dactylogyrus (monogenea: dactylogyridae) from seven...
TRANSCRIPT

Proc. Helminthol. Soc. Wash.55(2), 1988, pp. 214-223
Dactylogyrus (Monogenea: Dactylogyridae) from Seven Species ofNotropis (Pisces: Cyprinidae) from the Tennessee River Drainage:Descriptions of Four New Species and Remarks on Host Relationships
DONALD G. CLOUTMANDuke Power Company, Production Environmental Services, Route 4, Box 531,Huntersville, North Carolina 28078
ABSTRACT: Five previously described and 5 new species of Dactylogyrus are reported from Notropis speciesfrom the Tennessee River drainage: D. bulbus Mueller, 1938, D. lepidus Rogers, 1967, D. luxili Rogers, 1967,D. manicatus Rogers, 1967, and D. peiius Mueller, 1938, occurred on N. chrysocephalus chrysocephalus; Dac-tylogyrus sp. cf. sceptici Cloutman, 1980, occurred on N. ariommus; D. circumflexus sp. n. and D. delicatus sp.n. are described from N. c. chrysocephalus; D. dolus sp. n. is described from N. coccogenis; D. spatulus sp. n. isdescribed from N. telescopus and was also found on N. ariommus, N. coccogenis, N. leuciodus, and N. rubricroceus.Based on examination of type specimens, D. per/us is considered here to be a senior synonym of D. banghamiMizelle and Donahue, 1944. No Dactylogyrus species were found on N. rubellus. Similarities in the Dactylogyrusspecies infesting N. c. chrysocephalus, N. c. isolepis, and N. cornutus corroborate ichthyological evidence thatthese hosts are very close relatives in the N. cornutus species group of Notropis (Luxilus). The presence of D.dolus and D. spatulus (close relatives of D. manicatus and D. dubius Mizelle and Klucka, 1953, respectively,that occur on certain members of the N. cornutus species group) on N. coccogenis indicate a not so closerelationship between N. coccogenis and the N. cornutus species group within Notropis (Luxilus). The presenceof D. spatulus on N. ariommus, N. coccogenis, N. leuciodus, N. rubricroceus, and N. telescopus indicates a possiblephyletic link between these hosts, which has only in part been suggested through ichthyological studies. Thepresence of Dactylogyrus sp. cf. sceptici on N. ariommus corroborates ichthyological evidence of a close rela-tionship between N. ariommus and N. scepticus.
KEY WORDS: Monogenea, Dactylogyridae, morphology, systematics, Dactylogyrus circumflexus sp. n., Dac-tylogyrus delicatus sp. n., Dactylogyrus dolus sp. n., Dactylogyrus spatulus sp. n., Dactylogyrus banghami assynonym, Dactylogyrus bulbus, Dactylogyrus lepidus, Dactylogyrus luxili, Dactylogyrus manicatus, Dactylogyrusperlus, Notropis spp., Cyprinidae, shiners, prevalence.
Twenty-seven species of Dactylogyrus Dies- were made as presented by Mizelle and Klucka (1953);ing, 1850, on 18 species of hosts have been re- mcans are followed bY ran8es in parentheses. Al l type
, f. I T - TT j /T-> specimens of new species and representative specimensported from the Tennessee River drainage (Rog- Qpf previously descrPlbed species were deposited in theers, 1967; Chien, 1971, 1974a, b; Cloutman, helminthological collection of the National Museum1987). This paper reports on 5 previously de- of Natural History (USNM). Other nontype materialscribed and 5 new species of Dactylogyrus from is in the author's collection. For comparative purposes,7 species of the cyprinid genus Notropis Ran- f original descriptions and redemptions of North
* . . , , - _ . /. T* American Dactylogyrus species and specimens of thenesque, bringing the total to 35 species of Dae- foliowing species from the USNM, the Harold W.tylogyrus known from 22 species of hosts from Manter Laboratory, University of Nebraska State Mu-the Tennessee River drainage. Also, evolution- seum (HWML), and the collection of Dr. Wilmer A.ary relationships of hosts based on infesting Dae- R°8ers <WAR)were examined: D. acus Mueller, 1938,
, A- A 5 syntypes (USNM 71445, 71446); D. arcus Rogers,tylogyrus species are discussed. 1%7; holotype (U§NM 61369)> 6 paratypes (USNM
Material s and Methods 61370J1 specimen]and WAR [5 specimens]); I) tejg-harm Mizelle and Donahue, 1944, 3 syntypes (USNM
The species and numbers of hosts examined are list- 73552[1 specimen] and HWML 21545 [2 specimens]),ed in Table 1. With the exception of museum speci- 4 nontype specimens (USNM 73161); Z). bulbus Muel-mens of Notropis ariommus (Cope), the hosts were ler, 1938, 16 syntypes (USNM 71454, 71457); D. car-placed in jars containing a 1:4,000 formalin solution nutus Mueller, 1938, 6 syntypes (USNM 71454, 71457);immediately after capture; after approximately 1 hr, D. cursitans Rogers, 1967, 1 paratype (USNM 61380);enough formalin was added to make a 10% solution D. dubius Mizelle and Klucka, 1953, 4 nontype spec-(Putz and Hoffman, 1963). Al l parasites, collected from imens (HWML 20535); D. fulcrum Mueller, 1938, 2the gills of their hosts, were mounted in glycerin jelly, syntypes (USNM 71457); D. lepidus Rogers, 1967, ho-and observations were made with a Zeiss phase-con- lotype (USNM 61389) and 2 paratypes (USNM 61390);trast microscope. Drawings were made with the aid of D. luxili Rogers, 1967, 1 paratype (USNM 61394); D.a Zeiss drawing tube. Measurements, in micrometers, manicatus Rogers, 1967, 1 paratype (USNM 61397);
214
Copyright © 2011, The Helminthological Society of Washington

215
Table 1. Prevalence (°/o infestation), range, and relative density (total number of parasites/total number of hosts)of Dactylogyrus infesting 6 species of Notropis from the Tennessee River drainage. Numbers in parenthesesrepresent the number of host specimens examined.
Species Range Relative density
Notropis ariommus (Cope), popeye shiner (21)
Dactylogyrus sp. cf. sceptic/ Cloutman, 1980Dactylogyrus spatulus sp. n.
Notropis (Luxilus) chrysocephalus chrysocephalus (Rafmesque),
Dactylogyrus perlus Mueller, 1938Dactylogyrus bulbus Mueller, 1938Dactylogyrus circumflcxus sp. n.Dactylogyrus delicatus sp. n.Dactylogyrus lepidus Rogers, 1 967Dactylogyrus luxi/i Rogers, 1967Dactylogyrus manicatus Rogers, 1967
Notropis (Luxilus) coccogcnis (Cope), warpaint shiner (9)
Dactylogyrus dolus sp. n.Dactylogyrus spatulus sp. n.
Notropis (Hydrophlox) leuciodus (Cope), Tennessee shiner (21)
Dactylogyrus spatulus sp. n.
Notropis (Hydrophlox) ruhcllus (Agassi/), rosyfacc shiner (16)
Notropis (Hydrophlox) rubricroceus (Cope), saffron shiner (25)
Dactylogyrus spatulus sp. n.
Notropis telescopus (Cope), telescope shiner (10)
Dactylogyrus spatulus sp. n.
9.55.0
striped shiner (7)
71.471.485.757.157.1
100.042.9
66.733.3
19.0
8.0
90.0
0-50-7
0-70-90-60-30-21-30-2
0-140-23
0-2
0-16
0-22
0.30.3
2.93.03.01.40.71.90.6
5.04.9
0.2
0.8
8.2
D. perlus Mueller, 1938, 3 syntypes (USNM 71454,71457), and 1 nontype specimen (HWML 21305).
Results
Dactylogyrus bulbus Mueller, 1938
HOST: Notropis chrysocephalus chrysoceph-alus (Rafinesque), striped shiner.
LOCALITIES: Tennessee: Blount Co., Littl eRiver near Waland (USNM 80175, 2 speci-mens); Littl e River, Hwy. 411 bridge (USNM80176, 2 specimens).
REMARKS: Dactylogyrus bulbus is widely dis-tributed on Notropis chrysocephalus and N. cor-nutus (Mitchill ) in eastern North America (Muel-ler, 1938; Mizelle and Donahue, 1944; Mizelleand Klucka, 1953; Mizelle and Webb, 1953;Rogers, 1967; Molnar et al., 1974; Hanek et al.,1975). A record from Notemigonus crysoleucas(Mitchill ) (Cone, 1980) is considered here to rep-resent an abnormal infestation. Dactylogyrusbulbus appears to be specific for N. c. chrysoceph-alus in the Tennessee River drainage (Table 1;Rogers, 1967; Cloutman, 1987).
Dactylogyrus lepidus Rogers, 1967
HOST: Notropis chrysocephalus chrysoceph-alus (Rafinesque), striped shiner.
LOCALITIES: Tennessee: Blount Co., Littl eRiver near Waland (USNM 80177, 3 speci-mens); Littl e River, Hwy. 411 bridge.
REMARKS: This is the first report of Dacty-logyrus lepidus since its original description fromNotropis chrysocephalus isolepis Hubbs andBrown in Alabama (Rogers, 1967). Dactylogyruslepidus appears to be restricted to the 2 subspe-cies of TV. chrysocephalus. Its closest relative ap-pears to be D. fulcrum, found on N. cornutus inNew York (Mueller, 1938).
Dactylogyrus luxili Rogers, 1967
HOST: Notropis chrysocephalus chrysoceph-alus (Rafinesque), striped shiner.
LOCALITIES: Tennessee: Blount Co., Littl eRiver near Waland (USNM 80178, 4 speci-mens); Littl e River, Hwy. 411 bridge.
REMARKS: Dactylogyrus luxili has been re-ported on Notropis chrysocephalus, N. cornutus,and N. pilsbryi Fowler from Alabama, Ontario,and Arkansas, respectively (Rogers, 1967; Haneket al., 1975). Thus, all known hosts are speciesof Notropis (Luxilus). However, D. luxili doesnot appear to infest all Notropis (Luxilus); forexample, it has not been found on N. (L.) coc-cogenis (Rogers, 1967; present study).
Copyright © 2011, The Helminthological Society of Washington

216 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Dactylogyrus manicatus Rogers, 1967
HOST: Notropis chrysocephalus chrysoceph-alus (Rafinesque), striped shiner.
LOCALITIES: Tennessee: Blount Co., Littl eRiver near Waland (USNM 80179, 2 speci-mens); Littl e River, Hwy. 411 bridge.
REMARKS: This is the first report of Dacty-logyrus manicatus since its original descriptionfrom Notropis chrysocephalus isolepis in Ala-bama (Rogers, 1967). Dactylogyrus manicatusappears to parasitize only the 2 subspecies of N.chrysocephalus.
Dactylogyrus perlus Mueller, 1938
HOST: Notropis chrysocephalus chrysoceph-alus (Rafinesque), striped shiner.
LOCALITIES: Tennessee: Blount Co., Littl eRiver near Waland (USNM 80180, 4 speci-mens); Littl e River, Hwy. 411 bridge.
REMARKS: The nominal form Dactylogyrusperlus has been previously reported from Notro-pis cornutus from New York (Mueller, 1938) andOntario (Mizelle and Donahue, 1944). In thesame paper that they reported D. perlus basedon 1 specimen, Mizelle and Donahue (1944) de-scribed D. banghami based on 3 specimens. The2 species were supposedly distinguished by thepresence of an anteriorly directed process on thecirrus base of D. banghami and the lack of sucha process in D. perlus, as interpreted from Muel-ler's (1938) drawing. Mizelle and Donahue (1944)noted, however, that the accessory pieces of these2 species were of the same type. After examiningthe syntypes and Mizelle and Donahue's nontypespecimen of D. perlus, syntypes of D. banghami,and 13 specimens from the Tennessee Riverdrainage, I propose that D. perlus is a seniorsynonym of D. banghami. The specimens inMueller's (1938) syntype series possess a distinctanteriorly directed process on the cirrus, and oth-er sclerites are of the same general size and shapein both species.
Dactylogyrus perlus has been reported as D.banghami from several host species and locali-ties in eastern North America (Mizelle andMcDougal, 1970; Cloutman, 1974; Hanek et al.,1975; Beverley-Burton, 1984), but appears re-stricted to N. chrysocephalus chrysocephalus inthe Tennessee River drainage. Hanek et al. (1975)noted that some specimens reported as D. bang-hami from various host species exhibit differ-ences in the structure of the copulatory com-plexes and represent a species-group complex
comprising several species. After reexaminingspecimens that I reported as D. banghami fromN. lutrensis (Baird and Girard) and N. stramineus(Cope) (Cloutman, 1974), I believe they repre-sent D. beckeri Cloutman, 1987, from TV. lutren-sis and an undescribed species from N. strami-neus. Thus, I concur with the conclusion of Haneket al. (1975) that many specimens reported as D.banghami represent a complex of several species.A thorough review of this complex is needed.
Dactylogyrus sp. cf. sceptici Cloutman, 1980
HOST: Notropis ariommus (Cope), popeyeshiner.
LOCALITIES: Tennessee: Scott Co., North ForkHolston River, 0.5 km E of junction of Routes614 and 778. Virginia: specific locality unknown,Powell River.
REMARKS: This undescribed member of theDactylogyrus perlus species complex and its clos-est apparent relative, D. sceptici, have almostidentically shaped sclerites. The 2 species differmost notably in the size of the copulatory ap-paratus: Dactylogyrus sp. cirrus length 45 (43-48), accessory piece length 31 (30-32); D. scepticicirrus length 58 (53-61), accessory piece length43 (37-44) (Cloutman, 1980). Dactylogyrus sp.wil l be named and described when enough spec-imens suitable for type material become avail-able.
Dactylogyrus circumflexus sp. n.(Figs. 1-9)
TYPE HOST: Notropis chrysocephalus chryso-cephalus (Rafinesque), striped shiner.
TYPE LOCALITY: Tennessee: Blount Co., Lit -tle River near Waland.
TYPE SPECIMENS: Holotype, USNM 80181;10 paratypes, USNM 80182 (1 specimen), USNM80183 (3 specimens), and USNM 80184 (6 spec-imens).
OTHER LOCALITY: Tennessee: Blount Co.,Littl e River, Hwy. 411 bridge.
DESCRIPTION: With characters of the genus asemended by Mizelle and McDougal (1970). Bodywith thin tegument; length 287 (173-382), great-est width 71 (43-94). Two pairs of anterior ce-phalic lobes, lateral pair smaller than medial pair.Head organs not observed. Two pairs of eyesapproximately equal in size, anterior pair usuallyfarther apart than posterior pair. Pharynx cir-
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 55, NUMBER 2, JULY 1988 217
D. CIRCUMFLEXUS D. DELICATUS
20 urn
D. DOLUS D. SPATULUSFigures 1-37. Sclerotized parts of Dactylogyrus species (drawings are of holotypes unless otherwise specified).
1-9. Dactylogyrus circumflexus sp. n.: 1, 3 (USNM 80182), cirrus; 2, 4 (USNM 80182), accessory piece; 5,anchor; 6, dorsal bar; 7, ventral bar; 8, 9, hooks. 10-19. Dactylogyrus delicatus sp. n.: 10, 12 (USNM 80186),cirrus; 11, 13 (USNM 80186), accessory piece; 14, anchor; 15, dorsal bar; 16, 17 (USNM 80186), ventral bar;18, 19, hooks. 20-28. Dactylogyrus dolus sp. n.: 20, 22 (USNM 80194), cirrus; 21, 23 (USNM 80194), accessorypiece; 24, anchor; 25, dorsal bar; 26, ventral bar; 27, 28, hooks. 29-37. Dactylogyrus spatulus sp. n.: 29, 31(USNM 80196), cirrus; 30, 32 (USNM 80196), accessory piece; 33, anchor; 34, dorsal bar; 35, ventral bar; 36,37, hooks.
cular (dorsal view), transverse diameter 21 (18-26), gut not observed.
Peduncle usually present, 38 (14-91) long, 31(24-42) wide. Haptor 48 (36-70) long, 61 (49-70) wide. Single pair of dorsal anchors; each com-
posed of solid base with short deep root, elongatesuperficial root, and solid shaft curving to a sharppoint. Anchor length 30 (29-32), greatest widthof base 13(11-14). Dorsal bar length 21(17-24).Vestigial ventral bar length 20 (17-22). Sixteen
Copyright © 2011, The Helminthological Society of Washington

218 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
hooks (8 pairs), similar in shape (except 4A),normal in arrangement (Mizelle and Crane, 1964).Each hook composed of solid base, solid slendershaft, and sickle-shaped termination providedwith opposable piece (opposable piece lacking in4A). Hook lengths: No. 1, 18 (16-19); 2, 19 (15-20); 3, 23 (17-25); 4, 21 (20-22); 4A, 6; 5, 20(18-22); 6, 20 (17-22); 7, 20 (19-21). Cirrus anelongate arcuate tube with expanded base, length43 (38-46). Accessory piece a solid slender rodarching closely alongside cirrus shaft to a posi-tion approximately '/4-'/3 of the length of the cir-rus shaft from the terminus of the cirrus shaft,proximal end biramous, length 23 (20-27). Va-gina not observed. Vitellaria usually moderate,usually distributed from pharynx to haptor.
REMARKS: Dactylogyrus circumflexus mostclosely resembles D. arcus, D. dubius, and D.spatulus sp. n. by possessing an elongate arcuatecirrus with the accessory piece arching closelyalongside. However, D. circumflexus possessesa shorter accessory piece (extends approximatelyto '/4-1/? of the length of the cirrus shaft from theterminus of the cirrus shaft) than D. arcus, D.dubius, or D. spatulus (extends approximately toterminus of cirrus shaft). The cirrus shaft of D.arcus, D. circumflexus, and D. dubius is approx-imately the same diameter for its entire length,whereas that of D. spatulus gradually broadensinto a spatulate shape distally. Hanek et al. (1975)considered D. arcus to be a junior synonym ofD. dubius. However, based on examination ofnontype specimens conforming to the originaldescription by Mizelle and Klucka (1953) andthe redescription by Hanek et al. (1975) of D.dubius and type specimens of D. arcus, I considerD. arcus and D. dubius to be separate species.The proximal end of the accessory piece of D.arcus is biramous and not articulated with thecirrus base; the short ramus is sometimes notobserved, the long ramus is bent at a right anglefrom the accessory piece shaft (see Rogers, 1967).The proximal end of the accessory piece of D.dubius is uniramous and articulated with the cir-rus base, and a delicate ramus arises near themidlength of the accessory piece (see Mizelle andKlucka, 1953, and Hanek et al., 1975). I havebeen unable to locate the type specimen of D.dubius, which consists of a fragment containingthe cirrus and accessory piece but lacking thehaptor.
ETYMOLOGY: The species name is Latin (cir-cumflexus = arched), referring to the arcuate cir-rus shaft.
Dactylogyrus delicatus sp. n.(Figs. 10-19)
TYPE HOST: Notropis chrysocephalus chryso-cephalus (Rafinesque), striped shiner.
TYPE LOCALITY: Tennessee: Blount Co., Lit-tle River near Waland.
TYPE SPECIMENS: Holotype, USNM 80185; 3paratypes, USNM 80186 (1 specimen), USNM80187 (1 specimen), and USNM 80188 (1 spec-imen).
OTHER LOCALITY : Tennessee: Blount Co.,Littl e River, Hwy. 411 bridge.
DESCRIPTION: With characters of the genus asemended by Mizelle and McDougal (1970). Bodywith thin tegument; length 320 (252-425), great-est width 74 (50-94). Two pairs of anterior ce-phalic lobes, lateral pair smaller than medial pair.Head organs not observed. Two pairs of eyesapproximately equal in size, anterior pair fartherapart than posterior pair. Pharynx circular (dor-sal view), transverse diameter 21, gut not ob-served. Peduncle 24 (14-33) long, 25 (28-42)wide. Haptor 39 (35-42) long, 60 (52-74) wide.Single pair of dorsal anchors; each composed ofsolid base with short deep root and elongate su-perficial root, and solid shaft curving to a sharppoint. Anchor length 28 (27-28), greatest widthof base 13 (11-14).
Dorsal bar length 21 (20-22). Vestigial ventralbar length 20 (18-21). Sixteen hooks (8 pairs),similar in shape (except 4A), normal in arrange-ment (Mizelle and Crane, 1964). Each hook com-posed of solid base, solid slender shaft, and sick-le-shaped termination provided with opposablepiece (opposable piece lacking in 4A). Hooklengths: No. 1, 18 (16-19); 2, 21 (20-21); 3, 23(22-25); 4, 20 (18-21); 4A, 5; 5, 20 (18-22); 6,20 (19-21); 7, 19 (18-20). Copulatory complexcomposed of cirrus and articulated accessorypiece. Cirrus a curved tube with diagonally trun-cate point and expanded base, length 28 (26-29).Accessory piece bent near cirrus base and bentagain before bifurcating; distal ramus curved, at-tenuated; medial ramus possessing short, bluntprojection and long, thin, delicate, curved pro-jection attenuating to sharp point; length 15(13-16). Vagina not observed. Vitellaria moderate,distributed from pharynx to haptor.
REMARKS: Dactylogyrus delicatus mostclosely resembles D. lepidus in the shape of thecirrus, but the 2 species can be distinguished bythe morphology of the accessory piece. The bi-furcated accessory piece of D. delicatus possesses
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 55, NUMBER 2, JULY 1988 • 219
a medial ramus with a short blunt projection anda long, delicate, sharply pointed distal ramus,whereas the accessory piece of D. lepidus isU-shaped (Rogers, 1967).
ETYMOLOGY: The species name is Latin (del-icatus = delicate), referring to the delicate pro-jection on the medial ramus of the accessory piece.
Dactylogyrus dolus sp. n.(Figs. 20-28)
TYPE HOST: Notropis coccogenis (Cope), war-paint shiner.
TYPE LOCALITY : Tennessee: Monroe Co., Ci-tico Creek.
TYPE SPECIMENS: Holotype, USNM 80189;15 paratypes, USNM 80190(1 specimen), USNM80191 (1 specimen), USNM 80192 (3 speci-mens), USNM 80193 (9 specimens), and USNM80194 (1 specimen).
OTHER LOCALITIES: North Carolina: YanceyCo., South Toe River, Co. Rt. 1169 approxi-mately 8 km E of Micaville. Tennessee: BlountCo., Littl e River near Waland; Littl e River, Hwy.411 bridge.
DESCRIPTION: With characters of the genusDactylogyrus as emended by Mizelle andMcDougal (1970). Body with thin tegument;length 295 (173-396), greatest width 81 (43-115).Two pairs of anterior cephalic lobes, lateral pairsmaller than medial pair. Head organs not ob-served. Two pairs of eyes approximately equalin size, posterior pair usually farther apart thananterior pair. Pharynx circular (dorsal view),transverse diameter 21 (15-25), gut not ob-served. Peduncle 32 (15-60) long, 33 (22-46)wide. Raptor 38 (32-43) long, 61 (35-71) wide.Single pair of dorsal anchors; each composed ofsolid base with short deep root and elongate su-perficial root, and solid shaft curving to a sharppoint. Anchor length 29 (25-32), greatest widthof base 13(11-15). Dorsal bar length 20 (18-22).Vestigial ventral bar length 18 (17-21).
Sixteen hooks (8 pairs), similar in shape (ex-cept 4A), normal in arrangement (Mizelle andCrane, 1964). Each hook composed of solid base,solid slender shaft, and sickle-shaped termina-tion provided with opposable piece (opposablepiece lacking in 4A). Hook lengths: No. 1, 16(15-18); 2, 18 (16-22); 3, 22 (18-25); 4, 18 (16-20); 4A, 6 (5-7); 5, 17 (14-19); 6, 17 (14-19); 7,18 (15-20). Copulatory complex composed ofcirrus and articulated accessory piece. Cirrus withrounded base, bearing a slender slightly curved
process with a bulbous terminus, and curvingtubular shaft arched near distal end and withblunt terminus, length 25 (22-29). Accessory piecebifurcate with almost straight dagger-shaped ter-minus and curved ramus arising near distal thirdof shaft; a nonsclerotized "sleeve" often ob-served attached to ramus and enveloping end ofcirrus. Accessory piece length 18(16-21). Vaginanonsclerotized, opening dextrally at body marginposterior to cirrus. Vitellaria usually moderate,distributed from pharynx to haptor.
REMARKS: The closest apparent relative ofDactylogyrus dolus is D. manicatus. Both speciespossess a "sleeve" attaching to the ramus of theaccessory piece and enveloping the end of thecirrus. The major differences are: the cirrus shaftof D. dolus is arched most near the distal end,whereas the cirrus shaft of D. manicatus is highlyarched at the basal end; the terminus of the basalprocess of D. dolus possesses a small bulb, where-as that of D. manicatus does not; the terminusof the accessory piece of/), dolus is almost straightand dagger-shaped, whereas that ofD. manicatusis curved and attenuated to a point.
ETYMOLOGY: The name of this species is lat-inized Greek (dolos = dagger), referring to thedagger-shaped terminus of the accessory piece.
Dactylogyrus spatulus sp. n.(Figs. 29-37)
TYPE HOST: Notropis telescopus (Cope), tele-scope shiner.
TYPE LOCALITY : Tennessee: Blount Co., Lit-tle River near Waland.
TYPE SPECIMENS: Holotype, USNM 80195;19 paratypes, USNM 80196 (1 specimen) andUSNM 80197 (18 specimens).
OTHER HOSTS AND LOCALITIES: Notropisariommus (Cope)—Tennessee: Claiborne Co.,Powell River just upstream from Hwy. 25E bridge(USNM 80198, 2 specimens). Notropis cocco-genis (Cope)—North Carolina: Yancey Co., SouthToe River, Co. Rt. 1169 approximately 8 km Eof Micaville. Tennessee: Blount Co., Littl e Rivernear Waland; Littl e River, Hwy. 411 bridge(USNM 80199, 4 specimens); Monroe Co., Ci-tico Creek. Notropis leuciodus (Cope)—Tennes-see: Blount Co., Littl e River near Waland; Littl eRiver, Hwy. 411 bridge (USNM 80200, 1 spec-imen); Sevier Co., West Prong Littl e Pigeon Riv-er, 1.5 km downstream from Smoky Gap bridgebetween Gatlinburg and Pigeon Forge. Notropisrubricroceus (Cope)—Tennessee: Sevier Co., WestProng Littl e Pigeon River, 1.5 km downstream
Copyright © 2011, The Helminthological Society of Washington

220
from Smoky Gap bridge between Gatlinburg andPigeon Forge (USNM 80201, 2 specimens). No-tropis telescopus—Tennessee: Blount Co., Littl eRiver, Hwy. 411 bridge.
DESCRIPTION: With characters of the genusDactylogyrus as emended by Mizelle andMcDougal (1970). Body with thin tegument;length 308 (252-360), greatest width 106 (79-144). Two pairs of anterior cephalic lobes, lateralpair smaller than medial pair. Head organs notobserved. Two pairs of eyes approximately equalin size, posterior pair usually farther apart thananterior pair. Pharynx circular (dorsal view),transverse diameter 24 (20-31), gut not ob-served. Peduncle usually present, 20 (7-28) long,45 (27-73) wide. Haptor 53 (35-70) long, 67 (42-111) wide. Single pair of dorsal anchors; eachcomposed of solid base with short deep root andelongate superficial root, and solid shaft curvingto a sharp point. Anchor length 35 (28-39), great-est width of base 14 (11-18). Dorsal bar length24 (21-27). Vestigial ventral bar length 20 (18-22). Sixteen hooks (8 pairs), similar in shape (ex-cept 4A), normal in arrangement (Mizelle andCrane, 1964). Each hook composed of solid base,solid slender shaft, and sickle-shaped termina-tion provided with opposable piece (opposablepiece lacking in 4A). Hook lengths: No. 1, 18(14-21); 2, 20 (16-25); 3, 21 (18-24); 4, 20 (18-23); 4A, 6; 5, 20 (16-25); 6, 19 (15-22); 7, 19(16-22). Copulatory complex composed of cirrusand accessory piece. Cirrus with expanded baseand elongate arcuate shaft that gradually broad-ens toward distal end, length 42 (34-53). Acces-sory piece a solid slender rod arching closelyalongside approximately the entire length of thecirrus shaft, proximal end biramous, length 30(25-38). Vagina not observed. Vitellaria usuallymoderate, usually distributed from pharynx tohaptor.
REMARKS: The closest apparent relatives ofDactylogyrus spatulus are D. arcus, D. circum-flexus, and D. dubius (see Remarks for D. cir-cumflexus). Dactylogyrus arcus was describedfrom Notropis chrysocephalus and N. coccogenisby Rogers (1967). I examined the holotype and3 paratypes from N. chrysocephalus and 3 para-types from 7Y. coccogenis and found that the spec-imens from N. chrysocephalus are D. arcus, butthose from N. coccogenis are D. spatulus. Dac-tylogyrus spatulus exhibits a wider host rangethan most species of North American Dactylo-gyrus (Mizelle and McDougal, 1970). Compar-
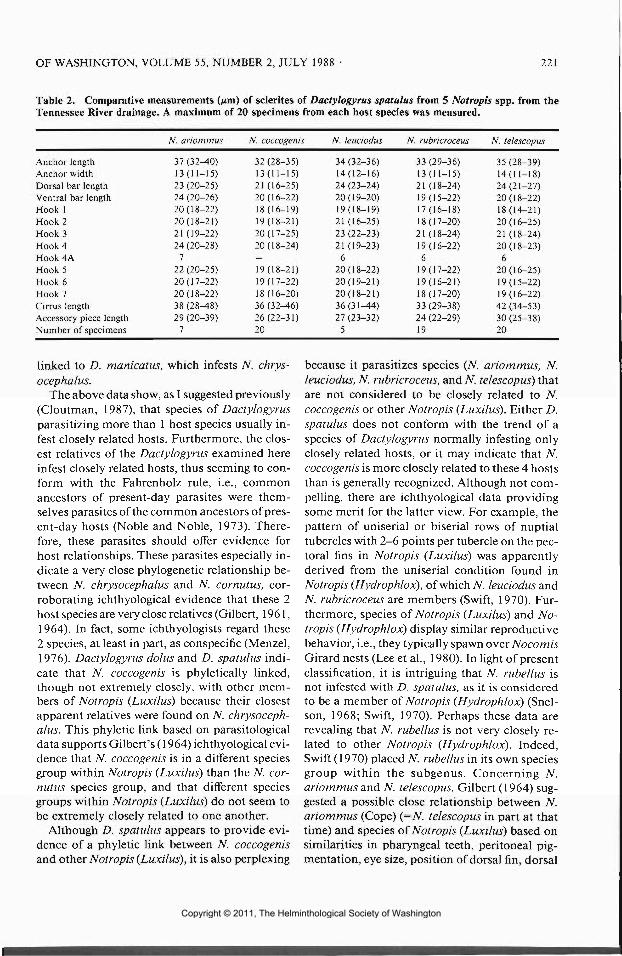
ative measurements of sclerites of D. spatulusfrom each host species are similar (Table 2).
ETYMOLOGY: The species name is Latin(spatula = shovel), referring to the broadenedcirrus shaft.
Discussion
Six of the 7 species of Notropis examined har-bored at least 1 species of Dactylogyrus (Table1). Notropis chrysocephalus chrysocephalus washost for 7 species of Dactylogyrus. Although thisis an unusually high number for most Notropishosts, it is typical for the N. cornutus speciesgroup comprising N. chrysocephalus and N. cor-nutus, 2 very closely related species of Notropis(Luxilus) (Gilbert, 1964). Notropis chrysoceph-alus and N. cornutus have been reported previ-ously as hosts for 8 and 11 species of Dacty-logyrus, respectively (Mizelle and McDougal,1970). In contrast, N. rubellus examined in thisstudy was not infested, although it has been re-ported as the host of D. orchis Mueller, 1938, inNew York (Mueller, 1938) and D. rubellus Muel-ler, 1938, in New York and Ontario (Mueller,1938;Haneketal., 1975).
The species of Dactylogyrus reported here, withthe exception of D. spatulus, displayed high hostspecificity, parasitizing only 1 species of host inthe Tennessee River drainage (Table 1). Al-though D. perlus (reported as D. banghami) hasbeen reported from several host species over awide geographical range (Mizelle and McDougal,1970), I suspect that it normally parasitizes onlythe N. cornutus species group (see Remarks forD. perlus). Otherwise, considering the wide hostrange reported elsewhere, I would expect severalof the species of Notropis examined here to har-bor D. perlus. Dactylogyrus bulbus normally in-fests only N. chrysocephalus and N. cornutuselsewhere (Mizelle and McDougal, 1970). Dac-tylogyrus circumflexus, D. delicatus, D. lepidus,and D. manicatus appear to be strictly specificfor N. chrysocephalus, and their closest apparentrelatives are known only from other Notropis(Luxilus) (Mizelle and McDougal, 1970). Dac-tylogyrus luxili has been reported from N. chrys-ocephalus, N. cornutus, and N. pilsbryi (Fowler)(Mizelle and McDougal, 1970; Hanek et al.,1975), indicating specificity for Notropis (Luxi-lus). However, it does not appear to infest N. (L.)coccogenis (Table 1). Dactylogyrus dolus appearsmonoxenous for N. coccogenis but is phyletically
Copyright © 2011, The Helminthological Society of Washington
OF WASHINGTON, VOLUME 55, NUMBER 2, JULY 1988 221
Table 2. Comparative measurements (^m) of sclerites of Dactylogyrus spatulus from 5 Notropis spp. from theTennessee River drainage. A maximum of 20 specimens from each host species was measured.
Anchor lengthAnchor widthDorsal bar lengthVentral bar lengthHook 1Hook 2Hook 3Hook 4Hook 4AHook 5Hook 6Hook?Cirrus lengthAccessory piece lengthNumber of specimens
N. ariommus
37 (32-40)13(11-15)23 (20-25)24 (20-26)20(18-22)20(18-21)21 (19-22)24 (20-28)7
22 (20-25)20(17-22)20(18-22)38 (28-48)29 (20-39)
7
N. coccogenis
32 (28-35)13(11-15)21 (16-25)20(16-22)18(16-19)19(18-21)20(17-25)20(18-24)—19(18-21)19(17-22)18(16-20)36 (32-46)26(22-31)20
N. leuciodm
34 (32-36)14(12-16)24 (23-24)20(19-20)19(18-19)21 (16-25)23 (22-23)21 (19-23)
620(18-22)20(19-21)20(18-21)36(31-44)27 (23-32)
5
N. rubricroceus
33 (29-36)13(11-15)21 (18-24)19(15-22)17(16-18)18(17-20)21 (18-24)19(16-22)6
19(17-22)19(16-21)18(17-20)33 (29-38)24 (22-29)19
N. telescopus
35 (28-39)14(11-18)24(21-27)20(18-22)18(14-21)20(16-25)21 (18-24)20(18-23)
620(16-25)19(15-22)19(16-22)42 (34-53)30 (25-38)20
linked to D. manicatus, which infests N. chrys-ocephalus.
The above data show, as I suggested previously(Cloutman, 1987), that species of Dactylogyrusparasitizing more than 1 host species usually in-fest closely related hosts. Furthermore, the clos-est relatives of the Dactylogyrus examined hereinfest closely related hosts, thus seeming to con-form with the Fahrenholz rule, i.e., commonancestors of present-day parasites were them-selves parasites of the common ancestors of pres-ent-day hosts (Noble and Noble, 1973). There-fore, these parasites should offer evidence forhost relationships. These parasites especially in-dicate a very close phylogenetic relationship be-tween N. chrysocephalus and N. cornutus, cor-roborating ichthyological evidence that these 2host species are very close relatives (Gilbert, 1961,1964). In fact, some ichthyologists regard these2 species, at least in part, as conspecific (Menzel,1976). Dactylogyrus dolus and D. spatulus indi-cate that N. coccogenis is phyletically linked,though not extremely closely, with other mem-bers of Notropis (Luxilus) because their closestapparent relatives were found on TV. chrysoceph-alus. This phyletic link based on parasitologicaldata supports Gilbert's (1964) ichthyological evi-dence that N. coccogenis is in a different speciesgroup within Notropis (Luxilus) than the N. cor-nutus species group, and that different speciesgroups within Notropis (Luxilus) do not seem tobe extremely closely related to one another.
Although D. spatulus appears to provide evi-dence of a phyletic link between N. coccogenisand other Notropis (Luxilus), it is also perplexing
because it parasitizes species (N. ariommus, N.leuciodus, N. rubricroceus, and TV. telescopus) thatare not considered to be closely related to N.coccogenis or other Notropis (Luxilus). Either D.spatulus does not conform with the trend of aspecies of Dactylogyrus normally infesting onlyclosely related hosts, or it may indicate that N.coccogenis is more closely related to these 4 hoststhan is generally recognized. Although not com-pelling, there are ichthyological data providingsome merit for the latter view. For example, thepattern of uniserial or biserial rows of nuptialtubercles with 2-6 points per tubercle on the pec-toral fins in Notropis (Luxilus) was apparentlyderived from the uniserial condition found inNotropis (Hydrophlox), of which N. leuciodus andTV. rubricroceus are members (Swift, 1970). Fur-thermore, species of Notropis (Luxilus) and No-tropis (Hydrophlox) display similar reproductivebehavior, i.e., they typically spawn overNocomisGirard nests (Lee et al., 1980). In light of presentclassification, it is intriguing that TV. rubellus isnot infested with D. spatulus, as it is consideredto be a member of Notropis (Hydrophlox) (Snel-son, 1968; Swift, 1970). Perhaps these data arerevealing that TV. rubellus is not very closely re-lated to other Notropis (Hydrophlox). Indeed,Swift (1970) placed TV. rubellus in its own speciesgroup within the subgenus. Concerning TV.ariommus and TV. telescopus, Gilbert (1964) sug-gested a possible close relationship between TV.ariommus (Cope) (=TV. telescopus in part at thattime) and species of Notropis (Luxilus) based onsimilarities in pharyngeal teeth, peritoneal pig-mentation, eye size, position of dorsal fin, dorsal
Copyright © 2011, The Helminthological Society of Washington

222 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
striping, and ecological conditions. However,Snelson (1968) considered N. ariommus and N.telescopus as members of Notropis (Notropis), andGilbert (1969) later argued against a close rela-tionship between Notropis (Luxilus) and N.ariommus and TV. telescopus. The presence of D.spatulus suggests that perhaps Gilbert (1964) wascorrect formerly in suggesting a relationship be-tween Notropis (Luxilus) and TV. ariommus andN. telescopus. The presence of Dactylogyrus sp.cf. sceptici on TV. ariommus indicates a close phy-letic link between TV. ariommus and TV. scepticus(Jordan and Gilbert), corroborating Gilbert's(1969) statement that TV. ariommus most closelyresembles TV. scepticus.
These parasitological data should not be usedalone to make inferences about host relation-ships. The possibility of host transfers that couldinvalidate inferences of close phyletic links be-tween host species cannot be ignored. However,it also seems reasonable that transfers of parasitespecies from one host species to another are mostlikely to occur between closely related hosts. Af-ter a more thorough study of the taxonomy ofDactylogyrus, cladistic analysis such as that pro-posed by Brooks (1981) should provide moreinsight into the coe volution of Dactylogyrus andNotropis. Further ichthyological studies, includ-ing genetic and biochemical analyses, are neededto confirm or reject the host relationships sug-gested here.
Acknowledgments
I thank Drs. J. Ralph Lichtenfels, USNM, MaryHanson Pritchard, HWML, and Wilmer A. Rog-ers, Auburn University, for loaning specimensof Dactylogyrus. Gary D. Hickman, TVA, andDr. Robert E. Jenkins, Roanoke College, pro-vided specimens of Notropis ariommus.
Literatur e Cited
Beverley-Burton, M. 1984. Monogenea and Turbel-laria. Pages 5-209 in L. Margolis and Z. Kabata,eds. Guide to the Parasites of Fishes of Canada.Part 1. Canadian Special Publication of Fisheriesand Aquatic Sciences 74. 209 pp.
Brooks, D. R. 1981. Hennig's parasitological method:a proposed solution. Systematic Zoology 30:229-249.
Chien, S. M. 1971. Dactylogyrids from North Amer-ican cyprinids of the genus Nocomis. The recipro-cus species group. Journal of Parasitology 57:1211-1214.
. 1974a. Dactylogyrids from North Americancyprinids of the genus Nocomis. The bellicus group.Journal of Parasitology 60:585-594.
. 1974b. Dactylogyrids from North Americancyprinids of the genus Nocomis: the limulus andthe mollis groups. Journal of Parasitology 60:773-776.
Cloutman, D. G. 1974. Monogenean and copepodparasites of fishes from the Smoky Hil l River, EllisCounty, Kansas. Transactions of the KansasAcademy of Science 77:225-229.
. 1980. Two new species of Dactylogyrus(Monogenea: Dactylogyridae) from Notropis scab-riceps (Cope) and N. scepticus (Jordan and Gilbert)(Pisces: Cyprinidae). Journal of Parasitology 66:160-161.-. 1987. Dactylogyrus (Monogenea: Dactylo-
gyridae) from Hybopsis and Notropis (Cyprinella)(Pisces: Cyprinidae) from the Tennessee Riverdrainage, with descriptions of three new speciesand remarks on host relationships. Proceedings ofthe Helminthological Society of Washington 54:78-83.
Cone, D. K. 1980. The Monogenea of some fresh-water fishes in New Brunswick, Canada. Journalof Parasitology 66:178-180.
Gilbert , C. R. 1961. Hybridization versus intergra-dation: an inquiry into the relationship of two cyp-rinid fishes. Copeia 1961:181-192.
. 1964. The American cyprinid fishes of thesubgenus Luxilus (genus Notropis). Bulletin of theFlorida State Museum (Biological Sciences) 8:95-194.-. 1969. Systematics and distribution of the
American cyprinid fishes Notropis ariommus andNotropis telescopus. Copeia 1969:474-492.
Hanek, G., K. Molnar , and C. H. Fernando. 1975.New and previously known Dactylogyrus spp. fromsouthern Ontario fishes. Journal of Parasitology61:421-426.
Lee, D. S., C. R. Gilbert , C. H. Hocutt, R. E. Jenkins,D. E. McAllister , and J. R. Stauffer, Jr. 1980.Atlas of North American freshwater fishes. NorthCarolina State Museum of Natural History, Ra-leigh. 854 pp.
Menzel, B. W. 1976. Biochemical systematics andevolutionary genetics of the common shiner speciesgroup. Biochemical Systematics and Ecology 4:281-293.
Mizelle, J. D., and J. W. Crane. 1964. Studies onmonogenetic trematodes, XXIII . Gill parasites ofMicroptems salmoides (Lacepede) from a Califor-nia pond. Transactions of the American Micro-scopical Society 83:343-348.
, and M. A. Donahue. 1944. Studies on mono-genetic trematodes. XI . Dactylogyridae from Al-gonquin Park fishes. American Midland Naturalist31:600-624.
, and A. R. Klucka. 1953. Studies on mono-genetic trematodes. XIV . Dactylogyridae fromWisconsin fishes. American Midland Naturalist49:720-733.
, and H. D. McDougal. 1970. Dactylogyrus inNorth America. Key to species, host-parasite andparasite-host lists, localities, emendations, anddescriptions of D. kritskyi sp. n. American Mid-land Naturalist 84:444-462.
, and F. O. Webb. 1953. Studies on monoge-
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 55, NUMBER 2, JULY 1988 223
netic trematodes XV. Dactylogyridae from Alas-ka, Wisconsin, and Wyoming. American MidlandNaturalist 50:206-217.
Molnar, K., G. Hanek, and C. H. Fernando. 1974.Parasites of fishes from Laurel Creek, Ontario.Journal of Fish Biology 6:717-728.
Mueller , J. F. 1938. Additional species of NorthAmerican Gyrodactyloidea (Trematoda). Ameri-can Midland Naturalist 19:220-235.
Noble, E. R., and G. A. Noble. 1973. Parasitology.The Biology of Animal Parasites, 3rd ed. Lea andFebiger, Philadelphia, Pennsylvania. 617 pp.
Putz, R. E., and G. L. Hoffman. 1963. Two new Gy-rodactylus (Trematoda: Monogenea) from cypri-nid fishes with synopsis of those found on NorthAmerican fishes. Journal of Parasitology 49:559-566.
Rogers, W. A. 1967. Studies on Dactylogyrinae(Monogenea) with descriptions of 24 new speciesof Dactylogyrus, 5 new species of Pellucidhaptor,and the proposal of Aplodiscus gen. n. Journal ofParasitology 53:501-524.
Snelson, F. F., Jr. 1968. Systematics of the cyprinidfish Notropis amoenns, with comments on the sub-genus Notropis. Copeia 1968:776-802.
Swift, C. C. 1970. A review of the eastern NorthAmerican cyprinid fishes of the Notropis texanusspecies group (subgenus Alburnops), with a defi-nition of the subgenus Hydrophlox, and materialsfor a revision of the subgenus Alburnops. Ph.D.Dissertation, Florida State University, Tallahas-see. 476 pp.
Copyright © 2011, The Helminthological Society of Washington