Limax flavus L. (Gastro-poda, Pulmonata). Acta Univ. N. Copern. (Biol. 45) 87:71–79.
DROZDOWSKI A., ZAWIEJA E. 1993. Studium nad sezonow¹zmiennoœci¹ budowy przewodu pokarmowego u wybra-nych gatunków Pulmonata. Acta Univ. N. Copern. (Biol.45) 87: 33–59.
EHRMANN P. 1933. Mollusken (Weichtiere). In: Die TierweltMitteleuropas (BROHMER P. & ULMER G., eds), vol. 2(1),Leipzig.
FEDOSEEVA E. A. 1994. Comparative morphology of darts inthe superfamily Helicoidea (Gastropoda, Pulmonata).Ruthenica 4: 103–110.
FISCHER P. H. 1931. Recherches sur la vie ralentie de l’escar-got (Helix pomatia L.). Thesis Faculté des Sciences, Paris.
FRÖMMING E. 1954. Biologie der mitteleuropäischen Land-gastropoden. Duncker & Humbolt, Berlin.
GASPARD B. 1823. Beiträge zur Physiologie der Garten-schnecke (Helix pomatia L.). Deutsches Arch. f. Physiol.8: 243–290.
GHOSE K. CH. 1962. Origin and development of the dige-stive system of the giant land snail Achatina fulicaBowdich. Proc. R. Soc. Edinb. 68 B: 186–207.
GIUSTI F., LEPRI A. 1980. Aspetti morphologici ed etologicidell’accoppiamento in alcune specie della famiglia Heli-cidae. IV Congresso della Societa Malacologica Italiana,Accademia delle Scienze di Siena detta de’Fisiocritici.Atti Accademia Fisiocririci. Siena: 11–71.
GÓMEZ B. J., SERRANO T., ANGULO E. 1996. Morphologyand fine structure of glands of the dart-sac complex inHelicoidea (Gastropoda, Stylommatophora). Invertebr.Reprod. Develop. 29: 47–55.
GOUDSMIT E. M. 1975. Neurosecretory stimulation of galac-togen synthesis within the Helix pomatia albumen glandduring organ culture. J. Exp. Zool. 191: 193–198.
GÖTTING K. J. 1974. Malakozoologie. Grundriss der Weich-tierkunde. Gustav Fischer Verl., Stuttgart.
GRÉGOIRE C. 1972. Structure of the molluscan shell. In: Che-mical Zoology, vol. 7 Mollusca (FLORKIN M. & SCHEER B.T., eds), pp. 45–102. Acad. Press, New York & London.
238 Koralewska-Batura E.

GROCHMALICKI J. 1932. Ochrona zwierz¹t ni¿szych. In: Skar-by Przyrody i ich ochrona, pp. 108–116, Warszawa.
GROSSU A. V. 1955. Fauna Republicii Populare Romíne.Mollusca. vol. 3 (1). Gastropoda Pulmonata. vol. 3 (1).Edit. Acad. Republicii Populare Romine, Bucuresti.
GROSSU A. V. 1983. Gastropoda Romaniae. Ordo Stylomma-tophora. Suprafam: Arionacea, Zonitacea, Ariophanta-cea si Helicacea. vol. 4. Editura Litera, Bucuresti.
HELLER J., MAGARITZ M. 1983. From where do land snailsobtain the chemicals to build their shells? J. Moll. Stud.49: 116–121.
HRYNIEWIECKA-SZYFTER Z. 1966. Copper in hepatopancreasof the snail Helix pomatia (L.). Bull. Soc. Amis Sci. Lett.Poznañ Ser. D. biol. 8: 119–123.
HRYNIEWIECKA-SZYFTER Z., RÊDZINIAK E. 1976. The localiz-ation of the lysosomal enzymes in the bursa copulatrix ofthe snail Helix pomatia L. Bull. Soc. Amis Sci. Lett. Po-znañ Ser. D. 16: 125–134.
HUNT S. 1979. The structure and composition of the lovedart (gypsobelum) in Helix pomatia. Tissue & Cell 11:51–61.
JACKIEWICZ M., KORALEWSKA-BATURA E. 1999. Pneumosto-me structure in Helix pomatia Linnaeus, 1758 and H. lute-scens Rossmässler, 1837 (Gastropoda: Pulmonata: Helici-dae) – another diagnostic character. Folia Malacologica7: 43–45.
JEPPESEN L. L. 1976. The control of mating behaviour in He-lix pomatia L. (Gastropoda: Pulmonata). Anim. Behav.24: 275–290.
JEPPESEN L. L., NYGÄRD K. 1976. The influence of photo-period, temperature and internal factors on the hiber-nation of Helix pomatia L. (Gastropoda, Pulmonata). Vi-densk. Meddr dansk naturh. Foren. 139: 7–20.
JEPPESEN L. L. 1977. Photoperiodic control of hibernationin Helix pomatia L. (Gastropoda: Pulmonata). Behav. Pro-cesses 2: 373–383.
JOLLY G. M. 1965. Explicit estimates from capture-recapturedata with both death and immigration-stochastic model.Biometrika 52: 225–247.
JUNGBLUTH J. H., LIKHAREV I. M., WIKTOR A. 1985. Verglei-chend morphologische Untersuchungen an der Radulader Landnacktschnecken. II. Arionoidea und Trigono-chlamydoidea (Gastropoda: Pulmonata). Arch. Moll.116: 25–45.
KARSZNIA I. 1997. Biologia rozrodu œlimaka ¿ó³tawego (Helixlutescens Rossm.) (diploma thesis). Bibl. Zak³. ZoologiiWSP, Kielce.
KERNEY M. P., CAMERON R. A. D., JUNGBLUTH J. H. 1983.Die Landschnecken Nord- und Mitteleuropas. Paul Pa-rey Verlag, Hamburg & Berlin.
KILIAS R. 1960. Weinbergschnecken. Ein Überblick überihre Biologie und wirtschaftliche Bedeutung. Veb Deu-tscher Verlag der Wissenschaften, Berlin.
KILIAS R. 1985. Die Weinbergschnecke – über Leben undNutzung von Helix pomatia. Die Neue Brehm-Bücherei,Wittenberg Lutherstadt.
KORALEWSKA-BATURA E. 1993a. Charakterystyka biometrycz-na skorupki œlimaka ¿ó³tawego (Helix lutescens Rossm.) wPolsce. Przegl. Zool. 37: 283–289.
KORALEWSKA-BATURA E. 1993b. Die Struktur der Liebes-pfeile der Gattung Helix Linnaeus (Gastropoda: Pulmo-nata: Helicidae). Folia Malacologica 5: 115–118.
KORALEWSKA-BATURA E. 1993c. Nowe dane o wystêpowaniuHelix lutescens Rossmässler, 1837 (Gastropoda: Pulmona-ta) w Polsce. Przegl. Zool. 37: 69–72.
KORALEWSKA-BATURA E. 1994a. Development of reproduc-tive organs of Helix lutescens Rossm. (Gastropoda, Pulmo-nata). Biol. Bull. Poznañ 31: 67–70.
KORALEWSKA-BATURA E. 1994b. Der Bau der Radula beieinigen Arten der Familie Helicidae aus Polen (Gastro-poda: Pulmonata). Biol. Bull. Poznañ 31: 71–82.
KORALEWSKA-BATURA E. 1997a. Analysis of lung haemal sys-tem in Helix lutescens Rossm. (Gastropoda, Pulmonata).Biol. Bull. Poznañ 34: 69–72.
KORALEWSKA-BATURA E. 1997b. Sinistrally coiled specimenof Helix lutescens Rossmässler, 1837 (Gastropoda: Stylom-matophora: Helicidae). Malak. Abh. 18: 233–237.
KÜHN W. 1914. Beiträge zur Biologie der Weinbergschnecke(Helix pomatia L.). Z. wiss. Zool. 109: 128-184.
LIKHAREV I. M., RAMMELMEIER E. C. 1952. Nazemnyemollyuski fauny SSSR. Izd. Akad. Nauk SSSR, Moskwa –Leningrad.
LIND H. 1973. The functional significance of the sperma-tophore and the fate of spermatozoa in the genital tractof Helix pomatia (Gastropoda: Stylommatophora). J.Zool. 169: 39–64.
LIND H. 1976. Causal and functional organization of the ma-ting behavior sequence in Helix pomatia. Behaviour 59:162–202.
LIND H. 1988. The behaviour of Helix pomatia L. (Gastropo-da, Pulmonata) in a natural habitat. Vidensk. Meddr.dansk naturh. Foren. 147: 67–92.
LISICKÝ M. J. 1991. Mollusca Slovenska. Vyd. Slov. akad. vied,Bratislava.
LOZEK V. 1956. Klíè Èeskoslovenských mékkýšù. Bratislava.
LOZEK V. 1964. Quartämollusken der Tschechoslowakei.Rozpr. Ústr. Úst. Geol. 31. Praha.
MEISENHEIMER J. 1912. Die Weinbergschnecke Helix pomatiaL. Monographien einheimischer Tiere. Band 4. Verlagvon Dr. Werner Klinkhardt, Leipzig.
NIELAND M., GOUDSMIT E. M. 1969. Ultrastructure of galac-togen in the albumen gland of Helix pomatia. J. Ultra-struct. Res. 29: 119–140.
NIETZKE G. 1970. Die Weinbergschnecke. Verlag Eugen Ul-mer, Stuttgart.
OKO£OWICZ W., MARTYN D. 1979. Regiony klimatyczne Pol-ski. In: Atlas geograficzny, PPWK, Warszawa.
OWEN G. 1966. Digestion. In: Physiology of Mollusca (WIL-BUR K. M. & YONGE C. M., eds), vol. 2, pp. 53–96, Acade-mic Press, New York & London.
PIECHOCKI A. 1979. Miêczaki (Mollusca). Œlimaki (Gastropo-da). Fauna S³odkowodna Polski 7. PWN, Poznañ.
PIECHOCKI A. 1990. Interesuj¹ce gatunki œlimaków l¹dowych(Gastropoda terrestria) na Roztoczu. Przegl. Zool. 34:237–247.
PIECHOCKI A. 1991. Terrestrial snails of the RoztoczeUpland (SE Poland): zoogeographical considerations.
Helix lutescens 239

Proc. Tenth Intern. Malacol. Congr. Tübingen 1989:469–470.
POKORA Z. 1989. Hormonalna kontrola procesów rozrod-czych u œlimaków p³ucodysznych. II. Stylommatophora.Przegl. Zool. 33: 207–229.
POLIÑSKI W. 1917. Materia³y do fauny malakozoologicznejKrólestwa Polskiego, Litwy i Polesia. Pr. TN Warsz. 27:1–130.
POLIÑSKI W. 1919. Rozsiedlenie geograficzne Helicidów wPolsce. Szkic zoogeograficzny, z mapk¹. Przegl. Geogr. 1:269–280.
POLIÑSKI W. 1924. Anatomisch-systematische und zoogeo-graphische Studien über die Heliciden Polens. Bull.Acad. Pol. Sci. Lett., Cl. Sci. Math. Nat., B, Cracovie:131–279.
POLIÑSKI W. 1927. Znaczenie zoogeograficzne miêczakówPolski i koniecznoœæ ochrony ich zespo³ów. OchronaPrzyrody 7: 45–53.
POLLARD E. 1975. Aspects of the ecology of Helix pomatia L.J. Anim. Ecol. 44: 305–329.
RAO B. 1975. Some observations on feeding, anatomy, histo-logy of the digestive tract and digestive enzymes in thelimpet, Gallena radiata (Born). Proc. malac. Soc. London41: 309–320.
REEDER R. L., ROGERS S. H. 1979. The histochemistry of thespermatheca in four species of Sonorella (Gastropoda:Pulmonata). Trans. Amer. Micros. Soc. 98: 267–271.
RIEDEL A. 1954. Miêczaki okolic Kazimierza nad Wis³¹.Fragm. Faun. 7: 147–185.
RIEDEL A. 1988. Œlimaki l¹dowe Gastropoda terrestria. Kata-log Fauny Polski 36. PWN, Warszawa.
ROSSMÄSSLER E. A. 1835–1842. Iconographie der Land-und Süsswasser-Mollusken, mit vorzüglicher Berück-sichtigung der europäischen noch nicht abgebildetenArten. 1835–1837: Band 1. 1838–1842: Band 2. Dresden& Leipzig.
ROSSMÄSSLER E. A. 1859. Iconographie der Land- undSüsswasser-Mollusken Europas, mit vorzüglicher Berück-sichtigung kritischer und noch nicht abgebildeter Arten.Band 3. Leipzig.
RUDOLPH P. H. 1975. Accumulation of galactogen in the al-bumen gland of Triodopsis multilineata (Pulmonata, Sty-lommatophora) after dormancy. Malacol. Rev. 8: 57–63.
SEBER G. A. F. 1982. The estimation of animal abundanceand related parameters. Griffin, London.
SOKAL R. R., ROHLF F. J. 1995. Biometry. Freeman, NewYork.
STÊPCZAK K. 1976. Wystêpowanie, zasoby, uzyskiwanie iochrona œlimaka winniczka (Helix pomatia L.) w Polsce.Wyd. Nauk. UAM Poznañ, Ser. zool. 3: 1–68.
STÊPCZAK K., £AWNICZAK H., WIELAND A. 1982. Dobowa i se-zonowa aktywnoœæ œlimaków winniczków (Helix pomatiaL.). Pr. Kom. Biol. PTPN 66: 13–35.
SUMNER A. T. 1965a. The cytology and histochemistry of thedigestion gland cells of Helix. Quart. J. micr. Soc. 106:173–192.
SUMNER A. T. 1965b. Experiments on phagocytosis and lipidabsorption in the alimentary system of Helix. J. Royal.micr. Soc. 84: 415–431.
SUTHERLAND W. J. 1996. Ecological census techniques: ahandbook. Cambridge University Press, Cambridge.
SHILEYKO A. A. 1978. Nazemnye mollyuski nadsemeystwaHelicoidea. Fauna SSSR. Mollyuski 3, 6. Nauka, Lenin-grad.
TISCHLER W. 1973. Zur Biologie und Ökologie der Wein-bergschnecke (Helix pomatia L.). Faun.-ökol. Mitt. 4:283–299.
TISCHLER W. 1974. Aussenfaktoren als Auslöser tageszeitli-chen Verhaltens und begrenzter Dormanz bei Helix po-matia L. Zool. Anz. 193: 251–255.
URBAÑSKI J. 1937. Bemerkenswerte Weichtierfunde aus Po-len. Fragm. Faun. 3: 11–20.
URBAÑSKI J. 1947. Krytyczny przegl¹d miêczaków (Mollusca)Polski. Ann. UMCS, Ser. C, 2: 1–35.
URBAÑSKI J. 1948. Reliktowe miêczaki ziem polskich i nie-których krajów przyleg³ych. Ochrona Przyrody 18: 66–95.
URBAÑSKI J. 1957. Krajowe œlimaki i ma³¿e. Klucz do ozna-czania wszystkich gatunków dot¹d w Polsce wykrytych.PZWS, Warszawa.
URBAÑSKI J. 1964. Œlimak winniczek Helix pomatia L. – jegosystematyka, biologia, znaczenie gospodarcze i ochrona.Ochrona Przyrody 29: 215–254.
URBAÑSKI J. 1973. Charakterystyka fauny œlimaków (Gastro-poda) Wy¿yny Krakowsko-Czêstochowskiej. Rocz. Muz.Czêstoch. 3: 217–232.
URBAÑSKI J. 1977. Miêczaki. In: Przyroda Ojcowskiego ParkuNarodowego. Stud. Naturae B, 28: 299–318.
URBAÑSKI J. 1989. Mollusca – miêczaki. In: Zoologia, bezkrê-gowce (GRABDA E., ed.), vol. 1: 3, pp. 621–921. PWN,Warszawa.
WAGGE L. E. 1951. The activity of amoebocytes and of alka-line phosphatases during the regeneration of the shell inthe snail, Helix aspersa. Quart. J. micr. Sci. 92: 307–321.
WIKTOR A. 1989. Limacoidea et Zonitoidea nuda. Œlimakipomrowiokszta³tne (Gastropoda: Stylommatophora).Fauna Polski 12. PWN, Warszawa.
WIKTOR A., RIEDEL A. 1992. Œlimaki l¹dowe Gastropoda ter-restria. In: Czerwona lista zwierz¹t gin¹cych i za-gro¿onych w Polsce (G£OWACIÑSKI Z., ed.), pp. 31–38.PAN, Kraków.
WILBUR K. M., YONGE C. M. (eds). 1964. Physiology of Mol-lusca. 1. Academic Press, New York & London.
ZAWIEJA E. 1979. Mechanizmy trawienia w ¿o³¹dku Pulmona-ta. Przegl. Zool. 23: 233–237.
ZAWISTOWSKI S. 1975. Technika histologiczna. Histologiaoraz podstawy histopatologii. PZWL, Warszawa.
¯BIKOWSKA E. 1997. Porównawcze badania hemocytówdwóch gatunków p³ucodysznych œlimaków ró¿ni¹cych siêœrodowiskiem ¿ycia. XIII Krajowe Seminarium Malako-logiczne, Œwinoujœcie: 99–107.
received: July 5th, 1999accepted: September 27th, 1999
240 Koralewska-Batura E.