00_gesamter text_2 - opus4.kobv.de
TRANSCRIPT

Aus der Abteilung für Transfusionsmedizin und Hämostaseologie
der Friedrich-Alexander-Universität Erlangen-Nürnberg
Direktor: Prof. Dr. R. Eckstein
Vergleichende Untersuchung der Zellqualität mononuk leärer Zellen aus
Leukozytenreduktionskammern und Leukapheresaten
Inaugural-Dissertation zur Erlangung der Doktorwürde
an der Medizinischen Fakultät
der Friedrich-Alexander-Universität Erlangen-Nürnberg
vorgelegt von
Thomas Michael Weidinger
aus Erlangen

Gedruckt mit Erlaubnis der
Medizinischen Fakultät der Friedrich-Alexander-Univ ersität Erlangen-Nürnberg
Dekan: Prof. Dr. med. Dr. h.c. J. Schüttler Referent: Priv.-Doz. Dr. med. E. Strasser Koreferent: Prof. Dr. med. R. Eckstein Tag der mündlichen Prüfung: 25.07.2012

Meiner Mutter und meiner Schwester
in tiefer Dankbarkeit gewidmet.

Inhaltsverzeichnis 4
I. Inhaltsverzeichnis
I. Inhaltsverzeichnis………………………………………….………………………….. ...4
II. Zusammenfassung und Abstract…………………….………………………… .........6
III. Einleitung……………………………………………………………………….……..... .10
1. Zelluläre Immuntherapien bei Tumorerkrankungen 10
2. Dendritische Zellen und Gewinnung dendritischer Zellen 11
3. Zellschädigung und Zelltod 12
3.1. Zelltod durch Nekrose 12
3.2. Zelltod durch Apoptose 13
3.2.1. Membranintegrität und Phosphatidylserin-Switch 14
4. Fragestellung 16
IV. Material und Methoden……………………………………………………………….. .17
1. Blutspender und Blutgewinnung 17
1.1 Blutspender 17
1.2 Leukapheresate 17
1.3 Thrombozytapherese und Leukozytenreduktionskammern 18
2. Lagerung, Bestimmung der Zellkonzentrationen und Blutgasanalyse 20
3. Durchflusszytometrische Messungen 20
3.1 Das Prinzip der Durchflusszytometrie 20
3.2 Färbungen 22
3.2.1 Oberflächenantigenfärbungen 22
3.2.2 Nachweis und Messung von Nekrose und Apoptose 23
3.2.2.1 Nekrose und 7-Aminoactinomycin D (7-AAD) 23
3.2.2.2 Apoptose, Phosphatidylserin-Switch und Annexin V 23
3.3 Instrument Settings 24
3.4 Auswertung der Messungen 25
3.4.1 Festlegung der Gates der durchflusszytometrischen Messungen 25
3.4.1.1 Antikörperfärbungen gegen Oberflächenantigene 25
3.4.1.2 Färbungen mit Annexin V und 7-AAD 26
3.4.2 Statistische Auswertung 28
4. Verwendetet Formeln 28

Inhaltsverzeichnis 5
5. Versuchsreihen zur Beantwortung der Fragestellung 28
5.1 Vergleich von Leukozytenreduktionskammern zweier Zellseparatoren 28
5.2 Zellgehalt und Zellschädigung in Leukozytenreduktionskammern 29
5.3 Blut aus Leukozytenreduktionskammern und aus Leukapheresaten 30
V. Ergebnisse……………………………………….………………………………………31
1. Vergleich von Leukozytenreduktionskammern zweier Zellseparatoren 31
2. Zellgehalt und Zellschädigung in Leukozytenreduktionskammern 33
3. Blut aus Leukozytenreduktionskammern und aus Leukapheresaten 38
VI. Diskussion………………………………………………………………………….…… 42
1. Vergleich von Leukozytenreduktionskammern zweier Zellseparatoren 42
2. Zellgehalt und Zellschädigung in Leukozytenreduktionskammern 43
3. Blut aus Leukozytenreduktionskammern und aus Leukapheresaten 46
VII. Literaturverzeichnis………………………………………………………………… … 50
VIII. Abkürzungen…………………………………………………………………………….57
IX. Verzeichnis der Vorveröffentlichungen………………………… ………………….58
X. Anhang……………………………………………………………………………………59
Danksagung

Zusammenfassung 6
II. Zusammenfassung und Abstract
Hintergrund und Ziele
Zur Herstellung dendritischer Zellen für zelluläre Immuntherapien werden mo-
nonukleäre Zellen des peripheren Bluts benötigt. Der Goldstandard zur Gewinnung
mononukleärer Zellen ist die Leukozytapherese. Die Kultivierungseffizienz von dendri-
tischen Zellen aus Monozyten beträgt aktuell ca. 30%. Eine Vorschädigung der Mo-
nozyten im Sinne der Apoptose bzw. Nekrose kann einen Einfluss auf die Kultivie-
rungsrate haben. Kürzlich wurde eine neue Quelle zur Gewinnung von mononukleären
Zellen mit ausgezeichneter Qualität beschrieben: die Leukozytenreduktionskammern
der Trima Accel® Thrombozytapheresemaschine. Ziel dieser Arbeit ist der Vergleich
zwischen Leukozytenreduktionskammern und Leukapheresaten hinsichtlich des Leu-
kozytengehalts und der Apoptose- und Nekroserate mononukleärer Zellen.
Methoden
Nach der Thrombozytapherese und nach der Leukozytapherese wurden Blut-
proben aus den Leukozytenreduktionskammern und Leukozytapheresaten entnommen
und sofort für Messungen verwendet. Blut aus den jeweiligen Apheresen wurde in
PVC-Beutel überführt und gelagert. Weitere Messungen erfolgten nach einer Lagerung
von 6, 24, 48 und 72 Stunden. Bestimmt wurden Zellkonzentration, Blutgase und
durchflusszytometrisch der Anteil an CD14-, CD45-, Annexin V- und 7-AAD-positiven
Zellen.
Ergebnisse und Beobachtungen
Der Gehalt mononukleärer Zellen der Leukozytenreduktionskammern des Trima
Accel® Zellseparators war um ein Vielfaches höher als in denen der COBE® Spectra. In
den Leukozytenreduktionskammern der Trima Accel® korrelierte der Leukozytengehalt
sehr gut mit der Dauer und dem prozessierten Blutvolumen der Thrombozytapherese;
nach doppelter Thrombozytapherese waren signifikant mehr Leukozyten in den Kam-
mern waren als nach einfacher. In den Leukozytenreduktionskammern des Trima Ac-
cel® Zellseparators war die Leukozytenkonzentration signifikant höher als in Leuko-
zytapheresaten. Auf Grund des höheren Produktvolumens der Leukozytapheresate

Zusammenfassung 7
war hier der Leukozytengehalt aber deutlich höher als in den Leukozytenreduktions-
kammern. Es wurde gefunden, dass der Anteil Annexin V+/7-AAD+ Zellen nach Kurz-
zeitlagerung über 72 Stunden in doppelten verglichen mit einfachen Thrombozyten-
spenden signifikant erhöht war. In den Leukozytapheresaten dagegen kam es während
der Lagerung zu einem früheren und stärkerem Anstieg der Apoptose- und Nekrosera-
te mononukleärer Zellen mit einem früheren und stärkeren Abfall des prozentuellen
Anteils mononukleärer Zellen verglichen mit den Leukozytenreduktionskammern der
Trima Accel®. Des Weiteren fiel der pH-Wert in gelagerten Leukozytapheresaten stär-
ker ab als in gelagerten Leukozytenreduktionskammern, wobei in den Leukozytaphere-
saten der pH-Wert sehr gut mit dem Anteil an nekrotischen Zellen korrelierte.
Praktische Schlussfolgerungen
Die Leukozytenreduktionskammern der Trima Accel® stellen eine sehr gute
Quelle für mononukleäre Zellen dar, während dies für die COBE Spectra® nicht zutrifft.
Die mononukleären Zellen der Trima Accel® Leukozytenreduktionskammern zeigten
während der Kurzzeitlagerung eine signifikant bessere Qualität verglichen mit Leuko-
zytapheresaten. Leukozytapheresate sind auf Grund des höheren Gehalts mononukle-
ärer Zellen der Goldstandard für die Gewinnung von mononukleären Zellen, die heute
für die klinische Versorgung mit Zelltherapeutika zunehmend benötigt werden. Leuko-
zytenreduktionskammern sind aber eine gute Leukozytenquelle für die experimentelle
Zellforschung, wobei vermutlich das schonende Absammeln der Zellen in den Leuko-
zytenreduktionskammern mit einem geringeren Zellstress und damit geringeren
Zellapoptose verbunden ist.

Zusammenfassung 8
Background and Objectives
Peripheral blood mononuclear cells are frequently used for dendritic cell culture
in adoptive immunotherapies. The gold standard to obtain monocytes for dendritic cell
therapies today is leukapheresis. Current culture efficiency is about 30%. Cell damage
of monocytes in terms of apoptosis or necrosis might influence the efficiency of
dendritic cell culture. A new source for peripheral blood mononuclear cells of good
quality, the leukoreduction system chambers of Trima Accel® plateletpheresis device,
has recently been described. This work compares leukoreduction system chambers
and leukapheresis products with regard to leukocytes cell yield and apoptosis and ne-
crosis of mononuclear cells.
Materials and Methods
Immediately after platelet- and leukapheresis blood samples were transferred
from leukoreduction system chambers and leukapheresis products into PVC bags
where they were stored for a maximum of 72 hours. Within one hour after apheresis
and after six, 24, 48 and 72 hours of storage cell counts were measured, blood gas
analysis was performed and the fractions of CD14-, CD45-, Annexin V- and 7-AAD-
positive events were analysed by flow cytometry.
Results
In leukoreduction system chambers of Trima Accel® mononuclear cell yield was
many times higher to those of COBE® Spectra. Leukocyte count of Trima Accel® leuko-
redution system chambers correlated very well with donation time and processed blood
volume. Thus, after double platelet units, leukoreduction system chambers contained a
significantly higher leukocyte count compared to those of single platelet units. Leuko-
cyte concentration of leukoreduction system chambers were higher compared to those
obtained from leukapheresis products. However, due to significantly higher product
volume leukocyte yields of leukapheresis products many times exceeded those con-
tained in leukoreduction system chambers. After 72 hours of storage, fraction of Annex-
in V+/7-AAD+ events were significantly higher in double compared to single platelet
units. In contrast, fractions of apoptotic and necrotic mononuclear cells obtained from
leukapheresis products increased earlier during storage and to higher levels accompa-
nied with an earlier drop of mononuclear cell counts compared to leukoreduction sys-
tem chambers of Trima Accel®. Furthermore, during short time storage pH decreased

Zusammenfassung 9
in leukapheresis products to lower levels compared to leukoreduction system cham-
bers showing a very good inverse correlation of pH and fraction of necrotic cells in
samples of leukapheresis products.
Conclusion
Leukoreduction system chambers of the Trima Accel® plateletpheresis device
are a very good source for peripheral blood mononuclear cells which does not apply for
those of the COBE® Spectra plateletpheresis device. Mononuclear cells of Trima
Accel® leukoreduction system chambers showed a better viablilty regarding apoptosis
and necrosis compared to stored samples of leukapheresis products. Due to high cell
yields leukapheresis is the gold standard to obtain mononuclear cells which today are
increasingly needed for cellular therapies. For experimental purpose leukoreduction
system chambers are a good source to obtain leukocytes while the gentle way to col-
lect cells in leukoreduction system chambers might be accompanied with lower cell
stress and thus lower cell apoptosis.
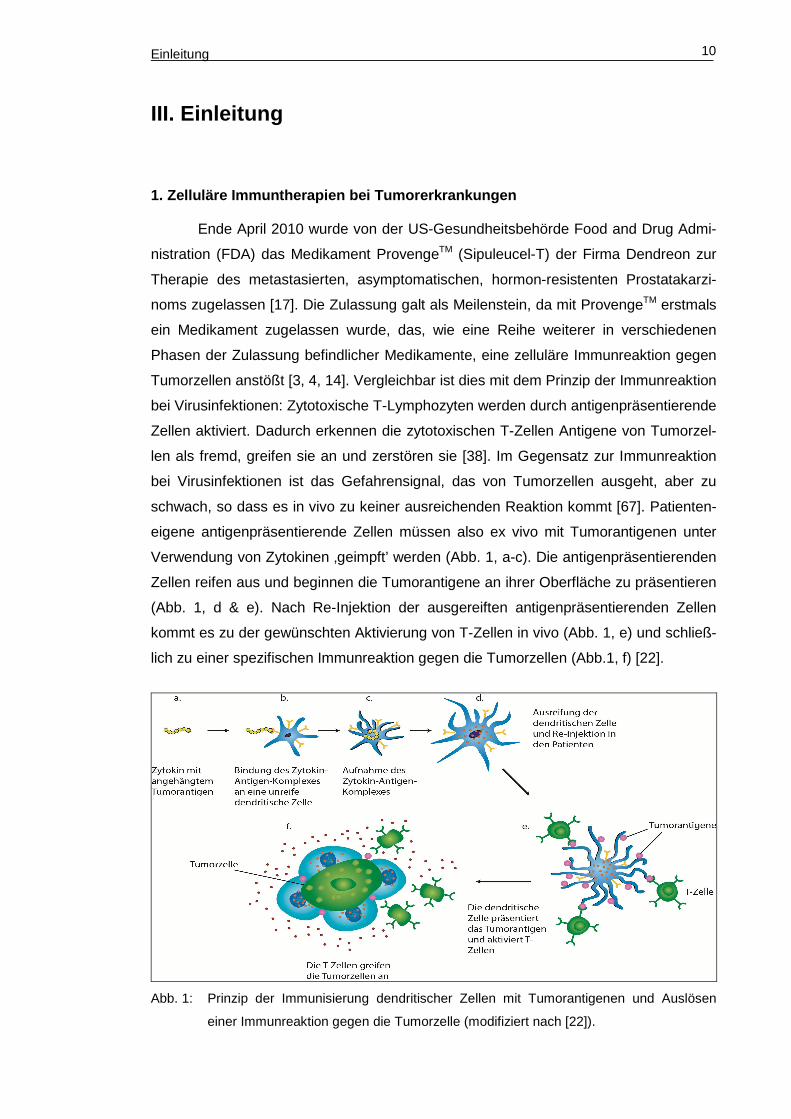
Einleitung 10
III. Einleitung
1. Zelluläre Immuntherapien bei Tumorerkrankungen
Ende April 2010 wurde von der US-Gesundheitsbehörde Food and Drug Admi-
nistration (FDA) das Medikament ProvengeTM (Sipuleucel-T) der Firma Dendreon zur
Therapie des metastasierten, asymptomatischen, hormon-resistenten Prostatakarzi-
noms zugelassen [17]. Die Zulassung galt als Meilenstein, da mit ProvengeTM erstmals
ein Medikament zugelassen wurde, das, wie eine Reihe weiterer in verschiedenen
Phasen der Zulassung befindlicher Medikamente, eine zelluläre Immunreaktion gegen
Tumorzellen anstößt [3, 4, 14]. Vergleichbar ist dies mit dem Prinzip der Immunreaktion
bei Virusinfektionen: Zytotoxische T-Lymphozyten werden durch antigenpräsentierende
Zellen aktiviert. Dadurch erkennen die zytotoxischen T-Zellen Antigene von Tumorzel-
len als fremd, greifen sie an und zerstören sie [38]. Im Gegensatz zur Immunreaktion
bei Virusinfektionen ist das Gefahrensignal, das von Tumorzellen ausgeht, aber zu
schwach, so dass es in vivo zu keiner ausreichenden Reaktion kommt [67]. Patienten-
eigene antigenpräsentierende Zellen müssen also ex vivo mit Tumorantigenen unter
Verwendung von Zytokinen ‚geimpft’ werden (Abb. 1, a-c). Die antigenpräsentierenden
Zellen reifen aus und beginnen die Tumorantigene an ihrer Oberfläche zu präsentieren
(Abb. 1, d & e). Nach Re-Injektion der ausgereiften antigenpräsentierenden Zellen
kommt es zu der gewünschten Aktivierung von T-Zellen in vivo (Abb. 1, e) und schließ-
lich zu einer spezifischen Immunreaktion gegen die Tumorzellen (Abb.1, f) [22].
Abb. 1: Prinzip der Immunisierung dendritischer Zellen mit Tumorantigenen und Auslösen
einer Immunreaktion gegen die Tumorzelle (modifiziert nach [22]).

Einleitung 11
In der onkologischen Therapie werden auch reine Antikörper gegen Tumoranti-
gene als Chemotherapeutika eingesetzt. Die Wirkweise ist mit der einer Immunreaktion
vergleichbar. Allerdings geht sie nicht vom Immunsystem aus. Beispiele dafür sind die
Medikamente Rituximab zur Therapie von B-Zell-Lymphomen oder Trastuzumab zur
Therapie des HER2-positiven Mammakarzinoms. Zwar erspart der Einsatz monoklona-
ler Antikörper in der Immuntherapie gegen Tumore den Aufwand allogene antigenprä-
sentierende Zellen zu sammeln und diese in vitro zu immunisieren. Im Gegensatz zu
einer reinen Antikörpertherapie bieten T-Zellen, die nach Aktivierung durch antigenprä-
sentierende Zellen gegen Tumorzellen gerichtet sind, aber einige Vorteile: T-Zellen
können besser zu Tumoren in tief liegendem Gewebe vordringen, sie proliferieren bis
alle Antigen tragenden Tumorzellen eliminiert sind und sie bilden ein immunologisches
Gedächtnis, so dass eine Immunreaktion bei Rezidiven möglich ist [16].
2. Dendritische Zellen und Gewinnung dendritischer Zellen
Die für die Immunisierung von Patienten gegen Tumorantigene verwendeten
antigenpräsentierenden Zellen sind Dendritische Zellen (griechisch dendron: Baum),
da sie als die potentesten antigenpräsentierenden Zellen des menschlichen Immunsys-
tems gelten [22, 38, 55]: Sie beeinflussen viele verschiedene Klassen der Lymphozy-
ten (B, NK, NKT) und viele Arten der T-Zell-Antwort (Th1/Th2, T-Zell-Regulierung).
Außerdem gelten sie als Wächter des Immunsystems, da sie dieses aktivieren und die
frühen Immunreaktionen beeinflussen [6, 56]. Man unterscheidet Vorläuferzellen, unrei-
fe dendritische Zellen und reife dendritische Zellen. Um eine ausreichende Immunisie-
rung gegen Tumorantigene zu erreichen sind ca. 1x107 reifer dendritischer Zellen nötig
[50]. Diese Menge ist durch Anreicherung von dendritischen Zellen aus peripherem
Blut schwer zu erreichen. In vivo können sich CD14+ Monozyten im Rahmen von Im-
munreaktionen bei Infekten in dendritische Zellen umwandeln. In vitro ist dies auch
möglich, indem man die CD14+ Monozyten mit verschiedenen Zytokinen, bevorzugt
mit GM-CSF und IL-4, kultiviert [46, 48, 50]. Der Goldstandard zur Gewinnung von Mo-
nozyten für die Zelltherapie mit dendritischen Zellen ist die Leukapherese, da bei einer
Leukapherese mehr als 1x109 CD14+ Monozyten gewonnen werden können [59, 62].
Dietz und Kollegen haben in einer 2007 veröffentlichten Arbeit berichtet, dass
aus Leukozytenreduktionskammern, die als Nebenprodukt bei Thrombozytapheresen
mit den Trima Accel® Apheresegeräten entstehen, mononukleäre Zellen von sehr gu-
ter Qualität gewonnen werden können [15, 35, 41]. Neron et al. beschrieben, dass der
Anteil mononukleärer Zellen in Leukozytenreduktionskammern um das Fünffache hö-
her ist als in Leukoreduktionsfiltern [41]. Leukozytenreduktionskammern werden auf
Grund des geringen Volumens von maximal 10ml vorrangig zur Forschung verwendet.

Einleitung 12
Abb. 2: Morphologischer Ablauf
der Nekrose (modifiziert
nach [30]).
Auch bei dem Thrombozytenspendegerät COBE® Spectra werden zur Leukozytenre-
duktion Leukozytenreduktionskammern verwendet. Über die Qualität des hierbei ge-
sammelten, an mononukleären Zellen reichen Blutes sind bisher noch keine Daten
veröffentlicht.
3. Zellschädigung und Zelltod
Aktuell beträgt die Ausreifungsrate von dendritischen Zellen aus Monozyten ca.
20% bis 40%. Es ist nahe liegend, dass hierbei eine Schädigung der Monozyten vor
Kultivierung Einfluss auf die Ausreifungsrate haben kann. Eine Schädigung der Zellen
kann zum Beispiel physiologisch im Sinne einer Zellalterung, aber auch unter patholo-
gischen Umständen wie Hypoxie, immunologischen Reaktionen, Gen-Defekten und
chemischen, physikalischen oder infektiösen Noxen stattfinden [38]. Abhängig vom
Ausmaß der Schädigung und der Art der Schädigung erholt sich die Zelle ad integrum
oder geht zugrunde. Im letzteren Fall gibt es zwei mögliche Wege: die Apoptose oder
die Nekrose [51, 65].
3.1. Zelltod durch Nekrose
Der Begriff „Nekrose“ leitet sich von dem
griechischen Wort nekrosis (ηεκροσισ), die Tötung,
ab. Jede abrupte, ausreichend intensive Zellschä-
digung kann zur Zellnekrose führen. Typisches
Beispiel hierfür ist die Zellnekrose beim akuten
Myokardinfarkt, bei der es auf Grund einer Hypoxie
zur irreversiblen Schädigung der Myokardzellen
kommt. Die wichtigsten morphologischen Merkma-
le einer Nekrose sind das Anschwellen der Zellen
und der Verlust der Integrität sowohl der Plasma-
membran als auch der Kernmembran („Karyor-
rhexis“). Der Zellkern schrumpft („Pyknosis“) und
wird aufgelöst („Karyolysis“). Gleichzeitig tritt der
Zellinhalt mitsamt verschiedener Proteasen durch
die löchrige Plasmamembran aus der Zelle aus
und verursacht eine Entzündungsreaktion im um-
liegenden Gewebe (Abb. 2) [63]. Ursprünglich galt
die Nekrose als plötzlich eintretendes Ereignis, mit
zufälligem, ungeregeltem Ablauf ohne Beteiligung

Einleitung 13
von Signalkaskaden. Neuere Studien zeigen, dass unter bestimmten Voraussetzun-
gen, vor allem in Abhängigkeit der auslösenden Stimuli, vorbestimmte Signalkaskaden
während der Nekrose, vergleichbar zur Apoptose, ablaufen [64]. Am morphologischen
Ablauf der Nekrose ändert sich dabei aber nichts, so dass diese Erkenntnis keinen
Einfluss auf die in dieser Arbeit dargestellten Ergebnisse haben wird.
3.2. Zelltod durch Apoptose
Die Apoptose beschreibt eine Form des Zelltodes, den man als ‚stiller Selbst-
mord’ bezeichnen könnte. Eingeführt wurde der Begriff „Apoptose“ 1972 von Kerr et al.,
die eine neue Form des Zelltodes beobachtet haben; ohne Anschwellen und Platzen
der Zelle und ohne Entzündungsreaktion des umliegenden Gewebes. Stattdessen lös-
ten sich die sterbenden Zellen aus dem Zellverband, zersetzten sich nach und nach in
von Plasmamembran umhüllte Vesikel und wurden von umgebenden Zellen phagozy-
tiert (Abb. 3) [30, 31]. Den diesen Vorgang beschreibenden Begriff „Apoptose“, bzw. im
englischen „Apoptosis“, prägte Professor J. Cormack (Lehrstuhl für klassische Philolo-
gie (Griechisch), Universität Aberdeen, Aberdeen, GB). Apoptosis (άπωπτωσισ) wird
im griechischen als Beschreibung des Abfallens von Laub oder Blütenblättern beim
Welken verwendet.
Abb. 3: Morphologischer Ablauf der Apoptose und Phagozytose der apoptotischen Vesikel
(modifiziert nach [30]).
Die Auslöser für die Apoptose sind vielfältig. Pathologische Ursachen sind
DNS-Schädigung, intrazelluläre Ansammlung fehlerhaft gefalteter Proteine oder Zell-
schädigung bei diversen Infektionen, beispielsweise mit Adenoviren. Es gibt aber auch

Einleitung 14
physiologische Ursachen für die Apoptose. Bei der positiven und negativen Selektion
während der Reifung der T-Lymphozyten, bei der Rückbildung von Zellen während der
Embryogenese, bei der Gewebshomöostase, aber auch beim Abbau von Zellen, deren
Funktionen nicht mehr gebraucht werden, spielt die Apoptose eine entscheidende Rol-
le [34].
Heute weiß man, dass die Apoptose auf molekularer Ebene ein streng regulier-
ter Prozess mit genau aufeinander abgestimmten Signalübertragungswegen und Er-
eignissen ist [25]. Ausgelöst wird sie entweder durch Liganden an Todesrezeptoren
(„extrinsische Kaskade“) oder durch Proteine mitochondriellen Ursprungs („intrinsische
Kaskade“), vorrangig Cytochrom C [18]. Bei beiden Wegen der Aktivierung wird eine
Kaskade proteolytischer Enzyme angestoßen, die letztlich für fast alle Veränderungen
im Verlauf der Apoptose verantwortlich zu sein scheint. Diese proteolytischen Enzyme
heißen „Caspasen“, hergeleitet vom Cystein im aktiven Zentrum des Enzyms und der
Eigenschaft Peptidbindungen immer nach einer Asparaginsäure im Substrat zu spalten
(engl.: cysteinyl-aspartate specific protease). In vitalen Zellen liegen die Caspasen in
inaktiver Form vor. Ihre Aktivierung erfolgt durch die proteolytische Abspaltung der
N-Terminalen Pro-Domäne. Bisher wurden sieben unterschiedliche Caspasen ent-
deckt, die für die Apoptose menschlicher Zellen bedeutsam sind. Kommt es zu einem
apoptogenen Stimulus, unabhängig ob intrinsisch oder extrinsisch, wird nach Ablauf
der jeweiligen Caspase-Kaskade schließlich die Caspase 3 als zentrale Caspase der
Apoptose aktiviert [42]. Die zentrale Rolle der Caspasen an der Apoptose wird daran
deutlich, dass sie für die Aktivierung von DNSen mit Zersetzung der DNS, für die
Fragmentierung des Zellkerns, für die Zersetzung des Zytoskeletts mit konsekutiver
Bildung membranumhüllter Vesikel und für die Aufhebung der Plasmamembranasym-
metrie verantwortlich sind.
3.2.1. Membranintegrität und Phosphatidylserin-Swit ch
Die Zellmembran einer menschlichen Zelle besteht aus Phospholipiden, die
sich entsprechend dem Fluid-Mosaik-Model von Singer und Nicolson in einer Doppel-
schicht anordnen, so dass die hydrophoben Lipidanteile im Inneren der Membran zu
einander stehen und die hydrophilen Phosphatanteile an den Außenseiten der Memb-
ran aneinander gereiht sind (Abb. 4, a) [54]. In und an diese Phospholipiddoppelschicht
sind weitere Moleküle wie zum Beispiel Proteine, Lipide oder Cholesterine gebunden.
Die Aufgaben dieser Moleküle sind verschieden; beispielsweise können Transmemb-
ranproteine als Ionenkanäle fungieren.

Einleitung 15
Physiologischer Weise ist die Plasmamembran asymmetrisch. Das bedeutet,
dass es bei vitalen Zellen Moleküle gibt, die nur auf der Innenseite der Membran zu
finden sein können, nicht aber auf der Außenseite und umgekehrt [36]. Diese Membra-
nintegrität ist für den physiologischen Ablauf vieler Prozesse, zum Beispiel die Auf-
rechterhaltung der Elektrolytverteilung zwischen dem intra- und extrazellulären Raum,
einer vitalen Zelle notwendig. Eines dieser Moleküle ist das Phospholipid Phos-
phatidylserin. In vitalen Zellen kommt Phosphatidylserin ausschließlich in der intrazellu-
lären Schicht der Plasmamembran vor. Die Aufrechterhaltung dieser Verteilung ist
energieabhängig [49]. Wird aber eine Zelle apoptotisch, wird das Phosphatidylserin
sehr früh im Verlauf der Apoptose von der inneren auf die äußere Seite der Membran
verschoben, unabhängig vom Auslöser der Apoptose (Abb. 4, b) [37]. Diesen Vorgang
nennt man Phosphatidylserin-Switch. Wahrscheinlich dient die Externalisation von
Phosphatidylserin den umliegenden Zellen, die Apoptose zu erkennen und die Pha-
gozytose der im Verlauf der Apoptose entstehenden membranumhüllten Zellreste zu
initiiren [9, 11].
Abb. 4: Phosphatidylserin-Switch im Verlauf der Apoptose mit Phosphatidylserin in vitalen
Zellen ausschließlich auf der Zytoplasma-Seite (a) und Switch im Verlauf der Apopto-
se auf die Außenseite der Membran (b) (modifiziert nach [40]).

Einleitung 16
4. Fragestellung
Leukozytenreduktionskammern scheinen eine vielversprechende Quelle für
mononukleäre Zellen des peripheren Blutes zu sein. Einige Daten zur Qualität der aus
den Leukozytenreduktionskammern gewonnenen Zellen liegen bereits vor [15, 35, 41].
Bisher wurden aber nur Kammern untersucht, die mit der Trima Accel® hergestellt wur-
den. Des Weiteren wurden bei allen bisher veröffentlichten Daten keine Angaben zur
Apoptose gemacht. Daher ergeben sich folgende Fragestellungen:
1. Nicht nur bei der Thrombozytapherese mit der Trima Accel® sondern auch wäh-
rend der Thrombozytapherese mit der COBE® Spectra werden Leukozytenredukti-
onskammern verwendet. Der Zellgehalt und gegebenenfalls die Qualität der Zellen
der Leukozytenreduktionskammern beider Thrombozytapheresemaschinen sollen
miteinander verglichen werden.
2. In den bisher veröffentlichten Daten wurde Blut von sehr homogenen Spender-
gruppen verwendet und Ergebnisse mit geringer Streubreiter erzielt. Zu prüfen ist,
ob es bei Einfach- gegenüber Doppelapharesen zu Unterschieden in der Leukozy-
tenzahl der Leukozytenreduktionskammern kommt. Sind im Verlauf der Kurzzeitla-
gerung über 72 Stunden Zeichen einer Schädigung im Sinne der Apoptose bzw.
Nekrose bei mononukleären Zellen des peripheren Blutes nachweisbar? Wie ver-
halten sich die Zellkonzentrationen im Verlauf der Lagerung?
3. Sind Unterschiede in der Qualität bezüglich der Apoptose- und Nekroserate zwi-
schen mononukleären Zellen aus Leukozytenreduktionskammern und mononukle-
ären Zellen der Leukapherese nachweisbar? Haben die unterschiedlichen Prinzi-
pien der Anreicherung mononukleärer Zellen bei der Leukapherese und in Leuko-
zytenreduktionskammern einen Einfluss auf die Qualität der Produkte? Kommt es
zu Unterschieden in über 48 Stunden gelagerten Produkten?

Material und Methoden 17
IV. Material und Methoden
1. Blutspender und Blutgewinnung
1.1 Blutspender
Das in dieser Forschungsarbeit verwendete Blut wurde entsprechend der Richt-
linien zur Gewinnung von Blut und Blutbestandteilen und zur Anwendung von Blutpro-
dukten (Hämotherapie) der Bundesärztekammer gewonnen. [2, 26, 27] Alle Spender
dieser Studien wurden nach Feststellung der Spendetauglichkeit über den Spendevor-
gang einschließlich der Risiken aufgeklärt und stimmten der Verwendung ihres Blutes
zu Forschungszwecken zu. Alle Spender waren entsprechend der Richtlinien gesund.
Keiner der Spender erfüllte ein oder mehrere der unter Punkt 2.2 der Richtlinien zur
Gewinnung von Blut und Blutbestandteilen und zur Anwendung von Blutprodukten
(Hämotherapie) der Bundesärztekammer genannten Ausschlusskriterien. Für das Wohl
der Spender wurde jederzeit Sorge getragen. Das Studienprotokoll war von der Ethik-
kommission der Friedrich-Alexander-Universität Erlangen-Nürnberg geprüft und gebil-
ligt worden. Verwendet wurde Blut aus Leukapherisaten des COM.TEC® Zellseparators
und Blut aus Leukozytenreduktionskammern der COBE® Spectra und der Trima Accel®
Thrombozytapherese-Maschinen.
1.2 Leukapheresate
In dieser Arbeit wurden ausschließlich Produkte verwendet, die mit dem
COM.TEC® Zellseparator (Fresenius HemoCare GmbH, Bad Homburg, Deutschland)
unter dem Separationsprogramm „autoMNC“ hergestellt wurden. Bei der Leukapherese
wird venöses Blut des Spenders über ein geschlossenes System in die Apheresema-
schine geführt. Durch Zentrifugation wird das Vollblut entsprechend der jeweiligen phy-
sikalischen Eigenschaften der Blutbestandteile in Plasma, den sogenannten „Buffy
Coat“ aus Leukozyten und Thrombozyten und in Erythrozyten aufgespaltet (Abb. 5).
Bei der Aufspaltung spielt das Gewicht der einzelnen Zellen die entscheidende Rolle.
Entsprechend der Zentripetalkraft und der Trägheit der Masse werden bei Bewegun-
gen auf einer gekrümmten Bahn und bei kreisförmigen Bewegungen Körper mit einer
höheren Masse weiter nach außen getragen als Körper mit einer geringeren Masse
[47, 52]. Entsprechend des durchschnittlichen Gewichts der einzelnen Zellen werden
daher die schwereren Erythrozyten weiter vom Mittelpunkt der Kreisbahn der Zentrifu-
ge getragen als Leukozyten. Am wenigsten werden Thrombozyten und das Blutplasma

Material und Methoden 18
durch die Zentrifugation beeinflusst, so dass es zur in Abb. 5 gezeigten Aufspaltung
des Blutes kommt. Durch die Aufteilung der zellulären Blutbestandteile während der
Zentrifugation in verschiedene Schichten lassen sich bestimmte Blutzellen durch ge-
zieltes Absammeln der jeweiligen Schicht gewinnen.
Ziel der Leukapherese ist das Sammeln leukozytenreicher Blutprodukte, abzu-
saugen ist daher der Buffy Coat. Das „autoMNC“-Separationsprogramm verwendet
eine einstufige Separationskammer, genannt PL1a. Die Blutzellen werden während
den einstellbaren „Überschuss-“ und „Buffy Coat - Phasen“ gesammelt. Die Trenn-
schicht zum Buffy Coat wurde durch acht optische Sensoren kontrolliert, wobei sieben
auf die Zellschichten ausgerichtet waren und einer auf die Plasmaschicht. Während der
Apherese betrug die Zentrifugendrehzahl 1500 Umdrehungen pro Minute. Die durch
die Zentrifugation auf die Zellen wirkende Kraft entsprach der Kraft, die durch das
297-fache der Erdbeschleunigung hervorgerufen werden würde.
Abb. 5: Schematische Darstellung der durch Zentrifugation aufgespaltenen Blutbestandteile
bei der Leukozytapherese (modifiziert nach [53]).
1.3 Thrombozytapherese und Leukozytenreduktionskamm ern
Während der Thrombozytapherese mit den Geräten Trima Accel® (Caridian
BCT, Lakewood, CO, USA) und COBE® Spektra (Caridian BCT) werden im Spender-
blut Thrombozyten von Leukozyten und Erythrozyten getrennt, damit die Thrombozyten
im Konzentrat so rein wie möglich angereichert werden. Die Trennung geschieht beim
Plasma
Buffy Coat:
Thrombozyten
Leukozyten
Erythrozyten

Material und Methoden 19
Abb. 6: Leukozytenreduktionskammer der
Trima Accel Thrombozytapherese
Maschine nach Apherese.
Durchlaufen einer konischen Leukozytenreduktionskammer, wobei die Trennung der
Blutbestandteile von der Geometrie und den hydrodynamischen Eigenschaften der
Leukozytenreduktionskammern abhängt (Abb. 6). Durch den konischen, stufenför-
migen Aufbau der Kammer sammeln
sich im Bereich der Kammerwand die
schweren Blutbestandteile, also Erythro-
zyten und Leukozyten, an, während die
Thrombozyten und das Blutplasma
durch die Kammer hindurch fließen. So
enthalten die Leukozytenreduktions-
kammern erythrozyten- und leukozyten-
reiches Blut [27].
Bisher wurden diese Leukozyten-
reduktionskammern nach Abschluss des
Spendevorgangs entsorgt. Das in der
Kammer der Trima Accel® verbleibende
Blut ist auf Grund des Leukozytenreich-
tums aber eine gute Quelle für mononuk-
leäre Zellen des peripheren Blutes, wie
kürzlich gezeigt werden konnte [15]. Es
konnte weiterhin gezeigt werden, dass
die gewonnenen Monozyten und Lym-
phozyten eine hervorragende Qualität
besitzen und gut zu dendritischen Zellen kultiviert werden können [15, 35, 41]. Daten
zur Eignung der Leukozytenreduktionskammer des COBE® Spectra Zellseparators als
Quelle für mononukleäre Zellen liegen bisher nicht vor.
Bei den Thrombozytapheresen, deren Leukozytenreduktionskammern für diese
Arbeit genutzt wurden, wurde an der Trima Accel® die Software Version 5.1, an der
COBE® Spectra die Software Version 7.0 der jeweiligen Hersteller verwendet. Die Ge-
räteeinstellungen waren bei beiden Separatoren gleich: ‚anticoagulant ratio’ 11:1, ‚draw
Management’ 6; ‚return Management’ 4, maximaler ‚draw flow’ mittel. Um zu einer
Thrombozytenspende zugelassen zu werden, musste die Thrombozytenkonzentration
der Spender vor der Spende mindestens 160 x 109 Thrombozyten pro Liter betragen.
Überstieg die Thrombozytenkonzentration 230 x 109 Thrombozyten pro Liter wurde
eine doppelte Thrombozytenspende durchgeführt (Doppel-Thrombozytapherese, D-
TA), sonst eine einfache (Einfach-Thrombozytapherese, E-TA). Sofort nach Abschluss
der Apherese wurden die Thrombozytenkonzentrate und im direkten Anschluss auch

Material und Methoden 20
die zu- und abführenden Schläuche der Leukozytenreduktionskammern abgeschweißt.
Das Blut der Leukozytenreduktionskammern wurde unter Wahrung aseptischer Bedin-
gungen über einen „sampling site coupler“ (Baxter, REF EMC1401) in Beutel aus Po-
lyvinylchlorid (PVC) mit einem Fassungsvolumen von je 150ml (Compoflex R Single
Bags, Fresenius Kabi, Bad Homburg, Deutschland) überführt und gelagert.
2. Lagerung, Bestimmung der Zellkonzentrationen und Blutgasanalyse
Die Blutprodukte wurden bei 22°C ± 2°C ohne Agitati on und vor Licht geschützt
im Thrombozyteninkubator gelagert. Lagermedien wurden nicht verwendet. Zur Lage-
rung wurden die Leukozytenreduktionskammern über ein Verbindungsstück (sampling
site coupler, Baxter, REF EMC1401) in einen 150ml PVC-Beutel (Compoflex R Single
Bags, Fresenius Kabi, Bad Homburg, Deutschland) überführt. Um vergleichbare Be-
dingungen zu schaffen wurden aus den Leukapherisaten Proben mit einem Volumen
von 10ml, dem maximalen Volumen der Leukozytenreduktionskammern, unter asepti-
schen Bedingungen entnommen und ebenfalls in den 150ml PVC-Beuteln gelagert. Die
Gasaustauschraten der PVC-Beutel betrugen: O2: 0.675l / m2 Beuteloberfläche / 24 h /
bar; CO2: 4.75l / m2 Beuteloberfläche / 24h / bar. Gelagert wurden die Beutel bis zu 72
Stunden. Um bei der Probenentnahme aus dem Beutel aseptische Bedingungen zu
wahren wurde der bei der Probenüberführung bereits verwendete sampling site coupler
desinfiziert und die Probe über eine Kanüle in eine 2ml Spritze abgezogen. Das jeweils
abgezogene Probevolumen betrug 0,7ml.
Die Zellkonzentrationen von Leukozyten, Thrombozyten und Erythrozyten in
den Blutprodukten wurde mit dem ADVIA120-Zellzähler (Bayer HealthCare Diagnos-
tics) bestimmt. Wurde eine Blutgasanalyse zur Bestimmung von pH-Wert, pCO2, pO2,
HCO3- - Konzentration, sO2 und Base Excess (BE) benötigt, so wurde diese mit dem
ABL-5 (Radiometer, Kopenhagen, Dänemark) durchgeführt.
3. Durchflusszytometrische Messungen
3.1. Das Prinzip der Durchflusszytometrie
Die Durchflusszytometrie, auch fluoreszenzaktivierte Zellanalyse („fluorescence
activated cell sorting“, FACS) genannt, ist ein opto-elektronisches Messsystem, bei
dem optische Signale in elektronische Impulse umgewandelt werden und so mit Hilfe
eines EDV-Systems verarbeitet werden können. Dabei werden Streulicht- und Fluores-
zenzeigenschaften von Einzelzellen in Suspension detektiert. Der Aufbau dieser Gerä-
te setzt sich aus drei Teilen zusammen (Abb. 7):

Material und Methoden 21
a. Als Lichtquelle dient ein Laser der Wellenlänge 488nm und ein Dioden-Laser der
Wellenlänge 635nm. In Geräten älterer Bauart werden Quecksilber- oder Xenon-
dampflampen verwendet.
b. In der Analysekammer werden die einzelnen Zellen durch eine schnelle laminare
Strömung perlschnurartig hintereinander aufgereiht und nacheinander durch den
Laserstrahl geführt.
c. Das Detektionssystem besteht aus einer Reihe optischer Filter und Sensoren. Die
emittierten Photonen jeder einzelnen, die Analysekammer passierenden Zelle wer-
den durch die optischen Filter nach Wellenlängen aufgeteilt und an den optischen
Sensoren (FSC, SSC und FL1-FL4) in elektronische Signale umgewandelt.
Abb. 7: Schematischer Aufbau eines Durchflusszytometers (modifiziert nach [19]).
Jede Zellart streut entsprechend ihrer Größe und ihrer Zellstruktur das Licht des
488nm-Lasers auf eine zellspezifische Weise. Das vorwärts gestreute Licht (foreward
light scatter, FSC) ist nach Ausblenden des Zentralstrahls ein Maß für die Größe einer
Zelle. Das im rechten Winkel zum Zentralstrahl gestreute Licht (sideward light scatter,
SSC) ist ein Maß für die Granularität [60, 61]. Die Grundlage der Fluoreszenzmessung
ist das Anfärben von Zellen bzw. der zu untersuchenden Moleküle mit Fluoreszenz-
farbstoffen, meist nach dem Prinzip der Antigen-Antikörper-Reaktion. Bei Bestrahlen
der Fluoreszenzfarbstoffe mit dem 635nm Dioden-Laser in der Durchflusszelle absor-
bieren die Farbstoffe eine für jeden Farbstoff spezifische Menge an Energie und wer-
den dadurch kurzzeitig auf ein höheres Energieniveau gehoben. Bei dem anschließen-
den Absinken auf das ursprüngliche Energieniveau wird die Energiedifferenz in Form
von Licht bestimmter Wellenlänge entsprechend der Planck’schen Wellen-Energie-
Relation E = h x f emittiert. Dabei steht E für die absorbierte und dadurch emittierbare
Energie, h für eine Naturkonstante, das Plancksche Wirkungsquantum, und f für die

Material und Methoden 22
Frequenz und damit indirekt für die Wellenlänge des emittierten Lichts [45]. Da jeder
Fluoreszenzfarbstoff ausschließlich eine bestimmte Energiemenge absorbiert, wird auf
Grund der Planck’schen Wellen-Energie-Relation nur Licht mit einem Intensitätsmaxi-
mum in einem bestimmtem Wellenlängenbereich, der für den Farbstoff charakteristisch
ist, emittiert. Die Sensoren des Durchflusszytometers registrieren das emittierte Licht
und digitalisieren das Signal. Da die Zellen einer Probe in der Durchflusszelle durch die
laminare Strömung hintereinander aufgereiht werden, lassen sich für jede einzelne
Zelle Aussagen bezüglich des Färbeverhaltens treffen. Auf Grund des vorbekannten
Emissionsmaximums jedes Farbstoffes lässt sich mit Hilfe einer geeigneten Computer-
Software das registrierte Signal mit den Messergebnissen der vorwärts und seitwärts
gestreuten Lichter kombinieren und so einem Ereignis, in diesem Fall einer Zelle, zu-
ordnen. Da bei konstanter Leistung des Lasers die Intensität der Lichtemission von der
Zahl der aktivierten Fluoreszenzfarbstoffe abhängt ist, ist mit dieser Methode nicht nur
ein qualitativer Nachweis, sondern auch eine (semi)-quantitative Messung möglich [29].
Die verwendeten Fluoreszenzfarbstoffe sind Fluorescein isothiocyanat (FITC), Pyco-
erythrin (PE) und Allophycocyanin (APC). Das Fluoreszenzmaximum von FITC liegt bei
519nm (gelb-grün), das von PE bei 578nm (rot-orange) und das von APC bei 660nm
(gelb). Für die hier dargestellten Analysen wurde das Durchflusszytometer
FACSCalibur mit der Software CellQuest™ Pro (beides BD Biosciences, San Jose,
CA, USA) verwendet.
3.2 Färbungen
Alle für die in dieser Arbeit durchgeführten Färbungen benötigten Geräte, Pro-
dukte, Chemikalien und Biochemikalien sind im Anhang, Tab. A1 und A2, aufgeführt.
3.2.1 Oberflächenantigenfärbungen
Jeder Zelltyp hat sein spezifisches Oberflächenantigenmuster entsprechend
dem er zu identifizieren ist. Diese Oberflächenantigene werden nach der „cluster of
differentiation“-Klassifikation (CD) in Kombination mit einer Zahl eingeteilt. Hierbei gibt
die zugeordnete Zahl die Reihenfolge der Zuordnung zur CD-Nomenklatur wieder. Alle
Leukozyten haben das an der Oberfläche exprimierte „leukocyte common antigen“,
kurz CD45. Monozyten besitzen zusätzlich das für sie typische Oberflächenantigen
CD14. Zur durchflusszytometrischen Bestimmung des Anteils an CD14+ Monozyten in
peripherem Blut wurde die Blutprobe mit fluoreszenzmarkierten Antikörpern gegen
CD14 und CD45 gefärbt. Die bei der Antikörperherstellung verwendeten Klone und die
an die Antikörper gekoppelten Fluoreszenzfarbstoffe sind in Tab. 1 aufgeführt.

Material und Methoden 23
Tab. 1: Fluoreszenzfarbstoffe und Klone der verwendeten Antikörper.
Antikö rper Klon Fluoreszenzfar bstoff
CD45 HI30 PE
CD14 M5E2 APC
Entsprechend des Färbeprotokolls des Herstellers werden vor der Färbung mit
Antikörpern gegen Oberflächenantigene die Erythrozyten mit PharM-LyseTM bei Raum-
temperatur im Dunkeln lysiert. Nach anschließendem zweimaligem Waschen mit PBS
(Phosphat-gepufferte Salzlösung) und Zentrifugieren mit 500x g für fünf Minuten wird
das Sediment mit 200µl PBS und 5µl des gewünschten Antikörpers pro 1x106 Leukozy-
ten bei 4°C im Dunkeln für 25 Minuten inkubiert. Ab schließend wird die Probe zweimal
mit PBS gewaschen und zur durchflusszytometrischen Messung mit PBS versetzt oder
das Sediment zu weiteren Färbungen verwendet.
3.2.2 Nachweis und Messung von Nekrose und Apoptose
3.2.2.1 Nekrose und 7-Aminoactinomycin D (7-AAD)
7-AAD ist ein Farbstoff, der hochspezifisch an DNS bindet. Bei intakter Plas-
mamembran dringen die Farbstoffe nicht in die Zelle ein. Es kommt zu keiner Färbung.
Bei Verlust der Membranintegrität können 7-AAD-Moleküle durch die löchrige Memb-
ran in die Zelle eindringen und eine Kernfärbung bewirken. Typischerweise ist bei der
Nekrose oder bei der späten Apoptose ein Verlust der Membranintegrität zu finden.
Daher dient diese Färbung als hochspezifischer Nachweis für tote Zellen. Die maxima-
le Fluoreszenzemission von 7-AAD liegt bei 610nm. Das Färbeprotokoll sieht eine Fär-
bung ausschließlich in Kombination mit Annexin V vor. Der Ablauf der Färbung wird
unter dem entsprechenden Unterpunkt erläutert.
3.2.2.2 Apoptose, Phosphatidylserin-Switch und Anne xin V
Wie unter I. 3.2.1 beschrieben kommt es schon früh im Verlauf der Apoptose zu
einem sogenannten Phosphatidylserin-Switch, bei dem Phosphatidylserin vom inneren
Blatt der Plasmamembran auf das äußere wechselt. Annexin V, ursprünglich „vascular
anticoagulant alpha“ (VAC alpha) genannt, bindet spezifisch an Phosphatidylserin [2].
Durch die Koppelung an FITC werden Zellen mit externalisiertem Phosphatidylserin,
also apoptotische Zellen, im Durchflusszytometer dedektierbar. Bei der Nekrose kommt
es zum Verlust der Membranintegrität, wobei die Membran porös wird. Annexin V bin-
det somit auch an nekrotische Zellen, im Gegensatz zur Apoptose aber von intrazellu-
lär. Um einen eventuell beginnenden Zerfall der Plasmamembran nachzuweisen und

Material und Methoden 24
so zwischen Apoptose und Nekrose unterscheiden zu können, erfolgt die gleichzeitige
Färbung von Annexin V und 7-AAD [13, 20, 24, 66]. Dadurch ergeben sich die vier in
Tab. 2 aufgezeigten Möglichkeiten, wie die Zielzellen gefärbt sein können.
Tab. 2: Mögliche Messergebnisse bei gleichzeitiger Färbung mit Annexin V und 7-AAD.
Mögliche Kombinationen für A nnexin -V und 7 -AAD Interpretation der Fä rbung
Annexin V– / 7-AAD– vitale Zellen
Annexin V+ / 7-AAD– (Früh-) Apoptose
Annexin V– / 7-AAD+ Messfehler
Annexin V+ / 7-AAD+ Nekrose / Spätapoptose
Aus den Bindungseigenschaften von Annexin V und 7-AAD ergibt sich, dass ei-
ne gleichzeitige Bindung an 7-AAD ohne Bindung von Annexin V nicht möglich ist.
Damit spricht das Ausbleiben von Annexin V–/7-AAD+ Ereignissen für eine qualitativ
hochwertige Messung. Die Färbung mit Annexin V erfolgt nach der Färbung mit Anti-
körpern gegen Oberflächenantigene. Nach letztmaligem Waschen der Zellen wird das
Sediment mit 1ml Bindungspuffer (1x) pro 1x106 Leukozyten versetzt. Davon werden
100µl bei Raumtemperatur im Dunkeln mit je 5µl Annexin V und 7-AAD inkubiert, an-
schließend mit 400µl unverdünntem Bindungspuffer versetzt und durchflusszytomet-
risch gemessen.
3.3 Instrument Settings
In dieser Arbeit wurden die Zellen mit vier Fluoreszenzfarbstoffen gefärbt. Wie
unter 3.1 angeführt hat jeder Farbstoff ein photometrisches Intensitätsmaximum mit der
Wellenlänge des nach Aktivierung emittierten Lichts. Die Farbstoffe wurden so gewählt,
dass sich die Wellenlängen der Intensitätsmaxima der einzelnen Farbstoffe unter-
scheiden. Da aber die Farbstoffe nicht monochromes Licht mit ausschließlich einer
Wellenlänge, sondern zusätzlich Licht verschiedener Wellenlängen mit reduzierter In-
tensität emittieren, kommt es zu Überschneidungen der zusätzlich zum Intensitätsma-
ximum emittierten Wellenlängen zwischen den verschiedenen Farbstoffen. Beispiels-
weise emittiert der Farbstoff A auch Licht der Wellenlänge des Intensitätsmaximums
des Farbstoffes B. Dadurch werden falsch positive Ereignisse am die Wellenlänge des
Farbstoffes B registrierenden Sensors aufgezeichnet. Um diese falsch positiven Mess-
werte auszuschließen müssen in der verwendeten Computer-Software die Messungen
der Sensoren gegeneinander abgeglichen werden. Diese gegenseitige Kompensation
der Sensoren wird in den Geräteeinstellungen des Durchflusszytometers festgelegt.
Hier kann auch festgelegt werden, ob Messereignisse bis zu einer bestimmten Größe
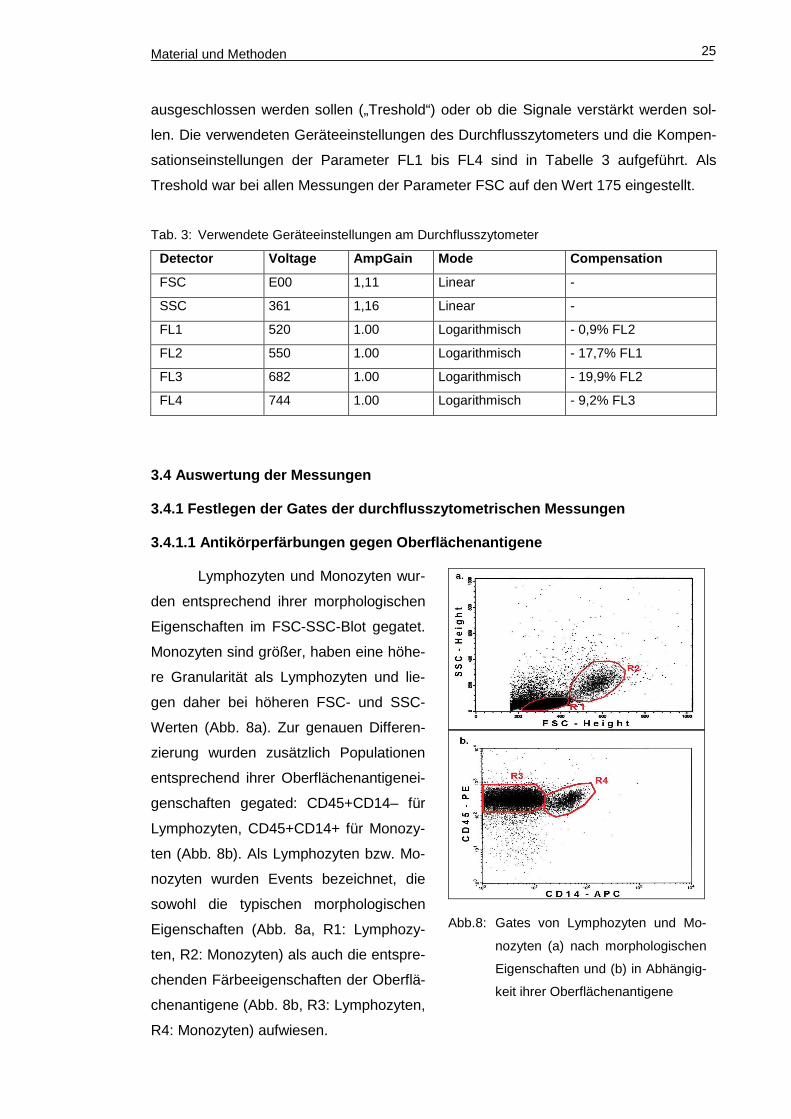
Material und Methoden 25
Abb.8: Gates von Lymphozyten und Mo-
nozyten (a) nach morphologischen
Eigenschaften und (b) in Abhängig-
keit ihrer Oberflächenantigene
ausgeschlossen werden sollen („Treshold“) oder ob die Signale verstärkt werden sol-
len. Die verwendeten Geräteeinstellungen des Durchflusszytometers und die Kompen-
sationseinstellungen der Parameter FL1 bis FL4 sind in Tabelle 3 aufgeführt. Als
Treshold war bei allen Messungen der Parameter FSC auf den Wert 175 eingestellt.
Tab. 3: Verwendete Geräteeinstellungen am Durchflusszytometer
Detector Voltage AmpGain Mode Compensation
FSC E00 1,11 Linear -
SSC 361 1,16 Linear -
FL1 520 1.00 Logarithmisch - 0,9% FL2
FL2 550 1.00 Logarithmisch - 17,7% FL1
FL3 682 1.00 Logarithmisch - 19,9% FL2
FL4 744 1.00 Logarithmisch - 9,2% FL3
3.4 Auswertung der Messungen
3.4.1 Festlegen der Gates der durchflusszytometrisc hen Messungen
3.4.1.1 Antikörperfärbungen gegen Oberflächenantige ne
Lymphozyten und Monozyten wur-
den entsprechend ihrer morphologischen
Eigenschaften im FSC-SSC-Blot gegatet.
Monozyten sind größer, haben eine höhe-
re Granularität als Lymphozyten und lie-
gen daher bei höheren FSC- und SSC-
Werten (Abb. 8a). Zur genauen Differen-
zierung wurden zusätzlich Populationen
entsprechend ihrer Oberflächenantigenei-
genschaften gegated: CD45+CD14– für
Lymphozyten, CD45+CD14+ für Monozy-
ten (Abb. 8b). Als Lymphozyten bzw. Mo-
nozyten wurden Events bezeichnet, die
sowohl die typischen morphologischen
Eigenschaften (Abb. 8a, R1: Lymphozy-
ten, R2: Monozyten) als auch die entspre-
chenden Färbeeigenschaften der Oberflä-
chenantigene (Abb. 8b, R3: Lymphozyten,
R4: Monozyten) aufwiesen.

Material und Methoden 26
3.4.1.2 Färbungen mit Annexin V und 7-AAD
Wie in Tab. 2 (Seite 24) aufgezeigt, gibt es vier mögliche Kombinationen bei der
Färbung mit Annexin V und 7-AAD. Die Einteilung der gemessenen Ereignisse erfolgt
daher am besten mit Hilfe von Quadranten (Abb. 9). Wie bereits dargestellt, gelten An-
nexin V+/7-AAD+ Zellen als spät-apoptotisch bzw. nekrotisch, da die Färbung mit 7-
AAD ein Kernfärbung ist. Diese nur stattfinden kann, wenn die Zellmembran einer Zelle
undicht wird, so wie es während der Nekrose oder in späten Stadien der Apoptose ge-
schieht. Zellen die Annexin V+, aber 7-AAD- sind, gelten somit als früh-apoptotisch
bzw. apoptotisch.
Da aber bei der Zuordnung der durchflusszytometrisch registrierten Ereignisse
zu Lymphozyten und Monozyten nicht nur die Färbeeigenschaften gegen Oberflä-
chenantigene der jeweiligen Zellen, sondern auch deren Größe und Granularität be-
achtet wurden, konnten keine Annexin V+/7-AAD+ Lymphozyten oder Monozyten dar-
gestellt werden. Wie unter Punkt 3 der Einleitung beschrieben, ändert sich im Verlauf
der Apoptose und der Nekrose die Zellmorphologie. Die Zellen fallen also aus den für
sie typischen Regionen (Abb. 8a, R1: Lymphozyten, R2: Monozyten) heraus und wer-
den so nicht in die Auswertung einbezogen. Dies hat zwei Folgen: zum einen können
die bei dieser Gating-Strategie die als Lymphozyten beziehungsweise Monozyten be-
zeichneten Events allein bezüglich ihrer Annexin V - Färbeeigenschaften bewertet wer-
den, da, wie eben beschrieben, weder Lymphozyten noch Monozyten 7-AAD-positiv
sein können. Bei der Differenzierung zwischen Annexin V-negativ und Annexin V-
positiv kann somit auch auf ein Histogramm, welches eine eindeutigere graphische
Abb. 9: Einteilung in Quadranten nach Annexin V-
und 7-AAD- Doppelfärbung.
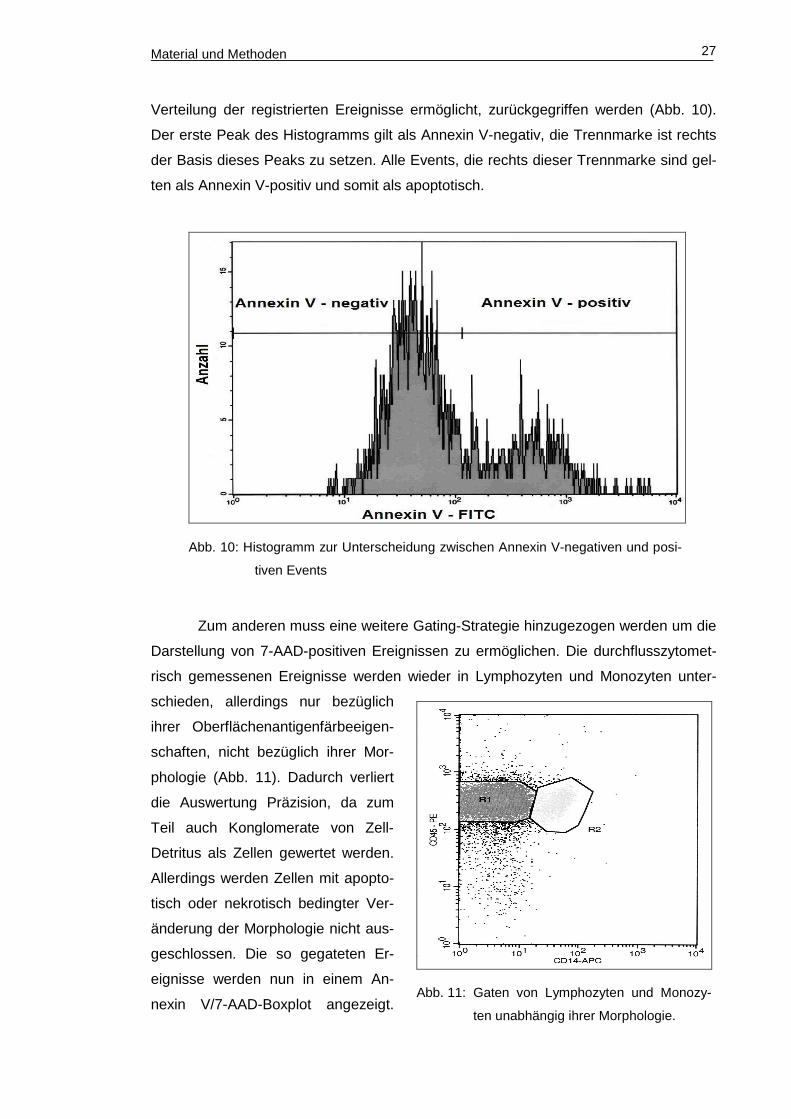
Material und Methoden 27
Abb. 11: Gaten von Lymphozyten und Monozy-
ten unabhängig ihrer Morphologie.
Verteilung der registrierten Ereignisse ermöglicht, zurückgegriffen werden (Abb. 10).
Der erste Peak des Histogramms gilt als Annexin V-negativ, die Trennmarke ist rechts
der Basis dieses Peaks zu setzen. Alle Events, die rechts dieser Trennmarke sind gel-
ten als Annexin V-positiv und somit als apoptotisch.
Zum anderen muss eine weitere Gating-Strategie hinzugezogen werden um die
Darstellung von 7-AAD-positiven Ereignissen zu ermöglichen. Die durchflusszytomet-
risch gemessenen Ereignisse werden wieder in Lymphozyten und Monozyten unter-
schieden, allerdings nur bezüglich
ihrer Oberflächenantigenfärbeeigen-
schaften, nicht bezüglich ihrer Mor-
phologie (Abb. 11). Dadurch verliert
die Auswertung Präzision, da zum
Teil auch Konglomerate von Zell-
Detritus als Zellen gewertet werden.
Allerdings werden Zellen mit apopto-
tisch oder nekrotisch bedingter Ver-
änderung der Morphologie nicht aus-
geschlossen. Die so gegateten Er-
eignisse werden nun in einem An-
nexin V/7-AAD-Boxplot angezeigt.
Abb. 10: Histogramm zur Unterscheidung zwischen Annexin V-negativen und posi-
tiven Events

Material und Methoden 28
Da die Aufteilung in Annexin V-/7-AAD- und Annexin V+/7-AAD- bereits erfolgte, wer-
den hierbei lediglich die Annexin V+/7-AAD+ Ereignisse bewertet und der prozentuelle
Anteil an Lymphozyten beziehungsweise Monozyten angegeben.
3.4.2 Statistische Auswertung
Die statistische Auswertung erfolgte mit dem Programm PASW, Version 18. Mit
Hilfe der explorativen Datenanalyse wurden Mittelwert, Standardabweichung, Spann-
weite und Median berechnet. Der Test auf Normalverteilung der Daten erfolgte nach
Kolmogorov [33]. Zum Vergleich der Daten der Apheresemethoden und der Daten zu
den verschiedenen Zeitpunkten nach Spende wurde das generalisierte lineare Model
verwendet, gefolgt von post-hoc-Tests entsprechend der Methode von Sidak zur An-
passung der p-Werte [23]. Um den Zusammenhang zwischen verschiedenen Variablen
zu erfassen wurde der Korrelationskoeffizient nach Pearson verwendet [43]. Alle Hypo-
thesentests waren zweiseitig und wurden als signifikant angesehen, wenn die p-Werte
kleiner als 0,05 waren und als hochsignifikant, wenn die p-Werte kleiner als 0,01 wa-
ren.
4. Verwendete Formeln
a. Totales Blutvolumen (TBV), Berechnung nach Nadler [39]:
♂: TBV = (0,3669 x (Größe in m)3 + 0,03219 x (Gewicht in kg) + 0,6041) x 1000 [l]
♀: TBV = (0,3561 x (Größe in m)3 + 0,03308 x (Gewicht in kg) + 0,1833) x 1000 [l]
b. Body Mass Index (BMI):
BMI = (Gewicht in kg) / (Größe in m)2 [kg/m2]
c. Zellgehalt eines Blutproduktes:
Zellgehalt = (Zell-Konzentration in Zellen / ml) x (Produktvolumen in ml)
5. Versuchsreihen zur Beantwortung der Fragestellun gen
5.1 Vergleich von Leukozytenreduktionskammern zweie r Zellseparatoren
Es wurde in einer 2007 veröffentlichten Studie über Leukozytenreduktionskam-
mern des Trima Accel® Zellseparators von einer hervorragenden Menge und Qualität
mononukleärer Zellen des peripheren Blutes in den Kammern berichtet. [6] Wie unter
Material und Methoden, Punkt 1.3 beschrieben wird beim COBE® Spectra Zellsepara-
tor ebenfalls eine geometrisch gleich geformte Leukozytenreduktionskammer zur Leu-
kozytenreduktion verwendet. Eventuelle Unterschiede zwischen dem Blut aus den

Material und Methoden 29
Leukozytenreduktionskammern beider Zellseparatoren wurden untersucht, indem der
Gehalt von Leukozyten, mononukleären Zellen und Restzellen von zehn Leukozyten-
reduktionskammern der COBE® Spectra und von 27 Leukozytenreduktionskammern
der Trima Accel® analysiert wurden. Die Auswahl der Spender der Thrombozytaphe-
resen, deren Leukozytenreduktionskammern verwendet wurden, war zufällig. Innerhalb
eines Zeitraums von zwei Stunden nach der Thrombozytapherese wurde der Inhalt der
gefüllten Leukozytenreduktionskammern in Röhrchen mit dem Volumen 20 ml über-
führt. Die Zellkonzentrationen wurden mit dem ADVIA120 bestimmt, der prozentuelle
Anteil mononukleärer Zellen durchflusszytometrisch entsprechend der oben beschrie-
benen Färbemethoden gegen Oberflächenantigene (Material und Methoden, Punkt
3.2.1).
5.2 Zellgehalt und Zellschädigung in Leukozytenredu ktionskammern
Um eventuelle Einflüsse auf den Gehalt mononukleärer Zellen in den Leukozy-
tenreduktionskammern sowie auf die Qualität der Zellen im Sinne einer Zellschädigung
durch unterschiedliche Spendeparameter wie z.B. die Spendedauer, zu erkennen soll-
ten die Leukozytenreduktionskammern des Trima Accel® Zellseparators nach einfa-
chen Thrombozytapheresen (E-TA) mit denen nach doppelten Thrombozytapheresen
(D-TA) des selben Geräts verglichen werden. Des Weiteren sollte das Blut der Leuko-
zytenreduktionskammern kurzzeitgelagert werden, um aus einer eventuell zu beobach-
tenden Dynamik innerhalb des Blutes, wie Änderungen des Zellgehalts oder des An-
teils geschädigter Zellen, Aussagen zur Qualität der aus Leukozytenreduktionskam-
mern gewonnenen Zellen zu treffen. Dazu wurden je neun Leukozytenreduktionskam-
mern nach E-TA und nach D-TA innerhalb eines Zeitraums von maximal einer Stunde
nach Spende in PVC-Beutel unter aseptischen Bedingungen umgefüllt. Die Auswahl
der Thrombozytapheresen, deren Leukozytenreduktionskammern verwendet wurden,
erfolgte zufällig. Die mit dem Blut der Leukozytenreduktionskammern gefüllten Beutel
wurden für 72 Stunden entsprechend der unter Material und Methoden, Punkt 2 ange-
führten Bedingungen gelagert. Zu den Messzeitpunkten 1 Stunde, 6 Stunden, 24 Stun-
den, 48 Stunden und 72 Stunden nach Ende der Apherese wurden über eine Kanüle
Proben mit je ca. 0,7 ml Blut aus dem gelagerten Material entnommen. Bestimmt wur-
den das Blutbild (Leukozyten, Thrombozyten und Erythrozyten) mit dem ADVIA120
Zellzähler und durchflusszytometrisch der Anteil der CD45+, CD14+, Annexin V+ und
7-AAD+ Events.

Material und Methoden 30
5.3 Blut aus Leukozytenreduktionskammern und aus Le ukapheresaten
Um Unterschiede zwischen der Qualität mononukleärer Zellen des peripheren
Bluts aus Leukozytenreduktionskammern und aus Leukapheresaten zu erforschen,
wurde Blut aus je sechs zufällig ausgewählten Leukozytenreduktionskammern nach
E-TA und nach D-TA sowie Proben aus sechs zufällig ausgewählten Leukapherese-
produkten entnommen. Innerhalb einer Stunde nach Ende der Apherese wurde dieses
Blut in PVC-Beutel umgefüllt und über 48 Stunden unter den unter Material und Me-
thoden, Punkt 2 beschriebenen Bedingungen gelagert. Zu den Messzeitpunkten 1
Stunde, 6 Stunden, 24 Stunden und 48 Stunden nach Ende der Apherese wurden über
eine Kanüle Proben mit je ca. 0,7 ml Blut aus dem gelagerten Material entnommen.
Bestimmt wurden das Blutbild mit dem ADVIA120 Zellzähler und der pH-Wert sowie
die Blutgase mit dem Abl-5. Außerdem wurden die Proben mit CD14-APC- und CD45-
PE- Antikörpern, mit Annexin-V-FITC und mit 7-AAD gefärbt und durchflusszytomet-
risch analysiert.
Ergebnisse 31
V. Ergebnisse
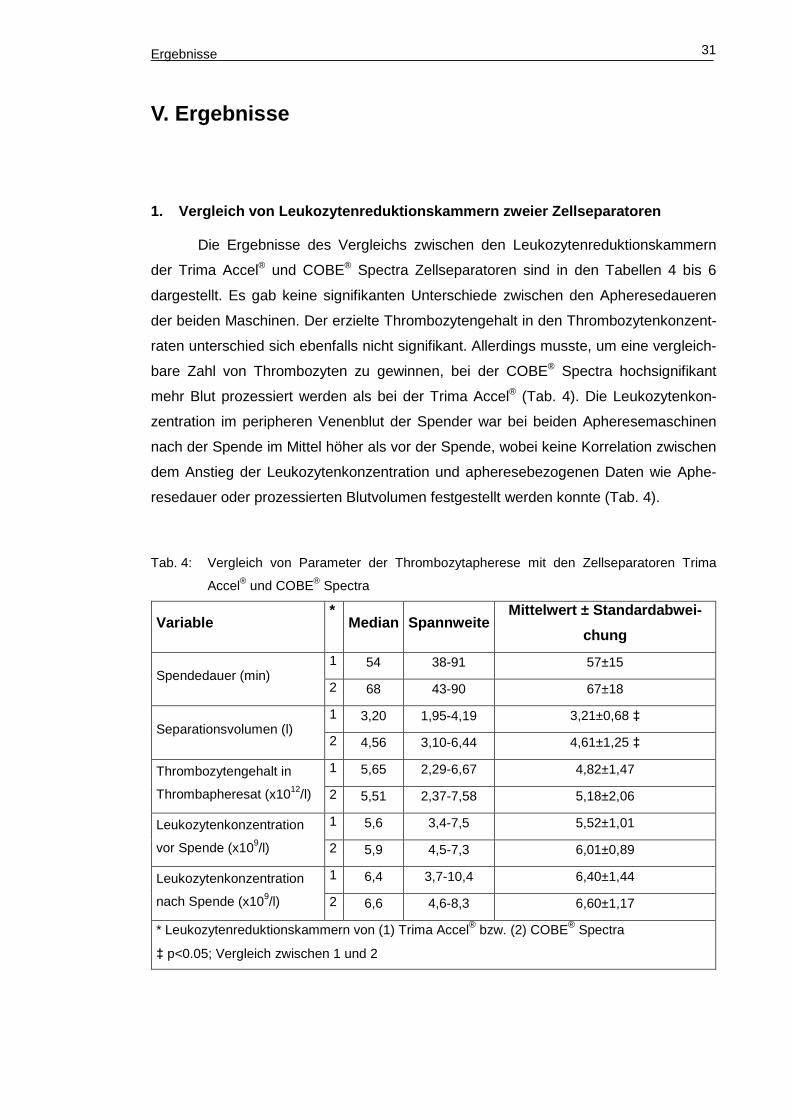
1. Vergleich von Leukozytenreduktionskammern zweie r Zellseparatoren
Die Ergebnisse des Vergleichs zwischen den Leukozytenreduktionskammern
der Trima Accel® und COBE® Spectra Zellseparatoren sind in den Tabellen 4 bis 6
dargestellt. Es gab keine signifikanten Unterschiede zwischen den Apheresedaueren
der beiden Maschinen. Der erzielte Thrombozytengehalt in den Thrombozytenkonzent-
raten unterschied sich ebenfalls nicht signifikant. Allerdings musste, um eine vergleich-
bare Zahl von Thrombozyten zu gewinnen, bei der COBE® Spectra hochsignifikant
mehr Blut prozessiert werden als bei der Trima Accel® (Tab. 4). Die Leukozytenkon-
zentration im peripheren Venenblut der Spender war bei beiden Apheresemaschinen
nach der Spende im Mittel höher als vor der Spende, wobei keine Korrelation zwischen
dem Anstieg der Leukozytenkonzentration und apheresebezogenen Daten wie Aphe-
resedauer oder prozessierten Blutvolumen festgestellt werden konnte (Tab. 4).
Tab. 4: Vergleich von Parameter der Thrombozytapherese mit den Zellseparatoren Trima
Accel® und COBE® Spectra
Variable *
Median Spannweite Mittelwert ± Standardabwe i-
chung
Spendedauer (min) 1 54 38-91 57±15
2 68 43-90 67±18
Separationsvolumen (l) 1 3,20 1,95-4,19 3,21±0,68 ‡
2 4,56 3,10-6,44 4,61±1,25 ‡
Thrombozytengehalt in
Thrombapheresat (x1012/l)
1 5,65 2,29-6,67 4,82±1,47
2 5,51 2,37-7,58 5,18±2,06
Leukozytenkonzentration
vor Spende (x109/l)
1 5,6 3,4-7,5 5,52±1,01
2 5,9 4,5-7,3 6,01±0,89
Leukozytenkonzentration
nach Spende (x109/l)
1 6,4 3,7-10,4 6,40±1,44
2 6,6 4,6-8,3 6,60±1,17
* Leukozytenreduktionskammern von (1) Trima Accel® bzw. (2) COBE® Spectra
‡ p<0.05; Vergleich zwischen 1 und 2
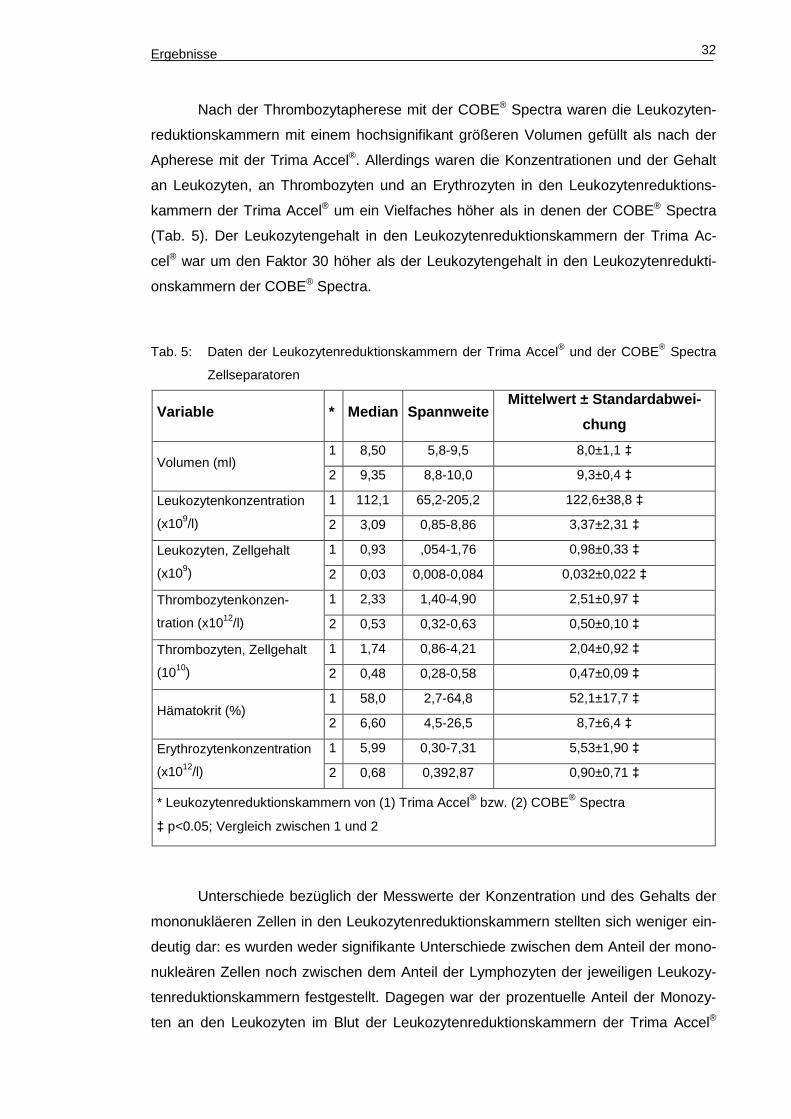
Ergebnisse 32
Nach der Thrombozytapherese mit der COBE® Spectra waren die Leukozyten-
reduktionskammern mit einem hochsignifikant größeren Volumen gefüllt als nach der
Apherese mit der Trima Accel®. Allerdings waren die Konzentrationen und der Gehalt
an Leukozyten, an Thrombozyten und an Erythrozyten in den Leukozytenreduktions-
kammern der Trima Accel® um ein Vielfaches höher als in denen der COBE® Spectra
(Tab. 5). Der Leukozytengehalt in den Leukozytenreduktionskammern der Trima Ac-
cel® war um den Faktor 30 höher als der Leukozytengehalt in den Leukozytenredukti-
onskammern der COBE® Spectra.
Tab. 5: Daten der Leukozytenreduktionskammern der Trima Accel® und der COBE® Spectra
Zellseparatoren
Variable * Median Spannweite Mittelwert ± Standardabwe i-
chung
Volumen (ml) 1 8,50 5,8-9,5 8,0±1,1 ‡
2 9,35 8,8-10,0 9,3±0,4 ‡
Leukozytenkonzentration
(x109/l)
1 112,1 65,2-205,2 122,6±38,8 ‡
2 3,09 0,85-8,86 3,37±2,31 ‡
Leukozyten, Zellgehalt
(x109)
1 0,93 ,054-1,76 0,98±0,33 ‡
2 0,03 0,008-0,084 0,032±0,022 ‡
Thrombozytenkonzen-
tration (x1012/l)
1 2,33 1,40-4,90 2,51±0,97 ‡
2 0,53 0,32-0,63 0,50±0,10 ‡
Thrombozyten, Zellgehalt
(1010)
1 1,74 0,86-4,21 2,04±0,92 ‡
2 0,48 0,28-0,58 0,47±0,09 ‡
Hämatokrit (%) 1 58,0 2,7-64,8 52,1±17,7 ‡
2 6,60 4,5-26,5 8,7±6,4 ‡
Erythrozytenkonzentration
(x1012/l)
1 5,99 0,30-7,31 5,53±1,90 ‡
2 0,68 0,392,87 0,90±0,71 ‡
* Leukozytenreduktionskammern von (1) Trima Accel® bzw. (2) COBE® Spectra
‡ p<0.05; Vergleich zwischen 1 und 2
Unterschiede bezüglich der Messwerte der Konzentration und des Gehalts der
mononukläeren Zellen in den Leukozytenreduktionskammern stellten sich weniger ein-
deutig dar: es wurden weder signifikante Unterschiede zwischen dem Anteil der mono-
nukleären Zellen noch zwischen dem Anteil der Lymphozyten der jeweiligen Leukozy-
tenreduktionskammern festgestellt. Dagegen war der prozentuelle Anteil der Monozy-
ten an den Leukozyten im Blut der Leukozytenreduktionskammern der Trima Accel®
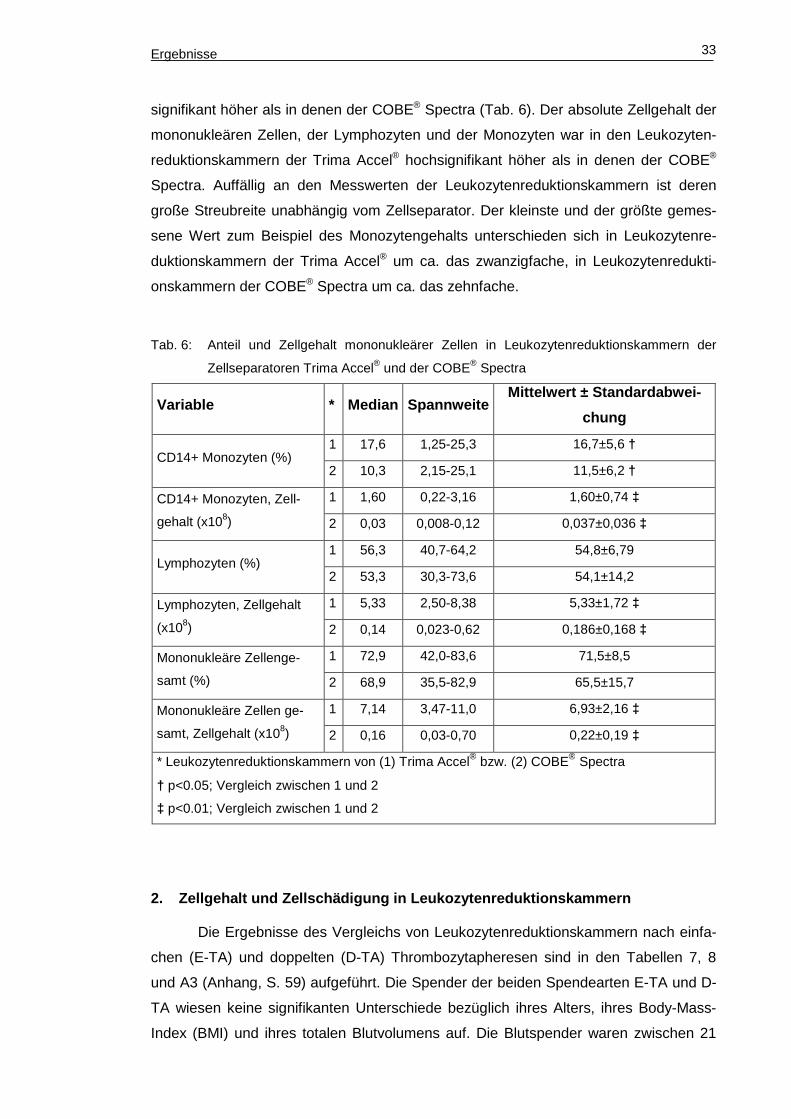
Ergebnisse 33
signifikant höher als in denen der COBE® Spectra (Tab. 6). Der absolute Zellgehalt der
mononukleären Zellen, der Lymphozyten und der Monozyten war in den Leukozyten-
reduktionskammern der Trima Accel® hochsignifikant höher als in denen der COBE®
Spectra. Auffällig an den Messwerten der Leukozytenreduktionskammern ist deren
große Streubreite unabhängig vom Zellseparator. Der kleinste und der größte gemes-
sene Wert zum Beispiel des Monozytengehalts unterschieden sich in Leukozytenre-
duktionskammern der Trima Accel® um ca. das zwanzigfache, in Leukozytenredukti-
onskammern der COBE® Spectra um ca. das zehnfache.
Tab. 6: Anteil und Zellgehalt mononukleärer Zellen in Leukozytenreduktionskammern der
Zellseparatoren Trima Accel® und der COBE® Spectra
Variable * Median Spannweite Mittelwert ± Standardabwe i-
chung
CD14+ Monozyten (%) 1 17,6 1,25-25,3 16,7±5,6 †
2 10,3 2,15-25,1 11,5±6,2 †
CD14+ Monozyten, Zell-
gehalt (x108)
1 1,60 0,22-3,16 1,60±0,74 ‡
2 0,03 0,008-0,12 0,037±0,036 ‡
Lymphozyten (%) 1 56,3 40,7-64,2 54,8±6,79
2 53,3 30,3-73,6 54,1±14,2
Lymphozyten, Zellgehalt
(x108)
1 5,33 2,50-8,38 5,33±1,72 ‡
2 0,14 0,023-0,62 0,186±0,168 ‡
Mononukleäre Zellenge-
samt (%)
1 72,9 42,0-83,6 71,5±8,5
2 68,9 35,5-82,9 65,5±15,7
Mononukleäre Zellen ge-
samt, Zellgehalt (x108)
1 7,14 3,47-11,0 6,93±2,16 ‡
2 0,16 0,03-0,70 0,22±0,19 ‡
* Leukozytenreduktionskammern von (1) Trima Accel® bzw. (2) COBE® Spectra
† p<0.05; Vergleich zwischen 1 und 2
‡ p<0.01; Vergleich zwischen 1 und 2
2. Zellgehalt und Zellschädigung in Leukozytenreduk tionskammern
Die Ergebnisse des Vergleichs von Leukozytenreduktionskammern nach einfa-
chen (E-TA) und doppelten (D-TA) Thrombozytapheresen sind in den Tabellen 7, 8
und A3 (Anhang, S. 59) aufgeführt. Die Spender der beiden Spendearten E-TA und D-
TA wiesen keine signifikanten Unterschiede bezüglich ihres Alters, ihres Body-Mass-
Index (BMI) und ihres totalen Blutvolumens auf. Die Blutspender waren zwischen 21
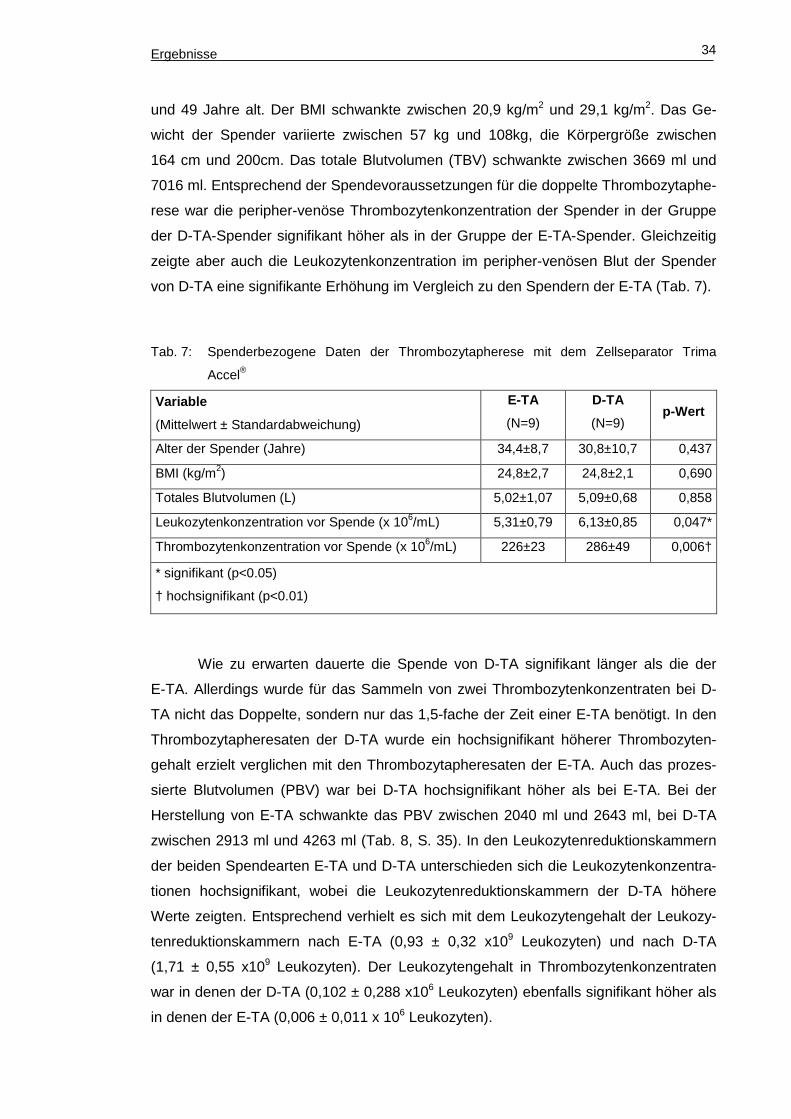
Ergebnisse 34
und 49 Jahre alt. Der BMI schwankte zwischen 20,9 kg/m2 und 29,1 kg/m2. Das Ge-
wicht der Spender variierte zwischen 57 kg und 108kg, die Körpergröße zwischen
164 cm und 200cm. Das totale Blutvolumen (TBV) schwankte zwischen 3669 ml und
7016 ml. Entsprechend der Spendevoraussetzungen für die doppelte Thrombozytaphe-
rese war die peripher-venöse Thrombozytenkonzentration der Spender in der Gruppe
der D-TA-Spender signifikant höher als in der Gruppe der E-TA-Spender. Gleichzeitig
zeigte aber auch die Leukozytenkonzentration im peripher-venösen Blut der Spender
von D-TA eine signifikante Erhöhung im Vergleich zu den Spendern der E-TA (Tab. 7).
Tab. 7: Spenderbezogene Daten der Thrombozytapherese mit dem Zellseparator Trima
Accel®
Variable
(Mittelwert ± Standardabweichung)
E-TA
(N=9)
D-TA
(N=9) p-Wert
Alter der Spender (Jahre) 34,4±8,7 30,8±10,7 0,437
BMI (kg/m2) 24,8±2,7 24,8±2,1 0,690
Totales Blutvolumen (L) 5,02±1,07 5,09±0,68 0,858
Leukozytenkonzentration vor Spende (x 106/mL) 5,31±0,79 6,13±0,85 0,047*
Thrombozytenkonzentration vor Spende (x 106/mL) 226±23 286±49 0,006†
* signifikant (p<0.05)
† hochsignifikant (p<0.01)
Wie zu erwarten dauerte die Spende von D-TA signifikant länger als die der
E-TA. Allerdings wurde für das Sammeln von zwei Thrombozytenkonzentraten bei D-
TA nicht das Doppelte, sondern nur das 1,5-fache der Zeit einer E-TA benötigt. In den
Thrombozytapheresaten der D-TA wurde ein hochsignifikant höherer Thrombozyten-
gehalt erzielt verglichen mit den Thrombozytapheresaten der E-TA. Auch das prozes-
sierte Blutvolumen (PBV) war bei D-TA hochsignifikant höher als bei E-TA. Bei der
Herstellung von E-TA schwankte das PBV zwischen 2040 ml und 2643 ml, bei D-TA
zwischen 2913 ml und 4263 ml (Tab. 8, S. 35). In den Leukozytenreduktionskammern
der beiden Spendearten E-TA und D-TA unterschieden sich die Leukozytenkonzentra-
tionen hochsignifikant, wobei die Leukozytenreduktionskammern der D-TA höhere
Werte zeigten. Entsprechend verhielt es sich mit dem Leukozytengehalt der Leukozy-
tenreduktionskammern nach E-TA (0,93 ± 0,32 x109 Leukozyten) und nach D-TA
(1,71 ± 0,55 x109 Leukozyten). Der Leukozytengehalt in Thrombozytenkonzentraten
war in denen der D-TA (0,102 ± 0,288 x106 Leukozyten) ebenfalls signifikant höher als
in denen der E-TA (0,006 ± 0,011 x 106 Leukozyten).
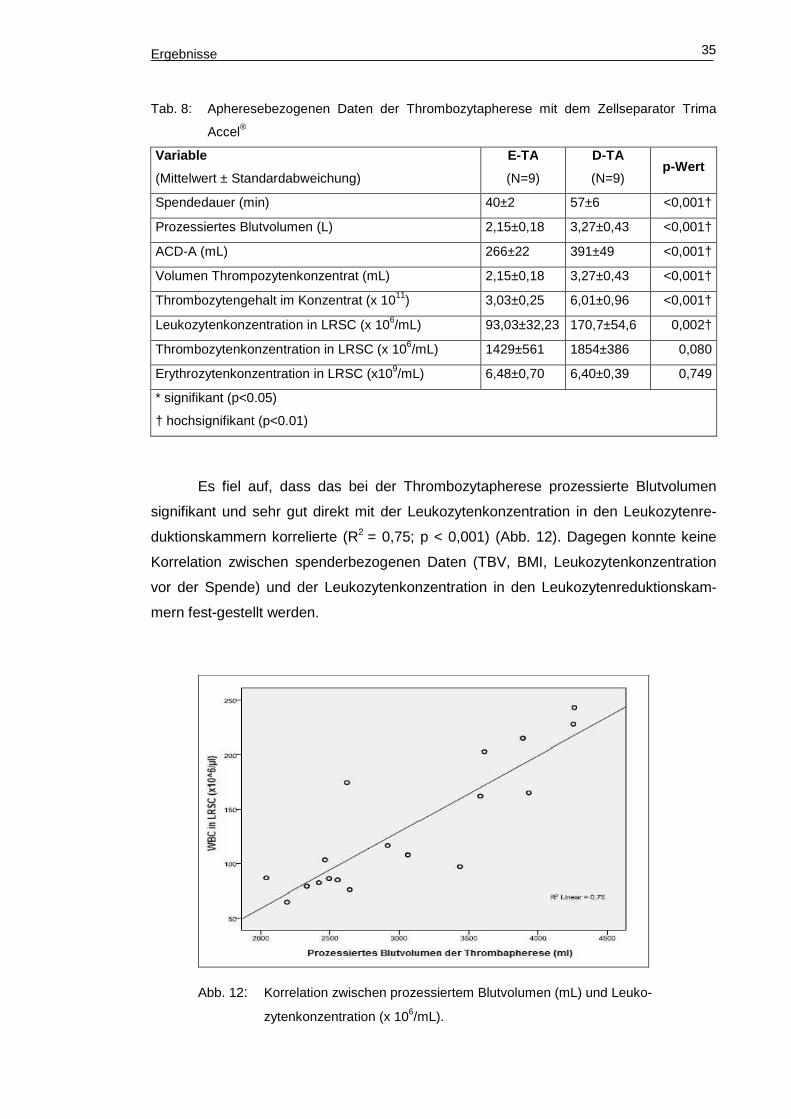
Ergebnisse 35
Tab. 8: Apheresebezogenen Daten der Thrombozytapherese mit dem Zellseparator Trima
Accel®
Variable
(Mittelwert ± Standardabweichung)
E-TA
(N=9)
D-TA
(N=9) p-Wert
Spendedauer (min) 40±2 57±6 <0,001†
Prozessiertes Blutvolumen (L) 2,15±0,18 3,27±0,43 <0,001†
ACD-A (mL) 266±22 391±49 <0,001†
Volumen Thrompozytenkonzentrat (mL) 2,15±0,18 3,27±0,43 <0,001†
Thrombozytengehalt im Konzentrat (x 1011) 3,03±0,25 6,01±0,96 <0,001†
Leukozytenkonzentration in LRSC (x 106/mL) 93,03±32,23 170,7±54,6 0,002†
Thrombozytenkonzentration in LRSC (x 106/mL) 1429±561 1854±386 0,080
Erythrozytenkonzentration in LRSC (x109/mL) 6,48±0,70 6,40±0,39 0,749
* signifikant (p<0.05)
† hochsignifikant (p<0.01)
Es fiel auf, dass das bei der Thrombozytapherese prozessierte Blutvolumen
signifikant und sehr gut direkt mit der Leukozytenkonzentration in den Leukozytenre-
duktionskammern korrelierte (R2 = 0,75; p < 0,001) (Abb. 12). Dagegen konnte keine
Korrelation zwischen spenderbezogenen Daten (TBV, BMI, Leukozytenkonzentration
vor der Spende) und der Leukozytenkonzentration in den Leukozytenreduktionskam-
mern fest-gestellt werden.
Abb. 12: Korrelation zwischen prozessiertem Blutvolumen (mL) und Leuko-
zytenkonzentration (x 106/mL).

Ergebnisse 36
Bezüglich der Thrombozytenkonzentration wurden weder in den Thrombozy-
tenkonzentraten (E-TA, 1208 ± 78 x106/ml; D-TA, 1128 ± 78 x106/ml; p=0,058) noch in
den Leukozytenreduktionskammern signifikante Unterschiede zwischen den beiden
Spendearten E-TA und D-TA gefunden. Allerdings ist sowohl im Blut der Thrombozy-
tenkonzentrate als auch im Blut der Leukozytenreduktionskammern eine Tendenz zu
höheren Werten der Thrombozytenkonzentrtaion nach doppelter Thrombozytapherese
zu erkennen. Bei der Herstellung von E-TA wurde in mehr Fällen und damit mit einem
größeren Gesamtvolumen Plasma parallel zum Thrombozytenkonzentrat gesammelt
(n=9, 400 ± 108 ml) als bei der Herstellung von D-TA (n=6, 194 ± 8 ml). In den Leuko-
zytenreduktionskammern nach E-TA mit unterschiedlichen Volumina von nebenher
gesammeltem Plasma (298 ± 52 ml im Gegensatz zu 482 ± 49 ml, p=0,01) unterschie-
den sich der Gehalt an Leukozyten und der Gehalt mononukleärer Zellen nur gering
und nicht signifikant.
Tabelle A3 (Anhang, S. 59) zeigt die Ergebnisse der durchflusszytometrischen
Messungen im Verlauf der Lagerung über 72 Stunden. Die Leukozytenkonzentration
blieb ebenso wie die Lymphozytenkonzentration über die gesamte Lagerdauer auf ei-
nem stabilen Niveau. Die Monozytenkonzentration fiel dagegen nach einer Lagerdauer
von 48 Stunden signifikant auf etwa die Hälfte des Ausgangswertes ab. Der Anteil an
Annexin V+/7-AAD– Lymphozyten aus Leukozytenreduktionskammern sowohl von E-
TA als auch von D-TA schwankte während der Lagerung ohne signifikante Unterschie-
de zwischen 21% und 28%. Zwischen E-TA und D-TA waren hier ebenfalls keine signi-
fikanten Unterschiede festzustellen. Dagegen stieg der Anteil an Annexin V+/7-AAD+
Lymphozyten signifikant während der Lagerung der Zellen aus Leukozytenreduktions-
kammern nach E-TA von 1,97% ± 0,96% (1 Stunde) auf 17,41% ± 10,02% (72 Stun-
den) beziehungsweise aus denen nach D-TA von 4,34% ± 2,28% (1 Stunde) auf
25,23% ± 18,35% (72 Stunden) an. Der prozentuelle Anteil von CD14+ Monozyten an
Leukozyten war in den Leukozytenreduktionskammern der D-TA hochsignifikant höher
als in denen der E-TA. Zwischen den Messzeitpunkten nach 48 Stunden und nach 72
Stunden fiel die Konzentration der gelagerten CD14+ Monozyten signifikant verglichen
zum Ausgangswert ab: in E-TA um 38% (p<0,01), in D-TA um 47% (p<0,01). Bezüglich
der Bindung von Annexin V an Monozyten zeigte sich direkt nach der Thrombozyta-
pherese eine hohe Bindungsrate (E-TA, 48,48% ± 8,83%; D-TA, 39,12% ± 10,97%),
die binnen 24-stündiger Lagerung in E-TA um 19% auf 29,88% ± 5,93% (p<0,01), in D-
TA um 8% auf 31,44% ± 7,50% abfiel. Im weiteren Verlauf der Lagerung stieg der An-
teil Annexin V+/7-AAD– Monozyten kontinuierlich bis auf ca. 70% in den Produkten
beider Spendearten an (Abb. 13).
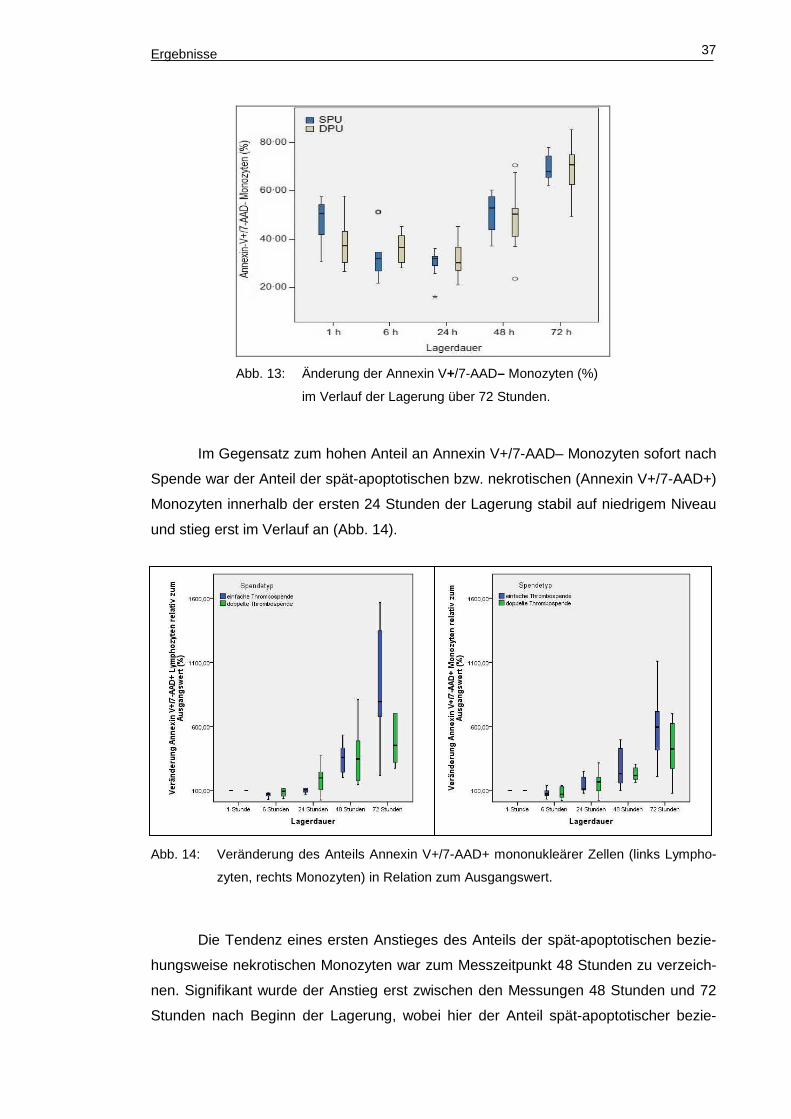
Ergebnisse 37
Im Gegensatz zum hohen Anteil an Annexin V+/7-AAD– Monozyten sofort nach
Spende war der Anteil der spät-apoptotischen bzw. nekrotischen (Annexin V+/7-AAD+)
Monozyten innerhalb der ersten 24 Stunden der Lagerung stabil auf niedrigem Niveau
und stieg erst im Verlauf an (Abb. 14).
Abb. 14: Veränderung des Anteils Annexin V+/7-AAD+ mononukleärer Zellen (links Lympho-
zyten, rechts Monozyten) in Relation zum Ausgangswert.
Die Tendenz eines ersten Anstieges des Anteils der spät-apoptotischen bezie-
hungsweise nekrotischen Monozyten war zum Messzeitpunkt 48 Stunden zu verzeich-
nen. Signifikant wurde der Anstieg erst zwischen den Messungen 48 Stunden und 72
Stunden nach Beginn der Lagerung, wobei hier der Anteil spät-apoptotischer bezie-
Abb. 13: Änderung der Annexin V+/7-AAD– Monozyten (%)
im Verlauf der Lagerung über 72 Stunden.
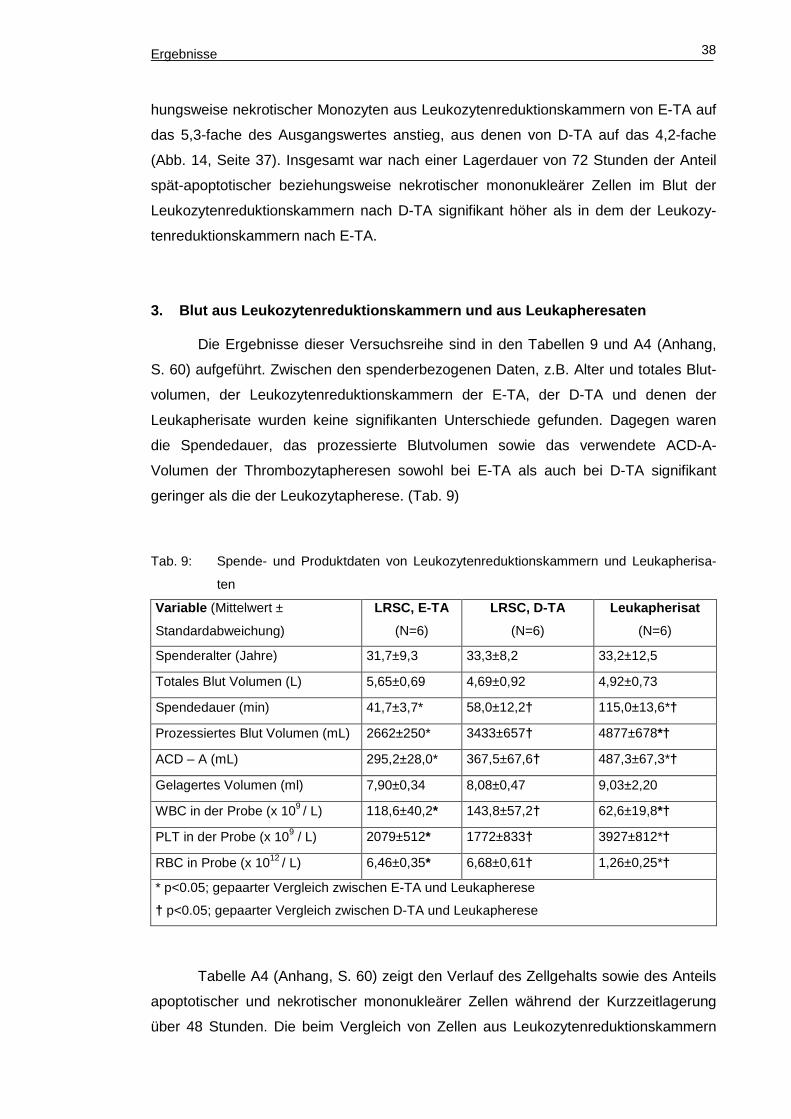
Ergebnisse 38
hungsweise nekrotischer Monozyten aus Leukozytenreduktionskammern von E-TA auf
das 5,3-fache des Ausgangswertes anstieg, aus denen von D-TA auf das 4,2-fache
(Abb. 14, Seite 37). Insgesamt war nach einer Lagerdauer von 72 Stunden der Anteil
spät-apoptotischer beziehungsweise nekrotischer mononukleärer Zellen im Blut der
Leukozytenreduktionskammern nach D-TA signifikant höher als in dem der Leukozy-
tenreduktionskammern nach E-TA.
3. Blut aus Leukozytenreduktionskammern und aus Le ukapheresaten
Die Ergebnisse dieser Versuchsreihe sind in den Tabellen 9 und A4 (Anhang,
S. 60) aufgeführt. Zwischen den spenderbezogenen Daten, z.B. Alter und totales Blut-
volumen, der Leukozytenreduktionskammern der E-TA, der D-TA und denen der
Leukapherisate wurden keine signifikanten Unterschiede gefunden. Dagegen waren
die Spendedauer, das prozessierte Blutvolumen sowie das verwendete ACD-A-
Volumen der Thrombozytapheresen sowohl bei E-TA als auch bei D-TA signifikant
geringer als die der Leukozytapherese. (Tab. 9)
Tab. 9: Spende- und Produktdaten von Leukozytenreduktionskammern und Leukapherisa-
ten
Variable (Mittelwert ±
Standardabweichung)
LRSC, E-TA
(N=6)
LRSC, D-TA
(N=6)
Leukapher isat
(N=6)
Spenderalter (Jahre) 31,7±9,3 33,3±8,2 33,2±12,5
Totales Blut Volumen (L) 5,65±0,69 4,69±0,92 4,92±0,73
Spendedauer (min) 41,7±3,7* 58,0±12,2† 115,0±13,6*†
Prozessiertes Blut Volumen (mL) 2662±250* 3433±657† 4877±678*†
ACD – A (mL) 295,2±28,0* 367,5±67,6† 487,3±67,3*†
Gelagertes Volumen (ml) 7,90±0,34 8,08±0,47 9,03±2,20
WBC in der Probe (x 109 / L) 118,6±40,2* 143,8±57,2† 62,6±19,8*†
PLT in der Probe (x 109 / L) 2079±512* 1772±833† 3927±812*†
RBC in Probe (x 1012 / L) 6,46±0,35* 6,68±0,61† 1,26±0,25*†
* p<0.05; gepaarter Vergleich zwischen E-TA und Leukapherese
† p<0.05; gepaarter Vergleich zwischen D-TA und Leukapherese
Tabelle A4 (Anhang, S. 60) zeigt den Verlauf des Zellgehalts sowie des Anteils
apoptotischer und nekrotischer mononukleärer Zellen während der Kurzzeitlagerung
über 48 Stunden. Die beim Vergleich von Zellen aus Leukozytenreduktionskammern
Ergebnisse 39
von E-TA mit denen von D-TA unter Ergebnisse, Punkt 2. beschriebenen Unterschiede
konnten tendenziell reproduziert werden, in dieser Stichprobe allerdings ohne den
Nachweis der vorbeschriebenen Signifikanzniveaus. Weiterhin wurden keine signifikan-
ten Unterschiede zwischen den Zellen der Leukozytenreduktionskammern der E-TA
und denen der D-TA gefunden.
Dagegen war in den Proben der Leukapheresate der sofort nach der Spende
gemessene Anteil von Annexin V+/7-AAD- Lymphozyten im Vergleich zu dem der Leu-
kozytenreduktionskammern doppelt so hoch (E-TA: p=0,005; D-TA: p=0,001). Der An-
teil an Annexin V+/7-AAD- Monozyten zeigte in Leukapherisaten verglichen mit den
Leukozytenreduktionskammern ebenfalls höhere Werte, ohne jedoch signifikante Un-
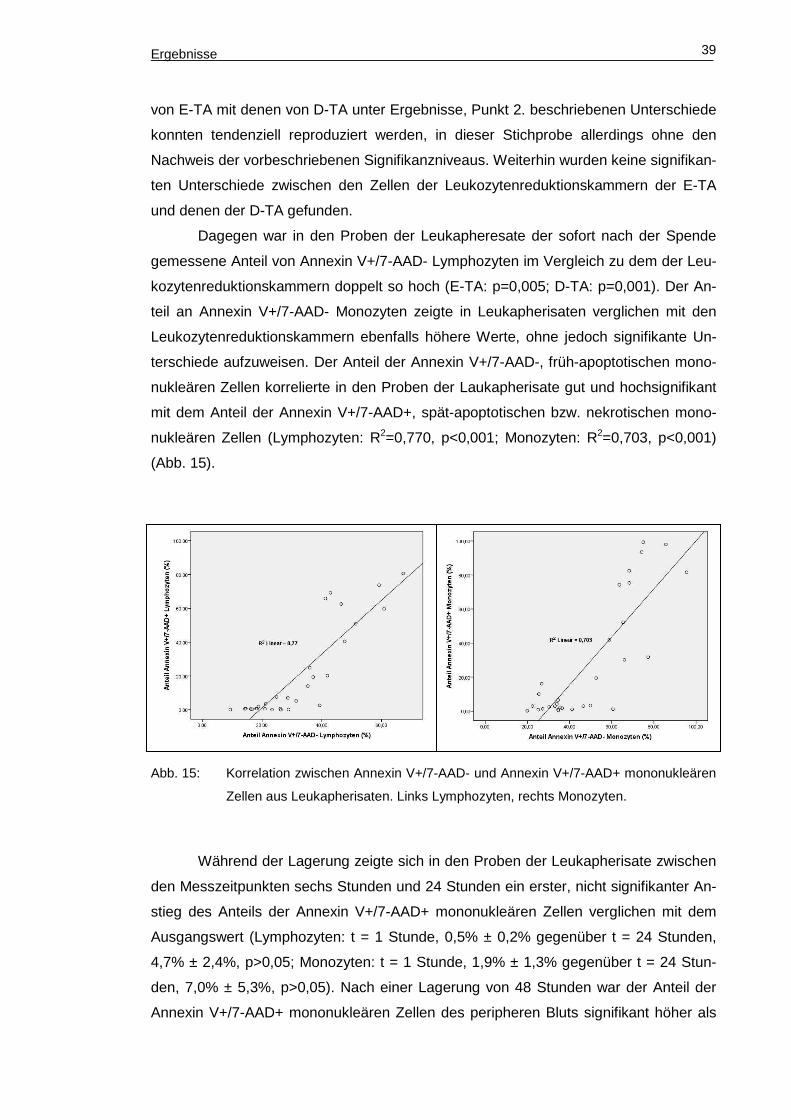
terschiede aufzuweisen. Der Anteil der Annexin V+/7-AAD-, früh-apoptotischen mono-
nukleären Zellen korrelierte in den Proben der Laukapherisate gut und hochsignifikant
mit dem Anteil der Annexin V+/7-AAD+, spät-apoptotischen bzw. nekrotischen mono-
nukleären Zellen (Lymphozyten: R2=0,770, p<0,001; Monozyten: R2=0,703, p<0,001)
(Abb. 15).
Abb. 15: Korrelation zwischen Annexin V+/7-AAD- und Annexin V+/7-AAD+ mononukleären
Zellen aus Leukapherisaten. Links Lymphozyten, rechts Monozyten.
Während der Lagerung zeigte sich in den Proben der Leukapherisate zwischen
den Messzeitpunkten sechs Stunden und 24 Stunden ein erster, nicht signifikanter An-
stieg des Anteils der Annexin V+/7-AAD+ mononukleären Zellen verglichen mit dem
Ausgangswert (Lymphozyten: t = 1 Stunde, 0,5% ± 0,2% gegenüber t = 24 Stunden,
4,7% ± 2,4%, p>0,05; Monozyten: t = 1 Stunde, 1,9% ± 1,3% gegenüber t = 24 Stun-
den, 7,0% ± 5,3%, p>0,05). Nach einer Lagerung von 48 Stunden war der Anteil der
Annexin V+/7-AAD+ mononukleären Zellen des peripheren Bluts signifikant höher als
Ergebnisse 40
der anfängliche Wert nach der Apherese (Lymphozyten: p<0,001; Monozyten:
p<0,001). Dagegen blieb der Anteil der Annexin V+/7-AAD+ mononukleären Zellen aus
dem Blut der Leukozytenreduktionskammern über die gesamte Lagerdauer auf einem
stabilen Niveau ohne signifikante Unterschiede (Tab. A4, Anhang S. 60). Nach einer
Lagerdauer von 48 Stunden waren sowohl der Anteil der Annexin V+/7-AAD– als auch
der Anteil der Annexin V+/7-AAD+ mononukleären Zellen aus Proben der Leukaphere-
sate signifikant höher als im gelagerten Blut der Leukozytenreduktionskammern (E-TA
und D-TA). Gleichzeitig fiel der Anteil und die Konzentration der CD 45+ Lymphozyten
und der CD14+ Monozyten im Blut der Leukapheresate signifikant ab Dagegen verän-
derten sich die Anteile der mononukleären Zellen in den Leukozytenreduktionskam-
mern während der gesamten Lagerdauer nicht.
Des Weiteren war zu beobachten, dass der pH-Wert sowohl im Blut der Leuko-
zytenreduktionskammern als auch im Blut der Leukapherisate innerhalb einer Lager-
dauer von 24 Stunden signifikant abfiel. Allerdings war bereits zu diesem Zeitpunkt der
pH-Wert in Proben der Leukapherisate signifikant tiefer als in denen der Leukozytenre-
duktionskammern (E-TA und D-TA). Im gelagerten Blut der Leukozytenreduktions-
kammern stieg das pCO2 binnen der Lagerung von 24 Stunden stetig an und fiel da-
nach bis zum Messzeitpunkt 48 Stunden signifikant ab. Der pO2-Wert verhielt sich ge-
gengleich. Im gelagerten Blut der Leukapherisate fiel dagegen der pCO2-Wert von Be-
ginn an stetig ab, bei einem gleichzeitigen Anstieg des pO2-Wertes auf mehr als das
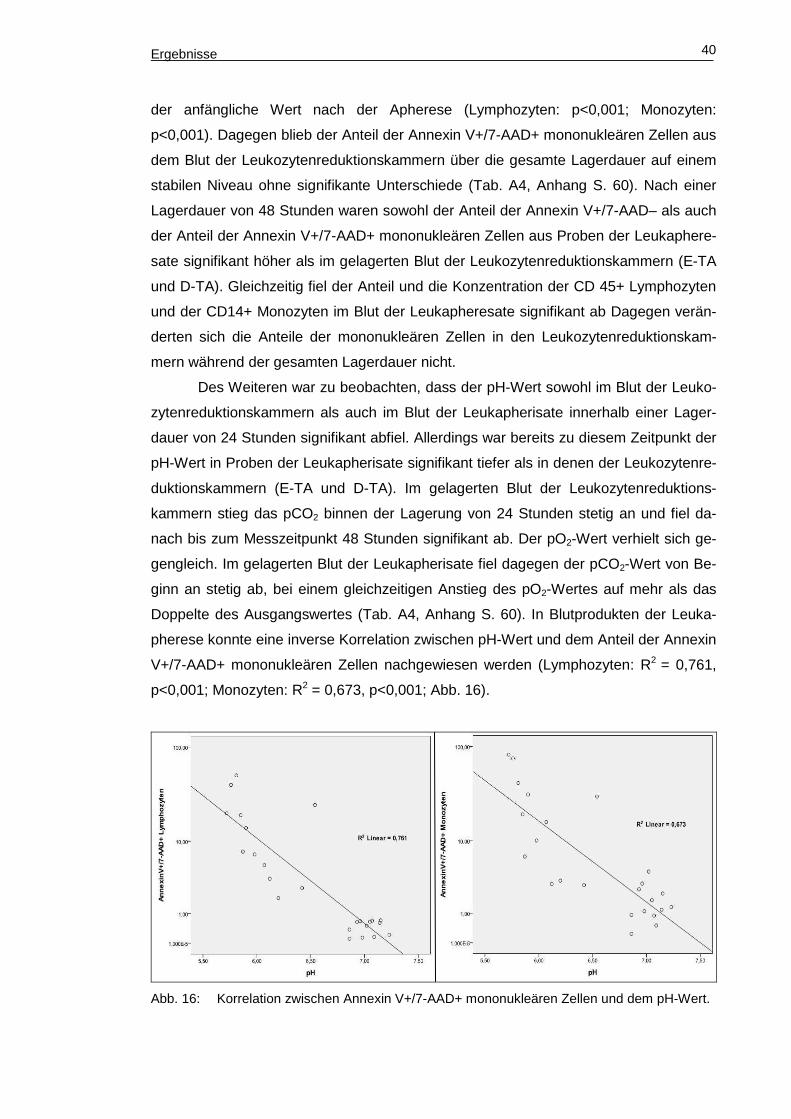
Doppelte des Ausgangswertes (Tab. A4, Anhang S. 60). In Blutprodukten der Leuka-
pherese konnte eine inverse Korrelation zwischen pH-Wert und dem Anteil der Annexin
V+/7-AAD+ mononukleären Zellen nachgewiesen werden (Lymphozyten: R2 = 0,761,
p<0,001; Monozyten: R2 = 0,673, p<0,001; Abb. 16).
Abb. 16: Korrelation zwischen Annexin V+/7-AAD+ mononukleären Zellen und dem pH-Wert.
Ergebnisse 41
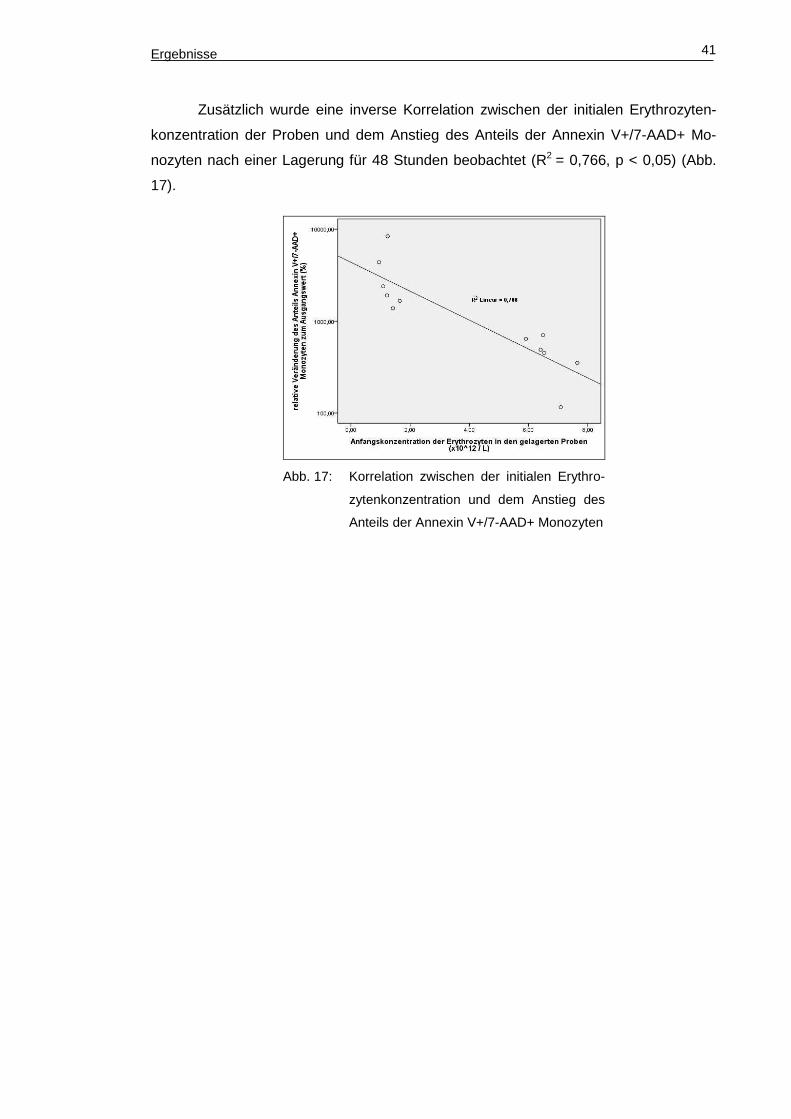
Zusätzlich wurde eine inverse Korrelation zwischen der initialen Erythrozyten-
konzentration der Proben und dem Anstieg des Anteils der Annexin V+/7-AAD+ Mo-
nozyten nach einer Lagerung für 48 Stunden beobachtet (R2 = 0,766, p < 0,05) (Abb.
17).
Abb. 17: Korrelation zwischen der initialen Erythro-
zytenkonzentration und dem Anstieg des
Anteils der Annexin V+/7-AAD+ Monozyten

Diskussion 42
VI. Diskussion
1. Vergleich von Leukozytenreduktionskammern zweie r Zellseparatoren
In einer Studie wurden die Leukozytenreduktionskammern der Trima Accel® und
der COBE® Spectra Zellseparatoren bezüglich ihres Zellgehalts verglichen. [59] Die am
häufigsten verwendete Quelle für Leukozyten des Menschen ist der Buffy Coat, der
Bestandteil des Blutes, den man nach Zentrifugation durch Abtrennen der Erythrozyten
erhält. Entsprechend Dietz et al. sind Leukozytenreduktionskammern eine weitere
Quelle, um Produkte mit einem hohen Leukozytengehalt und sehr guter Zellqualität zu
erhalten [15]. Allerdings konnten die Daten von Dietz et al. in dieser Arbeit nicht repro-
duziert werden. Die hier dargestellten Daten zeigen dagegen einen deutlich geringeren
Zellgehalt und eine deutlich stärkere Streuung innerhalb der Ergebnisse. Der Gehalt
mononukleärer Zellen in Leukozytenreduktionskammern des Trima Accel® Zellsepara-
tors war im Mittel 0,69 x109 Zellen, also um 66% geringer als die von Dietz et al. ge-
messenen Werte. Diese Unterschiede zur Studie von Dietz et al. könnten zum Beispiel
durch unterschiedliche Geräteeinstellungen der verwendeten Zellseparatoren hervor-
gerufen worden sein. Bei Dietz et al. waren das Antikoagulationsverhältnis 13:1 (hier
11:1), das „draw management“ war 3 (hier 6), das „return management“ 1 (hier 4) und
der maximale Fluss zum Gerät war auf „fast“ (schnell) eingestellt, in dieser Arbeit da-
gegen auf „medium“ (mittel). Auch unterschieden sich die Volumina des prozessierten
Blutes und die Spendedauer. Während Dietz et al. in 80 Minuten 4,25 l Blut prozessier-
ten, waren es hier nur 3,21 l in 54 Minuten. Dieser Unterschied ist wichtig, da die Leu-
kozytenreduktionskammern der TRIMA Accel® laut Herstellerangaben als voll gelten,
wenn ein Volumen von 4,7 l bei Spendern mit einem BMI < 30 kg/m2 beziehungsweise
von 4,1 l bei Spendern mit einem BMI > 30 kg/m2 prozessiert wurde. Durch die be-
schriebenen Unterschiede ist somit ein direkter Vergleich zwischen den hier dargestell-
ten Daten und denen von Dietz et al. nur bedingt möglich.
Durch die in dieser Arbeit gezeigten starken Schwankungen innerhalb der er-
zielten Zellkonzentrationen und dem erzielten Zellgehalt in den Leukozytenreduktions-
kammern muss die Eignung der Leukozytenreduktionskammern als Quelle für mono-
nukleäre Zellen zur Gewinnung dendritischer Zellen in Frage gestellt werden. Die Un-
beständigkeit des Zellgehalts kann zu einer Unbeständigkeit der Kultivierungsraten
führen. Mögliche Einflüsse auf die Beständigkeit des Zellgehalts in Leukozytenredukti-
onskammern könnten zum Beispiel im prozessierten Blutvolumen oder in unterschied-
lichen Patienteneigenschaften wie Alter oder BMI vermutet werden.

Diskussion 43
Zur Kultivierung von dendritischen Zellen aus mononukleären Zellen werden
nach dem derzeitigen Stand der Dinge mindestens 109 Monozyten benötigt [8]. In den
hier gezeigten Daten erzielten die Leukozytenreduktionskammern einen Monozytenge-
halt von maximal 31,6% des Mindestbedarfs, im Mittel sogar nur 16%. Damit reicht der
Zellgehalt der Leukozytenreduktionskammern der Trima Accel® aktuell nicht aus, um
aus ihnen eine komplette Impfserie mit dendritischen Zellen herstellen zu können. Al-
lerdings könnte mit verbesserten Kultivierungsraten dendritischer Zellender Mindest-
zellgehalt für die Herstellung von Impfstoffen aus dendritischen Zellen zukünftig sinken.
Bemerkenswert ist, dass der Gehalt an Leukozyten in Leukozytenreduktions-
kammern der TRIMA Accel® 30-fach höher war als in denen der COBE® Spetra. Ver-
mutlich kommt es während oder gegen Ende der Thrombozytapherese mit der COBE®
Spectra zu einem Auswaschen der Zellen aus den Leukozytenreduktionskammern.
Weitere mögliche Ursachen für den geringen Zellgehalt können allgemeine Unter-
schiede in den Spendeprinzipien wie zum Beispiel die Dauer der Apherese, die Fluss-
geschwindigkeit oder das prozessierte Blutvolumen sein. Unabhängig von der Ursache
für den geringen Zellgehalt ist zusammen zu fassen, dass die Leukozytenreduktions-
kammern der COBE® Spectra im Gegensatz zu denen der Trima Accel® keine suffizi-
ente Quelle für Leukozyten und mononukleäre Zellen des peripheren Blutes sind.
2. Zellgehalt und Zellschädigung in Leukozytenreduk tionskammern
In einer Folgestudie wurde verglichen, ob sich der Zellgehalt sowie die Apopto-
se- und Nekroserate während der Kurzzeitlagerung über 72 Stunden in Leukozytenre-
duktionskammern der Trima Accel® nach einfacher und nach doppelter Thrombozyta-
pherese unterscheiden. [58] Es konnte ein hochsignifikanter Unterschied zwischen den
Leukozytenkonzentrationen der Leukozytenreduktionskammern nach einfachen und
doppelten Thrombozytapheresen mit der Trima Accel® gefunden werden (Tab. 8, S.
35). Der Leukozytengehalt in den Leukozytenreduktionskammern war bei einem Füll-
volumen von 10 ml nach E-TA durchschnittlich 0,93 x109 ± 0,32 x109 Zellen und nach
D-TA durchschnittlich 1,71 x109 ± 0,55 x109 Zellen (p < 0,01). Die letzteren Werte ent-
sprechen denen, die Neron et al. in ihrer 2007 veröffentlichen Arbeit über den in Leu-
kozytenreduktionskammern vorliegenden Zellgehalt beschrieben haben [41]. Die signi-
fikanten Unterschiede zwischen den Leukozytenkonzentrationen und dem Leukozyten-
gehalt von E-TA und D-TA können durch das signifikant unterscheidliche PBV beider
Spendearten erklärt werden, da die Leukozytenkonzentration hochsignifikant mit dem
PBV korreliert (r2 = 0,75, p < 0,001) (Abb. 12, S. 35). Die unter Diskussion, Punkt 1.

Diskussion 44
beschriebenen Unterschiede zwischen den Ergebnissen dieser Arbeit und denen von
Dietz können durch Unterschiede im prozessierten Blutvolumen erklärt werden [59].
Auch in dieser Versuchsserie konnten die veröffentlichten Daten nicht reprodu-
ziert werden. Allerdings hatten im Gegensatz zu der Studie von Dietz et al. [15] die
Spender dieser Serie einen BMI von maximal 30 kg/m2. Damit konnte das Volumen, ab
welchem die Leukozytenreduktionskammern als voll angesehen werden, nicht prozes-
siert werden [10]. Das heißt, dass der Vergleich mit den Ergebnissen von Dietz et al.
auch bei diesen Daten nur eingeschränkt möglich ist. Es konnte keine Korrelation zwi-
schen der Leukozytenkonzentration und dem Anteil CD14+ Monozyten im Blut der
Leukozytenreduktionskammern nachgewiesen werden. Die Konzentration der Monozy-
ten war in Leukozytenreduktionskammern der D-TA signifikant höher als in denen der
E-TA. Dagegen wurden keine signifikanten Unterschiede bezüglich der Konzentration
der Lymphozyten gefunden.
Entsprechend der aktuellen Literatur konnten die aus Leukozytenreduktions-
kammern gewonnenen mononukleären Zellen erfolgreich zur Kultivierung dendritischer
Zellen verwendet werden [15, 35, 41]. Weitgehend unbekannt war bisher, ob die aus
Leukozytenreduktionskammern gewonnenen Zellen Zeichen von Apoptose oder Nek-
rose aufzeigen und wenn nicht, wie lange sie gelagert werden können, ohne dass eine
Schädigung auftritt. Dies ist von besonderem Interesse, da eine Zellschädigung zu
Zellverlust führen und so die Kultivierungsraten dendritischer Zellen beeinflussen kann.
Neron und Kollegen berichteten, dass 95% der aus Leukozytenreduktionskammern
gewonnenen Leukozyten vital sind [41]. Dies deckt sich mit den in Tabelle A3 (Anhang,
S. 59) aufgeführten Daten, die einen initialen Anteil von 7-AAD+ mononukleären Zellen
in Höhe von 2% bis 5% zeigen. Zusätzlich werden in dieser Arbeit Daten bezüglich der
Vitalität mononukleärer Zellen aufgeführt, zum einen im Verlauf der Kurzzeitlagerung,
zum anderen im Vergleich von Zellen aus Leukozytenreduktionskammern mit einer
niedrigen Leukozytenkonzentration (E-TA) und denen mit einer hohen Leukozytenkon-
zentration (D-TA). Der Anteil der Annexin V+/7-AAD+ Lymphozyten war in den Leuko-
zytenreduktionskammern der D-TA signifikant höher als in denen der E-TA. Man könn-
te daraus schließen, dass eine hohe Leukozytenkonzentration zu einem höheren Anteil
spät-apoptotischen bzw. nekrotischen Zellen führt. Dem widerspricht allerdings, dass
keine signifikanten Unterschiede zwischen den Anteilen Annexin V+/7-AAD+ Monozy-
ten in den Leukozytenreduktionskammern zu finden waren (Tab. A3, Anhang, S. 59).
Zwischen Lymphozyten und Monozyten gab es signifikante Unterschiede be-
züglich der Anteile der Annexin V+/7-AAD– Zellen. Der Anteil von Annexin V+/7-AAD–
Lymphozyten schwankte zwischen 20% und 27% und war während der gesamten La-
gerdauer stabil. Unterschiede zwischen dem Anteil der Annexin V+/7-AAD– Lymphozy-

Diskussion 45
ten aus Leukozytenreduktionskammern der E-TA mit denen der D-TA wurden nicht
beobachtet. Dagegen war der Anteil der Annexin V+/7-AAD– Monozyten intial mit Wer-
ten zwischen 39% und 48% erhöht und fiel nach einer Lagerung von 24 Stunden so-
wohl in den Leukozytenreduktionskammern der E-TA als auch in denen der D-TA auf
durchschnittlich 30%. Danach kam es zu einem signifikanten Anstieg des Anteils der
Annexin V+/7-AAD– Monozyten, der von einem Anstieg des Anteils der Annexin V+/7-
AAD+ Monozyten begleitet wurde. Sowohl bei Lymphozyten wie auch bei Monozyten
nahm der Anteil der Annexin V+/7-AAD+ Zellen zu, was auf eine irreversible Zellschä-
digung hinweist. Der erste signifikante Anstieg des Anteils der Annexin V+/7-AAD+
Zellen wurde bei Lymphozyten wie bei Monozyten nach einer Lagerdauer von 24 Stun-
den beobachtet. Diese Daten decken sich mit den Beobachtungen von Strasser et al.,
die in einer Studie über die Kurzzeitlagerung von Leukapherisaten in PVC-Beuteln be-
schrieben, dass es zu einem ersten signifikanten Abfall der Konzentration mononukleä-
rer Zellen nach einer Lagerdauer von mehr als 24 Stunden kommt [57].
Obwohl Monozyten einen hohen Anteil an Annexin V+ Zellen binnen der ersten
24 Stunden der Lagerung zeigten war der Anteil der 7-AAD+ Monozyten als Zeichen
der irreversiblen Zellschädigung in diesem Zeitraum gering. Möglicherweise wurden die
hohen Messwerte bezüglich des Anteils der Annexin V+/7-AAD– Monozyten durch un-
spezifische Bindungen von Annexin V an Monozyten hervor gerufen. Es ist bekannt,
dass Annexin V unspezifisch an Zellen binden kann [7, 12]. Allerdings ist dies als allei-
nige Ursache vor allem im Hinblick auf den Abfall des Anteils der Annexin V+ Monozy-
ten binnen der ersten 24 Stunden nach der Apherese unwahrscheinlich. Amer et al.
fanden in einer 2010 veröffentlichten Studie heraus, dass Lymphozyten wie Monozyten
während einer Apherese oxidativem Stress ausgesetzt sind. Dies führt zu einer Zell-
schädigung mit Externalisation von Phosphatidylserin und damit zur Annexin V-
Positivität nach Färbung [1]. Geske et al. zeigten, dass eine durch p53-induzierte
Apoptose in frühen Stadien reversibel ist, wenn die apoptotischen Stimuli entfernt wer-
den. Dabei kam es zum Rückgang der Phosphatidylserin-Externalisation und somit
auch zum Abfall des Anteils der Annexin V+ Zellen [21]. Vor dem Hintergrund der Be-
obachtungen von Amer und Geske könnte die Ursache für den initial nach der Aphere-
se erhöhten Anteil der Annexin V-positiven Monozyten durch die Apherese hervorgeru-
fener Zellstress sein. Nach Ende der Apherese und des Zellstresses wird der apoptoti-
sche Stimulus entfernt, die Zellen regenerieren sich und es kommt nicht obligatorisch
zum Zelltod der Monozyten. Die Regeneration der Monozyten scheint entsprechend
der hier dargestellten Daten binnen der ersten 24 Stunden nach Apherese statt zu fin-
den. Der im Vergleich zu Monozyten niedrige Anteil von Annexin-V+/7-AAD– Lympho-
zyten initial nach der Apherese scheint auf eine höhere Resistenz der Lymphozyten

Diskussion 46
gegenüber äußeren Einflüssen wie zum Beispiel Zellstress während der Apherese hin
zu weisen.
3. Blut aus Leukozytenreduktionskammern und aus Le ukapheresaten
In einer weiteren Folgestudie wurden Leukozytenreduktionskammern mit
Leukapheresaten bezüglich Zellgehalt und Zellapoptose/-nekrose während der Kurz-
zeitlagerung über 48 Stunden verglichen. [68] Die hier dargestellten Daten zeigen hin-
sichtlich der Zellviabilität eine deutliche Diskrepanz zwischen der Qualität der Leuka-
pheresate, dem Goldstandard für die Gewinnung mononukleärer Zellen, und der Quali-
tät der Leukozytenreduktionskammern. Bereits bei den Messungen sofort nach der
Apherese zeigte sich verglichen mit den Leukozytenreduktionskammern ein erhöhter
Anteil Annexin V+/7-AAD- mononukleärer Zellen in den Leukapherisaten. Die Färbung
mit Annexin V ist bekanntlich nicht unkritisch zu bewerten, da dieses nicht nur an
apoptotische Zellen bindet, sondern auch unspezifisch binden kann. Dagegen ist eine
mit 7-AAD gefärbte Zelle sicher irreversibel geschädigt. In den hier dargestellten Daten
korrelierte der Anteil der Annexin V+/7-AAD– mononukleären Zellen mit dem Anteil der
Annexin V+/7-AAD+ mononukleären Zellen hochsignifikant (Lymphozyten: R2=0,770,
p<0,001; Monozyten: R2=0,703, p<0,001) (Abb. 15). Es ist daher davon auszugehen,
dass die Bindung von Annexin V an mononukleäre Zellen durch eine Zellschädigung
hervorgerufen wurde.
Üblicherweise werden zur Kultivierung dendritischer Zellen aus Monozyten die
mononukleären Zellen der Leukapherisate binnen sechs bis 24 Stunden nach der A-
pherese in Zellkulturen angelegt. In dieser Zeitspanne war aber bereits ein erster ten-
denzieller Anstieg der Annexin V+/7-AAD+ Events in den Proben der Leukapherisate
zu erkennen, auch wenn noch keine signifikanten Unterschiede bestanden. Gleichzei-
tig fiel in den gelagerten Proben der Leukapherisate der pH-Wert verglichen mit den
Leukozytenreduktionskammern deutlich stärker ab. Im gelagerten Blut der Leukozyten-
reduktionskammern stieg der pCO2-Wert innerhalb der ersten 24 Stunden an, gleich-
zeitig kam es zum Abfall des pO2-Werts. Da die zur Lagerung verwendeten PVC-
Beutel eine feste Gasaustauschrate für O2 und für CO2 haben, kann diese Beobach-
tung als metabolischer Sauerstoffverbrauch unter CO2-Produktion gewertet werten. In
den Blutproben der Leukapherisate fiel dagegen der pCO2-Wert von Beginn der Lage-
rung stetig ab während der pO2-Wert stetig anstieg. Verglichen mit den Proben der
Leukozytenreduktionskammern sind in den Proben der Leukapherisate ein reduzierter
Sauerstoffverbrauch und eine gleichzeitig reduzierte CO2-Produktion, also eine anae-
robe Stoffwechselsituation, zu vermuten. Picker et al. beschrieben in einer Studie zur

Diskussion 47
Thrombozytenlagerung, dass der Sauerstoffverbrauch von Zellen invers mit der Zell-
schädigung korreliert [44]. Gleichzeitig ist aus der Literatur bekannt, dass während der
Lagerung mononukleärer Zellen der pH-Wert umso stärker abfällt, je mehr Laktat pro-
duziert wird [57]. Die Laktatkonzentration konnte in dieser Arbeit auf Grund des gerin-
gen Lagervolumens und des vergleichsweise großen Probenbedarfs zur Laktatbe-
stimmung nicht gemessen werden. Wie unter Ergebnisse, Punkt 3. beschrieben, korre-
liert im Blut der Leukapheresate der pH-Wert-Abfall hochsignifikant mit dem Anstieg
des Anteils der Annexin V+/7-AAD+ mononukleären Zellen (Abb. 16, Seite 40). Aus der
Zusammenschau all dieser Beobachtungen kann die Hypothese formuliert werden,
dass im Blut der Leukapherisate die mononukleären Zellen dermaßen geschädigt sein
könnten, dass der aerobe Stoffwechsel und dadurch der Sauerstoffverbrauch gestört
sind.
Die Diskrepanz zwischen der Viabilität mononukleärer Zellen aus Leukapherisa-
ten und derer aus Leukozytenreduktionskammern kann unterschiedliche Ursachen
haben. Zum einen unterscheidet sich die Art der Zellanreicherung während der Leuko-
zytapherese und in den Leukozytenreduktionskammern. Während der Leukozytaphe-
rese werden Leukozyten durch die Zentrifugation dicht zusammengepresst. Die auf sie
dabei wirkende Kraft entspricht bei den verwendeten Geräteeinstellungen und einer
Drehzahl von 1500 Umdrehungen/Minute 297 x g, also fast dem 300-fachen der Erd-
anziehungskraft. Dagegen kommt es in den Leukozytenreduktionskammern zu einer
Anreicherung von Leukozyten, indem die schweren Zellen, Leukozyten und Erythrozy-
ten, auf Grund der Bauweise der Kammern zur Kammerwand fließen. Dagegen können
die leichten Zellen, Thrombozyten, die Kammer passieren. Ursächlich dafür sind kön-
nen Strömungsturbulenzen an den Stufen des sich in Flussrichtung terrassenförmig
erweiternden Kegels sein. Die dabei auf die Zellen wirkende Kraft ist nicht bekannt.
Bekannt ist aber, dass sich dabei eine lockere Ansammlung von Zellen bildet, so dass
wahrscheinlich geringere Kräfte auf die Zellen in den Kegeln wirken als während der
Leukozytapherese [27, 41]. Des Weiteren haben Amer et al. wie bereits beschrieben
beobachtet, dass Apheresen oxidativen Stress verursachen und so zu einer erhöhten
Annexin V-Bindungsrate an Zellen führen [1]. Vor der Aussage der Studie von Amer ist
zu vermuten, dass neben der sanften Anreicherung der Leukozyten in den Leukozyten-
reduktionskammern weitere Faktoren der Apherese, wie zum Beispiel Unterschiede im
prozessierten Blutvolumen oder in der Spendedauer, Einfluss auf die Qualität der ge-
wonnen mononukleären Zellen haben können. Die Hypothese, dass diese Unterschie-
de in der Methode mononukleäre Zellen anzureichern unterschiedlich stark ausgepräg-
te Zellschädigungen hervorrufen, ist daher naheliegend.

Diskussion 48
Zum anderen unterscheiden sich die Proben der Leukozytenreduktionskam-
mern und der Leukapherisate nicht nur bezüglich ihrer Herstellung sondern auch be-
züglich ihres Restzellgehalts, wobei dieser die Qualität des Blutes weniger direkt nach
der Apherese beeinflusst haben könnte als mehr im Verlauf der Kurzzeitlagerung. In
Proben der Leukapherisate war die Thrombozytenkonzentration verglichen mit denen
der Leukozytenreduktionskammern doppelt so hoch (E-TA, p=0,002; D-TA, p<0,001).
Gleichzeitig war in den Proben der Leukapherisate die Erythrozytenkonzentration um
ein Vielfaches signifikant geringer als in den Proben der Leukozytenreduktionskam-
mern (Tab. 9, S. 38). Thrombozyten werden bekanntermaßen während einer Apherese
aktiviert und steigern dabei ihre Stoffwechselaktivität [5, 28]. Daher wäre es möglich,
dass in den Leukapherisaten durch die erhöhte Zahl der Thrombozyten die für den
Stoffwechsel der Leukozyten nötigen Substrate durch die Thrombozyten verbraucht
wurden, die gelagerten Zellen ihren Stoffwechsel nicht mehr aufrecht erhalten konnten
und es dadurch zu einer Zellschädigung im gelagerten Produkt kam. Es ist bekannt,
dass in gelagerten Thrombozytenkonzentraten der pH-Wert hypoxiebedingt signifikant
abfallen kann [32]. Allerdings kam es im Verlauf der Lagerung zu einem Anstieg des
Sauerstoffpartialdrucks in den Proben der Leukapherisate, so dass eine Hypoxie und
dadurch ein Einfluss der Thrombozyten auf den pH-Wert unwahrscheinlich sind. Dass
keine signifikanten Korrelationen zwischen der Thrombozytenkonzentration und den
Parametern der Zellschädigung beobachtet werden konnte, ist möglicherweise auf die
geringe Fallzahl dieser Studie zurück zu führen.
Amer et al. beschrieben, dass Apheresen zu Zellschädigung durch oxidativem
Stress und so zur Bindung von Annexin V führt [1]. Gleichzeitig wurde von der Arbeits-
gruppe beschrieben, dass durch Glutathion als Antioxidans die bei der Apherese ent-
stehenden Oxidantien reduziert werden. Der oxydative Stress war also umso geringer
je mehr Glutathion zur verfügung stand. Gluthation kommt im Blut vorrangig in Erythro-
zyten vor. In den gelagerten Proben der Leukozytenreduktionskammern war die Eryth-
rozytenkonzentration um ein Vielfaches höher als in denen der Leukapherisate. Außer-
dem konnte beobachtet werden, dass je geringer die initiale Erythrozytenkonzentration
war, umso stärker stieg der Anteil der Annexin V+/7-AAD+ Monozyten im Blut der
Leukapherisate an (Abb. 17, S. 41) Möglich wäre daher, dass die erhöhte Erythrozy-
tenkonzentration in den Leukozytenreduktionskammern einen protektiven Effekt ge-
genüber dem oxidativen Stress während der Apherese haben könnte. Auch könnte auf
Grund der physiologischen Pufferkapazität des Hämoglobins die erhöhte Erythrozyten-
konzentration der Leukozytenreduktionskammern Einfluss auf den, verglichen mit den
Leukapherisaten, geringeren Abfall des pH-Werts haben. Ob Erythrozyten auf die Qua-

Diskussion 49
lität gelagerter mononukleärer Zellen einen Einfluss haben, lässt sich mit dieser Arbeit
aber nicht klären.
Wahrscheinlich ist die Diskrepanz zwischen der Viabilität mononukleärer Zellen
aus Leukapherisaten und derer aus Leukozytenreduktionskammern mit einer Kombina-
tion der hier diskutierten Einflussgrößen zu erklären. Auf Grund des hohen Monozyten-
bedarfs zur Herstellung dendritischer Zellen für die Immuntherapie reichen Leukozyten-
reduktionskammern aktuell noch nicht als Quelle für mononukleäre Zellen aus. Aller-
dings scheint allein das Prinzip der Zellanreicherung in Leukozytenreduktionskammern
als neue und sanfte Methode vielversprechend für die Gewinnung mononukleärer Zel-
len zu sein.
Zusammenfassend sollten die hier bearbeiteten Themen Gegenstand weiterer
Forschung sein um die Qualität der Leukozytenprodukte als auch der darauf folgenden
Kultivierung von dendritischen Zellen zu verbessern. Interessant ist, ob der hier beo-
bachtete Abfall der Annexin V-Bindung an Monozyten aus Leukozytenreduktionskam-
mern innerhalb der ersten 24 Stunden nach Spende Einfluss auf die Kultivierungsrate
dendritischer Zellen hat. Dies hätte die Empfehlung zur Konsequenz, Blutprodukte zwi-
schenzulagern bevor sie für Zellkulturen verwendet werde. Auch sollte der Hypothese
nachgegangen werden, dass die Unterschiede der Zellanreicherung ursächlich für die
Qualitätsunterschiede zwischen Blut aus Leukapheresaten und Leukozytenreduktions-
kammern sind. Offen ist, ob die hier beobachteten Qualitätsunterschiede auch Einfluss
auf die Kultivierungsrate von dendritischen Zellen haben. Es gibt mittlerweile Zellsepa-
ratoren, wie z.B. die Spectra Optia® von CaridianBCT, die eine schonendere Zellanrei-
cherung verglichen mit den klassischen Zellseparatoren bei gleichem Zellgehalt im
Produkt versprechen. Diese neuen Zellseparatoren könnten nützlich für die Gewinnung
dendritischer Zellen sein. Schließlich ist fraglich, inwieweit oxydativer Stress während
der Apherese auf die Kultivierung dendritischer Zellen Einfluss hat. Sollte die hier ge-
machte Beobachtung eines scheinbar protektiven Effekts für Leukozyten durch einen
hohen Erythrozytengehalt in Blutprodukten verifizierbar sein, so müsste zukünftig bei
Leukapheresen ein höherer Zielzellgehalt von Erythrozyten angestrebt werden.

Literaturverzeichnis 50
VII. Literaturverzeichnis
1. Amer, J., Frankenburg, S., und Fibach, E., Apheresis induces oxidative stress
in blood cells. Ther Apher Dial, 2010. 14(2): p. 166-71.
2. Andree, H.A., Reutelingsperger, C.P., Hauptmann, R., Hemker, H.C., Hermens,
W.T., und Willems, G.M., Binding of vascular anticoagulant alpha (VAC alpha)
to planar phospholipid bilayers. The Journal of biological chemistry, 1990.
265(9): p. 4923-8.
3. Anonym, Cancer vaccine approval could open floodgates. Nat Med, 2010.
16(6): p. 615.
4. Anonym, An empiric victory. Nat Biotechnol, 2010. 28(6): p. 529.
5. Bakry, R., Sayed, D., Galal, H., und Shaker, S., Platelet function, activation and
apoptosis during and after apheresis. Ther Apher Dial, 2010. 14(5): p. 457-64.
6. Banchereau, J., Briere, F., Caux, C., Davoust, J., Lebecque, S., Liu, Y.J.,
Pulendran, B., und Palucka, K., Immunobiology of dendritic cells. Annu Rev
Immunol, 2000. 18: p. 767-811.
7. Bedner, E., Li, X., Gorczyca, W., Melamed, M.R., und Darzynkiewicz, Z.,
Analysis of apoptosis by laser scanning cytometry. Cytometry, 1999. 35(3): p.
181-95.
8. Berger, T.G., Strasser, E., Smith, R., Carste, C., Schuler-Thurner, B.,
Kaempgen, E., und Schuler, G., Efficient elutriation of monocytes within a
closed system (Elutra) for clinical-scale generation of dendritic cells. Journal of
immunological methods, 2005. 298(1-2): p. 61-72.
9. Bruckheimer, E.M. und Schroit, A.J., Membrane phospholipid asymmetry: host
response to the externalization of phosphatidylserine. J Leukoc Biol, 1996.
59(6): p. 784-8.
10. Bühl-Göpfert, M. und Eibel, P., Analysetechniken in der Immunbiologie, 2011.

Literaturverzeichnis 51
11. Cooper, G.M. und Hausman, R.E., The cell : a molecular approach. 5th ed2009,
Washington, D.C., Sunderland, Mass.: ASM Press, Sinauer Associates. xix,
820 p.
12. Darzynkiewicz, Z., Bedner, E., und Smolewski, P., Flow cytometry in analysis of
cell cycle and apoptosis. Semin Hematol, 2001. 38(2): p. 179-93.
13. Darzynkiewicz, Z., Juan, G., Li, X., Gorczyca, W., Murakami, T., und Traganos,
F., Cytometry in cell necrobiology: analysis of apoptosis and accidental cell
death (necrosis). Cytometry, 1997. 27(1): p. 1-20.
14. DeFrancesco, L., Landmark approval for Dendreon's cancer vaccine. Nat
Biotechnol, 2010. 28(6): p. 531-2.
15. Dietz, A.B., Bulur, P.A., Emery, R.L., Winters, J.L., Epps, D.E., Zubair, A.C.,
und Vuk-Pavlovic, S., A novel source of viable peripheral blood mononuclear
cells from leukoreduction system chambers. Transfusion, 2006. 46(12): p.
2083-9.
16. Disis, M.L., Bernhard, H., und Jaffee, E.M., Use of tumour-responsive T cells as
cancer treatment. Lancet, 2009. 373(9664): p. 673-83.
17. FDA, Malarkey, M.A., und Witten, C.M., Approval Letter - Provenge. 2010.
18. Finkel, E., The mitochondrion: is it central to apoptosis? Science, 2001.
292(5517): p. 624-6.
19. Fromm, A., Durchflusszytometrie thrombozytärer Oberflächenantigene
[Elektronische Ressource] : Änderungen im Langzeitverlauf in einer Population
gesunder Erwachsener / vorgelegt von Annette Fromm2006.
20. Gerke, V. und Moss, S.E., Annexins: from structure to function. Physiological
reviews, 2002. 82(2): p. 331-71.
21. Geske, F.J., Lieberman, R., Strange, R., und Gerschenson, L.E., Early stages
of p53-induced apoptosis are reversible. Cell Death Differ, 2001. 8(2): p. 182-
91.
22. Goldman, B. und DeFrancesco, L., The cancer vaccine roller coaster. Nat
Biotechnol, 2009. 27(2): p. 129-39.

Literaturverzeichnis 52
23. Hajek, J. und Sidák, Z., Theory of rank tests1967, New York, NY <etc.>:
Academic Press. 297 S.
24. Hasper, H.J., Weghorst, R.M., Richel, D.J., Meerwaldt, J.H., Olthuis, F.M., und
Schenkeveld, C.E., A new four-color flow cytometric assay to detect apoptosis
in lymphocyte subsets of cultured peripheral blood cells. Cytometry, 2000.
40(2): p. 167-71.
25. Hedrick, S.M., Ch'en, I.L., und Alves, B.N., Intertwined pathways of
programmed cell death in immunity. Immunol Rev, 2010. 236: p. 41-53.
26. Heim, M.U., [Guidelines of the German Medical Association for Therapy with
Blood Components and Plasma Derivatives--an introduction. Evidence-based
recommendations for the risk-benefit analysis in hemotherapy]. Anasthesiol
Intensivmed Notfallmed Schmerzther, 2009. 44(3): p. 186-97; quiz 199.
27. Hillyer, C.D., Blood banking and transfusion medicine : basic principles &
practice. 2nd ed2007, Philadelphia, PA: Churchill Livingstone/Elsevier. xxi, 887
p.
28. Holmsen, H., Platelet metabolism and activation. Semin Hematol, 1985. 22(3):
p. 219-40.
29. Ibrahim, S.F. und van den Engh, G., Flow cytometry and cell sorting. Adv
Biochem Eng Biotechnol, 2007. 106: p. 19-39.
30. Kerr, J.F., Neglected opportunities in apoptosis research. Trends Cell Biol,
1995. 5(2): p. 55-7.
31. Kerr, J.F., Wyllie, A.H., und Currie, A.R., Apoptosis: a basic biological
phenomenon with wide-ranging implications in tissue kinetics. Br J Cancer,
1972. 26(4): p. 239-57.
32. Kilkson, H., Holme, S., und Murphy, S., Platelet metabolism during storage of
platelet concentrates at 22 degrees C. Blood, 1984. 64(2): p. 406-14.
33. Kolmogorov, A.N., Grundbegriffe der Wahrscheinlichkeitsrechnung. Ergebnisse
der Mathematik und ihrer Grenzgebiete1933, Berlin: Springer. 62 S.
34. Kumar, V. und Robbins, S.L., Robbins basic pathology. 8th ed2007,
Philadelphia, PA: Saunders/Elsevier. xiv, 946 p.

Literaturverzeichnis 53
35. Lee, Y., Kim, S., Lee, S.T., Kim, H.S., Baek, E.J., Kim, H.J., Lee, M., und Kim,
H.O., [Analysis of characteristics of mononuclear cells remaining in the
leukoreduction system chamber of Trima Accel and their differentiation into
dendritic cells]. Korean J Lab Med, 2009. 29(4): p. 353-60.
36. Lodish, H.F., Molecular cell biology. 6th ed2008, New York: W.H. Freeman. 1 v.
(various pagings).
37. Martin, S.J., Reutelingsperger, C.P., McGahon, A.J., Rader, J.A., van Schie,
R.C., LaFace, D.M., und Green, D.R., Early redistribution of plasma membrane
phosphatidylserine is a general feature of apoptosis regardless of the initiating
stimulus: inhibition by overexpression of Bcl-2 and Abl. J Exp Med, 1995.
182(5): p. 1545-56.
38. Murphy, K.P., Travers, P., Walport, M., und Janeway, C., Janeway's
immunobiology. 7th ed2008, New York: Garland Science. xxi, 887 p.
39. Nadler, S.B., Hidalgo, J.H., und Bloch, T., Prediction of blood volume in normal
human adults. Surgery, 1962. 51(2): p. 224-32.
40. Nave, C.R. The Cell Membrane. HyperPhysics 2011; Available from:
http://hyperphysics.phy-astr.gsu.edu/hbase/biology/celmem.html.
41. Neron, S., Thibault, L., Dussault, N., Cote, G., Ducas, E., Pineault, N., und Roy,
A., Characterization of mononuclear cells remaining in the leukoreduction
system chambers of apheresis instruments after routine platelet collection: a
new source of viable human blood cells. Transfusion, 2007. 47(6): p. 1042-9.
42. Patel, T., Gores, G.J., und Kaufmann, S.H., The role of proteases during
apoptosis. FASEB J, 1996. 10(5): p. 587-97.
43. Pearson, K., Karl Pearson's early statistical papers1948, Cambridge: At the
University Press. 557 S.
44. Picker, S.M., Schneider, V., Oustianskaia, L., und Gathof, B.S., Cell viability
during platelet storage in correlation to cellular metabolism after different
pathogen reduction technologies. Transfusion, 2009. 49(11): p. 2311-8.
45. Planck, M., Zur Theorie des Gesetzes der Energieverteilung im
Normalspectrum (vorgetragen in der Sitzung vom 14. December 1900.).
Verhandlungen der Deutschen Physikalischen Gesellschaft1900.

Literaturverzeichnis 54
46. Randolph, G.J., Jakubzick, C., und Qu, C., Antigen presentation by monocytes
and monocyte-derived cells. Curr Opin Immunol, 2008. 20(1): p. 52-60.
47. Resnick, R., Physics Robert Resnick, David Halliday, ed. D. Halliday. Vol. 3. ed.
1977, New York [usw.]: Wiley.
48. Sallusto, F. und Lanzavecchia, A., Monocytes join the dendritic cell family. Cell,
2010. 143(3): p. 339-40.
49. Schlegel, R.A. und Williamson, P., Phosphatidylserine, a death knell. Cell Death
Differ, 2001. 8(6): p. 551-63.
50. Schuler, G., Dendritic cells in cancer immunotherapy. Eur J Immunol, 2010.
40(8): p. 2123-30.
51. Schwartzman, R.A. und Cidlowski, J.A., Apoptosis: the biochemistry and
molecular biology of programmed cell death. Endocr Rev, 1993. 14(2): p. 133-
51.
52. Seibt, W., Physik für Mediziner / Walter Seibt2009, Stuttgart ; New York, NY :
Thieme.
53. Seisenberger, W.S., Veränderungen im Immunsystem durch Leukapherese, ed.
W.S. Seisenberger2003.
54. Singer, S.J. und Nicolson, G.L., The fluid mosaic model of the structure of cell
membranes. Science, 1972. 175(23): p. 720-31.
55. Steinman, R.M., Dendritic cells: understanding immunogenicity. Eur J Immunol,
2007. 37 Suppl 1: p. S53-60.
56. Steinman, R.M., Some interfaces of dendritic cell biology. APMIS, 2003. 111(7-
8): p. 675-97.
57. Strasser, E.F., Keller, B., Hendelmeier, M., Ringwald, J., Zingsem, J., und
Eckstein, R., Short-term liquid storage of CD14+ monocytes, CD11c+, and
CD123+ precursor dendritic cells produced by leukocytapheresis. Transfusion,
2007. 47(7): p. 1241-9.
58. Strasser, E.F., Weidinger, T., Weiss, D.R., Strobel, J., Zimmermann, R., und
Eckstein, R., Storage induced apoptosis of peripheral blood mononuclear cells

Literaturverzeichnis 55
obtained from leucoreduction system chambers. Vox Sang, 2011. 101(2): p.
106-11.
59. Strasser, E.F., Weidinger, T., Zimmermann, R., Ringwald, J., und Eckstein, R.,
Recovery of white blood cells and platelets from leukoreduction system
chambers of Trima Accel and COBE Spectra plateletpheresis devices.
Transfusion, 2007. 47(10): p. 1943-4; author reply 1944-5.
60. Traganos, F., Flow cytometry: principles and applications. I. Cancer Invest,
1984. 2(2): p. 149-63.
61. Traganos, F., Flow cytometry: principles and applications. II. Cancer Invest,
1984. 2(3): p. 239-58.
62. Tuyaerts, S., Aerts, J.L., Corthals, J., Neyns, B., Heirman, C., Breckpot, K.,
Thielemans, K., und Bonehill, A., Current approaches in dendritic cell
generation and future implications for cancer immunotherapy. Cancer Immunol
Immunother, 2007. 56(10): p. 1513-37.
63. Van Furth, R. und Van Zwet, T.L., Immunocytochemical detection of 5-bromo-2-
deoxyuridine incorporation in individual cells. J Immunol Methods, 1988. 108(1-
2): p. 45-51.
64. Vanlangenakker, N., Vanden Berghe, T., und Vandenabeele, P., Many stimuli
pull the necrotic trigger, an overview. Cell Death Differ, 2011.
65. Vermes, I. und Haanen, C., Apoptosis and programmed cell death in health and
disease. Adv Clin Chem, 1994. 31: p. 177-246.
66. Vermes, I., Haanen, C., Steffens-Nakken, H., und Reutelingsperger, C., A novel
assay for apoptosis. Flow cytometric detection of phosphatidylserine expression
on early apoptotic cells using fluorescein labelled Annexin V. Journal of
immunological methods, 1995. 184(1): p. 39-51.
67. Villablanca, E.J., Raccosta, L., Zhou, D., Fontana, R., Maggioni, D., Negro, A.,
Sanvito, F., Ponzoni, M., Valentinis, B., Bregni, M., Prinetti, A., Steffensen,
K.R., Sonnino, S., Gustafsson, J.A., Doglioni, C., Bordignon, C., Traversari, C.,
und Russo, V., Tumor-mediated liver X receptor-alpha activation inhibits CC
chemokine receptor-7 expression on dendritic cells and dampens antitumor
responses. Nat Med, 2010. 16(1): p. 98-105.

Literaturverzeichnis 56
68. Weidinger, T.M., Keller, A.K., Weiss, D., Zimmermann, R., Eckstein, R., und
Strasser, E.F., Peripheral blood mononuclear cells obtained from
leukoreduction system chambers show better viability than those from
leukapheresis. Transfusion, 2011. 51(9): p. 2047-9.

Abkürzungen 57
VIII. Abkürzungsverzeichnis
7-AAD: 7-Amino-Actinomycin D
APC: Allophycocyanin
BMI: Body Mass Index
CD: engl. Cluster of Differentiation, Oberflächenantigene
D-TA: Doppelt-Thrombozytapherese
DNS: Desoxyribonucleinsäure
E-TA: Einfach-Thrombozytapherese
FACS: engl. Fluorescence activated cell sorting, Durchflusszytometrie
FDA: Food and Drug Administration, US-amerikanische Gesundheitsbe-
hörde
FITC: Fluorescein isothiocyanat
FL1 – 4: Fluorescence 1 – 4
FSC: Foreward light scatter
LRSC: engl. leukoreductionsystem chamber, Leukozytenreduktionskammer
PBMC: engl. Peripheral blood mononuclear cells, mononukleäre Zellen des
peripheren Bluts
PBS: engl. Phosphate buffered saline, Phosphat-gepufferte Salzlösung
PBV: prozessiertes Blutvolumen
PE: Pycoerythrin
PLT: engl. platelets, Thrombozyten
PVC: Polyvinylchlorid
RBC: engl. red blood cells, Erythrozyten
SSC: Sideward light scatter
TBV engl. total blood volume, gesamtes Blutvolumen
VAC alpha: vascular anticoagulant alpha
WBC: engl. white blood cells, Leukozyten

Verzeichnis der Vorveröffentlichungen 58
IX. Verzeichnis der Vorveröffentlichungen
1. Strasser EF, Weidinger T, Zimmermann R, Ringwald J, Eckstein R. Recovery of
white blood cells and platelets from leukoreduction system chambers of Trima Ac-
cel and COBE Spectra plateletpheresis devices. Transfusion 2007;47:1943-4.
(Letter)
2. Strasser EF, Weidinger T, Weiss DR, Strobel J, Zimmermann R, Eckstein R. Stor-
age induced apoptosis of peripheral blood mononuclear cells obtained from leu-
coreduction system chambers. Vox Sang 2011;101(2):106-11.
3. Weidinger TM, Keller AK, Weiss D, Zimmermann R, Eckstein R, Strasser EF. Pe-
ripheral blood mononuclear cells obtained from leukoreduction system chambers
show better viability than those from leukapheresis. Transfusion 2011;51:2047-9.
(Letter)

Anhang 59
X. Anhang
Tab. A1: Chemikalien und Biochemikalien Hersteller
- 7-AAD BD Pharmingen
- Annexin-V-FITC Apoptosis Detection Kit BD Pharmingen
- Annexin-V-FITC BD Pharmingen
- Aqua ad iniectabilia 100ml Braun
- CD14 – APC BD Pharmingen
- CD45 – PE BD Pharmingen
- FACSRinse / FACSFlow BD Biosciences
- PBS Wash Buffer (1X) Girco
- PharM-Lyse™ Lysis Buffer (10X concentrate) BD Pharmingen
Tab. A2: Geräte Hersteller
- Blutgasanalysator ABL5 Radiometer SA
- COBE® Spectra Zellseparator Caridian BCT
- Com.Tec® Zellseparator Fresenius HemoCare GmbH
- Durchflusszytometer: FACSCalibur BD Biosciences
- FACS Messröhrchen BD Biosciences
- FALCON 5 ml Round-Bottom Tubes BD Biosciences
- Kanüle Luer 21G/1,5“, 0,8x40mm BD Microlance 3
- Monovette 2,7ml K3E mit 1,6mg EDTA/ml Blut Sarstedt AG
- Monovettenkappe Sarstedt AG
- PVC-Beutel, 150ml Compoflex R Single Bags
- Sampling Site Baxter
- Spritze 2ml, 5ml, 10ml BD Discardit II
- Thrombozyteninkubator Forma Scientific
- Trima Accel® Zellseparator Gambro BCT
- Vortex Genie 2 Scientific Industries
- Zellzähler: ADVIA 120 Bayer HealthCare Diagnostics
- Zentrifugen: Rotina 48 S Hettich Zentrifugen
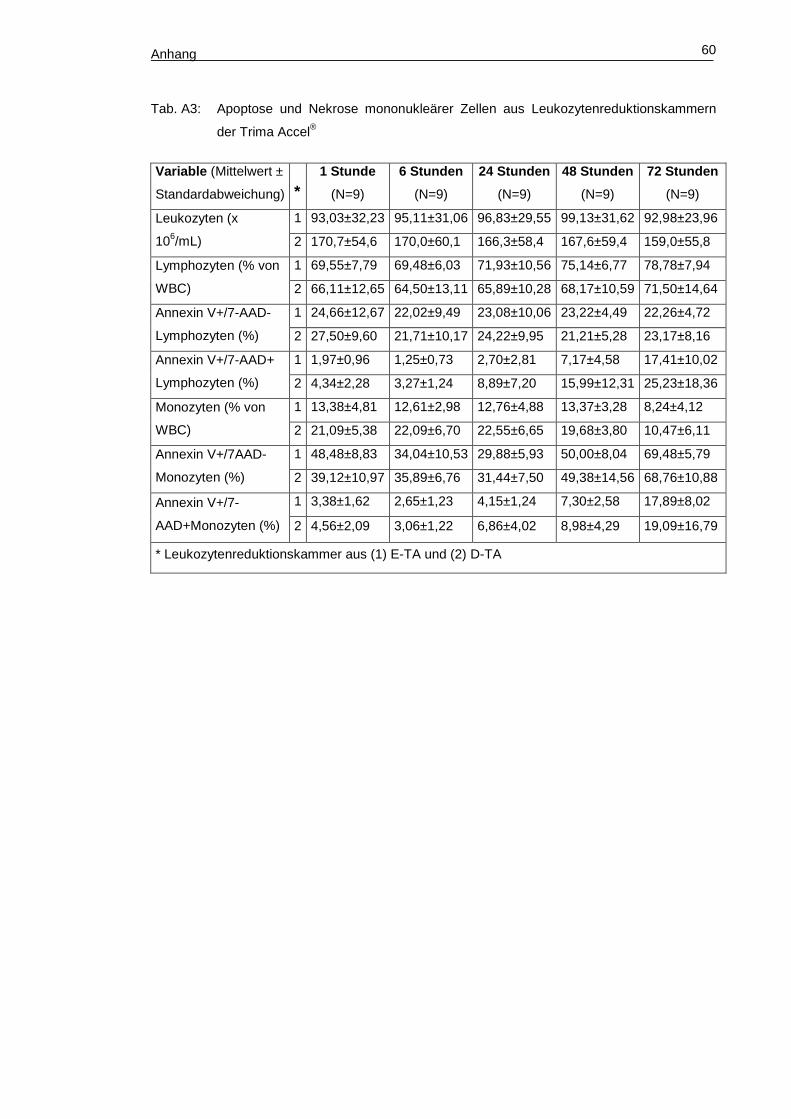
Anhang 60
Tab. A3: Apoptose und Nekrose mononukleärer Zellen aus Leukozytenreduktionskammern
der Trima Accel®
Variable (Mittelwert ±
Standardabweichung) *
1 Stunde
(N=9)
6 Stunden
(N=9)
24 Stunden
(N=9)
48 Stunden
(N=9)
72 Stunden
(N=9)
Leukozyten (x
106/mL)
1 93,03±32,23 95,11±31,06 96,83±29,55 99,13±31,62 92,98±23,96
2 170,7±54,6 170,0±60,1 166,3±58,4 167,6±59,4 159,0±5 5,8
Lymphozyten (% von
WBC)
1 69,55±7,79 69,48±6,03 71,93±10,56 75,14±6,77 78,78±7,94
2 66,11±12,65 64,50±13,11 65,89±10,28 68,17±10,59 71,50±14,64
Annexin V+/7-AAD-
Lymphozyten (%)
1 24,66±12,67 22,02±9,49 23,08±10,06 23,22±4,49 22,26±4,72
2 27,50±9,60 21,71±10,17 24,22±9,95 21,21±5,28 23,17±8,16
Annexin V+/7-AAD+
Lymphozyten (%)
1 1,97±0,96 1,25±0,73 2,70±2,81 7,17±4,58 17,41±10,02
2 4,34±2,28 3,27±1,24 8,89±7,20 15,99±12,31 25,23±18,36
Monozyten (% von
WBC)
1 13,38±4,81 12,61±2,98 12,76±4,88 13,37±3,28 8,24±4, 12
2 21,09±5,38 22,09±6,70 22,55±6,65 19,68±3,80 10,47±6 ,11
Annexin V+/7AAD-
Monozyten (%)
1 48,48±8,83 34,04±10,53 29,88±5,93 50,00±8,04 69,48±5,79
2 39,12±10,97 35,89±6,76 31,44±7,50 49,38±14,56 68,76±10,88
Annexin V+/7-
AAD+Monozyten (%)
1 3,38±1,62 2,65±1,23 4,15±1,24 7,30±2,58 17,89±8,02
2 4,56±2,09 3,06±1,22 6,86±4,02 8,98±4,29 19,09±16,79
* Leukozytenreduktionskammer aus (1) E-TA und (2) D-TA
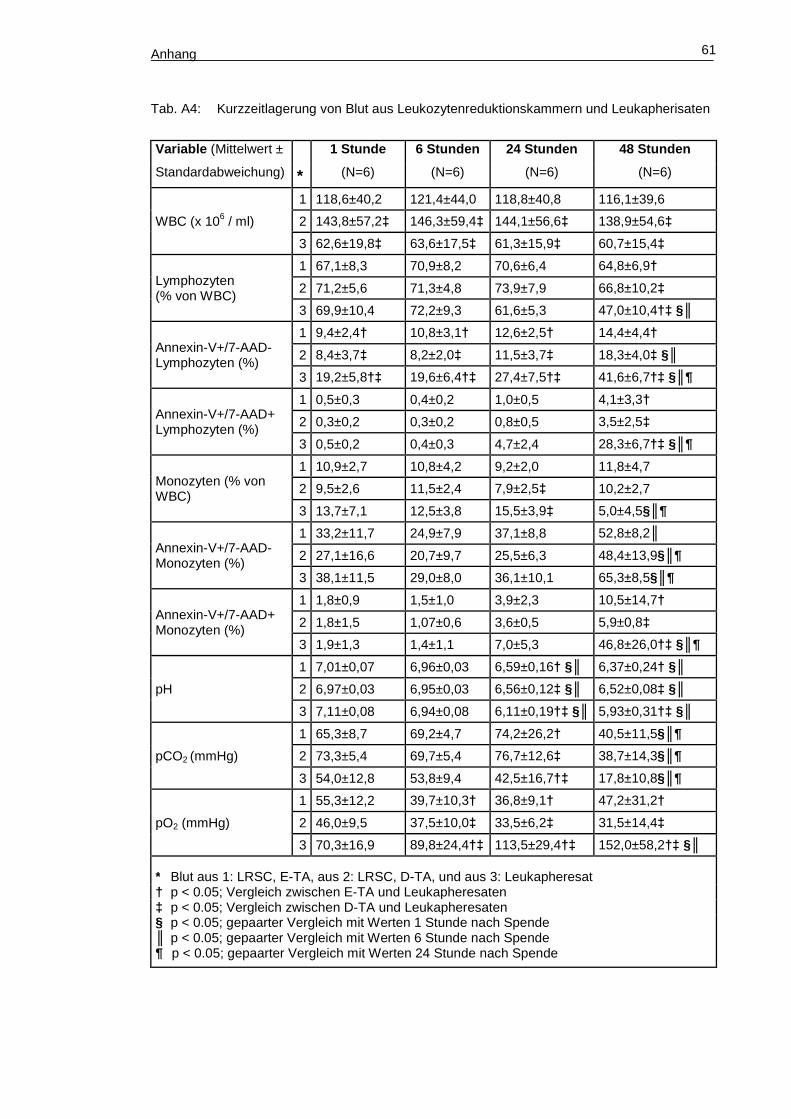
Anhang 61
Tab. A4: Kurzzeitlagerung von Blut aus Leukozytenreduktionskammern und Leukapherisaten
Variable (Mittelwert ±
Standardabweichung)
1 Stunde
(N=6)
6 Stunden
(N=6)
24 Stunden
(N=6)
48 Stunden
(N=6) *
WBC (x 106 / ml)
1 118,6±40,2 121,4±44,0 118,8±40,8 116,1±39,6
2 143,8±57,2‡ 146,3±59,4‡ 144,1±56,6‡ 138,9±54,6‡
3 62,6±19,8‡ 63,6±17,5‡ 61,3±15,9‡ 60,7±15,4‡
Lymphozyten (% von WBC)
1 67,1±8,3 70,9±8,2 70,6±6,4 64,8±6,9†
2 71,2±5,6 71,3±4,8 73,9±7,9 66,8±10,2‡
3 69,9±10,4 72,2±9,3 61,6±5,3 47,0±10,4†‡ §║
Annexin-V+/7-AAD- Lymphozyten (%)
1 9,4±2,4† 10,8±3,1† 12,6±2,5† 14,4±4,4†
2 8,4±3,7‡ 8,2±2,0‡ 11,5±3,7‡ 18,3±4,0‡ §║
3 19,2±5,8†‡ 19,6±6,4†‡ 27,4±7,5†‡ 41,6±6,7†‡ §║¶
Annexin-V+/7-AAD+ Lymphozyten (%)
1 0,5±0,3 0,4±0,2 1,0±0,5 4,1±3,3†
2 0,3±0,2 0,3±0,2 0,8±0,5 3,5±2,5‡
3 0,5±0,2 0,4±0,3 4,7±2,4 28,3±6,7†‡ §║¶
Monozyten (% von WBC)
1 10,9±2,7 10,8±4,2 9,2±2,0 11,8±4,7
2 9,5±2,6 11,5±2,4 7,9±2,5‡ 10,2±2,7
3 13,7±7,1 12,5±3,8 15,5±3,9‡ 5,0±4,5§║¶
Annexin-V+/7-AAD- Monozyten (%)
1 33,2±11,7 24,9±7,9 37,1±8,8 52,8±8,2║
2 27,1±16,6 20,7±9,7 25,5±6,3 48,4±13,9§║¶
3 38,1±11,5 29,0±8,0 36,1±10,1 65,3±8,5§║¶
Annexin-V+/7-AAD+ Monozyten (%)
1 1,8±0,9 1,5±1,0 3,9±2,3 10,5±14,7†
2 1,8±1,5 1,07±0,6 3,6±0,5 5,9±0,8‡
3 1,9±1,3 1,4±1,1 7,0±5,3 46,8±26,0†‡ §║¶
pH
1 7,01±0,07 6,96±0,03 6,59±0,16† §║ 6,37±0,24† §║
2 6,97±0,03 6,95±0,03 6,56±0,12‡ §║ 6,52±0,08‡ §║
3 7,11±0,08 6,94±0,08 6,11±0,19†‡ §║ 5,93±0,31†‡ §║
pCO2 (mmHg)
1 65,3±8,7 69,2±4,7 74,2±26,2† 40,5±11,5§║¶
2 73,3±5,4 69,7±5,4 76,7±12,6‡ 38,7±14,3§║¶
3 54,0±12,8 53,8±9,4 42,5±16,7†‡ 17,8±10,8§║¶
pO2 (mmHg)
1 55,3±12,2 39,7±10,3† 36,8±9,1† 47,2±31,2†
2 46,0±9,5 37,5±10,0‡ 33,5±6,2‡ 31,5±14,4‡
3 70,3±16,9 89,8±24,4†‡ 113,5±29,4†‡ 152,0±58,2†‡ §║ * Blut aus 1: LRSC, E-TA, aus 2: LRSC, D-TA, und aus 3: Leukapheresat † p < 0.05; Vergleich zwischen E-TA und Leukapheresaten ‡ p < 0.05; Vergleich zwischen D-TA und Leukapheresaten § p < 0.05; gepaarter Vergleich mit Werten 1 Stunde nach Spende ║ p < 0.05; gepaarter Vergleich mit Werten 6 Stunde nach Spende ¶ p < 0.05; gepaarter Vergleich mit Werten 24 Stunde nach Spende

62
Danksagung
Ich möchte mich ganz herzlich bei Herrn Prof. Dr. med. R. Eckstein für Überlassung
des Themas und die großzügige Bereitstellung des Arbeitsplatzes in seiner Abteilung
bedanken.
Für die hervorragende Betreuung, stetige Anleitung und Förderung während der Erstel-
lung der Arbeit und auch danach möchte ich Herrn PD Dr. med. E. Strasser danken.
Für die entgegenkommende Unterstützung bei der Lösung statistischer Probleme dan-
ke ich Frau Dipl.-Stat. FH A.K. Keller.
Auch bedanke ich mich bei den Schwestern und MTAs in der Blutspende und in den
Laboratorien der Transfusionsmedizinischen Abteilung der Universitätsklinik Erlangen
für deren Hilfe im Rahmen ihrer Möglichkeiten.